
A Plant Growth Promoting Rhizobacterium and Temperature Effects on Performance of 18 Clones of...
8
Amer J of Potato Res (1998) 75:145-152 145 A Plant Growth Promoting Rhizobacterium and Temperature Effects on Performance of 18 Clones of Potato Salah Bensalim, Jerzy Nowak 1, and Samuel I~ Asiedu 1Affiliation:Department of Plant Science, NovaScotiaAgriculturalCollege,Truro, NS, CanadaB2N 5E3. Correspondingauthor. Correspondence/communication address: Dr. Jerzy Nowak,Department of Plant Science, NovaScotiaA~ricultural College, Truro, NovaScotia~CanadaB2N 5E3;Tel: (902)893-6686; Fax: (902) 897-9762; E-mail: [email protected] ABSTRACT A survey of genotypic responses to beneficial bac- terium (Pseudomonas sp. strain PsJN) was conducted in vitro and ex vitro, under two temperature condi- tions, using eighteen clones of potato of different heat stress tolerance: temperate adapted cultivars Ken- nebec and Russet Burbank; heat tolerant DTO-2, DTO- 28, DTO-33, LT-1, LT-2, LT-5, LT-6, LT-7, LT-8, LT-9, Y84-02, NDD277-2, D~sir~e, and Maine-47; and heat sensitive abscissic acid (ABA)-deficient mutants 11401-01 and 9120-05. Nodal explants taken from 6- week-old bacterized and non-bacterized control plantlets were cultured in vitro on a hormone-free potato nodal cutting medium, and placed at either 20/15 C or 33/25 C day/night temperature, 12h pho- toperiod and 250 pE m -2 s-1 mixture of fluorescent and incandescent light, for six weeks. The tuberization response was studied ex vitro after two weeks accli- mation of 2-week old plantlets at 33/25C. The accli- mated plantlets were transplanted to 3L plastic nursery pots containing peat-based Pro-Mix growing medium and placed in growth chambers at either 20/15 or 33/25 C day/night temperature, 12 h photoperiod, 475 IrE m 2 s ~ light and =80% RH, for 12 weeks. Com- pared to the non-bacterized controls, bacterization sig- nificantly increased stem length of 12, shoot biomass of 9, and root biomass of 2 clones at 20/15C; and stem length of 14, shoot biomass of 15, and root biomass of 13 clones at 33/25C. High temperature increased length of internodes and had either no effect or slightly de- creased node number. Temperature increase had the most dramatic effect on root development. An average shoot to root ratio decreased from 3.7 at 20/15 C to 1.7 Acceptedfor publication February9, 1998. ADDITIONAL KEY WORDS:Beneficial bacterium, heat tolerance, Solanum ~berosum, in vitro, ex vitro. at 33/25 C for non-bacterized plantlets and, respec- tively, from 4.3 to 1.5 for bacterized. The beneficial effect of bacterization on root biomass was the most pronounced in LT-1 and Maine-47 at 20/15 C and LT-8, Maine 47, DTO-2, Kennebec, NDD277-2 and 11401-01 at 33/25C. The temperature elevation did not signi- ficantly affect root biomass of LT-6, DTO-28 and Ddsir~e. Temperature stress caused severe reduction in tuber number and tuber fresh weight. ABA-deficient mutants did not produce any tubers and LT-8, LT-9, Y84-027 and DTO-28 tuberized very poorly at 33/25C. DTO-33, D~sir~e, LT-1 and Kennebec gave the highest number of tubers per pot and Kennebec, LT-1, D~sir~e and LT-7 the highest yields at this temperature. There was no significant effect of bacterization on tuber- ization at 20/15 C but at 33/25 C bacterization signifi- cantly enhanced tuber number and weight in LT-7 and reduced tuber weight in DTO-2. Although there was no clear link between the in vitro response of particular clones to bacterization and their heat stress tolerance, improvement of ex vitro performance of heat tolerant LT-7 indicates that rhizosphere bacteria may play a role in clonal adaptation of potato to heat stress. INTRODUCTION Heat stress tolerance in potato is strongly influenced by genotype and environment (Mendoza and Estrada 1979, Mid- more 1992). Availability of water (Trebajo and Midmore 1990), humidity (Mendoza and Estrada 1979), irradiance (Menzel 1985), photoperiod and nitrogen are considered the most critical environmental factors affecting response to heat in potato (reviewed in Ewing 1995). in vitro experi- ments also indicate differential responses of potato clones to constituents of growing media (Nowak and Colborne 1989, Nowak and Asiedu 1992), and beneficial bacteria (Nowak et al. 1995 and 1997, Conn eta/. 1997). To our knowl- edge, there is no published information on the role of rhizo- sphere microflora in heat responses of potato.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of A Plant Growth Promoting Rhizobacterium and Temperature Effects on Performance of 18 Clones of...

Amer J of Potato Res (1998) 75:145-152 145
A Plant Growth Promoting Rhizobacterium and Temperature
Effects on Performance of 18 Clones of Potato
Salah Bensalim, J e r zy N o w a k 1, and Samuel I~ Asiedu
1Affiliation: Department of Plant Science, Nova Scotia Agricultural College, Truro, NS, Canada B2N 5E3. Corresponding author. Correspondence/communication address: Dr. Jerzy Nowak, Department of Plant Science, Nova Scotia A~ricultural College,
Truro, Nova Scotia~ Canada B2N 5E3; Tel: (902) 893-6686; Fax: (902) 897-9762; E-mail: [email protected]
ABSTRACT
A survey of genotypic responses to beneficial bac-
ter ium ( P s e u d o m o n a s sp. s train PsJN) was conducted
in v i t ro and e x v i t ro , unde r two t empe ra tu r e condi-
tions, using eighteen clones of pota to o f different heat
s t r e ss to le rance : t e m p e r a t e adap ted cul t ivars Ken-
nebec and Russet Burbank; heat to lerant DTO-2, DTO-
28, DTO-33, LT-1, LT-2, LT-5, LT-6, LT-7, LT-8, LT-9,
Y84-02, NDD277-2, D~sir~e, and Maine-47; and hea t
s ens i t i ve absc i ss ic ac id (ABA)-de f i c i en t m u t a n t s
11401-01 and 9120-05. Nodal explants t aken f rom 6-
week -o ld b a c t e r i z e d and n o n - b a c t e r i z e d c o n t r o l
p l an t l e t s were cu l tu red in v i t ro on a ho rm one - f r ee
p o t a t o nodal cu t t ing medium, and p laced a t e i t he r
20/15 C or 33/25 C day/night t empera tu re , 12h pho- toper iod and 250 pE m -2 s-1 mixture of f luorescent and
incandescen t light, fo r six weeks. The t ube r i za t i on
response was studied ex vi tro af ter two weeks accli-
mat ion of 2-week old p lant le ts a t 33/25C. The accli-
m a t e d p l a n t l e t s w e r e t r a n s p l a n t e d t o 3L p las t i c
nurse ry pots containing peat-based Pro-Mix growing
medium and placed in growth chambers a t e i ther 20/15
or 33/25 C day/night t empera ture , 12 h photoper iod,
475 IrE m 2 s ~ light and =80% RH, for 12 weeks. Com-
pared to the non-bacterized controls, bacterizat ion sig-
nificantly increased stem length of 12, shoot biomass of
9, and roo t biomass o f 2 clones at 20/15C; and s tem
length of 14, shoot biomass o f 15, and root biomass of
13 clones at 33/25C. High temperature increased length
o f internodes and had e i ther no effect or slightly de-
creased node number. Tempera ture increase had the
most dramatic effect on roo t development. An average
shoot to root rat io decreased from 3.7 at 20/15 C to 1.7
Accepted for publication February 9, 1998. ADDITIONAL KEY WORDS: Beneficial bacterium, heat tolerance, Solanum ~berosum, in vitro, ex vitro.
at 33/25 C for non-bacter ized p lant le ts and, respec-
tively, f rom 4.3 to 1.5 for bacter ized. The beneficial
effect o f bacter izat ion on roo t biomass was the most
pronounced in LT-1 and Maine-47 a t 20/15 C and LT-8,
Maine 47, DTO-2, Kennebec, NDD277-2 and 11401-01
a t 33/25C. The t empera tu re e levat ion did no t signi-
f i c an t ly a f f e c t r o o t b iomass o f LT-6, DTO-28 and
Ddsir~e. Temperature stress caused severe reduct ion
in tuber number and tuber fresh weight. ABA-deficient
mutan t s did no t produce any tubers and LT-8, LT-9,
Y84-027 and DTO-28 tuberized very poorly at 33/25C.
DTO-33, D~sir~e, LT-1 and Kennebec gave the highest
number o f tubers per pot and Kennebec, LT-1, D~sir~e
and LT-7 the highest yields a t this temperature. There
was no s ignif icant effect o f bac te r i za t ion on tuber-
izat ion a t 20/15 C but at 33/25 C bacterizat ion signifi-
cantly enhanced tuber number and weight in LT-7 and
reduced tuber weight in DTO-2. Although there was no
clear link between the in v i tro response of part icular
clones to bacterization and the i r heat stress tolerance,
improvement of ex vi tro performance of heat to le rant
LT-7 indicates tha t rh izosphere bac te r ia may play a
role in clonal adaptat ion of po ta to to heat stress.
INTRODUCTION
Heat stress tolerance in potato is strongly influenced by
genotype and environment (Mendoza and Estrada 1979, Mid-
more 1992). Availability of water (Trebajo and Midmore
1990), humidity (Mendoza and Estrada 1979), irradiance
(Menzel 1985), photoperiod and nitrogen are considered the
most critical environmental factors affecting response to
heat in potato (reviewed in Ewing 1995). in vitro experi-
ments also indicate differential responses of potato clones
to constituents of growing media (Nowak and Colborne
1989, Nowak and Asiedu 1992), and beneficial bacteria
(Nowak et al. 1995 and 1997, Conn eta/. 1997). To our knowl-
edge, there is no published information on the role of rhizo-
sphere microflora in heat responses of potato.

146 AMERICAN JOURNAL OF POTATO RESEARCH Vol. 75
Beneficial bacteria, designated as plant-growth promot- ing rhizobacteria (PGPR) (Kloepper et al. 1988), colonize
roots, enhance shoot emergence, and stimulate plant growth either directly, by producing plant hormones and improving
nutrient uptake, or indirectly, by changing the microbial bal-
ance in the rhizosphere in favor of the beneficial microor- ganisms (reviewed by: Brown 1974, Kloepper et al. 1988,
Glick 1995, Lazarovits and Nowak 1997). A non-fluorescent PGPR, P s ~ sp. strain PsJN, isolated in our labora-
tory, increases root branching, root and shoot dry weights, node number, stem length and lignification, leaf pubescence
and chlorophyll content in tissue culture grown potato plantlets (Frommel et al. 1991). Bacterized plantlets have
fully functioning stomata (Frommel et al. 1991) and tolerate transplant stress much better than non- bacterized controls
(Nowak et al. 1995, Nowak et al. 1997). Furthermore, they have higher cytokinin content and, after transplanting, initi-
ate stolons and tubers earlier than non-bacterized controls
(Lazarovits and Nowak 1997, Dunbar 1997). Clonal variation in the bacterium stimulation of root sys-
tem development, both, i n vitro (Conn et al., 1997) and ex vitro (Dunbar, 1997), and overall improvement of transplants
water management (Nowak et al., 1995; Lazarovits and Nowak, 1997) in particMar, lead us to hypothesize that rhizo-
sphere bacteria modify potato responses to heat stress. The objective of this study was to survey bacterization effects on
i n vitro and ex vitro performance of potato clones of varied heat stress tolerance under elevated temperatures.
MATERIALS AND METHODS
P l a n t Mater ia l a n d Cul ture
Disease-indexed tissue culture plantlets of 14 potato
clones selected for heat tolerance, LT-1, LT-2, LT-5, LT-6, LT- 7, LT-8, LT-9, DTO-2, DTO-28, DTO-33, NDD277-2, Y84-027, D6sir6e, and Maine-47, were obtained from the International
Potato Centre (CIP), Lima, Peru. Plantlets of two temperate adapted cultivars, Kennebec and Russet Burbank, were from
the Plant Propagation Centre, New Brunswick Department of Agriculture, Fredericton, NB, Canada. Two abscissic acid (ABA)-deficient mutants, diploids, 11401-01 and 9120-05, sim-
ilar to the "droopy" mutant (Simmonds 1965, Quarrie 1982), were supplied as tubers by Dr. Henry de Jong, Agriculture
and Agri-Food Canada Research Station, Fredericton, NB, Canada. Tissue culture of the mutants was initiated in our
laboratory. All clones were cultured in 25 X 150 mm test tubes, using single-node explants, on 10 ml of Murashige and
Skoog based, hormone-free, potato nodal cutting medium
(PNCM) as in Sipos et al. (1988). Unless specified otherwise, plantlets were maintained and multiplied under 16h photo-
period, 160-180 HE m -2 s -1 fluorescent light and 22/19 C day/
night growth room temperature.
I n v i t r o B a c t e r i z a t i o n
A plant growth promoting, non-fluorescent, Pseudomo-
nas sp. strain PsJN, originally isolated from surface sterilized
onion roots by J. Nowak (reported by Herman, 1987), was
maintained i n p lan ta , using i n v i t r o grown Kennebec plantlets. The bacterial inoculum was prepared as in Pillay
and Nowak (1997). About 1 cm long nodal explants, taken from 6-week-old plantlets, were dipped in the inoculum for 30 sec, blotted with sterile filter paper, and placed on PNCM.
Non-bacterized controls were immersed in phosphate buffered saline only. The cultures were grown in the growth
room as above.
Tempera ture a n d B a c t e r i z a t i o n Effects on P lan t l e t Growth
Tempera ture and bacter izat ion effects on i n v i t ro
growth of 18 potato clones were determined using nodal
explants taken from 6-week old bacterized and non-bacter- ized plantlets. The explants were cultured as above and
placed in two growth chambers, 20/15 C and 33/25 C day/
night temperatures, respectively, and 250 p E m ~ s -1 fluores- cent and incandescent lights mixture, at 12h photoperiod. After 6 weeks, plantlets were harvested and stem length,
node number, and shoot (stem and leaves) and root dry weights determined (after forced dry air drying at 60 C for 48h). The experiment was conducted twice.
Ex vitro Tuber i za t ion
Nodal explants of 18 bacterized and non-bacterized clones were grown i n vitro in 20/15 C growth chamber as
above, for two weeks. Two-week-old plantlets of the same size were randomly assigned to two groups and transplanted into 3L plastic nursery pots filled with a peat based Pro-Mix
growing medium (ASB Greenworld, Point Spain, NB,
Canada), one plant per pot. Group I transplants were placed in 20/15 C growth chamber and group II in 33/25C. Light intensity, provided by a mixture of fluorescent and incan-
descent lights and measured with LI-COR quantum sensor, was 475 pE m 2 s-', with 12h photoperiod and =80% RH. All
plants were watered daily, and fertilizer was applied weekly, starting one week after transplanting (WAT). Weekly fertil-

1998 BENSALIM, eta/.: PERFORMANCE OF CLONES OF POTATO 147
ization was as described in Dunbar (1997). During weeks 2-
5, a stock solution of 50 and 20 grams of 10-52-10 and 13-0-44
N:P:K, respectively, and 50 g of magnesium sulphate, and 3
g of the chelated micro-nutrients (Plant Products Co. Ltd.,
Brampton, ON, Canada) was mixed in 1L distilled water
(Stock A). In a separate flask, 20 g of calcium nitrate (15.5-
0-0) was dissolved in 1L distilled water (Stock B). Aliquats of
10 mL of each stock were mixed in 1L water prior to each
fertilization. From week 8 on, 13-0-44 (Stock A) and 15.5-0-0
(Stock B) were both increased to 50 g, and 10-52-10 was
decreased to 20 g (Stock A). To promote tuberization, during
the weeks 6 and 7 the plants did not receive any nutrients.
Fertilization was terminated on week 10. Tubers were har-
vested 12 WAT and tuber number and fresh weight per plant
were determined. The conditions for the experiment above
were established after a preliminary experiment with 11
potato clones and 4 replicates of each combination of treat-
ments was conducted.
E x p e r i m e n t a l D e s i g n a n d D a t a A n a l y s i s
A split plot factorial design, with two temperature levels
(20/15 C and 33/25 C), two bacterization levels (bacterized
and non-bacterized) and 18 potato clones was used to deter-
mine the effects of temperature and bacterization on in vitro
potato plantlet growth. Temperature was assigned to the
main plot, whereas the subplot contained the factorial com-
bination of bacterization and clones. Each treatment combi-
nation was replicated 12 times. To assess tuberization
responses to temperature and bacterization, a factorial
arrangement of two levels of bacterization and two tempera-
tures, with 18 potato clones was used in a randomized com-
plete block design, with 3 replicates. Analysis of variance (ANOVA) was performed using a
computer statistical analysis program (SAS Institute Inc.,
Cary, NC). Treatment means were separated using LSD.05. To
avoid the deflation of the variance in the ex vitro experiment,
ABA-deficient mutants response was excluded from the
ANOVA.
RESULTS
I n v i t r o P l a n t l e t G r o w t h
Temperature X bacterization X clone interactions were
highly significant (P=0.0001) for in vitro growth responses of
FIGURE 1 Ef fec t ofPseudomonas sp. s train PsJN, on growth o f DTO-2 (A),11401-01 (B) , and LT-7 (C) potato clones cultured in vitro at 20/15 C and 33/25C day/night temperatures.

148 AMERICAN JOURNAL OF POTATO RESEARCH Vol. 75
A 1 2 ,
LSD(0 05) = 0 8
11 ~
a. 10
i 9 ,
8 ,
-t I
20 33
B 12 .
LSD(0 05) = 0 8
lO
9 ,
8 ,
7 , I 33
6O
55
50
45
v 40
3o
2O
20
c LSD(0 G5) = 4 9 mg
/
33
4O
35
30
~ zs ~ 2o
I:1
F I G U R E 2
D
Temperature (C ~ 20 33 Temperature (C ~
Temperature X bacterization interaction effect on stem length (A), node number (B) , shoot dry weight (C) , and root dry weight (D) o f 18 clones o f potato co-cultured in vitro with strain PsJN. [ ] Non-bacterized, �9 Bacterized.
the tested potato clones. Response variation is represented by DTO-2, 11401-01, and LT-7 in Fig.1 (& B, and C, respec-
tively). Significant interaction between temperature and bac- terization was recorded for stem length (P=0.0352), and shoot (P=0.0013) and root (P=0.0001) dry weights (Fig. 2A~ C and D, respectively). There was no significant interaction between
these two factors and the node number (Fig. 2B). On average, at 20/15C, bacterized plantlets were taller (86%), had more nodes (19%), and contained higher shoot (62~ and root (57~ dry weights than non-bacterized controls (Table 1). The dif-
ference in growth responses between bacterized and control treatments was enhanced at 33/25C; the difference in the bio- mass accumulation was 162% and 129~ for root and shoot dry
weights, respectively. Compared to 20/15C, at 33f25 C bacter- ized plants had 3120/5 higher root and 54% higher shoot bio- mass and 27% longer stems. Node number was, however, 15%
greater at 20/15 C (Fig. 2B). Temperature increase caused reallocafion of biomass from shoot to root portion of plantlets (Table 1); an average value of the shoot to root ratio was 3.7
at 20/15 C and 1.7 at 33/25C. This reallocation was even more dramatic in bacterized plantlets; shoot/root ratio decreased
from 4.3 at 20/15 C to 1.5 at 33/25C. Compared to the non-bacterized controls, bacterization
at 20/15 C significantly increased shoot biomass in 1104-01,
LT-1, LT-5, LT-9, DTO-2, DTO-28, Maine-47, D~sir~e, and
NDD277-2, but not in 9120-05, LT-2, LT-7, LT-8, DTO-33, Ken- nebec, Russet Burbank, and Y84-027 (Table 1). Clones 1104- 01, DTO-2, Y84-027, LT-7 and NDD277-2 benefitted the most
and LT-9, LT-6 and LT-1, the least at 33/25C. The beneficial effect of bacterization on root biomass at
20/15 C was only significant in Maine-47 (160%) and LT-1 (156% stimulation). On the other hand, 13 clones benefitted
significantly from this treatment at 33f25 C (Table 1). Bacter- ized LT-8, Maine-47, DTO-2, Kennebec, NDD277-2 and 11401-
01 had the largest root mass at the elevated temperature and LT-6, DTO-28 and D~sir~e, the smallest. The most pronounced
difference in root biomass between bacterized and control plants at 33/25 C was in DTO-2, Kennebec and 11401-01 (54.9,
42.1 and 39.4 mg per plantlet, respectively). Root growth stim- ulation at the higher temperature lowered shoot/root (S/R) dry matter ratio in all clones (Table 1); in Y84-027 (278%), Ken-
nebec (250%), LT-8 (221%), LT-9 (207~ and Russet Burbank
(181%) the most and DTO-33 (13~ LT-5 (30~ LT-6 (60%), 11401-01 (66%) and LT-2 (71%), the least. This ratio was low- ered even further in clones which best responded to bacteri- zation at 33/25C, i.e. Kennebec (511%), NDD277-2 (391%), LT-8
(308%), Russet Burbank (300~ and 11401-01 (294%). High temperature (33/25C) significantly inc.reased total
biomass (shoot plus root) of non-bacterized plantlets only in LT-8, Maine-47, LT-9, and LT-1. There was no significant dif-

1998 BENSALIM, eta/.: P E R F O R M A N C E O F CLONES O F POTATO 149
TABLE 1.--Effects of temperature and bacterization wi th P s e u d o m o n a s sp. strain PsJN, on in vitro growth responses of 18
potato clones.
Clone SL NN SDW RDW TB S/R Trt T1 T2 T1 T2 T1 T2 T1 T2 T1 T2 T1 T2
11401-01
9120-05
LT-1
LT-2
LT-5
LT-6
LT-7
LT-8
LT-9
DTO-2
DTO-28
DTO-33
Kennebec
R.Burbank
Maine-47
D~sir~e
NDD277-2
Y84-027
C 3.3 7.0 7.0 10.8 8.0 13.7 2.0 5.8 10.0 19.5 4.0 2.4 B 13.8 12.5 17.2 12.2 63.3 70.2 5.7 45.2 42.0 115.4 6.3 1.6
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 5.5 8.1 9.3 10.8 14.5 22.8 2.7 8.3 17.2 31.1 5.4 2.7 B 8.8 15.5 8.4 12.9 25.3 64.3 4.9 28.5 30.2 92.8 5.2 2.3
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 5.8 5.7 11.1 6.8 28.3 31.6 9.9 29.3 38.2 60.9 2.9 1.1 B 11.2 8.9 11.3 8.2 52.9 36.6 25.4 34.4 78.3 71.0 2.1 1.1
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 5.9 7.6 11.3 10.2 22.3 30.1 7.7 17.7 30.0 47.8 2.9 1.7 B 6.3 9.1 9.9 9.1 24.1 43.5 6.1 34.6 30.2 78.1 3.9 1.3
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 6.6 9.8 12.3 9.8 23.0 30.3 6.6 11.3 29.6 41.6 3.5 2.7 B 10.6 13.7 12.8 10.1 36.5 83.8 5.9 48.0 42.4 131.6 6.2 1.8
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 4.3 4.9 8.9 7.2 15.4 14.3 6.5 9.3 21.9 23.6 2.4 1.5 B 4.3 5.9 8.5 6.8 16.7 15.7 8.0 9.9 24.7 25.6 2.1 1.6
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 2.0 2.2 6.4 4.5 20.7 16.7 4.4 7.3 25.1 24.0 4.7 2.3 B 3.3 9.8 7.8 8.9 30.0 68.4 5.4 37.2 35.4 105.6 5.5 1.8
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 8.3 7.4 11.4 8.0 41.3 53.3 9.2 38.6 50.5 91.9 4.5 1.4 B 9.1 13.2 12.1 9.2 48.0 92.7 9.6 75.4 57.6 168.1 4.9 1.2
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 5.8 7.7 11.3 8.1 31.2 37.0 7.8 28.1 39.0 65.1 4.0 1.3 B 11.4 8.3 12.3 8.3 50.1 40.1 11.3 30.7 61.4 70.8 4.4 1.3
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 3.8 4.6 11.1 6.3 19.5 15.8 5.0 7.3 24.5 23.1 3.9 2.2 B 9.9 14.2 12.4 11.8 35.2 78.2 9.3 62.2 44.5 140.4 3.8 1.3
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 3.1 3.6 7.3 5.9 20.3 14.7 5.2 8.6 25.5 23.3 3.9 1.7 B 8.6 7.0 11.1 7.9 35.3 29.6 8.6 16.6 43.9 46.2 4.1 1.8
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 4.1 4.5 9.1 7.1 16.9 22.3 6.8 10.0 23.7 32.3 2.5 2.2 B 6.3 10.9 11.1 10.0 24.3 48.8 6.4 30.1 30.7 78.9 3.8 1.6
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 6.2 5.0 12.2 6.6 30.3 23.8 7.2 9.8 37.5 43.6 4.2 1.2 B 10.2 11.7 12.6 9.4 42.0 47.6 7.7 52.3 49.7 99.9 5.5 0.9
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 5.0 4.9 11.3 7.2 25.5 20.2 5.7 12.8 31.2 33.0 4.5 1.6 B 8.4 13.4 12.3 10.1 31.6 54.6 5.7 39.3 37.3 93.9 5.6 1.4
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 6.5 8.5 11.9 11.2 38.1 47.4 10.0 28.4 48.1 75.8 3.8 1.7 B 10.0 15.4 11.9 13.5 57.6 77.6 26.0 62.6 83.6 140.2 2.2 1.2
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 3.9 5.4 8.1 6.6 13.4 17.7 4.8 11.7 18.2 29.4 2.8 1.5 B 10.2 10.4 11.0 9.4 40.2 44.8 13.8 21.3 54.0 66.1 2.9 2.1
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 2.6 4.8 9.2 8.2 13.9 13.7 3.9 10.4 17.8 24.1 3.6 1.3 B 7.8 8.4 10.0 10.0 30.0 52.1 5.5 47.9 35.5 100.0 5.4 1.1
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
C 3.8 3.5 6.3 6.3 24.0 11.6 7.0 12.0 31.0 23.6 3.4 0.9 B 5.5 9.9 8.7 8.8 32.1 48.0 7.7 35.5 39.8 83.5 4.2 1.4
Values per plantlet are means of 12 replicates. Treatment Crrt): T1, 20/15 C; T2, 33/25 C; C, control; B, bacterized plantlets. LSD 05 values: stem length (SL), 2.9 cm; node number (NN), 3.0; shoot dry weigh (SDW), 12.7 mg; root dry weight (RDW), 10.2 mg; total biomass (TB), 18.1 mg.
f e r e n c e in t h e b i o m a s s a c c u m u l a t e d in e i t h e r t h e h e a t to le r -
a n t DTO-33, DTO-28, DTO-2, LT-7, LT-6, D~sir~e, NDD277-2,
a n d Y84-027, t e m p e r a t e a d a p t e d R u s s e t B u r b a n k , o r t h e h e a t
s e n s i t i v e 11401-01 a n d 9120-05. B a c t e r i z a t i o n a t 20 /15 C
i m p r o v e d to ta l b i o m a s s a c c u m u l a t i o n in 7 c lones , Maine-47,
LT-1, LT-9, D~sir~e, DTO-2, DTO-28, a n d 11401-01 as c o m -
p a r e d to t he n o n - b a c t e r i z e d con t ro l s , w h e r e a s b a c t e r i z a t i o n
a t 33/25 C i n c r e a s e d to ta l b i o m a s s a c c u m u l a t i o n in 15 c l o n e s
(Tab le 1). Bac t e r i zed LT-8 a c c u m u l a t e d t h e h i g h e s t b i o m a s s ,
f o l l o w e d b y t he b a c t e r i z e d DTO-2, Maine-47, a n d LT-5.
A l t h o u g h t h e r e w e r e c l o n a l d i f f e r ences in r e s p o n s e to
33/25 C in vitro, s e p a r a t i o n o f h e a t t o l e r a n t a n d s e n s i t i v e
c l o n e s w a s n o t poss ib le .
Ex vitro Tuber iza t ion
A l t h o u g h t e m p e r a t u r e s t r e s s (33 /25C) c a u s e d s e v e r e
r e d u c t i o n in t u b e r n u m b e r a n d t u b e r f r e s h weight , h ighly sig-
n i f i can t (P=0.0001) t e m p e r a t u r e X c l o n e i n t e r a c t i o n s w e r e
r e c o r d e d for b o t h var iables . A s ign i f ican t (P=0.0365) t empe r -
a tu re X b a c t e r i z a t i o n i n t e r ac t i on fo r t u b e r n u m b e r w a s a lso
found . I n d e p e n d e n t l y o f bac t e r i za t ion , a t 33/25C, ABA-defi-
c i en t c lones d id n o t p r o d u c e a n y t u b e r s a n d c lones LT-8, LT-
9, Y84-027 a n d DTO-28 t u b e r i z e d v e r y p o o r l y ( T a b l e 2).
DTO-33, D~sir~e, LT-1, a n d K e n n e b e c gave t he h ighes t h u m -
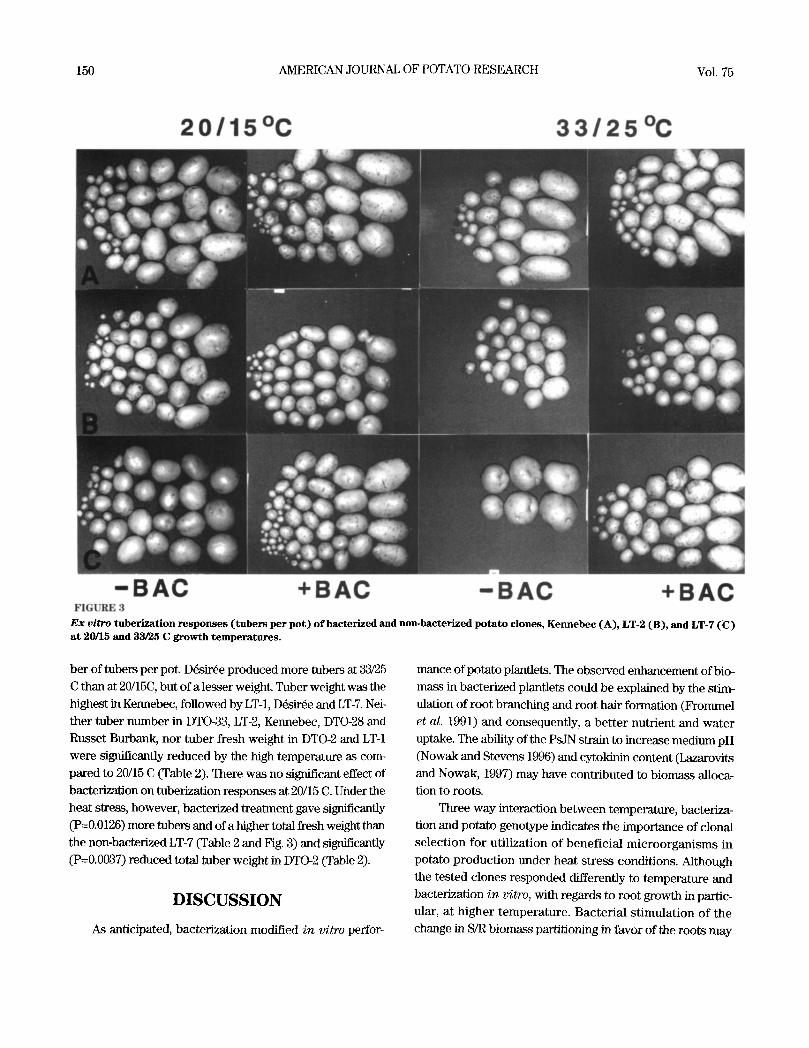
150 AMERICAN JOURNAL OF POTATO RESEARCH Vol. 75
E x vi tro tuberization responses (tubers per pot ) o f bacterized and non-bacterized potato clones, Kennebec (A) , LT-2 (B), and LT-7 (C) at 20/15 and 33/25 C growth temperatures.
ber of tubers per pot. D~sir~e produced more tubers at 33/25
C than at 20/15C, but of a lesser weight. Tuber weight was the highest in Kennebec, followed by LT-1, D~sir~e and LT-7. Nei- ther tuber number in DTO-33, LTb2, Kennebec, DTO-28 and
Russet Burbank, nor tuber fresh weight in DTO-2 and LT-1 were significantly reduced by the high temperature as com- pared to 20/15 C (Table 2). There was no significant effect of bacterization on tuberization responses at 20/15 C. Under the
heat stress, however, bacterized treatment gave significantly (P=0.0126) more tubers and of a higher total fresh weight than the non-bacterized LT-7 (Table 2 and Fig. 3) and significantly
(P=0.0037) reduced total tuber weight in DTO-2 (Table 2).
DISCUSSION
As anticipated, bacterization modified i n vitro perfor-
mance of potato plantlets. The observed enhancement of bio-
mass in bacterized plantlets could be explained by the stim- ulation of root branching and root hair formation (Frommel
et al. 1991) and consequently, a better nutrient and water uptake. The ability of the PsJN strain to increase medium pH (Nowak and Stevens 1996) and cytokinin content (Lazarovits
and Nowak, 1997) may have contributed to biomass alloca- tion to roots.
Three way interaction between temperature, bacteriza- tion and potato genotype indicates the importance of clonal selection for utilization of beneficial microorganisms in potato production under heat stress conditions. Although the tested clones responded differently to temperature and bacterization i n vitro, with regards to root growth in partic- ular, at higher temperature. Bacterial stimulation of the change in S/R biomass partitioning in favor of the roots may

1998 BENSALIM, eta/.: PERFORMANCE OF CLONES OF POTATO 151
TABLE 2. - -Temperature and bacterization effects on tuber
yield o f 18 clones o f potato at two dif ferent
temperatures.
Tuber Number Fresh weight (g)
Clone Trt 20/15 C 33/25 C 20/15 C 33/25 C
11401.011" C 21.3 0.0 34.7 0.0 B 19.7 0.0 16.7 0.0
9120-05 1" C 5.0 0.0 7.9 0.O B 8.0 0.0 15.2 0.0
LT-1 C 27.3 13.0 158.9 119.3 B 18.0 17.3 136.9 109.9
LT-2 C 16.0 11.0 122.9 62.1 B 14.3 9.7 120.1 86.2
LT-5 C 29.7 16.0 199.5 102.4 B 22.3 8.3 174.1 89.9
LT-6 C 23.3 13.0 182.6 76.6 B 21.0 J 13.0 168.2 71.0
LT-7 C 16.0 3.0 184.2 92.6 B 23.0 17.3 175.2 157.2
LT-8 C 28.3 4.7 185.7 50.9 B 16.3 4.7 148.4 33.8
LT-9 C 26.7 3.7 179.4 45.2 B 23.7 6.3 165.9 63.4
DTO-2 C 32.7 11.3 184.0 136.7 B 35.7 13.0 224.2 76.5
DTO-28 C 14.3 8.0 209.2 75.5 B 12.0 5.7 187.9 73.8
DTO-33 C 30.3 21.4 139.7 87.2 B 31.2 22.4 162.8 99.7
Kennebec C 16.7 10.3 175.0 120.6 B 15.7 16.3 197.0 150.6
R.Burbank C 14.7 7.3 154.6 81.2 B 12.3 10.3 162.8 78.8
Maine-47 C 14.7 7.7 148.2 76.5 B 10.7 10.7 138.9 72.7
Ddsirde C 14.0 19.7 116.5 94.8 B 20.7 18.0 165.4 94.2
NDD277-2 C 21.7 11.0 135.8 64.0 B 18.3 13.0 146.1 76.7
Y84-027 C 19.7 4.7 198.3 34.8 B 20.0 4.3 242.6 62.0
Values per plant are means of 3 replicates. LSD 05 values: Tuber number, 9.2; Tuber fresh weight, 51.9 g. Treatment (Trt): non-bacterized (C); bac- terized (B) plantlets. Heat acclimated and non-acclimated 4 week-old plants were potted for 12 weeks. * excluded from ANOVA (See text for explanation).
allow the plant to endure heat and water stress and eventu-
ally lead to greater yields. Indeed, our field and greenhouse
experiments confirmed better survival and performance of
bacterized plantlets compared to non-bacterized controls
when transplanted under water or heat stress (Nowak et al.
1995). Bacterization may also have practical implications for
t issue culture laboratories. Bacterized plant le ts no t only
grow faster but are sturdier, with better developed root mass
and produce more vigorous transplants (Nowak et al. 1995,
Nowak et al. 1997), capable of withstanding low fungal dis-
ease pressure significantly better than non-bacterized ones
(Nowak et al. 1997, Sharma and Nowak 1998). Moreover, the
bacterium establishes endophytic and epiphytic populations
in tissue culture plantlets (Frommel et al. 1991, Pillay and
Nowak 1997) and thus there is no need for re-inoculation if
the plantlets are multiplied by nodal explants.
The ABA-deficient mutants, 11401-01 and 9120-05, had
spindly appearance with small leaves both i n vitro and ex
vitro. They both benefitted from bacterization in vitro (Table
1). The bacterium induced shoot growth promotion in 11401-
01 at 20/15 C was so dramatic, that we think this clone has
potential for utilization in bioassays for plant growth pro-
moting ability of microorganisms. Despite such benefits i n
vitro, the transplants of the ABA-deficient mutants did not
benef i t f rom bacter izat ion ex v i t ro (Table 2). At 20/15 C
mutants produced small tubers with significantly lower f~esh
weight than the other clones, and at 33/25 C they both died 4
weeks after transplanting, independent ly of bacterization.
Plants of the ABA-deficient mutants are similar to the droopy
potato plants, which had high stomatal conductance due to
the very little ABA accumulation (Quarrie, 1982). This con-
firms the crucial role of abscissic acid in heat stress survival.
Our earlier study of the bacterization effect on i n vitro tuber-
ization responses in potato indica ted that the bac te r ium
induces synthesis of a tuberization factor, presumably ajas-
monate (a similar stimulation was achieved with jasmonic
acid) (Nowak et al. unpublished). The assumption was that
this compound may compensate for the ABA deficiency in
the mutants. The observed acceleration of tuberization (Fig.
3) and senescence in bacterized LT-7 plants (Bensalim 1997)
further supported this assumption.
Our data show that some clones originally selected for
heat stress tolerance, e.g. LT-1, LT-5, LT-6, LT-7, LT-8, LT-9,
DTO-2, and Y84-027, were more sensitive to high tempera-
ture than the temperate cultivars, Russet Burbank and Ken-
nebec (Table 2). The bacterization, which in greenhouse
experiments in fiats, enhanced stolon production and tuber
initiation (Dunbar 1997), under the heat stress significantly
increased tuber number in LT-7 (Table 2, Fig. 3). The non-
bacter ized LT-7 plants grown at 33/25 C produced m a n y
stolons but only a few tubers. On the contrary, stolon num-
ber in the bacterized LT-7 was reduced but the plants set
almost six times as many tubers. It is also interesting to note,
that in DTO-2 bacterization enhanced tuber fresh weight at
20/15 C and significantly reduced at 33/25 C (Table 2).

152 AMERICAN JOURNAL OF POTATO RESEARCH Vol. 75
The expe r imen t s ind ica te tha t ut i l izat ion o f benef ic ia l
bac ter ia has a great po ten t ia l in t i ssue cul ture p lan t le t pro-
duct ion as wel l as the i r adapta t ion to hea t stress. To utilize
the benefits of bac te r iza t ion to its full potent ial , one should
consider genotypic se lec t ions (both, plant and microbia l ) for
spec i f ic env i ronments . Benef ic ia l b a c t e r i a c o u l d be intro-
duced into t issue cul ture o f po ta to at the early mul t ipl icat ion
s tages and p ropaga ted th rough success ive genera t ions . Our
r ecen t data also ind ica te t ha t bac te r i a inhabi t ing p lan t le t s
can t r ans loca te to s e e d t u b e r s ( N o w a k and S turz u n p u b -
l ished) and potent ia l ly benef i t the n e x t genera t ion o f po ta to
plants.
ACKNOWLEDGMENT
The authors w o u l d l ike to thank Dr. Henry de J o n g f rom
the Agr icu l tu re and A g r i - F o o d C a n a d a R e s e a r c h Sta t ion,
Fredericton, New Bnmswick , for his kind supply o f the ABA-
deficient mutants and for the f requent d iscuss ions dur ing the
tenure of this project , and Dr. Tess Astatkie f rom the Depart-
men t of Mathemat ics and Physics, N o v a Scot ia Agricul tural
College, Truro, Nova Scotia, for his ass is tance wi th ~ c s .
LITERATURE CITED
Bensalim, S. 1997. Heat stress responses of in vitro bacterized (Pseudo- monas sp. strain PsJN) and non-bacterized clones of potato (Solanum tuberosum L). M.Sc. Thesis, Dalhousie Univ., Halifax, NS, Canada.
Brown, M.E. 1974. Seed and root bacterization. Ann Rev Phytopathol 12:181-197.
Conn, K.L, J. Nowak, and G. I.~azarovits. 1997. A gnotobiotic bioassay for studying interactions between potato and plant growth-pro- moimg rhizobacteri3. Can J Microbio143:801-808.
Dunbar, C. 1997. Utilization of seaweed extract and plant growth pro- mofing rhizobacterium in greenhouse production of potato mini- tubers. M.Sc. Thesis, Dalhousie Univ., Halifax, NS, Canada.
Ewing, E.E. 1995. The role of hormones in potato (Solanum tuberosum L) tuberizafion. In: "Plant hormones: Physiology, biochemistry and molecular biology." Edited by P.J. Davies. Kluwer Acad Publ, Dordrecht, NL, pp. 698-724.
Frommel, M.I., J. Nowak, and G. I~7~rovits. 1991. Growth enhancement and developmental modifications of in vitro grown potato (Solanum tuberosum spp. tuberosum) as affected by a non- fluorescent P s ~ sp. Plant Physio196:928-936.
Glick, B.I~ 1995. The enhancement of plant growth by free-living bacte- ri~ Can J Microbio141:109-117.
Herman, E.B. 1987. "Contaminants" promote potato micropropagation. Agricell Report 9:38.
Kloepper, J.W., R. Lffshitz, and M.N. Schroth. 1988. Pseudomonas inoc- ulants to benefit plant production. ISI Atlas of Science: Anim Plant Sci 1:60-64.
Lazarovits, G. and J. Nowak. 1997. Rhizobacteria for improvement of plant growth and establishment. HortScience 32:188-192.
Mendoza, H.A. and R.N. Estrada. 1979. Breeding potatoes for tolerance to stress: Heat and frost. In: "Stress Physiology in Crop Plants." Edited by I-L Mussel and R.C. Staples. John Wiley and Sons, Inc., N Y, pp. 227-262.
Menzel, C.M. 1985. Tuberization in potato at high temperatures: Inter- action between temperature and hTadiance. Ann Bot 55:35-39.
Midmore, DJ. 1992. Potato production in the tropics. In: "The potato". Edited by P. Han~s. Chapmann and Hall, London, UK, pp. 509- 569.
Nowak, J. and D. Colborne. 1989. In vitro tuberization and tuber pro- teins as indicators of heat stress tolerance in potato. Am Potato J 66:35-45.
Nowak, J. and S.I~ Asiedu. 1992. Gelling agent and light effects on in vitro tuberization of potato cultivars. Am Potato J 69:461-470.
Nowak, J., S.I~ Asiedu, G. Lazarovits, V. Pillay, A. Stewart, C.Smith, and V. Liu. 1995. Enhancement of in vitro growth and transplant stress tolerance of potato and vegetable plantlets co-cultured with a plant growth promoting pseudomonad bacterium. In: "Ecophysiology and photosynthetic in vitro cultures." Edited by F. Carre and P. Chagvardieff, CEA, Aix-en-Provence, France, pp. 173-180.
Nowak, J., S.K. Asiedu, S. Bensalim, J. Richards, A. Stewart, C. Smith, D. Stevens, and A.V. Sturz. 1997. From laboratory to applications: challenges and progress with in vitro dual cultures of potato and beneficial bacteria. In: "Bacteria and bacteria-like contaminants of plant tissue culture." Edited by A. Cassells. Kluwer Acad Publ, Dordrecht, NL (8 pages, in press).
Nowak, J. and D. Stevens. 1996. Root leakage and growth responses of potato cultivars to beneficial bacteria. Department of Plant Science Biennial Report, Nova Scotia Agricultural College, Truro, NS, Canada, pp. 80-82.
Pillay, V. and J. Nowak. 1997. Inoculum density, temperature and geno- type effects on epiphytic and endophytic colonization and in vitro growth promotion of tomato (Lycopersicon esculentum L) by a pseudomonad bacterium. Can J Microbio143:354-361.
Quarrie, S.A~ 1982. Droop~. a wilty mutant of potato deficient in abscisic acid. Plant Cell Environ 5:23-26
Shanna, V.K. and J. Nowak. 1998. Introduction of Verticillium wilt resis- tance in tomato transplants by in vitro co-culture of seedlings with a plant growth-promoiing rhizobacterium (Pseudomonas sp. strain PsJN). Can J Microbiol (in press).
Simmonds, N.W. 1965. Mutant expression in diploid potatoes. Heredity 20:65-72.
Sipos, J., J. Nowak, and G.Hicks. 1988. Effect of daminozide on survival, growth and yield of micropropagated potatoes. Am Potato J 65:353-364.
Trebejo, I. and D.J. Midmore. 1990. Effect of water stress on potato growth, yield and water use in a hot tropical climate. J Agric Sci 114:321-334.




















