
Biodegradation kinetics of phenol and catechol using Pseudomonas putida MTCC 1194

www.fems-microbiology.org
FEMS Microbiology Letters 237 (2004) 355–361
A 19F NMR study of fluorobenzoate biodegradation bySphingomonas sp. HB-1
F.G. Hidde Boersma a, W. Colin McRoberts b, Steven L. Cobb c, Cormac D. Murphy a,*
a Department of Industrial Microbiology, University College Dublin, Belfield, Dublin 4, Irelandb Department of Agriculture and Rural Development for Northern Ireland, Newforge Lane, Belfast BT9 5PX, UK
c School of Chemistry, University of St. Andrews, Fife KY16 9ST, UK
Received 27 April 2004; received in revised form 27 June 2004; accepted 30 June 2004
First published online 14 July 2004
Abstract
While several microorganisms readily degrade 2- and 4-fluorobenzoates, only a very small number appear to catabolise the 3-
fluoro isomer, owing to the accumulation of toxic intermediates. Here we describe the isolation of a bacterium capable of using
3-fluorobenzoate as a sole source of carbon and energy, and the experiments conducted to define the steps involved in the biodeg-
radation of this compound. The organism was identified as a strain belonging to the genus Sphingomonas by sequence analysis of its
16S rRNA gene. To date no other organism from this genus is known to degrade this compound. Using fluorine nuclear magnetic
resonance spectroscopy (19F NMR) to analyse the culture supernatant it was possible to observe the disappearance of 3-fluor-
obenzoate and the appearance of fluoride ion and four other fluorinated compounds. These were identified as 3-fluorocatechol,
2-fluoromuconic acid and 3- and 5-fluoro-1,2-dihydro-1,2-dihydroxybenzoates. Thus, the likely catabolic pathway involves dioxy-
genation of 3-fluorobenzoate yielding fluorocatechol and subsequent intra-diol cleavage to yield fluoromuconic acid. The organism
can also use 2- and 4-fluorobenzoates as growth substrates.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Dioxygenation; Fluorobenzoate; 19F NMR
1. Introduction
The widespread use of organofluorine compounds by
the agrochemical, pharmaceutical and plastic industries
has resulted in these compounds becoming significant
environmental contaminants [1]. However, while much
is known about the microbial degradation of chlori-
nated organic compounds, our understanding of the
biodegradation of organofluorine compounds is poor.Progress in this area has improved with the emergence
0378-1097/$22.00 � 2004 Federation of European Microbiological Societies
doi:10.1016/j.femsle.2004.06.052
* Corresponding author. Tel.: +353-1-716-1311; fax: +353-1-716-
1183.
E-mail address: [email protected] (C.D. Murphy).
of 19F NMR as an analytical tool for studying the me-tabolism of fluorinated compounds [2,3].
The catabolism of fluorobenzoates has been studied
in severalmicroorganisms andbacteria have been isolated
that are capable of growing on 2- and 4-fluorobenzoate as
a sole carbon and energy source. 4-Fluorobenzoate typi-
cally undergoes dioxygenation to yield 4-fluorocatechol
followed by intra-diol cleavage forming 3-fluoro-cis,cis-
muconic acid. Cycloisomerization yields 4-fluoromuc-onolactone (4-carboxymethyl-4-fluorobut-2-en-4-olide)
and fluoride ion is eliminated forming maleylacetic acid,
which is channelled into the TCA cycle via oxoadipate
[4,5]. An alternative pathway has been found in anAureo-
bacterium sp., whereby the fluoride is enzymatically
removed in the initial step of degradation to yield
. Published by Elsevier B.V. All rights reserved.

356 F.G. Hidde Boersma et al. / FEMS Microbiology Letters 237 (2004) 355–361
4-hydroxybenzoate [6]. Dioxygenation of 2-fluorobenzo-
ate can result in either the spontaneous elimination of
fluoride forming catechol, or the formation of 3-fluoro-
catechol, which is a very poor substrate for catechol 1,2-
dioxygenases and can accumulate in, and poison, cells.
Nevertheless, there are some examples of bacteria usingthis compound as a growth substrate [7,8].
The co-metabolism of 3-fluorobenzoate has been re-
ported [9,10], but this compound appears to be a less ame-
nable substrate for microbial growth than the 2- or
4-fluoro isomers. A partially characterised strain (Agro-
bacterium-Rhizobium branch) isolated from a Nepalese
soil [11] was subjected to metabolic investigations and it
was concluded that the ability of amicroorganism to growon 3-fluorobenzoate was dependent upon the regioselec-
tivity of the initial dioxygenation reaction; however, only
fluoride ion and 2-fluoromuconic acid were identified in
the culture fluid in this study. Here we report the isolation
and characterisation of a bacterium capable of growing
on all three isomers of fluorobenzoate and the identifica-
tion of several catabolic intermediates by 19F NMR, al-
lowing elucidation of the degradative pathways.
2. Materials and methods
2.1. Chemicals
3-Fluorobenzoate was obtained from Fluka, 2- and
4-fluorobenzoate were purchased from Fluorochem(Derbyshire, UK) and 3-fluorcatechol was acquired
from Acros Organics (NJ, USA). Fluoro-substituted
3,5-cyclohexadiene-1,2-diol-1-carboxylic acids (dihydro-
dihydroxybenzoates) were obtained from Alcaligenes
eutrophus B9 [12], which was cultured on a fructose
mineral medium containing 3-fluorobenzoate (1 mM).
The A. eutrophus strain was a gift from Andrew Myers
(Department of Chemistry and Chemical Biology, Har-vard University, USA).
2.2. Isolation and characterisation of a 3-fluorobenzoate
degrading bacterium
Soil from Co. Wexford, Ireland that had been previ-
ously treated with a fluorinated herbicide (Punch C) was
used to inoculate E2 mineral medium [13] containing 5mM 3-fluorobenzoate as the sole source of carbon. After
14 days incubation at 25 �C in 250 ml Erlenmeyer flasks
on an orbital shaker, the fluoride ion concentration was
measured using an ion-selective electrode (Orion Model
96.06) using the method of Cooke [14]. Agar plates con-
taining E2 medium and 3-fluorobenzoate were inoculat-
ed with 0.1 ml of the suspension and one of the colonies
(strain HB-1) was purified by streaking on the same agarmedium. Purity was assessed on nutrient agar plates and
by light microscopy (Gram staining).
The 16S rRNA gene was amplified by PCR with
Taq polymerase (Promega) as recommended by the
manufacturer using the oligonucleotides F27 (5 0-AGA-
GTTTGATCMTGGCTCAG-3 0) and R1492 (5 0-TAC-
GGYTACCTTGTTACGACTT-3 0). The reaction
mixture was incubated at 94 �C for 2 min and was sub-sequently subjected to 25 cycles of 94 �C for 45 s, 52 �Cfor 45 s, 74 �C for 2 min, followed by incubation at 74
�C for 7 min.
Dideoxy sequencing reactions were done with the
CEQ DCTS Kit as described by the manufacturer
(Beckman) using oligonucleotides complementary to
conserved regions in the 16S rRNA gene. The nucleotide
sequence was determined using a Beckman CEQ 2000automatic sequencer; nucleotide sequence data were
compiled using the Staden package [15].
2.3. Identification of intermediates
The isolated bacterium was grown in Erlenmeyer
flasks containing E2 medium (50 ml) supplemented with
fluorobenzoate (2.5 or 5 mM) incubated on an orbitalshaker at 25 �C. Portions of the culture were removed
and centrifuged at various intervals and centrifuged,
and the supernatants stored frozen.
The supernatants were analysed by 19F nuclear mag-
netic resonance spectroscopy (19F NMR) using a Bruker
Avance 500 MHz spectrometer. Supernatants (0.5 ml)
were transferred to NMR tubes and D2O (0.2 ml) was
added to provide a lock signal. Identification of com-pounds present in the supernatants was also determined
by gas chromatography–mass spectrometry (GC–MS)
using a Hewlett–Packard 6890 gas chromatograph
linked to a 5973 mass selective detector controlled by
Chemstation software. Culture supernatants (200 ll or1 ml) were freeze-dried and either resuspended in chloro-
form or derivatised by adding N-methyl-N-(trimethylsi-
lyl) trifluoroacetamide (MSTFA, 200 ll) and heating for1 h at 100 �C. Samples in chloroform (1 ll) were injectedin the splitless mode onto a Hewlett–Packard Ultra 1
column (12 m·0.2 mm·0.33 lm) and the oven temper-
ature held at 50 �C for 1 min then raised to 280 �C at 10
�Cmin�1. The derivatised samples (1 ll) were injected
onto a Chrompak CP-Sil 19 CB column (25 m·0.25mm·0.2 lm) employing a split ratio of 10:1 and the
oven temperature held at 120 �C for 1 min then raisedto 270 �C at 10 �Cmin�1.
3. Results
3.1. Isolation and characterisation of strain HB-1
Degradation of 3-fluorobenzoate in the soil enrich-ment cultures was determined by measuring the concen-
tration of free fluoride ion. Cultures containing elevated
F.G. Hidde Boersma et al. / FEMS Microbiology Letters 237 (2004) 355–361 357
concentrations of fluoride (>70% defluorination) were
visibly darker than those in which no free fluoride was
detected, probably as a consequence of fluorosubstituted
catechol accumulation. Upon further culturing on agar
supplemented with 3-fluorobenzoate one colony type
was predominant, which was subsequently isolated. Mi-croscopic analysis determined this bacterium to be a
small Gram negative rod. Sequence comparison of the
16S rRNA gene with sequences in the GenBank data-
base revealed that the organism was a sphingomonad,
which was very closely related to Sphingomonas yan-
oikuyae (100% sequence identity with 918 nucleotides
of 16S rRNA gene). The sequence has been deposited
in GenBank under the Accession No. AY387397.
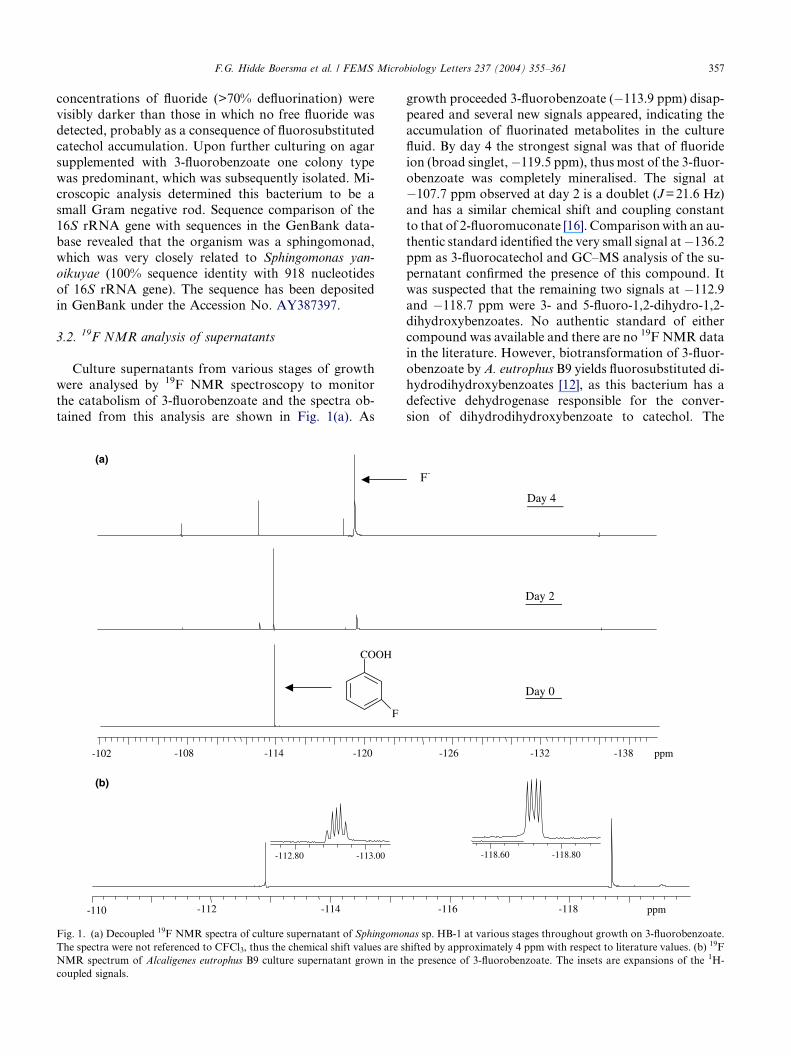
3.2. 19F NMR analysis of supernatants
Culture supernatants from various stages of growth
were analysed by 19F NMR spectroscopy to monitor
the catabolism of 3-fluorobenzoate and the spectra ob-
tained from this analysis are shown in Fig. 1(a). As
-113.00 -112.80
-114 -112 -110
-120-114-108-102
COOH
F
(a)
(b)
Fig. 1. (a) Decoupled 19F NMR spectra of culture supernatant of Sphingomo
The spectra were not referenced to CFCl3, thus the chemical shift values are s
NMR spectrum of Alcaligenes eutrophus B9 culture supernatant grown in t
coupled signals.
growth proceeded 3-fluorobenzoate (�113.9 ppm) disap-
peared and several new signals appeared, indicating the
accumulation of fluorinated metabolites in the culture
fluid. By day 4 the strongest signal was that of fluoride
ion (broad singlet,�119.5 ppm), thus most of the 3-fluor-
obenzoate was completely mineralised. The signal at�107.7 ppm observed at day 2 is a doublet (J=21.6 Hz)
and has a similar chemical shift and coupling constant
to that of 2-fluoromuconate [16]. Comparisonwith an au-
thentic standard identified the very small signal at�136.2
ppm as 3-fluorocatechol and GC–MS analysis of the su-
pernatant confirmed the presence of this compound. It
was suspected that the remaining two signals at �112.9
and �118.7 ppm were 3- and 5-fluoro-1,2-dihydro-1,2-dihydroxybenzoates. No authentic standard of either
compound was available and there are no 19F NMR data
in the literature. However, biotransformation of 3-fluor-
obenzoate byA. eutrophus B9 yields fluorosubstituted di-
hydrodihydroxybenzoates [12], as this bacterium has a
defective dehydrogenase responsible for the conver-
sion of dihydrodihydroxybenzoate to catechol. The
-118.80-118.60
ppm-118 -116
-138 -132-126 ppm
F-
Day 0
Day 2
Day 4
nas sp. HB-1 at various stages throughout growth on 3-fluorobenzoate.
hifted by approximately 4 ppm with respect to literature values. (b) 19F
he presence of 3-fluorobenzoate. The insets are expansions of the 1H-

-140-130-120-110-100-90-80 ppm
Day 2
Day 6
COOH
F
COOHCOOH
F
Fig. 2. Decoupled 19F NMR spectrum of culture supernatant of Sphingomonas sp. HB-1 grown on 2-fluorobenzoate. The metabolite at �107.7 ppm
is a doublet (J=21.6 Hz).
358 F.G. Hidde Boersma et al. / FEMS Microbiology Letters 237 (2004) 355–361
19F NMR spectrum of the culture supernatant from A.
eutrophus B9 grown in the presence of 3-fluorobenzoate
is shown in Fig. 1(b). It could be demonstrated by analy-
sing a mixture of the culture supernatants from both or-ganisms that the unidentified signals present in the
Sphingomonas sp. supernatant had identical chemical
shifts to the signals observed in the A. eutrophus superna-
tant. The coupling constants of both signals are very sim-
ilar indicating coupling of fluorine to hydrogen atoms on
the adjacent carbon (3JF–H=12 Hz) and to hydrogen at-
oms that are two carbons removed (4JF–H=6 Hz), but
the splitting patterns are different (Fig. 1(b)). The signalat �112.9 ppm (doublet of doublets) is probably that of
3-fluorodihydrodihydroxybenzoate, where the fluorine
is coupled to the hydrogen at C4 yielding a doublet, which
is further split by coupling to the hydrogen at either C5
or C2. The signal at �118.7 ppm is thus 5-fluor-
odihydrodihydroxybenzoate with fluorine coupling to
equivalent hydrogen atoms at C4 and C6 yielding a trip-
let, and to the hydrogen at C3, which further splits the sig-nal. Moreover, GC–MS analysis of culture supernatants
from Sphingomonas sp. HB-1 revealed two compounds
with almost identical mass spectra and molecular ions
that corresponded to the calculated mass of silylated flu-
orosubstituted dihydrodihydroxybenzoates; compounds
with the same retention times and mass spectra were de-
tected in A. eutrophus culture supernatant.
When E2 medium containing 2- or 4-fluorobenzoateas sole carbon and energy sources was inoculated with
Sphingomonas sp. HB-1 (grown on 3-fluorobenzoate),
growth was visible after 4–6 days incubation. Analysis
of the supernatants revealed that 2-fluoromuconate
and fluoride accumulated in the 2-fluorobenzoate-grown
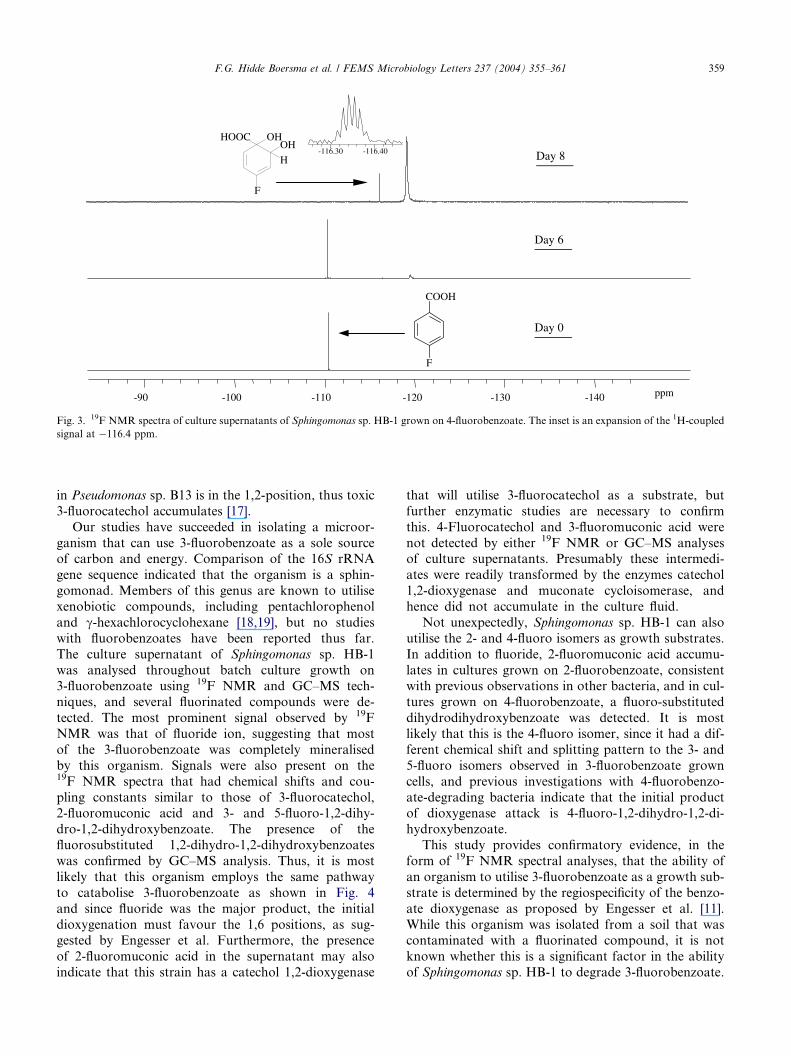
culture (Fig. 2). Fluoride accumulated in the 4-fluor-
obenzoate-grown culture along with a small amount of
another compound at �116.4 ppm (Fig. 3), which wasnot readily identifiable from data in the literature. The
splitting pattern and coupling constants indicated that
it might be 4-fluorodihydrodihydroxybenzoate: fluorine
coupling to equivalent hydrogens at C3 and C5 yielding
a triplet (J=11.9 Hz) and longer range coupling to the
hydrogen at C6 or C2 resulting in further splitting(J=6 Hz). GC–MS analysis of the derivatised superna-
tant detected the presence of a fluorosubstituted
dihydrodihydroxybenzoate. Interestingly, no 2- and 3-
fluorobenzoate remained in the supernatants after 4
days incubation, but cultures grown on 4-fluorobenzo-
ate took much longer (8 days) to fully catabolise the
substrate. This was consistent with the appearance of
turbidity in the culture flasks.
4. Discussion
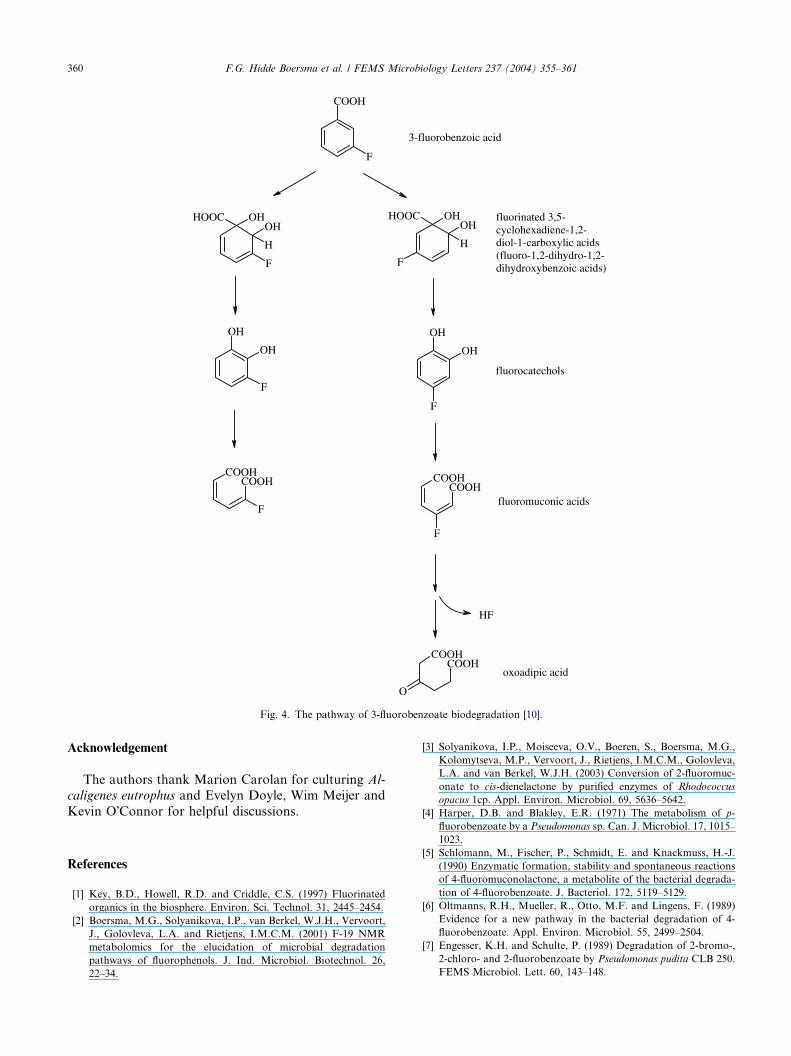
A catabolic pathway for the degradation of 3-fluor-
obenzoate by Pseudomonas sp. B13 was proposed by
Schreiber et al. [10] and is shown in Fig. 4. When grown
on 3-chlorobenzoate this strain could co-metabolise 3-fluorobenzoate; 2-fluoromuconic and 3- and 4-fluoro-
catechol were detected in the culture medium. However,
the organism could not use 3-fluorobenzoate as a sole
carbon and energy source. Engesser et al. [11] described
the isolation of a bacterial strain, FLB 300, which could
use 3-fluorobenzoate as a sole source of carbon and en-
ergy. Fluoride and 2-fluoromuconic acid were detected
in the culture fluid, and it was suggested that the samepathway as that proposed by Schreiber et al. (Fig. 4)
was responsible for the catabolism of the 3-fluorobenzo-
ate in this strain. However, it was also concluded that on
the basis of the ratio of fluoride and 2-fluoromuconate,
the predominant pathway was via initial 1,6-dioxygen-
ation, yielding 4-fluorocatechol. It was further suggested
that the regioselectivity of the initial dioxygenation was
the critical step in determining whether or not an organ-ism could use 3-fluorobenzoate as a sole source of car-
bon and energy, for example, the initial dioxygenation
-140 -130 -120 -110 -100 -90 ppm
Day 0
Day 6
Day 8
OHOH
H
HOOC
F
COOH
F
-116.40 -116.30
Fig. 3. 19F NMR spectra of culture supernatants of Sphingomonas sp. HB-1 grown on 4-fluorobenzoate. The inset is an expansion of the 1H-coupled
signal at �116.4 ppm.
F.G. Hidde Boersma et al. / FEMS Microbiology Letters 237 (2004) 355–361 359
in Pseudomonas sp. B13 is in the 1,2-position, thus toxic
3-fluorocatechol accumulates [17].
Our studies have succeeded in isolating a microor-
ganism that can use 3-fluorobenzoate as a sole sourceof carbon and energy. Comparison of the 16S rRNA
gene sequence indicated that the organism is a sphin-
gomonad. Members of this genus are known to utilise
xenobiotic compounds, including pentachlorophenol
and c-hexachlorocyclohexane [18,19], but no studies
with fluorobenzoates have been reported thus far.
The culture supernatant of Sphingomonas sp. HB-1
was analysed throughout batch culture growth on3-fluorobenzoate using 19F NMR and GC–MS tech-
niques, and several fluorinated compounds were de-
tected. The most prominent signal observed by 19F
NMR was that of fluoride ion, suggesting that most
of the 3-fluorobenzoate was completely mineralised
by this organism. Signals were also present on the19F NMR spectra that had chemical shifts and cou-
pling constants similar to those of 3-fluorocatechol,2-fluoromuconic acid and 3- and 5-fluoro-1,2-dihy-
dro-1,2-dihydroxybenzoate. The presence of the
fluorosubstituted 1,2-dihydro-1,2-dihydroxybenzoates
was confirmed by GC–MS analysis. Thus, it is most
likely that this organism employs the same pathway
to catabolise 3-fluorobenzoate as shown in Fig. 4
and since fluoride was the major product, the initial
dioxygenation must favour the 1,6 positions, as sug-gested by Engesser et al. Furthermore, the presence
of 2-fluoromuconic acid in the supernatant may also
indicate that this strain has a catechol 1,2-dioxygenase
that will utilise 3-fluorocatechol as a substrate, but
further enzymatic studies are necessary to confirm
this. 4-Fluorocatechol and 3-fluoromuconic acid were
not detected by either 19F NMR or GC–MS analysesof culture supernatants. Presumably these intermedi-
ates were readily transformed by the enzymes catechol
1,2-dioxygenase and muconate cycloisomerase, and
hence did not accumulate in the culture fluid.
Not unexpectedly, Sphingomonas sp. HB-1 can also
utilise the 2- and 4-fluoro isomers as growth substrates.
In addition to fluoride, 2-fluoromuconic acid accumu-
lates in cultures grown on 2-fluorobenzoate, consistentwith previous observations in other bacteria, and in cul-
tures grown on 4-fluorobenzoate, a fluoro-substituted
dihydrodihydroxybenzoate was detected. It is most
likely that this is the 4-fluoro isomer, since it had a dif-
ferent chemical shift and splitting pattern to the 3- and
5-fluoro isomers observed in 3-fluorobenzoate grown
cells, and previous investigations with 4-fluorobenzo-
ate-degrading bacteria indicate that the initial productof dioxygenase attack is 4-fluoro-1,2-dihydro-1,2-di-
hydroxybenzoate.
This study provides confirmatory evidence, in the
form of 19F NMR spectral analyses, that the ability of
an organism to utilise 3-fluorobenzoate as a growth sub-
strate is determined by the regiospecificity of the benzo-
ate dioxygenase as proposed by Engesser et al. [11].
While this organism was isolated from a soil that wascontaminated with a fluorinated compound, it is not
known whether this is a significant factor in the ability
of Sphingomonas sp. HB-1 to degrade 3-fluorobenzoate.
COOH
F
F
OHOH
H
HOOC OHOH
H
F
HOOC
OH
OH
F
OH
F
OH
COOHCOOH
F
COOHCOOH
F
COOHCOOH
O
HF
3-fluorobenzoic acid
fluorinated 3,5-cyclohexadiene-1,2-diol-1-carboxylic acids(fluoro-1,2-dihydro-1,2-dihydroxybenzoic acids)
fluorocatechols
fluoromuconic acids
oxoadipic acid
Fig. 4. The pathway of 3-fluorobenzoate biodegradation [10].
360 F.G. Hidde Boersma et al. / FEMS Microbiology Letters 237 (2004) 355–361
Acknowledgement
The authors thank Marion Carolan for culturing Al-
caligenes eutrophus and Evelyn Doyle, Wim Meijer and
Kevin O�Connor for helpful discussions.
References
[1] Key, B.D., Howell, R.D. and Criddle, C.S. (1997) Fluorinated
organics in the biosphere. Environ. Sci. Technol. 31, 2445–2454.
[2] Boersma, M.G., Solyanikova, I.P., van Berkel, W.J.H., Vervoort,
J., Golovleva, L.A. and Rietjens, I.M.C.M. (2001) F-19 NMR
metabolomics for the elucidation of microbial degradation
pathways of fluorophenols. J. Ind. Microbiol. Biotechnol. 26,
22–34.
[3] Solyanikova, I.P., Moiseeva, O.V., Boeren, S., Boersma, M.G.,
Kolomytseva, M.P., Vervoort, J., Rietjens, I.M.C.M., Golovleva,
L.A. and van Berkel, W.J.H. (2003) Conversion of 2-fluoromuc-
onate to cis-dienelactone by purified enzymes of Rhodococcus
opacus 1cp. Appl. Environ. Microbiol. 69, 5636–5642.
[4] Harper, D.B. and Blakley, E.R. (1971) The metabolism of p-
fluorobenzoate by a Pseudomonas sp. Can. J. Microbiol. 17, 1015–
1023.
[5] Schlomann, M., Fischer, P., Schmidt, E. and Knackmuss, H.-J.
(1990) Enzymatic formation, stability and spontaneous reactions
of 4-fluoromuconolactone, a metabolite of the bacterial degrada-
tion of 4-fluorobenzoate. J. Bacteriol. 172, 5119–5129.
[6] Oltmanns, R.H., Mueller, R., Otto, M.F. and Lingens, F. (1989)
Evidence for a new pathway in the bacterial degradation of 4-
fluorobenzoate. Appl. Environ. Microbiol. 55, 2499–2504.
[7] Engesser, K.H. and Schulte, P. (1989) Degradation of 2-bromo-,
2-chloro- and 2-fluorobenzoate by Pseudomonas pudita CLB 250.
FEMS Microbiol. Lett. 60, 143–148.

F.G. Hidde Boersma et al. / FEMS Microbiology Letters 237 (2004) 355–361 361
[8] Vora, K.A., Singh, C. and Modi, V.V. (1988) Degradation of 2-
fluorobenzoate by a pseudomonad. Curr. Microbiol. 17, 249–254.
[9] Clarke, K.F., Callely, A.G., Livingstone, A. and Fewson, C.A.
(1975) Metabolism of monofluorobenzoates by Acinetobacter
calcoaceticusN.C.I.B. 8250. Biochim. Biophys. Acta 404, 169–179.
[10] Schreiber, A., Hellwig,M., Dorn, E., Reineke,W. andKnackmuss,
H.-J. (1980) Critical reactions in fluorobenzoate degradation by
Pseudomonas sp. B13. Appl. Environ. Microbiol. 39, 58–67.
[11] Engesser, K.H., Auling, G., Busse, J. and Knackmuss, H.-J.
(1990) 3-Fluorobenzoate enriched bacterial strain FLB 300
degrades benzoate and all three isomeric monofluorobenzoates.
Arch. Microbiol. 153, 193–199.
[12] Reineke, W., Otting, W. and Knackmuss, H.-J. (1978) cis-
Dihydrodiols microbially produced from halo- and methylbenzoic
acids. Tetrahedron 34, 1707–1714.
[13] Vogel, H.J. and Bonner, D.M. (1956) Acetylornithase of E. coli:
partial purification and some properties. J. Biol. Chem. 218, 97–
106.
[14] Cooke, J.A. (1972) Fluoride compounds in plants: their occur-
rence, distribution and effects. Ph.D. Thesis, University of
Newcastle upon Tyne.
[15] Staden, R., Beal, K.F. and Bonfield, J.K. (2000) The Staden
package, 1998. Methods Mol. Biol. 132, 115–130.
[16] Boersma, M.G., Dinarieva, T.Y., Middelhoven, W.J., van
Berkel, W.J.H., Doran, J., Vervoort, J. and Rietjens,
I.M.C.M. (1998) 19F Nuclear magnetic resonance as a tool to
investigate microbial degradation of fluorophenols to fluoroca-
techols and fluoromuconates. Appl. Environ. Microbiol. 64,
1256–1263.
[17] Schmidt, E. and Knackmuss, H.-J. (1984) Production of cis,cis-
muconate from benzoate and 2-fluoro-cis,cis-muconate from 3-
fluorobenzoate by 3-chlorobenzoate degrading bacteria. Appl.
Microbiol. Biotechnol. 20, 351–355.
[18] Imai, R., Nagata, Y., Senoo, K., Wada, H., Fukuda, M.,
Takagi, M. and Yano, K. (1989) Dehydrochlorination of
c-hexachlorocyclohexane (cBHC) by c-BHC-assimilating
Pseudomonas paucimobilis. Agric. Biol. Chem. 53, 2015–
2017.
[19] Colores, G.M., Radehaus, P.M. and Schmidt, S.K. (1995) Use
of a pentachlorophenol degrading bacterium to bioremediate
highly contaminated soil. Appl. Biochem. Biotechnol. 54, 271–
275.
Copyright © 2022 FDOKUMEN




















