
2007 Dubois Zootaxa Ranks zt01519p068
42
Accepted by Z.-Q. Zhang: 23 May 2007; published: 2 Jul. 2007 27 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2007 · Magnolia Press Zootaxa 1519: 27–68 (2007) www.mapress.com/ zootaxa/ Phylogeny, taxonomy and nomenclature: the problem of taxonomic categories and of nomenclatural ranks ALAIN DUBOIS Vertébrés: Reptiles & Amphibiens, USM 0602 Taxonomie & Collections, Département de Systématique & Evolution, Muséum national d’Histoire naturelle, 25 rue Cuvier, 75005 Paris, France. E-mail: [email protected] Tables of contents Abstract ............................................................................................................................................................................. 27 Introduction ....................................................................................................................................................................... 28 Ranks and nominal-series in zoological nomenclature ..................................................................................................... 29 The problem of equivalence between taxa of the same nomenclatural rank .................................................................... 31 The confusion between nomenclatural ranks and taxonomic categories .......................................................................... 35 The usefulness of nomenclatural ranks in taxonomy ........................................................................................................ 38 Clarity and ambiguity in the meaning of nomina .............................................................................................................. 39 Systems of allocation of nomina to taxa, monosemy and polysemy ................................................................................ 41 Extensional definitions of nomina ............................................................................................................................. 41 Intensional definitions of nomina .............................................................................................................................. 42 Ostensional allocation of nomina to taxa ................................................................................................................... 44 Discussion ......................................................................................................................................................................... 46 The distinction between nomenclatural ranks and taxonomic categories .................................................................. 46 Compulsory and optional nomenclatural ranks ......................................................................................................... 48 Should the Code be fundamentally modified to become fully monosemic? ............................................................. 50 The need of clarifications in the Code ....................................................................................................................... 52 Phylogenetic definitions of taxa and the Code ........................................................................................................... 52 Conclusion ......................................................................................................................................................................... 54 Acknowledgements ........................................................................................................................................................... 55 References ......................................................................................................................................................................... 55 Appendix. Cladognoses, apognoses and diagnoses: the example of the family Megophryidae (Amphibia, Anura) ....... 60 Abstract The use of ranks and nominal-series in zoological nomenclature has recently been challenged by some authors who sup- port unranked systems of nomenclature. It is here shown that this criticism is based on a double misunderstanding: (1) the confusion between nomenclatural ranks and taxonomic categories; (2) the request for a monosemic nomenclatural system, not for scientific reasons, but to please non-taxonomists, especially customers of the web. It is here argued that nomenclatural ranks and taxonomic categories should be clearly distinguished and designated by different terms, and that the Code should be modified in order to make this distinction clear. Whereas taxonomic categories have biological defi- nitions, nomenclatural ranks do not, as they express only a position in a taxonomic hierarchy. If used consistently (which is not always the case), the system of nomenclatural ranks is very useful for the storage and retrieval of taxonomic and phylogenetic information. Taxa referred to a given rank in different groups cannot therefore be considered equivalent by any criterion, so that using ranks for comparisons between taxa (e.g., for biodiversity richness assessment) is irrelevant and misleading. Although the current Code needs to be improved in several respects, the superiority of this nomencla- tural system, which is theory-free regarding taxonomy as it relies on ostensional allocation of nomina to taxa rather than
Transcript of 2007 Dubois Zootaxa Ranks zt01519p068

Accepted by Z.-Q. Zhang: 23 May 2007; published: 2 Jul. 2007 27
ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2007 · Magnolia Press
Zootaxa 1519: 27–68 (2007) www.mapress.com/zootaxa/
Phylogeny, taxonomy and nomenclature:the problem of taxonomic categories and of nomenclatural ranks
ALAIN DUBOISVertébrés: Reptiles & Amphibiens, USM 0602 Taxonomie & Collections, Département de Systématique & Evolution, Muséum national d’Histoire naturelle, 25 rue Cuvier, 75005 Paris, France. E-mail: [email protected]
Tables of contents
Abstract .............................................................................................................................................................................27Introduction .......................................................................................................................................................................28Ranks and nominal-series in zoological nomenclature ..................................................................................................... 29The problem of equivalence between taxa of the same nomenclatural rank .................................................................... 31The confusion between nomenclatural ranks and taxonomic categories ..........................................................................35The usefulness of nomenclatural ranks in taxonomy........................................................................................................ 38Clarity and ambiguity in the meaning of nomina ..............................................................................................................39Systems of allocation of nomina to taxa, monosemy and polysemy ................................................................................41
Extensional definitions of nomina ............................................................................................................................. 41Intensional definitions of nomina ..............................................................................................................................42Ostensional allocation of nomina to taxa ...................................................................................................................44
Discussion .........................................................................................................................................................................46The distinction between nomenclatural ranks and taxonomic categories ..................................................................46Compulsory and optional nomenclatural ranks ......................................................................................................... 48Should the Code be fundamentally modified to become fully monosemic? .............................................................50The need of clarifications in the Code .......................................................................................................................52Phylogenetic definitions of taxa and the Code ...........................................................................................................52
Conclusion .........................................................................................................................................................................54Acknowledgements ...........................................................................................................................................................55References .........................................................................................................................................................................55Appendix. Cladognoses, apognoses and diagnoses: the example of the family Megophryidae (Amphibia, Anura) .......60
Abstract
The use of ranks and nominal-series in zoological nomenclature has recently been challenged by some authors who sup-port unranked systems of nomenclature. It is here shown that this criticism is based on a double misunderstanding: (1)the confusion between nomenclatural ranks and taxonomic categories; (2) the request for a monosemic nomenclaturalsystem, not for scientific reasons, but to please non-taxonomists, especially customers of the web. It is here argued thatnomenclatural ranks and taxonomic categories should be clearly distinguished and designated by different terms, and thatthe Code should be modified in order to make this distinction clear. Whereas taxonomic categories have biological defi-nitions, nomenclatural ranks do not, as they express only a position in a taxonomic hierarchy. If used consistently (whichis not always the case), the system of nomenclatural ranks is very useful for the storage and retrieval of taxonomic andphylogenetic information. Taxa referred to a given rank in different groups cannot therefore be considered equivalent byany criterion, so that using ranks for comparisons between taxa (e.g., for biodiversity richness assessment) is irrelevantand misleading. Although the current Code needs to be improved in several respects, the superiority of this nomencla-tural system, which is theory-free regarding taxonomy as it relies on ostensional allocation of nomina to taxa rather than

DUBOIS28 · Zootaxa 1519 © 2007 Magnolia Press
on intensional definitions of nomina, is again stressed. It is suggested that all taxonomists should follow the Code for theallocation and validity of nomina, whatever taxonomic theory they favour, and in particular whatever kinds of definitionsor diagnoses they wish to use for taxa. This would avoid the considerable loss of manpower, time and energy that wouldbe required by the implementation of a new nomenclatural system (e.g., in order to require “phylogenetic definitions” fornomina, or to make nomenclature fully monosemic), and the confusion that would result for most users of nomina. Thenew paradigm imposed to biology by the combination of the taxonomic impediment and of the biodiversity crisisrequires from taxonomists, who are already considerably much less numerous than required by this new situation, to con-centrate on what should be their priority at the beginning of the century of extinctions, namely the inventory of the livingspecies of our planet before they get extinct.
Key words: Taxonomic impediment, Biodiversity crisis, Nomenclatural ranks, Taxonomic categories, Equivalencebetween taxa, Nomenclatural redundancy, Rule of Coordination, Nomenclatural parsimony, Polysemy, Monosemy, Goo-gle taxonomy, Onomatophores, Code, Phylocode
Introduction
We are today at the beginning of the century of extinctions (Dubois 2003). Although this has not been noticedby many yet, the whole of biology in this century will be confronted with a new paradigm, which results froma double reality: (1) the taxonomic impediment (Anonymous 1994), i.e., the fact that, both quantitatively andqualitatively, our knowledge of the species and other taxa of our planet is very unsatisfying, in fact muchbelow the standard required today by our society for all other scientific disciplines; (2) the biodiversity crisis(Wilson, 1985), which, despite all our (laudable) efforts to limit it, will inevitably result in mass extinctions,especially in the tropical regions of the world. To give just one example, more than 30 % of the total numberof described amphibian species of the planet are threatened with extinction and 30 % are data deficient (Stuartet al. 2004), but the number of undescribed species of this group is still gigantic, possibly larger than the num-ber of described species (Dubois 2004a; Köhler et al. 2005), and threats on the unknown species are impossi-ble to estimate.
Confronted to this situation, the only appropriate reply from zoology would be to speed up considerablythe collection, inventory and description of the living species of our planet before they get extinct. However,this message is difficult to pass to our society as a whole: “Funds nominally allocated to taxonomy go largelyto reconstruct molecular phylogenies, while thousands of species are threatened by imminent extinction. (...)In the face of the biodiversity crisis, the need for urgency could be no greater. (...) Our generation is the first tofully comprehend the threat of the biodiversity crisis and the last with the opportunity to explore and docu-ment the species diversity of our planet. Time is rapidly running out. (...) The grand biological challenge ofour age is to create a legacy of knowledge for a planet that is soon to be biologically decimated.” (Wheeler etal. 2004). However, as aptly pointed out by de Carvalho et al. (2005), rather than facing these facts, some lookfor “miracle solutions” (such as molecular taxonomy, informatics or the internet) that would avoid having togo to the field and collect specimens, study them carefully in the laboratory, compare them with collections,describe them and publish their descriptions. Taxonomists and all citizens who feel concerned by biodiversityshould face the following basic problem: our societies (and especially those of Europe and North America)are ready to invest money into technologies, material equipments and products (an important part of thesefunds being in the end restituted to the stockholders of the industries producing the latter) but not into salaries(which are “lost” for the stockholders). However, what we primarily need to describe the vanishing species ofthe globe is not molecular phylogenies, online databases or workshops on the “globalization” of taxonomy,but above all manpower, i.e., permanent positions for well-trained professionals, brains and arms to do thedaily field and laboratory work in all countries of the world, well-supported museums and publications: “sys-tematics needs theoretical training, more professionals, a lasting commitment to collections, and recognitionas a robust science by peers and policy-makers, without which taxonomy itself may fall victim to extinction”

Zootaxa 1519 © 2007 Magnolia Press · 29PHYLOGENY, TAXONOMY AND NOMENCLATURE
(de Carvalho et al. 2005). According to Wilson (2004), there are at present about 6000 working taxonomistsworldwide, i.e., “a tiny slice of the biological community as a whole”, and their discipline is “one of the weak-est and most underfounded”. The least that members of this small community can do is to identify properlytheir priorities, and to devote most of their time and energy to these priorities.
However, some signs exist that this is not exactly what is going on. For example, in the last decade, manydiscussions, meetings and papers were devoted to the conception and promotion of a new conception of bio-logical nomenclature, as well as to its rebuttal. Although these controversies have had the merit to renewinterest in the topic of biological nomenclature as well as on taxonomy in general, this has already divertedthousands of working hours (i.e., collectively, months or years) of competent taxonomists, who would havebeen better inspired to use this precious time (provided by their salaries) and their energy to collect, study anddescribe the biodiversity of our planet that is getting extinct at an accelerated rate in our indifference. It ismore than time to stop this (Carpenter 2003), but this requires to provide replies to the questions and criti-cisms raised by the promoters of these new ideas. As will be shown below, whereas some of these criticismsare based on misunderstandings of the nomenclatural Rules currently in force, some others are tackling genu-ine problems, and must be addressed seriously by considering bringing changes in the current Rules.
Ranks and nominal-series in zoological nomenclature
In order to be able to deal with the millions of different kinds of organisms of our planet, we need a classifica-tion of these organisms. Although different kinds of classifications can be used in different situations (e.g., ofanimals as kind of food, as commercial items, as pets, for hunting and fishing, etc.), the most useful one,which allows the highest number of generalisations to be made, is a scientific classification or taxonomy,based on the recognition of units called taxa (singular taxon), such as those recognized in traditional taxon-omy as species, genera or families. Nowadays, taxa are recognized on the basis of our current understandingof biological evolution, and especially of hypotheses about cladistic relationships between organisms asexpressed in phylogenetic trees. The most convenient system for the designation of taxa is through scientificnames or nomina (singular nomen; Dubois 2000, 2005a), and the discipline in charge for naming taxa is callednomenclature.
In the recent years, many discussions have been devoted to the theoretical bases of biological nomencla-ture (Dubois 2005a, Polaszek & Wilson 2005) and to their practical implications on the exploration of biodi-versity. Particular attention has been given to a comparison of the pros and cons of nomenclature followingthe zoological (Anonymous 1999; “the Code” hereafter) and botanical (McNeill et al. 2006) internationalcodes, as opposed to the recent proposal of several new nomenclatural systems all called “phylogenetic” (seeDubois 2006c: 3), one of which, the Phylocode (de Queiroz & Gauthier 1990, 1994; Laurin & Cantino 2004,2006), has received the heaviest promotion and support (for references on recent debates on this complexquestion, see Dubois 2005a, 2006b-c; Kuntner & Agnarsson 2006). The present paper provides a new discus-sion of several basic nomenclatural questions, with particular emphasis on zoological nomenclature, andexamples from recent works on the taxonomy of amphibians. This discussion stems from the fact that the use-fulness of nomenclatural ranks in zoological taxonomy has recently been challenged by several authors (e.g.,Smith 1988; Sundberg & Pleijel 1994; Minelli 2000; Pleijel & Rouse 2003; Kluge 2005; Laurin 2005; Hillis2006), mostly among supporters of the Phylocode. Because they are all linked in nomenclatural theory andpractice, several points are addressed below: onomatophores, nomenclatural hierarchy and ranks, monosemyand polysemy in nomenclature, nomenclatural redundancy and parsimony, and the nomenclatural Rule ofCoordination. After study of these questions, new proposals for the improvement of the current Code are pre-sented.
In zoology, since the beginning of the 20th century a vast majority of taxonomists and almost all zoologistsand biologists have followed the Rules of the Code. The object of the latter is not to deal with the theory and

DUBOIS30 · Zootaxa 1519 © 2007 Magnolia Press
practice of classification of organisms (taxonomy), but to provide Rules for the automatic and universal estab-lishment of the unique valid nomen of a given taxon (nomenclature) in any given taxonomic frame. Thisestablishment is a three-step process that includes Rules first for the nomenclatural availability of nomina(first floor of the “nomenclatural house”; Dubois 2005a), then for their allocation to taxa (second floor) andfinally for their validity (third floor): once made nomenclaturally available through publication following cer-tain conditions, a given nomen is unambiguously allocated to a given taxon in any given taxonomy throughthe use of an onomatophore or “name-bearing type” (Dubois & Ohler 1997; Dubois 2005a); whenever,according to these Rules, two or more distinct nomina apply to the same taxon, or have the same spelling, thevalid one among these synonyms or homonyms is determined by a simple Rule, that of priority of publication.
A major characteristic of the Code is that nomina are referred to ranks and the latter to nominal-series(Dubois 2000, 2005a). Ranks are levels in the taxonomic hierarchy, such as species, genus, family or order.Nominal-series (“groups of names” in the Code) are sets of ranks: within each of these series, nomina interactconcerning coordination (and eponymy; see below), priority, synonymy and homonymy, but they do not do sobetween series (a genus nomen cannot have priority over a family nomen, and there cannot be homonymybetween them: the superfamily nomen RANOIDEA Rafinesque-Schmaltz, 1814 is not a homonym of the genusnomen Ranoidea Tschudi, 1838). Nominal-series are five in number. Three are recognized by the Code: thespecies-series (species, subspecies, groups of species and of subspecies), the genus-series (genus and subge-nus) and the family-series (family, tribe, and ranks just above and below, such as superfamily or subtribe).Two additional series can be recognized, although for the time being they are outside the Code: the class-series (phylum, class, order, and other higher ranks) and the variety-series (natio, variety, form, and otherlower ranks).
Two different kinds of ranks can be distinguished in zoological nomenclature (Dubois 2006a): key ranks(such as family, genus or species) and subsidiary ranks (such as superfamily or subfamily). Among key ranks,some (such as class or family), called primary key ranks by Dubois (2006a), are more important than second-ary key ranks (such as legio or phalanx), being more universally used than them. The most often used keyranks in zoology (Simpson 1961: 16–17; Blackwelder 1967: 220–221; Mayr & Ashlock 1991: 133) are king-dom, phylum, class and order (class-series), family and tribe (family-series), genus (genus-series) and species(species-series). The ranks variety and form (variety-series), although of normal and frequent use in botany,being recognized by the botanical code (McNeill et al. 2006), are seldom used by zoologists, being then“informal ranks” without “official” status under the Code.
Nomenclatural ranks have been in use in zoology since the book of Linnaeus (1758), which has been arbi-trarily chosen as the starting date for zoological nomenclature. At the beginning of this book, Linnaeus (1758:6–7) presented the general pattern of his taxonomic hierarchy, with six nomenclatural ranks: regnum, classis,ordo, genus, species and varietas. However, and although this has been ignored by most subsequent authors, inthe book itself he used in a few cases several additional, unnamed, ranks (Dubois, in preparation): one belowclassis and above ordo, one between ordo and genus, up to four below genus and above species, and onebetween species and varietas. Therefore, altogether, over the whole animal kingdom, Linnaeus (1758) recog-nized as many as 13 nomenclatural ranks, not only six. This shows that the idea of having a complex hierarchyof ranks, some of which are “compulsory” (the six mentioned in the introduction of the book), whereas theothers are optional and can be used when the needs appears, is as old as the “Linnaean” system itself.
Of course, the meaning of ranks has changed with the evolution of ideas in biology. Under a fixist, typo-logical thinking like that prevalent at the time of Linnaeus, ranks were meant to express a hierarchy of “noble-ness” of organisms, supposed to reflect God’s plans at the creation of the world. Nowadays, they are used toexpress hypothesized cladistic relationships among organisms, as formulated in phylogenetic trees. However,if taxonomy is to remain a useful practical tool, not all hypothetical nodes of such trees need to be named(Dubois 2006a), and this particularly applies to groups composed only or mostly of fossil taxa, many of whichare poorly known. Among named hypothetical nodes, those considered more ancestral are given higher ranks

Zootaxa 1519 © 2007 Magnolia Press · 31PHYLOGENY, TAXONOMY AND NOMENCLATURE
than those considered more recent. The exactly same ideas can be expressed in terms of hierarchical inclu-sions of members into sets (e.g., Hennig 1966: fig. 18, Dubois 2006a: fig. 3), more inclusive taxa (correspond-ing to more ancestral hypothetical nodes) being afforded higher ranks in the nomenclatural hierarchy. Strictlyspeaking, of course, “clades” are not included in one another as clades are not classes or sets of members buthistorical entities or lineages, i.e., ontologically, individuals. But the object of taxonomy is not the real cladesof the real world, which will probably remain unknown to us forever, but taxa, i.e., hypotheses, models or the-ories supposed to account for the real process of evolution (Dubois 2005a: 378). Nowadays, most taxonomistswill only recognize a taxon for a group considered to be “monophyletic” sensu Hennig (1950, 1966) necHaeckel (1866), or preferably holophyletic (Ashlock 1971; Dubois 1986, 1988; Mayr & Ashlock 1991). Forsuch a concept (holophyletic taxon), the synonymous terms phylon (Dubois 1991), cladon (Mayr 1995) andphylo-taxon (Joyce et al. 2004) have been coined. Such taxa are hypotheses about the real clades of the realworld, but not these clades themselves, that no one has ever seen or will ever see. Thus, if clades may beviewed ontologically as individuals, it is not the case of taxa, that are classes of organisms, defined by criteriasuch as characters or hypothetic cladistic relationships (Dubois 2006b-c), and whose hierarchical relationshipsmay legitimally be represented in terms of hierarchical inclusions of members into sets.
Dubois (2006a: 217) suggested that, using the traditional nomenclatural hierarchy, almost all animalorganisms of the planet (except a few poorly known ones, especially among extinct taxa) can be referred totaxa at seven primary key ranks (regnum, phylum, classis, ordo, familia, genus, species), which could thus beconsidered virtually “compulsory” for all animals, the other primary key ranks (tribus and varietas), all sec-ondary key ranks and all subsidiary ranks being then considered optional. The need to use more or less numer-ous ranks in any given zoological group is related, not directly to the number of species in the group, but to thecomplexity of the cladistic relationships hypothesized among the latter, i.e., to the quality of “resolution” ofthe cladogram, and to the symmetry of the tree (Dupuis 1979). Whenever taxonomists decide to name allnodes in their trees (which is not compulsory but is sometimes the case in recent works that deal with rathersmall groups: e.g., Frost et al. 2006), the number of ranks increases directly with the number of levels ornodes. Thus this number tends also to increase with the number of species in the group, as their mutual cladis-tic relationships become better and better “resolved” and polytomies disappear. It is therefore inevitable thatthe number of ranks be different in different zoological taxa, and this difference must be respected in our tax-onomic schemes as it tells us something about the patterns of organismic evolution (rates of speciation, spe-cies diversity, etc.; see e.g. Dubois 2004a). The current nomenclatural system with primary and secondary keyranks, which may be expanded with the use of subsidiary ranks, is excellent in this respect and should be kept.Dubois (2006a) proposed a detailed standard nomenclatural hierarchy which altogether allows for 209 poten-tial ranks, i.e., probably much more than will ever be needed, even in very species-rich zoological groups withvery detailed cladistic relationships, except if all nodes had to be named, an idea that has very few supporters,if any. In actual recent taxonomies (see e.g. Minelli 1991), only a very small proportion of these 209 potentialranks (with various denominations given to them) were indeed used, and this will probably remain the case fora long time yet, before robust detailed cladistic relationships are available at all levels in all zoological groups.
The problem of equivalence between taxa of the same nomenclatural rank
As mentioned above, some recent authors challenged the usefulness of ranks, as implemented in the Code, inzoological nomenclature. Two different criticisms were expressed in this respect: (1) the absence of equiva-lence between taxa of same rank in different taxonomies; (2) the ambiguity caused by the existence, under theRule of Coordination, of different taxa bearing the same nomen. Let us start with the first question.
It has long been known by competent zoologists that “Taxa of the same rank (genera, families, etc.) arenot equivalent, especially in different higher groups” (Bănărescu 1973: 217). A genus of birds and a genus ofinsects, for example, are not equivalent by any criterion, be it their “taxonomic size” (number of included spe-

DUBOIS32 · Zootaxa 1519 © 2007 Magnolia Press
cies), their diversity (either morphological, genetic, ecological or karyological), their geological age, etc.There exists no definition, biological or other, of most categories used in zoological taxonomy at the ranks oftribe, family, order, etc. As pointed out already (e.g., Minelli 2000, Laurin 2005), ranks have been misused inseveral works to make unwarranted comparisons, but this is not a valid reason to reject ranks (Dubois 2005a:392): “Just because some doctors misprescribe a particular drug, should other doctors be banned from pre-scribing it? Certainly not.” (Moore 2003: 13).
However, definitions do indeed exist for a few taxonomic categories, mostly at the ranks of species (andconsequently subspecies) and genus (and consequently subgenus). In the first case, numerous definitions ofthe category “species” have been proposed (see e.g. Claridge et al. 1997a; Wheeler & Meier 2000) and noagreement exists among members of the community of systematists as to which one is “best”. In contrast, inthe second case, only a few proposals were made regarding the category “genus” (Dubois 1988, 2004b), andthey have not been widely discussed so far. It is therefore quite clear that, for the time being but also probablyfor a long time still, no criterion of “equivalence” exists between taxa of same rank in different taxonomicgroups.
After a few interesting comments by Schaefer (1976), this question was explored in detail by Dubois(1988: 46–51, 59–67). Several kinds of criteria can a priori be considered as potentially useful for establish-ing such an equivalence. Most of them are biological criteria. They include species richness (or more gener-ally, at each rank, richness in taxa of just subordinate rank), or various measures of “diversity” or “similarity”,e.g., based on morphological data (“phenetic distances”), or on ecological, ethological, karyological or otherinformation, including structural (“genetic distances”) or functional genetic comparisons (e.g., through organ-ismal hybridization results; for details on the distinction between these two kinds of genetic distances, seeDubois 1988). Another approach to equivalence between taxa in different groups is not through biological, buthistorical criteria, particularly the absolute or relative ages of taxa.
As shown in detail by the analysis provided in Dubois (1988), few of the criteria just listed could be con-sidered as a basis for standardising the use of categories in zoological taxonomy, because few of them havegenerality over the whole animal kingdom. The criterion of taxonomic richness, as implemented for examplein Van Valen’s (1973) metataxonomic criterion, is an interesting one for comparing the taxonomies of differ-ent groups as it allows some inferences on the evolutionary modalities within the groups (high or low rates ofspeciation or rates of divergence between species), but it does not allow building up taxonomies (Dubois1988: 64–65). Unlike most data in taxonomy which are based on character analysis (characters being studiedindependently in each organism or taxon to be compared), hybridization data can be useful as “relational tax-onomic criteria” (Dubois 1988) or more shortly relacters (Dubois 2004b) to define taxonomic categories, butonly for a few of them. Thus, at the species level, the absence of natural hybridization in sympatry or parapa-try, and therefore of gene flow between two entities (whatever the reasons for this absence), can be used asevidence that these two entities must be treated as species under the “biological species” concept (this is anon-arbitrary taxonomic criterion for exclusion, as defined by Simpson 1961: 115). Similarly, at the genuslevel, the existence of adult hybrids, even if obtained in artificial conditions, can be used as evidence that twospecies must be placed in the same genus (non-arbitrary taxonomic criterion for inclusion). But such a crite-rion can hardly be used for higher, more inclusive categories, as the “scale” of the possible results of hybrid-ization between species is rather limited (Dubois 1988: 51). The only other important category which couldpossibly be defined in such a way across the whole of zoology could be family, if it was agreed that any twospecies liable to produce together genuine fertilized diploid eggs, even if in artificial conditions and even ifdevelopment does not proceed further, should never be placed in different families (non-arbitrary taxonomiccriterion for inclusion). But no use of this criterion for still higher categories (order, class, etc.) is possible asno hybridization can even start between too distantly related organisms.
An interesting approach to the problem of equivalence was suggested by Hennig (1936, 1950, 1966) andsupported by several subsequent authors (Kiriakoff 1954, 1965; Crowson 1970; Sibley & Ahlquist 1982,

Zootaxa 1519 © 2007 Magnolia Press · 33PHYLOGENY, TAXONOMY AND NOMENCLATURE
1990; Avise & Johns 1999; Avise & Mitchell 2007): the use of the absolute age of taxa to determine theirrank. This proposal is very promising as this criterion has more generality than others, including the hybrid-ization criterion, over both the whole animal kingdom and the whole nomenclatural hierarchy. However, forthe time being it faces two major problems that make it premature to envisage its being shortly implementedin zoological taxonomy.
The first one is that we still miss robust data on the absolute ages of many taxa. This could be in part rem-edied by using relative or approximate ages (just comparing the relative ages of some cladogeneses, withoutbeing able to date them; or just using gross stratigraphic levels and periods as landmarks for major taxonomicranks). No doubt, information about the ages of taxa will keep growing and, in the next decades, is likely tobecome much more reliable than now, thus being potentially usable to calibrate ranks over the whole animalkingdom. But then we come to the second problem, which is the “strength of tradition”. Strangely enough,although their work would seem to prepare them to finding new results and to be ready to accept novelties,scientists, and particularly evolutionary biologists and systematists, are very strongly attached to tradition. Asshown with some nice examples by Avise & Johns (1999) and Avise & Mitchell (2007), adopting the age oftaxa as a basis for allocating taxa to ranks would result in major changes in the ranks traditionally given tomany taxa (much more than implementation of the hybridization criterion for genera as proposed by Dubois1988, 2004b), and it is unlikely that most taxonomists would be willing to take this step today. They mightchange their mind in a few decades, as many more data on the ages of taxa will then be available, but, eventhen, it is unlikely that this could be obtained by an addition of individual actions. It would probably requireholding one or several large international meetings specially for this purpose.
Be it as it may, for the time being there exists no criterion universally agreed upon among zootaxonomistsregarding “equivalence” of taxa at the same nomenclatural rank. Consequently, taxa of the same rank in dif-ferent zoological groups are by no way “comparable”, and the use, for example, of numbers of genera or offamilies in comparative works of biogeography, evolution, paleontology, etc., is completely misleading andunjustified. This observation, however, which is by no way new, does not imply in the least that ranks bythemselves are “useless”, “harmful” or even “dangerous” (Minelli 2000). Any competent taxonomist “knowswell” that a genus of beetles has nothing to do with a genus of ducks, but the problem is to disseminate thisinformation outside the specialised milieu of taxonomists: rather than a problem of taxonomy, this is a prob-lem of pedagogy (Dubois 2005a, 2006b–c).
Taxonomic ranks are indeed subjective and arbitrary, and do not warrant comparisons between taxa of thesame rank in different groups. This does not mean that they should be abandoned (Moore 2003: 13; Dubois2005a). As discussed e.g. by Knox (1998), taxonomic hierarchies as reflected in nomenclatural ranks are“organizational models of relationships” that are useful for keeping track of inter-level relationships amongentities. Abandoning ranks in zoological nomenclature would make the latter much less efficient as a tool forindexation of information and for communication about taxonomies, its basic purpose. But the problem maycome in part from a recent use of ranks in some publications that is not consistent with the system of ranking,as will be shown below with two examples.
Ascribing a given rank to a taxon in a given taxonomy means that: (1) this taxon has at least one sister-taxon of same rank (more than one if the cladistic relationships are still problematic, some polytomies beingunresolved), which may be called parordinate (Dubois 2006b); (2) it is subordinate to a single taxon of justhigher rank; (3) it may be superordinate to two or more taxa of just lower rank. In any taxonomy based on aphylogenetic tree, these three pieces of information have a phylogenetic meaning, as they inform on the hier-archical relations between taxa, i.e. on the structure of the tree. They are not provided by unranked nomencla-tural systems. Of course, this information may be found in cladograms, or in taxonomies presented with asystem of indentations relative to margin, but, as convincingly discussed by Wiley (1981: 203–204), such asystem is not convenient in use whenever a taxonomy spreads over several pages, and above all it cannot beused in a text. Although many taxonomists seem to forget it sometimes, nomina of taxa are not for the private

DUBOIS34 · Zootaxa 1519 © 2007 Magnolia Press
use of systematists only, but are necessary for all users of biological material who want to designate this mate-rial briefly and unambiguously: in the course of a paper on physiology, behaviour or ecology, one cannotexpect an author to provide a cladogram or an indented taxonomy, and only nomina can be used.
The important point in ranking is not the absolute rank given to any taxon, which has no meaning byitself, but the relative ranks of the different taxa, and in particular the fact that sister-taxa in a phylogenetictaxonomy have the same rank: “In any Linnaean classification, the taxa are arranged in a nested hierarchy ofprogressively more inclusive ranks or categories. In cladistic classification, the pattern of cladistic relation-ships, usually taken to hypothesize genealogy, is the basis for ranking. The clades are recognized as taxa andtheir rank is determined by their position. More inclusive groups are ranked at higher category levels than lessinclusive groups. In its simplest form, a cladistic classification places all sister taxa at the same rank. This istotally unambiguous; the classification exactly expresses the genealogy.” (Raikow 1985: 195).
However, probably in part as a result of the recent criticisms against ranking, it is striking that these verysimple and sound principles were ignored in some publications, including recent ones. A very enlighteningone in this respect is the “amphibian tree of life” of Frost et al. (2006). In this work, parordinate taxa (taxaconsidered sister-groups according to the cladistic hypothesis adopted) are in many cases afforded differentranks, and in many cases also they belong in different nominal-series. Thus, in some cases they are both mem-bers of the family-series, but at different ranks (e.g., superfamily PYXICEPHALOIDEA and family PETROPE-DETIDAE), in other cases one of them belongs in the family-series and the other one in the genus-series (e.g.,subfamily MELANOBATRACHINAE and genus Ramanella), in many cases one of them belongs in the family-series and the other one in the class-series (e.g., family DICROGLOSSIDAE and “taxon” AGLAIOANURA), and inmany other cases they both belong in the class-series but with different endings, which suggests that they arenot parordinate (e.g., “taxon” AFRICANURA and “taxon” SAUKROBATRACHIA). Besides, although theseauthors followed the Code’s Rule of Priority for family-series and genus-series nomina, they took advantageof the absence of binding Rules in the Code for class-series nomina to apply ad hoc nomina to taxa referred toranks above superfamily, ignoring in some cases available nomina and coining their “own” nomina for someof their taxa, or using in other cases existing nomina but in a sense different from that they had in the publica-tion where they had been coined. However, no nomenclatural system can be stable if based on subsequent def-initions of existing nomina (a virtually endless process), and nomenclatural Rules in the class-series, whichare badly needed (Dubois 2005a,c, 2006a,c), ought to follow an important device of the Code in this respect,which can be called its “founder effect” (Dubois 2005b: 201): nomina are defined once and for all in the pub-lications where they are coined, and subsequent uses of “the same” nomina with different definitions amountsin fact to the creation of new nomina, junior homonyms of the original ones (Dubois 2006a). Independentlyfrom the questionable taxonomic scheme of this book, the suprageneric nomenclature used by Frost et al.(2006) opens the door for considerable chaos and will have to be corrected to become compatible with theRules of the Code (Dubois, in preparation). Frost et al. (2006: 143) “justified” this Rule-free nomenclature bythe following statement: “we follow the International Code of Zoological Nomenclature (…) for regulatedtaxa (family group and down) and apply an unranked taxonomy for unregulated taxa (above family group)”.In fact, inasmuch as they afford in many cases different ranks to taxa considered sister-taxa in their taxonomicscheme, even in the family-series their nomenclature does not follow the Code and is in fact unranked. Such anomenclatural system can be called pseudoranked, as ranks used in this system provide no information on thestructure of the tree adopted as a basis for the taxonomy, and by way of consequence on the structure of thetaxonomy itself. It would be better and more consistent if all supraspecific taxa in this taxonomy had beencalled “taxa” rather than “genera”, “subfamilies” or “families”.
Another example of pseudoranked nomenclatural system is the genus-series nomenclature used in theworks of Hillis et al. (2001) and Hillis & Wilcox (2005), respectively in their works on the salamander genusEurycea and on the frog genus Rana. As analysed in detail elsewhere (Dubois 2006b,d, 2007), these nomen-clatures are not acceptable under the Code as they recognize several taxa under the rank of subgenus although

Zootaxa 1519 © 2007 Magnolia Press · 35PHYLOGENY, TAXONOMY AND NOMENCLATURE
they are included in one another. Against all evidence, Hillis (2006) argued that he considered such a systemacceptable under the Code because this is, according to him, a matter of “interpretation” of the latter. How-ever, he still did not make a clear distinction between availability and validity of nomina, i.e., between floors1 and 3 of the “nomenclatural house” (Dubois 2005a,c). As explained in detail in Dubois (2006b,d), one firstpoint is that, regarding availability, following Article 10.4 of the Code (in its Chapter 4, “Criteria of availabil-ity”), the nomen of any genus-series division of a genus, whatever its original designation, is deemed to havebeen created at the subgenus rank. But this does not mean that, regarding validity, several subgenera can be“nested hierarchically within one another”. This would be equivalent to stating that several genera can benested hierarchically within one another, or several species within one another, several families within oneanother, etc., a course which would be completely contradictory with the hierarchical structure of zoologicalnomenclature as implemented in the Code. Under the Code, if a family includes two or more subordinate fam-ily-series taxa, these must have the rank of subfamily, then the rank below is tribe, then subtribe, etc., withoutlimitation. Particular problems exist in the genus-series and in the species-series of nomina because of therestriction in the number of ranks allowed by the Code in these nominal-series (Dubois, 2006b), but Hillis etal. (2001) and Hillis & Wilcox (2005) ignored these problems. Hillis (2006) was still not convinced by myanalysis, and wrote: “Readers are welcome to read the rules and decide for themselves which interpretation iscorrect.” This is easy, as no problem of “interpretation” exists, as explained in detail elsewhere (Dubois 2007:
6)1.If these papers, by prominent phylogeneticists and taxonomists, were to be taken as examples by other
colleagues, pseudoranked nomenclatures could become more and more frequent, and this would be a consid-erable source of confusion for communication between taxonomists, and, above all (and more importantly)between the latter and the rest of the community of biologists, and even non-biologists. In a way, pseudo-ranked nomenclatures may be more confusing than unranked nomenclatures like the Phylocode, which at leastare consistent as not including ranks in the definition of nomina (see e.g. comparisons of a Phylocode nomen-clature with a pseudoranked one and a nomenclature respecting the Code, for the same taxonomy of North-American ranid frogs, in Dubois 2006b,d). As some authors still pretend that “interpretation problems” doexist, the Code should be modified in order to fully clarify this point: it should state in full words that nomen-clatural ranks are (and are only) hierarchical levels in a hierarchy, that parordinate taxa should always have thesame rank and that no two taxa can be given the same nomenclatural rank if included into one another. Thisquestion is addressed again below.
The confusion between nomenclatural ranks and taxonomic categories
Most of the problems discussed above derive, as many other ones in systematics (Dubois 2005a), from confu-sion between the fields of taxonomy (recognizing or diagnosing taxa) and nomenclature (naming taxa). Thebasic confusion at stake here is between the nomenclatural concept of nomenclatural rank and the taxonomic
1. The paper by Hillis (2006) contains many other confusions, misinterpretations and misleading statements, but my
reply to this paper was rejected by Derek E. Wildman, Associate Editor of the journal Molecular Phylogenetics & Evolu-
tion, without having been sent out to referees, for the following reason: “The manuscript is mostly about nomenclature,
and as such I feel it is not entirely appropriate for the journal Molecular Phylogenetics & Evolution. (…) While I agree
that differences and misunderstandings surrounding ‘The Code’ and ‘Phylocode’ can cause confusion and misunderstan-
ding in classification and taxonomy, I also feel that debates regarding these differences are better suited to nomenclature
journals.” (18 September 2006). [Which are these journals is unclear to me]. Thus my reply (Dubois 2007) had to be
published elsewhere, and readers of MPE will continue to have misleading information about some basic Rules and con-
cepts of the Code.

DUBOIS36 · Zootaxa 1519 © 2007 Magnolia Press
concept of taxonomic category. This confusion is well illustrated by the use in some recent publications (e.g.,Kluge 2005, de Queiroz 2006) of the fully ambiguous formula “categorical ranks”. However, it is true that theclear distinction between category and rank was made for the first time only very recently, although briefly, inDubois (2005a: 412–413, 2006a: 219–220).
A nomenclatural rank is “nothing but” a place in a hierarchy of taxa included into one another, or subordi-nate to one another, in particular in order to express a hypothesis of cladistic relationships. It does not tell any-thing more about the particularities, qualities or history of these taxa.
A taxonomic category is a class of taxa that are defined by some criteria which may be biological (e.g.,panmixy or hybridizability) or historical (e.g., absolute age), independently from their cladistic relationships.
Any given taxon may be referred both to a nomenclatural rank and to a taxonomic category, but the twoprocesses are distinct. In higher-ranked taxonomy, as discussed above, no taxonomic category has ever beendefined, but such categories have been in use at lower nomenclatural levels, especially at genus and specieslevel and even below. In such cases, categories do not correspond to hierarchically superordinate, parordinateor subordinate ranks, but to alternative categories of the same rank in the nomenclatural hierarchy.
This distinction is particularly clear at the species level. The endless so-called “species problem” of sys-tematics has several dimensions. Although it is possible to list as many as 92 (and probably ten times more)different “definitions of species” (Lherminier & Solignac 2000) or to recognize 22 “species concepts” (May-den 1997), many of these “definitions” or “concepts” are largely or partly equivalent, and the number can bereduced to a few major groups. But, first of all, it is crucial to distinguish between several fully distinct mean-ings of the term “species”. Mayr (1997: 133) recognized three major meanings of this term: “The word ‘spe-cies’ is applied to three very different objects or phenomena: (1) the species concept; (2) the species category,and (3) species taxa. Endless confusion in the literature has resulted from the failure of some authors to dis-criminate among these three very different meanings of the word ‘species’ (...) The species category is a par-ticular rank in the Linnaean hierarchy”. These distinctions are useful, but insufficient, as the confusionbetween nomenclatural rank and taxonomic category still persists.
As a nomenclatural rank, the species is the “brick” of zoological taxonomy. All living organisms can andmust be referred to a taxon of species rank, whatever their reproductive mode or other biological characteris-tics. Such taxa of species rank are indispensable to our society as a whole, as they are those which are listed inmany official documents used in domains sometimes very far away from biological taxonomy, such as redlists for biological conservation, commercial documents, juridical texts, custom forms, etc. On the other hand,several concepts of species as a taxonomic category have been proposed and are defended by different taxon-omists or schools of taxonomists. This diversity of “species concepts” (e.g., Claridge et al. 1997a, Wheeler &Meier 2000) is in part artificial, as several of these concepts can be united into more general ones (Mayr &Ashlock 1991: 24–28), but it also reflects a double reality.
First, it reflects the existence of different schools of thought in taxonomy. It is not, and should not be, thepurpose of the Code to choose one of these schools against the other ones. As stated in the Preamble of theCode, the latter is not aiming at restricting the freedom of taxonomic thought or actions. Although everyauthor argues that his concept is “best” (Cracraft 2000), the debate will probably rage still for long. A basicassumption of many participants to this debate is very strange from the part of people whose major study fieldis the diversity of organisms. This assumption clearly does not belong in the comparative and evolutionarymode of thought, and comes rather from “reductionist” biology: it is the idea that there should be a single con-
cept of species (as a category)1. However, there is no doubt that different kinds of biological entities do existin nature, corresponding to different modes of speciation, of reproductive mode, of evolutionary pattern, etc.
1. For a long time, the same reductionist attitude resulted in most evolutionary biologists seeking the mode of specia-
tion, although it is now clear that there is not one, but several modes of speciation (e.g., White 1978, Barigozzi 1982,
Hubbell 2001).

Zootaxa 1519 © 2007 Magnolia Press · 37PHYLOGENY, TAXONOMY AND NOMENCLATURE
This is the second reality behind the existence of several species concepts. Except if one wants to “force” rea-lity into an artificial unitary, reductionist concept, one has to accept that different kinds of basic evolutionaryentities do exist in nature and that different taxonomic concepts should be applied to them. The aim of a uni-tary taxonomic concept may be wrong altogether, and at any rate, no consensus exists nowadays among prac-ticing systematists as to the possibility to agree even on a “working concept”. For the time being, the practical(and not necessarily temporary) solution to this problem may be to use different taxonomic concepts of spe-cies in different taxonomic groups and particularly in different evolutionary situations (Claridge et al. 1997b).As a matter of fact, widely different evolutionary situations can be distinguished and are worth being recogni-zed taxonomically.
Thus, Bernardi (1956, 1957, 1980) provided very useful reviews of the concepts proposed to account fordifferent “kinds” of “species” and of infraspecific taxa, which he called “evolutionary taxonomic” categories.Additional data in this respect can be found in many other works (e.g., Vuilleumier 1976, Eck 1977, Dubois &Günther 1982, Haffer 1986, Dubois 1991). Some of these categories are designated by terms including theterm species combined with modifiers (e.g., quasispecies), some by different terms. Let us consider brieflysome of these categories.
The well-known category superspecies, for example, is a supraspecific but infrageneric category thatgroups species of recent origin, having a parapatric distribution and that are considered vicariant (Bernardi1980). Such species can be called prospecies (also sometimes semispecies, allospecies, etc.). The taxonomiccategory prospecies is nomenclaturally referred to the nomenclatural rank species, but the functions of bothallocations are distinct: the nomenclatural rank refers to a place in a hierarchy of taxa, whereas the taxonomiccategory refers to some (in this case, evolutionary and distributional) properties of these entities. Any taxon ofthe category superspecies, being superspecific and infrageneric, can nomenclaturally be recognized either as ataxon of the rank subgenus, or, preferably, of a supraspecific rank in the species-series, called “aggregate ofspecies” in the Code, and supraspecies by Génermont & Lamotte (1980) and Dubois (2006a-b).
Similarly, dualspecies (or cryptic species, sibling species, etc.) are “young” species that are morphologi-cally indistinguishable or very difficult to recognize visually (at least by man), but that can be told by othercharacters (molecular, karyological, bioacoustic, etc.). A group of several dualspecies can be recognized as asupraspecific infrageneric taxon called ultraspecies (Bernardi 1980). The category dualspecies is nomencla-turally referred to the rank species, whereas the category ultraspecies is supraspecific, being preferablyreferred to the rank supraspecies.
Some taxonomic categories designate entities that do not reproduce through normal meiosis (with recom-bination between parental genomes) and free panmictic breeding among members of the population. Someauthors (e.g., Mayden 1997) use the general term agamospecies for all such entities, but this is confusing as itindiscriminately covers widely different situations. It is clearer to distinguish the latter by recognizing severaldistinct taxonomic categories. Thus, Dubois & Günther (1982) proposed the term klepton (emended tokleptospecies by Mayr & Ashlock 1991: 33) for an evolutionary taxonomic category accomodating taxa ofhybrid origin and with special meiosis and reproductive mode, where the taxon can reproduce only throughsexual parasitism on another taxon. Kleptons are “particular kinds of species” that are of the nomenclaturalspecies rank. The taxonomic category synklepton designates supraspecific taxa including one or several klep-ton(s) and two or more “normal” species that are involved in genetic interactions with the klepton(s). Dubois(1991) distinguished between gynokleptons and zygokleptons, and introduced the term klonon for taxa withclonal transmission of genome, including both “species” with parasexual clonal reproduction (e.g., partheno-genesis, self-fertilization) and “species” with truly asexual reproduction (e.g., vegetative reproduction).
Evolutionary taxonomic categories can also be recognized between the nomenclatural ranks of speciesand subspecies, e.g., quasispecies, vicespecies or exerge (see Bernardi 1957, 1980), and between subspecies,e.g. morpha or cline (see Bernardi 1957).
In the other direction, going towards higher, more inclusive, taxa, it is possible to recognize a taxonomic

DUBOIS38 · Zootaxa 1519 © 2007 Magnolia Press
category for taxa that group together species liable to produce hybrids, either in natural or in artificial condi-tions. This category has been called coenospecies (Turesson, 1922) or syngameon, sensu Cuénot & Tétry(1951: 455) nec Lotsy (1918) (see Bernardi 1980: 396, 398). It can be arbitrarily decided to make this taxo-nomic category coincide with the nomenclatural rank of genus, for reasons explained by Dubois (1988). Onecould also contemplate the recognition of a taxonomic category for taxa composed of species liable to pro-duce fertilized eggs, even in artificial conditions and even if development does not proceed further, and tomake this category coincide with a major primary nomenclatural rank, e.g., family. As discussed above, suchdecisions could provide partial equivalence between taxa at these ranks, but this would be limited in effect, asonly positive results could be used for this purpose, for reasons explained in detail by Dubois (1988: 65, 73),so in many cases the criterion could not be used at all.
In sum, all examples of taxonomic categories briefly mentioned above, such as prospecies, dualspecies,klepton, klonon, superspecies or coenospecies, are categories defined by biological criteria, but they provideno information on the place of the taxa so recognized in the tree of hypothetic cladistic relationships. The lat-ter information is provided by the nomenclatural ranks of the taxa, which reflect the hierarchical structure ofthe taxonomy. At higher taxonomical levels, as no common criterion allows to provide a biological definitionof taxonomic categories, only nomenclatural ranks can be used, but the situation is more complex at lowerlevels, genus and above all species. For taxa referred to the nomenclatural species- and variety-series, nomen-clatural rank and taxonomic category should be better used to play distinct roles. Ranks provide informationon the hierarchical structure of the taxonomy, whereas categories may provide information on the kind oftaxon recognized. Being distinct, these two functions can be dissociated. For many purposes, it is only neces-sary that organisms be referred to a taxon of a given rank, whatever its special biological properties. Thus, forcommercial, medical, agricultural, legal or conservation biology purposes, it is necessary that all organisms bereferred to a taxon of the species rank or in some cases of a lower rank. This is distinct from the fact that someof these organisms may be referred to “particular” kinds of species, of infraspecific or of supraspecific taxa.Taxa at the same nomenclatural rank may be referred to different taxonomic categories.
The usefulness of nomenclatural ranks in taxonomy
Taxonomy needs nomenclatural ranks. Ranked taxonomies are convenient systems of storage and retrieval oftaxonomic information, which avoid having to provide a phylogenetic tree or an indented taxonomic list everytime one wishes to write or speak about taxa. Ranks do not provide any information on the biological proper-ties of the organisms placed in any given taxa, but they provide hierarchised information on the mutual rela-tionships between the latter within the frame of a given taxonomy. In a phylogenetic taxonomy, nomenclaturalsubordination, parordination and superordination reflect the structure of the tree, but this requires that two sis-ter-group (parordinate) taxa be afforded the same nomenclatural rank, and that superordinate taxa have ahigher rank and subordinate taxa a lower rank.
Writing that a taxon is a subfamily is stating that it belongs in a more inclusive taxon of rank family andonly one (immediately superordinate taxon), that at least one taxon of same rank exists, which is supposed tobe its sister-taxon (parordinate taxon), and that it includes, or may include (for taxa at species rank) subordi-nate taxa. In the nomenclatural family-series, this information (rank) is provided by the ending of the nomen(RANIDAE, RANINAE), and in the other two nominal-series recognized by the Code, it is provided by the ony-morph (see Dubois 2000): Rana (Rana), Rana temporaria temporaria.
In contrast, nomenclatural ranks do not provide any information on the biological properties of taxa, or onthe “nature” or “importance” of the differences or divergences between them. They do not allow any compar-ison between taxa but they allow comparisons within taxa: it is for example meaningful to compare the geneticor morphological diversity within the same family in two different ecosystems.

Zootaxa 1519 © 2007 Magnolia Press · 39PHYLOGENY, TAXONOMY AND NOMENCLATURE
If the usefulness of nomenclatural ranks is recognized, then the Code should not impose any limitation totheir use. Such a limitation could legitimately be considered to restrict the freedom of taxonomic thought oraction—well, this is exactly what the Code does. The current Code excludes from its domain all nominaabove the superfamily rank and below the subspecies rank, and it limits the number of ranks that can be usedin the genus- and species-series of nomina. There is no theoretical reason for limiting this hierarchy of nomen-clatural ranks either upwards, downwards or inwards. Furthermore, this unjustified limitation places the Codein a situation of weakness relatively to other alternative nomenclatural systems (e.g., Papavero et al. 2001,Kluge 2005, Laurin & Cantino 2006) which do not have these problems and which allow to name all nodes ofthe “tree of life”.
For this reason, it seems necessary and urgent to consider the need for updating the Code by removing allits current limitations regarding the number of ranks “allowed” and “covered” by this text. This would includeincorporation into the Code of the nomina of higher taxa (above superfamily) (Dubois 2006a,c), recognitionby the Code of the nomina of lower taxa (below subspecies) (Dubois 2006a–b,d) and elimination of limita-tions regarding the number of ranks allowed in the genus- and species-series (Dubois 2006a–b,d). Thisincrease in the number of potential ranks would be useful for taxonomic purposes, as it would allow a moredetailed taxonomic expression of phylogenetic hypotheses at all levels, including at low levels for phylogeo-graphic analyses (Dubois 2006b). It could also be useful in other domains, especially in conservation biology:in zoology, unlike in botany, the Code forbids naming taxa below the subspecies rank, which precludes plac-ing them on red lists or other documents dealing with conservation biology (Dubois 2006b). For all these rea-sons, Dubois (2006a–b) proposed a standard hierarchy for ranks in zoological nomenclature, including 19 keyranks (9 primary and 10 secondary) and 10 subsidiary ranks (5 above and 5 below each key rank), thus provid-ing in the end 209 potential ranks. As mentioned above, this is probably much more than any zoological tax-onomy will ever need, even in very species-rich and rank-rich groups.
Clarity and ambiguity in the meaning of nomina
In the recent decades, a myth has developed among taxonomists, probably as a result of the “pressure” of“society”, or more exactly of some “users” of nomina: this is the myth of taxonomic and nomenclatural stabil-ity. As a matter of fact, some of these “users” (biologists of various disciplines, conservationists, administra-tions, etc.) complain about the seemingly frequent changes of nomina which affect many taxonomic groups.They tend to blame “nomenclatural rules” for this instability and they therefore demand “official”, and, if pos-sible, “closed” and “final”, lists of nomina, i.e., a “stable canonized system” of taxonomy and nomenclature(Reif 2003: 135). However, despite a persistent legend, the responsibility for most of these changes lies notwith the Code, but with the progress in phylogenetic and taxonomic research. Today, taxonomists have col-lected, studied, described and named only 10–20 %, or even perhaps much less, of the living species of ourplanet (see Dubois 2003, and references therein), and, by way of consequence, our knowledge of their phylo-genetic relationships is still very preliminary. Among the factors of error in cladistic analysis, incomplete spe-cies sampling has a major influence (e.g., Lecointre et al. 1993), and it is therefore no surprise if, in mostcases, when a new cladistic analysis of a group previously studied is carried out, the results are slightly togrossly different. In many cases, these new results require changes in our taxonomic schemes and, by way ofinevitable consequence, in the nomina given to organisms. This has nothing surprising or shocking for scien-tists who know that knowledge is always increasing and changing. The request for taxonomic and nomencla-tural stability is not a scientific one. As claimed already by several authors (Gaffney 1977, 1979; Dominguez& Wheeler 1997; Benton 2000; Dubois 2005a; Padial & De la Riva 2006), “taxonomic stability is ignorance”,and few researchers in any scientific field would be ready to accept freezing of research in order to preserveignorance. Given the current level of our ignorance in taxonomy (the taxonomic impediment), the promulga-

DUBOIS40 · Zootaxa 1519 © 2007 Magnolia Press
tion today of “definitive lists” of taxa and nomina could be done only at the expense of stopping all research insystematics (Dubois 1998). The rate of mistakes usually considered acceptable in science is below 5 %, and inno other research field our society would be satisfied with a percentage of missing data of 80–90 %. The dutyof every taxonomist or evolutionary biologist would therefore appear to do all efforts to pass this message toour society. The misleading idea that “final lists of species” or “final taxonomies” are or should be available,and that, in order not to disturb the peace of mind of non-taxonomist users of data on biodiversity (including“conservation biologists”), no significant changes should be brought to the classifications and nomina, is notdoing a service to the discipline of taxonomy and to our knowledge of our planet’s biodiversity: “Rather thantrying to comply with the requests for ‘final lists’ that are often presented to them, taxonomists should explainthe reasons for this instability, and should try and convince our ‘social partners’ that, rather than asking for a‘freezing’ of the scientific activity of exploration and analysis of biodiversity, they should support and encour-age it. In the long run, it may prove more interesting and useful to better understand the biodiversity on ourplanet than to have ‘final’ and ‘stable’, i.e. wrong and incomplete, lists of this biodiversity for the peace ofminds of administrators and technocrats.” (Dubois 1998: 22). Whatever we may wish or regret, nomenclaturalstability could proceed only from a taxonomic stability which is still in limbo. In the meanwhile, the alloca-tion of a nomen to an organism cannot proceed from the nomenclatural Rules themselves: it can result onlyfrom the taxonomy adopted. As Rieppel (2006: 186) put it, “the composition of the taxon will naturally deter-mine the meaning of its name”.
A recent avatar of the request for taxonomic and nomenclatural stability is the plea for a “Google taxon-omy” well exemplified in Hillis’s (2006) paper. It has two dimensions. The first one is to claim that, in ordernot to upset users of online databases, taxa and nomina should not be changed. This statement, which onceagain relies on a confusion between taxonomy and nomenclature, deserves comment, especially as the philos-ophy behind it is likely to carry a very misleading message, not only to taxonomists, but also to other biolo-gists and to society as a whole.
In his rebuttal to my paper on his use (Hillis & Wilcox 2005) of infrageneric ranks in the genus Rana(Dubois 2006d), Hillis (2006) stated that the decision of Frost et al. (2006) to raise the previous subgenusRana (Lithobates) to the rank of genus Lithobates was unacceptable because it would confuse users of Googleor other search engines, as a search on Google allowed him to find many uses of the nomen Rana and few ofthe nomen Lithobates. However, the decision whether Rana and Lithobates should be treated as genera or sub-genera is a taxonomic one, which has nothing to do with the nomenclatural Rules. Because of the need of“freedom of taxonomic thought or actions”, such a decision cannot be imposed by any kind of Rules but mustbe left to the personal appreciation of taxonomists and to free discussions among them. In the long run, as hasalways been the case in the history of taxonomy, a consensus will no doubt appear among practitioners of thetaxonomy of any given group. This matter should be left in the hands of specialists, not of laymen outside thediscipline of taxonomy.
In this respect, Hillis (2006) proposed a very strange, and in my opinion very dangerous, approach, hisplea for “Google taxonomy”. He suggested that taxonomists should not change the nomina of taxa in order notto upset users of electronic databases such as Google. This suggestion was discussed in detail elsewhere(Dubois 2007: 11). In the future, the imperialism of “Google thinking” may possibly try to impose a freezingof science in order not to disturb “Google users”, and such a “manufacturing of consent” would probablyplease those who think that science is too costly for our society and that ongoing research should be replacedby a “definitive truth”, i.e., a dogma. But this would soon have such a destructive effect on knowledge andaction that this practice would soon have to be abandoned. Nevertheless, scientists should remain vigilantabout the possibility of such drifts.
The second aspect of Hillis’s (2006) proposals is that he thinks nomenclatural Rules should be changed inorder to avoid different taxa bearing the same nomen, with a seemingly simple idea in mind, which could beformulated: “one taxon, one nomen”. For the time being, in zoological nomenclature as regulated by the

Zootaxa 1519 © 2007 Magnolia Press · 41PHYLOGENY, TAXONOMY AND NOMENCLATURE
Code, the Rule of Coordination requires that, as soon as several coordinate taxa are referred to the same nom-inal-series (family-, genus- or species-series), then one of the subordinate taxa bears the same nomen (eitheridentical, or slightly modified in ending, in the family-series) as the superordinate one. The Code calls suchsubordinate taxa “nominotypical taxa”. Dubois (2006b: 828) called eponymy the relationships between suchnomina. There are two kinds of eponyms: hyponyms are subordinate, epinyms are superordinate. Without men-tioning these terms, Hillis (2006) claimed that eponymy should be abandoned as it is a source of confusion forGoogle customers, who will not distinguish between “genus Rana” and “subgenus Rana”.
Despite Hillis’s (2006) statements, the Rule of Coordination, and its correlate eponymy, has severaladvantages. First, it allows “nomenclatural parsimony”, as it requires many less nomina to account for thesame taxonomy than a system like the Phylocode which requires one distinct nomen for each taxon (Dubois,2006b). Second, this system is “self-obvious” and “mnemonic” in the sense that eponyms clearly refer to eachother: there is no need to know a zoological group and its taxonomy to understand that, whenever there aretwo families, each of which includes two subfamilies, the hyponymous subfamily of each family, togetherwith its parordinate subfamily, will belong in its epinymous family. Taxonomists familiar with the Code andits philosophy know that the function of nomina is not to provide by themselves information on the taxa theydesignate, but just to be labels pointing to these taxa: they therefore have no special feelings about the fact thatsome of these taxa bear “the same nomen”, as long as no ambiguity exists on the meaning of each nomen inthe frame of the nomenclatural hierarchy.
However, there is indeed a basic disagreement here. Borrowing a terminology used in linguistics andgrammar, the nomenclatural system of the Code can be described as polysemic (from the Greek roots poly,“many”, and sema, “sign”): in such a system, the same word can have several meaning. In contrast, in amonosemic (from the Greek roots mono, “many”, and sema, “sign”) system, each word has only one meaning.
Have we already entered the era of “Google taxonomy”? If so, to shift from a polysemic to a monosemicnomenclatural system, we will have to change a large proportion of all existing zoological nomina becausethey are eponyms. Contrary to a frequent, but wrong, statement, repeated by Hillis (2006), eponymy is not aconsequence of the existence of ranks by themselves, but of the existence of nominal-series (which are morecomprehensive than ranks), within which a Rule of Coordination is in force. The theoretical possibility indeedexists of a nomenclature following the basic principles of the Code, i.e., the principles of onomatophores andof priority (the valid nomen of a taxon being the oldest available one among those the onomatophore of whichis included in the taxon), but monosemic, working without ranks and nominal-series. This possibility wasexemplified in the Rules proposed by Dubois (2005b, 2006a) for the nomenclature of higher-ranked taxa. Asdiscussed in details in Dubois (2006c), the need and possibility to use, or not, nominal-series and eponyms,depends on the system adopted for the allocation of nomina to taxa. Dubois (2006c) recognized and describedthree major such systems and several subsystems, which may be briefly surveyed again here, particularly asregards their compatibility with monosemic and polysemic nomenclatures.
Systems of allocation of nomina to taxa, monosemy and polysemy
Extensional definitions of nomina
A nomen may be defined by the extension (or circumscription, or content) of the taxon to which it is meant toapply. In such a system, the definition refers to a list of members of the taxon (inclusive extension) or of non-members excluded from the taxon (exclusive extension). Extension may be closed (unchangeable) or open(liable to be modified, by addition or subtraction of members).
(1) Closed extension is a very rigid system that would not meet the needs of zoological nomenclature, asany addition or subtraction of even a single specimen would result in the need to create a new nomen for the

DUBOIS42 · Zootaxa 1519 © 2007 Magnolia Press
taxon (Dubois 2005a). Such a system would be useful in taxonomy, at least in one group or a few, only if weknew all its/their organisms, a situation from which we are currently incredibly far over most of zoology. Itcan work for some entities, e.g., the planets of the solar system (see Ohler 2005), but not for living organismsmost of which are still unknown to us.
A system of closed extension is of course monosemic, even caricaturally so: this system is so rigid that itcannot move in the least, so that, applied to biological taxonomy, such a system would be condemned to paral-ysis and death.
(2) In a system of definition of nomina through open extension, the list of members (or non-members) ofthe taxon may be modified. Any such system allows to have several distinct taxa, possibly coordinate, desig-nated by the same nomen, and the rule “one taxon, one nomen” is therefore not respected.
In this system, the list of members (or of non-members) of the taxon can be emended in two differentways.
(a) If this occurs by addition of members to the taxon (e.g., new subordinate taxa which were previouslyunknown), there is not necessarily a change in the intension of the taxon (see below), and there is nothingshocking, in zoological taxonomy, if the nomen is not changed. As a matter of fact, pushed to its extreme (i.e.,with an original extension that can be limited to one member), this system is equivalent to that of ostension(see below).
(b) However, if subtraction of members from the taxon is considered, the situation is more complex.Members may be withdrawn from the group because the information we had previously about them waswrong (because of incorrect observations or of incomplete data): in this case, there may be no modification ofthe intension, and it is normal to keep the same nomen. But members may be also removed because a taxono-mist has changed his/her opinion, or disagrees with previous authors. In such a case, the intension of the taxonis modified and it is not justified to keep the same nomen.
In conclusion, the only nomenclatural system based on extensional definitions of nomina which seemsuseful in zoological taxonomy is that of open extension with the possibility to increase membership in thetaxon without changing the nomen. This system then amounts to ostension and it will be discussed below.
Intensional definitions of nomina
Definition of nomina by intension is what is often understood by the word “definition”: it consists in a state-ment of some of the properties (or characteristics, or specificities) supposedly shared by the members of thetaxon designated by this nomen. Such a system is in all cases monosemic. In zoological taxonomy, two majorkinds of intensions have been used to define nomina.
(1) Phenetic intension consists in a list of character states that are considered diagnostic of a taxon. In tra-ditional taxonomy, a diagnosis is a particular combination of character states that is considered to occur onlyin members of this group, and in no non-member of the group. As long as it is not associated with a phyloge-netic hypothesis of relationships between members of the taxon and non-members, it is an essentialistic, non-evolutionary concept, which proves inefficient to deal with phenomena of homoplasy, convergence and paral-lel evolution, and it is therefore not appropriate for a taxonomy taking the fact of evolution into account.
(2) In contrast, phylogenetic or cladistic intension refers to a hypothesis of relationships. Following thepaper of de Queiroz & Gauthier (1990), an important theoretical reflection on the way phylogenetic defini-tions can be formulated was carried out, especially by supporters of the project of Phylocode, and the resultsof this work are summarized in Article 9.4.1 of the draft Phylocode ([http://www.ohiou.edu/phylocode]). Thisclarification is important, and might remain in the future as the major contribution of the “Phylocode revolu-tion” to the theory of taxonomy. There are two major kinds of phylogenetic intensions to define taxa (whichmay be combined, but are distinct).

Zootaxa 1519 © 2007 Magnolia Press · 43PHYLOGENY, TAXONOMY AND NOMENCLATURE
The first kind of phylogenetic definition is based, like in the preceding case, on characters: the intensionis then expressed as a list of character states that are considered apomorphies supporting the hypothesis thatthis taxon is holophyletic. Such a list of character states can be known as an apognosis (Dubois 1997, 2006d).In a phylogenetic taxonomy or cladonomy (Brummitt 1997; Dubois 1997), a taxon may be defined as the mostinclusive holophyletic group whose members share a given apognosis. This is equivalent to an “apomorphy-based definition” as defined in Article 9.4.1 of the draft Phylocode.
Phylogenetic intension, however, can do without characters. Such a definition may rely only upon phylo-genetic hypotheses, without mentioning the kind of information or analysis used to reach the latter (whichultimately also relies on characters). Such an intensional definition of taxon based on a tree but not directly oncharacters may be called a cladognosis (plural cladognoses; from Greek klados, “branch” and gignosko, “Iknow”), a term shorter than “phylogenetic definition” and similar in aspect to diagnosis and apognosis. Arti-cle 9.4.1 of the draft Phylocode recognizes two major kinds of cladognoses, so-called “node-based defini-tions” and “branch-based definitions”, and two other derived ones.
The Appendix below provides an example showing in detail how, within the frame of the Code, the sametaxon can be defined by a diagnosis, an apognosis and a cladognosis. It is however important to stress that, inthis example, these definitions point to the taxa, not to the nomina, as the latter under the Code are attached tothese taxa by the system of ostension described below.
Although strongly promoted by supporters of the Phylocode project or other systems of “phylogeneticnomenclature”, the idea of defining nomina by phylogenetic intension is certainly not a good one, for severalreasons. The first one is that this is tied to a philosophy of taxonomy. Thus, Article 9.4.2 of the draft Phylo-code states: “The application of a phylogenetic definition, and thus also a phylogenetically defined cladename, requires an [sic] hypothesized phylogeny”. Therefore, nomenclatural rules based on phylogeneticintensional definitions of nomina do not respect the “freedom of taxonomic thought or actions” which theCode claims to respect. This is not a problem as long as most taxonomists agree that the taxonomy of livingorganisms should be based on hypotheses of cladistic relationships between them, but it might become a realdifficulty if our ideas about taxonomy were to change, e.g., if the “tree of life” paradigm had to be abandoned– for example, for not being universal, as it does not apply to the organisms between which “lateral gene trans-fer” plays a major role (see Doolittle 1999): nomina created or re-defined with cladognoses would becomeunavailable under a different taxonomic paradigm.
The major strength of the nomenclatural rules of the Code is that they are theory-free regarding taxonomy(Dubois 2007): the Code does not prescribe the use of any kind of definitions for taxa, so that the latter can bedefined in many ways, phenetically, phylogenetically, or other. The Code only provides Rules for automati-cally naming taxa once they have been defined by taxonomists, through a process of ostensional pointing toonomatophores that is discussed in more details below. Nomina created under a given taxonomic paradigmcan still be used even after taxonomic concepts and taxonomies have drastically changed: this is the case ofmost nomina currently used in taxonomy with definitions of taxa relying on phylogeny, although these nom-ina had been proposed under a typological or phenetic taxonomic paradigm. This is all the more justified thatin many cases, although the “definitions” (intensions) of the taxa have changed, the contents (extensions) ofthe latter have remained virtually the same. In contrast, under the Phylocode, the definitions of taxa and nom-ina are contemporaneous. Both are qualified by “phylogenetic definitions”. Such definitions rely on “specifi-ers”, which are not equivalent to onomatophores in two distinct respects.
Any nomenclatural system defining nomina through phylogenetic intension but without mentioning spec-imens is disconnected from the real world of organisms, being a purely circular system “inside language”dealing with abstractions (intensions), relying only on concepts and hypotheses. In contrast, the traditionalsystem of the Code has a strong connection with organisms through the use of onomatophores which areeither directly or indirectly based on specimens (Dubois & Ohler 1997, Dubois 2005a, Pickett 2005). Thesespecimens have a basic function in biology, that of providing a system for objective and unambiguous alloca-

DUBOIS44 · Zootaxa 1519 © 2007 Magnolia Press
tion of nomina to taxa, thus allowing universality and stability in the use of nomina in scientific publications.Under the Phylocode, although specimens may in some cases be used as specifiers to help defining the nom-ina, this is not compulsory as, under this system, a specifier can be “a species, specimen, or apomorphy citedin a phylogenetic definition of a name as a reference point that serves to specify the clade to which the nameapplies” (Glossary of the project of Phylocode), with the following definition given for apomorphy: “Aderived character state; a new feature that arose during the course of evolution”. Apomorphies are concepts,hypotheses based on a cladistic analysis, and using concepts as a way to define nomina sounds like a reversalto the early days of taxonomy, when “the method of concepts” or “conceptual types” was used instead of “themethod of types” or “nomenclatural types” (Moore 1998, Dubois 2005a). Even in the case when specifiers arespecimens, and even if the Phylocode decided to make such specimens compulsory for the definition of nom-ina, specifiers are not equivalent to onomatophores, for the very reason discussed above, i.e., that the use ofonomatophores is theory-free, not that of specifiers.
Under the Code, ostensional pointing to onomatophores only requires that an onomatophore be referred toa taxon (i.e., included in it) to allocate the nomen based on this onomatophore to this taxon. In the Phylocodesystem, an additional condition is required to allocate a nomen to a taxon, and this is the existence of a “phy-logenetic hypothesis” or speculations regarding the relationships between taxa. This has an important conse-quence, that has been pointed out repeatedly (Lidén et al. 1997, Anonymous 2005, Dubois 2005a, Frost et al.2006). If new organisms are discovered, or significant (although not necessarily very important) changes arebrought to phylogenetic hypotheses or speculations, a widespread and well-known nomen may have to shiftfrom its traditional taxon to another one. The discovery of a single new species, basal to a clade, may requirerenaming the latter (Anonymous 2005). More generally, if allocation of a nomen N to a taxon A + (B + C) isgoverned by the definition “the ancestor of A and B and all of its decendants”, and if new data suggest the newphylogenetic hypothesis C + (A + B), the nomen N has to apply to a taxon including A + B and excluding C,even though the content of the taxon A + B + C has not changed (Lidén et al. 1997, Frost et al. 2006). For thissimple reason, it would be much better to abandon the direct linkage between nomina and “phylogenetic defi-nitions” of taxa in “phylogenetic nomenclature”: taxa could well be defined according to the principles of“phylogenetic definitions” of the Phylocode, but allocation of nomina to taxa should remain governed byostensional pointing to onomatophores, as suggested above and as practiced under the Code.
In any consistent and explicit taxonomy, intensional and extensional definitions of nomina should result inthe exactly same nomenclature, as the content of taxa is defined through the use of explicit criteria and charac-ters. However, in practice, this is true to a certain extent only, as allocation of organisms to taxa depends onthe amount and quality of the information we have about them: not rarely, the collection of new specimens,the discovery of new characters, or the mere re-examination of known specimens or characters, results in tax-onomic reallocation of specimens or taxa, and thus in nomenclatural changes.
All the discussion above shows that the allocation of nomina through definitions of nomina, be themextensional or intensional, is not a good system: it is bound to a conception of taxonomy, and therefore liableto become obsolete if taxonomic paradigms change; it is, at least partly, a circular system inside language, dis-connected from specimens; it is fragile relative to changes in our phylogenetic hypotheses, as in such cases thenomina, even if they have been in use for a long time, have to be either abandoned or “re-defined”, both solu-tions that are not desirable if nomenclature is to remain a stable reference system for biology. We then have toabandon the idea of “defining” nomina, but to use a system of ostensional allocation of nomina to taxa. Butthen this puts limitations on the possibility to use a monosemic nomenclatural system, as will now be shown.
Ostensional allocation of nomina to taxa
In this system, which is in force in the Code, nomina are not defined, but are attached to taxa through a toolspecial to biological nomenclature, the onomatophore. The role of the onomatophore is not to provide taxo-

Zootaxa 1519 © 2007 Magnolia Press · 45PHYLOGENY, TAXONOMY AND NOMENCLATURE
nomic information on characters (although it may “incidentally” do so), but to tie the nomen to a living (oronce living) population of organisms. Allocation of the nomen to a taxon is made through the taxonomic allo-cation of the onomatophore to a taxon, within the frame of a taxonomic arrangement adopted as valid by a tax-onomist. Under this system, nomina are independent from the definitions (either by diagnosis, apognosis orcladognosis, or other) given to the taxa they designate, and also of the taxonomic paradigm under which thesedefinitions were provided. Contrary to the misleading allegations of some (e.g., de Queiroz & Gauthier 1990,1994; Pennisi 2001; Joyce et al. 2004; Sluys et al. 2004), onomatophores (so-called “types”) do not allow todefine taxa and their use is not a survival of typological ideas in evolutionary biology. To definitely avoid suchbasic confusions, it is better to abandon the use of the term type, and its derivatives, in biological nomencla-ture, and to replace it by Simpson’s (1940, 1961) term onomatophore and other new terms, which do not havethis misleading connotation (Dubois 2005a).
Ostension through an onomatophore is not a definition of the taxon to which the nomen is attached, as itdoes not provide any limit for this taxon, it does not state which are the non-members of the taxon, and it onlyprovides information on membership in the taxon of just one or a few specimens or taxa. As a result, thenomen attached to this onomatophore can potentially apply to several, more or less inclusive, taxa, and, with-out additional Rules, all zoological nomina could potentially apply to all zoological taxa including their ono-matophores, upwards until the kingdom ANIMALIA. Rules are therefore needed to make a nomenclaturalsystem based on onomatophores monosemic, or only “slightly” polysemic.
Dubois (2006c) discussed in detail this question and identified five systems which have been in use, orcould possibly be used, to play this function.
The first one is the system in use in the Code, with nominal-series and a Rule of Coordination. In this sys-tem, the nomenclatural scale necessary to name all zoological taxa from the least to the most inclusive isdivided into five levels, three of which (the family-, genus- and species-series) are recognized in the Code,and two (the class- and the variety-series) excluded from the latter. Nomina in the five nominal-series areindependent, so that a given nomen is limited in potential use to the ranks used in its nominal-series. Withineach nominal-series recognized by the Code, the nomenclatural system is polysemic, at least for some nomina(the first published among several nomina which apply to a taxon and to several of its subordinate taxa). Asremembered above, this system, known in the Code as the system of “nominotypical taxa”, is better renamed(to contribute to extirpating the use of the term “type” from zoological nomenclature) system of “coordinatetaxa with eponyms”.
This system of eponymy has been criticized in the recent years by supporters of monosemic nomencla-tures. Their criticism is understandable, especially if scientific nomina are to be used “blindly” by many actorswho are not part of the community of taxonomists, such as many “Google users”. However, there is a basicquestion here. Do “Google users” require from physicists to make the laws of physics “simpler” and “easier touse by laymen”? Certainly not, and if they did physicists would simply ignore this request. Many other exam-ples could be given, where it is clear that “Google users” do not ask for an internal change in the domain inwhich they may be interested, but ask simply for clearer, more detailed, pedagogical explanations of what isgoing on in this domain. Strangely enough, this is not what is required from taxonomists, but to deeply modifytheir own discipline, to “please” lazy outsiders (or sometimes insiders?) of the field who do not wish to spendtime learning what is going on. The system of coordinate taxa with eponyms has been in use for more than onecentury and a half in millions of publications by considerable numbers of zoologists who have apparently nothad insuperable difficulties in mastering its use. Much more importantly, the whole building of zoologicalnomenclature has been constructed under these Rules, so modifying it now would require to change consider-able numbers of nomina, a work which would require considerable manpower, time and funds. Sure, this kindof “game” could be thought of if zootaxonomists had “nothing else to do”, but, as reminded above, they wouldbe better inspired to use their energy, working time and money to collect, study and describe the vanishingbiodiversity of our planet before most of it has gone...

DUBOIS46 · Zootaxa 1519 © 2007 Magnolia Press
Be it as it may, it seems that the existence of eponyms is a major reason why some zoologists want to getrid of the Rules of the Code to adopt new nomenclatural rules that would respect the dogma “one taxon, onenomen”. From what appears in papers like that of Hillis (2006), it seems that these people blame the recogni-tion of ranks in the Code for the existence of eponymy, but this is not true. Eponymy exists in the Code notbecause of ranks, but of nominal-series and of the Rule of Coordination. As was shown elsewhere in detail(Dubois 2006c), the basic Rules of the Code (onomatophores and priority) could fully work in a nomencla-tural system with ranks but without nomenclatural-series and coordination. Four systems at least would allowthis: the system of absolute ranks, the system of additivity of onomatophores, the system of indissoluble setsof nominal taxa as onomatophores, and the combined system of indissoluble sets of nominal taxa as onomato-phores and as onomatostases. Dubois (2006a,c) further suggested that the latter nomenclatural system (or, insome cases, the just preceding one) should be used to incorporate the nomina of higher-ranked taxa (nomen-clatural class-series) into the Code. This system can be qualified as a system of bidirectional ostension, beinga combination of inclusive and exclusive ostension (Dubois 2006c): whereas inclusive and exclusive osten-sions alone do not provide upper limits for taxa designated by the nomen, bidirectional ostension does, and anomen so defined can apply to only one taxon in a given taxonomy, even in a system using ranks.
The system of bidirectional ostension could theoretically be used in the three nominal-series of the Codeto produce an unambiguous, monosemic nomenclature at all levels. Whether this would be a good idea is,however, another question, that needs to be discussed now.
Discussion
The distinction between nomenclatural ranks and taxonomic categories
As discussed in many works already (see references in Dubois 2005a, 2006b) and shown again above, nomen-clatural ranks are useful in zoological taxonomy. They help for the storage and retrieval of taxonomic andphylogenetic information. The nomenclatural hierarchy is based nowadays upon the hypothesized structure ofthe tree of life, but other taxonomic paradigms could be used which would also rely on a hierarchical arrange-ment of information, as this is a very common and convenient system of ordering information in databases ofall kinds. It would be “masochist” for zoologists to abandon ranks and to deal only with “unranked taxa”. Thiswould not be understood by ecologists, conservationists and all other biologists and laymen. As Godfray &Knapp (2004b: 561) put it, we need “a hierarchical series of partitions of biodiversity, however imperfect thatmight be. An ecologist can work with the generic and family diversity of insects in a field site, and comparethis with other field sites. He or she knows that genera or family distinctions are arbitrary, but this matters lessin comparative studies using the same taxa. (...) Were taxonomists to retreat from ranked taxa, they would bere-invented by ecologists, naturalists and horticulturalists, and we believe that it would be better done by tax-onomists.”
But, in parallel with their continued use of nomenclatural ranks, taxonomists have the duty to explainwhat ranks are and are not, and to explain the difference between nomenclatural ranks and taxonomic catego-ries as clarified above. There is no doubt that, today, this pedagogical work is made difficult by the use of thesame terms to designate both ranks and categories. This is particularly true and worrying in the case of theterm species. Clarification of this problem would be made much easier by using different terms for the twokinds of entities. A few proposals in this respect can be offered for thought and discussion.
It would seem better and clearer to continue using the terms species, genus, family, order, class, etc., todesignate nomenclatural ranks. The main reason for this is that nomenclatural ranks are of universal use overthe whole tree of life (both in all taxonomic groups, and at all hierarchical taxonomic levels), whereas catego-ries are used only for taxa at some ranks and in some taxonomic groups only. Nomenclatural ranks are to befound in most non-taxonomic uses of nomina, e.g., in lists of taxa for conservation biology, commerce and

Zootaxa 1519 © 2007 Magnolia Press · 47PHYLOGENY, TAXONOMY AND NOMENCLATURE
international regulations. In contrast, taxonomic categories are used only in some cases, particularly in spe-cialized biological literature, to draw attention to peculiar evolutionary situations.
Let us consider briefly the many taxonomic categories that have been recognized so far at the ranks spe-cies and subspecies or just between, above or below them. My purpose here is not review them again, but justto consider the terminology used to designate them. Many terms have been coined to designate these catego-ries. It is here suggested that the standardized use of these terms, or of new, related ones, should be general-ized in zoological taxonomy. My preference, in this case like in others (Dubois 2006a), would go to simpleuninominal terms, rather than compound names, periphrases or formulae. This is not only for sake of brevity(although this also counts), but to point out that these terms are precise designations with a well-definedmeaning, not approximate wordings that can be used with variable senses. Thus, particularly misleading des-ignations like “species group” or “species complex”, which have been used in many senses, most often with-out being defined, should be avoided. I am also personally reluctant to the use of acronyms, such as “LITU”(Pleijel & Rouse 2000), or of abbreviations (abusively called nowadays by some “acronyms”, whereas anacronym is an abbreviation that can be read like a word, not spelled as a sequence of letters), like “BSC” or
“PSC”1. Simple words, precise technical terms, like many of those listed and studied by Bernardi (1956, 1957,1980), should in my opinion be preferred in scientific publications.
Many more terms have been coined than are necessary to designate really different concepts. There arenot 92 genuinely different “definitions of species” or even 22 “species concepts” (see above), and Mayr &Ashlock (1991: 24–28) are certainly closer to the truth when they recognize only four distinct concepts of thetaxonomic category species. Thus we will probably need only a handful of terms to designate these main cat-egories. Beside the plurinominal designations, formulae or abbreviations like “biological species concept” or“BSC”, many of the terms coined for this purpose have endings in -species: e.g., biospecies, morphospecies,agamospecies, superspecies or quasispecies. This would indeed become misleading if use of the term specieswas restricted, as suggested above, to designate a nomenclatural rank. If this is accepted, for full clarity, com-posed terms using this root should also designate nomenclatural ranks: subspecies, but also supraspecies,epispecies, infraspecies or hypospecies (see Dubois 2006a: 211–225, 2006b: 835–837). The other simplewords introduced in the literature to designate taxonomic categories are either unique terms without resem-blance among them (e.g., exerge, grex, morpha, forma or cline; see Bernardi 1957), or terms ending in -on, bysimilarity with the term taxon (e.g., linneon, jordanon or syngameon, as used by Cuénot & Tétry 1951; klep-ton or klonon, as used by Dubois 1991; a similar derivation also resulted in the term plesion of Patterson &Rosen 1977). It is here suggested that generalisation of endings in -on to designate taxonomic categories couldbe a good way to point to the difference between the latter and taxonomic ranks.
For taxa at ranks species, subspecies and close, one possibility would be to coin terms ending in -specion,a barbarism obtained from the Latin word species and the Greek suffix -on, frequent in neuter substantivations
1. A reviewer of this paper asked me to justify this opinion, so I comply with this request. I am well aware that nowadays
many people like to use acronyms and abbreviations, but this is mostly a language of newspapers, official circulars and
documents, or other short-lived writings. Scientists, who write publications with a longer potential life, are (or should be)
intellectuals, not administrators or journalists, and should in my opinion rather use a correct language, with complete
words, complete sentences with verbs (rather than “telegraphic style”), correct vocabulary and grammar. A newspaper
headline can read: “UNO to WWF: OK!”, but I do not think this style should be used in scientific publications. Funnily,
in one of his films, the movie-maker Michael Moore blamed the USA, the most powerful country in the world, for not
having even a simple name to designate it, unlike even the smallest village in the world. In the same country also, streets
often have mere numbers instead of names, etc. Who knows, perhaps this unusual tradition reflects a mistrust or aversion
for names and naming, that might have something to do with the difficulties of some recent authors with nomenclatural
problems, which can go as far as proposing to get completely rid of nomina in biological taxonomy, and replace them by
numbers or codes!

DUBOIS48 · Zootaxa 1519 © 2007 Magnolia Press
of Greek adjectives ending in -os. Similarly barbaric, but clear in use once defined, could be terms ending in-genion for taxonomic categories at rank genus, or in -familion at rank family. Using such endings for emen-dation of terms previously introduced in the literature would allow to produce terms of similar aspect (e.g.,biospecion, morphospecion or agamospecion), thus easy to understand. Terms already coined with this ending(e.g., syngameon, klepton or klonon) could be kept. Another possibility would be to revive an old approach inevolutionary systematics, to dedicate taxonomic categories to colleagues who played a role in the discovery,definition or acceptance of these categories: thus Lotsy’s (1916) linneon and jordanon, dedicated respectivelyto Carl von Linné and Karl Jordan. One could generalize this practice and dedicate the most important con-cepts of categories at rank species to researchers who played a major role in their recognition: e.g., mayron,dedicated to Ernst Mayr, for the “biological species concept” (species as a “protected gene pool”), or simpson,simply borrowed unchanged from the surname of George Gaylord Simpson, for the “evolutionary speciesconcept” (species as a “separate lineage”).
Similar terms in -on could be revived or coined for the few taxonomic categories that have been recog-nized for taxa at ranks genus or above. Thus, to designate the taxonomic category of rank genus defined byDubois (1981, 1988) as a group of species potentially hybridizable, at least in artificial conditions, the termcoenospecies of Turesson (1922) could be emended to coenogenion.
This proposal is not explored in more details here, but it is clear that adopting a different terminology todesignate taxonomic categories and nomenclatural ranks would considerably clarify the current situation inthis domain.
Under such a paradigm, allocation of a taxon to a nomenclatural rank and to a taxonomic category are dis-tinct, independent processes. Whereas all taxa (except so-called plesions and incertae sedis) are currentlyreferred to nomenclatural ranks, only a few taxa, at the lower nomenclatural ranks, are referred or referable totaxonomic categories, and higher taxa (order, class and above) may only be referred to ranks, never to catego-ries, for reasons given above. Under this approach, comparisons between taxa (e.g., for biodiversity richnessassessment) would be relevant in some cases using taxonomic categories, but would remain inappropriate inall cases using nomenclatural ranks.
Compulsory and optional nomenclatural ranks
The tree of life is not a symmetrical tree. Some branches are richer in terminal taxa and in intermediatebranches than others. If this is to be reflected in the taxonomic scheme and in nomenclature, this means thatsuch branches should be richer in nomenclatural ranks than others. This does not raise any special difficulty.As discussed above, the function of ranks is to express the hierarchical relationships between taxa, i.e. toexpress relations of subordination, superordination and parordination. However, it is important to realize that,because there is currently no criterion of equivalence between taxa in different groups, the fact that two taxabear the same rank in different branches of the cladogram does not imply any kind of “homology” or equiva-lence between them: such taxa are not really parordinate (Dubois 2006b), but pseudoparordinate, and the factthat they share a rank does not provide any information on their relationships or characteristics. But, within agiven taxon (or branch of the tree), ranks provide important information: whereas relations of coordination(superordination-subordination) express ancestor-descendants relationships, relations of parordination expresssister-group relationships.
It is not necessary to have taxa at all ranks of the nomenclatural hierarchy in all groups. As shown in fig-ure 1, to have a fully informative nomenclature, only two kinds of taxa require nomenclatural recognition (i.e.,naming): (1) taxa including several subtaxa; (2) taxa that are sister-groups of the latter, even if they do notinclude subtaxa. The latter should be taxonomically and nomenclaturally recognized at the same rank as theirsister-taxa. Not doing so, as done by partisans of various kinds of unranked or pseudoranked nomenclatures
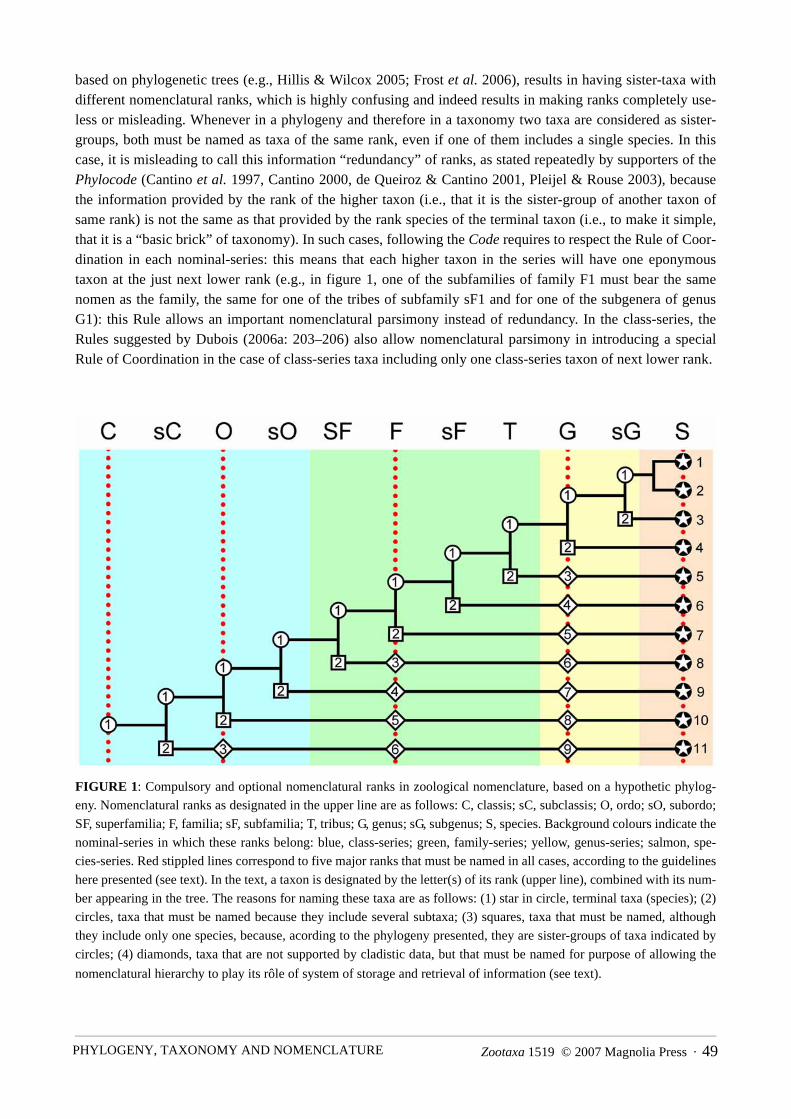
Zootaxa 1519 © 2007 Magnolia Press · 49PHYLOGENY, TAXONOMY AND NOMENCLATURE
based on phylogenetic trees (e.g., Hillis & Wilcox 2005; Frost et al. 2006), results in having sister-taxa withdifferent nomenclatural ranks, which is highly confusing and indeed results in making ranks completely use-less or misleading. Whenever in a phylogeny and therefore in a taxonomy two taxa are considered as sister-groups, both must be named as taxa of the same rank, even if one of them includes a single species. In thiscase, it is misleading to call this information “redundancy” of ranks, as stated repeatedly by supporters of thePhylocode (Cantino et al. 1997, Cantino 2000, de Queiroz & Cantino 2001, Pleijel & Rouse 2003), becausethe information provided by the rank of the higher taxon (i.e., that it is the sister-group of another taxon ofsame rank) is not the same as that provided by the rank species of the terminal taxon (i.e., to make it simple,that it is a “basic brick” of taxonomy). In such cases, following the Code requires to respect the Rule of Coor-dination in each nominal-series: this means that each higher taxon in the series will have one eponymoustaxon at the just next lower rank (e.g., in figure 1, one of the subfamilies of family F1 must bear the samenomen as the family, the same for one of the tribes of subfamily sF1 and for one of the subgenera of genusG1): this Rule allows an important nomenclatural parsimony instead of redundancy. In the class-series, theRules suggested by Dubois (2006a: 203–206) also allow nomenclatural parsimony in introducing a specialRule of Coordination in the case of class-series taxa including only one class-series taxon of next lower rank.
FIGURE 1: Compulsory and optional nomenclatural ranks in zoological nomenclature, based on a hypothetic phylog-
eny. Nomenclatural ranks as designated in the upper line are as follows: C, classis; sC, subclassis; O, ordo; sO, subordo;
SF, superfamilia; F, familia; sF, subfamilia; T, tribus; G, genus; sG, subgenus; S, species. Background colours indicate the
nominal-series in which these ranks belong: blue, class-series; green, family-series; yellow, genus-series; salmon, spe-
cies-series. Red stippled lines correspond to five major ranks that must be named in all cases, according to the guidelines
here presented (see text). In the text, a taxon is designated by the letter(s) of its rank (upper line), combined with its num-
ber appearing in the tree. The reasons for naming these taxa are as follows: (1) star in circle, terminal taxa (species); (2)
circles, taxa that must be named because they include several subtaxa; (3) squares, taxa that must be named, although
they include only one species, because, acording to the phylogeny presented, they are sister-groups of taxa indicated by
circles; (4) diamonds, taxa that are not supported by cladistic data, but that must be named for purpose of allowing the
nomenclatural hierarchy to play its rôle of system of storage and retrieval of information (see text).

DUBOIS50 · Zootaxa 1519 © 2007 Magnolia Press
Although this is a legitimate matter of discussion, my personal opinion is that a third kind of taxa shouldalso be recognized and named. As reminded and commented above, Dubois (2006a: 217) suggested that mostanimal taxa of the planet are or should be referable to seven primary key ranks (regnum, phylum, classis,ordo, familia, genus, species), which could thus be considered virtually “compulsory” for all animal organ-isms. The same proposal was published independently (during review of this paper) by Kuntner & Agnarsson(2006), who justified it by “purely practical reasons”, and this is a strong point of agreement between our pro-posals although I fully disagree with their suggestion of a “compromise” between the Code and the Phylocodewhich would only increase confusions. This proposal follows a long tradition in zootaxonomy. It is supportedby the important idea that biological classifications have two major and distinct functions (Mayr 1982, 1997),a practical one (i.e., providing a universal system of storage and retrieval of information) and an explanatoryone (i.e., providing an evolutionary interpretation and explanation of the diversity of organisms). Ignoring thefirst of these functions to concentrate only on the second may seem appealing as a “purer” approach and mayplease some professional taxonomists and theoreticians, but is not doing a service to taxonomy and its innu-merable users in all domains of human activity (Cracraft 1974, Ashlock 1984, Benton 2000, Dubois 2005a).
Using these seven standard nomenclatural ranks for all taxa (i.e., placing all species in a taxon of each ofthe six ranks above) would in particular facilitate the search for taxonomic information in databases, whichcould have a simplified structure including only these seven main ranks. This is illustrated in figure 1 whichshows that, if this “principle” is adopted, some taxa additional to the sister-groups mentioned above must alsobe named. In this case, taxonomic redundancy is genuine (in extreme cases, a single “phylogenetically iso-lated” species must be referred to several subsequent monospecific taxa of ranks genus, family, order, etc.),but this concerns only a very small proportion of all animal taxa (although, given the incompleteness of ourknowledge, this proportion is higher in entirely fossil groups). The taxonomy illustrated in figure 1 is by nomeans “revolutionary”, but rather corresponds to a very frequent way of naming taxa in zootaxonomy follow-ing the Rules of the Code.
A final, but important recommendation made by Dubois (2006a: 224) is that, in any taxonomy, very wellknown taxa (i.e., nomina used in many textbooks or as titles of volumes of databases or series like the Zoolog-ical Record) should, as far as possible, be ascribed primary key ranks such as phylum, classis or ordo. Thiscan be obtained by deciding which ranks should be given in the cladogram to the taxa designated by thesenomina under the nomenclatural Rules. The ranks of all other taxa just above and below then depend on theranks of these “major” taxa. This does not raise any theoretical difficulty as: (1) ranks are arbitrary and meannothing by themselves, they just have a relative meaning, that of designating a place in a hierarchy of taxa; (2)there is no need of a robustness in the ranks of taxa such as there is for the nomina of taxa: many new cladisticanalyses result in the addition of deletion of nodes in the cladograms, and by way of consequence of ranks inthe nomenclatural hierarchy, but these changes are trivial as long as the extensions and intensions of most taxado not change.
Should the Code be fundamentally modified to become fully monosemic?
It has been shown above that a nomenclatural system using bidirectional ostension for the allocation of nom-ina to taxa could theoretically be used in the three nominal-series of the Code to produce an unambiguous,monosemic nomenclature at all levels. As far as I am concerned, I think the benefit that this change couldbring (“to please Google users”) would not counterbalance the gigantic loss of time and energy that it wouldcause to professional taxonomists in order to implement the new system, not to mention the errors and the lossof information that would be caused by the transfer of data from one system to the other, and the problems ofcommunication that would be caused by the parallel existence of several nomenclatural systems in the litera-ture, on the world wide web, etc. (see Sluys et al. 2004). For the nomenclatures in family-, genus- and species-

Zootaxa 1519 © 2007 Magnolia Press · 51PHYLOGENY, TAXONOMY AND NOMENCLATURE
series to become monosemic would require coining considerable numbers of new nomina, or resurrecting syn-onyms, in order to rename hyponymous taxa of all subdivided taxa within these series. Furthermore, thiswould pose particular problems in the family-, and sometimes in the genus-series, when the hyponymoustaxon that has to be renamed contains only one taxon of next lower level.
Thus, in the example of the Appendix below, the tribe LEPTOBRACHIINI contains only one genus, Lepto-brachium, which is also the nucleogenus of the subfamily LEPTOBRACHIINAE: renaming the tribe would requireusing the nomen of a synonym (or in this case of the subgenus Vibrissaphora) as the basis for the tribe nomen,but how to proceed when there is no synonym or lower taxon? Of course, some would probably argue thatLEPTOBRACHIINAE and LEPTOBRACHIINI are not the same nomen, as they have different spellings, but this iswrong. Both are different morphonyms of the same nomen, and they have the same date, author and onomato-phore (Dubois 2000). A given nomen can have different morphonyms, including its protonym (original mor-phonym) and its aponyms (derived morphonyms). An aponym differs from the protonym in one at least of thethree following characteristics: its spelling, rank and onymorph (for more details, see Dubois 2000). Confu-sion between nomen (an individual nomenclatural entity) and morphonym (any avatar of this entity) is arather frequent one in taxonomic literature, and this is why precise technical terms are needed to designatenomenclatural concepts (Dubois 2000, 2005a, 2006a). The different eponyms of a nomen that can be used todesignate coordinate taxa are different morphonyms, but they are the same nomen. Under the Code, the samenomen, under different avatars, can designate different taxa at different ranks. This again stresses the differ-ence between nomen and taxon.
This is even clearer for genus-series and species-series nomina, as there is no spelling difference betweengeneric and subgeneric, or specific and subspecific, eponymous nomina. In the Appendix below, the genusLeptobrachium and the subgenus Leptobrachium are different taxa, but they bear the same nomen, as differentmorphonyms. And in this nominal-series, difficulties similar to that described above would arise if nomencla-ture had to be made monosemic. In the case of a genus including two subgenera, both of which include a sin-gle species, without synonyms or subspecies, a new nomen should be coined for one of the subgenera, basedon one of the two species, but then, according to the Rules now in force, one of the two subgeneric nominawould be an invalid junior objective synonym of the genus nomen, so that the Rules would have to be modi-fied to make this possible.
As can be seen from these short examples, which could be developed in more details, making the whole ofzoological nomenclature monosemic would require extensive changes in the Code and creation of many newnomina.
My opinion is that, for the time being (i.e., for our generation and for the next ones), all priorities of tax-
onomy should be put on the inventory of the species of our planet, and not on nomenclatural issues1. However,if militants for a “Google taxonomy” think their cause is worth enough, and if they are supported in theirendeavour by “users” and “societal demand”, I think they should consider the two following suggestions: (1)they should refrain from adopting the project of the Phylocode in its present form, and other projects in whichnomenclatural rules are linked to a taxonomic theory, to adhere to a theory-free system of allocation ofnomina to taxa through bidirectional ostension with onomatophores and onomatostases, which could allow foran evolution of the Code toward a fully monosemic nomenclatural system; (2) they should seek funding from“society” to pay manpower, bibliographic research, database implementations, etc., required to realize theseheavy changes in millions of nomina without putting the burden of this work on the shoulders of professionaltaxonomists, who are already considerably much less numerous than required by the taxonomic impediment
1. This statement may seem contradictory to the long discussion in this paper, but it is not. One of the aims of this dis-
cussion is to show that the current Code is excellent in most of its provisions and Rules and that it should not be thrown
to the garbage: it only needs to be updated in several respects, which would be much less time- and energy-consuming
than replacing it altogether by brand new rules.

DUBOIS52 · Zootaxa 1519 © 2007 Magnolia Press
and should not be diverted from the work of inventory of living organisms of the planet before most of the lat-ter are extinct (Wilson, 2004).
The need of clarifications in the Code
As discussed above, some colleagues have difficulties understanding the way ranks work under the Code.Some of them think ranks have or should have an absolute meaning by themselves, although they just providerelative information on the hierarchical relationships between taxa. Some of them think that sister-taxa in ataxonomy can be referred to different ranks, or that taxa at the same rank can be embedded into one another.Worse even, when shown that they are wrong under the Rules of the Code, such authors are ready to write(and this is accepted by the referees and editors of journals where they submit such statements): “Readers arewelcome to read the rules and decide for themselves which interpretation is correct” (Hillis 2006: 335).Although in my opinion, and as shown by the detailed analyses that I presented elsewhere (Dubois 2006d,2007), there is no question of “interpretation” here, I suggest the Code’s wording should be even more clari-fied to avoid such futile and uselessly time-consuming discussions in the future.
I suggest that this could be done by the addition of two new Articles in the Code, better defining nomen-clatural ranks and describing their use in zoological nomenclature. These Articles should be at the beginningof the Code, e.g. as Articles 1.2.3 and 1.2.4, or as an additional Article between the current Articles 1.1 and1.2. Here are provisional proposals regarding these two Articles, which use in part some of the wordings thatappear in the current Glossary of the Code:
“Article 1.2.3. For the purpose of the Code, a nomenclatural rank is the level of a taxon in a nomen-clatural hierarchy (e.g., a family is at a rank lying between superfamily and subfamily). This is anomenclatural concept, distinct from the taxonomic concept of category.”
“Article 1.2.4. In a nomenclatural hierarchy, any taxon immediately subordinate to another one mustbe referred to the next lower nomenclatural rank (e.g., if a family includes two subordinate taxa and ifthe latter are referred to the nominal family-series, they must be given the rank of subfamily). Twoparordinate taxa (e.g., sister-taxa in a taxonomy based on hypothesized cladistic relationships) mustalways be given the same nomenclatural rank, even if they do not include the same number of lower-ranked taxa (e.g., if one taxon subordinate to a family is given the rank subfamily, its sister-taxonmust also be given this rank, even if the former includes several tribes, genera and species, and thelatter only one species).”
Other suggested changes in the Code regarding this question of ranks are to replace the formula “hierar-chy of classification” (which appears in the current wording of Article 42.1) by the formula “nomenclaturalhierarchy” (Dubois 2006b: 835), and, throughout the Code, the terms “family group”, “genus group” and“species group” by “family-series”, “genus-series” and “species-series” (Dubois 2000, 2005a, 2006a).
Phylogenetic definitions of taxa and the Code
The nomenclatural system of the Code relies on two major concepts that in my opinion make it by far superiorto all other nomenclatural systems that have been proposed so far for a taxonomy of living organisms: (1) ono-matophores, which are permanent, objective references for the allocation of nomina to taxa, independent fromthe taxonomic philosophy used to recognize and define the latter; (2) priority, which is a permanent, objective,

Zootaxa 1519 © 2007 Magnolia Press · 53PHYLOGENY, TAXONOMY AND NOMENCLATURE
indisputable criterion for establishing the validity of one nomen among several synonyms (Ohler 2005). Asdiscussed above, this system needs to be modified and updated, but certainly not abandoned, especially asmillions of nomina in current use in zoology have been established under this system and as practicising tax-onomists, who are far from being numerous enough to do their work, have better to do than “redefining” thesemillions of nomina. Unfortunately, rather than improving the current Code, some biologists recently decidedto devote considerable time and energy to work on alternative nomenclatural systems, although this “exten-sive literature” and “special symposia” are of highly questionable utility compared to other discussions thathave so far attracted little interest (Avise & Michell 2007: 133).
Following the ideas of Ghiselin (1966), de Queiroz & Gauthier (1990: 308) stated that “definitions aregiven to words, including to names, not to the things that the names represent, that is, the taxa themselves”.Such a statement makes the confusion between “things” that no doubt exist, or existed, in the real world, i.e.,clades, and the concepts we use to account for these (unknown) clades, i.e., taxa. Taxa are not things. Evenwhen based on cladograms, all taxa are concepts (i.e., tools used by man’s mind to interpret reality), they areworking hypotheses that are defined by science, not discovered: “taxonomic definitions (or diagnoses, if youprefer) are hypotheses about entities in nature. Those hypotheses are subject to test and rejection or revision.”(Schuh 2003: 70). Science usually proceeds this way. Scientific theories allow predictions to be made, on thebasis of such predictions some concepts may be proposed, and names given to objects or features of realitythat have not yet been observed or demonstrated to exist. When research allows to falsify these hypotheses,they must be replaced by others. The Mendeleev periodic table of elements allowed to predict the existence ofseveral elements, some of their properties, and to name them before their discovery: such definitions andnames were then hypotheses or theories and would have to have been abandoned if these elements had notbeen observed. As organic evolution is not a teleological process, there is no (and cannot be any) Mendeleevtable of species or taxa (“clades”) (Dubois 2003), but, using scientific methodologies, we can formulatehypotheses about these taxa and give them nomina according to these hypotheses. Just like for the elements,these nomina rely on hypotheses, but as it is considerably more difficult to demonstrate the existence of aclade than that of a chemical element, decades or centuries will be needed before these hypotheses can turn toestablished facts.
Thus, the discussion about whether taxa or nomina are defined (Stuessy 2000, 2001; Jørgensen, 2000; deQueiroz & Cantino 2001) is a largely sterile one, as the term “taxon” is given different meanings by the con-tradictors: either to designate a “thing of the real world” (a “clade”), or to designate the concept we use for thisthing (also called by some cladon, phylon and phylo-taxon; see above). But this theoretical, largely irrelevant,debate, has a strong practical consequence: it was the theoretical foundation on which the Phylocode projectwas built.
Because they claim that nomina, but not taxa, can be defined, supporters of the Phylocode use intensionaldefinitions for the nomina of zoological taxa. In consequence, as shown above, their nomenclature is tied to ataxonomic paradigm and their nomina do not consistently have the connection to reality that under the Code isprovided by onomatophores. The idea promoted here is that nothing of their interesting and useful theoreticalwork (especially concerning the way “phylogenetic definitions” can be formulated) would be lost if they reck-oned that “phylogenetic definitions may well be used to define taxa, and the allocation of nomina to these taxamade by ostension through onomatophores” (Dubois 2006c: 17). Under such an interpretation, taxa can bedefined under different kinds of apognoses and cladognoses corresponding to the various categories of “phy-logenetic definitions” like those listed in Article 9.4 of the draft Phylocode. This is exemplified in the Appen-dix below. Taxa so defined are deemed to be holophyletic groups and are compatible with the principles of“phylogenetic taxonomy” (de Queiroz & Gauthier 1992). According to such definitions, we can build a list oforganisms and taxa that are referred to this group, and the Rules of the Code can be used to know whichamong these included members is the onomatophore of the oldest available nomen for the taxon, thus provid-ing unambiguously the nomen for this taxon. Of course, in a few cases, because the current state of our knowl-

DUBOIS54 · Zootaxa 1519 © 2007 Magnolia Press
edge may be incomplete or wrong (see e.g. Delorme et al. 2004: 60–61), we will erroneously refer a specimenor a taxon to this group, or fail to refer an organism or a taxon to this group, which may result in having tochange the nomen of this taxon, even if the apognosis or cladognosis is not changed. But such cases shouldbecome rarer and rarer as phylogenetic and taxonomic work progresses. Furthermore, this potential cause oftaxonomic instability is not more important than that caused by bad original definitions of nomina under thePhylocode, which would require either to abandon or to emend them (Bryant 1996: 186) when this is discov-ered.
We are at the beginning of the century of extinctions, and if Phylocode militants accepted that their “phy-logenetic definitions” can apply to taxa, and decided to follow the Code for the allocation of nomina to taxa,this could be extremely beneficial to taxonomy, and more generally to the study and preservation of biodiver-sity. The present confusion due to the parallel existence of several nomenclatural systems would stop. Aboveall, considerable time, energy and money could be saved, rather than to be used to “redefine nomina”, toaccelerate the inventory of the living species of our planet before they get extinct. It is true that most of thetheoreticians who developed the Phylocode paradigm are involved in rather general phylogenetic analyses,and that few of them are working on alpha-taxonomy and species descriptions. But the continued discussionof these questions in the literature is a source of confusion that may become highly detrimental to the disci-pline of taxonomy, if decision-makers retain the impression that this domain is only a battle-field amongincompatible schools, rather than a productive field of research (see also Sluys et al. 2004, Polaszek & Wilson2005). The Code and the Phylocode are indeed based on incompatible philosophies, mostly because theformer one is theory-free regarding taxonomy whereas the latter tries to impose a taxonomic paradigm (“phy-logenetic taxonomy”) to all taxonomists of the present and future. Trying to reconcile both philosophies(Kuntner & Agnarsson 2006) can only increase temporarily the confusion, because in the long run taxono-mists will have to choose between these two options. My opinion is that they should choose the Code, mostlyin order to be able to concentrate on the taxonomic work of collection and description of taxa, but that theyshould take advantage of the theoretical work and of the discussions that have developed recently, mainlyaround the Phylocode project, to improve substantially the current Code, which really needs it.
Conclusion
The use of ranks and nominal-series in zoological nomenclature, recently criticized by some authors, is veryuseful for the storage and retrieval of taxonomic and phylogenetic data. The nomenclatural hierarchy of ranksreflects the phylogenetic tree, although in a simplified and partial way only (because not all nodes can andmust be named). Nomenclatural ranks, however, only provide information on the structure of the taxonomichierarchy, not on particularities of the taxa ascribed to given ranks, which are not equivalent by any criterionand therefore cannot be used for comparisons between taxa (e.g., for biodiversity richness assessment). Part ofthe confusion that surrounds this question is avoided if a clear distinction is made between nomenclaturalranks and taxonomic categories, the latter being in use only for some taxa at lower levels (around species andgenus) and in some zoological groups.
The detailed analysis of this question provided here suggests that the basic Rules of the Code, using osten-sional allocation of nomina to taxa through onomatophores, priority, nominal-series and ranks for the alloca-tion of nomina to taxa, although not perfect in some respects, is the best one nowadays, as it has been in forcefor about 250 years, for millions of nomina, and as it is theory-free regarding taxonomy. The Code shouldhowever be improved in several respects, among others in providing clearer definitions of ranks and descrip-tions of their use in zoological nomenclature. It should make clear that two parordinate taxa (e.g., sister-taxain a taxonomy based on hypothesized cladistic relationships) must always be given the same nomenclaturalrank, even if they do not include the same number of lower-ranked taxa, and that a subordinate taxon cannot

Zootaxa 1519 © 2007 Magnolia Press · 55PHYLOGENY, TAXONOMY AND NOMENCLATURE
share the same rank as a taxon superordinate to it (or vice versa). The request for a monosemic nomenclatural system, not for scientific reasons, but to please non-taxono-
mists, especially customers of the web, is not supported, especially as its implementation over the whole zoo-logical taxonomy would require considerable time and manpower, at the expense of what should beconsidered the priority for taxonomists in the century of extinctions, i.e., the inventory of the living species ofthe planet before they vanish. For the same reason, the replacement of the Code by a new nomenclatural sys-tem like the Phylocode, and the confusion that would result for most users of nomina, could be very detrimen-tal to taxonomy as a whole. However, the theoretical work developed around the Phylocode project hasproduced interesting ideas and proposals, especially regarding the way “phylogenetic definitions” can be for-malised. It is suggested that this methodology can be used to formulate apognoses and cladognoses for taxa,but that the latter should continue to be named according to the Rules of the Code.
Acknowledgements
I am grateful to Michel Laurin, who, despite our basic disagreements, devoted considerable time to commenton a previous manuscript of this paper and provided useful suggestions. Many thanks to Roger Bour for pre-paring figure 1 for publication.
References
Anonymous [Systematics Agenda 2000] (1994) Charting the biosphere: a global initiative to discover, describe andclassify the world’s species. Technical report. American Museum of Natural History, American Society of Plant Tax-onomy, Society of Systematic Biologists and the Willi Hennig Society, NewYork, 34 pp.
Anonymous [International Commission on Zoological Nomenclature] (1999) International code of zoological nomencla-ture. Fourth edition. International Trust for zoological Nomenclature, London, 335 pp.
Anonymous (2005) Discussion. In: Pleijel, F., Le projet de “Phylocode” et la nomenclature des taxons supérieurs, in:Dubois, A., Poncy, O., Malécot, V. & Léger, N. (Eds.), Comment nommer les taxons de rang supérieur en zoologieet en botanique?, Biosystema, 23, 67–71.
Ashlock, P.D. (1971) Monophyly and associated terms. Systematic Zoology, 20, 63–69.Ashlock, P.D. (1984) Monophyly: its meaning and importance. In: Duncan, T. & Stuessy, T. F. (Eds.), Cladistics: per-
spectives on the reconstruction of evolutionary history, Columbia University Press, New York, pp. 39–46.Avise, J.C. & Johns, G.C. (1999) Proposal for a standardized temporal scheme of biological classification for extant spe-
cies. Proceeding of the national Academy of Sciences of the USA, 96, 7358–7363.Avise, J.C. & Mitchell, D. (2007) Time to standardize taxonomies. Systematic Biology, 56, 130–133.Bănărescu, P. (1973) Principiile şi metodele zoologiei sistematice. Editura Academiei Republicii Socialiste România,
Bucureşti, 220 pp.Barigozzi, C. (Ed.) (1982) Mechanisms of speciation. Liss, New York, 560 pp.Benton, M.J. (2000) Stems, nodes, crown clades, and rank-free lists: is Linnaeus dead? Biological Reviews, 75, 633–648.Bernardi, G. (1956) Contribution à l’étude des catégories taxonomiques. I. Avant-propos, nomenclature et définitions.
Bulletin de la Société entomologique de France, 61, 194–200. Bernardi, G. (1957) Contribution à l’étude des catégories taxonomiques. II. Les Règles internationales de la nomencla-
ture zoologique et la notation des catégories taxonomiques. Bulletin de la Société entomologique de France, 62,224–250.
Bernardi, G. (1980) Les catégories taxonomiques de la systématique évolutive. In: Bocquet, C., Génermont, J. &Lamotte, M. (Eds.), Les problèmes de l’espèce dans le règne animal, Vol. 3, Mémoires de la Société zoologique deFrance, 40, 373–425.
Blackwelder, R.E. (1967) Taxonomy: a text and reference book. John Wiley & Sons, New York, 713 pp.Brummitt, R.K. (1997) Taxonomy versus cladonomy, a fundamental controversy in biological systematics. Taxon, 46,
723–734.Bryant, H.N. (1996) Explicitness, stability, and universality in the phylogenetic definition and usage of taxon names: a
case study of the phylogenetic taxonomy of the Carnivora (Mammalia). Systematic Biology, 45, 174–189.Cantino, P.D. (2000) Phylogenetic nomenclature: addressing some concerns. Taxon, 49, 85–93.

DUBOIS56 · Zootaxa 1519 © 2007 Magnolia Press
Cantino, P.D., Olmstead, R.G. & Wagstaff, S.J. (1997) A comparison of phylogenetic nomenclature with the current sys-tem: a botanical case study. Systematic Biology, 46, 313–331.
Carpenter, J.M. (2003) Critique of pure folly. The botanical Review, 69, 79–92.Claridge, M.F., Dawah, H.A. & Wilson, M.R. (Eds.) (1997a) Species, the units of biodiversity. Chapman & Hall, London,
455 pp.Claridge, M.F., Dawah, H.A. & Wilson, M.R. (1997b) Practical approaches to species concepts for living organisms. In:
Claridge et al. (1997a), pp. 1–15.Cracraft, J. (1974) Phylogenetic models and classification. Systematic Zoology, 23, 71–90.Cracraft, J. (2000) Species concepts in theoretical and applied biology: a systematic debate with consequences. In:
Wheeler & Meier (2000), pp. 3–14.Crowson, R.A. (1970) Classification and biology. Heinemann, London, 359 pp.Cuénot, L. & Tétry, A. (1951) L’évolution biologique. Les faits, les incertitudes. Masson, Paris, 597 pp.De Carvalho, M.R., Bockmann, F.A., Amorim, D.S., de Vivo, M., de Toledo-Piza, M., Menezes, N.A., de Figueiredo,
J.L., Castro, R.M.C., Gill, A.C., McEachran, J.D., Compagno, L.J.V., Schelly, R.C., Britz, R., Lundberg, J.G., Vari,R.P. & Nelson, G. (2005) Revisiting the taxonomic impediment. Science, 307, 353.
De Queiroz, K. (2006) The PhyloCode and the distinction between taxonomy and nomenclature. Systematic Biology, 55,160–162.
De Queiroz, K. & Cantino, P.D. (2001) Taxon names, not taxa, are defined. Taxon, 50, 821–826.De Queiroz, K. & Gauthier, J. (1990) Phylogeny as a central principle in taxonomy: phylogenetic definitions of taxon
names. Systematic Zoology, 39, 307–322.De Queiroz, K. & Gauthier, J. (1992) Phylogenetic taxonomy. Annual Review of Ecology & Systematics, 23, 449–480.De Queiroz, K. & Gauthier, J. (1994) Toward a phylogenetic system of biological nomenclature. Trends in Ecology &
Evolution, 9, 27–31.Delorme, M., Dubois, A., Grosjean, S. & Ohler, A. (2006) Une nouvelle ergotaxinomie des Megophryidae (Amphibia,
Anura). Alytes, 24, 6–21.Delorme, M., Dubois, A., Kosuch, J. & Vences, M. (2004) Molecular phylogenetic relationships of Lankanectes
corrugatus from Sri Lanka: endemism of South Asian frogs and the concept of monophyly in phylogenetic studies.Alytes, 22, 53–64.
Dominguez, E. & Wheeler, Q.D. (1997) Taxonomic stability is ignorance. Cladistics, 13, 367–372. Doolittle, W.F. (1999) Phylogenetic classification and the universal tree. Science, 284, 2124–2128.Dubois, A. (1981) Hybridation interspécifique et notion de genre en zoologie. Comptes rendus de l’Académie des Sci-
ences, (3), 292 (A), 201–203.Dubois, A. (1986) A propos de l’emploi controversé du terme “monophylétique”: nouvelles propositions. Bulletin men-
suel de la Société linnéenne de Lyon, 55, 248–254.Dubois, A. (1988) The genus in zoology: a contribution to the theory of evolutionary systematics. Mémoires du Muséum
national d’Histoire naturelle, (A), 140, 1–123.Dubois, A. (1991) Nomenclature of parthenogenetic, gynogenetic and “hybridogenetic” vertebrate taxons: new propos-
als. Alytes, 8, 61–74.Dubois, A. (1997) An evolutionary biologist’s view on the science of biology. Alytes, 15, 133–136.Dubois, A. (1998) List of European species of amphibians and reptiles: will we soon be reaching “stability”? Amphibia-
Reptilia, 19, 1–28.Dubois, A. (2000) Synonymies and related lists in zoology: general proposals, with examples in herpetology. Dumerilia,
4, 33–98.Dubois, A. (2003) The relationships between taxonomy and conservation biology in the century of extinctions. Comptes
rendus Biologies, 326 (suppl. 1), S9–S21.Dubois, A. (2004a) Developmental pathway, speciation and supraspecific taxonomy in amphibians. 1. Why are there so
many frog species in Sri Lanka? Alytes, 22, 19–37.Dubois, A. (2004b) Developmental pathway, speciation and supraspecific taxonomy in amphibians. 2. Developmental
pathway, hybridizability and generic taxonomy. Alytes, 22, 38–52.Dubois, A. (2005a) Proposed Rules for the incorporation of nomina of higher-ranked zoological taxa in the International
Code of Zoological Nomenclature. 1. Some general questions, concepts and terms of biological nomenclature. Zoo-systema, 27, 365–426.
Dubois, A. (2005b) Proposals for the incorporation of nomina of higher-ranked taxa into the Code. Bulletin of zoologicalNomenclature, 62, 200–209.
Dubois, A. (2006a) Proposed Rules for the incorporation of nomina of higher-ranked zoological taxa in the InternationalCode of Zoological Nomenclature. 2. The proposed Rules and their rationale. Zoosystema, 28, 165–258.
Dubois, A. (2006b) New proposals for naming lower-ranked taxa within the frame of the International Code of Zoologi-cal Nomenclature. Comptes rendus Biologies, 329, 823–840.

Zootaxa 1519 © 2007 Magnolia Press · 57PHYLOGENY, TAXONOMY AND NOMENCLATURE
Dubois, A. (2006c) Incorporation of nomina of higher-ranked taxa into the International Code of Zoological Nomencla-ture: some basic questions. Zootaxa, 1337, 1–37.
Dubois, A. (2006d) Naming taxa from cladograms: a cautionary tale. Molecular Phylogenetics & Evolution, 42, 317–330.
Dubois, A. (2007) Naming taxa from cladograms: some confusions, misleading statements, and necessary clarifications.Cladistics, in press [doi: 10.1111/j.1096-0031.2007.00151.x].
Dubois, A. & Günther, R. (1982) Klepton and synklepton: two new evolutionary systematics categories in zoology. Zool-ogischer Jahrbuch Systematik, 109, 290–305.
Dubois, A. & Ohler, A. (1997) Early scientific names of Amphibia Anura. I. Introduction. Bulletin du Muséum nationald’Histoire naturelle, (4), 18, 297–320.
Dupuis, C. (1979) Permanence et actualité de la systématique: la “systématique phylogénétique” de W. Hennig (his-torique, discussion, choix de références). Cahiers des Naturalistes, Bulletin des Naturalistes parisiens, (n.s.),“1978”, 34 (1), 1–69.
Eck, S. (1978) Die Entwicklung superspezifischer Begriffe in der zoologischen Taxonomie seit der Jahrhundertwende.Biologische Rundschau, 15, 98–103.
Frost, D.R., Grant, T., Faivovich, J., Bazin, R.H., Haas, A., Haddad, C.F.B., de Sá, R.O., Channing, A., Wilkinson, M.,Donnellan, S.C., Raxworthy, C.J., Campbell, J.A., Blotto, B.L., Moler, P., Drewes, R.C., Nussbaum, R.A., Lynch,J.D., Green, D.M. & Wheeler, W.C. (2006) The amphibian tree of life. Bulletin of the American Museum of NaturalHistory, 297, 1–370.
Gaffney, E.S. (1977) The side-necked turtle family Chelidae: a theory of relationships using shared derived characters.American Museum Novitates, 2620, 1–28.
Gaffney, E.S. (1979) An introduction to the logic of phylogeny reconstruction. In: Cracraft, J. & Eldredge, N. (Eds.),Phylogenetic analysis and paleontology, Columbia University Press, New York, pp. 79–111.
Génermont, J. & Lamotte, M (1980) Le concept biologique de l’espèce dans la zoologie contemporaine. In: Bocquet, C.,Génermont, J. & Lamotte, M. (Eds.), Les problèmes de l’espèce dans le règne animal, Vol. 3, Mémoires de laSociété zoologique de France, 40, 427–452.
Ghiselin, M.T. (1966) An application of the theory of definitions to systematic principles. Systematic Zoology, 15, 127–130.
Godfray, H.C.J. & Knapp, S., (eds.) (2004a) Taxonomy for the twenty-first century. Philosophical Transactions of theroyal Society of London, (B), 359, 559–739.
Godfray, H.C.J. & Knapp, S. (2004) Introduction. In: Godfray & Knapp (2004b), pp. 559–569.Haeckel, E. (1866) Generelle Morphologie der Organismen. Zweiter Band. Allgemeine Entwickelunggsgeschichte der
Organismen. Georg Kramer, Berlin, 630 pp.Haffer, J. (1986) Superspecies and species limits in vertebrates. Zeitschrift für zoologische Systematik- und Evolutions-
forschung, 24, 169–190.Hennig, W. (1936) Beziehungen zwischen geographischer Verbreitung und systematischer Gliederung bei einigen Dip-
terenfamilien: ein Beitrag zum Problem der Gliederung systematischer Kategorien höherer Ordnung. ZoologischerAnzeiger, 116, 161–175.
Hennig, W. (1950) Grundzüge einer Theorie der phylogenetischen Systematik. Deutscher Zentralverlag, Berlin, 370 pp.Hennig, W. (1966) Phylogenetic systematics. Univ. Illinois Press, Urbana, Chicago & London, 270 pp.Hillis, D.M. (2006) Constraints in naming parts of the tree of life. Molecular Phylogenetics & Evolution, 42, 331–338.Hillis, D.M., Chamberlain, D.A., Wilcox, T.P. & Chippindale, P.T. (2001) A new species of subterranean blind sala-
mander (Plethodontidae: Hemidactyliini: Eurycea: Typhlomolge) from Austin, Texas, and a systematic revision ofcentral Texas paedomorphic salamanders. Herpetologica, 57, 266–280.
Hillis, D.M. & Wilcox, T.P. (2005) Phylogeny of the New World true frogs (Rana). Molecular Phylogenetics and Evolu-tion, 34, 299–314.
Hubbell, S.P. (2001) The unified neutral theory of biodiversity and biogeography. Princeton University Press, Princeton,458 pp.
Jørgensen, P.M. (2000) Names are defined, but not as taxa. Taxon, 49, 779.Joyce, W.G., Parham, J.F. & Gauthier, J.A. (2004) Developing a protocol for the conversion of rank-based taxon names to
phylogenetically defined clade names, as exemplified by turtles. Journal of Paleontology, 78, 989–1013.Kiriakoff, S.G. (1954) Chorologie et systématique phylogénétique. Bulletin et Annales de la Société royale d’Entologie
de Belgique, 90, 185–198.Kiriakoff, S.G. (1965) La vicariance géographique et la taxonomie. Comptes rendus de la Société de Biogéographie, 41,
103–115.Kluge, A.G. (2005) Taxonomy in theory and practice, with arguments for a new phylogenetic system of taxonomy. In:
Donnelly, M.H., Crother, B.I., Guyer, C., Wake, M.H. & White, M.E. (Eds.), Ecology and evolution in the tropics: aherpetological perspective, University of Chicago Press, Chicago, pp. 39–46.

DUBOIS58 · Zootaxa 1519 © 2007 Magnolia Press
Knox, E. (1998) The use of hierarchies as organizational models in systematics. Biological Journal of the Linnean Soci-ety, 63, 1–49.
Köhler, J., Vieites, D.R., Bonett, R.M., Hita García, F., Glaw, F., Steinke, D. & Vences, M. (2005) New amphibians andglobal conservation: a boost in species discoveries in a highly endangered vertebrate group. BioScience, 55, 693–696.
Kuntner, M. & Agnarsson, I. (2006) Are the Linnean and phylogenetic nomenclatural systems combinable? Recommen-dations for biological nomenclature. Systematic Biology, 55, 774–784.
Laurin, M. (2005) The advantages of phylogenetic nomenclature over Linnean nomenclature. In: Minelli, A., Ortalli, G.& Sanga, G. (Eds.), Animal names, Istituto Veneto di Scienze, Lettere ed Arti, Venezia, pp. 67–97.
Laurin, M. & Cantino, P.D. (2004) First International Phylogenetic Nomenclature Meeting: a report. Zoologica scripta,33, 475–479.
Laurin, M. & Cantino, P.D. (2006) Second Meeting of the International Society for Phylogenetic Nomenclature: a report.Zoologica scripta, 36, 109–117.
Laurin, M., de Queiroz, K. & Cantino, P.D. (2006) Sense and stability in taxon names. Zoologica scripta, 35, 113–114.Lecointre, G., Philippe, H., Lê, H.L.V. & Le Guyader, H. (1993) Species sampling has a major impact on phylogenetic
inference. Molecular Phylogenetics & Evolution, 2, 205–224.Lherminier, P. & Solignac, M. (2000) L’espèce: définitions d’auteurs. Comptes rendus de l’Académie des Sciences de
Paris, Sciences de la Vie, 323, 153–165.Lidén, M., Oxelman, B., Backlund, A., Andersson, L., Bremer, B., Eriksson, R., Moberg, R., Nordal, I., Persson, K.,
Thulin, M. & Zimmer, B. (1997) Charlie is our darling. Taxon, 46, 735–738.Linnaeus, C. (1758) Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum charac-
teribus, differentiis, synonymis, locis. Editio decima, reformata. Tomus I. Laurentii Salvii, Holmiae, 828 pp.Lotsy, J.P. (1916) Evolution by means of hybridization. Martinus Nijhoff, The Hague, 176 pp.Lotsy, J.P. (1918) La quintessence de la théorie du croisement. Archives néerlandaises des Sciences exactes et naturelles,
(3B), 3, 351–353.Mayden, R.L. (1997) A hierarchy of species concepts: the denouement of the saga of the species problem. In: Claridge et
al. (1997), pp. 381–424.Mayr, E. (1982) The growth of biological thought. Belknap Press, Cambridge, Mass. & London, 987 pp.Mayr, E. (1995) Systems of ordering data. Biology & Philosophy, 10, 419–434.Mayr, E. (1997) This is biology. The science of the living world. Belknap Press, Cambridge, Mass. & London, 344 pp.Mayr, E. & Ashlock, P.D. (1991) Principles of systematic zoology. Second edition. McGraw-Hill, New York, 495 pp.McNeill, J., Barrie, F.R., Burdet, H.M., Demoulin, V., Hawksworth, D.L., Marhold, K., Nicolson, D.H., Prado, J., Silva,
P.C., Skog, J.E., Wiersema, J.H. & Turland, N.J., (Eds.) (2006) International code of botanical nomenclature(Vienna Code) adopted by the Seventeenth International Botanical Congress, Vienna, Austria, July 2005. Koeltz,Königstein, 586 pp.
Minelli, A. (1991) Biological systematics: the state of the art. Chapman & Hall, London, 403 pp.Minelli, A. (2000) The ranks and the names of species and higher taxa, or a dangerous inertia of the language of natural
history. In: Ghiselin, M. T. & Leviton, A. E. (Eds.), Cultures and institutions of natural history: essays in the historyand philosophy of sciences, California Academy of Sciences, San Francisco, pp. 339–351.
Moore, G. (1998) A comparison of traditional and phylogenetic nomenclature. Taxon, 47, 561–579.Moore, G. (2003) Should taxon names be explicitly defined? The botanical Review, 69, 2–21.Ohler, A. (2005) Le mode de fonctionnement des systèmes nomenclaturaux en zoologie et en botanique. In: Dubois, A.,
Poncy, O., Malécot, V. & Léger, N. (Eds.), Comment nommer les taxons de rang supérieur en zoologie et en bota-nique?, Biosystema, 23, 11–16.
Padial J.M. & De la Riva, I. (2006) Taxonomic inflation and the stability of species lists: the perils of ostrich’s behavior.Systematic Biology, 55, 859–867.
Papavero, N., Llorente-Bousquets, J. & Abe, J.M. (2001) Proposal of a new system of nomenclature for phylogeneticsystematics. Arquivos de Zoologia, São Paulo, 36, 1–145.
Patterson, C. & Rosen, D.E. (1977) Review of ichthyodectiform and other Mesozoic teleost fishes and the theory andpractice of classifying fossils. Bulletin of the american Museum of natural History, 158, 81–172.
Pennisi, E. (2001) Linnaeus’s last stand? Science, 291, 2304–2305, 2307.Pickett, K.M. (2005) Is the PhyloCode now roughly analogous to the actual codes? A reply to Laurin et al. Cladistics,
21, 608–610.Pleijel, F. & Rouse, G.W. (2000) Least-inclusive taxonomic unit: a new taxonomic concept for biology. Proceedings of
the royal Society of London, (B), 267, 627–630.Pleijel, F. & Rouse, G.W. (2003) Ceci n’est pas une pipe: names, clades and phylogenetic nomenclature. Journal of sys-
tematic and evolutionary Research, 41, 162–174.Polaszek, A. & Wilson, E.O. (2005) Sense and stability in animal names. Trends in Ecology and Evolution, 20, 421–422.

Zootaxa 1519 © 2007 Magnolia Press · 59PHYLOGENY, TAXONOMY AND NOMENCLATURE
Raikow, R.J. (1985) Problems in avian classification. In: Johnston, R. F. (Ed.), Current Ornithology, Vol. 2, Plenum, NewYork, pp. 187–212.
Reif, W.-E. (2003) Problematic issues of cladistics. 1. Ancestor recognition and phylogenetic classification. Neues Jahr-buch für Geologie und Paläontologie Abhandlungen, 230, 97–143.
Rieppel, O. (2006) The PhyloCode: a critical discussion of its theoretical foundation. Cladistics, 22, 186–197. Schaefer, W.W. (1976) The reality of the higher taxonomic categories. Zeitschrift für zoologischer Systematik und Evolu-
tions-Forschung, 14, 1–10.Schuh, R.T. (2003) The Linnaean system and its 250-year persistence. The botanical Review, 69, 59–78.Sibley, C.G. & Ahlquist, J.E. (1982) The relationships of the yellow-breasted chat (Icteria virens) and the alleged slow-
down in the rate of macromolecular evolution in birds. Postilla, 187, 1-19.Sibley, C.G. & Ahlquist, J.E. (1990) Phylogeny and classification of birds. A study in molecular evolution. Yale Univer-
sity Press, New Haven & London, 999 pp.Simpson, G.G. (1940) Types in modern taxonomy. American Journal of Science, 238, 413–431.Simpson, G.G. (1961) Principles of animal taxonomy. Columbia Univ. Press, New York, 259 pp.Sluys, R., Martens, K. & Schram, F.R. (2004) The PhyloCode: naming of biodiversity at a crossroads. Trends in Ecology
and Evolution, 19, 280–281.Smith, A.B. (1988) Patterns of diversification and extinction in early Palaeozoic echinoderms. Palaeontology, 31, 799–
828.Stuart, S.N., Chanson, J.S., Cox, N.A., Young, B.E., Rodrigues, A.S.L., Fischman, D.L. & Waller, R.W. (2004) Status
and trends of amphibian declines and extinctions worldwide. Science, 306, 1783–1786.Stuessy, T.F. (2000) Taxon names are not defined. Taxon, 49, 231–233.Stuessy, T.F. (2001) Taxon names are still not defined. Taxon, 50, 185–186.Sundberg, P. & Pleijel, F. (1994) Phylogenetic classification and the definition of taxon names. Zoologica scripta, 23,
19–25.Turesson, G. (1922) The genotypical response of the plant species to the habitat. Hereditas, 3, 211–350.Van Valen, L. (1973) Are categories in different phyla comparable? Taxon, 22, 333–373.Vuilleumier, F. (1976) La notion d’espèce en ornithologie. In: Bocquet, C., Génermont, J. & Lamotte M. (Eds.), Les
problèmes de l’espèce dans le règne animal, Vol. 1, Mémoires de la Société zoologique de France, 38, 29-65.Wheeler, Q.D. & Meier, R. (Eds.) (2000) Species concepts and phylogenetic theory. Columbia University Press, New
York, 243 pp.Wheeler, Q.D., Raven, P.H. & Wilson, E.O. (2004) Taxonomy: impediment or expedient? Science, 303, 285.White, M.J.D. (1978) Modes of speciation. Freeman, San Francisco, 463 pp.Wiley, E.O. (1981) Phylogenetics. The theory and practice of phylogenetic systematics. Wiley, New York, 454 pp.Wilson, E.O. (1985) The global biodiversity crisis:a challenge to science. Issues in Science & Technology, 2, 20–29. Wilson, E.O. (2004) Taxonomy as a fundamental discipline. In: Godfray & Knapp (2004a), p. 739.

DUBOIS60 · Zootaxa 1519 © 2007 Magnolia Press
Appendix. Cladognoses, apognoses and diagnoses: the example of the family Megophryidae (Amphibia, Anura)
The taxonomy of the family Megophryidae (Amphibia, Anura) recently proposed by Delorme et al. (2006), based on thetree shown in Fig. 2 (which results from both morphological and molecular data), can be taken as an example of the useof cladognoses, apognoses and diagnoses for the same taxa.
For each taxon recognized in a phylogenetic taxonomy, a cladistic hypothesis can be provided in the form either of aphylogenetic tree or of a taxonomic hierarchy. The latter can be expressed as an indented list of taxa or, for each taxon, asa statement of its ametoangiotaxon (taxon immediately superordinate), its ametoxenotaxon/a (taxon/a of same rankimmediately parordinate, i.e., subordinate to the same ametoangiotaxon) and its ametoendotaxon/a (taxon/a immediatelysubordinate) (Dubois 2005a, 2006a). Besides, it can be characterized by its cladognosis, its apognosis and/or its diagno-sis.
A cladognosis is a branch- or stem-based phylogenetic definition (de Queiroz & Gauthier, 1990) of taxon, i.e., usingonly inclusion and exclusion of lower ranked taxa for delimitation of a group considered holophyletic. Such a definitioncan be presented in such a way as allowing also unambiguous allocation of a nomen to this taxon, by bidirectional osten-sion, using both inclusive ostension of onomatophore and exclusive ostension of onomatostase (Dubois 2006c andabove). No characters are mentioned in cladognoses.
FIGURE 2: Phylogenetic tree of the MEGOPHRYIDAE (AMPHIBIA, ANURA) inferred from morphological and moleculardata (Delorme et al. 2006). The numbers on the right preceded by letters designate the genera (G) and subgenera (sG) inthe text. The numbers on the branches correspond to the following taxa: (1) family MEGOPHRYIDAE Bonaparte, 1850(1931); (2) subfamily LEPTOBRACHIINAE Dubois, 1983; (3) tribe LEPTOBRACHIINI Dubois, 1983 and genus LeptobrachiumTschudi, 1838; (4) tribe OREOLALAGINI Tian & Hu, 1985; (5) genus Scutiger Theobald, 1868; (6) subfamily LEPTOLALAG-INAE Delorme, Dubois, Grosjean & Ohler, 2006; (7) genus Leptolalax Dubois, 1980; (8) subfamily MEGOPHRYINAE
Bonaparte, 1850 (1931); (9) tribe XENOPHRYINI Delorme, Dubois, Grosjean & Ohler, 2006; (10) tribe MEGOPHRYINI
Bonaparte, 1850 (1931).

Zootaxa 1519 © 2007 Magnolia Press · 61PHYLOGENY, TAXONOMY AND NOMENCLATURE
An apognosis is an apomorphy-based phylogenetic definition of taxon (de Queiroz & Gauthier, 1990), i.e., relyingon supposed apomorphy/ies shared by members of a group considered holophyletic.
A diagnosis is a definition of taxon based on character states (either apomorphies or plesiomorphies) that are differ-ent from the character states in the parordinate taxon/a or sister-group(s).
The diagnoses of two parordinate taxa are “parallel” or “symmetrical”, therefore approximately of the same length,as they use the same characters, but with different character states in the two taxa. In contrast, apognoses of two parordi-nate taxa usually do not use the same characters, except in the rather rare case where both taxa are characterized by dif-ferent apomorphies for the same character. Apognoses can be of very variable length, some being very short, others verylong, according to the amount of apomorphies identified for the taxon at stake. In the diagnoses and apognoses of thetaxa listed below, only morphological characters are listed, although some of these taxa are also supported by molecularapomorphies (Delorme et al. 2006). Consequently, some taxa recognized below do not have apognoses, because for thetime being they are not known to have morphological apomorphies; in all such cases however, the parordinate taxa arealways characterized by apognoses which apply to all their examined members (see Delorme et al. 2006).
An interesting pattern of this example is that the phylogenetic tree on which the taxonomy is based is not completelyresolved, as it contains two polytomies, including a basal one for the subfamilies LEPTOBRACHIINAE, LEPTOLALAGINAE
and MEGOPHRYINAE. What would happen if this polytomy was resolved, let us say just through the use of molecular data(i.e., without additional morphological characters), and if, for example, the LEPTOLALAGINAE in the taxonomy below wereshown to be the sister-group of the LEPTOBRACHIINAE, and these two groups altogether the sister-group of the MEGOPHRY-INAE, without any other change? Following the Rules of the Code, the changes in the ranks and contents of some taxawould be immediate and automatic, and changes in some of their nomina also. Under such a hypothesis, only two sub-families would remain (LEPTOBRACHIINAE and MEGOPHRYINAE), the first of which would include two tribes (LEPTO-BRACHIINI and LEPTOLALAGINI), the first of which again would contain two subtribes (LEPTOBRACHIINA andOREOLALAGINA). All the genera and subgenera would remain unchanged, as would the morphological apognoses of allthe taxa. However, the diagnoses and the cladognoses of the two new subfamilies, and those of the two tribes of the LEP-TOBRACHIINAE, would have to be changed, as they rely in each case respectively on comparisons between sister-taxa andon inclusions and exclusions in/from these two sister-taxa, and as the patterns of sister-relationships have changed.
The following terms used below were proposed by Dubois (2005a) to avoid the use of the root “type” in nomencla-tural terminology: monophory for monotypy; nucleogenus for type-genus; nucleospecies for type-species.
Family MEGOPHRYIDAE Bonaparte, 1850 (1931)
Subfamily LEPTOBRACHIINAE Dubois, 1983
Nucleogenus. – Leptobrachium Tschudi, 1838, by implicit etymological designation.Ametoangiotaxon. – Family MEGOPHRYIDAE Bonaparte, 1850 (1931).Ametoxenotaxa. – Subfamilies LEPTOLALAGINAE Delorme, Dubois, Grosjean & Ohler, 2006 and MEGOPHRYINAE
Bonaparte, 1850 (1931).Ametoendotaxa. – Tribes LEPTOBRACHIINI Dubois, 1983 and OREOLALAGINI Tian & Hu, 1985.Cladognosis. – The most inclusive holophyletic taxon including the genus Leptobrachium Tschudi, 1838 and
excluding the genera Leptolalax Dubois, 1980 and Megophrys Kuhl & Van Hasselt, 1822.Apognosis. – No known morphological apomorphy, taxon defined only by plesiomorphic character states. Diagnosis. – (1) Inner metacarpal tubercle rounded, not extending on first metacarpal; (2) inner palmar tubercle of
medium size; (3) axillary glands wide and flat, on flanks below arm insertions or on sides of breast; (4) head wide; (5)heels not in contact or just in contact when thighs are placed perpendicular to body; (6) nuptial spines on fingers ofbreeding male absent or present; (7) nostrils of tadpole not tubular, surrounded by an elevated edge; (8) rostrodont of tad-pole keratinised, visible exteriorly; (9) oral disk of tadpole ventral, disk-like; (10) lower labium of oral disk of tadpoleunilobated, with a medial extension; (11) keratodonts of tadpole present; (12) vent of tadpole dextral.
Tribe LEPTOBRACHIINI Dubois, 1983
Nucleogenus. – Leptobrachium Tschudi, 1838, by implicit etymological designation.Ametoangiotaxon. – Subfamily LEPTOBRACHIINAE Dubois, 1983.

DUBOIS62 · Zootaxa 1519 © 2007 Magnolia Press
Ametoxenotaxon. – Tribe OREOLALAGINI Tian & Hu, 1985.Ametoendotaxon. – Genus Leptobrachium Tschudi, 1838.Cladognosis. – The most inclusive holophyletic taxon including the genus Leptobrachium Tschudi, 1838 and
excluding the genus Oreolalax Myers & Leviton, 1962.Apognosis. – (1) Presence of a network on skin of body and limbs; (2) presence of numerous small glands on flank. Diagnosis. – (1) Glands on eyelids absent; (2) canthus rostralis very obtuse, loreal region convex; (3) supratympanic
fold narrow; (4) network on skin of body and limbs present; (5) numerous small glands on flank present; (6) pectoralglands absent; (7) small round and flat glands below arms absent; (8) nuptial spines on fingers of breeding male absent;(9) eggs pigmented.
G1. Genus Leptobrachium Tschudi, 1838
Nucleospecies. – Leptobrachium hasseltii Tschudi, 1838, by original monophory. Ametoangiotaxon. – Tribe LEPTOBRACHIINI Dubois, 1983.Ametoxenotaxon. – None.Ametoendotaxa. – Subgenera Leptobrachium Tschudi, 1838 and Vibrissaphora Liu, 1945.Cladognosis. – Same as for tribe.Apognosis. – Same as for tribe. Diagnosis. – Same as for tribe.
sG1a. Subgenus Leptobrachium Tschudi, 1838
Nucleospecies. – Leptobrachium hasseltii Tschudi, 1838, by original monophory. Ametoangiotaxon. – Genus Leptobrachium Tschudi, 1838.Ametoxenotaxon. – Subgenus Vibrissaphora Liu, 1945.Ametoendotaxa. – Fifteen species: Leptobrachium (Leptobrachium) abbotti (Cochran, 1926); L. (L.) banae Lathrop,
Murphy, Orlov & Ho, 1998; L. (L.) buchardi Ohler, Tenyié & David, 2004; L. (L.) chapaense (Bourret, 1937); L. (L.)gunungense Malkmus, 1996; L. (L.) hainanense Ye & Fei, 1993; L. (L.) hasseltii Tschudi, 1838; L. (L.) hendricksoni Tay-lor, 1962; L. (L.) huashen Fei & Ye, 2005; L. (L.) montanum Fischer, 1885; L. (L.) mouhoti Stuart, Sop & Neang, 2006;L. (L.) nigrops Berry & Hendrickson, 1963; L. (L.) pullum (Smith, 1921); L. (L.) smithi Matsui, Nabhitabhata & Panha,1998; L. (L.) xanthospilum Lathrop, Murphy, Orlov & Ho, 1998.
Cladognosis. – The most inclusive holophyletic taxon including the species Leptobrachium hasseltii Tschudi, 1838and excluding the species Vibrissaphora boringii Liu, 1945.
Apognosis. – No known morphological apomorphy, taxon defined only by plesiomorphic character states. Diagnosis. – (1) Tympanum visible; (2) spines on upper lip of breeding male absent; (3) arms of breeding male not
enlarged; (4) males smaller than females; (5) nostrils of tadpole surmounted by a small middorsal projection.
sG1b. Subgenus Vibrissaphora Liu, 1945
Nucleospecies. – Vibrissaphora boringii Liu, 1945, by original monophory. Ametoangiotaxon. – Genus Leptobrachium Tschudi, 1838.Ametoxenotaxon. – Subgenus Leptobrachium Tschudi, 1838.Ametoendotaxa. – Six species: Leptobrachium (Vibrissaphora) ailaonicum (Yang, Chen & Ma, 1983); L. (V.) bor-
ingii (Liu, 1945); L. (V.) echinatum Dubois & Ohler, 1998; L. (V.) leishanense (Liu & Hu, 1973); L. (V.) liui (Pope, 1947);L. (V.) ngoclinhensis (Orlov, 2005).
Cladognosis. – The most inclusive holophyletic taxon including the species Vibrissaphora boringii Liu, 1945 andexcluding the species Leptobrachium hasseltii Tschudi, 1838.
Apognosis. – (1) Presence of spines on upper lip of breeding male; (2) males smaller than females; (3) nostrils of tad-pole surrounded by ten small projections.
Diagnosis. – (1) Tympanum hidden; (2) spines on upper lip of breeding male present; (3) arms of breeding maleenlarged; (4) males larger than females; (5) nostrils of tadpole surrounded by ten small projections.

Zootaxa 1519 © 2007 Magnolia Press · 63PHYLOGENY, TAXONOMY AND NOMENCLATURE
Tribe OREOLALAGINI Tian & Hu, 1985
Nucleogenus. – Oreolalax Myers & Leviton, 1962, by original designation.Ametoangiotaxon. – Subfamily LEPTOBRACHIINAE Dubois, 1983.Ametoxenotaxon. – Tribe LEPTOBRACHIINI Dubois, 1983.Ametoendotaxa. – Genera Oreolalax Myers & Leviton, 1962 and Scutiger Theobald, 1868.Cladognosis. – The most inclusive holophyletic taxon including the genus Oreolalax Myers & Leviton, 1962 and
excluding the genus Leptobrachium Tschudi, 1838.Apognosis. – Presence of pectoral glands.Diagnosis. – (1) Glands on eyelids present; (2) canthus rostralis little distinct, loreal region vertical or slightly con-
cave; (3) supratympanic fold wide; (4) network on skin of body and limbs absent; (5) numerous small glands on flankabsent; (6) pectoral glands present; (7) small round and flat glands below arms present; (8) nuptial spines on fingers ofbreeding male present; (9) eggs unpigmented.
G2. Genus Oreolalax Myers & Leviton, 1962
Nucleospecies. – Scutiger pingii Liu, 1943, by original designation. Ametoangiotaxon. – Tribe OREOLALAGINI Tian & Hu, 1985. Ametoxenotaxon. – Genus Scutiger Theobald, 1868.Ametoendotaxa. – Seventeen species: Oreolalax chuanbeiensis Tian, 1983; O. granulosus Fei, Ye & Chen, 1991; O.
jingdongensis Ma, Yang & Li, 1983; O. liangbeiensis Liu & Fei, 1979; O. lichuanensis Hu & Fei, 1979; O. major (Liu &Hu, 1960); O. multipunctatus Wu, Zhao, Inger & Schaffer, 1993; O. nanjiangensis Fei & Ye, 1999; O. omeimontis (Liu& Hu, 1960); O. pingii (Liu, 1943); O. popei (Liu, 1947); O. puxiongensis Liu & Fei, 1979; O. rhodostigmatus Hu & Fei,1979; O. rugosus (Liu, 1943); O. schmidti (Liu, 1947); O. weigoldi (Vogt, 1924); O. xiangchengensis Fei & Huang,1983.
Cladognosis. – The most inclusive holophyletic taxon including the species Oreolalax pingii (Liu, 1943) and exclud-ing the species Scutiger sikimmensis (Blyth, 1854).
Apognosis. – No known morphological apomorphy, taxon defined only by plesiomorphic character states. Diagnosis. – (1) Tympanic annulus present; (2) supratympanic fold moderately wide; (3) maxillary teeth present,
well developed; (4) axillary glands on flanks below arm insertion; (5) nuptial spines of finger I of breeding male extend-ing over forearm; (6) heels in contact when thighs are placed perpendicular to body; (7) femoral glands present; (8) nos-trils of tadpole surmounted by two or three small middorsal projections.
G3. Genus Scutiger Theobald, 1868
Nucleospecies. – Bombinator sikimmensis Blyth, 1854, by original monophory. Ametoangiotaxon. – Tribe OREOLALAGINI Tian & Hu, 1985. Ametoxenotaxon. – Genus Oreolalax Myers & Leviton, 1962.Ametoendotaxa. – Subgenera Aelurophryne Boulenger, 1919 and Scutiger Theobald, 1868.Cladognosis. – The most inclusive holophyletic taxon including the species Scutiger sikimmensis (Blyth, 1854) and
excluding the species Oreolalax pingii (Liu, 1943).Apognosis. – (1) Tympanic annulus absent; (2) supratympanic fold very wide, glandular. Diagnosis. – (1) Tympanic annulus absent; (2) supratympanic fold very wide, glandular; (3) maxillary teeth absent,
or present but vestigial; (4) axillary glands on sides of breast; (5) nuptial spines of finger I of breeding male extending tofirst articulation of finger I; (6) heels far apart when thighs are placed perpendicular to body; (7) femoral glands absent;(8) nostrils of tadpole surmounted by a single middorsal projection.
sG3a. Subgenus Aelurophryne Boulenger, 1919
Nucleospecies. – Bufo mammatus Günther, 1896, by original monophory. Ametoangiotaxon. – Genus Scutiger Theobald, 1868.

DUBOIS64 · Zootaxa 1519 © 2007 Magnolia Press
Ametoxenotaxon. – Subgenus Scutiger Theobald, 1868.Ametoendotaxa. – Eight species: Scutiger (Aelurophryne) adungensis Dubois, 1979; S. (A.) bhutanensis Delorme &
Dubois, 2001; S. (A.) glandulatus (Liu, 1950); S. (A.) gongshanensis Yang & Su, 1978; S. (A.) jiulongensis Fei, Ye &Jiang, 1996; S. (A.) mammatus (Günther, 1896); S. (A.) muliensis Fei & Ye, 1986; S. (A.) tuberculatus Liu & Fei, 1979.
Cladognosis. – The most inclusive holophyletic taxon including the species Scutiger mammatus (Günther, 1896) andexcluding the species Scutiger sikimmensis (Blyth, 1854).
Apognosis. – Nuptial spines on fingers of breeding male very large. Diagnosis. – (1) Pectoral glands larger than axillary glands; (2) nuptial spines on fingers of breeding male very large;
(3) nuptial spines on finger III of breeding male absent.
sG3b. Subgenus Scutiger Theobald, 1868
Nucleospecies. – Bombinator sikimmensis Blyth, 1854, by original monophory. Ametoangiotaxon. – Genus Scutiger Theobald, 1868. Ametoxenotaxon. – Subgenus Aelurophryne Boulenger, 1919.Ametoendotaxa. – Ten species: Scutiger (Scutiger) boulengeri (Bedriaga, 1898); S. (S.) chintingensis Liu & Hu,
1960; S. (S.) liupanensis Huang, 1985; S. (S.) maculatus (Liu, 1950); S. (S.) nepalensis Dubois, 1974; S. (S.) ningshanen-sis Fang, 1985; S. (S.) nyingchiensis Fei, 1977; S. (S.) occidentalis Dubois, 1978; S. (S.) pingwuensis Liu & Tian, 1978;S. (S.) sikimmensis (Blyth, 1854).
Cladognosis. – The most inclusive holophyletic taxon including the species Scutiger sikimmensis (Blyth, 1854) andexcluding the species Scutiger mammatus (Günther, 1896).
Apognosis. – Nuptial spines on finger III of breeding male present, similar to those on finger I. Diagnosis. – (1) Pectoral glands of same size as axillary glands; (2) nuptial spines on fingers of breeding male rather
small; (3) nuptial spines on finger III of breeding male present, similar to those on finger I.
Subfamily LEPTOLALAGINAE Delorme, Dubois, Grosjean & Ohler, 2006
Nucleogenus. – Leptolalax Dubois, 1980, by original designation.Ametoangiotaxon. – Family MEGOPHRYIDAE Bonaparte, 1850 (1931).Ametoxenotaxa. – Subfamilies LEPTOBRACHIINAE Dubois, 1983 and MEGOPHRYINAE Bonaparte, 1850 (1931).Ametoendotaxa. – Genera Leptobrachella Smith, 1925 and Leptolalax Dubois, 1980.Cladognosis. – The most inclusive holophyletic taxon including the genus Leptolalax Dubois, 1980 and excluding
the genera Leptobrachium Tschudi, 1838 and Megophrys Kuhl & Van Hasselt, 1822.Apognosis. – (1) Head narrow, longer than wide; (2) outer metacarpal tubercle rounded, much smaller than inner
metacarpal tubercle; (3) oral disk of tadpole ventral, cup-like; (4) both labia of oral disk of tadpole bilobated; (5) a spacebetween marginal and submarginal papillae of oral disk of tadpole; (6) eight pairs or more of median papillae on ventralvelum of tadpole; (7) two groups of pustules posteriorly ta arena of buccal roof of tadpole.
Diagnosis. – (1) Inner metacarpal tubercle rounded, not extending on first metacarpal; (2) inner palmar tubercle oflarge size; (3) axillary glands wide and flat or small and tit-like, on sides of breast; (4) head narrow; (5) heels largelyoverlapping when thighs are placed perpendicular to body; (6) nuptial spines on fingers of breeding male absent; (7) nos-trils of tadpole not tubular, surrounded by an elevated edge; (8) rostrodont of tadpole keratinised, visible exteriorly; (9)oral disk of tadpole ventral, cup-like; (10) lower labium of oral disk of tadpole bilobated; (11) keratodonts of tadpolepresent or absent; (12) vent of tadpole dextral.
G4. Genus Leptobrachella Smith, 1925
Nucleospecies. – Leptobrachium natunae Günther, 1895, by original monophory.Ametoangiotaxon. – Subfamily LEPTOLALAGINAE Delorme, Dubois, Grosjean & Ohler, 2006.Ametoxenotaxon. – Genus Leptolalax Dubois, 1980.Ametoendotaxa. – Seven species: Leptobrachella baluensis Smith, 1931; L. brevicrus Dring, 1984; L. mjobergi
Smith, 1925; L. natunae (Günther, 1895); L. palmata Inger & Stuebing, 1992; L. parva Dring, 1984; L. serasanae Dring,1984.

Zootaxa 1519 © 2007 Magnolia Press · 65PHYLOGENY, TAXONOMY AND NOMENCLATURE
Cladognosis. – The most inclusive holophyletic taxon including the species Leptobrachella natunae (Günther, 1895)and excluding the species Leptolalax gracilis (Günther, 1872).
Apognosis. – (1) Adult of very small size; (2) finger II much longer than finger I; (3) tips of digits with lanceolatedisks; (4) rostrodont of tadpole weakly keratinised, flexible.
Diagnosis. – (1) Glands on eyelids absent; (2) axillary glands small and tit-like; (3) finger II much longer than fingerI; (4) tips of digits with lanceolate disks; (5) supernumerary tubercules under toes absent; (6) rostrodont of tadpoleweakly keratinised, flexible; (7) keratodonts of tadpole absent.
G5. Genus Leptolalax Dubois, 1980
Nucleospecies. – Leptobrachium gracile Günther, 1872, by original designation.Ametoangiotaxon. – Subfamily LEPTOLALAGINAE Delorme, Dubois, Grosjean & Ohler, 2006.Ametoxenotaxon. – Genus Leptobrachella Smith, 1925.Ametoendotaxa. – Subgenera Lalax Delorme, Dubois, Grosjean & Ohler, 2006 and Leptolalax Dubois, 1980. Cladognosis. – The most inclusive holophyletic taxon including the species Leptolalax gracilis (Günther, 1872) and
excluding the species Leptobrachella natunae (Günther, 1895).Apognosis. – Supernumerary tubercles present under toes, absent under fingers. Diagnosis. – (1) Glands on eyelids present; (2) axillary glands wide and flat; (3) fingers II and I subequal; (4) tips of
digits rounded; (5) supernumerary tubercules under toes present; (6) rostrodont of tadpole very keratinised, breakable;(7) keratodonts of tadpole present.
sG5a. Subgenus Lalax Delorme, Dubois, Grosjean & Ohler, 2006
Nucleospecies. – Leptolalax bourreti Dubois, 1983, by original designation.Ametoangiotaxon. – Genus Leptolalax Dubois, 1980.Ametoxenotaxon. – Subgenus Leptolalax Dubois, 1980.Ametoendotaxa. – Eleven species: Leptolalax (Lalax) alpinus Fei, Ye & Li, 1991; L. (L.) bourreti Dubois, 1983; L.
(L.) lateralis (Anderson, 1871); L. (L.) liui Fei & Ye, 1991; L. (L.) nahangensis Lathrop, Murphy, Orlov & Ho, 1998; L.(L.) oshanensis (Liu, 1950); L. (L.) pelodytoides (Boulenger, 1893); L. (L.) pluvialis Ohler, Marquis, Swan & Grosjean,2000; L. (L.) sungi Lathrop, Murphy, Orlov & Ho, 1998; L. (L.) tuberosus Inger, Orlov & Darevsky, 1999; L. (L.) ven-tripunctatus Fei, Ye & Li, 1991.
Cladognosis. – The most inclusive holophyletic taxon including the species Leptolalax bourreti Dubois, 1983 andexcluding the species Leptolalax gracilis (Günther, 1872).
Apognosis. – (1) Latero-ventral glandular fold present; (2) nostrils of tadpole surrounded by four lobes in juxtaposi-tion.
Diagnosis. – (1) Latero-ventral glandular fold present; (2) nostrils of tadpole surrounded by four lobes in juxtaposi-tion; (3) rostrodont of tadpole bearing coarse denticulations.
sG5b. Subgenus Leptolalax Dubois, 1980
Nucleospecies. – Leptobrachium gracile Günther, 1872, by original designation.Ametoangiotaxon. – Genus Leptolalax Dubois, 1980.Ametoxenotaxon. – Subgenus Lalax Delorme, Dubois, Grosjean & Ohler, 2006.Ametoendotaxa. – Eight species: Leptolalax (Leptolalax) arayai Matsui, 1997; L. (L.) dringi Dubois, 1987; L. (L.)
gracilis (Günther, 1872); L. (L.) hamidi Matsui, 1997; L. (L.) heteropus Boulenger, 1900; L. (L.) kajangensis Grismer,Grismer & Youmans, 2004; L. (L.) maurus Inger, Lakim, Biun & Yambun, 1997; L. (L.) pictus Malkmus, 1992.
Cladognosis. – The most inclusive holophyletic taxon including the species Leptolalax gracilis (Günther, 1872) andexcluding the species Leptolalax bourreti Dubois, 1983.
Apognosis. – No known morphological apomorphy, taxon defined only by plesiomorphic character states. Diagnosis. – (1) Latero-ventral glandular fold absent; (2) nostrils of tadpole surmounted by a middorsal projection;
(3) rostrodont of tadpole bearing fine denticulations.

DUBOIS66 · Zootaxa 1519 © 2007 Magnolia Press
Subfamily MEGOPHRYINAE Bonaparte, 1850 (1931)
Nucleogenus. – Megophrys Kuhl & Van Hasselt, 1822, by implicit original designation (as Megalophrys Wagler, 1930).Ametoangiotaxon. – Family MEGOPHRYIDAE Bonaparte, 1850 (1931).Ametoxenotaxa. – Subfamilies LEPTOBRACHIINAE Dubois, 1983 and LEPTOLALAGINAE Delorme, Dubois, Grosjean &
Ohler, 2006.Ametoendotaxa. – Tribes MEGOPHRYINI Bonaparte, 1850 (1931) and XENOPHRYINI Delorme, Dubois, Grosjean &
Ohler, 2006.Cladognosis. – The most inclusive holophyletic taxon including the genus Megophrys Kuhl & Van Hasselt, 1822 and
excluding the genera Leptobrachium Tschudi, 1838 and Leptolalax Dubois, 1980.Apognosis. – (1) Presence of two clear lines connecting lower jaw and arm; (2) two fine glandular crests on back; (3)
inner metatarsal tubercle elongated, extending on first metacarpal; (4) outer palmar tubercle elongated and little distinct;(5) nuptial pads on inner dorsal side of fingers I et II of breeding male, extending to wrist of finger I, less extended onfinger II; (6) nostrils of tadpole dorso-lateral, closer to eyes than to tip of snout, in shape of elongated external tubes; (7)tip of tail of tadpole rounded; (8) oral disk of tadpole funnel-shaped, in dorsal position, entirely surrounded by minutepapillae; (9) tadpole with ridges on labia and a flexible rostrodont, bearing denticulations in shape of comb teeth; (10)tadpole with prelingual papillae undigitated and convex; (11) tadpole without papillae on arena of buccal floor, with lon-gitudinal ridges delimiting anteriorly this arena, with veils covering posterior part of longitudinal ridges, without medianpapillae on ventral velum, with a U-shaped prenarial ridge, a thickening of anterior wall of choanae, postnarial papillaelinked to a ridge, very long lateral ridges, a phallic-shaped median ridge, excrescences continuing the longitudinal ridges,and inside of arena of buccal roof smooth.
Diagnosis. – (1) Inner metacarpal tubercle oval, extending on first metacarpal; (2) inner palmar tubercle of mediumsize; (3) axillary glands small and tit-like, on sides of breast; (4) head wide or narrow; (5) heels not in contact, in contactor overlapping when thighs are placed perpendicular to body; (6) nuptial spines on fingers of breeding male present; (7)nostrils of tadpole tubular and elongated; (8) rostrodont of tadpole not keratinised, not visible exteriorly; (9) oral disk oftadpole dorsal, funnel-like, with lateral extensions and crests; (10) lower labium of oral disk of tadpole unilobated; (11)keratodonts of tadpole absent; (12) vent of tadpole medial.
Tribe MEGOPHRYINI Bonaparte, 1850 (1931)
Nucleogenus. – Megophrys Kuhl & Van Hasselt, 1822, by implicit original designation (as Megalophrys Wagler, 1930).Ametoangiotaxon. – Subfamily MEGOPHRYINAE Bonaparte, 1850 (1931).Ametoxenotaxon. – Tribe XENOPHRYINI Delorme, Dubois, Grosjean & Ohler, 2006.Ametoendotaxa. – Genera Borneophrys Delorme, Dubois, Grosjean & Ohler, 2006, Brachytarsophrys Tian & Hu,
1983 and Megophrys Kuhl & Van Hasselt, 1822.Cladognosis. – The most inclusive holophyletic taxon including the genus Megophrys Kuhl & Van Hasselt, 1822 and
excluding the genus Xenophrys Günther, 1864.Apognosis. – (1) Skin fold on the back of head and thickening of head skin present; (2) glandular spine-like excres-
cences at lips commissure and on back present; (3) two fine parallel glandular lines on the sides of back. Diagnosis. – (1) Head very wide; (2) skin fold on the back of head and thickening of head skin present; (3) glandular
spine-like excrescences present at lips commissure and on back present; (4) two fine parallel glandular lines on the sidesof back.
G6. Genus Borneophrys Delorme, Dubois, Grosjean & Ohler, 2006
Nucleospecies. – Megophrys edwardinae Inger, 1989, by original designation.Ametoangiotaxon. – Tribe MEGOPHRYINI Bonaparte, 1850 (1931).Ametoxenotaxa. – Genera Brachytarsophrys Tian & Hu, 1983 and Megophrys Kuhl & Van Hasselt, 1822.Ametoendotaxon. – One species: Borneophrys edwardinae (Inger, 1989).Cladognosis. – The most inclusive holophyletic taxon including the species Borneophrys edwardinae (Inger, 1989)
and excluding the species Brachytarsophrys carinensis (Boulenger, 1889) and Megophrys montana Kuhl & Van Hasselt,1822.

Zootaxa 1519 © 2007 Magnolia Press · 67PHYLOGENY, TAXONOMY AND NOMENCLATURE
Apognosis. – No known morphological apomorphy, taxon defined only by plesiomorphic character states. Diagnosis. – (1) Snout projecting beyond lower jaw; (2) eyelid projections composed of two small tubercles; (3)
vomerine teeth absent; (4) tympanum present, very small; (5) gland in contact with supratympanic glandular foldpresent; (6) hind limbs rather thin.
G7. Genus Brachytarsophrys Tian & Hu, 1983
Nucleospecies. – Leptobrachium carinense Boulenger, 1889, by original designation.Ametoangiotaxon. – Tribe MEGOPHRYINI Bonaparte, 1850 (1931).Ametoxenotaxa. – Genera Borneophrys Delorme, Dubois, Grosjean & Ohler, 2006 and Megophrys Kuhl & Van Has-
selt, 1822.Ametoendotaxa. – Five species: Brachytarsophrys carinensis (Boulenger, 1889); B. chuannanensis Fei, Ye &
Huang, 2001; B. feae (Boulenger, 1887); B. intermedia (Smith, 1921); B. platyparietus Rao & Yang, 1997. Cladognosis.– The most inclusive holophyletic taxon including the species Brachytarsophrys carinensis (Boulenger,
1889) and excluding the species Borneophrys edwardinae (Inger, 1989) and Megophrys montana Kuhl & Van Hasselt,1822.
Apognosis. – (1) Snout not projecting beyond lower jaw; (2) hind limbs massive. Diagnosis. – (1) Snout not projecting beyond lower jaw; (2) eyelid projections composed of three to five small tuber-
cles; (3) vomerine teeth present; (4) tympanum absent; (5) gland in contact with supratympanic glandular fold present;(6) hind limbs massive.
G8. Genus Megophrys Kuhl & Van Hasselt, 1822
Nucleospecies. – Megophrys montana Kuhl & Van Hasselt, 1822, by original monophory.Ametoangiotaxon. – Tribe MEGOPHRYINI Bonaparte, 1850 (1931).Ametoxenotaxa. – Genera Borneophrys Delorme, Dubois, Grosjean & Ohler, 2006 and Brachytarsophrys Tian &
Hu, 1983.Ametoendotaxa. – Five species: Megophrys kobayashii Malkmus & Matsui, 1997; M. ligayae (Taylor, 1920); M.
montana Kuhl & Van Hasselt, 1822; M. nasuta Schlegel, 1858; M. stejnegeri (Taylor, 1920).Cladognosis. – The most inclusive holophyletic taxon including the species Megophrys montana Kuhl & Van Has-
selt, 1822 and excluding the species Borneophrys edwardinae (Inger, 1989) and Brachytarsophrys carinensis (Bou-lenger, 1889).
Apognosis. – (1) Snout bearing a pointed projection; (2) eyelid projection unique and long. Diagnosis. – (1) Snout not projecting beyond lower jaw; (2) eyelid projection unique and long; (3) vomerine teeth
present; (4) tympanum present, very small; (5) gland in contact with supratympanic glandular fold absent; (6) hind limbsrather thin.
Tribe XENOPHRYINI Delorme, Dubois, Grosjean & Ohler, 2006
Nucleogenus. – Xenophrys Günther, 1864, by original designation.Ametoangiotaxon. – Subfamily MEGOPHRYINAE Bonaparte, 1850 (1931).Ametoxenotaxon. – Tribe MEGOPHRYINI Bonaparte, 1850 (1931).Ametoendotaxa. – Genera Ophryophryne Boulenger, 1903 and Xenophrys Günther, 1864.Cladognosis. – The most inclusive holophyletic taxon including the genus Xenophrys Günther, 1864 and excluding
the genus Megophrys Kuhl & Van Hasselt, 1822.Apognosis. – No known morphological apomorphy, taxon defined only by plesiomorphic character states. Diagnosis. – (1) Head as long as wide; (2) skin fold on the back of head and thickening of head skin absent; (3) glan-
dular spine-like excrescences at lips commissure and on back absent; (4) two fine glandular lines on back forming a moreor less complete Y, H or X.

DUBOIS68 · Zootaxa 1519 © 2007 Magnolia Press
G9. Genre Ophryophryne Boulenger, 1903
Nucleospecies. – Ophryophryne microstoma Boulenger, 1903, by original monophory.Ametoangiotaxon. – Tribe XENOPHRYINI Delorme, Dubois, Grosjean & Ohler, 2006.Ametoxenotaxon. – Genus Xenophrys Günther, 1864.Ametoendotaxa. – Five species: Ophryophryne gerti Ohler, 2003; O. hansi Ohler, 2003; O. microstoma Boulenger,
1903; O. pachyproctus Kou, 1985; O. synoria Stuart, Sok & Neang, 2006. Cladognosis. – The most inclusive holophyletic taxon including the species Ophryophryne microstoma Boulenger,
1903 and excluding the species Xenophrys parva (Boulenger, 1893).Apognosis. – (1) Head narrow (less than 25 % of snout-vent length); (2) tip of snout truncated without dermal projec-
tion; (3) pupil horizontal; (4) maxillary teeth absent. Diagnosis. – (1) Head narrow (less than 25 % of snout-vent length); (2) snout truncated, without crests between nos-
trils and tip of snout; (3) pupil horizontal.
G10. Genus Xenophrys Günther, 1864
Nucleospecies. – Xenophrys monticola Günther, 1864, by original monophory.Ametoangiotaxon. – Tribe XENOPHRYINI Delorme, Dubois, Grosjean & Ohler, 2006.Ametoxenotaxon. – Genus Ophryophryne Boulenger, 1903.Ametoendotaxa. – Thirty-three species: Xenophrys aceras (Boulenger, 1903); X. auralensis (Ohler, Swan & Daltry,
2002); X. baluensis (Boulenger, 1899); X. binchuanensis (Ye & Fei, 1995); X. boettgeri (Boulenger, 1899); X. brachyko-los (Inger & Romer, 1961); X. caudoprocta (Shen, 1994); X. daweimontis (Rao & Yang, 1997); X. dringi (Inger, Stuebing& Tan, 1995); X. gigantica (Liu, Hu & Yang, 1960); X. glandulosa (Fei, Ye & Huang, 1991); X. huangshanensis (Fei &Ye, 2005); X. jingdongensis (Fei & Ye, 1983); X. kuatunensis (Pope, 1929); X. longipes (Boulenger, 1886); X. major(Boulenger, 1908); X. mangshanensis (Fei & Ye, 1991); X. medogensis (Fei, Ye & Huang, 1983); X. minor (Stejneger,1926); X. nankiangensis (Liu & Hu, 1966); X. omeimontis (Liu, 1950); X. pachyproctus (Huang, 1981); X. palpebralespi-nosa (Bourret, 1937); X. parallelus (Inger & Iskandar, 2005); X. parva (Boulenger, 1893); X. robusta (Boulenger, 1908);X. shapingensis (Liu, 1950); X. shuichengensis (Tian, Gu & Sun, 2000); X. spinata (Liu & Hu, 1973); X. wawuensis (Fei,Jiang & Zheng, 2001); X. wuliangshanensis (Ye & Fei, 1995); X. wushanensis (Ye & Fei, 1995); X. zhangi (Ye & Fei,1992).
Cladognosis. – The most inclusive holophyletic taxon including the species Xenophrys parva (Boulenger, 1893) andexcluding the species Ophryophryne microstoma Boulenger, 1903.
Apognosis. – No known morphological apomorphy, taxon defined only by plesiomorphic character states. Diagnosis. – (1) Head wide (more than 25 % of snout-vent length); (2) snout not truncated, with crests between nos-
trils and tip of snout; (3) pupil vertical.




















