
2013, Orta-Zavalza, Local silencing controls the oxidative stress response and the multidrug...
14
Local silencing controls the oxidative stress response and the multidrug resistance in Candida glabrata Emmanuel Orta-Zavalza, Gehenna Guerrero-Serrano, Guadalupe Gutiérrez-Escobedo, Israel Cañas-Villamar, Jacqueline Juárez-Cepeda, Irene Castaño and Alejandro De Las Peñas* IPICYT, División de Biología Molecular, Instituto Potosino de Investigación Científica y Tecnológica, Camino a la Presa San José 2055, Lomas 4 a sección, San Luis Potosí 78216, México. Summary In Candida glabrata, the sirtuins Sir2 and Hst1 control the expression of a wide number of genes including adhesins required for host colonization and niacin transporters needed for growth. Given that these sirtuins can be inactivated during infection, we asked if their inhibition could modify the response of C. glabrata to other stressful conditions. Here, we found that deletion of HST1 decreases susceptibility of C. glabrata to fluconazole and hydrogen peroxide. The transcription factor Pdr1 and the ABC transporter Cdr1 mediated the fluconazole resistance phenotype of the hst1Δ cells, whereas the transcriptional activator Msn4 and the catalase Cta1 are necessary to provide oxida- tive stress resistance. We show that the transcription factor Sum1 interacts with Hst1 and participate in the regulation of these genes. Interestingly, even though C. glabrata and Saccharomyces cerevisiae are closely related phylogenetically, deletion of HST1 decreased susceptibility to fluconazole and hydrogen peroxide only in C. glabrata but not in S. cerevisiae, indicating a different transcriptional control by two similar sirtuins. Our findings suggest that Hst1 acts as a regulator of stress resistance associated-genes. Introduction Candida glabrata is a commensal yeast that acts as an opportunistic pathogen in immunocompromised patients. In the last three decades, C. glabrata has emerged as the second most frequent causative agent of mucosal and bloodstream fungal infections (Richardson and Lass-Florl, 2008; Pfaller and Diekema, 2010). Changes in the tran- scription program are a major strategy of C. glabrata to adapt to hostile environments. For example, induction of extracellular proteases, siderophore transporters, ele- ments of the oxidative stress response (OSR), autophagy, shift in metabolism and chromatin remodelling are required for the survival of C. glabrata inside phagocytes (Kaur et al., 2007; Cuellar-Cruz et al., 2008; Roetzer et al., 2010; 2011; Nevitt and Thiele, 2011; Seider et al., 2011; Rai et al., 2012; Fukuda et al., 2013). In addition, C. glabrata induces membrane-spanning efflux pumps in the presence of flu- conazole (Sanglard et al., 1999; Vermitsky and Edlind, 2004; Sanguinetti et al., 2005), which are associated with multidrug resistance (MDR) (Pfaller et al., 2010). The response to environmental stimuli is essential not only to ensure the survival of C. glabrata within the host but also to detect conditions that favour infection. C. glabrata induces the transcription of virulence-related adhesins when availability of nicotinamide adenine dinucleotide (NAD + ) precursors in the environment is limited (Domergue et al., 2005; Ma et al., 2009). This response is due to the derepression of epithelial adhesin genes encoded in sub- telomeric regions that are subject to chromatin silencing (De Las Penas et al., 2003; Castano et al., 2005; Domergue et al., 2005). In C. glabrata, the Sir complex (Sir2, Sir3 and Sir4) is essential for subtelomeric silencing (De Las Penas et al., 2003; Castano et al., 2005; Rosas- Hernandez et al., 2008). The enzymatic component of the Sir complex is Sir2, a histone deacetylase that uses NAD + as cofactor to keep the chromatin in a transcriptionally repressive state. Interestingly, since C. glabrata is unable to de novo synthesize NAD + (Ma et al., 2007), Sir2 relies on the environmental supply of NAD + precursors (niacin) to maintain silencing. Thus, environmental limitation of niacin causes inactivation of Sir2 and derepression of normally silent adhesins that result in increased adherence to epi- thelial cells (Domergue et al., 2005). Sirtuins are a family of NAD + -dependent protein dea- cetylases present in all organisms, from bacteria to humans (Sauve et al., 2006). C. glabrata contains five sirtuin genes that include SIR2 and four homologues of SIR2 called HST1 – HST4. HST1 is the closest paralogue of SIR2 and both are involved in transcriptional repres- sion, however these proteins display non-overlapping functions (Ma et al., 2009). In S. cerevisiae, the paral- Accepted 26 April, 2013. *For correspondence. E-mail cano@ ipicyt.edu.mx; Tel. (+52) 444 8342000; Fax (+52) 444 8342010. The authors declare no conflict of interest. Molecular Microbiology (2013) ■ doi:10.1111/mmi.12247 © 2013 John Wiley & Sons Ltd
Transcript of 2013, Orta-Zavalza, Local silencing controls the oxidative stress response and the multidrug...

Local silencing controls the oxidative stress response andthe multidrug resistance in Candida glabrata
Emmanuel Orta-Zavalza, Gehenna Guerrero-Serrano,Guadalupe Gutiérrez-Escobedo,Israel Cañas-Villamar, Jacqueline Juárez-Cepeda,Irene Castaño and Alejandro De Las Peñas*IPICYT, División de Biología Molecular, InstitutoPotosino de Investigación Científica y Tecnológica,Camino a la Presa San José 2055, Lomas 4a sección,San Luis Potosí 78216, México.
Summary
In Candida glabrata, the sirtuins Sir2 and Hst1 controlthe expression of a wide number of genes includingadhesins required for host colonization and niacintransporters needed for growth. Given that thesesirtuins can be inactivated during infection, weasked if their inhibition could modify the responseof C. glabrata to other stressful conditions. Here, wefound that deletion of HST1 decreases susceptibility ofC. glabrata to fluconazole and hydrogen peroxide. Thetranscription factor Pdr1 and the ABC transporter Cdr1mediated the fluconazole resistance phenotype of thehst1Δ cells, whereas the transcriptional activator Msn4and the catalase Cta1 are necessary to provide oxida-tive stress resistance. We show that the transcriptionfactor Sum1 interacts with Hst1 and participate in theregulation of these genes. Interestingly, even thoughC. glabrata and Saccharomyces cerevisiae are closelyrelated phylogenetically, deletion of HST1 decreasedsusceptibility to fluconazole and hydrogen peroxideonly in C. glabrata but not in S. cerevisiae, indicating adifferent transcriptional control by two similar sirtuins.Our findings suggest that Hst1 acts as a regulator ofstress resistance associated-genes.
Introduction
Candida glabrata is a commensal yeast that acts as anopportunistic pathogen in immunocompromised patients.In the last three decades, C. glabrata has emerged as thesecond most frequent causative agent of mucosal andbloodstream fungal infections (Richardson and Lass-Florl,
2008; Pfaller and Diekema, 2010). Changes in the tran-scription program are a major strategy of C. glabrata toadapt to hostile environments. For example, induction ofextracellular proteases, siderophore transporters, ele-ments of the oxidative stress response (OSR), autophagy,shift in metabolism and chromatin remodelling are requiredfor the survival of C. glabrata inside phagocytes (Kauret al., 2007; Cuellar-Cruz et al., 2008; Roetzer et al., 2010;2011; Nevitt and Thiele, 2011; Seider et al., 2011; Rai et al.,2012; Fukuda et al., 2013). In addition, C. glabrata inducesmembrane-spanning efflux pumps in the presence of flu-conazole (Sanglard et al., 1999; Vermitsky and Edlind,2004; Sanguinetti et al., 2005), which are associated withmultidrug resistance (MDR) (Pfaller et al., 2010).
The response to environmental stimuli is essential notonly to ensure the survival of C. glabrata within the host butalso to detect conditions that favour infection. C. glabratainduces the transcription of virulence-related adhesinswhen availability of nicotinamide adenine dinucleotide(NAD+) precursors in the environment is limited (Domergueet al., 2005; Ma et al., 2009). This response is due to thederepression of epithelial adhesin genes encoded in sub-telomeric regions that are subject to chromatin silencing(De Las Penas et al., 2003; Castano et al., 2005;Domergue et al., 2005). In C. glabrata, the Sir complex(Sir2, Sir3 and Sir4) is essential for subtelomeric silencing(De Las Penas et al., 2003; Castano et al., 2005; Rosas-Hernandez et al., 2008). The enzymatic component of theSir complex is Sir2, a histone deacetylase that uses NAD+
as cofactor to keep the chromatin in a transcriptionallyrepressive state. Interestingly, since C. glabrata is unableto de novo synthesize NAD+ (Ma et al., 2007), Sir2 relies onthe environmental supply of NAD+ precursors (niacin) tomaintain silencing. Thus, environmental limitation of niacincauses inactivation of Sir2 and derepression of normallysilent adhesins that result in increased adherence to epi-thelial cells (Domergue et al., 2005).
Sirtuins are a family of NAD+-dependent protein dea-cetylases present in all organisms, from bacteria tohumans (Sauve et al., 2006). C. glabrata contains fivesirtuin genes that include SIR2 and four homologues ofSIR2 called HST1 – HST4. HST1 is the closest paralogueof SIR2 and both are involved in transcriptional repres-sion, however these proteins display non-overlappingfunctions (Ma et al., 2009). In S. cerevisiae, the paral-
Accepted 26 April, 2013. *For correspondence. E-mail [email protected]; Tel. (+52) 444 8342000; Fax (+52) 444 8342010. Theauthors declare no conflict of interest.
Molecular Microbiology (2013) ■ doi:10.1111/mmi.12247
© 2013 John Wiley & Sons Ltd

ogues Sir2 and Hst1 have sub-functionalized after geneduplication (Mead et al., 2007; Froyd and Rusche, 2011)so that Sir2 operates through regional transcriptionalsilencing (Moazed et al., 1997; Tanny et al., 1999),whereas Hst1 acts as a local transcriptional repressor (Xieet al., 1999; McCord et al., 2003). The functional diver-gence between these sirtuins is likely conserved inC. glabrata (Domergue et al., 2005; Rosas-Hernandezet al., 2008; Ma et al., 2009). Hst1 controls the transcrip-tion of genes encoding for high-affinity niacin transporters(TNA1, TNR1 and TNR2) required to maintain NAD+
homeostasis in C. glabrata (Ma et al., 2009). WhenC. glabrata is starved for niacin, Hst1 losses its activityand the expression of these genes is derepressed, toensure proper levels of NAD+ precursors that are requiredfor growth (Ma et al., 2009). In S. cerevisiae, Hst1 alsoacts as a sensor that regulates the NAD+ levels, butmainly by controlling the biosynthetic NAD+ genes(Bedalov et al., 2003). It is worth noting that the transcrip-tional profile of the hst1Δ mutants between S. cerevisiaeand C. glabrata is notably different (Hirao et al., 2003; Maet al., 2009).
The auxotrophy of C. glabrata for NAD+ links the inac-tivation of sirtuins with the environment and the coloniza-tion of specific niches in the host, such as the urinary tract(Domergue et al., 2005; Ma et al., 2009). Given that sir-tuins participate in transcriptional repression and can beinactivated in some niches of the host, we asked if theinactivation of sirtuins could modify the adaptive capacityof C. glabrata against stressful conditions. In this study,we decided to evaluate the role of the sirtuins in the OSRand the MDR of C. glabrata. The involvement of sirtuins inthe control of either of the two processes remains largelyunexplored. Here, we show the existence of a layer ofepigenetic regulation that represses genes involved in theMDR and the OSR of C. glabrata. The induction of theseresponses is mediated by the specific inactivation of Hst1and this control is present only in C. glabrata but not in thephylogenetically related yeast S. cerevisiae. Furthermore,Hst1 interacts with Sum1 and this interaction is required torepress its target genes in C. glabrata.
Results
Inhibition of Hst1 decreases susceptibility to fluconazolein C. glabrata
In C. glabrata, sirtuins participate in the transcriptionalregulation of a wide number of genes including the majorvirulence-associated adhesins (EPA genes) and transport-ers required for niacin assimilation (Domergue et al., 2005;Ma et al., 2009). Inactivation of sirtuins may occur in someniches of the host and can lead to the derepression ofadditional genes. We asked whether derepression of theremaining genes regulated by sirtuins affects the capacityof C. glabrata to adapt to other environmental stresses. Weevaluated the involvement of sirtuins in two stress condi-tions: fluconazole exposure and oxidative stress. Wespotted serial dilutions of stationary phase cells of theparental strain (BG14) on YPD plates in the absence orpresence of 10 mM nicotinamide [NAM, a physiologicalnon-competitive inhibitor of the NAD+-dependent histonedeacetylases (Bitterman et al., 2002)] and different con-centrations of fluconazole (see Experimental procedures).Interestingly, the parental strain (BG14) was able to grow athigher concentrations of fluconazole (32 μg ml−1) in thepresence of NAM (Fig. 1A). This result indicated that sir-tuins could be repressing genes involved in fluconazoleresistance.
We then determined whether the sirtuins Sir2, Hst1 andHst2, could participate in the resistance to fluconazole (seeExperimental procedures). In principle, if one of thesedeacetylases controls fluconazole resistance genes, thedeletion of the sirtuin will display a resistant phenotype.The sir2Δ and the hst2Δ mutant strains were able to growat 8 μg ml−1 of fluconazole, the same concentration as theparental strain BG14 (Fig. 1A – NAM and data not shown).These results indicate that the reduced susceptibility tofluconazole observed in the presence of NAM, is not due toSir2 or Hst2. In contrast, deletion of HST1 (hst1Δ) showedthe same reduced susceptibility to fluconazole as theparental strain in the presence of NAM (Fig. 1A). Theanalysis of the single mutants, hst1Δ and sir2Δ treated withNAM, and the double mutant hst1Δ sir2Δ suggested that
Fig. 1. C. glabrata and S. cerevisiae susceptibility to fluconazole in the absence of sirtuins. (A) C. glabrata parental strain (BG14), hst1Δ(CGM84) and sir2Δ (BG1048) mutant strains and (B) S. cerevisiae parental strain (BY4742), hst1Δ (CGM1094), sir2Δ (L66), sum1Δ (L101)and rfm1Δ (L104) mutant strains were grown for 48 h at 30°C with constant shaking in YPD broth. Cultures were adjusted to an OD = 0.5 indistilled water and logarithmically diluted. Dilutions were spotted on YPD agar plates containing different fluconazole concentrations (0, 8, 16and 32 μg ml−1) and 10 mM of NAM where indicated. See Experimental procedures.
2 E. Orta-Zavalza et al. ■
© 2013 John Wiley & Sons Ltd, Molecular Microbiology

the reduced susceptibility to fluconazole is due to the reliefof the Hst1-mediated repression (Figs 1Aand S1). We thenperformed a quantitative fluconazole susceptibility assayand found that the hst1Δ mutant MIC50 was 6.3 timeshigher than BG14, consistent with the genetic data (Table 1and Fig. 1A). Furthermore, given that S.c.Hst1 andC.g.Hst1 are highly similar (76%), we asked if S. cerevisiaemutant strains, Schst1Δ or Scsir2Δ, could also showreduced susceptibility to fluconazole. We found that S. cer-evisiae parental strain (BY4742), Schst1Δ, and Scsir2Δ didnot alter the fluconazole sensitivity to the extent caused bythe hst1Δ mutant in C. glabrata (Fig. 1B). In summary,these results suggest that Hst1 acts as a negative regulatorof genes that decrease susceptibility to fluconazole andthat despite the high similarity between Sir2 and Hst1(72%), these proteins have non-overlapping functions (Maet al., 2009).
PDR1 and CDR1 are necessary for fluconazoledecreased susceptibility in the absence of HST1
The overexpression of ABC efflux pumps represents themajor mechanism responsible for high-level azole resist-ance in C. glabrata isolates (Bennett et al., 2004; Brunet al., 2004; Borst et al., 2005; Cannon et al., 2009;Ferrari et al., 2009). To date there is one report suggestingthat genes of the pleiotropic drug resistance (PDR) couldbe negatively regulated by sirtuins (Ma et al., 2009). Wedecided to analyse the sensitivity to fluconazole when thetranscriptional factor (PDR1) and the efflux pumps (CDR1or CDR2) were deleted in the hst1Δ mutant backgroundand in the presence of NAM. Consistent with previousreports (Sanglard et al., 1999; Vermitsky and Edlind,2004), the pdr1Δ and cdr1Δ mutants were hypersensitiveto fluconazole (Fig. 2A and Table 1, compare BG14,pdr1Δ and cdr1Δ). However, deletion of CDR2 (alsoknown as PDH1; Izumikawa et al., 2003) did not alter thefluconazole sensitivity in the presence or absence ofHST1 (Fig. 2A, compare BG14 and cdr2Δ and hst1Δ withcdr2Δ hst1Δ), possibly because Cdr1 is compensatingfor the absence of Cdr2 (Izumikawa et al., 2003). Thedecreased susceptibility to fluconazole of the hst1Δ strainwas abolished in the absence of PDR1 or CDR1 (Fig. 2A,compare hst1Δ with pdr1Δ hst1Δ and cdr1Δ hst1Δ andTable 1). These results indicate that both genes arerequired to mediate azole resistance when Hst1 is absent.Notably, even when the pdr1Δ hst1Δ and cdr1Δ hst1Δdouble mutants were more sensitive to fluconazole thanthe hst1Δ mutant strain, their sensitivity was not the sameas the exhibited by the single mutant pdr1Δ or cdr1Δ
Table 1. Fluconazole susceptibility of C. glabrata strains.
Strain MIC50 (μg ml−1)a
BG14 14.93 ± 2.951hst1Δ 93.68 ± 21.8sum1Δ 100.2 ± 21.07rfm1Δ 101.3 ± 23.88pdr1Δ 2.427 ± 0.6116cdr1Δ 2.803 ± 0.6957hst1Δ pdr1Δ 3.933 ± 0.8991hst1Δ cdr1Δ 3.94 ± 0.6811hst1 C344A C347A 82.19 ± 11.26
a. MIC50 were determined by a modified microdilution method, seeExperimental procedures.
Fig. 2. Fluconazole susceptibility of C. glabrata under nicotinamide treatment and in the absence of HST1 or multidrug resistance genes.C. glabrata parental strain (BG14), and hst1Δ (CGM84), pdr1Δ (CGM1094), cdr1Δ (CGM1096), cdr2Δ (CGM1098), pdr1Δ hst1Δ (CGM1117),cdr1Δ hst1Δ (CGM1118) and cdr2Δ hst1Δ (CGM1120) mutant strains were grown and treated with fluconazole (A) and fluconazole with 10 mMNAM (B) as described in the legend to Fig 1. See Experimental procedures.
Hst1 controls azole and H2O2 resistance in C. glabrata 3
© 2013 John Wiley & Sons Ltd, Molecular Microbiology
(Fig. 2A, compare pdr1Δ with pdr1Δ hst1Δ and cdr1Δ withcdr1Δ hst1Δ), suggesting that CDR1 could be dere-pressed in pdr1Δ hst1Δ and PDR1 could be derepressedin cdr1Δ hst1Δ inducing the expression of other drugtransporter encoding gene like CDR2; thus providing theadditional reduced sensitivity. Quantification of the MIC50of the single and double mutants indicated that the differ-ence between these mutants is less evident than thegenetic data (compare MIC50 of hst1Δ, pdr1Δ, and cdr1Δ,with hst1Δ pdr1Δ and hst1Δ cdr1Δ from Table 1 andFig. 2A). The MIC50 value for the set of mutants shown inTable 1 was highly reproducible in YPD medium, howeverwe could not confirm these phenotypes using RPMI-1640medium as described in the NCCLS M-27 A2 method.Furthermore, the analysis of the susceptibility to flucona-zole of the hst1Δ, pdr1Δ, cdr1Δ and the double mutants inthe presence of NAM, were consistent with NAM inhibitingHst1 activity. The presence of NAM mimics the absence ofHst1 (Fig. 2B, compare pdr1Δ and pdr1Δ hst1Δ with pdr1Δhst1Δ -NAM in Fig. 2A and cdr1Δ and cdr1Δ hst1Δ withcdr1Δ hst1Δ -NAM in Fig. 2A). These data suggest thatthe decreased susceptibility due to the absence of HST1is mainly mediated by CDR1 and PDR1.
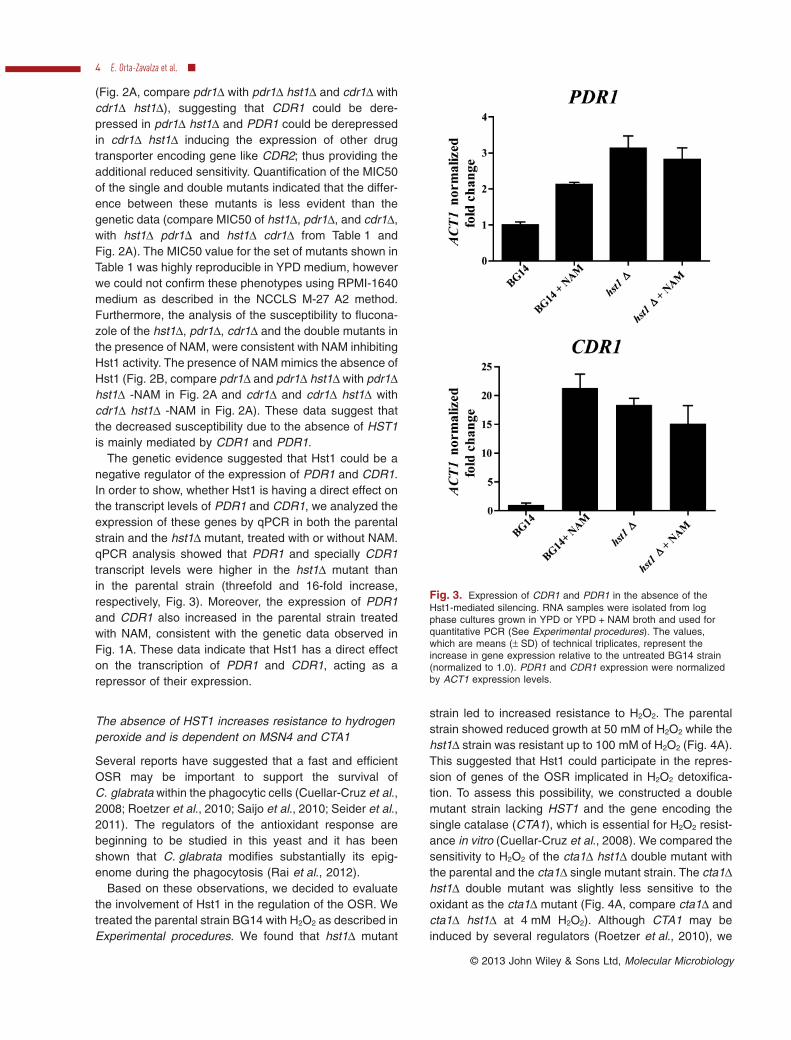
The genetic evidence suggested that Hst1 could be anegative regulator of the expression of PDR1 and CDR1.In order to show, whether Hst1 is having a direct effect onthe transcript levels of PDR1 and CDR1, we analyzed theexpression of these genes by qPCR in both the parentalstrain and the hst1Δ mutant, treated with or without NAM.qPCR analysis showed that PDR1 and specially CDR1transcript levels were higher in the hst1Δ mutant thanin the parental strain (threefold and 16-fold increase,respectively, Fig. 3). Moreover, the expression of PDR1and CDR1 also increased in the parental strain treatedwith NAM, consistent with the genetic data observed inFig. 1A. These data indicate that Hst1 has a direct effecton the transcription of PDR1 and CDR1, acting as arepressor of their expression.
The absence of HST1 increases resistance to hydrogenperoxide and is dependent on MSN4 and CTA1
Several reports have suggested that a fast and efficientOSR may be important to support the survival ofC. glabrata within the phagocytic cells (Cuellar-Cruz et al.,2008; Roetzer et al., 2010; Saijo et al., 2010; Seider et al.,2011). The regulators of the antioxidant response arebeginning to be studied in this yeast and it has beenshown that C. glabrata modifies substantially its epig-enome during the phagocytosis (Rai et al., 2012).
Based on these observations, we decided to evaluatethe involvement of Hst1 in the regulation of the OSR. Wetreated the parental strain BG14 with H2O2 as described inExperimental procedures. We found that hst1Δ mutant
strain led to increased resistance to H2O2. The parentalstrain showed reduced growth at 50 mM of H2O2 while thehst1Δ strain was resistant up to 100 mM of H2O2 (Fig. 4A).This suggested that Hst1 could participate in the repres-sion of genes of the OSR implicated in H2O2 detoxifica-tion. To assess this possibility, we constructed a doublemutant strain lacking HST1 and the gene encoding thesingle catalase (CTA1), which is essential for H2O2 resist-ance in vitro (Cuellar-Cruz et al., 2008). We compared thesensitivity to H2O2 of the cta1Δ hst1Δ double mutant withthe parental and the cta1Δ single mutant strain. The cta1Δhst1Δ double mutant was slightly less sensitive to theoxidant as the cta1Δ mutant (Fig. 4A, compare cta1Δ andcta1Δ hst1Δ at 4 mM H2O2). Although CTA1 may beinduced by several regulators (Roetzer et al., 2010), we
Fig. 3. Expression of CDR1 and PDR1 in the absence of theHst1-mediated silencing. RNA samples were isolated from logphase cultures grown in YPD or YPD + NAM broth and used forquantitative PCR (See Experimental procedures). The values,which are means (± SD) of technical triplicates, represent theincrease in gene expression relative to the untreated BG14 strain(normalized to 1.0). PDR1 and CDR1 expression were normalizedby ACT1 expression levels.
4 E. Orta-Zavalza et al. ■
© 2013 John Wiley & Sons Ltd, Molecular Microbiology
decided to evaluate the involvement of the transcriptionfactor MSN4 because it was overexpressed to a greaterextent in a hst1Δ mutant strain (Ma et al., 2009). Weconstructed the msn4Δ hst1Δ double mutant strain andevaluated its resistance to H2O2. The msn4Δ mutant strainhad the same sensitivity to H2O2 as the parental strain(Fig. 4A, compare msn4Δ with BG14). However, themsn4Δ mutation in the hst1Δ background abolishedthe H2O2 resistant phenotype. This result indicates thatthe oxidative stress resistance induced by the absence ofHST1 is predominantly due to the presence of the cata-lase and might be regulated by Msn4. Furthermore, wealso found that in S. cerevisiae, the response to oxidativestress is unchanged in the absence of ScHST1 or ScSIR2(Fig. 4B). Along with the results described above, CgHst1is regulating a different set of genes than those controlledby the orthologue ScHst1.
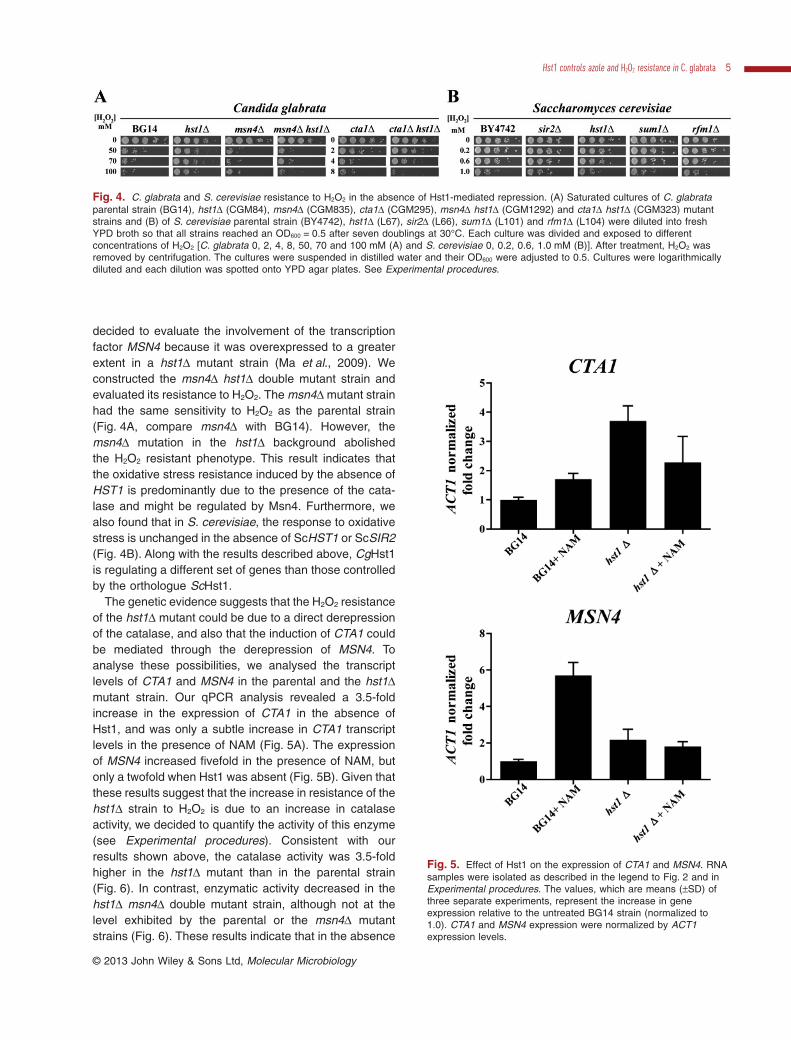
The genetic evidence suggests that the H2O2 resistanceof the hst1Δ mutant could be due to a direct derepressionof the catalase, and also that the induction of CTA1 couldbe mediated through the derepression of MSN4. Toanalyse these possibilities, we analysed the transcriptlevels of CTA1 and MSN4 in the parental and the hst1Δmutant strain. Our qPCR analysis revealed a 3.5-foldincrease in the expression of CTA1 in the absence ofHst1, and was only a subtle increase in CTA1 transcriptlevels in the presence of NAM (Fig. 5A). The expressionof MSN4 increased fivefold in the presence of NAM, butonly a twofold when Hst1 was absent (Fig. 5B). Given thatthese results suggest that the increase in resistance of thehst1Δ strain to H2O2 is due to an increase in catalaseactivity, we decided to quantify the activity of this enzyme(see Experimental procedures). Consistent with ourresults shown above, the catalase activity was 3.5-foldhigher in the hst1Δ mutant than in the parental strain(Fig. 6). In contrast, enzymatic activity decreased in thehst1Δ msn4Δ double mutant strain, although not at thelevel exhibited by the parental or the msn4Δ mutantstrains (Fig. 6). These results indicate that in the absence
Fig. 4. C. glabrata and S. cerevisiae resistance to H2O2 in the absence of Hst1-mediated repression. (A) Saturated cultures of C. glabrataparental strain (BG14), hst1Δ (CGM84), msn4Δ (CGM835), cta1Δ (CGM295), msn4Δ hst1Δ (CGM1292) and cta1Δ hst1Δ (CGM323) mutantstrains and (B) of S. cerevisiae parental strain (BY4742), hst1Δ (L67), sir2Δ (L66), sum1Δ (L101) and rfm1Δ (L104) were diluted into freshYPD broth so that all strains reached an OD600 = 0.5 after seven doublings at 30°C. Each culture was divided and exposed to differentconcentrations of H2O2 [C. glabrata 0, 2, 4, 8, 50, 70 and 100 mM (A) and S. cerevisiae 0, 0.2, 0.6, 1.0 mM (B)]. After treatment, H2O2 wasremoved by centrifugation. The cultures were suspended in distilled water and their OD600 were adjusted to 0.5. Cultures were logarithmicallydiluted and each dilution was spotted onto YPD agar plates. See Experimental procedures.
Fig. 5. Effect of Hst1 on the expression of CTA1 and MSN4. RNAsamples were isolated as described in the legend to Fig. 2 and inExperimental procedures. The values, which are means (±SD) ofthree separate experiments, represent the increase in geneexpression relative to the untreated BG14 strain (normalized to1.0). CTA1 and MSN4 expression were normalized by ACT1expression levels.
Hst1 controls azole and H2O2 resistance in C. glabrata 5
© 2013 John Wiley & Sons Ltd, Molecular Microbiology
of HST1, C. glabrata is more resistant to H2O2 through theincrease of the expression of CTA1 that requires the tran-scription factor Msn4.
Cysteine residues 344 and 347 are essential forHst1 activity
Several amino acids have been identified as important forthe structure and enzymatic activity of sirtuins in otherorganisms. Similar to other sirtuins, CgHst1 contains aCys-X-X-Cys-(X)15–20-Cys-X-X-Cys conserved motif thatco-ordinates a Zn2+ ion (Sherman et al., 1999). Interest-ingly, this same sequence motif (Cys-Pro-Tyr-Cys) is alsolocated in the active site of glutaredoxins, which is atarget for oxidation (Foloppe and Nilsson, 2004). Weenvisioned the possibility that Hst1 could be oxidized andinactivated in the presence of oxidative stress. To deter-
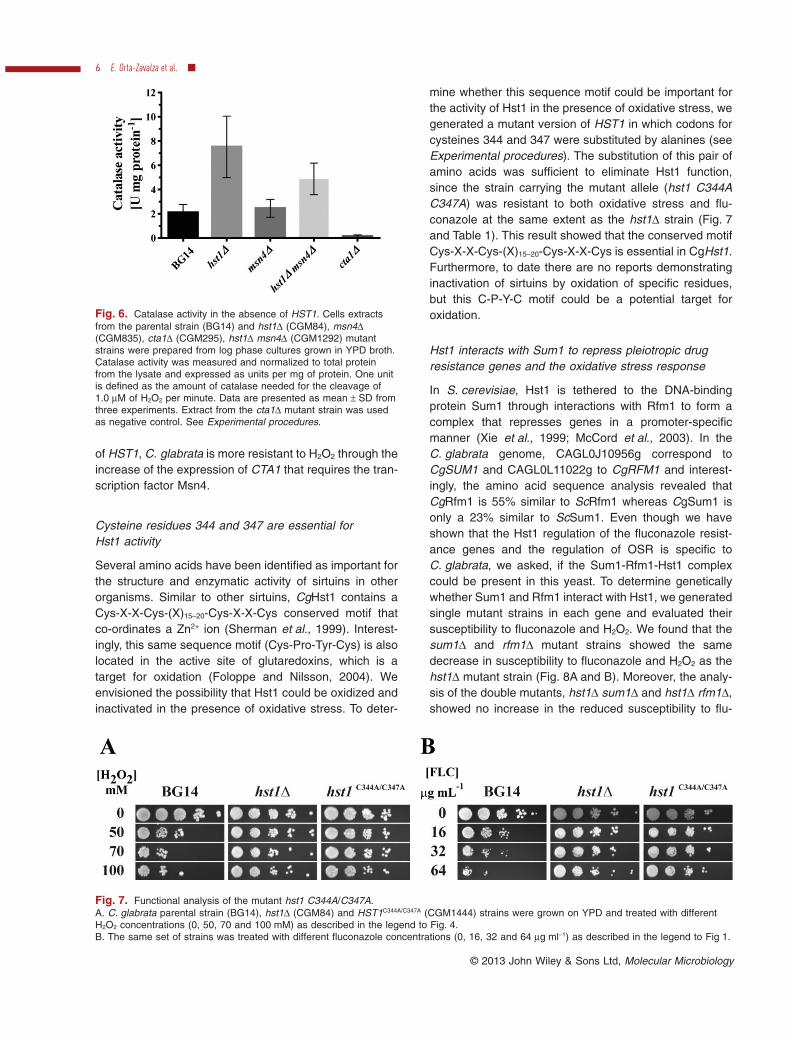
mine whether this sequence motif could be important forthe activity of Hst1 in the presence of oxidative stress, wegenerated a mutant version of HST1 in which codons forcysteines 344 and 347 were substituted by alanines (seeExperimental procedures). The substitution of this pair ofamino acids was sufficient to eliminate Hst1 function,since the strain carrying the mutant allele (hst1 C344AC347A) was resistant to both oxidative stress and flu-conazole at the same extent as the hst1Δ strain (Fig. 7and Table 1). This result showed that the conserved motifCys-X-X-Cys-(X)15–20-Cys-X-X-Cys is essential in CgHst1.Furthermore, to date there are no reports demonstratinginactivation of sirtuins by oxidation of specific residues,but this C-P-Y-C motif could be a potential target foroxidation.
Hst1 interacts with Sum1 to repress pleiotropic drugresistance genes and the oxidative stress response
In S. cerevisiae, Hst1 is tethered to the DNA-bindingprotein Sum1 through interactions with Rfm1 to form acomplex that represses genes in a promoter-specificmanner (Xie et al., 1999; McCord et al., 2003). In theC. glabrata genome, CAGL0J10956g correspond toCgSUM1 and CAGL0L11022g to CgRFM1 and interest-ingly, the amino acid sequence analysis revealed thatCgRfm1 is 55% similar to ScRfm1 whereas CgSum1 isonly a 23% similar to ScSum1. Even though we haveshown that the Hst1 regulation of the fluconazole resist-ance genes and the regulation of OSR is specific toC. glabrata, we asked, if the Sum1-Rfm1-Hst1 complexcould be present in this yeast. To determine geneticallywhether Sum1 and Rfm1 interact with Hst1, we generatedsingle mutant strains in each gene and evaluated theirsusceptibility to fluconazole and H2O2. We found that thesum1Δ and rfm1Δ mutant strains showed the samedecrease in susceptibility to fluconazole and H2O2 as thehst1Δ mutant strain (Fig. 8A and B). Moreover, the analy-sis of the double mutants, hst1Δ sum1Δ and hst1Δ rfm1Δ,showed no increase in the reduced susceptibility to flu-
Fig. 6. Catalase activity in the absence of HST1. Cells extractsfrom the parental strain (BG14) and hst1Δ (CGM84), msn4Δ(CGM835), cta1Δ (CGM295), hst1Δ msn4Δ (CGM1292) mutantstrains were prepared from log phase cultures grown in YPD broth.Catalase activity was measured and normalized to total proteinfrom the lysate and expressed as units per mg of protein. One unitis defined as the amount of catalase needed for the cleavage of1.0 μM of H2O2 per minute. Data are presented as mean ± SD fromthree experiments. Extract from the cta1Δ mutant strain was usedas negative control. See Experimental procedures.
Fig. 7. Functional analysis of the mutant hst1 C344A/C347A.A. C. glabrata parental strain (BG14), hst1Δ (CGM84) and HST1C344A/C347A (CGM1444) strains were grown on YPD and treated with differentH2O2 concentrations (0, 50, 70 and 100 mM) as described in the legend to Fig. 4.B. The same set of strains was treated with different fluconazole concentrations (0, 16, 32 and 64 μg ml−1) as described in the legend to Fig 1.
6 E. Orta-Zavalza et al. ■
© 2013 John Wiley & Sons Ltd, Molecular Microbiology

conazole or H2O2 when compared with the single mutants,suggesting that these genes are components of the samepathway (Fig. 8A and B). As control, reconstitution of boththe HST1 and SUM1 in their corresponding loci in thehst1Δ and sum1Δ mutant strains, reverted the fluconazolephenotype (data not shown). Finally and consistent withthe results shown above for S. cerevisiae, deletion ofScHST1, ScSUM1 and ScRFM1 did not alter the suscep-tibility to fluconazole or H2O2 in this yeast (Figs 1B and4B).
Our genetic analysis points toward an interactionbetween Hst1, Sum1 and Rfm1 in C. glabrata. In orderto demonstrate this association, we performed a co-immunoprecipitation experiment using cMyc-tagged Hst1and Flag-tagged Sum1 proteins. First, to determine if thetagged proteins were functional, we analysed the sensitiv-ity to fluconazole of the HST1::cMyc SUM1::FLAG strain(CGM982). We found that the strain carrying the epitope-tagged alleles had the same fluconazole sensitivity pheno-type as the parental strain, indicating that both proteins arefunctional (Fig. S2). We then performed the Co-IP usingstrain CGM982 (HST1::cMyc SUM1::FLAG) as describedin the legend to Fig. 9 and in Experimental procedures. Wefound that Sum1-Flag co-immunoprecipitated with Hst1-Myc (Fig. 9, lane 4) and that this interaction is largelydependent on Rfm1, since the co-immunoprecipitationdecreased when the RFM1 gene was deleted in the back-ground of the double–tagged strain CGM1294 (Fig. 9, lane5) and the majority of the Sum1-Flag remained in thesupernatant. Taken together, our findings support a modelin which Sum1, Rfm1 and Hst1 interact to form a complexthat represses genes of the MDR and the OSR inC. glabrata.
Discussion
Sirtuins are NAD+-dependent deacetylases highly con-served from bacteria to humans (Greiss and Gartner,2009). In S. cerevisiae, sirtuins participate in gene silen-cing, DNA repair, replication, recombination, cell cycle
Fig. 8. Analysis of HST1, SUM1 and RFM1 in the fluconazole sensitivity and oxidative stress response. C. glabrata parental strain (BG14),hst1Δ (CGM84), sir2Δ (BG1048), sum1Δ (CGM673), rfm1Δ (CGM943), hst1Δ sum1Δ (CG1607) and hst1Δ rfm1Δ (CG1609) mutant strainswere grown and treated with fluconazole and H2O2 as described in the legends to Figs 1 and 4 respectively. See Experimental procedures.
Fig. 9. Analysis of Hst1 and Sum1 interaction. Total extracts wereobtained from log phase cells of the parental strain (BG14,untagged, lane 1), from the single tagged strains: CGM934(Hst1-Myc, lane 2) and CGM1342 (Sum1-Flag, lane 3); and fromthe double tagged strains: CGM982 (Hst1-Myc Sum1-Flag, lane 4)and CGM1294 (Hst1-Myc Sum1-Flag tagged rfm1Δ, lane 5).Extracts from these strains were analysed by Western blot using anantibody specific to the Flag and Myc epitope for the detection ofSum1 and Hst1 respectively (Input). Extracts from these strainswere co-immunoprecipitated with antibodies specific to the Mycepitope and probed with antibodies specific to the Flag epitope(IP α-Myc, WB α-Flag). The remaining supernatant afterimmunoprecipitation with antibodies specific to the Myc epitope wasanalysed using antibodies against Flag (SN α-Myc, WB α-Flag).IP, immunoprecipitation; WB, Western blot; SN, supernatant.
Hst1 controls azole and H2O2 resistance in C. glabrata 7
© 2013 John Wiley & Sons Ltd, Molecular Microbiology

control and longevity (Finkel et al., 2009; Vaquero, 2009).In the pathogenic yeast C. glabrata, sirtuins are involved inthe transcriptional repression of genes encoding cell wallproteins which mediate adhesion and also genes encodingtransporters of niacin, both processes are important forcolonization of the host (Domergue et al., 2005; Ma et al.,2009). However, given the auxotrophy of C. glabrata forNAD+, the activity of sirtuins is completely dependent onthe environmental availability of NAD+ precursors (niacin)(Ma et al., 2009). In host niches that are limited in niacin(urinary tract), sirtuins are inactivated and C. glabrataincreases its adherence and niacin uptake. In this work, wedecided to evaluate if inactivation of sirtuins modifies thecapacity of this yeast to adapt to different stress conditions.
Inhibition of Hst1 decreases the susceptibility tofluconazole in C. glabrata
Here, we showed that C. glabrata decreases its suscepti-bility to fluconazole through the inhibition of Hst1 in thepresence of NAM (Fig. 1). Based on our data, we proposethat this phenotype is mediated primarily by the ABCtransporter CDR1 and the transcription factor PDR1(Figs 2 and 3; Table 1). However it is possible that othertransporters could be derepressed as well, although withless affinity to fluconazole (Fig. 2). The main mechanisminvolved in the acquisition of resistance to azoles inC. glabrata, is the overexpression of genes belonging tothe family ofABC transporters [Reviewed in (Cannon et al.,2009)]. Interestingly, to date almost all reported mecha-nisms involving overexpression of those transportersengage genetic modifications, such as the acquisition ofgain of function mutations in Pdr1(Ferrari et al., 2009), lossof mitochondrial function (Kaur et al., 2004; Ferrari et al.,2011) and increased gene dosage of transporters bychromosomal rearrangements (Polakova et al., 2009).Recently it was demonstrated that some genes thatprovide azole resistance in C. glabrata are under the nega-tive regulation of Stb5, a zinc cluster transcription factor(Noble et al., 2013). Here, we propose, based on ourgenetic and biochemical analysis, that Hst1 represses thetranscription of PDR1 and CDR1. This hypothesis is furthersupported by the microarray analysis of the C. glabratastrain lacking Hst1 (Ma et al., 2009), where the absence ofHST1 allows the induction of CDR1, PDR1 and CDR2(PDH1).
Elimination of the Hst1-mediated silencing increases theoxidative stress resistance of C. glabrata
We found that C. glabrata increased the transcript levelsof CTA1, its catalase activity and its resistance to H2O2 inthe absence of HST1 (Figs 4, 5 and 6). Thus, it is probablethat CTA1 is subject to negative control mediated by Hst1.
The purpose of this negative regulation on the catalase isstill unclear; although it has been shown that an increasein catalase activity is associated with premature aging ofS. cerevisiae (Mesquita et al., 2010). This pro-aging activ-ity has not been found in C. glabrata but silencing of CTA1could contribute to maintain low level of transcript andlimit the deleterious effect of this enzyme on viability. Inaddition, our results also suggest that the resistanceobserved in the absence of HST1 requires the transcrip-tion factor MSN4 (Figs 4 and 6). While it was true that theincrease in the transcript levels of MSN4 were subtle, islikely to be sufficient to activate transcription of CTA1(Fig. 5). Nevertheless, this finding apparently contrastswith previous observations that proposed a lack of partici-pation by Msn2/4 in the control of CTA1 (Roetzer et al.,2011) although we detect the role of Msn4 only in theabsence of Hst1. Thus, the resistance to oxidative stressobserved in the hst1Δ cells may be due to at least two notmutually exclusive possibilities: 1) direct derepression ofCTA1 or 2) derepression of one or several activators ofCTA1 (such as MSN4) that are under the control of Hst1.Transcriptional regulation of CTA1 is a fine-tuned processthat is subject to a complex interaction of factors thatinfluence its activation and repression.
Sum1, Rfm1 and Hst1 repress genes of stressresistance in C. glabrata but not in S. cerevisiae
The precise mechanism by which Hst1 regulates thegenes of the MDR and OSR is still not completely eluci-dated. However, here we showed that Hst1 could berepressing the transcription of genes that participate inboth processes (Figs 3 and 5). Moreover, the putativetranscriptional factor Sum1 and the connector proteinRfm1 were also required for the activity of Hst1 (Fig. 8)and the physical interaction of Hst1 with Sum1 wasdependent on Rfm1 (Fig. 9). These findings are consist-ent with the results reported for S. cerevisiae (Xie et al.,1999; McCord et al., 2003) and suggest that the complexis conserved in both organisms. In S. cerevisiae, Hst1and Sum1 control the NAD+ biosynthetic genes (Bedalovet al., 2003) and Hst1 has retained the role of regulatingniacin uptake in C. glabrata (Ma et al., 2009). Surprisingly,deletion of any of the genes encoding the proteins thatcomprise the Hst1-Rfm1-Sum1 complex decreased sus-ceptibility to fluconazole and H2O2 in C. glabrata but not inS. cerevisiae (Figs 1 and 4). These similar deacetylases(74% similar) control different genes in phylogeneticallyrelated organisms. In agreement with this observation, thetranscriptional profiles of S. cerevisiae and C. glabratastrains carrying a mutation in HST1, show that only 19genes are regulated in common by this sirtuin in these twoyeasts [Comparing the upregulated genes found in themicroarrays reported by Ma and Hirao (Hirao et al., 2003;
8 E. Orta-Zavalza et al. ■
© 2013 John Wiley & Sons Ltd, Molecular Microbiology

Ma et al., 2009)]. It is possible that the reason for thisdifferential regulation might lie in Sum1. At the proteinlevel, CgSum1 is only 23% similar to ScSum1. BLASTanalysis shows no putative conserved motifs over theentire length of Sum1 proteins, but the N-domain appearsto be the most conserved region in C. glabrata and otherphylogenetically related yeasts (data not shown). Despitethe divergence that CgSum1 has undergone, our resultssuggest that it may conserve important amino acid resi-dues needed for its interaction with CgRfm1 and probablyalso for DNA binding. Based in our data we propose amodel in which Sum1 interacts with Rfm1 and directs Hst1to the promoters of the MDR and OSR genes to silencethem (Fig. 10).
Hst1 as a regulator of genes associated with survivalwithin the host
Our data also confirm that despite the high similaritybetween Sir2 and Hst1 (74%), only the absence of Hst1-
mediated silencing decreases the susceptibility ofC. glabrata to fluconazole and oxidative stress. In addi-tion, deletion of HST1 also increases the in vitroadherence of C. glabrata to Lec2 epithelial cells(B.P. Cormack and I. Castano, pers. comm.) and thetranscription of EPA6 (Domergue et al., 2005), suggest-ing that Hst1 also participates in the repression of someepithelial adhesins. Therefore, it appears that Hst1represses genes that could be advantageous to thispathogen (Fig. 10), thus Hst1 could be inactivated duringinfection. C. glabrata colonizes the genitourinary tractwhere the levels of niacin are limited so it is feasible thatHst1 loses its activity and allow derepression of its targetgenes under this condition. Consistent with this, it hasbeen suggested that MSN4 and CTA1 are upregulatedunder limitation of niacin and in human urine (Ma et al.,2009).
According to our model, inactivation of Hst1 could be anadvantage for C. glabrata because it would allow coloni-zation of the host through the derepression of specificadhesins and also would increase resistance to xenobio-tics and oxidative stress (Fig. 10). This work uncoversa novel level of regulation in the MDR and OSR circuitsof C. glabrata. We think that this epigenetic regulationof resistance could be advantageous for C. glabratabecause, either it might allow the stochastic appearanceof resistant cells within a genetically identical populationwithout permanent genetic alterations that may compro-mise the fitness of the yeast or the Hst1-Rfm1-Sum1complex could be a target of different environmentalsignals. It remains to be evaluated whether these mecha-nisms are relevant to the acquisition of resistance to flu-conazole during therapy of the patients and to endure therespiratory burst during phagocytosis. It will also be worthexploring whether maintaining the activity of Hst1 couldbe beneficial to the host, as this condition would keepsilent the genes that mediate adhesion and resistance ofC. glabrata.
Experimental procedures
Strains and growth conditions
Escherichia coli strain DH10 (Gibco BRL) was used forplasmid transformation, isolation and storage in 10% glyc-erol. All mutant C. glabrata strains (Table 2) were generatedin the BG14 strain background and here referred to as paren-tal strain (Cormack and Falkow, 1999). The S. cerevisiaemutant strains were all derivatives from BY4742 (Table 3). Allyeast strains were stored in 15% glycerol stocks at −80°C.Yeast cultures were incubated at 30°C in YPD [1% yeastextract, 2% peptone, 2% glucose and supplemented with25 mg ml−1 of uracil (Sigma Aldrich®)] or in synthetic completemedium (SC) (yeast nitrogen base at 1.7 g l−1 and (NH4)2SO4
at 5 g l−1, supplemented with 0.6% Casamino Acids and 2%glucose). For solid media, 2% agar was added.
Fig. 10. Model of the Hst1-Rfm1-Sum1 complex that controlgenes involved in stress resistance of C. glabrata. Under normalconcentrations of NAD+, Sum1 interacts with Rfm1 to recruit thehistone deacetylase Hst1 and repress their target genes. WhenC. glabrata is exposed to the sirtuins inhibitor (NAM), limitingamounts of niacin, or oxidative stress, Hst1 loses its activity andallows derepression of genes involved in niacin uptake, adherence,resistance to azoles and oxidative stress. There are probablyadditional signals that can inactivate the complex. Loss of theHst1-mediated silencing leads to the induction of the ABCtransporter CDR1, the catalase CTA1, and the transcriptionalfactors PDR1 and MSN4. As a result, (i) C. glabrata decreases itssusceptibility to fluconazole through active efflux of the drug, whichis mainly mediated by Cdr1, (ii) derepression of CTA1 increasesthe resistance of C. glabrata to H2O2, (iii) C. glabrata could increaseits adherence to epithelial cells through the derepression of EPA6,and (iv) derepression of the high-affinity niacin transporters (TNA1,TNR1 and TNR2) increases the niacin uptake in order to restoreNAD+ homeostasis, probably also to preserve the function of therest of the sirtuins and to avoid detrimental effects of theirinactivation.
Hst1 controls azole and H2O2 resistance in C. glabrata 9
© 2013 John Wiley & Sons Ltd, Molecular Microbiology

Plasmid and strain construction
Plasmids used in this study are described in Table S1. Allplasmids were purified from E. coli using the Qiagen® PlasmidKit QIAprep. All gene fragments for plasmid constructs and forgene disruption were generated by PCR using the oligonu-cleotides detailed in Table S2. Fusion PCR for gene disruptionwas performed as previously described (Baudin et al., 1993)and all PCR products were purified before transformationusing Qiagen® Gel Extraction Kit. C. glabrata was transformed
using the lithium acetate protocol and the knockout mutationsand the epitope-tagging were constructed by two-step recom-bination; these methods had been described previously(Castano et al., 2003; De Las Penas et al., 2003; Cuellar-Cruzet al., 2008). To construct the null mutant strains, the openreading frame (ORF) of each target gene was replaced with ahygromycin resistance cassette or with the nourseothricinresistance cassette. The plasmids used for epitope taggingcontain a C-terminal segment of the gene without stop codon(HST1 or SUM1) placed in phase with the epitope sequence
Table 2. Candida glabrata strains.
Strain Parent Relevant genotype Reference
BG2 Clinical isolate Fidel et al. (1996)BG14 BG2 ura3Δ::Tn903 G418R Cormack et al. (1999)BG1048 BG14 ura3Δ::Tn903 G418R sir2Δ::hph HygR Domergue et al. (2005)BG1073 BG14 ura3Δ::Tn903 G418R hst1Δ::hph HygR Domergue et al. (2005)CGM84 BG1073 ura3Δ::Tn903 G418R hst1Δ This studyCGM295 BG14 ura3Δ::Tn903 G418R cta1Δ::hph HygR Cuellar-Cruz et al. (2008)CGM323 CGM84 ura3Δ::Tn903 G418R hst1Δ/cta1Δ::hph HygR This studyCGM673 BG14 ura3Δ::Tn903 G418R sum1Δ::hph HygR This studyCGM712 CGM84 ura3Δ::Tn903 G418R hst1Δ/sir2Δ::hph HygR
(Made with pAP596)This study
CGM943 BG14 ura3Δ::Tn903 G418R rfm1Δ::NatMX4 NatR
(Fusion PCR, oligonucleotides #27-#32)This study
CGM1094 BG14 ura3Δ::Tn903 G418R pdr1Δ::hph HygR
(Fusion PCR, oligonucleotides #13-#18)This study
CGM1096 BG14 ura3Δ::Tn903 G418R cdr1Δ::hph HygR
(Fusion PCR, oligonucleotides #13-#14, #19-#22)This study
CGM1098 BG14 ura3Δ::Tn903 G418R CDR2Δ::hph HygR
(Fusion PCR, oligonucleotides #13-#14, #23-#26)This study
CGM1117 CGM84 ura3Δ::Tn903 G418R hst1Δ/pdr1Δ::hph HygR
(Fusion PCR, oligonucleotides #13-#18)This study
CGM1118 CGM84 ura3Δ::Tn903 G418R hst1Δ/cdr1Δ::hph HygR
(Fusion PCR, oligonucleotides #13-#14, #19-#22)This study
CGM1120 CGM84 ura3Δ::Tn903 G418R hst1Δ/CDR2Δ::hph HygR
Fusion PCR, oligonucleotides #13-#14, #23-#26)This study
BG1740 BG14 ura3Δ::Tn903 G418R msn4Δ::hph HygR
(pRD97/Bcg I)Cormack lab collection
CGM835 BG1740 ura3Δ::Tn903 G418R msn4Δ::hph HygS This studyCGM1292 CGM84 ura3Δ::Tn903 G418R hst1Δ/msn4Δ::hph HygR
(pRD97/Bcg I)This study
CGM1336 BG14 ura3Δ::Tn903 G418R hst1 C344A/C347A::URA3 Ura+
(pOZ86/Msc I )This study
CGM1444 CGM1336 ura3Δ::Tn903 G418R hst1 C344A/C347A This studyCGM823 BG14 ura3Δ::Tn903 G418R HST1-cMyc13::hph HygR
(pOZ32/Xmn I-Blp I)This study
CGM934 CGM823 ura3Δ::Tn903 G418R HST1-cMyc13 This studyCGM1342 BG14 ura3Δ::Tn903 G418R SUM1-FLAG::hph HygR
(pOZ72/Pvu II-Bgl II)This study
CGM982 CGM934 ura3Δ::Tn903 G418R HST1-cMyc13 SUM1-FLAG::hph HygR
(pOZ72/Pvu II-Bgl II)This study
CGM1294 CGM982 ura3Δ::Tn903 G418R HST1-cMyc13 SUM1-FLAG::hph HygR
rfm1Δ::NatMX4 NatR (Fusion PCR, oligonucleotides #27–#32)This study
CGM1232 CGM673 ura3Δ::Tn903 G418R sum1Δ This studyCGM1234 CGM943 ura3Δ::Tn903 G418R rfm1Δ This studyCGM1607 CGM1232 ura3Δ::Tn903 G418R sum1Δ hst1Δ::hph HygR
(Made with pAP628)This study
CGM1609 CGM1234 ura3Δ::Tn903 G418R rfm1Δ hst1Δ::hph HygR
(Made with pAP628)This study
CGM1605 CGM84 ura3Δ::Tn903 G418R HST1(pOZ102/Blp I)
This study
CGM1611 CGM1232 ura3Δ::Tn903 G418R SUM1(pOZ100/Bgl II)
This study
10 E. Orta-Zavalza et al. ■
© 2013 John Wiley & Sons Ltd, Molecular Microbiology

(cMyc or FLAG). This translational fusion is followed by the 3'UTR of the catalase gene (3' UTRCTA1) and a hygromycinresistance cassette for selection of the transformants. The 3'UTRCTA1 and the hygromycin cassette are flanked by two FRTdirect repeats. Downstream the hygromycin cassette, theconstructs contain a fragment of the 3' UTR of the gene. Themutant or epitope tagged strains were selected on YPDplates containing 440 μg ml−1 hygromycin (A. G. Scientific®) or100 μg ml−1 nourseothricin (Werner BioAgents®). The correctreplacement in the chromosome was confirmed by PCR.For each target gene, two independent transformants wereobtained and tested. To place the HST1-cMyc and SUM1-FLAG with their native 3' UTR, the epitope-tagged HygR strainswere transformed with pMZ21 (Table S1), and transformantswere selected on SC-Ura plates. pMZ21 is a replicative vectorexpressing ScFLP1 that encodes a recombinase that recog-nizes the FRT sites flanking the hygromycin marker. After Flp1recognizes the FRT sites, the hph marker is deleted from thechromosome and the cognate 3' UTR is placed immediatelydownstream of the tagged gene and leaves one copy of theFRT site (35 bp). Transformants were purified on SC-Uraplates. Single colonies were then grown on non-selectivemedium (YPD agar) and screened for HygS for the loss of theHyg cassette and for Ura− for the loss of pMZ21. This protocolallows the construction of multiple mutants or multiple taggedstrains.
For site directed mutagenesis, the coding region of HST1was mutated using PCR so that Cys residues were replacedby Ala. Expand Long Template PCR System (Roche®) wasused for amplification of the fragments. Sequences of themutagenic oligonucleotides used for the generation of thehst1 C344A C347A mutant are listed in Table S2. The muta-genic oligonucleotides were designed to mutate the selectedcodons and simultaneously introduce a novel Nhe I restrictionsite, which facilitates distinction of mutant from wild-type frag-ment at subsequent cloning steps. To confirm the introductionof the mutation, plasmid pOZ50 was sequenced. To replaceHST1 with the mutant allele (C344A C347A), WT-strain wastransformed with linearized pOZ86 using the pop-in/pop-outmethod. Segregants were selected on SC-Ura plates. Topromote recombination, colonies were cultured in YPD brothand plated on YPD agar plates. Loss of URA3 was selectedby streaking colonies on 5-FOA plates. Finally identification ofthe mutants was achieved using PCR, restriction analysisand sequencing.
Fluconazole susceptibility assays
To test for susceptibility to fluconazole (Pfizer®), C. glabratacells were grown for 48 h at 30°C in YPD broth, diluted to
OD600 = 0.5 in distilled water, and 10-fold serial dilutions werespotted onto YPD agar plates containing different fluconazoleconcentrations (Sanglard et al., 1999). To assess fluconazolesusceptibility under inhibition of sirtuins, 10 mM of NAM(Sigma Aldrich®) was added to the plates containing flucona-zole. Spots were made using a VP 407AH 48-pin multi-blotreplicator (V&P Scientific Inc.). Plates were incubated at 30°Cfor 36 h or 48 h. Experiments were repeated at least threetimes.
To quantify the sensitivity to fluconazole, stationary phasecultures of the indicated strains were diluted to obtain a cellularsuspension in YPD. One hundred and fifty microlitres of thissuspension (104 cells ml−1) was transferred to multi-wellsplates (Oy Growth Curves Ab Ltd) where each well contained150 μl of broth with different concentrations of fluconazole(0.0078, 0.0156, 0.0312, 0.0625, 0.125, 0.250, 0.5, 1, 2, 4, 8,16, 32, 64, 72, 80, 96, 112, 128, 160, 192, 224 or 256 μg ml−1).Growth was then continuously monitored in a Bioscreen Csystem (Oy Growth Curves Ab Ltd) at 600 nm, for each 15 minat 35°C during 24 h. Sensitivities were determined as mean50% Minimal Inhibitory Concentrations (MIC50), calculated onthe basis of the dose-response relationship in the GraphPadPrism software (GraphPad Software Inc., San Diego, CA,USA). The CLSI microdilution method M27-A2 was also usedfor susceptibility testing but RPMI-1640 broth was supple-mented with 25 mg ml−1 of uracil.
H2O2 susceptibility assay
Sensitivity to hydrogen peroxide (H2O2) of logarithmic phasecells was determined as described previously (Cuellar-Cruzet al., 2009). A 35% wt H2O2 solution (Sigma Aldrich®) wasused for the assay. Cultures were grown for 48 h at 30°C inYPD broth, diluted into fresh YPD medium in such way that allthe strains went through seven duplications to reach 0.5 OD600.After this period, the cultures were divided, exposed to differ-ent H2O2 concentrations and incubated with shaking at 30°Cfor three hours. After the treatment, the H2O2 was removed bycentrifugation and the cells were suspended in 1 ml of distilledwater. The cultures were adjusted to an OD600 = 0.5, seriallydiluted and spotted on YPD plates as described above. Theplates were incubated at 30°C for 36 h or 48 h. The experi-ments were performed at least three times.
Catalase activity assay
Strains were grown at 30°C for seven duplications in YPDbroth until cultures reached an OD600 = 0.5. Cells were col-lected by centrifugation, washed twice with distilled water and
Table 3. Saccharomyces cerevisiae strains.
Strain Parent Relevant genotype Reference
BY4742 MATα his3Δ1, leu2Δ, lys2Δ, ura3Δ Brachmann et al. (1998)L66 BY4742 MATα his3Δ1, leu2Δ, lys2Δ, ura3Δ, sir2Δ::KanMX KanR Rine lab collectionL67 BY4742 MATα his3Δ1, leu2Δ, lys2Δ, ura3Δ, hst1Δ::KanMX KanR Rine lab collectionL101 BY4742 MATα his3Δ1, leu2Δ, lys2Δ, ura3Δ, sum1Δ::KanMX KanR Rine lab collectionL104 BY4742 MATα his3Δ1, leu2Δ, lys2Δ, ura3Δ, rfm1Δ::KanMX KanR Rine lab collection
Hst1 controls azole and H2O2 resistance in C. glabrata 11
© 2013 John Wiley & Sons Ltd, Molecular Microbiology

suspended in lysis buffer [pottasium phosphate (pH 7.0)supplemented with 1× general protease inhibitors [SigmaFAST®]). A volume of zirconia beads (BioSpec®) was addedand cells were disrupted by vortexing at 4°C for 1 min andplaced on ice for another minute (repeated 20 times). Thelysate was centrifuged at 25 000 g for 30 min at 4°C toremove cell debris and zirconia beads. The supernatant wasused for measure of catalase activity and its protein contentwas determined by Bradford assay (Fermentas®). Bovineserum albumin (Sigma®) was used as standard. Catalaseactivity was determined by the spectrophotometric methodthat measures the breakdown of H2O2 by catalase (Aebi,1984) and the assays were performed three times. The activ-ity was normalized to total content of protein from the lysateand expressed as units per mg of protein. One unit is definedas the amount of catalase required for degradation of1.0 μmol H2O2 per minute.
Quantitative PCR (qPCR)
RNA was extracted from log phase cells (grown in YPD or inYPD broth containing 10 mM NAM) using TRIzol reagent(Invitrogen) according to the manufacturer’s instructions andtreated with DNase I (Invitrogen) to ensure removal of con-taminating genomic DNA. Synthesis of cDNA was carried outusing SuperScript® II reverse transcriptase and the reverseoligonucleotides for each gene (Table S2) and the reactionwas carried out at 42°C for 50 min. Quantitative PCR (qPCR)was performed using the Fast SYBR® Green Master Mix(Invitrogen) in the ABI 7500 Fast Real-Time PCR System(Applied Biosystems). The reverse and forward oligonucleo-tides used for the qRT-PCR were designed using the primerexpress 3.0 software (Table S2, ACT1 #41–42, CTA1 #43–44, CDR1 #45–46, MSN4 #47–48 and PDR1 #49–50). ACT1was used as an internal control for normalization and thethreshold cycle (2−ΔΔCt) method was used to calculate thedifferences in gene expression. qPCR experiments were per-formed in technical triplicates and from three independentRNA extractions.
Co-immunoprecipitation experiments
The CoIP protocol was performed as described (McCordet al., 2003). Strains were grown at 30°C for seven duplica-tions in YPD broth until cultures reached an OD600 = 1.0. Cellswere collected by centrifugation and suspended in 300 μlof lysis buffer (45 mM HEPES-KOH [Promega®, pH 7.5],400 mM potassium acetate, 1 mM EDTA, 0.5% Nonidet P-40substitute [Fluka Biochemica®], 1 mM DTT, 10% glycerol,1 mM PMSF, 1 × general protease inhibitors [Sigma FAST®]and 1 × Complete protease inhibitors [ROCHE®]). A volumeof 300 μl of zirconia beads (BioSpec®) was added and cellswere lysed as described for in the catalase activity protocol.The lysate was centrifuged at 25 000 g for 30 min at 4°C. Thesupernatant of each sample was transferred to a new tubeand protein content was determined by Bradford assay(Fermentas®). For each sample, 100 μg of protein were incu-bated at 4°C for 1 h with 30 μl of sepharose beads containingcross-linked protein G (Sigma®). The samples were centri-fuged at 25 000 g for 5 min at 4°C and the clarified lysate was
transferred to a clean tube. For immunoprecipitation, thelysate was combined with 50 μl of anti-cMyc or anti-FLAGagarose (Sigma®) and incubated at 4°C for two hours withconstant inversion. The supernatant was recovered and IPswere collected by centrifugation, washed three times withLysis buffer, suspended in 40 μl of 2× sodium dodecyl sul-phate (SDS) loading buffer and heated at 95°C for 5 min. Theinput (20 μg of protein), supernatant (one twentieth of theimmunoprecipitated extract) and the IPs were loaded onto a10% SDS-polyacrylamide gel. After electrophoresis, the pro-teins were blotted onto PVDF membranes (BIO-RAD®) andprobed with either mouse anti-cMyc (Millipore®) or mouseanti-FLAG (Sigma®) for two hours at room temperature.After washing, the membranes were probed with a goatanti-mouse horseradish peroxidase-conjugated secondaryantibody (Amersham®). Signal detection was achieved usingthe ECL chemiluminescence reagents (Amersham®) andX-OMAT (Kodak®) films.
Acknowledgements
We thank Jasper Rine for kindly providing S. cerevisiaestrains (University of California, Berkeley, CA, USA). We aregrateful to Sergio Casas and Lina Riego (IPICYT, San LuisPotosí, SLP, Mexico) for providing reagents. We thank CandyY. Ramírez, Omar Arroyo and Marcela Briones for technicalsupport, Jorge Folch (UAEM) and members of the A.D.L.P.laboratory for their suggestions and careful reading of themanuscript. This work was funded by CONACYT fellowshipsto E.O.Z. (233455), G.G.S. (230938), M.G.G.E. (48880),I.C.V. (224300), J.J.C. (48549) and by CONACYT Grant No.CB-2010-01-153929 and UC Mexus Grant No. CN-07-53 toA.D.L.P.
References
Aebi, H. (1984) Catalase in vitro. Methods Enzymol 105:121–126.
Baudin, A., Ozier-Kalogeropoulos, O., Denouel, A., Lacroute,F., and Cullin, C. (1993) A simple and efficient method fordirect gene deletion in Saccharomyces cerevisiae. NucleicAcids Res 21: 3329–3330.
Bedalov, A., Hirao, M., Posakony, J., Nelson, M., and Simon,J.A. (2003) NAD+-dependent deacetylase Hst1p controlsbiosynthesis and cellular NAD+ levels in Saccharomycescerevisiae. Mol Cell Biol 23: 7044–7054.
Bennett, J.E., Izumikawa, K., and Marr, K.A. (2004) Mecha-nism of increased fluconazole resistance in Candidaglabrata during prophylaxis. Antimicrob Agents Chemother48: 1773–1777.
Bitterman, K.J., Anderson, R.M., Cohen, H.Y., Latorre-Esteves, M., and Sinclair, D.A. (2002) Inhibition of silencingand accelerated aging by nicotinamide, a putative negativeregulator of yeast Sir2 and human SIRT1. J Biol Chem 277:45099–45107.
Borst, A., Raimer, M.T., Warnock, D.W., Morrison, C.J., andArthington-Skaggs, B.A. (2005) Rapid acquisition of stableazole resistance by Candida glabrata isolates obtainedbefore the clinical introduction of fluconazole. AntimicrobAgents Chemother 49: 783–787.
12 E. Orta-Zavalza et al. ■
© 2013 John Wiley & Sons Ltd, Molecular Microbiology

Brachmann, C.B., Davies, A., Cost, G.J., Caputo, E., Li, J.,Hieter, P., and Boeke, J.D. (1998) Designer deletion strainsderived from Saccharomyces cerevisiae S288C: a usefulset of strains and plasmids for PCR-mediated gene disrup-tion and other applications. Yeast 14: 115–132.
Brun, S., Berges, T., Poupard, P., Vauzelle-Moreau, C.,Renier, G., Chabasse, D., and Bouchara, J. (2004) Mecha-nisms of azole resistance in petite mutants of Candidaglabrata. Antimicrob Agents Chemother 48: 1788–1796.
Cannon, R.D., Lamping, E., Holmes, A.R., Niimi, K., Baret,P.V., Keniya, M.V., et al. (2009) Efflux-mediated antifungaldrug resistance. Clin Microbiol Rev 22: 291–321.
Castano, I., Kaur, R., Pan, S., Cregg, R., Penas Ade, L., Guo,N., et al. (2003) Tn7-based genome-wide random inser-tional mutagenesis of Candida glabrata. Genome Res 13:905–915.
Castano, I., Pan, S.J., Zupancic, M., Hennequin, C., Dujon,B., and Cormack, B.P. (2005) Telomere length control andtranscriptional regulation of subtelomeric adhesins inCandida glabrata. Mol Microbiol 55: 1246–1258.
Cormack, B.P., and Falkow, S. (1999) Efficient homologousand illegitimate recombination in the opportunistic yeastpathogen Candida glabrata. Genetics 151: 979–987.
Cormack, B.P., Ghori, N., and Falkow, S. (1999) An adhesinof the yeast pathogen Candida glabrata mediating adher-ence to human epithelial cells. Science 285: 578–582.
Cuellar-Cruz, M., Briones-Martin-del-Campo, M., Canas-Villamar, I., Montalvo-Arredondo, J., Riego-Ruiz, L.,Castano, I., and De Las Penas, A. (2008) High resistanceto oxidative stress in the fungal pathogen Candida glabratais mediated by a single catalase, Cta1p, and is controlledby the transcription factors Yap1p, Skn7p, Msn2p, andMsn4p. Eukaryot Cell 7: 814–825.
Cuellar-Cruz, M., Castano, I., Arroyo-Helguera, O., andDe Las Penas, A. (2009) Oxidative stress response tomenadione and cumene hydroperoxide in the opportunisticfungal pathogen Candida glabrata. Mem Inst OswaldoCruz 104: 649–654.
De Las Penas, A., Pan, S.J., Castano, I., Alder, J., Cregg, R.,and Cormack, B.P. (2003) Virulence-related surface glyco-proteins in the yeast pathogen Candida glabrata areencoded in subtelomeric clusters and subject to RAP1- andSIR-dependent transcriptional silencing. Genes Dev 17:2245–2258.
Domergue, R., Castano, I., De Las Penas, A., Zupancic, M.,Lockatell, V., Hebel, J.R., et al. (2005) Nicotinic acid limi-tation regulates silencing of Candida adhesins during UTI.Science 308: 866–870.
Ferrari, S., Ischer, F., Calabrese, D., Posteraro, B.,Sanguinetti, M., Fadda, G., et al. (2009) Gain of functionmutations in CgPDR1 of Candida glabrata not only mediateantifungal resistance but also enhance virulence. PLoSPathog 5: e1000268.
Ferrari, S., Sanguinetti, M., De Bernardis, F., Torelli, R.,Posteraro, B., Vandeputte, P., and Sanglard, D. (2011) Lossof mitochondrial functions associated with azole resistancein Candida glabrata also results in enhanced virulence inmice. Antimicrob Agents Chemother 55: 1852–1860.
Fidel, P.L., Jr, Cutright, J.L., Tait, L., and Sobel, J.D. (1996) Amurine model of Candida glabrata vaginitis. J Infect Dis173: 425–431.
Finkel, T., Deng, C.X., and Mostoslavsky, R. (2009) Recentprogress in the biology and physiology of sirtuins. Nature460: 587–591.
Foloppe, N., and Nilsson, L. (2004) The glutaredoxin-C-P-Y-C-motif: influence of peripheral residues. Structure 12:289–300.
Froyd, C.A., and Rusche, L.N. (2011) The duplicated dea-cetylases Sir2 and Hst1 subfunctionalized by acquiringcomplementary inactivating mutations. Mol Cell Biol 31:3351–3365.
Fukuda, Y., Tsai, H.F., Myers, T.G., and Bennett, J.E. (2013)Transcriptional profiling of Candida glabrata during phago-cytosis by neutrophils and in the infected mouse spleen.Infect Immun 81: 1325–1333.
Greiss, S., and Gartner, A. (2009) Sirtuin/Sir2 phylogeny,evolutionary considerations and structural conservation.Mol Cells 28: 407–415.
Hirao, M., Posakony, J., Nelson, M., Hruby, H., Jung, M.,Simon, J.A., and Bedalov, A. (2003) Identification of selec-tive inhibitors of NAD+-dependent deacetylases usingphenotypic screens in yeast. J Biol Chem 278: 52773–52782.
Izumikawa, K., Kakeya, H., Tsai, H.F., Grimberg, B., andBennett, J.E. (2003) Function of Candida glabrata ABCtransporter gene, PDH1. Yeast 20: 249–261.
Kaur, R., Castano, I., and Cormack, B. (2004) Functionalgenomic analysis of fluconazole susceptibility in the patho-genic yeast Candida glabrata: roles of calcium signalingand mitochondria. Antimicrob Agents Chemother 48:1600–1613.
Kaur, R., Ma, B., and Cormack, B. (2007) A family ofglycosylphosphatidylinositol-linked aspartyl proteases isrequired for virulence of Candida glabrata. Proc Natl AcadSci USA 104: 7628–7633.
Ma, B., Pan, S.J., Zupancic, M.L., and Cormack, B.P. (2007)Assimilation of NAD+ precursors in Candida glabrata. MolMicrobiol 66: 14–25.
Ma, B., Pan, S.J., Domergue, R., Rigby, T., Whiteway, M.,Johnson, D., and Cormack, B.P. (2009) High affinity trans-porters for NAD+ precursors in Candida glabrata are regu-lated by Hst1 and induced in response to niacin limitation.Mol Cell Biol 29: 4067–4079.
McCord, R., Pierce, M., Xie, J., Wonkatal, S., Mickel, C., andVershon, A.K. (2003) Rfm1, a novel tethering factorrequired to recruit the Hst1 histone deacetylase for repres-sion of middle sporulation genes. Mol Cell Biol 23: 2009–2016.
Mead, J., McCord, R., Youngster, L., Sharma, M.,Gartenberg, M.R., and Vershon, A.K. (2007) Swapping thegene-specific and regional silencing specificities of theHst1 and Sir2 histone deacetylases. Mol Cell Biol 27:2466–2475.
Mesquita, A., Weinberger, M., Silva, A., Sampaio-Marques,B., Almeida, B., Leao, C., et al. (2010) Caloric restriction orcatalase inactivation extends yeast chronological lifespanby inducing H2O2 and superoxide dismutase activity. ProcNatl Acad Sci USA 107: 15123–15128.
Moazed, D., Kistler, A., Axelrod, A., Rine, J., and Johnson,A.D. (1997) Silent information regulator protein complexesin Saccharomyces cerevisiae: a SIR2/SIR4 complex andevidence for a regulatory domain in SIR4 that inhibits its
Hst1 controls azole and H2O2 resistance in C. glabrata 13
© 2013 John Wiley & Sons Ltd, Molecular Microbiology

interaction with SIR3. Proc Natl Acad Sci USA 94: 2186–2191.
Nevitt, T., and Thiele, D.J. (2011) Host iron withholdingdemands siderophore utilization for Candida glabrata tosurvive macrophage killing. PLoS Pathog 7: e1001322.
Noble, J.A., Tsai, H.F., Suffis, S.D., Su, Q., Myers, T.G., andBennett, J.E. (2013) STB5 Is a Negative Regulator of AzoleResistance in Candida glabrata. Antimicrob Agents Chem-other 57: 959–967.
Pfaller, M.A., and Diekema, D.J. (2010) Epidemiology of inva-sive mycoses in North America. Crit Rev Microbiol 36:1–53.
Pfaller, M.A., Diekema, D.J., Gibbs, D.L., Newell, V.A., Ellis,D., Tullio, V., et al. (2010) Results from the ARTEMIS DISKGlobal Antifungal Surveillance Study, 1997 to 2007: a 10.5-year analysis of susceptibilities of Candida Species to flu-conazole and voriconazole as determined by CLSIstandardized disk diffusion. J Clin Microbiol 48: 1366–1377.
Polakova, S., Blume, C., Zarate, J.A., Mentel, M., Jorck-Ramberg, D., Stenderup, J., and Piskur, J. (2009) Forma-tion of new chromosomes as a virulence mechanism inyeast Candida glabrata. Proc Natl Acad Sci USA 106:2688–2693.
Rai, M.N., Balusu, S., Gorityala, N., Dandu, L., and Kaur, R.(2012) Functional genomic analysis of Candida glabrata-macrophage interaction: role of chromatin remodeling invirulence. PLoS Pathog 8: e1002863.
Richardson, M., and Lass-Florl, C. (2008) Changing epide-miology of systemic fungal infections. Clin Microbiol Infect14 (Suppl. 4): 5–24.
Roetzer, A., Gratz, N., Kovarik, P., and Schuller, C. (2010)Autophagy supports Candida glabrata survival duringphagocytosis. Cell Microbiol 12: 199–216.
Roetzer, A., Klopf, E., Gratz, N., Marcet-Houben, M., Hiller,E., Rupp, S., et al. (2011) Regulation of Candida glabrataoxidative stress resistance is adapted to host environment.FEBS Lett 585: 319–327.
Rosas-Hernandez, L.L., Juarez-Reyes, A., Arroyo-Helguera,O.E., De Las Penas, A., Pan, S.J., Cormack, B.P., andCastano, I. (2008) yKu70/yKu80 and Rif1 regulate silenc-ing differentially at telomeres in Candida glabrata. EukaryotCell 7: 2168–2178.
Saijo, T., Miyazaki, T., Izumikawa, K., Mihara, T., Takazono,
T., Kosai, K., et al. (2010) Skn7 is involved in oxidativestress response and virulence of Candida glabrata. Myco-pathologia 169: 81–90.
Sanglard, D., Ischer, F., Calabrese, D., Majcherczyk, P.A.,and Bille, J. (1999) The ATP binding cassette transportergene CgCDR1 from Candida glabrata is involved in theresistance of clinical isolates to azole antifungal agents.Antimicrob Agents Chemother 43: 2753–2765.
Sanguinetti, M., Posteraro, B., Fiori, B., Ranno, S., Torelli, R.,and Fadda, G. (2005) Mechanisms of azole resistance inclinical isolates of Candida glabrata collected during a hos-pital survey of antifungal resistance. Antimicrob AgentsChemother 49: 668–679.
Sauve, A.A., Wolberger, C., Schramm, V.L., and Boeke, J.D.(2006) The biochemistry of sirtuins. Annu Rev Biochem 75:435–465.
Seider, K., Brunke, S., Schild, L., Jablonowski, N., Wilson, D.,Majer, O., et al. (2011) The facultative intracellular patho-gen Candida glabrata subverts macrophage cytokine pro-duction and phagolysosome maturation. J Immunol 187:3072–3086.
Sherman, J.M., Stone, E.M., Freeman-Cook, L.L.,Brachmann, C.B., Boeke, J.D., and Pillus, L. (1999) Theconserved core of a human SIR2 homologue functions inyeast silencing. Mol Biol Cell 10: 3045–3059.
Tanny, J.C., Dowd, G.J., Huang, J., Hilz, H., and Moazed, D.(1999) An enzymatic activity in the yeast Sir2 protein that isessential for gene silencing. Cell 99: 735–745.
Vaquero, A. (2009) The conserved role of sirtuins in chroma-tin regulation. Int J Dev Biol 53: 303–322.
Vermitsky, J.P., and Edlind, T.D. (2004) Azole resistance inCandida glabrata: coordinate upregulation of multidrugtransporters and evidence for a Pdr1-like transcriptionfactor. Antimicrob Agents Chemother 48: 3773–3781.
Xie, J., Pierce, M., Gailus-Durner, V., Wagner, M., Winter, E.,and Vershon, A.K. (1999) Sum1 and Hst1 repress middlesporulation-specific gene expression during mitosis in Sac-charomyces cerevisiae. EMBO J 18: 6448–6454.
Supporting information
Additional supporting information may be found in the onlineversion of this article at the publisher’s web-site.
14 E. Orta-Zavalza et al. ■
© 2013 John Wiley & Sons Ltd, Molecular Microbiology



















