
159_ GALLARDO et al.
14
Original article Nutrient cycling in deciduous forest ecosystems of the Sierra de Gata mountains: nutrient supplies to the soil through both litter and throughfall Juan F. Gallardo Alejandro Martín b Gerardo Moreno’ Ignacio Santa Regina a C.S.I.C., Aptdo. 257, Salamanca 37071, Spain b Area de Edafología, Facultad de Farmacia, Salamanca 37080, Spain (Received 2 September 1997; accepted 17 October 1997) Abstract - The present work fits into a general study on nutrient cycling in four Quercus pyrenaica oak forests and one Castanea sativa chestnut coppice located in the Sierra de Gata mountains (Cen- tral System, western Spain). The work consists of an estimation of bioelement supplies to the soil by the litter of these species and by throughfall from the canopy with a view to defining their role in the soil and, more generally, in ecosystem bioelement dynamics. It is concluded that the greatest differences between the oak stands and the chestnut coppice lie in the fact that in the latter ecosystem potentially more N, P, K, Mg, Na and Mn return through the litter owing to greater production in the chestnut cop- pice (and/or root uptake). Additionally, the relative importance of some bioelements (N, P, K and Mn) in the chestnut coppice is different from that of the oak forests. It is also possible to differentiate three groups of bioelements: 1) those that potentially return almost exclusively through the litter (C and N); 2) those for which both litter and throughfall must be taken into account to determine the potential return of bioelements (Ca, Mg, P, K, Fe and Mn); and 3) those that return almost exclusively through canopy leaching (Na, Cu and Zn). Despite this, on attempting to calculate the actual minimum annual returns, the three groups must be reduced to two: bioelements that almost exclusively return by throughfall (Na, Cu and Zn), and bioclements that return through litter decay and canopy leach- ing. Exceptionally, Fe behaves in a special way in the sense that it tends to be immobilized by decay- ing leaf litter. (© Inra/Elsevier, Paris) nutrient cycling / throughfall / bioelement return / forest litter / broadleaf forest ecosystems Résumé - Cycle des bioéléments dans des écosystèmes forestiers de la Sierra de Gata : apport d’éléments nutritifs au sol par le pluviolessivage et la décomposition de la litière. Le recyclage de bioéléments dans quatre chênaies à Quercus pyrenaica et dans une châtaigneraie à Castanea sativa, localisées dans la Sierra de Gata (Système Central, ouest de l’Espagne) a fait l’objet de cette étude. Il s’agit d’une estimation des éléments biogènes qui retournent au sol par décomposition de la * Correspondence and reprints E-mail: [email protected]
Transcript of 159_ GALLARDO et al.

Original article
Nutrient cycling in deciduous forest ecosystemsof the Sierra de Gata mountains: nutrient supplies
to the soil through both litter and throughfall
Juan F. Gallardo Alejandro Martínb Gerardo Moreno’
Ignacio Santa Regina
aC.S.I.C., Aptdo. 257, Salamanca 37071, SpainbArea de Edafología, Facultad de Farmacia, Salamanca 37080, Spain
(Received 2 September 1997; accepted 17 October 1997)
Abstract - The present work fits into a general study on nutrient cycling in four Quercus pyrenaicaoak forests and one Castanea sativa chestnut coppice located in the Sierra de Gata mountains (Cen-tral System, western Spain). The work consists of an estimation of bioelement supplies to the soil bythe litter of these species and by throughfall from the canopy with a view to defining their role in thesoil and, more generally, in ecosystem bioelement dynamics. It is concluded that the greatest differencesbetween the oak stands and the chestnut coppice lie in the fact that in the latter ecosystem potentiallymore N, P, K, Mg, Na and Mn return through the litter owing to greater production in the chestnut cop-pice (and/or root uptake). Additionally, the relative importance of some bioelements (N, P, K and Mn)in the chestnut coppice is different from that of the oak forests. It is also possible to differentiatethree groups of bioelements: 1) those that potentially return almost exclusively through the litter(C and N); 2) those for which both litter and throughfall must be taken into account to determine thepotential return of bioelements (Ca, Mg, P, K, Fe and Mn); and 3) those that return almost exclusivelythrough canopy leaching (Na, Cu and Zn). Despite this, on attempting to calculate the actual minimumannual returns, the three groups must be reduced to two: bioelements that almost exclusively returnby throughfall (Na, Cu and Zn), and bioclements that return through litter decay and canopy leach-ing. Exceptionally, Fe behaves in a special way in the sense that it tends to be immobilized by decay-ing leaf litter. (© Inra/Elsevier, Paris)
nutrient cycling / throughfall / bioelement return / forest litter / broadleaf forest ecosystems
Résumé - Cycle des bioéléments dans des écosystèmes forestiers de la Sierra de Gata : apportd’éléments nutritifs au sol par le pluviolessivage et la décomposition de la litière. Le recyclagede bioéléments dans quatre chênaies à Quercus pyrenaica et dans une châtaigneraie à Castaneasativa, localisées dans la Sierra de Gata (Système Central, ouest de l’Espagne) a fait l’objet de cetteétude. Il s’agit d’une estimation des éléments biogènes qui retournent au sol par décomposition de la
* Correspondence and reprintsE-mail: [email protected]

litière et par le pluviolessivage des arbres. L’objet de l’étude est de définir le rôle et la dynamique desbioéléments dans le sol et l’écosystème.On peut conclure que la plus grande différence existante entre les peuplements de Q. pyrenaica et deC. sativa est que ce dernier écosystème peut potentiellement restituer davantage de N, P, K, Mg,Na et Mn par la litière, à cause d’une plus forte production de biomasse aérienne chez le châtaignier(et/ou plus forte absorption par les racines). On observe, en outre, que la relative importance dequelques bioéléments (N, P, K et Mn) est différente dans la châtaigneraie et les autres chênaies.Il est ainsi possible de différencier trois groupes d’élément biogènes : tout d’abord, ceux qui peuventpotentiellement retourner majoritairement par la litière (C et N); en deuxième lieu, ceux qui retour-nent soit par la décomposition de la litière soit par le pluviolessivage (Ca, Mg, P, K, Fe et Mn); et fina-lement, ceux qui retournent presque exclusivement par pluviolessivage (Na, Cu et Zn).En revanche, en ce qui concerne l’apport réel annuel de bioéléments, deux groupes peuvent se dif-férencier : d’une part, celui des bioéléments qui retournent par pluviolessivage (Na, Cu et Zn); et d’autrepart, les bioéléments qui retournent soit par décomposition de la litière, soit par pluviolessivage.Le Fe, au contraire, a un comportement spécial car il est immobilisé dans la litière en décomposition.(© Inra/Elsevier, Paris)
cycle des bioéléments / pluviolessivage / retour d’éléments nutritifs / litière / forêt caducifoliée/ écosystème
1. INTRODUCTION
Plant litter returns the nutrients and
energy stocked in the vegetation to soils,with the important participation of microor-ganisms; nutrients circulate in the ecosys-tem and play a special and important role,essential for the life of all components [17,19, 25, 45]. Litter quality, litter decomposi-tion and quantitative inputs to the soil affectpedogenesis and the productivity of ecosys-tems. Knowledge of these different aspectsis a determining factor for understanding the functions of nutrient flows in ecosys-tems.
Bioelement inputs from throughfall tothe forest floor, and then to the soil, are theresult of a complex interaction of atmo-spheric, hydrological and biogeochemicalprocesses [34]. The final composition of thewater flowing from the canopy is determinedby the initial composition of the rainfallwater, the wash-off of dry atmospheric dust,and water interception, leaching and/oruptake of ions by the forest canopy [39].
The quantities of bioelements brought tothe soil through these processes and also the
quality of solubilized substances are ofmajor interest for ecosystem function andproductivity [5, 25]. Knowledge of thesenutrient contributions (mostly in availableform for plants) is of great importance forplant nutrition [17, 37].
Studies on the inputs of biogenous ele-ments in broadleaf forest populations havebeen carried out by Lossaint [24], Rapp [40],Aussenac et al. [1], Lemee [21], SantaRegina et al. [43, 44], Hernández et al. [18],Moreno et al. [34] and Martin et al. [27, 28],among others.
The present work fits into a general studyon nutrient cycling in four Quercus pyre-naica oak forests and one Castanea sativa
coppice located in the Sierra de Gata moun-tains (Central System, western Spain). Thework aims at estimating total bioelementsupplies to the soil by the litter of thesespecies and by throughfall with a view todefining their role in the soil and, more gen-erally, in ecosystem bioelement dynamics. Afurther aim is to attempt to estimate the trueminimum annual nutrient input to the soil.

2. MATERIALS AND METHODS
2.1. Characteristics of the study site
The study area is located in the El Rebollardistrict (Sierra de Gata mountains, westernSpain); its coordinates are 40° 19’ N and6° 43’ W [14, 16]. The wooded area is mainlyformed of Quercus pyrenaica Wilid. (deciduousoak), Pinus pinaster Ait. (maritime pine) and, atthe southern border of the El Rebollar district,Castanea sativa Miller (chestnut).
The four selected Q. pyrenaica oak plots sit-uated at Navasfrías (NF), El Payo (EP), Vi-llasrubias (VR) and Fuenteguinaldo (FG) accord-ing to a decreasing rainfall transect, display thefollowing characteristics: a tree density rangingfrom 1 040 trees ha-1 at VR to 406 trees ha-1 atEP (table I). The plot with the lowest density(EP) has the highest mean trunk diameter(25.4 cm) and greatest tree height (17 m); thelowest values of these parameters are in VR plot(11 cm and 8.5 m, respectively; table 1). The leafarea index (L.A.I.) ranges from 1.8 to 2.6 m2 m-2on the NF and FG plots, respectively. Basal arearanges from 0.135 and 0.212 m2 m-2 on the VRand FG plots, respectively (table I).
The selected coppice of Castanea sativachestnut is situated in San Martin de Trevejo
(SM) and has a density of 3 970 trees ha-1, witha mean diameter of 10 cm and a height of 13 m.The mean basal area of 0.306 m2 m-2 and theL.A.I. is 3.7 m2 m-2 (table I).
The climate of the area is characterized byrainy winters and hot dry summers, falling underthe classification of humid Mediterranean, withan average rainfall and temperature of approxi-mately 1 580 mm year-1 and 10.4 °C for NF and720 mm yeat-1 and 12.9 °C for FG (table I).
The soils of these areas are generally humicCambisols [11] developed on slates and
graywackes at NF and VR and on Ca-alcalinegranite at EP and FG [13]. At SM, owing to thestrong slope (approximately 45 %), granitic sandspredominate, sometimes with man-made terraces.
2.2. Chemical compositionsof litterfall and throughfall
The litter fallen over the year was sampled atvarying intervals depending on its rate of fall(between 2 weeks and 1 month [18]. After col-lection, the litter was separated into differentfractions (leaves, branches, flowers, fruits, barks,etc.) and then dried, air cleaned and weighed[29].

Study of the decomposition of oak and chest-nut leaves was followed using the classic lit-terbag method [27, 30]. Field material (leaves,branches, twigs, water) was suitably treated priorto determining the following bioelement con-centrations: litter organic C by a Carmhograph 12Wösthoff; litter N by a Heraeus Macro-N-ana-lyzer; P by spectrophotometry (Varian DMS 90)using either the vanadomolybdophosphoric yel-low method for determining litter P or the ascor-bic acid method for determining water P; waterpH was determined with a Beckman 3500 pHmeter; water-dissolved total and organic C by aBeckman 315A T.O.C.A. Water-dissolved anionswere determined by ionic chromatography(Dionex 350). The determination of dissolvedcations and these bioelements in litter was carriedout by atomic absorption spectroscopy (Varian1475) and water-dissolved micronutrients byplasma spectrometry (Perkin-Elmer ICP-2).
We use the term ’potential return of onebioelement’ to refer to the total content of thiselement in the litterfall [17]; that is, the totalquantity of one bioelemenl which is releasedfrom the decomposing litter when it has com-pletely decomposed (including the more recal-citrant fractions of the litter); then, the potentialreturn of each bioelement is estimated by multi-plying the litterfall by its composition (weightingthe different fractions [ 17]).
As is known, significant fractions of bioele-ments are usually retained in the organic remains.The potential return of nutrients generally has ahigher value than the actual return. We use theterm ’actual minimum input of one nutrient tothe soil’ to refer to the calculated minimum realcontribution of the decomposing litter, accordingto the pattern of release of each element as deter-mined by the litterbag method [30, 45].
It should be mentioned that the contribution
by the roots to potential bioelement return wasnot taken into account in the present work.Khanna and Ulrich [19] estimated that the rootbioelement content represents about 20 % of thetotal potential return.
The contributions of bioelements reachingthe forest floor through canopy leaching maycome from three different sources: rainfall, drydeposition and throughfall, each of them havinga different degree of quantitative importance forall the elements considered. In this article
throughfall and canopy leaching are used as syn-onymous, even though they are not exactly thesame [35]. Furthermore, according to Moreno etal. [36] rainfall water represents the main source
of bioelements reaching the forest floor throughcanopy leaching.
3. RESULTS
3.1. Total nutrient inputto the soil by litter
Aspects related to aboveground produc-tion of these forests will be discussed else-where [16], although some figures areshown in table I.
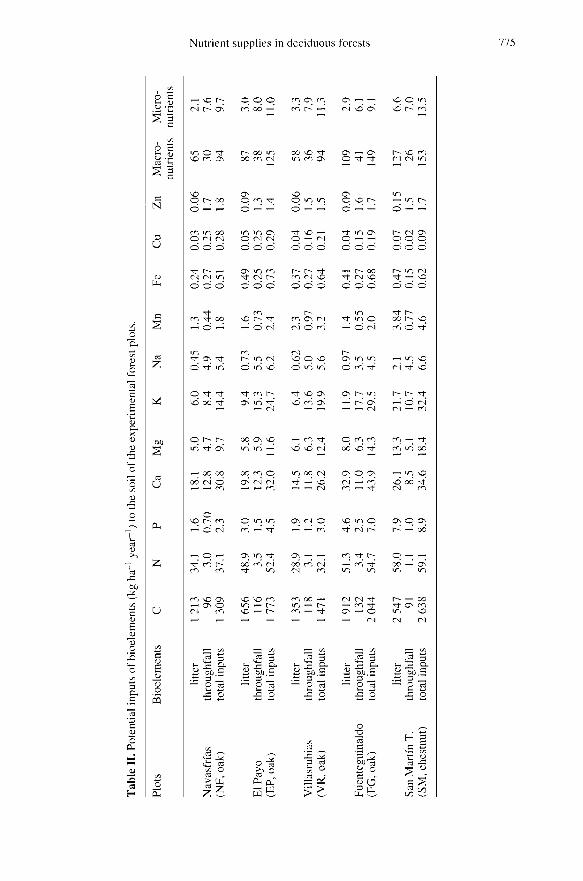
The data concerning the annual poten-tial return of bioelements through litterfall tothe forest soil of the five forest systems areshown in table II.
The oak forest at FG has the highestpotential bioelement return and litterfall pro-duction (4.1 Mg ha-1 year-1, equivalent to1.9 Mg ha-1 year-1 of C). VR and NF arethe plots with the lowest potential bioele-ment return and also the lowest litterfall (2.8and 2.6 Mg ha-1 year-1, respectively, equiv-alent to 1.3 and 1.2 Mg ha-1 year-1 of C,respectively). Additionally, significant dif-ferences were found (table II) in the poten-tial return of bioelements between forests
developed on slates and those on granites.Because the chestnut coppice is the most
productive forest (table I), the highest levelsof potential return (table II) of macronutri-ents (sum of N, P, Ca, K and Mg) andmicronutrients (Na, Fe, Mn, Cu and Zn)were obtained there (127 and 6.6 kg ha-1year-1, respectively); 2.6 Mg ha-1 year-1 of
organic C were also returned at the chest-nut coppice (table II). By contrast, 108, 87,65 and 57 kg ha-1 year-1 of macronutrientsand 2.9, 3.0, 2.1 and 3.3 kg ha-1 year-1 of
micronutrients were returned annuallythrough the litterfall at FG, EP, NF and VRoak forests, respectively (table II).
3.2. Inputs of bioelements to soilsby throughfall (canopy leaching)
The results are shown in table II.

In canopy leaching, C was the elementwith the greatest contribution to soil, farhigher than those observed for the other ele-ments.
The major cations were Ca and K, whileN had lower values (above 3 kg ha-1 year-1,lower at SM). Ca values were close to12 kg ha-1 year-1, lower at SM. K and Mghad values close to 15 and 5 kg ha-1 year-1,respectively, lower at NF and higher at SM.
Phosphorus had a very low concentra-tion, at values close to those of micronutri-ents.
In general, in terms of mass, the order ofimportance of the different elements pre-sent in the thoughfall would be:
No large differences can be seen betweenthe different forest ecosystems, except for Kand P in FG (higher soil pH), and Ni andCu in SM (chestnut coppice).
4. DISCUSSION
4.1. Litterfall versus litterfall inputs
Annual litterfall production (table I) isthe main factor governing the annual poten-tial return of macronutrients (except K;table II). A noteworthy observation wasthat VR, being the oak forest with the poor-est soil (see soil pH, table I; also Martin et al.[28]) had a high potential return of Mn; thechestnut coppice also has a relatively highpotential return of Mn.
Leaves are the litter fraction accountingfor about 75 % of total bioelement return
by litter [16]. This value ranged from 70 to85 % of the total return; for Mg and Mn,the percentage of total return through theleaves may increase to 88 % in soils on
slates, indicating nutrition imbalance [27,28]. The opposite trend is seen for K in thechestnut coppice and this is consistent withthe findings of Pires et al. [38], who reported
that this percentage depends on forest man-agement.
Data relating to the water and bioelementfluxes in the four oak forests have previ-ously been discussed by Moreno et al. [34,35], and those concerning the chestnut cop-pice by Gallardo et al. [15]. These authorstook into account the monthly variation inwater composition and bioelement uptakeor leaching. In the contribution of bioele-ments through canopy leaching it is possibleto distinguish different sources [34]: bulkprecipitation, dry deposition, stemflow andthroughfal.
The measured contributions of C bythroughfall are similar to those reported bySanta Regina and Gallardo [42], Edmonds etal. [10] and Krivosonova et al. [20], butmuch greater than those described byStevens et al. [46] and Van Breemen et al.[48].
The high C input indicates a possiblelocal source of nutrient elements found inthe bulk precipitation, due to deposition ofsuspended particles coming from the forestitself [23, 39]. However, as estimated byMoreno [32], atmospheric dust only repre-sents 2-3 % of the N and Ca measured andless than 1 % of the remaining nutrients.
In general, the values of these macronu-trients are very similar to those obtained byother authors on the Iberian Peninsula [2,4, 8, 41] or slightly lower [42]. In all cases,the contributions may be considered mod-erate to low [37]. The lowest inputs ofbioelements by canopy leaching occurredon the plot on slates, pointing to lower soilfertility at this plot [28].
Nitrogen values are very similar to thosereported by Likens et al. [22] and Belillasand Roda [3], and clearly lower than thosefound in industrialized areas [7, 48].
Phosphorus is an element that tends tobe present at very low concentrations in bulk
precipitation [36], at values close to thoseof micronutrients; it is a bioelement with avery closed plant-soil cycle and the contri-bution from the atmosphere is low. Accord-

ingly, the contribution from bulk precipita-tion is very low, as reported by other authors(e.g. [22, 37, 42, 46]).
The oligoelements Fe, Mn, Zn and Cu,like N, have very low values, lower thanthose obtained in more industrialized areasbecause the precipitation in the area stud-ied (as pointed out) comes almost entirelyfrom the West (Atlantic Ocean), with littleor no influence from air masses comingfrom the East (continent).
The total contribution of all the elements
analysed in the throughfall water are greaterthan those of the rain water [36], indicatingthat the content of these elements in the rainwater increases as the water passes throughthe forest canopy. The elements showingthe greatest increase in concentration in thethroughfall water with respect to rain waterare P, K, Mn, Mg and C [34]. This increasein concentration is a fairly common phe-nomenon observed in forests [32, 37] andis due to different factors: thus, the evapo-ration of intercepted water contributes to asmall increase in concentration (about 19 %,[33]) and to a large extent the enrichmentin nutrients is due to the washing of drydepositions (mainly Ca, P, Fe, Cu and Zn,and Mg on some plots) and leaching pro-cesses (mainly C, Mg, K and Mn). The lat-ter four elements are considered to be read-
ily leachable by Tukey [47]. However, thesedata are not consistent with the findingsreported by Ferres et al. [ 12], for whom onlyK is clearly enriched during its passagethrough the canopy.
The results for Ca contrast with thosefound for Mg (table II); in this sense, in thefirst case low throughfall values are obtainedwith respect to those found in the literaturecited; by contrast, the values found for Mgare higher. This suggests that Mg replacesthe role of Ca owing to the scarceness ofthe latter element in the acid soils studied
[27].
The entry of C to the soil depends almostexclusively on the litter (table II), whilethroughfall input of C is not very relevant.
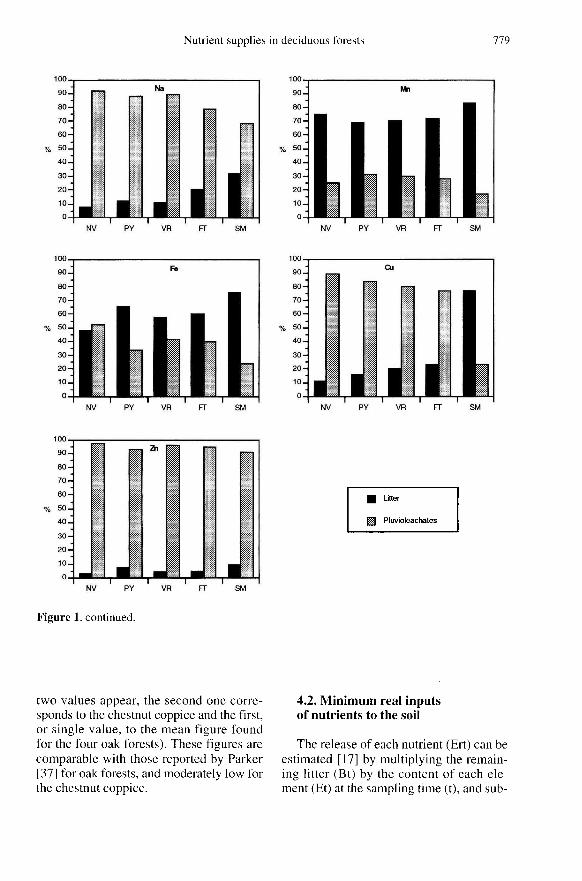
Nitrogen reaches the soil mainly throughthe litter (92 % at the oak forest and 98 % atthe chestnut coppice; figure 1), the canopynot contributing to the release of this ele-ment.
Ca and P also reach the soil mainlythrough the litter (table II), as found byParker [37]; there is also an important con-tribution of Ca by throughfall and of P bydry deposition [36].
The contributions of Mg by both routes(litter and throughfall) are very similar(except in the chestnut coppice, where thelitter contribution prevails; figure 1),throughfall being very important. By con-trast, in the case of K the major contribu-tions are due to throughfall (except at thechestnut coppice), although the litter isimportant (38 %;figure 1) and canopy leach-ing is also relevant (table II).
Na, Cu (except at the chestnut coppice)and Zn are mainly contributed by through-fall (table II); according to Moreno et al.[36] incident rainfall is very important asregards Na and Zn, and leaf leaching forCu.
To a large extent, Mn comes from the lit-ter [37], throughfall being relatively unim-portant (approximately 3 % on the oakstands and 1% at the chestnut coppice).Finally, the return of Fe is very similarthrough both routes (figure 1), the contri-bution due to throughfall being unimpor-tant.
Accordingly, the most important return ofC and N is through the litter, whereas thereturn of Na, Cu and Zn is greater throughthroughfall. Regarding the other elements,the contributions through both routes arebalanced, with the exception of P and Mn,which are slightly higher in the litter and Kin oak stand throughfall (figure 1). In thelight of these general characteristics, itshould be stressed that the return of Ca
through the litter, both at FG and at thechestnut coppice, represents 75 % of thetotal (figure 1) owing to the larger amountsreturned by this litter. Moreover, the higher

concentrations of Mg, P and K in chestnutleaves mean that the contributions are higherin the litter (table II), with percentages of72, 89 and 67 %, respectively.
In the light of the data offered in table II,it could be concluded that throughfall (a fac-
tor indicating nutrient exchange at canopylevel) represents mean percentages, with
respect to the total contribution, of 3 % forC; 4 % for Ca; 23-15 % for Mg; 12-1 %for P; 35-7 % for K; 15-8 % for Mn; 8 %for Fe; 45 % for Cu and 8 % for Zn (where
two values appear, the second one corre-
sponds to the chestnut coppice and the first,or single value, to the mean figure foundfor the four oak forests). These figures arecomparable with those reported by Parker[37] for oak forests, and moderately low forthe chestnut coppice.
4.2. Minimum real inputsof nutrients to the soil
The release of each nutrient (Ert) can beestimated [17] by multiplying the remain-ing litter (Bt) by the content of each ele-ment (Et) at the sampling time (t), and sub-

tracting the result from the initial contentof that nutrient in the litterfall biomass (Bo):
The minimum real contributions reachingthe soil annually through the leaves can beestimated since the leaves, as is known, rep-resent the main source of return in the lit-ter [30]. The data offered in table III arebased on knowledge of the mean potentialreturn through the leaf litter (table II) andthe capacity to release bioelements over3 years from the leaves contained in litter
bags [30, 45]. It should be noted that this isan underestimation of the actual return ofbioelements because in bags the degrada-tion processes are slowed down, and alsobecause, of the total litter, only the leavesare considered; one is thus referring to theminimum real inputs of available nutrientsto the soil.
The chestnut litter is the one that releasesthe largest amounts of bioelements over the3 years (table III) due both to a greaterpotential return (table II) and to a fasterdecomposition rate [30]. Despite this, thereare two elements (Na and Fe) that are notreleased in net form (negative sign intable III) owing to the strong degree of accu-mulation undergone during the first year ofdecomposition [30]. The greatest returnoccurs in the chestnut coppice at SM (themost demanding species). Among the oakforests, return depends on the elements(table III), although the lowest return val-ues are seen at the oak forest in VR sincethis is the most dystrophic ecosystem (seesoil pH, table I); such dystrophy is alsoreflected in the possible Ca/Mg nutritionalimbalance [28] since it is on this latter plot(VR) where the least return of Ca and thegreatest return of Mg occurred (table III) inthe oak forests.
The amount of P released by the leaves ishigher on granite soils than on soils devel-oped over slates; undoubtedly, the scarce-
ness of this (except at the FG and SM plots)must be the factor governing its retentionby microbial activity (biological immobi-lization [9]).
The losses of K from decomposing leavesare slightly higher in the oak forests devel-oped over granites than those located onslates (table III), although much lower thanthose seen at the chestnut coppice. Despitethe greater richness in K of the chestnut cop-pice floor [25], the greater requirement ofK on this plot leads its external cycle tobecome more fluid and its internal cycle tobecome more intense, canopy leaching beinglower (table II), with a more marked releaseduring decomposition (table III; [45]).
The behaviour of elements considered tobe minor ones in this study (Na, Mn, Fe, Cuand Zn) to a large extent depends on thecontributions through canopy leaching(table II) and soil conditions [28]. Thus, itmay be seen (table III) that in manyinstances these elements are accumulatedin the decomposing litter after 3 yearsbecause the needs of the plants for them arelow and are largely or even wholly supple-mented by the atmosphere [31].
It is possible to estimate the minimumannual amount of bioelements reaching thesoil by adding the amount of nutrientsreleased by decomposing leaves during thefirst 3 years of leaf decomposition (table III)to those afforded by throughfall (table II).These amounts will be underestimated if
only the leaf fraction of the litter is consid-ered and if one estimates what is releasedin 3 years [30, 45]. Accordingly, the actualamount of nutrients reaching the soil willrange between the values offered in table II
(maximum) and those shown in table III(minimum).
It should be stressed that the valuesobtained for the actual return of Na, Fe, Cuand Zn (negative values) by the leaves aredue to enrichment of the litters undergoingdecomposition due to external contributionsafter their emplacement [36, 45]. Thus, inthese cases no real return is produced by the


leaf litter after 3 years; additionally, owingto the high contents of these elements (Na,Cu and Zn) in throughfall, it could beassumed, as has been commented above,that such enrichments would be a result of
canopy leaching and would therefore rep-resent an amount of nutrients coming fromthe atmosphere or from the canopy that doesnot reach the soil but rather is retained inthe humus layer [26]. In view of this, oncalculating the total contribution of nutri-ents to the soil, it would be necessary to sub-tract that accumulation from the contribu-tion by throughfall. Fe, by contrast, shows adifferent trend since its contribution throughthroughfall is not sufficient to account forthe enrichment (negative values of the totalcontribution) at VR, FG and SM and hencean origin in the soil should be sought [28]. Inacid medium, the Fe content increases inthe soil solution [6], favouring greater immo-bilization by organisms, and hence the neg-ative value of the total contribution repre-sents the annual enrichment of the humusdue to the soil, apart from the fact that theactivity of the soil mesofauna also has acontaminating effect on the decomposinglitter by Fe.
Overall, it may be seen that the contri-butions of nutrients by throughfall are verysimilar to (and sometimes even higher than)the minimum contributions received by thesoil through litter decomposition, high-lighting the importance of this flow in forestnutrition.
5. CONCLUSIONS
The following main conclusions can bedrawn from the present work.
1) The greatest differences between theoak forests and the chestnut coppice lie inthe fact that in the latter ecosystem more N,P, K, Mg, Na and Mn potentially returnthrough the litter, undoubtedly due to agreater degree of tree uptake and/or pro-duction in the chestnut coppice. Among the
oak forests, VR shows a nutritional imbal-ance.
2) It is possible to differentiate threegroups of bioelements, namely: a) those thatpotentially return through the litter almostexclusively (C and N); b) those for whichboth the litter and throughfall must be takeninto account to explain their potential return(Ca, Mg, P, K, Fe and Mn); and c) thosethat return almost exclusively throughthroughfall (Na, Cu and Zn).
3) Despite the foregoing, after calcula-tion of the minimum annual returns, theabove three groups become reduced to two:bioelements that almost all return effectivelythrough canopy leaching (Na, Cu and Zn);and bioelements that return through bothlitter decomposition and throughfall. How-ever, Fe behaves in a special fashion in thesense that it tends to be immobilized by thedecomposing litter.
ACKNOWLEDGEMENTS
The authors thank the collaboration of theJunta de Castilla y León. This work was also
sponsored by the European Union (CAST/ENVI-RONMENT and MEDCOP/AIR Projects) andthe Spanish National Funds (CICYT/INIA). Tech-nical support was received from M. Tapia, M.L.Cosme, J. Hernández and C. Pérez. The Englishversion has been revised by N. Skinner and D.Garvey.
REFERENCES
[1] Aussenac G., Bonneau M., Le Tacon F., Resti-tution des éléments minéraux au sol par l’intermédi-aire de la litière et des précipitations dans quatre peu-plements forestiers de l’Est de la France, Oecol. Plant.7 (1972) 1-21.
[2] Avila A., Balanç d’aigua i nutrients en unaconca d’alzinar del Montseny. Estudis i Monografies,Diputación de Barcelona, 1988, 219 pp.
[3] Belillas M.C., Rodá F., Nutrient budgets in adry heatland watershed in northeastern Spain, Bio-geochemistry 13 (1991) 137-157.
[4] Bellot J., Análisis de los flujos de deposiciónglobal, translocación, escorrentía cortical y deposiciónseca en el encinar mediterráneo de L’Avic (Sierra de

Prades, Tarragona), thesis, University of Alicante,1989.
[5] Berg B., Lundmark J.E., Decomposition of nee-dle in Pinus contorta and P. sylvestris monocultures: acomparison, Scand. J. For. Res. 2 (1987) 3-12.
[6] Black C.A., Relaciones suelo-planta, HemisferioSur, Buenos Aires, 1975, 866 pp.
[7] Brechtel H.M.., Balazs F., Lehnardt F., Precip-itation input of inorganic chemicals in the open fieldand in forest stands. Results of investigations in theState of Hesse, in: Georgii H.W. (Ed.), AtmosphericPollutants in Forest Areas, D. Reidel, Dordrecht, 1986,pp. 47-67.
[8] Calvo D.E. Anta R.M., Paz A., Díaz-Fierros F.,Nuevos datos de la influencia en la vegetación en laformación de suelo en Galicia. II. Aportes de elemen-tos por lavado de la cubierta y tronco, An. Edaf. Agro-biol. 39 (1979) 1675-1691.
[9] Duchaufour P., Edafología: Edafogénesis yclasificación, Masson, Barcelona, 1984, 494 pp.
[10] Edmonds R.L., Thomas T.B., Rhodes J.J.,Canopy and soil modifications of precipitation chem-istry in a temperate rain forest, Soil Sci. Soc. Am. J. 55(1991) 1685-1693.
[11] F.A.O., Soil Map of the World: The Legend,F.A.O., Rome, 1991.
[12] Ferres L.L., Rodá F., Verdú A.M.C., TerradasJ., Circulación de nutrientes en algunos ecosistemasforestales del Monseny (Barcelona), Mediterranea, Ser.Biol. 7 (1984) 139-166.
[13] Gallardo J.F., Egido J.A., Prat L., Suelos fore-stales de El Rebollar (Salamanca). I. Consideracionesgenerales, Anu. Cent. Edafol. Biol. Apl. 6 (1980)193-213.
[14] Gallardo J.F., Cuadrado S., Egido J.A.,González M.I., Rico M., Santa Regina I., Gallego H.A.,Martin A., Menéndez I., Moreno G., Quilchano C,Schneider K, Turrión B., Forteza J., Saavedra J., Moy-ano A., Memoria final del Proyecto STEP/CEE: Nutri-ent cycling in degenerate natural forests in Europe inrelation to their rehabilitation, CSIC/University of Sala-manca, Salamanca, 1992, 201 pp.
[15] Gallardo J.F., Egido J.A., González M.I., RicoM., Santa Regina I., Gallego H., Martin A., Menén-dez I., Moreno G., Schneider K., Turrión B., Saave-dra J., Nutrient cycles in chestnut ecosystems of Sierrade Gata (western-central Spain), in: Romane F. (Ed.),Biological Criteria for Sustainable Developments inNatural Degenerate Forests of Mediterranean Europe:A Case Study of Chestnut Ecosystems, CNRS, Mont-pellier, 1994, pp. 23-44.
[16] Gallardo J.F., Martin A., Santa Regina I., Nutri-ent cycling in deciduous forest ecosystems of the Sierrade Gata mountains: aboveground litter production andpotential nutrient return, Ann. Sci. For. 55 (1998)749-769.
[ 17] Gallardo J.F., Santa Regina I., Harrison A.F.,Howard D.M., Organic matter and nutrient dynamics inthree ecosystems of the Sierra de Béjar mountains,Acta Oecol. 16 (1995) 447-459.
[18] Hernández I.M., Gallardo J.F., Santa Regina I.,Dynamic of organic matter in forests subject to aMediterranean semi-arid climate in the Duero Basin,Acta Oecol. 13 (1992) 55-65.
[19] Khanna P.K., Ulrich B., Ecochemistry of tem-perate deciduous forests, in: Röhrig E., Ulrich B. (Eds.),Ecosystems of the World. 7. Temperate DeciduousForests, Elsevier, Amsterdam, 1991, pp. 121-163.
[20] Krivonosova G.M., Yanova G.N., VerveykoY.I., Composition of precipitation and inputs of chem-ical elements into the soils of the Western forest steppeof the Ukraine, Agrokhimiya 5 (1990) 130-134.
[21] Lemee G., Recherches sur les écosystèmesdes reserves biologiques de la forêt de Fontainebleau.IV. Entrées d’éléments minéraux par les precipitationset transfert au sol par le pluviolessivage, Oecol. Plant.9 (1974) 187-200.
[22] Likens G.E., Bormann F.H., Pierce R.S., EatonJ.S., Johnson N.M., Biogeochemistry of a ForestedEcosystem, Springer Verlag, New York, 1977, 148 pp.
[23] Lindberg S.E., Lovett G.M., Richter D.D.,Johnson D.W., Atmospheric deposition and canopyinteraction of major ions in a forest, Science 231 ( 1986)141-145.
[24] Lossaint P., Influence de la composition chi-mique des litières forestières sur la vitesse de décom-position, these, Univesity of Strabourg, 1953, 107 pp.
[25] Mangenot F., Toutain F., Les litières, in: Pes-son P. (Ed.), Actualités d’écologie forestière, Gauthier-Villars, París, 1980, pp. 3-69.
[26] Martín A., Reciclado de bioelementos a travésde la hojarasca en ecosistemas forestales de la Sierra deGata (Sistema Central español), thesis, University ofSalamanca, 1995, 356 pp.
[27] Martin A., Rapp M., Santa Regina I., GallardoJ.F., Leaf litter decomposition dynamics in somemediterranean deciduous oaks, Eur. J. Soil Biol. 30(1994) 119-124.
[28] Martín A., Santa Regina I., Gallardo J.F., Inter-action between litter and soil epipedons in forest ecosys-tems of the Sierra de Gata mountains, Province deSalamanca, Spain, Arid Soil Res. Rehabil. 10 (1995)299-305.
[29] Martín A., Gallardo J.F., Santa Regina I.,Aboveground litter production and bioelement poten-tial return in an evergreen oak (Q. rotundifolia) wood-land near Salamanca, Ann. Sci. For. 53 (1996) 811-818.
[30] Martín A., Gallardo J.F., Santa Regina I., Long-term decomposition process of leaf litter from Quercuspyrenaica forest across a rainfall gradient (SpanishCentral System), Ann. Sci. For. 54 (1997) 191-202.
[31] Miller H.G., Cooper J.M., Miller J.D., Effectof nitrogen supply on nutrients in litter fall and crownleaching in a stand of Corsican pine, J. Appl. Ecol. 13(1976) 233-248.
[32] Miller H.G., Miller J.D., Cooper J.M., Trans-formation in rainwater chemistry on passing throughforested ecosystems, in: Coughtrey M., Martin M.,Unsworth M. (Eds.), Pollutant Transport and Fate inEcosystems, Blackwell, Oxford, 1987, pp. 171-180.

[33] Moreno G., Balances de agua y nutrientes enrebollares (Quercus pyrennica Willd.) de la vertientesalmantina de la Sierra de Gata, thesis, University ofSalamanca, 1994, 470 pp.
[34] Moreno G., Gallardo J.F., Cuadrado S.,Deposición atmosférica de bioelementos y su modifi-cación por la cubierta vegetal en bosques de Quercuspyrenaica de la Sierra de Gata (Salamanca), in: Gal-lardo J.F. (Ed.), Biogeoquímica de ecosistemas, Juntade Castilla y León, Valladolid, 1994, pp. 201-215.
[35] Moreno G., Gallardo J.F., Ingelmo F., Hernán-dez J., Soil water budget in four Quercus pyrenaicaforests across a rainfall gradient. Arid Soil Res. Reha-bil. 10 (1996) 65-84.
[36] Moreno G., Gallardo J.F., Schneider K.,Ingelmo F., Water and bioelement fluxes in four Quer-cus pyrenaica forests along a pluviometric gradient,Ann. Sci. For. 53 (1996) 625-639.
[37] Parker G.G., Throughfall and stemflow in theforest nutrient cycle, Adv. Ecol. Res. 13 (1983) 57-134.
[38] Pires A.L., Portela E., Martins A., Nutrientcycling in chestnut groves in the Tras-Os-Montesregion, in: Romane F. (Ed.), Biological Criteria forSustainable Developments in Natural DegenerateForests of Mediterranean Europe: A Case Study ofChestnut Ecosystems, CNRS, Montpellier, 1994, pp.9-22.
[39] Potter C.S., Ragsdale H.L., Swank W.T.,Atmospheric deposition and foliar leaching in a regen-erating Southern Appalachian forest canopy, J. Ecol. 79(1991) 97-115.
[40] Rapp M., Cicle de la matière organique et deséléments minéraux dans quelques écosystèmes méditer-ranéens, CNRS, Paris, 1971, 184 pp.
[41] Rodá F., Biogeoquímica de las aguas de Iluviay drenaje en algunos ecosistemas forestales delMontseny, thesis, Universidad Autónoma of Barcclona,1983, 428 pp.
[42] Santa Regina I., Gallardo J.F., Biogeochemi-cal cycles in forests of the Sierra de Béjar: return ofbioelements in rainfall, Acta Oecol., Oecol. Plant. 10( 1989 433-438.
[43] Santa Regina I., Gallardo J.F., San Miguel C.,Ciclos biogeoquímicos en bosques de la Sierra de Béjar(Salamanca, España): 2. Retorno potencial de bioele-mentos por medio de la hojarasca, Rev. Écol. Biol.Sol. 26 (1989) 155-170.
[44] Santa Regina I., Gallardo J.F., San Miguel C.,Ciclos biogeoquímieos en bosques de la Sierra de Béjar(Salamanca, España). 3. Descomposición de lahojarasca, Rev. Écol. Biol. Sol. 26 (1989) 407-416.
[45] Santa Regina I., Rapp M., Martin A., GallardoJ.F., Nutrient release dynamics in decomposing leaflitter in two Mediterranean decidous oak species, Ann.Sci. For. 54 (1997) 747-760.
[46] Stevens P.A., Hornung M., Hughes S., Soluteconcentration fluxes and major nutrient cycles in aMataure Stitka-Spruce plantation in Beddgelert forest,North Wales, For. Ecol. Manag. 27 (1989) 1-20.
[47] Tukey H.B., The leaching of substances fromplants, Annu. Rev. Plant Physiol. 21 (1970) 305-324.
[48] Van Breemen N., Visser W.F.J., Pape T.H.,Biogeochemistry of an oak-woodland ecosystem inthe Netherlands affected by acid atmospheric deposi-tion, Agric. Res. Repp., PUDOC, Wageningen, 1989,197 pp.




















