
A LINE2 repetitive DNA sequence from the cichlid fish, Oreochromis niloticus: sequence analysis and...
12
Abstract. We report the cloning and characterization of a long interspersed nucleotide element (LINE) from a cichlid fish, Oreochromis niloticus, and show the distri- bution of this element, called CiLINE2 for cichlid LINE2, in the chromosomes of this species. The identifi- cation of an open reading frame in CiLINE2 with amino acid sequence similarity to reverse transcriptases encod- ed by LINE-like elements in Caenorhabditis elegans, Platemys spixii, Schistosoma mansoni, Gallus gallus (CRI), Drosophila melanogaster (I factor), and Homo sapiens (LINE2), as well as the structure of the element, suggest it is a member of this family of non-long ter- minal repeat-containing retrotransposons. Search of a DNA sequence database identified sequences similar to CiLINE2 in four other fish species (Haplotaxodon mi- crolepis, Oreochromis mossambicus, Pseudotropheus ze- bra, and Fugu rubripes). Southern blot hybridization ex- periments revealed the presence of sequences similar to CiLINE2 in all Tilapiini species analyzed from the gen- era Oreochromis, Tilapia, and Sarotherodon, and gave an estimated copy number of about 5500 for the haploid genome of O. niloticus. Fluorescent in situ hybridization showed that CiLINE2 sequences were organized in small clusters dispersed over all chromosomes of O. ni- loticus, with a higher concentration near chromosome ends. Furthermore, the long arm of chromosome 1 was strikingly enriched with this sequence. The distribution of LINE2-related elements might underlie the difference in chromosome banding patterns observed between cold- blooded vertebrates and mammals. Introduction Recent studies have shown that much of the dispersed moderately repeated DNA of eukaryotes consists of mo- bile elements, sequences that have moved from one ge- nomic location to another (Kazazian and Moran 1998). They can comprise a significant proportion of the ge- nome, for example, 36% of the human genome is esti- mated to be derived from these elements (Smit 1996). Mobile elements are classified according to their mode of transposition, either as DNA transposons that trans- pose directly through a ‘cut and paste’ mechanism or as retroelements that transpose through an RNA intermedi- ate (Charlesworth et al. 1994). Short interspersed nucle- otide elements (SINEs) and long interspersed nucleotide elements (LINEs) are two general types of retroelement that are distinguished from each other on the basis of their length and structure, and are found in eukaryotic genomes as diverse as plants and mammals (reviewed in Singer 1982; Martin 1991; Okada 1991; Charlesworth et al. 1994; Smit 1996; Okada et al. 1997). SINEs have arisen by reverse transcription of short stable RNAs, such as tRNAs and 7SL RNA, and have no protein-cod- ing capacity, whereas complete LINE elements poten- tially encode some or all of the enzymes, including re- verse transcriptase, necessary for retrotransposition (Martin 1991). Both LINEs and SINEs lack the long ter- minal repeats (LTRs) characteristic of the third major class of retroelements, the retroviral-like retrotranspo- sons, and instead end in a characteristic poly(A) or sim- ple AT-rich repeat tract (Martin 1991). Retrotransposition of retroviral-like retrotransposons involves tRNA-primed reverse transcription of the RNA of the element, with the DNA copy being inserted at a nonspecific target site (Boeke and Corces 1989). In con- trast, LINEs have been proposed to transpose by using a nick in the target DNA to prime reverse transcription of their RNA, with the copied element becoming directly integrated at the site of the nick (Luan and Eickbush 1995). This process, termed target primed reverse tran- scription (TPRT), has been demonstrated for a LINE- GenBank accession numbers: AF016498, AF016499, AF074485, AF074486, AF074487, AF074488 Permanent address: C. Oliveira, F. Porto-Foresti, Departamento de Morfologia, Instituto de Biociências, UNESP, Botucatu, São Paulo, Brazil Edited by: P.B. Moens Correspondence to: J.M. Wright e-mail: [email protected] Chromosoma (1999) 108:457–468 A LINE2 repetitive DNA sequence from the cichlid fish, Oreochromis niloticus: sequence analysis and chromosomal distribution Claudio Oliveira 1 , Joyce S.K. Chew 1,2 , Fábio Porto-Foresti 1 , Melanie J. Dobson 2 , Jonathan M. Wright 1 1 Department of Biology, Dalhousie University, Halifax, Nova Scotia, Canada B3H 4J1 2 Department of Biochemistry, Dalhousie University, Halifax, Nova Scotia, Canada B3H 4H7 Received: 4 March 1999; in revised form: 28 May 1999 / Accepted: 2 July 1999 © Springer-Verlag 1999
-
Upload
independent -
Category
Documents
-
view
8 -
download
0
Transcript of A LINE2 repetitive DNA sequence from the cichlid fish, Oreochromis niloticus: sequence analysis and...

Abstract. We report the cloning and characterization ofa long interspersed nucleotide element (LINE) from acichlid fish, Oreochromis niloticus, and show the distri-bution of this element, called CiLINE2 for cichlidLINE2, in the chromosomes of this species. The identifi-cation of an open reading frame in CiLINE2 with aminoacid sequence similarity to reverse transcriptases encod-ed by LINE-like elements in Caenorhabditis elegans,Platemys spixii, Schistosoma mansoni, Gallus gallus(CRI), Drosophila melanogaster (I factor), and Homosapiens (LINE2), as well as the structure of the element,suggest it is a member of this family of non-long ter-minal repeat-containing retrotransposons. Search of aDNA sequence database identified sequences similar toCiLINE2 in four other fish species (Haplotaxodon mi-crolepis, Oreochromis mossambicus, Pseudotropheus ze-bra, and Fugu rubripes). Southern blot hybridization ex-periments revealed the presence of sequences similar toCiLINE2 in all Tilapiini species analyzed from the gen-era Oreochromis, Tilapia, and Sarotherodon, and gavean estimated copy number of about 5500 for the haploidgenome of O. niloticus. Fluorescent in situ hybridizationshowed that CiLINE2 sequences were organized insmall clusters dispersed over all chromosomes of O. ni-loticus, with a higher concentration near chromosomeends. Furthermore, the long arm of chromosome 1 wasstrikingly enriched with this sequence. The distributionof LINE2-related elements might underlie the differencein chromosome banding patterns observed between cold-blooded vertebrates and mammals.
Introduction
Recent studies have shown that much of the dispersedmoderately repeated DNA of eukaryotes consists of mo-bile elements, sequences that have moved from one ge-nomic location to another (Kazazian and Moran 1998).They can comprise a significant proportion of the ge-nome, for example, 36% of the human genome is esti-mated to be derived from these elements (Smit 1996).Mobile elements are classified according to their modeof transposition, either as DNA transposons that trans-pose directly through a ‘cut and paste’ mechanism or asretroelements that transpose through an RNA intermedi-ate (Charlesworth et al. 1994). Short interspersed nucle-otide elements (SINEs) and long interspersed nucleotideelements (LINEs) are two general types of retroelementthat are distinguished from each other on the basis oftheir length and structure, and are found in eukaryoticgenomes as diverse as plants and mammals (reviewed inSinger 1982; Martin 1991; Okada 1991; Charlesworth etal. 1994; Smit 1996; Okada et al. 1997). SINEs havearisen by reverse transcription of short stable RNAs,such as tRNAs and 7SL RNA, and have no protein-cod-ing capacity, whereas complete LINE elements poten-tially encode some or all of the enzymes, including re-verse transcriptase, necessary for retrotransposition(Martin 1991). Both LINEs and SINEs lack the long ter-minal repeats (LTRs) characteristic of the third majorclass of retroelements, the retroviral-like retrotranspo-sons, and instead end in a characteristic poly(A) or sim-ple AT-rich repeat tract (Martin 1991).
Retrotransposition of retroviral-like retrotransposonsinvolves tRNA-primed reverse transcription of the RNAof the element, with the DNA copy being inserted at anonspecific target site (Boeke and Corces 1989). In con-trast, LINEs have been proposed to transpose by using anick in the target DNA to prime reverse transcription oftheir RNA, with the copied element becoming directlyintegrated at the site of the nick (Luan and Eickbush1995). This process, termed target primed reverse tran-scription (TPRT), has been demonstrated for a LINE-
GenBank accession numbers: AF016498, AF016499, AF074485,AF074486, AF074487, AF074488
Permanent address: C. Oliveira, F. Porto-Foresti, Departamento de Morfologia, Instituto de Biociências, UNESP, Botucatu, São Paulo, Brazil
Edited by: P.B. Moens
Correspondence to: J.M. Wrighte-mail: [email protected]
Chromosoma (1999) 108:457–468
A LINE2 repetitive DNA sequence from the cichlid fish, Oreochromis niloticus: sequence analysis and chromosomal distributionClaudio Oliveira1, Joyce S.K. Chew1,2, Fábio Porto-Foresti1, Melanie J. Dobson2, Jonathan M. Wright1
1 Department of Biology, Dalhousie University, Halifax, Nova Scotia, Canada B3H 4J12 Department of Biochemistry, Dalhousie University, Halifax, Nova Scotia, Canada B3H 4H7
Received: 4 March 1999; in revised form: 28 May 1999 / Accepted: 2 July 1999
© Springer-Verlag 1999

like element from insects (Luan et al. 1993). The best-studied LINE element, human L1, is 6–7 kb in length,comprises about 15% of the human genome, and con-tains two open reading frames (ORFs), one of which en-codes the endonuclease/reverse transcriptase responsiblefor nicking the target DNA and for making the DNAcopy of the LINE transcript (Martin 1991; Smit 1996;Kazazian and Moran 1998). More than 95% of L1 cop-ies are truncated at their 5’ end and about 10% of theseare internally rearranged, perhaps reflecting the lack ofprocessivity of the LINE-encoded reverse transcriptaseor some other general feature of TPRT-mediated retro-transposition (Kazazian and Moran 1998). A second,less abundant LINE family, LINE2 has also been identi-fied in the human genome (Smit 1996). No intact LINE2elements have been isolated but the element is character-ized by sharing 50 bp of its 3’ end with a tRNA-derivedSINE named MIR (mammalian-wide interspersed re-peat) found in all mammalian genomes, including thoseof marsupials and monotremes (Jurka et al. 1995; Smitand Riggs 1995; Smit 1996). It has been proposed thatthe LINE2-encoded reverse transcriptase, by recognizingthis sequence in the MIR transcript and promoting MIRretrotransposition by TPRT into sites at which it wouldotherwise copy LINE2 RNA, caused the amplification ofMIR elements prior to the radiation of mammals (Jurkaet al. 1995; Smit and Riggs 1995; Smit 1996; Okada etal. 1997). The discovery of other families of SINEs,each sharing a common 3’ end with a given LINE fami-ly, has led to the suggestion that all SINEs have arisenand spread owing to their mobilization by a partnerLINE (Okada et al. 1997).
Among fishes, fragments of LINE-like sequences,largely truncated, have been identified in rainbow trout(Winkfein et al. 1988), eel (Ohshima et al. 1996), Japa-nese medaka, desert pupfish (Duvernell and Turner1998), and zebrafish (Okada et al. 1997). In this study,we have identified a LINE2-like element from the cich-lid fish Oreochromis niloticus. Species from the cichlidtribe Tilapiini, commonly referred to as tilapia, are nota-ble for being members of one of the largest taxonomicgroups of fishes, and for their ecological diversity(Trewavas 1983). Tilapia represent the predominant fishfauna of Africa’s Great Lakes, but as a result of intro-ductions, they are now also distributed worldwide inboth tropical and subtropical regions (Trewavas 1983).Some Tilapinii, such as O. niloticus, are of particular im-portance to aquaculture (Pullin 1991).
Cytogenetic studies have shown that the haploid ge-nome of O. niloticus consists of 22 chromosomes, 20 ofwhich are small, similarly sized, and submedian or sub-telomeric, making chromosome assignment difficult(Kornfield et al. 1979; Majumdar and McAndrew 1986).Fluorescent in situ hybridization using cloned satelliteDNA sequences as probes and conventional chromo-some staining techniques have been used to investigatethe heterochromatin distribution on the chromosomes,which has allowed further resolution of the karyotype(Majumdar and McAndrew 1986; Oliveira and Wright1998). Recently, a linkage map covering all 22 chromo-somes has been constructed using polymorphic DNA
markers (Kocher et al. 1998). This genetic map will beinvaluable for mapping quantitative trait loci of impor-tance for aquacultural strain improvement. Current ef-forts are focused on the construction of a physical mapof the chromosomes of O. niloticus. As part of that ef-fort, we are examining the distribution of repetitive se-quences within the O. niloticus chromosomes. Differen-tial hybridization patterns obtained with repetitive DNAprobes (Oliveira and Wright 1998) may be useful inidentifying a particular chromosome from several othersimilarly sized chromosomes, such as those observed inthe O. niloticus karyotype (Majumdar and McAndrew1986). The identification of chromosomes on the basisof the DNA sequences they contain is an essential stepin integrating the physical and genetic maps for any ge-nome. For the cichlids, a molecular karyotype will pro-vide the basis for analyzing chromosomal changes thatmight underlie their recent rapid adaptive radiation(Greenwood 1991). Here we report the cloning, se-quence analysis and chromosomal distribution of aLINE2-like element in O. niloticus and demonstrate its presence in other cichlid genomes. This element, CiLINE2, is similar to LINE2 in mammals and sharesidentity with a LINE-like element recently reported fromanother cichlid species, Haplotaxodon microlepis (Teraiet al. 1998).
Materials and methods
Animals and isolation of genomic DNA
Fifteen adult O. niloticus (nine females and six males) were ob-tained from the tilapia culture facility at Dalhousie University(courtesy of Dr. Jim Wright). Blood samples of Oreochromis au-reus, Oreochromis mossambicus, Oreochromis placidus, Tilapiarendalli, Tilapia zillii, and Sarotherodon galileus were obtainedfrom Dr. B. J. McAndrew, Institute of Aquaculture at the Univer-sity of Stirling, UK. Genomic DNA was extracted from blood ac-cording to Wright (1989).
Cloning and sequencing of a LINE-like element
Two Sau3AI restriction fragments of 1977 bp and 1634 bp, eachencoding part of a LINE-like element, were derived from O. nilot-icus yeast artificial chromosome (YAC) genomic DNA clones(Chew 1997). The Sau3AI fragments were ligated to the vector,pTZ19R (Pharmacia), by standard methods (Sambrook et al.1989) to produce the recombinant plasmids, 6-2 and 12-5, respec-tively. Additional clones of putative LINE-like elements, hereafternamed CiLINE2 elements, were obtained from O. niloticus ge-nomic DNA by polymerase chain reaction (PCR) amplificationusing the primers A (5’-ATG GTG TTC CAC AGG GTT CA-3’)and B (5’-TAG GAT AAG ATG GGG CCT GA-3’) based on thesequence of the 6-2 CiLINE2 clone (nucleotides 221–240 and921–940, respectively) (Fig. 1). The PCRs were performed with5–20 ng of genomic DNA, 150 pmol of each primer, 200 µMdNTPs, and 2.5 U of Taq polymerase in the buffer provided by thesupplier (Pharmacia). The PCR conditions were as follows: initialdenaturation at 94°C for 3 min, 35 cycles of denaturation at 94°Cfor 30 s, annealing at 50°C for 1 min, extension at 72°C for 3 min,and then a final 5 min extension at 72°C. The PCR-amplifiedproducts were cloned in the plasmid pGEM-T Easy (Promega).Transformants of Escherichia coli strain JM109 were screened ac-cording to the procedure recommended by the supplier (Promega).Inserts from amplified products were prepared for sequencing us-
458

ing a Nucleobond AX kit (Vector BioSystems). The purifiedDNAs were primed by fluorescent tagged primers (LICOR) andsequenced with the T7 Sequenase 7-deaza-dGTP sequencing kit(Amersham). The reactions were separated on a LICOR 4200 automated sequencer. GenBank Accession numbers are: 6-2(AF016498), 12-5 (AF016499), and for the four cloned PCRamplicons: L3 (AF074486), L9 (AF074488), L30 (AF074487),L37 (AF074485).
Sequence analysis
Alignment of sequences was performed using CLUSTALW1.7(Thompson et al. 1994). Nucleic acid sequences were subjected toBLASTN and BLASTP (Altschul et al. 1990) searches at the Na-tional Center for Biotechnology Information (www2.ncbi.nlm.nih.gov). The sequences were also screened for repetitive se-quences using the RepeatMasker program (Smit and Green 1997)at the website of the University of Washington (ftp.genome.wash-ington.edu/cgi-bin/RepeatMasker). Nucleotide sequences weresurveyed for ORFs using the DNA sequence translation softwareat the website of the University of Minnesota (alces.med.umn.edu/cuse.html).
Genomic distribution and copy number of CiLINE2
The distribution of CiLINE2 elements in Tilapiini species was de-termined by electrophorectically separating in a 0.9% agarose gel10 µg aliquots of MboI-digested genomic DNA from each of thefollowing cichlid fish species: O. niloticus, O. aureus, O. moss-ambicus, O. placidus, T. rendalli, T. zillii, and S. galileus. To de-termine whether CiLINE2 sequences could be detected in otherspecies, DNA from representative orders of other fishes and mam-mals was also subjected to Southern blot hybridization analysis(Southern 1975). These DNAs were: Oncorhynchus mykiss (rain-bow trout); Salmo salar (Atlantic salmon); Melanogrammus aeg-lefinus (haddock); Gadus morhua (Atlantic cod); Lophius ameri-canus (goosefish); Urophycis tenuis (whitehake); Pollachius vir-ens (pollock); Oryctolagus cuniculus (rabbit); Canis familiaris(domestic dog); Phoca vitulina concolour (harbor seal); Physetermacrocephalus (sperm whale). Fractionated DNA was transferredto Hybond N membranes (Amersham) by vacuum blotting usingstandard methodologies (Sambrook et al. 1989). Hybridizationprobes were prepared from the insert DNA in plasmid 6-2 andfrom L3 (one of the cloned PCR amplicons described above that only contains 852 bp from within the putative reverse tran-scriptase coding region of CiLINE2) by radiolabeling with [α-32P]-dCTP (3000 Ci/mmol, ICN) using a random priming kitfrom Boehringer Mannheim. Hybridization was conducted in 10 ml of hybridization solution [6×SSPE (1 M NaCl, 60 mMNaH2PO4.H2O, 60 mM EDTA), 5×Denhardt’s (0.1% Ficoll Type400, 0.1% polyvinylpyrrolidone, 0.1% BSA), 1.0% SDS, and 100 µg/ml yeast RNA] for 16 h at 65°C. After hybridization, themembrane was washed twice in 2×SSC, 0.1% SDS at 65°C for 30 min. (1×SSC is 0.15 M NaCl, 0.015 M sodium citrate.) Thenumber of copies of the CiLINE2 element in the O. niloticus ge-nome was estimated by screening a genomic library cloned inbacteriophage EMBL-3 (Harris and Wright 1995) using the L3 in-sert as a hybridization probe (Sambrook et al. 1989).
Chromosome techniques
Mitotic chromosomes were prepared as described by Foresti et al.(1993). Fluorescent in situ hybridization (FISH) analyses wereperformed with plasmid L3. The whole plasmid was labeled withbiotin-14-dATP using a commercially available nick translationkit (Life Technologies). For in situ hybridization, slides were in-cubated in 2×SSC and 100 µg/ml RNase at 37°C for 1 h and dehy-drated in an ethanol series (75%, 80%, 95%). Chromosomal DNAwas denatured by immersing slides for 2 min at 70°C in 70%formamide, 2×SSC, and then dehydrated in an alcohol series(75%, 80%, 95%). The hybridization mixture was composed of
50% formamide in 2×SSC, 10% dextran sulfate, 500 µg/ml carrierRNA, and 1.5 µg/ml biotinylated DNA probe. After 10 min dena-turation at 75°C, 15 µl of hybridization mixture was applied toeach slide under a sealed coverslip. Hybridization was performedovernight in a moist chamber at 37°C. Slides were then washedfor 30 min in formamide (50%) at 42°C, twice for 10 min in2×SSC (42°C), and twice for 10 min in 4×SSC (42°C). Hybridiza-tion was detected with fluorescein-labeled avidin (Oncor) and thesignal was enhanced by incubation with anti-avidin antibody (On-cor) followed by the application of fluoresceinated avidin. Chro-mosomes were counterstained with propidium iodide/antifade so-lution (Oncor).
Results
Identification of a dispersed O. niloticusrepeated DNA sequence
Two O. niloticus genomic Sau3AI fragments, each froma different YAC clone, were subcloned (plasmids 6-2and 12-5), and shown to contain a repetitive DNA se-quence based on intense hybridization to radiolabeled O. niloticus genomic DNA in a Southern blot analysis.Preliminary hybridization studies using the Sau3AI frag-ment from plasmid 6-2 as a probe indicated that the re-peated sequence was not present in the yeast genome,but was present on at least three independently isolatedO. niloticus YAC clones (data not shown).
Nucleotide sequence of CiLINE2 elements
Sequencing of the O. niloticus clones, 6-2 and 12-5, re-vealed a short overlap of nearly identical sequence heldin common between the two Sau3AI fragments (Fig. 1).A BLASTN search of GenBank (Altschul et al. 1990)with the complete nucleotide sequence of clone 6-2 re-vealed significant matches [P(N) <10–10] to several arc-hived Oreochromis DNA sequences, including a regiondownstream of the growth hormone genes in O. niloticus(M97765–66, M84774) and O. mossambicus (Y11732)and a region upstream of the prolactin I gene in O. moss-ambicus (X92380), confirming the presence of a repeti-tive sequence in the 6-2 clone shared with, but extendingbeyond the region of overlap with, the 12-5 clone. The6-2 repeat also shares approximately 95% sequenceidentity with a LINE-like element recently reported from another cichlid species, H. microlepis (Terai et al.1998). This Haplotaxodon element has been designatedCiLINE2 (Terai et al. 1998) owing to its similarity to hu-man LINE2 elements (Smit 1996). Analysis by the RepeatMasker program of the insert sequence of the O. niloticus 6-2 clone identified a 157 bp sequence atposition 196–352 and a 131 bp sequence at position817–947, as well as a 135 bp sequence in clone 12-5 thatshowed sequence identity to mammalian LINE2 ele-ments (Fig. 1) (Smit and Riggs 1995; Smit 1996).
Lower significance matches were obtained with the6-2 CiLINE2 repeat to a region downstream of a(CTAT)19 microsatellite-containing region in the cichlidPseudotropheus zebra (U14397), and to sequences fromsalamander (D17442), and snake (D31777). A BLASTN
459

460

GenBank search with the sequence of clone 12-5 thatcorresponded to the CiLINE2 element also revealed asignificant match with sequences upstream of the growthhormone gene of pufferfish, Fugu rubripes (U63807).
The most striking feature of the 6-2 sequence was thepresence of an 885 bp ORF. A BLASTP search of Gen-Bank (Altschul et al. 1990) using a conceptual transla-tion of this ORF gave significant matches to putative re-verse transcriptases encoded by LINE-like elementsfrom species as diverse as Caenorhabditis elegans (nem-atode), Platemys spixii (turtle), Schistosoma mansoni(bloodfluke), Gallus gallus (chicken), Drosophila mel-anogaster, and Homo sapiens (data not shown), confirm-ing the classification of the 6-2 repeat as a LINE.
To determine a consensus sequence for O. niloticusCiLINE2, other copies were cloned by PCR using prim-ers based on the sequence of two regions within the 6-2 ORF that correspond to conserved domains in the di-verse reverse transcriptases identified in the BLASTPsearch. Agarose gel electrophoresis revealed two majorPCR products of about 750 and 900 bp (data not shown).The smaller product was approximately the same size asthe 719 bp product expected for the 6-2 CiLINE2 clone.All products from the PCR were included in the ligationto the cloning vector. The insert sizes of 42 clones(L1–L42) obtained from this pooled ligation were deter-mined by PCR amplification and ranged in size from200 to 1200 bp.
Alignment of the DNA sequences of genomic clones6-2 and 12-5, and four of the PCR clones, L3, L9, L30,and L37, (Fig. 1) showed that the PCR clones were verysimilar to each other and to the genomic sequencethroughout most of their length with occasional inser-tions/deletions and nucleotide substitutions. The greatersize of two of the longer PCR products, L37 and L9, isaccounted for by duplicated blocks of sequence at their5’ ends, reflecting either a rearrangement during thecloning step or amplification of a rearranged CiLINE2.Unexpectedly, all four PCR clones ended at the same po-sition, 132 bp downstream of the expected 6-2 targetsite. The sequence of the 12-5 genomic clone revealed anucleotide stretch sharing 9 bp in common with the tar-get site that must have been sufficient to allow annealingof the primer at this position.
The aligned DNA sequences from the genomic andPCR clones were used to derive a consensus for the O. niloticus CiLINE2 ORF (Fig. 1). Neither of the genom-ic sequences cloned in 6-2 or 12-5 contained a completecopy of the CiLINE2 element. Sequences beyond thoseshown for 12-5 show no similarity to CiLINE2 elements.In contrast, the 3’ end of the CiLINE2 ORF is deleted in 6-2, but sequences corresponding to the 3’ untranslated re-gion of the element are present in this clone (data notshown). The amino acid sequence of this putative reversetranscriptase (Fig. 1) was then aligned with reverse tran-scriptase sequences encoded by other representative LINE-like elements (Fig. 2). The alignment revealed that the O.niloticus CiLINE2 sequence corresponded to conserved re-verse transcriptase domains 3–7 found in all non-LTRretrotransposon reverse transcriptases (Xiong and Eick-bush 1990) and was most similar to the reverse transcrip-tase encoded by the LINE2 elements in mammals and aLINE-like element from the zebrafish, Danio rerio (Okadaet al. 1997). These three putative reverse transcriptasesalso share several stretches of conserved or identical se-quence downstream of the region containing the domainsthat are usually conserved between different reverse tran-scriptases (Xiong and Eickbush 1990). The Sau3AI clon-ing site used in generating the O. niloticus 6-2 clone trun-cates the CiLINE2 reverse transcriptase ORF such that theregion encoding the amino-terminal portion containing do-mains 1 and 2 was not present on this clone. It is, there-fore, unclear how much further 5’ the reverse transcriptaseORF extends in this species.
Genomic distribution of CiLINE2
The presence of CiLINE2-related sequences on morethan one, independently isolated, genomic DNA clone ofO. niloticus suggested that the CiLINE2 element was arelatively abundant repeated sequence in this species.Moreover, the identification of sequences similar to CiLINE2 in various species, revealed by databasesearches, indicated that this sequence might be presentin the genomes of other cichlids. Southern blot analysisof genomic DNA from O. niloticus and six other Tilapiini species revealed that the 6-2 sequence hybrid-ized to numerous MboI restriction fragments (Fig. 3).These fragments ranged in size from 0.3 kb to more than4.0 kb and were detected in all seven cichlid species, in-dicating that CiLINE2-related sequences are present inthe genomes of all species sampled from the genera Ore-ochromis, Tilapia, and Sarotherodon. Although bands ofsimilar size hybridized to the 6-2 probe in all genomes,the relative intensity of the predominant bands differedbetween genera. In addition, other bands not shared incommon between different species were observed. Hy-bridization analysis of these same genomic DNAs toplasmid L3 produced identical results to those obtainedwith the 6-2 probe. No related sequences were detectedby Southern blot analysis in genomic DNA of other representative vertebrate species, teleost fishes or mam-mals under the same hybridization conditions (data notshown, see Materials and methods).
461
Fig. 1. Alignment of nucleotide sequences corresponding to thereverse transcriptase-encoding region of six CiLINE2 elementsfrom Oreochromis niloticus. Dots indicate sequence identity anddashes indicate no sequence for that clone or gaps that were intro-duced to maximize sequence alignment. Termination codons aredenoted by an asterisk (*). Arrows above the sequence show theextent of imperfect direct repeats found only in the polymerasechain reaction (PCR) clones L37 and L9. Text highlighted in boldshows sequence similarity to LINE2 elements in mammals (Smit1996). Numbers 1–984 are the 6-2 nucleotide sequence(AF016498), while 985–1165 are the complement of nucleotides937–652 from clone 12-5 (AF016499). The complete sequencesfor 6-2 and 12-5 are available from GenBank. Only the regionscorresponding to the CiLINE2 open reading frame (ORF) areshown here. The amino acid sequence of the putative reverse tran-scriptase (RT) derived from the CiLINE2 consensus ORF isshown

462

Copy number of CiLINE2
The copy number of CiLINE2 elements and their trun-cated derivatives in the O. niloticus genome was esti-mated by screening an EMBL-3 bacteriophage library ofO. niloticus genomic DNA (Harris and Wright 1995)with the radiolabeled L3 insert. Of 10,000 plaques (0.15genome equivalents), 809 hybridized to the L3 probe.With a genome size of approximately 109 bp and an av-erage insert size of 15 kb for each bacteriophage clone,and assuming a single copy per clone (Harris and Wright1995; Bryden et al. 1998), we conservatively estimatethat the CiLINE2 element is reiterated about 5500 timesper haploid genome.
Chromosomal distribution of CiLINE2 elements
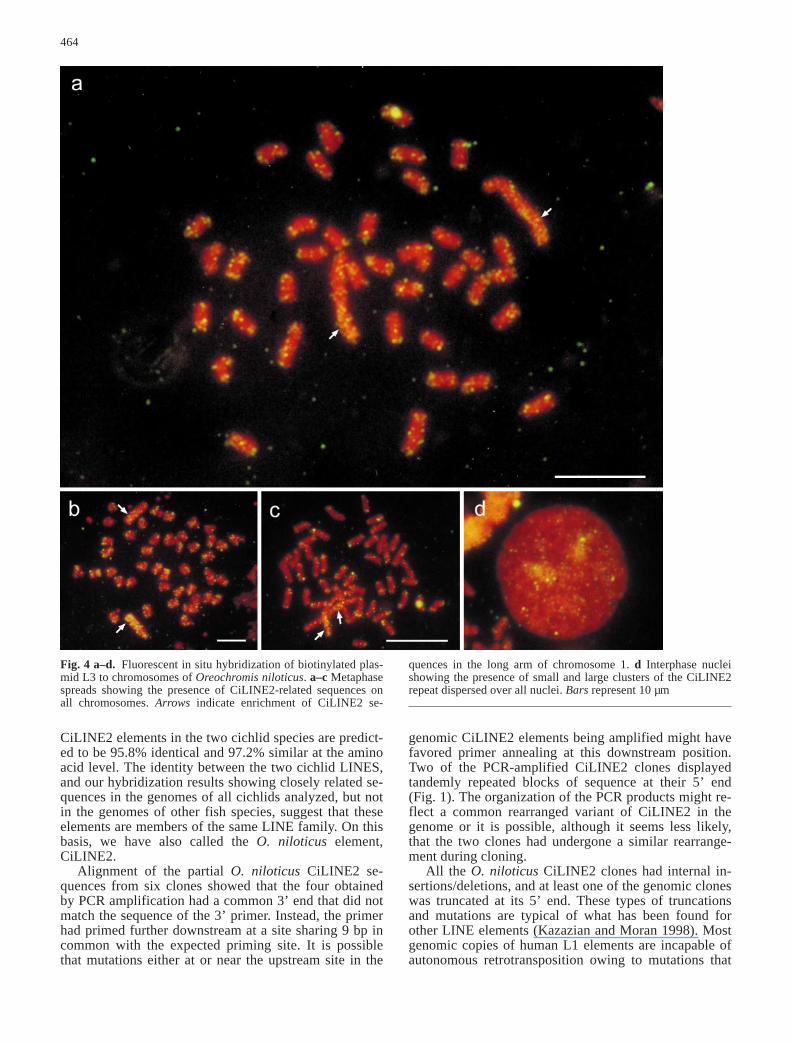
The chromosomal distribution of the CiLINE2 elementwas determined by FISH of plasmid L3 to metaphasespreads of O. niloticus chromosomes (Fig. 4). Hybrid-ization signals were observed as small clusters dispersedthroughout all chromosomes, with an enrichment nearmost chromosomal termini (Fig. 4a–c). Moreover, astriking concentration of CiLINE2 elements was seendistributed along the terminal two-thirds of the long armof chromosome 1, the only chromosome in O. niloticusthat can be readily distinguished from the others, on the basis of its much greater length (Majumdar andMcAndrew 1986). This higher concentration of CiLINE2 elements made possible the identification ofthis region of chromosome 1 in interphase nuclei (Fig.4d).
Discussion
Database searches with a dispersed repetitive sequencefrom the cichlid fish O. niloticus revealed sequence sim-ilarity to LINE-like elements from a range of other spe-cies. LINEs were originally identified in mammaliangenomes, with the L1 element being the most abundantmember of a widespread monophyletic group, andLINE2 representing a second, less abundant family(Smit 1996). The O. niloticus LINE DNA sequence dis-played identity with human LINE2 elements (Smit1996) and with a LINE-like element, named CiLINE2,discovered in another cichlid, H. microlepis (Terai et al. 1998). The reverse transcriptases encoded by the
463
Fig. 2. Alignment of the amino acid sequence encoded by theOreochromis niloticus CiLINE2 consensus ORF (CiLINE2) withreverse transcriptase sequences from other representative LINE-like elements. Termination codons are denoted by an asterisk (*),while dashes indicate gaps inserted to improve the alignment.Identical amino acids are highlighted in black, while conservativechanges are shaded in gray, if shared by at least half of the taxa.The numbers correspond to the amino acid residues for reversetranscriptases from Drosophila I factor (Ml4954), Caenorhabditiselegans (P34620), Platemys spixii (AB005891), Gallus gallus(L22146), Schistosoma mansoni (U66331), and the deduced ami-no acid sequences encoded by a Danio rerio LINE-like sequence(AB004653) and LINE2 (nucleotides 1338–2750 in RepeatMask-er database). The conserved reverse transcriptase domains definedby Xiong and Eickbush (1990) are denoted by brackets with con-served residues (RT cons.) indicated by their single letter aminoacid designation, or h hydrophobic, c charged, and p polar resi-dues
Fig. 3a,b Southern blot hybridization of ge-nomic DNA from fish of the tribe Tilapiini toclone 6-2. a Photograph of an ethidium bro-mide-stained 0.9% agarose gel on which MboI-digested DNA from the following specieswas separated by electrophoresis: (1) Oreochro-mis aureus, (2) Oreochromis mossambicus, (3)Tilapia rendalli, (4) Sarotherodon galileus, (5)Tilapia zillii, (6) Oreochromis niloticus, and (7)Oreochromis placidus. b Autoradiogram ofSouthern blot of the gel in a after hybridizationto radiolabeled DNA from plasmid 6-2. Sizes ofmolecular weight markers (in kb) are shown onthe left
CiLINE2 elements in the two cichlid species are predict-ed to be 95.8% identical and 97.2% similar at the aminoacid level. The identity between the two cichlid LINES,and our hybridization results showing closely related se-quences in the genomes of all cichlids analyzed, but notin the genomes of other fish species, suggest that theseelements are members of the same LINE family. On thisbasis, we have also called the O. niloticus element, CiLINE2.
Alignment of the partial O. niloticus CiLINE2 se-quences from six clones showed that the four obtainedby PCR amplification had a common 3’ end that did notmatch the sequence of the 3’ primer. Instead, the primerhad primed further downstream at a site sharing 9 bp incommon with the expected priming site. It is possiblethat mutations either at or near the upstream site in the
genomic CiLINE2 elements being amplified might havefavored primer annealing at this downstream position.Two of the PCR-amplified CiLINE2 clones displayedtandemly repeated blocks of sequence at their 5’ end(Fig. 1). The organization of the PCR products might re-flect a common rearranged variant of CiLINE2 in thegenome or it is possible, although it seems less likely,that the two clones had undergone a similar rearrange-ment during cloning.
All the O. niloticus CiLINE2 clones had internal in-sertions/deletions, and at least one of the genomic cloneswas truncated at its 5’ end. These types of truncationsand mutations are typical of what has been found forother LINE elements (Kazazian and Moran 1998). Mostgenomic copies of human L1 elements are incapable ofautonomous retrotransposition owing to mutations that
464
Fig. 4 a–d. Fluorescent in situ hybridization of biotinylated plas-mid L3 to chromosomes of Oreochromis niloticus. a–c Metaphasespreads showing the presence of CiLINE2-related sequences onall chromosomes. Arrows indicate enrichment of CiLINE2 se-
quences in the long arm of chromosome 1. d Interphase nucleishowing the presence of small and large clusters of the CiLINE2repeat dispersed over all nuclei. Bars represent 10 µm

465
disrupt their ORFs, both of which are required for L1retrotransposition in vivo (Moran et al. 1996). Similarly,to date, no intact human LINE2 elements have beenidentified. The DNA sequence of the longest O. niloticusgenomic CiLINE2 clone, 6-2, predicted the presence ofa stop codon (Fig. 1) within the portion of the elementthat would encode a domain (motif 3) conserved in allknown reverse transcriptases and RNA-directed RNApolymerases (Xiong and Eickbush 1990). Also, clone 6-2 contained an internal deletion, nucleotide position984, that removed the 3’ end of the reverse transcriptaseORF (Fig. 1), indicating that this copy of CiLINE2would not be capable of autonomous retrotransposition.Furthermore, since Sau3AI was used as the cloning en-zyme, it is unclear whether sequences 5’ to the Sau3AIsite in this genomic copy of the element extended to in-clude the endonuclease-encoding domain and the up-stream ORF that might be expected for a full-length ele-ment (Malik and Eickbush 1998). Nonetheless, the O. niloticus CiLINE2 6-2 clone contains 516 bp more 5’sequence than any of the copies of the element identifiedin Haplotaxodon (Terai et al. 1998), and has confirmedthat the majority of the reverse transcriptase-encodingdomain is present in this cichlid element.
The most convincing evidence that CiLINE2 belongsto a class of retrotransposable elements is the presenceof an ORF in CiLINE2 displaying significant amino acidsequence similarity to reverse transcriptases and RNA-dependent DNA polymerases (Fig. 2). The partial se-quence for the O. niloticus CiLINE2 reverse transcrip-tase showed the greatest similarity to the reverse tran-scriptases predicted from the human LINE2 element(Smit 1996) and from LINE-like elements in zebrafish(Fig. 2) as well as salamander and snake (data notshown). In addition to the conserved reverse transcrip-tase domains 3–7 found in all non-LTR retrotransposonreverse transcriptases (Xiong and Eickbush 1990), thesefour reverse transcriptase sequences share other blocksof sequence in common. For example, they contain aLDYCN motif downstream of reverse transcriptase do-main 7 instead of the PHLEY motif shared by the LINE-like elements from P. spixii, G. gallus and S. mansoni(Fig. 2). These similarities support the grouping of theO. niloticus CiLINE2 element as a member of theLINE2 clade of non-LTR retrotransposons.
Southern blot hybridization analysis detected numer-ous restriction fragments of O. niloticus genomic DNAthat hybridized to the cloned CiLINE2 sequence. Thisresult indicates that CiLINE2 is a highly repetitive ele-ment predominantly dispersed in the genome, a findingthat is consistent with the genomic organization of most,if not all, LINEs studied in other organisms. CiLINE2elements were detected by hybridization in the genomicDNA of all Tilapiini species tested from the genera Oreochromis, Tilapia, and Sarotherodon (Fig. 3). It is in-teresting to note that DNA from Oreochromis and Sarot-herodon species produced a hybridization pattern dis-tinct from that of the genomic DNA from Tilapia spe-cies. Thus, if this pattern is verified by Southern blotanalysis of genomic DNA for other tilapiine species, theCiLINE2 probe could be used to distinguish fish from
the genus Tilapia from those of Oreochromis and Sarot-herodon.
GenBank searches and the study by Terai et al.(1998) detected the presence of CiLINE2 elements inspecies from the Haplochromini, Tylochromini and Perissodini cichlid tribes of East Africa. Given that EastAfrican cichlids have recently undergone a rapid adap-tive radiation (Sturmbauer and Meyer 1993), it is notsurprising that the CiLINE2 elements from O. niloticusand H. microlepis are virtually identical at the nucleotidelevel as the sequences would not have had sufficienttime to accumulate many mutations. The presence of CiLINE2 elements in New World, Indian, Madagascan,and African cichlids (Terai et al. 1998) suggests that theCiLINE2 element must have been present in an ancestralcichlid species before the breakup of Gondwanaland 180million years ago (MYA).
No hybridization of the partial CiLINE2 sequencefrom clone L3 was seen to the genomic DNA of otherrepresentative teleost fishes and mammals despite theevidence from GenBank searches that closely relatedDNA sequences should exist in the genomes of mam-mals and other vertebrates. The hybridization conditionsused in the Southern blot analysis reported here wereprobably too stringent to allow detection of sequencesdisplaying a significant degree of mismatch with theprobe. If the ancestral LINE2 element was present in thelast common ancestor of all vertebrates, it would not besurprising if most existing LINE2 elements had now di-verged sufficiently from this sequence to be undetectableby hybridization. It is also possible that the copy numberof LINE2-related elements in some species might be toolow to be detected, even if the sequences have retainedsufficient sequence identity.
The estimate of about 5500 copies of CiLINE2 in thegenome of O. niloticus, as determined by the number ofgenomic DNA fragments in a bacteriophage λ librarythat hybridized to a CiLINE2 probe, contrasts with themuch greater abundance of 593,000 copies of L1 and271,000 copies of LINE2 estimated for the human ge-nome (Smit 1996). A relatively low abundance ofCiLINE2 in the tilapia genome compared with the abun-dance of LINEs in mammalian genomes would be con-sistent with the general observation that the number ofSINEs and LINEs in fish is much less than that observedin mammals (Okada et al. 1997). However, it might alsoreflect the inability of the CiLINE2 probe to detect cop-ies of the elements that have undergone either significantmutations or rearrangements.
Fluorescent in situ hybridization showed that CiLINE2 sequences are organized in O. niloticus chro-mosomes as small clusters, somewhat punctate, and dis-persed over all chromosomes, but with higher concentra-tions being found in the termini of most chromosomes.Most striking was a large concentration of CiLINE2 ele-ments throughout the long arm of chromosome 1, theonly chromosome in this species that can be readily dis-tinguished on the basis of its size. If CiLINE2 elementsrepresent a class of telomere-associated repeat, the exis-tence of an elevated number of copies of CiLINE2 inchromosome 1 is consistent with the long arm having

originated from the fusion of three smaller chromosomesand, therefore, retaining the remnants of the telomere-as-sociated regions at internal positions within the newchromosome (Chew, unpublished results).
The dispersed pattern of organization observed forthe CiLINE2 element contrasts with the pattern observedfor L1 in several mammalian species where the elementsare distributed in large clusters corresponding to the AT-rich, late-replicating Giemsa-positive (G) bands or thehuman Alu SINE family, which is found almost exclu-sively in regions corresponding to the GC-rich, earlyreplicating, Giemsa-light (R) chromosome bands (Manuelidis and Ward 1984; Korenberg and Rykowski1988; Chen and Manuelidis 1989; Boyle et al. 1990;Baker and Kass 1994). LINE2 elements are sevenfoldless abundant than L1 elements, comprising approxi-mately 2% of the human genome, and although their dis-tribution on the chromosomes has not been reported, ananalysis of LINE2 sequences found in database entrieshas revealed that they are embedded in genomic se-quences with a much less biased GC content than thosecontaining either L1 or Alu elements (Smit 1996). Fur-thermore, the MIR family of SINEs, which share a 3’end with LINE2 elements, also show this distributionwith respect to the GC content of their surrounding ge-nomic sequences (Smit 1996).
Differences in the distribution of LINEs have previ-ously been reported for other species. For example,among insects, two LINE elements identified as R1 andR2, are integrated only within the 28S rDNA transcrip-tion units (Jakubczak et al. 1991). In situ hybridizationstudies with LINE-like elements in the plant Hordeumvulgare (Moore et al. 1991) and in the silkmoth, Bombyxmori, (Ogura et al. 1994) revealed a punctate pattern oflabeling dispersed along all chromosomes, similar to thepattern observed for CiLINE2 in O. niloticus.
The almost mutually exclusive distribution of the AT-rich (58%) L1 elements and GC-rich (56%) AluSINEs has been suggested as the molecular basis for theobserved banding of human chromosomes with fluores-cent dyes (Korenberg and Rykowski 1988). Cytogeneticstudies have shown that fish chromosomes exhibit G andR bands, although these are much less pronounced thanthose seen for mammalian chromosomes (Delany andBloom 1984; Almeida-Toledo et al. 1988; Gold et al.1990; Gold and Li 1991; Bertollo et al. 1997), but thesebands are usually not detected by base-specific fluoro-chromes such as 4′,6-diamidino-2-phenylindole (an AT-specific fluorochrome) (Gold et al. 1990; Gold andLi 1991). The absence of base-specific fluorescent bandsin fish chromosomes has been attributed to their lack ofGC-rich isochores and to the more homogeneous DNAbase composition found in fish genomes relative tomammalian genomes (Bernardi and Bernardi 1990;Bernardi 1995).
If LINE2-related elements with their less-biased dis-tribution are the major family of LINEs in fish genomes,rather than L1-related elements, this could explain thepoor banding observed for chromosomes from O. nilot-icus. Similarly, if SINEs have been mobilized by a part-ner LINE (Okada et al. 1997) of the LINE2 family, these
should also show a less biased distribution in fish chro-mosomes. Two SINE-like elements that have been iden-tified in O. niloticus are less abundant than mammalianSINEs and also display a punctate pattern of staining onall chromosomes, with the exception of a large cluster ofthe SINE ROn-1 in an interstitial region of chromosome1 (C. Oliveira, unpublished results). The relatively smallnumber of LINE and SINE elements, and perhaps somefeature of the particular families of elements that haveamplified in the cichlid genomes, may underlie the poorchromosome banding. This hypothesis may also be validfor other cold-blooded vertebrates, so further analysis ofthe structure and distribution of other LINE and SINEelements in chromosomes of these species might welllead to a better understanding of the role of these ele-ments in the development of the AT- and GC-rich chro-mosome bands.
The presence of intense CiLINE2 hybridization sig-nals near the telomeres of all O. niloticus chromosomesresembles the distribution of a LINE-like element foundin Chlorella chromosomes (Higashiyama et al. 1997).Considering the pattern of distribution observed forthese elements, Higashiyama et al. (1997) suggested thatthe concentration of LINEs near the chromosome endsmay play a role in stabilizing the telomere, or that the el-ements transposed into subtelomeric regions and thenaccumulated, resulting in a structural buffer that protectsthe internal gene-rich region. In Drosophila, two non-LTR retrotransposons, HeT-A and TART, are only foundin tandem arrays at chromosome ends where they main-tain and protect the telomere in a manner analogous tothe simple TG-rich telomeric repeats found on chromo-some ends of other eukaryotic organisms (Pardue et al.1997). The simple telomeric repeats are synthesized bytelomerase, a ribonucleoprotein enzyme, which is a spe-cialized reverse transcriptase using its RNA componentas a template to direct synthesis of the species-specifictelomeric repeats directly onto the chromosome ends(Blackburn 1992). This mode of action is very similar tothe TPRT mechanism proposed for the retrotranspositionof LINE elements (Luan et al. 1993), the only significantdifference being that the telomerase does not have tonick the target DNA prior to using it to prime reversetranscription of its RNA. Pardue et al. (1997) suggestedthat the Het-A and TART elements are evolutionarily re-lated to telomerase and, in this context, it is interestingthat we have found another member of a non-LTR retro-transposon family, CiLINE2, that appears by FISH to beenriched near chromosome ends. The chromosomes ofO. niloticus terminate with tandem arrays of the ver-tebrate-specific (TTAGGG)n telomeric repeat (Chew1997), so further analysis will be needed to determinethe arrangement of the CiLINE2 sequences and theirproximity to the chromosome end.
LINE2-like elements have now been found in all ver-tebrates from fish to mammals (Jurka et al. 1995; Smit1996; Okada et al. 1997; Terai et al. 1998; this study).This finding suggests that the functional LINE2 elementwas present in a common ancestor over 400 MYA be-fore the divergence of bony fishes from other verte-brates. It is also during this time period that the verte-
466

467
brate telomeric repeat sequence (TTAGGG)n is believedto have been fixed in its function and location at theends of vertebrate chromosomes (Meyne et al. 1989).The concentration of CiLINE2 elements near the telo-meres makes it tempting to speculate that the LINE2 el-ements could have been fixed in telomere-associated re-gions at the same time as the fixation of the vertebratetelomere.
Acknowledgements. C.O. was supported by a fellowship fromFundação de Amparo à Pesquisa do Estado de São Paulo. F.P.F.was supported by a UNESCO Biotechnology Fellowship. J.S.K.C.was supported by a postgraduate scholarship from the NationalSciences and Engineering Research Council (NSERC) of Canada.This work was supported by grants from NSERC of Canada toJ.M.W. and from the Canadian Genome Analysis and TechnologyProgram to M.J.D. (GO-12404).
Note added in proofA LINE element, Maui, has recently been reported in Fugurubripes (Poulter et al. 1999, Gene 227:169–179). An alignmentof the putative reverse transcriptase with that of the CiLINE2element reported here, revealed that they share 79.8% similarity atthe amino acid level. This finding supports the inclusion of theFugu LINE in the LINE2 family.
References
Almeida-Toledo LF, Viegas-Pequignot E, Foresti F, Toledo SA,Dutrillaux B (1988) BrdU replication patterns demonstratingchromosome homeologies in two fish species, genusEigenmannia. Cytogenet Cell Genet 48:117–120
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Ba-sic local alignment search tool. J Mol Biol 215:403–410
Baker RJ, Kass DH (1994) Comparison of chromosomal distribu-tion of a retroposon (LINE) and a retrovirus-like element mysin Peromyscus maniculatus and P. leucopus. Chromosome Res2:185–189
Bernardi G (1995) The human genome: organization and evolu-tionary history. Annu Rev Genet 29:445–476
Bernardi G, Bernardi G (1990) Compositional patterns in the nu-clear genome of cold-blooded vertebrates. J Mol Evol 31:265–281
Bertollo LAC, Fontes MS, Fennocchio AS, Cano J (1997) TheX1X2Y sex chromosome system in the fish Hoplias malabar-icus. I. G-, C- and chromosome replication banding. Chromo-some Res 5:493–499
Blackburn EH (1992) Telomerases. Annu Rev Biochem 61:113–129
Boeke JD, Corces VG (1989) Transcription and reverse transcrip-tion of retrotransposons. Annu Rev Microbiol 43:403–434
Boyle AL, Ballard SG, Ward DC (1990) Differential distributionof long and short interspersed element sequences in the mousegenome: chromosome karyotyping by fluorescence in situ hy-bridization. Proc Natl Acad Sci U S A 87:7757–7761
Bryden L, Denovan-Wright EM, Wright JM (1998) ROn-1SINES: a tRNA-derived, short interspersed repetitive DNA el-ement from Oreochromis niloticus and its species-specific dis-tribution in Old World cichlid fishes. Mol Mar Biol Biot-echnol 7:48–54
Charlesworth B, Sniegowski P, Stephan W (1994) The evolution-ary dynamics of repetitive DNA in eukaryotes. Nature371:215–220
Chen TL, Manuelidis L (1989) SINEs and LINEs cluster in dis-tinct DNA fragments of Giemsa band size. Chromosoma 98:309–316
Chew JSK (1997) Isolation and characterization of Oreochromisniloticus DNA yeast artificial chromosomes clones. MSc The-sis, Dalhousie University, Canada
Delany ME, Bloom SE (1984) Replication banding patterns in thechromosomes of the rainbow trout. J Hered 75:431–434
Duvernell DD, Turner BJ (1998) Swimmer 1, a new LINE familyin teleost genomes with sequence similarity by mammalianL1. Mol Biol Evol 15:1791–1793
Foresti F, Oliveira C, Almeida-Toledo LF (1993) A method forchromosome preparations from large specimens of fishes us-ing in vitro short treatment with colchicine. Experientia 49:810–813
Gold JR, Li YC (1991) Trypsin G-banding of North American cy-prinid chromosomes: phylogenetic considerations, implica-tions for fish chromosome structure, and chromosome poly-morphism. Cytologia 56:199–208
Gold JR, Li CY, Shipley NS, Powers PK (1990) Improved meth-ods for working with fish chromosomes with a review ofmetaphase chromosome banding. J Fish Biol 37:563–575
Greenwood PH (1991) Speciation. In: Keenleyside MHA (ed)Cichlid fishes: behaviour, ecology and evolution. Chapmanand Hall, London, pp 86–102
Harris AS, Wright JM (1995) Nucleotide sequence and genomic or-ganization of cichlid fish minisatellites. Genome 38:177–184
Higashiyama T, Noutoshi Y, Fujie M, Yamada T (1997) Zepp, aLINE-like retrotransposon accumulated in the Chlorella telo-meric region. EMBO J 16:3715–3723
Jakubczak JL, Burke WD, Eickbush TH (1991) Retrotransposableelements R1 and R2 interrupt the rRNA genes of most insects.Proc Natl Acad Sci U S A 88:3295–3299
Jurka J, Zietkiewicz E, Labuda D (1995) Ubiquitous mammalian-wide interspersed repeats (MIRS) are molecular fossils fromthe mesozoic era. Nucleic Acids Res 23:170–175
Kazazian HH Jr, Moran JV (1998) The impact of L1 retrotranspo-sons on the human genome. Nat Genet 19:19–24
Kocher TD, Lee W-J, Sobolewska H, Penman D, McAndrew B(1998) A genetic linkage map of a cichlid fish, the tilapia(Oreochromis niloticus). Genetics 148:1225–1232
Korenberg JR, Rykowski MC (1988) Human genome organiza-tion: Alu, Lines, and the molecular structure of metaphasechromosome bands. Cell 53:391–400
Kornfield IL, Ritte U, Richler C, Wahrman J (1979) Biochemicaland cytological differentiation among cichlid fishes of the Seaof Galilee. Evolution 33:1–14
Luan DD, Eickbush TH (1995) RNA template requirements fortarget DNA-primed reverse transcription by the R2 retrotrans-posable element. Mol Cell Biol 15:3882–3891
Luan DD, Korman MH, Jakubczak JL, Eickbush TH (1993) Re-verse transcription of RM2Bm RNA is primed by a nick at thechromosomal target site: a mechanism for nonLTR retrotrans-position. Cell 72:595–605
Majumdar KC, McAndrew BJ (1986) Relative DNA content ofsomatic nuclei and chromosomal studies in three genera, Tila-pia, Sarotherodon, and Oreochromis of the tribe Tilapiini (Pisces, Cichlidae). Genetica 68:175–188
Malik HS, Eickbush TH (1998) The RTE class of non-LTR retr-otransposons is widely distributed in animals and is the originof many SINEc. Mol Biol Evol 15:1123–1134
Manuelidis L, Ward DC (1984) Chromosomal and nuclear distri-bution of the HindIII 1.9-kb human DNA repeat segment.Chromosoma 91:28–38
Martin SL (1991) LINEs. Curr Opin Genet Dev 1:505–508Meyne J, Ratliff RL, Moyzis RK (1989) Conservation of the hu-
man telomere sequence (TTAGGG)n among vertebrates. ProcNatl Acad Sci U S A 86:7049–7053
Moore G, Cheung W, Schwarzacher T, Flavell R (1991) BIS 1, amajor component of the cereal genome and a tool for studyinggenomic organization. Genomics 10:469–476
Moran JV, Holmes SE, Kazazian HH Jr (1996) High frequencyretrotransposition in cultured mammalian cells. Cell 87:917–927

Smit AFA, Riggs AD (1995) MIRs are classic, tRNA-derivedSINEs that amplified before the mammalian radiation. Nucle-ic Acids Res 23:98–102
Southern EM (1975) Detection of specific sequences among DNAfragments separated by gel electrophoresis. J Mol Biol 98:503–517
Sturmbauer C, Meyer A (1993) Mitochondrial phylogeny of theendemic mouthbrooding lineages of cichlid fishes from LakeTanganyika in Eastern Africa. Mol Biol Evol 10:751–768
Terai Y, Takahashi K, Okada N (1998) SINE cousins: the 3’endtails of the two oldest and distantly related families of SINEsare descended from the 3’ ends of LINEs with the same gene-alogical origin. Mol Biol Evol 15:1460–1471
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: im-proving the sensitivity of progressive multiple sequence align-ment through sequence weighting, positions-specific gap penal-ties and weight matrix choice. Nucleic Acids Res 22:4673–4680
Trewavas E (1983) Tilapiine fishes of the genera Sarotherodon,Oreochromis and Danakilia. British Museum (Natural Histo-ry), London
Winkfein RJ, Moir RD, Krawetz AS, Blanco J, States JC, DixonGH (1988) A new family of repetitive, retroposon-like se-quences in the genome of the rainbow trout. Eur J Biochem176:255–264
Wright JM (1989) Nucleotide sequence, genomic organization andevolution of a major repetitive DNA family in tilapia (Ore-ochromis mossambicuslhomorum). Nucleic Acids Res 17:5071–5079
Xiong Y, Eickbush TH (1990) Origin and evolution of retroele-ments based upon their reverse transcriptase sequences.EMBO J 9:3353–3362
468
Ogura T, Okano K, Tsuchida K, Miyajima N, Tanaka H, TakadaN, Izumi S, Tomino S, Maekawa H (1994) A defective non-LTR retrotransposon is dispersed throughout the genome ofthe silk worm, Bombyx mori. Chromosoma 103:311–323
Ohshima K, Hamada M, Terai Y, Okada N (1996) The 3’ end ofrRNA-derived short interspersed repetitive elements are de-rived from the 3’ ends of long interspersed repetitive ele-ments. Mol Cell Biol 16:3756–3764
Okada N (1991) SINEs. Curr Opin Genet Dev 1:498–504Okada N, Hamada M, Ogiwara I, Ohshima K (1997) SINEs and
LINEs share common 3’ sequences: a review. Gene 205:229–243
Oliveira C, Wright JM (1998) Molecular cytogenetic analysis ofheterochromatin in the chromosomes of tilapia, Oreochromisniloticus (Teleostei: Cichlidae). Chromosome Res 6:205–211
Pardue ML, Danilevskaya ON, Traverse KL, Lowenhaupt K(1997) Evolutionary links between telomeres and transposableelements. Genetica 100:73–84
Pullin RSV (1991) Cichlids in aquaculture. In: Keenleyside MHA(ed) Cichlid fishes: behaviour, ecology and evolution. Chap-man and Hall, London, pp 280–309
Sambrook J, Maniatis T, Fritsch EF (1989) Molecular cloning. Alaboratory manual, 2nd edn. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, New York
Singer MF (1982) SINEs and LINEs: highly repeated short andlong interspersed sequences in mammalian genomes. Cell28:433–434
Smit AFA (1996) The origin of interspersed repeats in the humangenome. Curr Opin Genet Dev 6:743–748
Smit AFA, Green P (1997) RepeatMasker. Unpublished data.http://ftp.genome.washington.edu/RM/RepeatMaster.html




















