2 3 Responses of epibenthic algal assemblages to water abstraction in Hong Kong streams
15
1 23 Hydrobiologia The International Journal of Aquatic Sciences ISSN 0018-8158 Volume 703 Number 1 Hydrobiologia (2013) 703:225-237 DOI 10.1007/s10750-012-1362-z Responses of epibenthic algal assemblages to water abstraction in Hong Kong streams Tao Tang, Sophia Qian Niu & David Dudgeon
Transcript of 2 3 Responses of epibenthic algal assemblages to water abstraction in Hong Kong streams

1 23
HydrobiologiaThe International Journal of AquaticSciences ISSN 0018-8158Volume 703Number 1 Hydrobiologia (2013) 703:225-237DOI 10.1007/s10750-012-1362-z
Responses of epibenthic algal assemblagesto water abstraction in Hong Kong streams
Tao Tang, Sophia Qian Niu & DavidDudgeon

1 23
Your article is protected by copyright and all
rights are held exclusively by Springer Science
+Business Media Dordrecht. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

PRIMARY RESEARCH PAPER
Responses of epibenthic algal assemblages to waterabstraction in Hong Kong streams
Tao Tang • Sophia Qian Niu • David Dudgeon
Received: 23 August 2012 / Accepted: 22 October 2012 / Published online: 1 November 2012
� Springer Science+Business Media Dordrecht 2012
Abstract We studied the effects of flow reduction on
epibenthic algal assemblages by comparing up- and
down-stream reaches of ten Hong Kong streams
subject to different degrees of water abstraction during
2007–2008. Downstream discharge declined by 71
and 54% during the wet and dry seasons, respectively.
Algal responses varied seasonally and according to
morphological guild and reflecting reductions in
discharge and current velocity, or changes in total
nitrogen and phosphate, or their combined effects.
Significant inter-reach assemblage differences were
observed during the both seasons, but wet-season
assemblage structure was not directly influenced by
any flow-related variable. During the dry season,
scouring-tolerant prostrate/adnate diatoms (especially
Achnanthes, Cocconeis) were relatively abundant
upstream, whereas intolerant stalked (Eunotia, Gom-
phonema) and mobile diatoms were more numerous
downstream. Both low- and high-profile (erect dia-
toms, Chamaesiphon, Calothrix) guilds were sensitive
to changes in nutrients. Low-flow conditions down-
stream, surprisingly, enhanced algal diversity, and
richness. Based on the linear response of Cocconeis
(mainly C. placentula) to discharge reduction, we
recommend that[65% of dry-season discharge should
be maintained in downstream reaches in order to
sustain near-natural algal assemblages; 32% of dis-
charge is required to avoid substantial alterations in
assemblage structure.
Keywords Flow reduction � Hydro–ecological
relationship � Diatoms � Morphological guilds �Hierarchical partitioning � Environmental flow
allocations
Introduction
Instream flows are fundamental drivers of spatiotem-
poral patterns and functions in lotic epibenthic algal
communities (Martınez de Fabricius et al., 2003;
Uehlinger et al., 2003; Luce et al., 2010). However,
globally many rivers and streams have been subjected
Handling editor: Judit Padisak
Electronic supplementary material The online version ofthis article (doi:10.1007/s10750-012-1362-z) containssupplementary material, which is available to authorized users.
T. Tang (&)
Institute of Hydrobiology, Chinese Academy of Sciences,
Wuhan 430072, People’s Republic of China
e-mail: [email protected]
S. Q. Niu
Department of Biology, Saint Louis University,
St. Louis, MO, USA
e-mail: [email protected]
D. Dudgeon
Chair of Ecology and Biodiversity, School of Biological
Sciences, The University of Hong Kong, Hong Kong
SAR, People’s Republic of China
e-mail: [email protected]
123
Hydrobiologia (2013) 703:225–237
DOI 10.1007/s10750-012-1362-z
Author's personal copy

to hydrologic regulation and dam construction for
various purposes (e.g., March et al., 2003; Nilsson
et al., 2005), causing diversity declines and composi-
tional changes in aquatic taxa. Such responses have
been widely documented to inform management
decisions about the implementation of environmental
flows; i.e., the flows left in streams to sustain
biodiversity, ecological integrity and ecosystems
service (Acreman & Dunbar, 2004; Brisbane Decla-
ration, 2007; Arthington et al., 2010). Nonetheless
studies on the relationships between flow reduction
and responses of stream algal assemblages are scarce
compared with those dealing with macroinvertebrates
and fishes, and most such studies are focused mainly
on single streams in temperate or subtropical latitudes
(Clausen & Biggs, 1997; Downes et al., 2003;
Soininen, 2003; Suren et al., 2003; Wang et al.,
2009; Wu et al., 2009). Comparable studies on tropical
or monsoonal streams—with markedly seasonal flow
regimes—are scarce (Niu & Dudgeon, 2011a).
Responses of epibenthic algal assemblages to
altered flow regimes can be mediated by biotic as
well as abiotic processes. Physical shear stress
imposed by current strongly influences the relative
abundance of algae with different growth forms or
morphologies (e.g., erect, prostrate, or filamentous)
and also affects colonization dynamics, thereby influ-
encing species composition (Stevenson, 1996; Hart &
Finelli, 1999). With reduced current speed and flow
volume, algal community development is less con-
strained by shear stress, and compositional shifts can
take place from, for instance, low-stature diatoms to
nuisance growths of filamentous green algae, although
the precise outcome depends on nutrients and temper-
ature regimes (Biggs & Price, 1987; Suren et al., 2003;
Berthon et al., 2011). The strength of grazing pressure
upon epibenthic algae can vary according to flow
conditions (Poff & Ward, 1995; Chester & Norris,
2006), potentially influencing the intensity of top–
down control of assemblage composition and biomass
(Opsahl et al., 2003). Because of the complex
interactions among abiotic factors, such as discharge
and nutrients, and biotic factors (Peterson & Steven-
son, 1992; Ledger et al., 2008), the effects of flow on
epibenthic algal assemblages vary substantially
according to stream conditions (Jowett & Duncan,
1990; Downes et al., 2003; Wu et al., 2010).
Hong Kong streams suffer extensive and aggressive
water extraction to meet the demands of municipal
users, with no apparent concern for downstream impacts
or consideration of the need to sustain in-stream
environmental flows (Dudgeon, 1996). Reduced flow
or even dewatering may occur in downstream reaches
(typically at elevations below 100 m) especially during
the dry season, leading to increased algal biomass,
declines in macroinvertebrate species richness and
compositional changes, as well as decreased litter
breakdown rates (Dudgeon, 1992; Niu & Dudgeon,
2011a, b). This occurs against a background of seasonal
variation in epibenthic algal assemblages in stream
reaches that do not experience water extractions. These
may be induced by flow (Yang et al., 2009) as well as
top–down control of biomass and composition by
grazing fish (Yang & Dudgeon, 2010b). In this study,
we compared epibenthic algal assemblages between up-
and down-stream reaches of a number of streams
experiencing varying degrees of water abstraction. We
attempted to test the hypotheses that: (1) there will be
substantial differences in assemblage composition in
terms of both taxonomic diversity and growth forms—
between upstream (unaffected) and downstream
reaches; (2) this difference would vary seasonally due
to the monsoonal climate; and (3) the magnitude of algal
assemblage changes would be related to the magnitude
or degree of flow reduction.
Methods
Study sites and sampling design
We investigated 11 second- to fourth-order stony
hillstreams (see Supplementary material—Appendix 1
for detail). Sampling was conducted at approximately
monthly intervals on six occasions: three during the
wet season in 2007 (June to September 2007) and three
during the following dry season (December 2007 to
March 2008). Most streams were situated within or
adjacent to country parks (conservation areas), and
vegetation cover was mainly secondary forests and
shrubland. Although all streams were unpolluted (Niu
& Dudgeon, 2011a, b give details), they experienced
various degrees of water extraction. Ten streams, all
ext Shek Mun Kap Stream (SMK), were sampled in
the 2007 wet season. Wong Lung Hang stream (WLH)
was abandoned and substituted by adjacent SMK in
following dry season due to a fish-poisoning episode
on January 2008. Therefore, only wet season data from
226 Hydrobiologia (2013) 703:225–237
123
Author's personal copy

WLH and only dry-season data from SMK were used,
resulting in datasets for ten streams for both seasons.
At each stream, we selected two reaches 100–200 m
apart with comparable physical conditions and stream-
bed characteristics: one above (upstream) and one
below (downstream) a water abstraction point (a small
dam or catchment channel). The upstream reach served
as a reference site with natural flow conditions, and
downstream reach received reduced flows. To avoid
direct impacts arising from water impoundment or
scouring, both reaches were least 50 m away from
dams or water-abstraction infrastructure.
Measurements of flow and physicochemical
variables
At each sampling occasion and each study reach, flow
parameters were measured from three representative
cross-sections (at least 15 m apart), and a water sample
was collected. We measured wetted width, and water
depths at 50-cm intervals and current velocities at
10-cm intervals (at 40% vertical water depth above the
bottom) using a Swoffer Model 2100 current meter
(Swoffer Instruments Inc., Seattle, Washington, USA).
Discharge (Q in l s-1), the volume of flow moving past
a given cross-section per second, was calculated using
the velocity-wetted-area method (Gore, 2006), and the
mean value based on three cross-sections was used to
characterise each reach. Water samples were collected
with an opaque Nalgene bottle and transported to the
laboratory in a cool-box, where dissolved oxygen
(DO), conductivity, and pH were measured as soon as
possible using YSI 85 and YSI pH 100 m meters (YSI
Inc., Yellow Springs, Ohio, USA). Water samples for
nutrient measurements (nitrate, nitrite, ammonia com-
bines as total inorganic nitrogen [TN]; phosphate
[PO4-P]) were collected from both reaches of each
study stream once during each season. Nutrients were
analysed following APHA (1995) using the automated
phenate method for ammoniacal-N, and colorimetric
methods for nitrate-N, nitrite-N, and PO4-P on a flow
injection analyser (QuickChem 8000; Lachat Instru-
ments, Inc., Milwaukee, Wisconsin, USA).
Sampling and laboratory procedures for epibenthic
algae
In each reach, epibenthic algae were collected from
three randomly selected stones of 10–15 cm diameter
(Stewart et al., 2005; Yang & Dudgeon, 2010a; Niu &
Dudgeon, 2011a). Each stone was brushed vigorously
with a brass-bristled brush to remove all attached
periphyton, and the three perpendicular parameters
measured to the nearest in order to estimate surface
area. The sample was stored in an opaque Nalgene
bottle with 500 ml of with stream and transported to
the laboratory in a dark cool-box. A well-mixed 10 ml
sample of periphyton preserved with 1% formalin was
drawn from each bottle for further processing.
Algae in each sample were first identified and
counted to genus-level using a 0.1 ml counting chamber
at 4009 magnification under a compound microscope
(Olympus� BX50F-3: Olympus Optical Co., Japan). At
least 300 units were tallied, where unit represented
individual cells for unicellular taxa, a 10 lm length for
filaments, and 10 9 10 lm2 area for colonies (Steven-
son & Bahls, 1999). All diatoms were grouped into one
category during this step. Subsequently, species-level
identification and enumeration of diatoms was con-
ducted, after they had been acid-cleaned (Hu & Wei,
2006) and slide-mounted, at 10009 magnification with
an oil immersion objective. At least 500 valves were
counted for each sample. Density of all taxa was
quantified in terms of numbers counted per unit area of
stone surface (cm2), with stone area calculated from
measures of perpendicular perimeters of each stones
using a regression model applicable to Hong Kong
streams (see Salas & Dudgeon, 2003). Relative abun-
dance was calculated also. Algal identification was
based on Patrick & Reimer (1966, 1975), Jao (1988), Qi
(1995), Li & Bi (1998), Shi (2004), and Zhu (2007),
while species names for diatoms were updated follow-
ing the Catalogue of Diatom Names of California
Academy of Sciences (On-line Version, updated 19
Sept. 2011, http://research.calacademy.org/research/
diatoms/names/index.asp).
Data analysis
Wet- and dry-season data were analyzed separately
because of substantial variations in water flow char-
acteristics between the two seasons. All streams were
treated as replicates in those analyses.
Differences in assemblage composition between
up- and down-stream reaches across all streams were
examined using non-metric multi-dimensional scaling
(NMDS, with Bray–Curtis similarities) and two-way
crossed analysis of similarities (ANOSIM). Where
Hydrobiologia (2013) 703:225–237 227
123
Author's personal copy

significant variation was detected, Similarity Percent-
ages (SIMPER), a procedure examining the contribu-
tion of species to average resemblances/dissimilarity
between sample groups based on Bray–Curtis simi-
larities (Clarke & Warwick, 2001), was used to
estimate the percentage contribution of each taxon to
the community dissimilarity. The analyses were
performed using PRIMER version 6 (PRIMER-E
Ltd, Plymouth, UK), with arcsine square root trans-
formations applied to relative abundance data to
reduce the influences of abundant taxa.
A variety of algal metrics were used to assess the
response to flow changes. Algal taxa were grouped into
three ecological guilds: high-profile (tall-statured
unicellular, erect, stalked, tube-forming, or filamen-
tous algae), low-profile (short-statured prostrate,
adnate, small erect, solitary centrics species) and
mobile guild (fast moving taxa) following Passy
(2007) and Passy & Larson (2011). Diatoms were
further classified into four growth groups, namely:
prostrate/adnate, erect, stalked, and filamentous
(Wang et al., 2005; Steuer et al., 2009; Berthon et al.,
2011) (see supplementary material—Appendix 3). In
addition, four diatom genera, each with distinctive
morphology, were enumerated and investigated: Ach-
nanthes sensu lato spp. (i.e., as defined before sepa-
rating some species into other genera, henceforth
Achnanthes spp.: usually small in size and prostrate
orientation to substrate), Cocconeis (prostrate taxon
with special adherence to the substrate by large
quantity of mucilage), Navicula sensu lato spp.
(henceforth Navicula spp.: generally prostrate, fre-
quently motile taxon), and Eunotia (adhere to the
substrate by one or more jelly pores) (Molloy, 1992;
Kutka & Richards, 1996).
The abundance for each guild, diatom growth form,
or genus was the sum of relative abundance of all taxa
within that group. Morphospecies richness (henceforth
species richness) and the exponent of Shannon–
Wiener diversity (exp. H0), which is more sensitive
to environmental change than H0 (Gray, 2000), were
calculated also.
Four dry-season samples (one from Tung Chung
and Tai Po Kau stream, two from SMK stream) were
spilled during transportation, resulting in an unbal-
anced repeated-measures dataset. Accordingly, linear
mixed models (LMM), adjusted by the restricted
maximum likelihood (REML), were used to investi-
gate the between-reach differences in flow upon algal
metrics. LMM applies to unequal repeated measures,
and can model various types of repeated covariance
structures (correlations in the repeated measures) that
are assumed to unstructured in conventional repeated
measures analyses of variance (Jiang, 2007; Goldstein,
2010). We used stream reaches as fixed factor, with
streams and sampling times as repeated measures, and
all algal metrics including abundance of ecological
guilds, diatom growth forms, the four diatom genera,
algal density, richness, and exp. H0, as well as pH, DO,
and conductivity, as dependent variables. We ran
LMM repeatedly by alternately selecting various
covariance structures (ante-dependence, first-order
autoregressive, compound symmetry, diagonal, and
so on) with the model yielding a value of Akaike’s
information criterion (AIC) closest to zero chosen as
the best-fit (SPSS Inc., 2008). Arcsine square root and
log(x ? 1) transformation was applied for percentage
and non-percentage data to improve the normality of
data sets. Nutrient concentrations, which were only
measured once per season, were analyzed by repeated-
measures ANOVA without replication. The analyses
were conducted using SPSS 17 (SPSS Inc., Chicago,
IL, USA).
Algal taxa contributing *40% of accumulated
community dissimilarity revealed by SIMPER, and
algal metrics that identified by LMM as exhibiting
significant between-reach differences in each season
were regarded as indicative of the effects of flow
reduction. We then calculated the between-reach dif-
ferences of those indicator taxa and metrics, and related
them to between-reach differences in flow (i.e., the
focus was on the magnitude of relative difference, not
the absolute values). Particular attention was paid to
differences in current velocity and discharge given that
they are both known to influence algal assemblages
(Biggs et al., 1998; Stanish et al., 2011; Villeneuve et al.,
2011). Differences in percentage data between reaches
were calculated by simple subtraction (upstream minus
downstream), while ratio data were calculated from the
percentage difference (difference between reaches
divided by upstream value).
To confirm whether responses of indicative algal
metrics and taxa (4Y) were caused by changes of flow
parameters, we use hierarchical partitioning (HP) to
identify significant explanatory factors (based on
upper 0.95 confidence limit, Z C 1.65) (Mac Nally,
2002). HP computes all possible hierarchical multiple
regression models that can be developed with a set of
228 Hydrobiologia (2013) 703:225–237
123
Author's personal copy

predictive variables and estimates independent effects
of each predictor by averaging contribution of a given
variable to the goodness of fit for all involved models.
Averaging is likely to alleviate multicolinearity prob-
lems so that the actual casual variables can be revealed
(Chevan & Sutherland, 1991; Mac Nally, 2000).
Besides percentage reductions of discharge (4Q)
and current velocity (4V), percentage changes in TN
(4TN), and PO4-P (4PO4-P) were also used as
predictive variables, irrespective of whether or not
there were statistically significant inter-reach differ-
ences in nutrient concentrations, because nutrients can
strongly influence algal assemblages (reviewed by
Larned, 2010). Quadratic terms for the four variables
were also included as predictors to detect possible
nonlinear relationships (Quinn & Keough, 2002). HP
was conducted in R (version 2.15) with the hier.part
package (version 1.03, Walsh & Mac Nally, 2008).
Whenever 4Q, 4V, or their quadratic terms was
detected as the unique significant causal variable for
the dependent variable 4Y, regression was then used
to uncover a possible relationship between % flow
reduction and the response of the algal metric.
Kolmogorov–Smirnov and Durbin–Watson tests were
used to check dataset normality, variance, and the
independence of residuals. Regressions and associated
tests were conducted with SigmaPlot 11.0 (Systat
Software Inc., Chicago, IL, USA).
Results
Effects of water abstraction on flow
and physicochemical variables
Water abstraction substantially altered stream flow in
both seasons. During the wet season, mean discharge
was reduced from 107 l s-1 (upstream, range:
21–245 l s-1) to 30 l s-1 (downstream, range: 1–220
l s-1) or a 71% reduction (range: 15–98%). Current
velocity decreased by 46% (39–83%), wetted width by
29% (3–53%) and depth by 23% (5–56%). Dry-season
discharge declined from 10 l s-1 (2–25 l s-1) to
4 l s-1 (0.1–13 l s-1), or by 54% (9–98%), with
current velocity reduced by 18% (ranged from a 79%
reduction to an increase of 50%) wetted width reduced
by 11%, and water depth slightly increased by 12%
(Table 1).
Concentrations of TN or PO4-P did not differ
significantly between reaches during either season,
while differences among streams varied inconsistently
according to season. Mean TN loading increased from
9.5 (upstream) to 11 lg l-1 (downstream), with
Table 1 Mean (±1 SE) and the percentage differences (%
change) of hydrological characteristics and stream water
between up- (U) and downstream (D) reaches of ten Hong
Kong streams during the wet and dry seasons
Wet season Dry season
Discharge (l s-1)
U 106.9 ± 13.5 9.6 ± 1.3
D 29.9 ± 10.9 3.7 ± 0.77
% Change -71.1 ± 6.9 -53.8 ± 6.3
Velocity (cm s-1)
U 20.2 ± 1.7 10.2 ± 0.7
D 9.4 ± 1.4 7.7 ± 0.8
% Change -46.4 ± 9.1 -17.8 ± 10.7
Wetted width (m)
U 5.5 ± 0.3 3.9 ± 0.3
D 3.7 ± 0.2 3.13 ± 0.2
% Change -29.2 ± 4.7 -11.1 ± 5.7
Depth (cm)
U 23.4 ± 1.1 14.3 ± 1.3
D 17.6 ± 1.4 15.2 ± 1.5
% Change -23.4 ± 5.6 11.8 ± 9.5
DO (mg l-1)
U 7.2 ± 0.1 9.0 ± 0.2
D 6.9 ± 0.1 8.7 ± 0.2
% Change -3.6 ± 1.6 -3.2 ± 1.8
Conductivity (ls cm-1)
U 22.8 ± 2.9 43.2 ± 3.1
D 24.5 ± 3.1 50.0 ± 3.8
% Change 10.6 ± 12.3 16.8 ± 7.0
pH
U 6.8 ± 0.1 7.0 ± 0.1
D 6.51 ± 0.2 6.8 ± 0.1
% Change -4.6 ± 1.9 -1.7 ± 1.0
Total nitrogen (lg l-1)
U 9.5 ± 3.3 65.2 ± 21.0
D 11.0 ± 3.4 41.3 ± 13.5
% Change 62.3 ± 55.7 -21.7 ± 31.8
Phosphate (lg l-1)
U 2.2 ± 0.5 3.4 ± 0.7
D 2.7 ± 0.4 3.1 ± 0.5
% Change 33.4 ± 17.7 2.8 ± 16.1
Hydrobiologia (2013) 703:225–237 229
123
Author's personal copy

absolute differences among streams ranging from
-20.3 to 19.9 lg l-1 during the wet season, and from
65.2 (upstream) to 41.3 lg l-1 (downstream, differ-
ences range: -96.4 to 69.5 lg l-1) during the dry
season. The equivalent values for PO4-P were 2.2 to
2.7 lg l-1 (wet, -1.3 to 1.7 lg l-1) and 3.4 to
3.1 lg l-1 (dry, -2.5 to 2.4 lg l-1). In addition to a
lack of significance in between-reach differences in
nutrients, DO, conductivity, and pH did not vary
between reaches in either season (Table 1).
Responses of algal variables to water abstraction
Out of a total of 196 taxa, comprising 160 Bacilla-
riophyta, 17 Cyanophyta, 17 Chlorophyta, and 2
Rhodophyta were collected during the study; 162
during the dry season and 142 in the wet season
(supplementary material—Appendix 2). The vast
majority were diatoms: 96% of total counts during
the wet season, 89% in the dry season. Achnanthidium
minutissimum (Kutzing) Czarnecki (with a relative
abundance of 22.4% in the wet season and 25.8% in
the dry season), Cocconeis placentula Ehrenberg (wet:
15.4%; dry: 16.5%), Karayevia oblongella (Østrup)
Aboal (wet: 13.3%; dry: 7.5%), Gomphonema parv-
ulum (Kutzing) Kutzing (wet: 5.7%; dry: 7.0%) and
Gomphonema clevei Fricke (wet: 5.4%; dry: 3.2%)
were the dominant taxa, collectively contributing to
[60% of total algal abundance during both seasons.
Despite substantial among-stream differences in
algal assemblage structure (ANOSIM: stream effects,
wet season: global R = 0.759, P = 0.001; dry season:
global R = 0.946, P = 0.001) (Fig. 1), there were
nonetheless strong between-reach differences in
assemblage composition (reach effects, wet season:
global R = 0.205, P = 0.031; dry season: global
R = 0.532, P = 0.001). SIMPER showed that, in
combination, 11 taxa contributed*40% to inter-reach
assemblage dissimilarity during each season (Table 2).
Among these indicator taxa, eight were important in
both seasons, including the five most abundant taxa
which collectively contributed[20% dissimilarity.
LMM revealed that none of the algal metrics
differed between reaches during the wet season. In
contrast, ten metrics showed significant between-
reach differences during the dry season (Table 3). At
downstream reaches, % low profile guild, % prostrate
diatoms, % Achnanthes spp., and % Cocconeis spp.
were reduced by 6–11% and % high profile guild,
% erect diatoms, % Eunotia spp., and % Navicula spp.
increased by 1.4–5%. Diversity increased from 6.7 to
8.9 (difference: 46.7%) and this was reflected in
species richness (23–28 species; difference: 23.2%).
Regression relationships between water flow
change and algal responses
4Q and 4V were identified as unique or combined
causal variables for the responses of 11 algal metrics
or taxa by HP, and these causal relations may have
been simple or nonlinear. In addition, 4TN and
quadratic terms of4PO4-P were significant predictors
for 7 and 2 algal responses, respectively (Table 4).
The independent contributions of all of these predic-
tors to the explained variance of algal responses
ranged from 23 to 44%.
No response of algae was caused solely by discharge
or current velocity reduction during the wet season.
However, % Achnanthes spp., % Cocconeis spp.,
% Eunotia spp., % Navicula spp., exp. H0, Cocconeis
placentula, and Gomphonema clevei were affected by
flow conditions during the dry season and were used to
analyze algal responses to flow reduction. % Cocconeis
placentula and % Cocconeis spp. decreased linearly
Fig. 1 Non-metric multi-dimensional scaling (NMDS) ordina-
tion of epibenthic algal assemblages in up- and down-stream
reaches of ten Hong Kong streams during the a wet and b dry
seasons
230 Hydrobiologia (2013) 703:225–237
123
Author's personal copy
with DQ (Fig. 2), while % Achnanthes spp., and
diversity showed quadratic responses to DQ. % Ach-
nanthes spp. initially declined as discharge was
reduced, but increased when DQ exceeded 89%
(Fig. 2). Diversity increased quadratically with greater
% reduction in DQ (DY [ 0 when DQ exceeded 15%).
Only % Navicula spp. apparently increased linearly
when current velocity declined (Fig. 3).
Discussion
Partitioning the physical and chemical effects
of flow reduction
Despite reductions in flow beneath water abstraction
points during both seasons, the effects of decreased
flow on algal assemblages were only apparent during
the dry season. Presumably this result reflects the fact
that water abstraction during the wet season was not
sufficient to cause a response from the majority of
algae, because the ‘‘background’’ stream discharge
during this season was relatively high due to the high
precipitation. Indeed, downstream discharge in the
five streams with the most depleted flows during the
wet season were approximately the same as their
upstream (natural) flows during the dry season. In
other words, flow reduction did not reach a critical
threshold that triggered responses of any algae.
Algal responses were mainly caused by between-
reach changes of the two flow parameters: discharge
and current velocity. However, 7 metrics and taxa
responded to PO4-P and TN changes, despite the fact
that there were only minor and non-significant
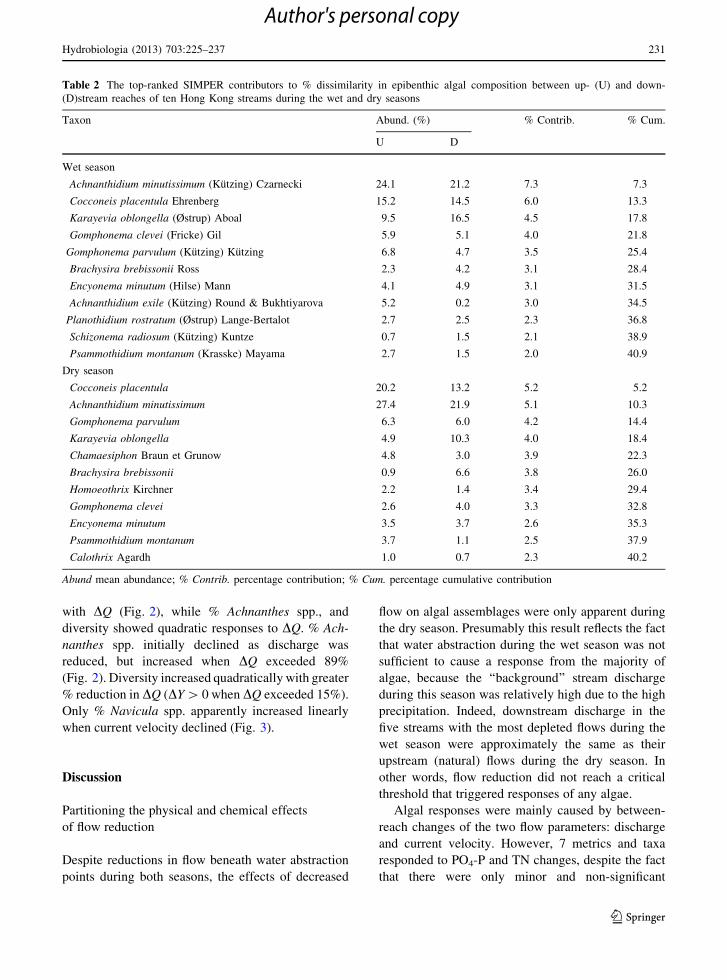
Table 2 The top-ranked SIMPER contributors to % dissimilarity in epibenthic algal composition between up- (U) and down-
(D)stream reaches of ten Hong Kong streams during the wet and dry seasons
Taxon Abund. (%) % Contrib. % Cum.
U D
Wet season
Achnanthidium minutissimum (Kutzing) Czarnecki 24.1 21.2 7.3 7.3
Cocconeis placentula Ehrenberg 15.2 14.5 6.0 13.3
Karayevia oblongella (Østrup) Aboal 9.5 16.5 4.5 17.8
Gomphonema clevei (Fricke) Gil 5.9 5.1 4.0 21.8
Gomphonema parvulum (Kutzing) Kutzing 6.8 4.7 3.5 25.4
Brachysira brebissonii Ross 2.3 4.2 3.1 28.4
Encyonema minutum (Hilse) Mann 4.1 4.9 3.1 31.5
Achnanthidium exile (Kutzing) Round & Bukhtiyarova 5.2 0.2 3.0 34.5
Planothidium rostratum (Østrup) Lange-Bertalot 2.7 2.5 2.3 36.8
Schizonema radiosum (Kutzing) Kuntze 0.7 1.5 2.1 38.9
Psammothidium montanum (Krasske) Mayama 2.7 1.5 2.0 40.9
Dry season
Cocconeis placentula 20.2 13.2 5.2 5.2
Achnanthidium minutissimum 27.4 21.9 5.1 10.3
Gomphonema parvulum 6.3 6.0 4.2 14.4
Karayevia oblongella 4.9 10.3 4.0 18.4
Chamaesiphon Braun et Grunow 4.8 3.0 3.9 22.3
Brachysira brebissonii 0.9 6.6 3.8 26.0
Homoeothrix Kirchner 2.2 1.4 3.4 29.4
Gomphonema clevei 2.6 4.0 3.3 32.8
Encyonema minutum 3.5 3.7 2.6 35.3
Psammothidium montanum 3.7 1.1 2.5 37.9
Calothrix Agardh 1.0 0.7 2.3 40.2
Abund mean abundance; % Contrib. percentage contribution; % Cum. percentage cumulative contribution
Hydrobiologia (2013) 703:225–237 231
123
Author's personal copy

between-reach differences in concentrations of both
nutrients. Stream flow has opposing subsidy-stress
effects on algae assemblages; i.e., flow continually
brings nutrients but simultaneously exerts shear stress
(Allan & Castillo, 2007). Their joint influence can vary
with circumstances, with nutrient supply dominating at
low current speeds and the physical effects of shear
stress becoming more important when flow is high
(Biggs et al., 1998). The interactions between flow and
nutrients are complex and compound specific. In this
Table 3 Comparison of
algal metrics between
upstream (U) and
downstream (D) reaches
during the dry season in
ten Hong Kong streams
based on LMM (see text)
Only metrics with
significant between-reach
differences (P \ 0.05)
are presented
Algal metrics Mean (range) LMM tests of reach effects
% High profile guild
U 17.8 (1.4 to 65.3)
D 22.7 (6.2 to 51.4)
Change 5.0 (-28.5 to 48.0) F1,15.142 = 10.276, P = 0.006
% Low profile guild
U 79.9 (34.2 to 98.4)
D 73.0 (46.4 to 91.0)
Change -6.9 (-49.1 to 27.9) F1,21.059 = 11.291, P = 0.003
% Prostrate/adnate diatoms
U 70.0 (26.9 to 98.4)
D 59.0 (16.2 to 92.1)
Change -11.1 (-50.1 to 28.5) F1,28.238 = 19.470, P \ 0.001
% Erect diatoms
U 0.8 (0 to 2.4)
D 3.3 (0 to 18.8)
Change 2.5 (-.07 to 18.8) F1,7.454 = 69.366, P \ 0.001
% Achnanthes
U 47.7 (23.7 to 81.4)
D 41.4 (13.8 to 75.0)
Change -6.2 (-31.6 to 17.9) F1,40.918 = 5.465, P = 0.024
% Cocconeis
U 20.7 (0 to 65.8)
D 14.6 (0 to 61.2)
Change -6.1 (-44.9 to 30.6) F1,43.004 = 5.032, P = 0.030
% Eunotia
U 0.3 (0 to 2.2)
D 2.0 (0 to 15.5)
Change 1.7 (-0.7 to 15.5) F1,6.213 = 11.003, P = 0.015
% Navicula
U 1.7 (0 to 10.1)
D 2.7 (0 to 12.3)
Change 1.4 (0.5 to 2.4) F1,7.558 = 76.915, P \ 0.001
Shannon–Wiener diversity (exp. H’)
U 6.7 (3.2 to 11.6)
D 8.9 (4.5 to 16.5)
% Change 46.7 (-52.7 to 192.5) F1,48.604 = 8.302, P = 0.006
Richness
U 23 (17 to 33)
D 28 (17 to 42)
% Change 23.2 (-29.2 to 82.6) F1,14.446 = 12.359, P = 0.003
232 Hydrobiologia (2013) 703:225–237
123
Author's personal copy

study, TN and PO4-P loads both increased during the
dry season when discharge was relatively low, and
there was a slight, but non-significant, increase in
PO4-P in downstream reaches. These changes are
highly likely to be due to a decrease in dilution capacity
and increase in residence times during under condi-
tions of reduced flow (Caruso, 2002; Lake, 2003).
Algal responses to flow reduction
Algal morphology strongly influenced their responses
to flow. Algae that are tolerant to scouring (Peterson,
1996; Biggs & Smith, 2002), such as members of
prostrate/adnate diatoms (especially Achnanthes spp.,
Cocconeis spp.), were more abundant in relative high
velocities or discharge upstream during the dry season.
Conversely, members of relative flow-intolerant
stalked species (Eunotia spp., Gomphonema clevei)
and mobile diatoms (Navicula spp., Brachysira breb-
issonii Ross) that able to crawl, avoid siltation and
select suitable habitats (Stevenson & Bahls, 1999)
were more abundant downstream. Other algal taxa and
metrics were more sensitive to nutrient changes than to
flow. Taxa in the high profile guild taxa (especially
erect diatoms, arborescent Chamaesiphon spp., Calo-
thrix spp.) were enhanced by small increases in PO4-P
or TN concentrations, while % low profile guild
diatoms in algal assemblages (including adnate Ach-
nanthidium exile (Kutzing) Round & Bukhtiyarova)
declined when PO4-P concentrations increased. This
result confirms the observation that low profile taxa
are more tolerant to resource-limited environments
(Passy, 2007; Berthon et al., 2011) where a complex
3D algal community cannot form due to low nutrient
availability (Passy, 2007; Passy & Larson, 2011).
Gomphonema parvulum and Psammothidium monta-
num (Krasske) Mayama were sensitive to both flow-
induced stresses, leading to combined responses.
Flow reduction appears to generate relatively stable
habitats that allow species with various attributes and
competitive abilities to coexist, resulting in increased
species richness (Biggs, 1996: Ghosh & Gaur, 1998)
as seen in downstream reaches of study streams. Flow-
related responses of diversity can be positive, negative
or shift from one to the other, depending on the studied
flow ranges (Wendker, 1992; Acs & Kiss, 1993;
Mackay et al., 2011) and it is notable that response of
diversity to discharge in this study was quadratic
(‘‘humped’’) rather than linear. Given the increased
species diversity downstream of water abstraction
points during the dry season, and higher periphyton
biomass reported by Niu & Dudgeon (2011a), it
appears that low-flow conditions have stimulated algal
assemblages in Hong Kong streams. In such cases
elsewhere, especially where nutrient supply is high.
Such conditions can give rise to excessive or nuisance
growths (Flinders & Hart, 2009).
Environmental flow allocation based on
eco–hydrological relationships
There have been few attempts to derive quantitative
eco–hydrological relationships between the extent of
Table 4 The significant predictors identified by hierarchical
partitioning and their independent contributions (I) to explain
between-reach difference of algal metrics and indicator taxa
during the wet and dry seasons (only present the metrics and
taxa whose causal variable(s) are detected)
Significant predictors I (%)
Algal metrics
Dry season
% High profile guild DPO4-P2 39.8
% Low profile guild DPO4-P2 44.0
% Erect diatoms DPO4-P2 38.1
% Achnanthes DQ2 43.5
% Cocconeis DQ 32.8
% Eunotia DV2 35.1
% Navicula DV 32.6
Exp. H0 DQ2 30.4
Richness DV 29.1
DQ 26.8
Indicator taxa
Wet season
Achnanthidium exile DPO4-P2 25.9
Dry season
Cocconeis placentula DQ 32.4
Gomphonema parvulum DQ 24.4
DPO4-P2 23.9
Chamaesiphon DTN 37.2
Brachysira brebissonii DV 35.6
DQ 29.4
Homoeothrix DPO4-P2 33.1
Gomphonema clevei DV2 37.4
Psammothidium montanum DPO4-P2 35.1
DV2 22.9
Calothrix DTN 26.5
Hydrobiologia (2013) 703:225–237 233
123
Author's personal copy

flow alternation and algal responses, and the manage-
ment implications of derived relationships can be
unclear partially because of the inconsistent responses
of ecological metrics (Larned, 2010; Poff & Zimmer-
man, 2010). The discharge reduction versus algal
response relationships reported herein, based on ten
streams with various degrees of flow reduction, could
be applied to determine the environmental flow
(e-flow) allocation needed to maintain epibenthic
algal assemblages in a close-to-natural condition in
Hong Kong, following a similar approach to that used
elsewhere (Arthington et al., 2006; Richter, 2010).
Using data from the same set of ten Hong Kong
streams, Niu and Dudgeon (2011a) proposed an e-flow
framework based on macroinvertebrates: a ‘‘thresh-
old’’ intended to maintain near-natural stream condi-
tions (4Y = 0), and a ‘‘degradation limit’’ that
allowed no more than 25% of the maximum indicator
Fig. 2 Linear or quadratic relationships between % discharge reduction (4Q) and between-reach differences in algal metrics during
the dry season. Only significant relationships (P \ 0.05) are presented
Fig. 3 Linear regressions between reductions in current veloc-
ity (4V, negative values representing increases) and between-
reach difference in algal metric during the dry season. Only
significant relationship (P \ 0.05) is presented
234 Hydrobiologia (2013) 703:225–237
123
Author's personal copy

response to flow reduction (4Y = 25%). We applied
this method to calculate acceptable % discharge
reduction based on the linear responses of relative
abundance of Cocconeis placentula and % Cocconeis
spp. during the dry season (estimating from the fitted
4Q versus 4Y regression equations in Fig. 2).
Calculated threshold e-flows required that C65%
natural discharge be maintained, and the degradation
limit was C32% natural discharge for the dry season.
A threshold or limit for the wet season could not be
calculated because discharge reduction was not the
causal factors to algal responses in the context of high
background levels of wet-season flow. Flow require-
ments derived from studies of benthic macroinverte-
brates in Hong Kong streams indicated a C74%
threshold discharge for both seasons, and degradation
limit discharges of C27% in the dry season (Niu &
Dudgeon, 2011a). Thus, a rough rule-of-thumb is that
at least three-quarters of the downstream flow should
be maintained to protect benthic macroinvertebrates
and algal assemblages in Hong Kong streams during
the dry season. Furthermore, retaining one-third of
natural discharge should be sufficient to limit the
serious impacts of water abstraction on macroinver-
tebrates and algal assemblages.
We note that the direction of the responses of most
algae and macroinvertebrates to discharge reduction
was inconsistent: macroinvertebrate diversity and
abundance decreased as discharge declined, while
algal metrics tended to exhibit the opposite trend.
However, e-flow management decisions need to be
based around sustaining natural assemblage structure,
and not maximizing diversity of a single group.
Among the algae, the linear response of Cocconeis
placentula to discharge reduction may have value as
indicator for monitoring success of an e-flow imple-
mentation program in monsoonal streams based
around the rules-of-thumb set out above, although
we recommend that any such monitoring include
macroinvertebrates as well as diatoms, and might also
be combined with process-based measurements that
reflect ecosystem functioning (Niu & Dudgeon,
2011b). The information thereby generated could be
used to inform adaptive management and thereby
refine e-flow allocations for regulated monsoonal
streams such as those in Hong Kong.
Acknowledgments We thank Yixin Zhang for facilitating this
collaborative research, Lily C. Y. Ng for technical support in the
laboratory, and the Field Work Support Team of the Division of
Ecology and Biodiversity at the University of Hong Kong for
logistical assistance.
References
Acreman, M. & M. J. Dunbar, 2004. Defining environmental
river flow requirements—a review. Hydrology and Earth
System Sciences 8: 861–876.
Acs, E. & K. T. Kiss, 1993. Effects of the water discharge on
periphyton abundance and diversity in a large river (River
Danube, Hungary). Hydrobiologia 249: 125–133.
Allan, J. & M. Castillo, 2007. Stream Ecology: Structure and
Function of Running Waters. Springer, Dordrecht, the
Netherlands.
APHA, 1995. Standard Methods for the Examination of Water
and Wastewater, 19th ed. American Public Health Asso-
ciation, New York.
Arthington, A., S. Bunn, N. Poff & R. Naiman, 2006. The
challenge of providing environmental flow rules to sustain
river ecosystems. Ecological Applications 16: 1311–1318.
Arthington, A. H., R. J. Naiman, M. E. McClain & C. Nilsson,
2010. Preserving the biodiversity and ecological services
of rivers: new challenges and research opportunities.
Freshwater Biology 55: 1–16.
Berthon, V., A. Bouchez & F. Rimet, 2011. Using diatom life-
forms and ecological guilds to assess organic pollution and
trophic level in rivers: a case study of rivers in south-
eastern France. Hydrobiologia 673: 259–271.
Biggs, B. J. F. & G. M. Price, 1987. A survey of filamentous
algal proliferations in New Zealand Rivers. New Zealand
Journal of Marine and Freshwater Research 21: 175–191.
Biggs, B. J. F. & R. A. Smith, 2002. Taxonomic richness of
stream benthic algae: effects of flood disturbance and
nutrients. Limnology and Oceanography 47: 1175–1186.
Biggs, B. J. F., 1996. Hydraulic habitat of plants in streams.
Regulated Rivers: Research & Management 12: 131–144.
Biggs, B. J. F., D. G. Goring & V. I. Nikora, 1998. Subsidy and
stress responses of stream periphyton to gradients in water
velocity as a function of community growth form. Journal
of Phycology 34: 598–607.
Biggs, B. J. F., R. Stevenson & R. Lowe, 1998. A habitat matrix
conceptual model for stream periphyton. Archiv fur Hy-
drobiologie 143: 21–56.
Brisbane Declaration, 2007. The Brisbane Declaration. Envi-
ronmental flows are essential for freshwater ecosystem
health and human well-being. Declaration of the 10th
International River symposium and International Envi-
ronmental Flows Conference, Brisbane, Australia, 3–6
Sept 2007.
Caruso, B., 2002. Temporal and spatial patterns of extreme low
flows and effects on stream ecosystems in Otago,
New Zealand. Journal of Hydrology 257: 115–133.
Chester, H. & R. Norris, 2006. Dams and flow in the Cotter
River, Australia: effects on instream trophic structure and
benthic metabolism. Hydrobiologia 572: 275–286.
Chevan, A. & M. Sutherland, 1991. Hierarchical partitioning.
The American Statistician 45: 90–96.
Hydrobiologia (2013) 703:225–237 235
123
Author's personal copy

Clarke, K. R. & R. M. Warwick, 2001. Change in Marine
Communities: An Approach to Statistical Analysis and
Interpretation, 2nd ed. PRIMER-E, Plymouth.
Clausen, B. & B. Biggs, 1997. Relationships between benthic
biota and hydrological indices in New Zealand streams.
Freshwater Biology 38: 327–342.
Downes, B. J., T. J. Entwisle & P. Reich, 2003. Effects of flow
regulation on disturbance frequencies and in-channel bry-
ophytes and macroalgae in some upland streams. River
Research and Applications 19: 27–42.
Dudgeon, D., 1992. Effects of water transfer on aquatic insects
in a stream in Hong Kong. Regulated Rivers: Research &
Management 7: 369–377.
Dudgeon, D., 1996. Anthropogenic influences on Hong Kong
streams. GeoJournal 40: 53–61.
Flinders, C. A. & D. D. Hart, 2009. Effects of pulsed flows on
nuisance periphyton growths in rivers: a mesocosm study.
River Research and Applications 25: 1320–1330.
Ghosh, M. & J. P. Gaur, 1998. Current velocity and the estab-
lishment of stream algal periphyton communities. Aquatic
Botany 60: 1–10.
Goldstein, H., 2010. Multilevel Statistical Models, 4th ed.
Wiley, Chichester, UK.
Gore, J., 2006. Discharge measurements and streamflow anal-
ysis. In Hauer, F. R. & G. A. Lamberti (eds), Methods in
Stream Ecology. Academic Press, San Diego: 51–77.
Gray, J. S., 2000. The measurement of marine species diversity,
with an application to the benthic fauna of the Norwegian
continental shelf. Journal of Experimental Marine Biology
and Ecology 250: 23–49.
Hart, D. & C. Finelli, 1999. Physical-biological coupling in
streams: the pervasive effects of flow on benthic organ-
isms. Annual Review of Ecology and Systematics:
363–395.
Hu, H. & Y. Wei, 2006. The Freshwater Algae of China: Sys-
tematics, Taxonomy and Ecology. Science Press, Beijing
(in Chinese).
Jao, C., 1988. Flora Algarum Sinigrum Aquae Dulcis (Tomus I):
Zygnemataceae. Science Press, Beijing (in Chinese).
Jiang, J., 2007. Linear and Generalized Linear Mixed Models
and Their Applications. Springer Science?Business
Media, New York, USA.
Jowett, I. G. & M. J. Duncan, 1990. Flow variability in New
Zealand Rivers and its relationship to in-stream habitat and
biota. New Zealand Journal of Marine and Freshwater
Research 24: 305–317.
Kutka, F. J. & C. Richards, 1996. Relating diatom assemblage
structure to stream habitat quality. Journal of the North
American Benthological Society 15: 469–480.
Lake, P. S., 2003. Ecological effects of perturbation by drought
in flowing waters. Freshwater Biology 48: 1161–1172.
Larned, S. T., 2010. A prospectus for periphyton: recent and
future ecological research. Journal of the North American
Benthological Society 29: 182–206.
Ledger, M., R. Harris, P. Armitage & A. Milner, 2008. Distur-
bance frequency influences patch dynamics in stream
benthic algal communities. Oecologia 155: 809–819.
Li, S. & L. Bi, 1998. Flora Algarum Sinicarum Aquae Dulcis
(Tomus V): Ulothricales Ulvales Chaetophorales Tren-
tepohliales Sphaeropleales. Science Press, Beijing
(in Chinese).
Luce, J., A. Cattaneo & M. Lapointe, 2010. Spatial patterns in
periphyton biomass after low-magnitude flow spates:
geomorphic factors affecting patchiness across gravel-
cobble riffles. Journal of the North American Benthologi-
cal Society 29: 614–626.
Mac Nally, R., 2000. Regression and model-building in con-
servation biology, biogeography and ecology: the distinc-
tion between—and reconciliation of—‘predictive’ and
‘explanatory’ models. Biodiversity and Conservation 9:
655–671.
Mac Nally, R., 2002. Multiple regression and inference in
ecology and conservation biology: further comments on
identifying important predictor variables. Biodiversity and
Conservation 11: 1397–1401.
Mackay, A. W., T. Davidson, P. Wolski, S. Woodward, R.
Mazebedi, W. R. L. Masamba & M. Todd, 2011. Diatom
sensitivity to hydrological and nutrient variability in a
subtropical, flood-pulse wetland. Ecohydrology 5:
491–502. doi:10.1002/eco.242.
March, J. G., J. P. Benstead, C. M. Pringle & F. N. Scatena,
2003. Damming tropical island streams: problems, solu-
tions, and alternatives. BioScience 53: 1069–1078.
Martınez de Fabricius, A. L., N. Maidana, N. Gomez &
S. Sabater, 2003. Distribution patterns of benthic diatoms
in a Pampean river exposed to seasonal floods: the Cuarto
River (Argentina). Biodiversity and Conservation 12:
2443–2454.
Molloy, J. M., 1992. Diatom communities along stream longi-
tudinal gradients. Freshwater Biology 28: 59–69.
Nilsson, C., C. A. Reidy, M. Dynesius & C. Revenga, 2005.
Fragmentation and flow regulation of the world’s large
river systems. Science 308: 405–408.
Niu, Q. & D. Dudgeon, 2011a. Environmental flow allocations
in monsoonal Hong Kong. Freshwater Biology 56:
1209–1230.
Niu, Q. & D. Dudgeon, 2011b. The influence of flow and season
upon leaf-litter breakdown in monsoonal Hong Kong
streams. Hydrobiologia 663: 205–215.
Opsahl, R. W., T. Wellnitz & N. L. Poff, 2003. Current velocity
and invertebrate grazing regulate stream algae: results of an
in situ electrical exclusion. Hydrobiologia 499: 135–145.
Passy, S. I., 2007. Diatom ecological guilds display distinct and
predictable behavior along nutrient and disturbance gra-
dients in running waters. Aquatic Botany 86: 171–178.
Passy, S. I. & C. Larson, 2011. Succession in stream biofilms is
an environmentally driven gradient of stress tolerance.
Microbial Ecology 62: 414–424.
Patrick, R. & C. W. Reimer, 1966. The Diatoms of the United
States, Exclusive of Alaska and Hawaii, Vol. 1. Monograph
of the Academy of Natural Sciences of Philadelphia,
Number 13: 688 pp.
Patrick, R. & C. W. Reimer, 1975. The Diatoms of the United
States, Exclusive of Alaska and Hawaii. Vol. 2, Part 1.
Monograph of the Academy of Natural Sciences of Phila-
delphia, Number 13: 213 pp.
Peterson, C. G., 1996. Response of benthic algal communities to
natural physical disturbance. In Stevenson, R. J., M.
L. Bothwell & R. L. Lowe (eds), Algal Ecology: Fresh-
water Benthic Ecosystems. Academic Press, San Diego:
375–402.
236 Hydrobiologia (2013) 703:225–237
123
Author's personal copy

Peterson, C. G. & R. J. Stevenson, 1992. Resistance and resil-
ience of lotic algal communities: importance of disturbance
timing and current. Ecology 73: 1445–1461.
Poff, N. L. & J. V. Ward, 1995. Herbivory under different flow
regimes: a field experiment and test of a model with a
benthic stream insect. Oikos 72: 179–188.
Poff, N. L. & J. K. H. Zimmerman, 2010. Ecological responses
to altered flow regimes: a literature review to inform the
science and management of environmental flows. Fresh-
water Biology 55: 194–205.
Qi, Y., 1995. Flora Algarum Sinicarum Aquae Dulcis (Tomus
IV): Bacillariophyta Centreae. Science Press, Beijing (in
Chinese).
Quinn, G. P. & M. J. Keough, 2002. Experimental Design and
Data Analysis for Biologists. Cambridge University Press,
Cambridge.
Richter, B. D., 2010. Re-thinking environmental flows: from
allocations and reserves to sustainability boundaries. River
Research and Applications 26: 1052–1063.
Salas, M. & D. Dudgeon, 2003. Life histories, production
dynamics and resource utilisation of mayflies (Epheme-
roptera) in two tropical Asian forest streams. Freshwater
Biology 48: 485–499.
Shi, Z., 2004. Flora Algarum Sinicarum Aquae Dulcis (Tomus
XII): Bacillariophyta Gomphonemacea. Science Press,
Beijing (in Chinese).
Soininen, J., 2003. Heterogeneity of benthic diatom communi-
ties in different spatial scales and current velocities in a
turbid river. Archiv fur Hydrobiologie 156: 551–564.
SPSS Inc., 2008. SPSS Advanced Statistics 17.0. SPSS Inc.,
Chicago, IL.
Stanish, L. F., D. R. Nemergut & D. M. McKnight, 2011.
Hydrologic processes influence diatom community com-
position in Dry Valley streams. Journal of the North
American Benthological Society 30: 1057–1073.
Steuer, J. J., J. D. Bales & E. Giddings, 2009. Relationship of
stream ecological conditions to simulated hydraulic met-
rics across a gradient of basin urbanization. Journal of the
North American Benthological Society 28: 955–976.
Stevenson, R. J., 1996. The stimulation and drag of current. In
Stevenson, R. J., M. L. Bothwell, & R. L. Lowe (eds), Algal
Ecology: Freshwater Benthic Ecosystems. Academic
Press, San Diego: 321–340.
Stevenson, R. J. & L. L. Bahls, 1999. Periphyton protocols. In
Barbour, M. T., J. Gerritsen, B. D. Snyder & J. B. Stribling
(eds), Rapid Bioassessment Protocols for Use in Streams
and Wadeable Rivers: Periphyton, Benthic Macroinverte-
brates, and Fish, 2nd edn. EPA 841-B-99–002. Office of
Water. US Environmental Protection Agency, Washing-
ton, DC.
Stewart, K. A., S. F. Lamoureux & A. C. Forbes, 2005. Hydro-
logical controls on the diatom assemblage of a seasonal
arctic river: boothia Peninsula, Nunavut, Canada. Hydro-
biologia 544: 259–270.
Suren, A. M., B. J. F. Biggs, C. Kilroy & L. Bergey, 2003.
Benthic community dynamics during summer low-flows in
two rivers of contrasting enrichment 1. Periphyton. New
Zealand Journal of Marine and Freshwater Research 37:
53–70.
Uehlinger, U., B. Kawecka & C. T. Robinson, 2003. Effects of
experimental floods on periphyton and stream metabolism
below a high dam in the Swiss Alps (River Spol). Aquatic
Sciences 65: 199–209.
Villeneuve, A., A. Bouchez & B. Montuelle, 2011. In situ
interactions between the effects of season, current velocity
and pollution on a river biofilm. Freshwater Biology 56:
2245–2259.
Walsh C, Mac Nally R., 2008. hier.part: hierarchical partition-
ing. R package. [Available at http://cran.r-project.org/web/
packages/hier.part/index.html].
Wang, Y., R. Stevenson & L. Metzmeier, 2005. Development
and evaluation of a diatom-based Index of Biotic Integrity
for the Interior Plateau Ecoregion, USA. Journal of the
North American Benthological Society 24: 990–1008.
Wang, Q., C. Zhi, P. B. Hamilton & F. Kang, 2009. Diatom
distributions and species optima for phosphorus and cur-
rent velocity in rivers from ZhuJiang Watershed within a
Karst region of south-central China. Fundamental and
Applied Limnology/Archiv fur Hydrobiologie 175:
125–141.
Wendker, S., 1992. Influence of current velocity on diatoms of a
small soft water stream. Diatom Research 7: 387–396.
Wu, N., T. Tang, S. Zhou, X. Jia, D. Li, R. Liu & Q. Cai, 2009.
Changes in benthic algal communities following con-
struction of a run-of-river dam. Journal of the North
American Benthological Society 28: 69–79.
Wu, N., T. Tang, X. Fu, W. Jiang, F. Li, S. Zhou, Q. Cai & N.
Fohrer, 2010. Impacts of cascade run-of-river dams on
benthic diatoms in the Xiangxi River, China. Aquatic
Sciences 72: 117–125.
Yang, G. Y. & D. Dudgeon, 2010a. Dietary variation and food
selection by an algivorous loach (Pseudogastromyzonmyersi: Balitoridae) in Hong Kong streams. Marine &
Freshwater Research 61: 49–56.
Yang, G. Y. & D. Dudgeon, 2010b. Response of grazing impacts
of an algivorous fish (Pseudogastromyzon myersi: Balito-
ridae) to seasonal disturbance in Hong Kong streams.
Freshwater Biology 55: 411–423.
Yang, G. Y., T. Tang & D. Dudgeon, 2009. Spatial and seasonal
variations in benthic algal assemblages in streams in
monsoonal Hong Kong. Hydrobiologia 632: 189–200.
Zhu, H., 2007. Flora Algarum Sinicarum Aquae Dulcis (Tomus
IX): Cyanophyta Hormogonophyceae. Science Press,
Beijing (in Chinese).
Hydrobiologia (2013) 703:225–237 237
123
Author's personal copy