
repository.ipb.ac.idrepository.ipb.ac.id/jspui/bitstream/123456789/58953/1/...Judul Penelitian:...
228
DISERTASI MODEL PENGELOLAAN POPULASI BADAK JAWA (Rhinoceros sondaicus) BERDASARKAN ANALISIS NUTRISI DAN TINGKAT CEKAMAN SEBAGAI PARAMETER KESEHATAN ADHI RACHMAT SUDRAJAT HARIYADI B361080011 ILMU BIOMEDIS HEWAN SEKOLAH PASCA SARJANA INSTITUT PERTANIAN BOGOR 2012
Transcript of repository.ipb.ac.idrepository.ipb.ac.id/jspui/bitstream/123456789/58953/1/...Judul Penelitian:...

DISERTASI
MODEL PENGELOLAAN POPULASI BADAK
JAWA (Rhinoceros sondaicus) BERDASARKAN
ANALISIS NUTRISI DAN TINGKAT CEKAMAN
SEBAGAI PARAMETER KESEHATAN
ADHI RACHMAT SUDRAJAT HARIYADI
B361080011
ILMU BIOMEDIS HEWAN
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
2012

MODEL PENGELOLAAN POPULASI BADAK
JAWA (Rhinoceros sondaicus) BERDASARKAN
ANALISIS NUTRISI DAN TINGKAT CEKAMAN
SEBAGAI PARAMETER KESEHATAN
ADHI RACHMAT SUDRAJAT HARIYADI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Ilmu Biomedis hewan
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
2012

Judul Penelitian: Model Pengelolaan Populasi Badak Jawa (Rhinoceros sondaicus)
berdasarkan Analisis Nutrisi dan Tingkat Cekaman sebagai
Parameter Kesehatan
Nama: Adhi Rachmat Sudrajat Hariyadi
NRP: B361080011
Program Studi: Ilmu Biomedis Hewan
Menyetujui:
Komisi Pembimbing
Prof. Drh. Dondin Sajuthi, MST, PhD
Ketua
Dr. Drh. Hera Maheshwari, MSc Prof. Dr. Ir. Hadi S Alikodra MS
Anggota Anggota
Prof. Dr. Ir. Dewi Apri Astuti, MS
Anggota
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
drh.Agus Setiyono, MS, PhD, ApVet Dr. Ir. Dahrul Syah, M.Sc.Agr
Tanggal Ujian: Tanggal Lulus:

SURAT PERNYATAAN
Saya menyatakan dengan sebenar-benarnya bahwa segala pernyataan dalam
disertasi saya yang berjudul:
Model Pengelolaan Populasi Badak Jawa (Rhinoceros sondaicus)
Berdasarkan Analisis Nutrisi dan Tingkat Cekaman Sebagai Parameter
Kesehatan
Merupakan gagasan atau hasil penelitian disertasi saya sendiri dengan arahan
Komisi Pembimbing, kecuali dengan jelas ditunjukkan rujukannya. Disertasi ini
belum pernah diajukan untuk memperoleh gelar pada program sejenis di
perguruan tinggi lain. Semua data dan informasi yang digunkan telah dinyatakan
secara jelas dan dapat dipertanggungjawabkan kebenarannya.
Bogor, September 2012
Adhi Rachmat Sudrajat Hariyadi
NIM B361080011


ABSTRACT
ADHI RACHMAT SUDRAJAT HARIYADI. Model of population management for
javan rhinoceros (Rhinoceros sondaicus) based on analysis of nutrition and
stress levels as health parameters. Under the guidance DONDIN SAJUTHI,
HERA MAHESHWARI, HADI SUKADI ALIKODRA, and DEWI APRI ASTUTI.
This research was designed to establish a model for population
management for javan rhinoceros (Rhinoceros sondaicus), a critically
endangered species inhabiting Ujung Kulon National Park. Population increase
by 3% every year had been assigned by the Ministry of forestry and this rate of
population increase heavily relied on the ability of this species to survive and to
reproduce naturally. The habitat of the rhino has conditions that were not
always favourable to the rhinos and these conditions could potentially trigger
stress for these animals. The research was divided into three stages consisting of
analysis of nutritions and digestibility profiles of three male rhinoceros, analysis
of relationship between stress levels on hormone profile of these animals, and
stress induction to model animals. Two horses were selected as models for stress
on rhinoceros, as the stress induction could not be done on the rhinoceros
directly due to their small numbers worldwide; thus designated as a criticaly
endangered species. The results of this research showed that the feed profile
directly correlated with the home range and the vegetation structure around each
individual rhinoceros and there were indications that the rhinos experienced feed
deficit in quantity and quality. Analysis using 3α, 11β-dihydroxy-CM as an
indicator of glucocorticoid stress hormon levels suggested that the rhinos
experienced greater stress during the dry season when the water source was
significantly depleted. This fact was also consistent with the result from stress
induction on horse as model animal suggesting that feed deficit created acute
type stress while water deficit created a chronic type stress. Compilations of the
results from the research stages were formulated into a model of population
management for javan rhinoceros with recommendations such as: habitat
enrichment using feed plants high in protein (Moringa citrifolia, Callicarpa
longifolia, Chisocheton microcarphus), high in fat (Alstonia angustiloba,
Callicarpa longifolia, Macaranga spp), high in energy (Derris thyorsifolia,
Pterospermum javanicum, Percampyulus glances) with density of 15 per hectare
(tree form) to 5,406 per hectare (seedling form); water availability can be
enhanced by replenishing feed plants high in water contents such as: Paederia
scandens, Alstonia scholaris, Costus speciosus, as well as maintaining access
and trais to water sources that were available year round. The use of 3α, 11β-
dihydroxy-CM needs to be validated using biological and chemical measures to
investigate its potential use in monitoring stress levels of javan rhinos in the wild;
horse can be used as relevant model animal for research in stress and its
implications to the javan rhinoceros.
Keywords: javan rhinoceros, nutrients, digestibility, hormon, stress, model for
population management

ABSTRAK
ADHI RACHMAT SUDRAJAT HARIYADI. Model pengelolaan populasi
badak jawa (Rhinoceros sondaicus) berdasarkan analisis nutrisi dan tingkat
cekaman sebagai parameter kesehatan. Dibawah bimbingan DONDIN
SAJUTHI, HERA MAHESHWARI, HADI SUKADI ALIKODRA, dan DEWI
APRI ASTUTI.
Penelitian ini dirancang untuk membangun suatu model pengelolaan
populasi untuk badak jawa (Rhinoceros sondaicus) yang merupakan satwa langka
yang hidup di Taman Nasional Ujung Kulon. Peningkatan populasi sebesar 3%
setiap tahun bergantung pada kemampuan spesies ini untuk bertahan hidup dan
berkembang biak secara alami. Habitat yang dihuni oleh badak memiliki kondisi
yang tidak selalu mendukung terhadap kesehatan satwa ini dan berpotensi untuk
memicu cekaman. Penelitian dibagi menjadi tiga tahap yaitu kajian status nutrisi
dan kecernaan, kajian hubungan tingkat cekaman terhadap profil hormon pada
tiga ekor badak jantan, serta uji coba cekaman terhadap hewan model. Kuda
dipilih sebagai hewan model untuk mewakili fisiologi badak terhadap cekaman
karena uji coba cekaman tidak dapat dilakukan langsung terhadap badak jawa
yang merupakan satwa yang terancam punah karena jumlahnya yang sangat
sedikit di seluruh dunia. Profil pakan badak berkorelasi dengan ruang jelajah
yang ditempuhnya dan habitat tumbuhan di sekitarnya. Analisis menggunakan
3α, 11β-dihydroxy-CM sebagai indikator profil hormon glukokortikoid yang
merupakan respons terhadap cekaman menunjukkan bahwa badak cenderung
menunjukkan tingkat cekaman yang tinggi pada musim kering dimana
ketersediaan air sangat berkurang. Hal ini juga konsisten dengan uji coba
kuantifikasi yang dilakukan pada hewan model kuda dan berdasarkan parameter
hematologi (netrofil:limfosit) hasilnya menunjukkan bahwa defisit pakan
merupakan sumber cekaman yang memiliki karakteristik akut sementara defisit
air menunjukkan karakteristik kronis. Kompilasi dari simpulan dalam tahapan
penelitian ini dirumuskan menjadi suatu model pengelolaan populasi dengan
rekomendasi yang terdiri dari: pengkayaan habitat dengan tumbuhan yang kaya
protein (Moringa citrifolia, Callicarpa longifolia, Chisocheton microcarphus),
kaya lemak (Alstonia angustiloba, Callicarpa longifolia, Macaranga spp), dan
kaya energi (Derris thyorsifolia, Pterospermum javanicum, Percampyulus
glances) dalam kerapatan 15 individu per hektar (untuk jenis pohon) sampai
5,406 individu per hektar (untuk jenis semai); pemenuhan kebutuhan air
dilakukan dengan memperbanyak tumbuhan pakan dengan kadar air tinggi seperti
Paederia scandens, Alstonia scholaris, Costus speciosus serta menjamin
akses/jalur ke air yang tersedia sepanjang tahun; 3α, 11β-dihydroxy-CM perlu
menjalani validasi lebih lanjut (biologi dan kimia) untuk menjajagi potensi
penggunaannya dalam implementasi pemantauan cekaman pada badak;
penggunaan kuda sebagai hewan model yang relevan bagi badak dalam penelitian
cekaman serta implikasinya.
Kata kunci: badak jawa, nutrien, kecernaan, hormon, cekaman, hewan model,
pengelolaan

RINGKASAN
ADHI RACHMAT SUDRAJAT HARIYADI. Model pengelolaan populasi
badak jawa (Rhinoceros sondaicus) berdasarkan analisis nutrisi dan tingkat
cekaman sebagai parameter kesehatan. Dibawah bimbingan DONDIN
SAJUTHI, HERA MAHESHWARI, HADI SUKADI ALIKODRA, dan DEWI
APRI ASTUTI.
Badak jawa (Rhinoceros sondaicus) merupakan satwa langka yang hanya
terdapat di Taman Nasional Ujung Kulon, Banten Indonesia. Kondisi habitat
yang ada saat ini diduga mengalami perubahan perlahan akibat suksesi alami
yang berakibat pada berubahnya struktur vegetasi yang ada. Sebagai hewan
herbivora yang sangat bergantung pada vegetasi sebagai sumber makanannya,
maka faktor nutrisi menjadi bagian yang penting untuk dipantau dalam populasi
saat ini. Penelitian ini bertujuan untuk dapat mengetahui pola pergerakan badak
di habitatnya dan mengukur kualitas, palatabilitas, serta asupan nutrien dan
antinutrisi (toksin) dari badak yang diikuti. Parameter yang dicatat terdiri dari:
jalur lintasan badak (trajektori) beserta korelasinya dengan keragaman,
palatabilitas, jumlah konsumsi pakan, kualitas nutrien tumbuhan pakan, risiko
toksisitas lantaden dari tumbuhan Lantana camara, kecernaan, serta ketersediaan
garam di sekitar jalur lintasan dan di dalam ruang jelajah badak tersebut.
Palatabilitas ditentukan dengan memilih maksimal lima jenis tumbuhan pakan
dengan estimasi jumlah ragutan terbanyak dari setiap badak. Analisis proksimat
digunakan untuk mengukur dan menghitung kualitas nutrien yang ada pada setiap
pakan badak, sementara risiko toksisitas lantaden dari konsumsi Lantana camara
dihitung berdasarkan estimasi jumlah konsumsi kering tumbuhan Lantana
camara setiap hari. Acid insoluble ash (AIA) digunakan untuk mengukur tingkat
kecernaan dan menunjukkan kompatibilitas dengan hasil penghitungan kecernaan
berdasarkan koleksi total.

Hasil pengamatan menunjukkan korelasi yang kuat antara ruang jelajah
dengan keragaman pakan serta jumlah kubangan, tumbuhan dengan palatabilitas
tinggi bukan merupakan tumbuhan dengan kualitas nutrisi terbaik, serta ada
fluktuasi komposisi pakan yang menyebabkan rendahnya kualitas asupan air,
nutrien, dan energi pada waktu-waktu tertentu. Dinamika komposisi pakan yang
terjadi adalah: asupan air dari pakan berada pada tingkat yang rendah pada bulan
November (setelah musim kering di bulan Oktober); asupan protein menunjukkan
titik terendah di bulan Oktober (musim kering); asupan lemak menunjukkan titik
terendah pada bulan-bulan Oktober, November, Februari, Maret; dan asupan
energi terlihat ada pada tingkat terendah pada bulan Desember. Perbandingan
konsumsi lantaden pada badak jawa dengan referensi dari literatur yang
menyatakan konsumsi lantaden dapat mulai menimbulkan gejala klinis pada
kadar 4,000 mg/ekor/hari menunjukkan bahwa risiko toksisitas dari komsumsi
lantaden pada badak jawa berada pada tingkatan yang rendah yaitu: 174.15
mg/ekor/hari.
Salah satu aspek penting dalam pengelolaan populasi badak jawa
(Rhinoceros sondaicus) di Taman Nasional Ujung Kulon Banten adalah upaya
untuk mempelajari dan memantau tingkat cekaman yang dihadapi oleh badak
jawa di habitatnya. Tingkat kesulitan untuk mendapatkan sampel darah
mengharuskan peneliti untuk menggunakan sampel feses untuk assay hormon
cekaman. 3α, 11β-dihydroxy-CM merupakan salah satu metabolit glukokortikoid
yang layak digunakan untuk analisis tingkat cekaman pada badak jawa. Kajian
cekaman dilakukan antar individu dengan tingkat asupan pakan dan nutrien yang
berbeda dan juga antar musim untuk mempelajari variasi kadar glukokortikoid di
musim kering dan musim hujan. Hasil yang diperoleh menunjukkan bahwa
individu 18 (individu muda) mengalami cekaman yang fluktuatif yang mungkin
disebabkan oleh cekaman akibat interaksi sosial dengan badak dewasa. Badak
menunjukkan kecenderungan cekaman tinggi pada musim kering yang memiliki
jumlah kejadian hujan yang rendah yaitu rataan 0.2 kejadian hujan setiap harinya.
Penelitian ini menunjukkan bahwa keterbatasan air merupakan sumber cekaman
yang besar bagi badak jawa.

Setelah mempelajari faktor cekaman yang dihadapi oleh badak jawa
akibat defisit pakan dan defisit air, maka penelitian ini dirancang untuk lebih
mendalami respons yang ditunjukkan oleh kuda (sebagai hewan model untuk
badak) terhadap cekaman tersebut. Penelitian ini bertujuan untuk melihat respons
perilaku, fisiologis (hematologi dan respirasi) pada hewan model kuda akibat
cekaman defisit pakan dan air 50% dari kebutuhan normal. Perlakuan yang
diberikan adalah pakan kontrol sesuai kebutuhan (K) dan 50% dari kebutuhan
(50%K). Jumlah kuda yang dipakai terdiri dari dua ekor kuda jantan berusia
dewasa dan muda dengan bobot badan masing-masing 104 dan 98 kg. Induksi
cekaman dilakukan pada dua ekor kuda model dengan mengurangi asupan pakan
secara bertahap sampai asupan pakan hanya 2.65% dari berat tubuh kuda (defisit
pakan) dan pengurangan air minum sampai 50% dari jumlah ad libitum yang
biasanya tersedia di kandang. Parameter yang diamati terdiri dari hematologi
rutin dan kadar kortisol darah yang diukur selama periode kontrol dan perlakuan
induksi cekaman. Hasil penelitian menunjukkan bahwa kadar kortisol dan
hematologi (rasio netrofil:limfosit) menunjukkan bahwa defisit pakan merupakan
cekaman akut yang dapat diatasi oleh kuda (ditandai dengan penurunan kortisol
pada penghujung perlakuan). Kadar kortisol dan hematologi (rasio
netrofil:limfosit) menunjukkan bahwa defisit air merupakan cekaman yang
bersifat kronis dan tidak dapat diatasi dengan mudah. Dengan defisit pakan dan
air, kedua ekor kuda menunjukkan perubahan-perubahan: perilaku (mengurangi
aktifitas berjalan dan melakukan aktifitas menggigit ember/pagar / crib biting),
peningkatan kadar kortisol, hematologi (perubahan rasio Netrofil/limfosit yang
menunjukkan adanya migrasi limfosit/imunosupresi saat cekaman), dan
peningkatan respirasi pada kedua ekor kuda saat mengalami cekaman. Penelitian
ini menunjukkan bahwa tingginya cekaman (sebagaimana ditunjukkan oleh
tingginya kadar indikator hormon cekaman) terjadi bersamaan dengan
keterbatasan air di musim kering, dan ini dapat disimpulkan bahwa keterbatasan
air tersebut cenderung dapat mengakibatkan cekaman bagi badak jawa.
Untuk memastikan kuantitas dan kualitas pakan yang memadai, perlu ada
upaya pengkayaan habitat dengan mengurangi dominasi tumbuhan langkap
(Arenga obtusifolia) yang mendominasi hampir 70% dari ruang jelajah badak di

semenanjung Taman Nasional Ujung Kulon, serta pengendalian tumbuhan
Lantana camara yang menempati hampir 60% areal pakan badak (rumpang)
terutama di daerah utara. Intervensi seperti ini akan memicu pertumbuhan jumlah
ketersediaan tumbuhan pakan yang disukai badak. Kualitas nutrisi pakan badak
dapat ditingkatkan dengan menanam tumbuhan pakan yang terdiri dari: Moringa
citrifolia, Callicarpa longifolia, Chisocheton microcarphus, (protein tinggi);
Alstonia angustiloba, Callicarpa longifolia, Macaranga spp, (lemak tinggi);
Derris thyorsifolia, Pterospermum javanicum, Percampyulus glances, (energy
tinggi); Paederia scandens, Alstonia scholaris, Costus speciosus (kandungan air
tinggi).
Tumbuhan-tumbuhan ini idealnya ditanam dalam komposisi kerapatan
tertentu agar ketersediannya berimbang. Ketersediaan air sepanjang tahun dapat
dipastikan dengan membuat jalur lintasan untuk badak ke lokasi-lokasi air dan
kubangan yang tersedia sepanjang tahun serta membuat kubangan ataupun
sumber air buatan yang dapat memberikan pasokan air bagi badak yang
menjelajah ke lokasi tersebut.

© Hak cipta milik Institut Pertanian Bogor, tahun 2012
Hak cipta dilindungi
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisankritik, atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan
yang wajar IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis
dalam bentuk apapu tanpa izin tertulis dari
IPB


MODEL PENGELOLAAN POPULASI BADAK
JAWA (Rhinoceros sondaicus) BERDASARKAN
ANALISIS NUTRISI DAN TINGKAT
CEKAMAN
SEBAGAI PARAMETER KESEHATAN
ADHI RACHMAT SUDRAJAT HARIYADI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Ilmu Biomedis hewan
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
2012


Judul Penelitian: Model Pengelolaan Populasi Badak Jawa (Rhinoceros sondaicus)
berdasarkan Analisis Nutrisi dan Tingkat Cekaman sebagai Parameter
Kesehatan
Nama: Adhi Rachmat Sudrajat Hariyadi
NRP: B361080011
Program Studi: Ilmu Biomedis Hewan
Menyetujui:
Komisi Pembimbing
Prof. Drh. Dondin Sajuthi, MST, PhD
Ketua
Dr. Drh. Hera Maheshwari, MSc Prof. Dr. Ir. Hadi S Alikodra MS
Anggota Anggota
Prof. Dr. Ir. Dewi Apri Astuti, MS
Anggota
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
drh.Agus Setiyono, MS, PhD, ApVet Dr. Ir. Dahrul Syah, M.Sc.Agr
Tanggal Lulus:

RIWAYAT HIDUP
Penulis lahir di Jakarta pada tanggal 28 April 1971 dan merupakan anak
pertama dari 2 bersaudara dari ayah yang bernama Hariyadi Suwandar dan ibu bernama
Mita Djuwita. Sejak kecil penulis tertarik dengan kegiatan alam terbuka dan juga
mempelajari ilmu biologi. Dengan berbekal latar belakang ini, pendidikan S1 di bidang
biologi dari University of Alberta, Kanada diselesaikan pada tahun 1994 dengan gelar
Bachelor of Science (BSc). Selepas pendidikan, penulis menempuh berbagai
pengalaman kerja antara lain sebagai teknisi karantina dan kesehatan ikan di SeaWorld
Indonesia, konsultan biologi, sampai akhirnya menangani kegiatan pelestarian satwa
badak jawa sebagai manajer proyek WWF Indonesia di Taman Nasional Ujung Kulon.
Didorong oleh keinginan untuk mempelajari kesehatan pada satwa liar, penulis
mengikuti pendidikan Magister Pascasarjana Program Studi Sains Veteriner di Institut
Pertanian Bogor sejak tahun 2004, lulus pada tahun 2006, dan setelah itu membimbing
beberapa mahasiswa kedokteran hewan IPB yang memiliki minat untuk mempelajari
kesehatan satwa liar.
Untuk lebih memperdalam pengetahuan di bidang kesehatan satwa liar,
khususnya badak jawa, penulis bergabung dengan program Doktor dalam bidang Ilmu
Biomedis hewan di Sekolah Pascasarjana IPB Bogor sejak tahun 2008 sambil
melibatkan diri untuk bekerja di kawasan konservasi Taman Nasional Ujung Kulon-
Banten, Bukit Barisan Selatan-Lampung, dan Kawasan ekosistem Leuser-Aceh dalam
upaya melestarikan spesies badak di Indonesia.
Penulis telah menerbitkan beberapa publikasi mengenai badak yang terdiri dari:
dua buah artikel jurnal ilmiah di jurnal Pachyderm milik Rhino Specialist Group, dua
artikel di jurnal veteriner sebagai penulis kedua, dan dua buah artikel yang sedang
dalam proses peer review di jurnal Pachyderm dan jurnal on-line Biomed Research
Notes


PRAKATA
Penulis mengucapkan puji syukur kepada Allah SWT yang telah memberi jalan,
petunjuk, dan kekuatan selama penulis menempuh studi di sekolah pasca sarjana
Institut Pertanian Bogor. Disertasi ini tidak mungkin dapat tersusun tanpa bantuan dan
dukungan moral dari keluarga tercinta yaitu Istri: Indriani Noverita, serta ananda:
Rahadrian Ksatria, dan Ayodhya Tangguh yang selalu memberikan motivasi dan juga
inspirasi.
Rasa terimakasih dan penghormatan yang sedalam-dalamnya penulis ucapkan
kepada para dosen pembimbing: Prof. drh. Dondin Sajuthi, MST, PhD, Prof. Dr. Dewi
Apri Astuti, MSc, Prof. Dr. Ir. Hadi S. Alikodra, MS, dan Dr. drh. Hera maheshwari,
MSc, yang dengan penuh kesabaran dan dedikasi memberikan pengarahan dan
masukan yang sangat berarti dalam penelitian ini. Terimakasih dan penghormatan juga
penuli sberikan kepada Dr. drh. M. Agil yang telah banyak memberi masukan dan
arahan selama pendidikan, dan kepada Handayani, drh. Dedi Setiadi, Msi, dan Ir.
Ghalib, Msc yang telah banyak membantu kerja penulis di laboratorium, serta kepada
ibu Nani di PSSP atas perhatian dan kesabarannya dalam menyusun keadministrasian
dan penjadwalan..
Penelitian ini didukung pula oleh kekompakan rekan-rekan WWF proyek Ujung
Kulon: Timer, Andri, Nina, Yuyun, Mutia, Ngatiman, Iwan, Mawi, Rani, Makmun, dan
Neng yang berhasil menjalankan kegiatan kantor secara sempurna selama penulis
meninggalkan tugas-tugas kantor untuk menempuh pendidikan dan juga terimakasih tak
terhingga pada Maryono dan Diki yang telah mengantar penulis bepergian dari tempat
kerja ke kampus tanpa kenal lelah mulai sejak dini hari sampai sore, dan bahkan malam
hari.

Penghargaan yang besar penulis sampaikan kepada tim ROAM Taman Nasional
Ujung Kulon: Daryan, Hendra Purnama, Asep Yayusm dan masyarakat desa Ujung
Jaya, kecamatan Sumur, Kabupaten Pandeglang, Provinsi Banten memberikan bantuan
dan dukungan selama pengambilan data di lapangan dan mendampingi perjalanan
menembus belantara Taman Nasional Ujung Kulon untuk mengikuti gerak langkah
badak jawa.
Semoga Allah memberikan yang terbaik
Bogor, September 2012
Adhi Rachmat Sudrajat Hariyadi

i
DAFTAR ISI
Halaman
DAFTAR ISI......................................................................................................... i
DAFTAR TABEL................................................................................................. v
DAFTAR GAMBAR............................................................................................. vii
DAFTAR LAMPIRAN......................................................................................... x
PENDAHULUAN................................................................................................ 1
Permasalahan............................................................................................ 2
Pendekatan Konservasi Melalui Aspek Medis.......................................... 3
Tujuan Penelitian............................................................................... ........ 4
Hipotesis Penelitian................................................................................... 5
Novelty/kebaruan............................................................................... ........ 5
Alur Penelitian........................................................................................... 6
Metode Umum........................................................................................... 7
Tahap I: Habitat Badak dan Profil Nutrisi................................................. 8
Tahap II: Habitat Badak dan Profil Cekaman........................................... 8
Tahap III: Simulasi Cekaman pada Hewan Model.................................... 9
TINJAUAN PUSTAKA........................................................................................ 10
Habitat Badak Jawa di Taman Nasional Ujung Kulon............................... 10
Informasi Umum tentang Badak Jawa........................................................ 11
Jenis-jenis Cekaman.................................................................................. 14
Cekaman Akut........................................................................................... 14
Cekaman Kronis........................................................................................ 15
Penyakit-penyakit pada Badak........................................................... ........ 15
Pengaruh Perubahan Iklim......................................................................... 16
Mekanisme Fisiologis Terhadap Cekaman Kekeringan................................ 18
Termoregulasi............................................................................................ 18
Kondisi Kulit............................................................................................. 18
Defisit Glukosa........................................................................................... 24
Lapar Serum............................................................................................... 26
Ambang Batas dan Proses Perbaikan dalam Kondisi Lapar Sel................. 27
Peranan dan Interaksi Hormon Kortisol dan Tiroksin sebagai Caraka...... 29
Lantana camara sebagai Bahan Toksin / Antinutrisi................................. 31
Hewan-hewan Model dan Pembanding untuk Badak Jawa......................... 33
Badak Sumatra (Dicerorhinus bicornis).................................................... 33
Badak India (Rhinoceros unicornis)........................................................... 34
Kuda............................................................................................................ 34
Analogi Perbandingan................................................................................. 36
Daftar pustaka.............................................................................................. 38
STUDI POLA PERGERAKAN, PALATABILITAS, KUALITAS PAKAN, DAN
KECERNAAN NUTRIEN SERTA RISIKO TOKSISITAS LANTADEN PADA
BADAK JAWA DI TAMAN NASIONAL UJUNG
KULON................................................................................................................. 47
Abstrak...................................................................................................... 47
Abstract...................................................................................................... 48
Pendahuluan............................................................................................... 49
Tujuan Penelitian............................................................................ 50

ii
Bahan dan Metode...................................................................................... 51
Ruang Jelajah Badak Jawa............................................................... 51
Palatabilitas.................................................................................... 54
Kualitas Pakan............................................................................. 55
Konsumsi Pakan............................................................................. 56
Kecernaan....................................................................................... 58
Metode AIA (Acid insoluble ash) menurut Van Keulen & Young.. 60
Risiko toksisitas dari konsumsi Lantana camara........................... 61
Ketersediaan Garam......................................................................... 61
Analisis Data..................................................................................... 62
Hasil dan Pembahasan................................................................................ 63
Karakteristik Ruang Jelajah Badak................................................... 63
Palatabilitas dan Keragaman Pakan................................................. 66
Kualitas dan Kuantitas Asupan Nutrien......................................... 67
Identifikasi Tumbuhan Pakan dengan Kualitas Nutrisi Tinggi....... 68
Kecernaan....................................................................................... 71
Kajian Risiko Toksisitas Lantaden................................................... 72
Ketersediaan Garam dan Air di Lokasi Kubangan............................ 73
Simpulan dan Saran.................................................................................... 75
Simpulan......................................................................................... 75
Saran................................................................................................ 76
Daftar pustaka............................................................................................. 77
PENENTUAN ASAI HORMON GLUKOKORTIKOID UNTUK MEMANTAU
INDIKATOR CEKAMAN PADA BADAK JAWA
JANTAN.................................................................................................................... 79
Abstrak........................................................................................................ 79
Abstract........................................................................................................ 80
Pendahuluan.................................................................................................. 81
Mekanisme dan Respons Individu terhadap Cekaman..................... 81
Definisi Cekaman............................................................................. 83
Hormon Terkait Cekaman Beserta Metabolitnya............................. 83
Tujuan penelitian............................................................................. 85
Bahan dan Metoda...................................................................................... 86
Penetapan Faktor Cekaman di Habitat Badak Jawa........................ 86
Penetapan Sampel Feses.................................................................. 86
Pengumpulan Sampel Feses............................................................ 87
Pemilihan Indikator Kadar Hormon Cekaman Asal Feses.............. 87
Persiapan Sampel Feses................................................................... 88
Asai Kortisol.................................................................................... 89
Prosedur Asai Kortisol..................................................................... 89
Asai Kortikosteron........................................................................... 90
Prosedur Asai Kortikosteron............................................................ 90
Asai 3α, 11β-dihydroxy-CM............................................................ 91
Prosedur Asai 3α, 11β-dihydroxy-CM............................................. 91
Analisis Data.................................................................................... 93
Hasil dan pembahasan............................................................................... 94

iii
Hasil Uji Paralelisme......................................................................... 94
Faktor Pengenceran untuk Kuantifikasi Metabolit
Glukokortikoid (3α, 11β-dihydroxy-
CM).................................................................................................... 94
Profil 3α, 11β-dihydroxy-CM pada Individu Badak & Populasi....... 95
Kondisi cekaman............................................................................... 97
Faktor cekaman dan Profil Glukokortikoid..................................... 98
Simpulan dan saran..................................................................................... 100
Simpulan.......................................................................................... 101
Saran................................................................................................. 101
Daftar pustaka.............................................................................................. 102
KAJIAN KUANTITATIF MENGGUNAKAN HEWAN MODEL SEBAGAI
PEMBANDING UNTUK BADAK JAWA DALAM PENELITIAN CEKAMAN
AKIBAT DEFISIENSI PAKAN DAN DEFISIENSI AIR...................................... 105
Abstrak......................................................................................................... 105
Abstract........................................................................................................ 106
Pendahuluan................................................................................................. 107
Proses Pemilihan Hewan Model........................................................ 108
1) Kelompok Hewan Model Konvensional..................................... 108
2) Kelompok Hewan Badak (Rhinoceratidae)................................ 109
Tujuan Penelitian.............................................................................. 110
Bahan dan metoda........................................................................................ 112
Perbandingan Perilaku...................................................................... 112
Pengambilan Data Pakan dan Air Minum (Fase Kontrol)............... 113
Induksi Cekaman.............................................................................. 114
1) Perlakuan Defisiensi Pakan........................................................ 114
2) Perlakuan Defisiensi Air.............................................................. 115
Periode “Pencucian”......................................................................... 116
Pengamatan Pola Respirasi................................................................ 116
Penghitungan Konsumsi dan Kecernaan Kuda................................. 116
Analisis Data..................................................................................... 117
Hasil dan Pembahasan.................................................................................. 118
Pemilihan Hewan Model.................................................................... 118
Perbandingan Perilaku Hewan Model............................................... 118
Proses Adaptasi Kuda di Dalam Kandang........................................ 120
1) Perilaku..................................................................................... 120
2) Tingkat Cekaman....................................................................... 121
3) Hematologi............................................................................... 122
Induksi Cekaman dengan Defisit Pakan............................................ 123
Respons Terhadap Perlakuan Defisiensi Pakan................................ 124
1) Perilaku.................................................................................... 124
2) Tingkat Cekaman...................................................................... 124
3) Hematologi............................................................................... 125
Respons Terhadap Perlakuan Defisiensi Air.................................... 126
1) Perilaku.................................................................................... 126
2) Tingkat Cekaman...................................................................... 127
3) Hematologi............................................................................... 129
Pola Respirasi dalam Kondisi Cekaman............................................ 130

iv
Identifikasi Komponen Pemicu Cekaman…………………………. 132
Kecernaan dalam Kondisi Cekaman................................................. 132
Adaptasi Fisiologis terhadap Cekaman............................................. 135
Simpulan dan saran..................................................................................... 136
Simpulan.......................................................................................... 136
Saran................................................................................................ 136
Daftar pustaka.............................................................................................. 137
PEMBAHASAN UMUM: MODEL DALAM PENGELOLAAN POPULASI BADAK
JAWA...................................................................................................................... 141
Pendahuluan............................................................................................... 141
Tekanan yang dihadapi badak jawa................................................. 143
1) Defisit Energi Asal Pakan.......................................................... 143
2) Defisit Air................................................................................. 144
Model Pengelolaan Badak Jawa....................................................... 145
Komponen 1: Teknik Pemantauan................................................... 146
Pemantauan Status Nutrisi.................................................... 146
Pemantauan Cekaman Berdasarkan Profil Hormon.............. 146
Komponen 2: Pengkayaan Nutrisi.................................................... 147
Menekan Laju Invasi dan Dominasi Tumbuhan
Langkap (Arenga obtusifolia)............................................... 147
Pengkayaan Air, Nutrien, & Energi dari Tumbuhan Pakan... 149
Pengendalian Lantana camara............................................. 150
Komponen 3: Mitigasi Cekaman Akibat Defisit Air......................... 151
Komponen 4: Penggunaan Hewan Model Untuk Riset..................... 152
Kecernaan dan Nutrisi........................................................... 152
Pencegahan Cekaman dan Implikasi Kesehatan.................... 153
Faktor-faktor lain yang relevan......................................................... 153
1) Faktor Habitat.............................................................................. 153
2) Faktor Populasi............................................................................ 153
3) Faktor Individu dan Perilaku Hewan........................................... 154
SIMPULAN DAN SARAN UMUM....................................................................... 155
Simpulan umum.......................................................................................... 155
Saran umum................................................................................................ 156
DAFTAR PUSTAKA.............................................................................................. 157

v
DAFTAR TABEL
Halaman
1 Daftar karakteristik antara badak jawa dengan hewan model yang
digunakan dalam penelitian (kuda dan badak sumatra)...........................
37
2 Jenis vegetasi tumbuhan paling dominan di lokasi ruang jelajah badak
jawa………………………………………………………………………
52
3 Rangkuman luas ruang jelajah dan jarak tempuh tiap ekor badak........... 64
4 Badak jawa, lokasi, dan jenis pakan dengan palatabilitas tertinggi
(persentase konsumsi basah rata-rata per hari) selama pengamatan bulan
Oktober 2009 sampai April 2010..............................................................
67
5 Komposisi nutrien dan energi dari tumbuhan pakan yang disukai badak
jawa (palatabilitas tinggi)........................................................................
67
6 Daftar tumbuhan pakan dengan kandungan air, nutrien, dan energi
tertinggi.....................................................................................................
68
7 Data asupan nutrisi harian badak
jawa............................................................................................................
70
8 Rataan asupan air, protein, lemak, dan energi per ekor per hari pada tiga
ekor badak yang diamati dalam penelitian...............................................
71
9 Persen kecernaan berat kering dari tiga ekor badak dengan koreksi 10%
menurut Mainka et al. (1989).....................................................................
71
10 Estimasi penghitungan asupan antinutrisi lantaden harian melalui
konsumsi tumbuhan Lantana camara pada badak jawa
(g/ek/h)......................................................................................................
73
11 Perbandingan hewan model serta kriteria pengamatan yang relevan
dengan penelitian pada badak jawa (dari berbagai sumber).....................
110
12 Pemberian air minum untuk setiap ekor kuda pada periode perlakuan
defisit air.....................................................................................................
115

vi
13 Ethogram (daftar perilaku) berdasarkan pengamatan kuda, badak jawa,
dan badak sumatra. Ethogram badak sumatra disusun berdasarkan
Siswandi et al.
(2005).........................................................................................................
119
14 Konsumsi pakan (% bahan kering per berat badan kuda)......................... 123
15 Konsumsi nutrien setiap ekor kuda selama periode perlakuan defisiensi
pakan...........................................................................................................
123
16 Persen kecernaan dua ekor kuda dengan koreksi 10 % menurut Mainka
et al. (1989).................................................................................................
133

vii
DAFTAR GAMBAR
Halaman
1 Skema alur penelitian serta kontribusi yang dihasilkan terhadap
konservasi badak jawa. Garis putus mewakili aspek konservasi
(Pemanfaatan dan Riset) yang tidak secara langsung difokuskan dalam
penelitian ini..............................................................................................
7
2 Badak jawa jantan yang tertangkap oleh kamera otomatis (kamera jebak)
di Taman Nasional Ujung Kulon, Banten. Foto: WWF Indonesia –
Balai Taman Nasional Ujung Kulon.........................................................
12
3 Struktur kulit badak afrika dalam keadaan normal yang terdiri dari
lapisan keratin pada bagian terluar. Pada gambar ini terlihat lapisan
keratin mengelupas dan digantikan dengan lapisan yang baru. Sumber:
Munson et al. (1998).................................................................................
19
4 Histopatologi pada epidermis badak (badak afrika) yang menunjukkan
adanya degenerasi hidroskopik, dan pembentukan celah/retakan pada
kulit. Tidak ada inflamasi ditemukan pada lesio ini. Pewarnaan HE, bar
= 100 mikron. Sumber: Munson et al. (1998)..........................................
20
5 Rangkaian reaksi yang dipicu oleh kondisi lapar sel (menurunnya kadar
gula dalam sel) yang berujung pada pemisahan sel (cell detachments),
dan kematian sel. Sumber: Suzuki et al (2003), Wu et al (2001), dan
Cavaliere et al (2001)...............................................................................
29
6 Tumbuhan perdu Lantana camara di habitat badak di Taman Nasional
Ujung Kulon memiliki bunga berwarna cerah. Foto: Ahariyadi – WWF
Indonesia...................................................................................................
32
7 Filogram yang menunjukkan kekerabatan berbagai spesies berdasarkan
variasi pada DNA 12S RNA. Angka menunjukkan kedekatan
kekerabatan berdasarkan perhitungan parsimoni maksimum. Sumber:
Fernando et al. (2005)...............................................................................
35
8 Tiga ekor badak jantan yang dipilih sebayak obyek penelitian: badak
nomor 12 (A), nomor 13(B), dan nomor 18(C). Foto: Balai Taman
Nasional Ujung Kulon-WWF Indonesia..................................................
53
9 Kondisi lahan pakan badak (rumpang) dengan perbandingan tinggi
tumbuhan pakan dan manusia (A) serta kondisi rumpang yang relatif
terbuka dibandingkan areal pepohonan di sekitarnya (B). Foto:
RSetiawan – WWF Indonesia...................................................................
54

viii
10 Contoh jejak badak yang relatif segar. Badak berada di lokasi ini 12 jam
sebelum jejak ditemukan. Foto: Ahariyadi-WWF Indonesia...................
59
11 Pola lintasan badak no 13 di Utara (A), badak 18 di Barat (B), dan badak
no 12 di Selatan (C) beserta aktifitas dari masing-masing badak
sebagaimana tercatat dalam survey lapangan..........................................
65
12 Beberapa contoh tumbuhan pakan badak Jawa yang tercatat dalam
pengamatan di lapangan. Foto di atas menunjukkan jenis tumbuhan
tepus: Amomum sp (A), Rotan: Calamus sp (B), dan Mara: Mallotus
floribundus (C). Foto: Rsetiawan – WWF Indonesia...............................
66
13 Rentang kandungan NaCl dalam tanah di empat lokasi kubangan yang
berada dalam lingkup ruang jelajah tiga ekor badak jawa yang diamati
dalam penelitian........................................................................................
74
14 Reaksi biokimia yang merupakan proses tanggap terhadap cekaman bagi
mamalia (sumber: Coenen 2005)..............................................................
82
15 Grafik paralelisme antara kadar 5-beta androstandiol (5-beta adiol) pada
feses dengan kurva standard berdasarkan hasil uji paralelisme......
95
16 Fluktuasi kadar hormon glukokortikoid pada semua individu badak....... 96
17 Rentang kadar metabolit glukokortikoid (5-beta adiol) antar individu
badak..........................................................................................................
96
18 Karakteristik musim pada periode pengamatan (bulan Oktober 2009-
April 2010) berdasarkan rata-rata jumlah kejadian hujan, cerah, ataupun
berawan dalam satu hari............................................................................
97
19 Perbandingan tingkat konsumsi energi per berat badan (sumbu x)
dengan kadar metabolit glukokortikoid pada feses………………………
99
20 Perbandingan kadar hormon cekaman antar individu badak dengan
asupan berat kering yang berbeda (A) dan antar musim/curah hujan
(B)...............................................................................................................
99
21 Perbandingan anatomi system pencernaan kuda (A) dan badak (B) yang
menunjukkan karakteristik hewan monogastrik (lambung satu) dan
keberadaan sekum untuk pencernaan selulosa. Sumber:
www.wren.aps.uo.guelph.ca......................................................................
108
22 Kuda jantan dewasa bernama Garuda (A) dan kuda jantan dewasa muda
bernama Elang (B) yang digunakan sebagai hewan model dalam
pengamatan di Rumah Sakit Hewan IPB Darmaga...................................
113
23 Komposisi aktifitas badak 18 (A), badak 12 (B), dan badak 13 (C)
berdasarkan durasi yang terekam dengan menggunakan kamera
otomatis.....................................................................................................
120

ix
24 Komposisi perilaku kuda jantan muda (A) dan jantan dewasa (B) yang
tercatat dalam pengamatan perilaku........................................................
120
25 Komposisi perilaku kuda Dewasa (A) dan kuda remaja (B) selama
periode aklimatisasi di kandang................................................................
122
26 Penurunan kadar kortisol dalam darah pada kedua ekor hewan (Garuda
dan Elang) selama periode aklimatisasi di kandang.................................
122
27 Fluktuasi kadar kortisol pada hewan pada perlakuan minus pakan......... 125
28 Perbandingan perilaku hewan model kuda jantan dewasa (A) dan kuda
jantan muda (B) dalam berbagai perlakuan..............................................
128
29 Trend peningkatan kadar kortisol dalam darah pada perlakuan minus air. 128
30 Trend peningkatan respirasi pada kuda dalam perlakuan defisit pakan
(A) dan defisit air (B) dibanding dengan kondisi normal istirahat.........
131
31 Perbandingan peningkatan respirasi badak dan kuda dalam kondisi
normal dan cekaman (stress).....................................................................
131
32 Trend kecernaan badak dan kuda dalam kondisi normal dan cekaman
(stress)........................................................................................................
134
33 Lokasi temuan kematian badak Jawa dalam periode tahun 2000-2010.
Titik hijau menunjukkan kasus dengan informasi yang relatif lengkap
mengenai penyebab kematian...................................................................
142
34 Model pengelolaan populasi dan habitat badak jawa dengan pendekatan
aspek nutrisi, cekaman, dan kesehatan......................................................
146
35 Distribusi badak jawa di semenanjung Taman Nasional Ujung Kulon
(titik merah) berdasarkan hasil survey 2005-2010. (sumber: WWF
Indonesia & Balai TNUK).........................................................................
148
36 Lokasi ideal untuk plot pengendalian langkap (Arenga obtusifolia) di
semanjung Ujung Kulon ditandai dengan poligon berwarna merah........
149
37 Usulan areal pengendalian tumbuhan Lantana camara di areal pakan
badak ditandai dengan poligon berwarna kuning......................................
151

x
DAFTAR LAMPIRAN
1 IZIN KOMISI ETIK HEWAN................................................................... 171
2 DATA NUTRISI BADAK JAWA............................................................. 175
3 DATA HEWAN MODEL.......................................................................... 181
4 DATA PARALELISME KIT HORMON.................................................. 191

PENDAHULUAN
Badak jawa (Rhinoceros sondaicus) merupakan mamalia besar yang tergolong
langka karena jumlahnya tidak melebihi 60 ekor di seluruh dunia, sehingga IUCN
memasukan badak jawa dalam kategori terancam punah. Badak jawa berada pada dua
lokasi yaitu: sekitar delapan ekor di Taman Nasional Cat Tien Vietnam dan sekitar 50
ekor di Taman Nasional Ujung Kulon di Indonesia. Tentang jumlah dan keragaman
genetika yang ada saat ini, para ahli berpendapat bahwa populasi di Taman Nasional
Ujung Kulon adalah satu-satunya populasi yang berpotensi untuk berkembang biak,
dan merupakan kunci kelestarian spesies ini di muka bumi. Hal ini bukanlah hal yang
ideal dari segi konservasi, karena dengan hanya bertumpu pada satu populasi saja,
justru risiko kepunahan akan semakin besar. Satu populasi memiliki tingkat
kerentanan yang tinggi, sehingga bencana alam katastropik (gunung berapi, tsunami)
dan/atau epidemik yang melanda populasi di Ujung Kulon akan menyebabkan
mortalitas massal yang berpotensi melenyapkan spesies ini.
Badak jawa yang telah bertahan hidup di Ujung Kulon dalam waktu yang lama
sempat mengalami penurunan jumlah populasi di tahun 1962 mencapai jumlah hanya
25 ekor saja pada saat itu. Saat ini badak jawa telah mengalami peningkatan dari
jumlah populasi di tahun 1962 ke jumlah sekitar 50 ekor, dan terbentuk paradigma
yang beranggapan bahwa badak jawa merupakan satwa yang tangguh dan selama
mereka berada di habitat alami, kesehatan mereka termasuk baik. Upaya pelestarian
badak jawa umumnya dititik beratkan pada upaya pengamanan dan pengelolaan
habitat, sementara aspek medis konservasi yang dibutuhkan untuk meningkatkan
peluang keberlangsungan spesies ini menjadi terabaikan.
Kelestarian spesies badak jawa ini terletak pada kemampuan populasi yang
ada pada saat ini untuk mempertahankan jumlahnya (stable population) dan
meningkatkan jumlahnya (viable population). Data terkini menunjukkan bahwa
populasi badak jawa di Taman Nasional Ujung Kulon berada pada jumlah yang stabil
serta tidak menunjukkan perkembang biakkan yang signifikan. Pertumbuhan populasi
secara signifikan ditetapkan pada angka pertumbuhan minimal 3% setiap tahun
sebagaimana dimandatkan dalam strategi konservasi badak Indonesia (PHKA 2007).

2
Sebagai populasi tertutup dimana tidak terjadi proses migrasi keluar atau
masuk ke dalam Taman Nasional Ujung Kulon, pertumbuhan populasi badak jawa
dapat dicapai dengan dua cara yaitu: meredam tingkat mortalitas dan/atau
meningkatkan reproduksi yang ada pada populasi saat ini.
Permasalahan
Dua hal yang besar pengaruhnya terhadap peluang reproduksi dan juga risiko
mortalitas adalah aspek asupan nutrisi dan tingkat cekaman. Sejak awal tahun 70an
tidak ada lagi terjadi kasus perburuan badak jawa, oleh karena itu slah satu ancaman
terbesar yang dapat menyebabkan kematian bagi badak jawa muncul dari risiko
penyakit dan perubahan pada komposisi ekosistem baik yang berupa suksesi alami
maupun perubahan yang terjadi sebagai dampak dari aktifitas manusia seperti
perubahan iklim. Dari sudut pandang epidemiologi, nutrisi dan cekaman merupakan
dua aspek yang ada dalam lingkungan / habitat badak yang berperan penting dalam
menentukan kemampuan badak untuk menurunkan risiko kematian dengan cara
mengatasi penyakit berupa agen infeksius dan non-infeksius.
Dalam dunia medis, agen infeksius didefinisikan sebagai penyebab penyakit
yang dapat berpindah atau ditularkan dari satu individu kepada individu lainnya baik
secara langsung maupun melalui perantara (vektor). Contoh-contoh agen infeksius
terdiri dari: virus, bakteri, dan parasit yang dapat membahayakan kehidupan satwa liar
dan juga manusia (Daszak 2000). Sebaliknya, agen non-infeksius didefinisikan
sebagai penyebab penyakit yang tidak dapat berpindah atau menular dari satu individu
ke individu lainnya seperti toksin (racun) yang menyebabkan penyakit akibat
konsumsi ataupun kontaminasi bahan toksin yang berasal dari lingkungan.
Oleh karena itu, fokus kajian ini dititik beratkan pada upaya untuk
mengidentifikasi status kualitas nutrisi dan profil cekaman serta dampaknya terhadap
individu badak agar upaya untuk menurunkan tingkat cekaman terhadap badak jawa
dapat dirancang. Harapannya spesies ini akan memiliki peluang yang lebih besar
untuk mampu bertahan terhadap penyakit serta peluang yang lebih besar pula untuk
meningkatkan kapasitas reproduksinya.

3
Pendekatan Konservasi Melalui Aspek Medis
Teknik medis konservasi mulai diperlukan dengan mempertimbangkan adanya
berbagai ancaman yang dapat mempengaruhi kesehatan dan kelangsungan hidup
badak jawa. Secara kualitatif, mortalitas akibat gangguan kesehatan badak jawa
pernah terjadi pada lima ekor badak di tahun 1982 (WWF-IUCN 1982), dan satu ekor
badak di tahun 2003. Semua kasus kematian diawali dengan gejala klinis kolik yang
mengindikasikan adanya gangguan pada sistem pencernaan. Beberapa kemungkinan
gangguan kesehatan dapat muncul karena adanya infeksi silang antar satwa liar
(banteng dan badak menggunakan areal yang sama di semenanjung Taman Nasional
Ujung Kulon), dan infeksi dari hewan ternak yang lepas dan masuk ke dalam
kawasan. Selain ancaman dari agen infeksius, ancaman dari sumber non-infeksius
seperti: asupan metabolit sekunder dari sumber makanan dan bahan-bahan yang
bersifat toksik juga merupakan aspek yang harus dicermati dalam pengelolaan
populasi.
Habitat di Ujung Kulon secara perlahan namun pasti mengalami perubahan
akibat berbagai kejadian seperti: invasi tumbuhan langkap Arenga obtusifolia (Putro
1997) dan perubahan iklim. Kedua faktor di atas menyebabkan berkurangnya
ketersediaan tumbuhan pakan yang dibutuhkan oleh badak, sementara perubahan
iklim ditengarai dapat mengakibatkan perubahan pada iklim mikro (Suprayogi et al.
2006), keragaman jenis tumbuhan (Huxman & Scott 2007), serta perubahan pada
mekanisme penyebaran penyakit (Harvell et al. 2002). Suksesi vegetasi alami dapat
mendorong terjadinya dominasi tumbuhan tertentu dan dapat mengakibatkan
keterbatasan pakan. Beberapa jenis tumbuhan yang mendominasi struktur vegetasi
tersebut mengandung antinutrisi/toksin seperti pada jenis Lantana cammara yang
mengandung zat yang dikenal dengan nama: lantaden (Sharma et al. 2000). Zat
lantaden ini merupakan suatu senyawa yang diproduksi secara alami oleh tumbuhan
Lantana cammara dan bersifat racun hepatotoksin, atau senyawa racun yang dapat
mengganggu sistem pencernaan (McSweeney & Pass 1983; Sagar et al. 2005), dan
merusak hati (liver) berbagai hewan ternak (Pass et al. 1979). Selain keterbatasan
pakan, ketersediaan air merupakan faktor lain yang dapat mempengaruhi kualitas
hidup dan kesehatan badak jawa. Aspek pakan dan air tersebut di atas berpotensi
mengakibatkan stres akut (yang terjadi dalam periode waktu singkat) maupun kronik
(yang terjadi dalam waktu yang relatif panjang atau) karena badak sepenuhnya
bergantung pada sumber tumbuhan pakan dan air yang terbatas di habitat alaminya.

4
Sampai saat ini belum ada metode aplikatif yang dapat digunakan untuk
memantau kualitas asupan nutrisi dan tingkat cekaman yang dihadapi olah badak
jawa. Tingkat kesulitan untuk menemukan badak di habitatnya dan tidak adanya
badak jawa di penangkaran mengharuskan peneliti untuk mengembangkan metode
pengamatan dan pemantauan dengan menggunakan sampel feses badak yang relatif
lebih mudah didapat. Tantangan berikutnya ada pada proses untuk menggali
informasi serta memperoleh parameter kesehatan dari feses badak tersebut sebagai
indikator yang akurat dan sensitif untuk mencerminkan kesehatan secara umum dan
kondisi pencernaan secara khusus. Indikator ini dapat digunakan sebagai baseline,
namun informasi mengenai fluktuasi dan ambang batas indikator-indikator terkait
masih diperlukan sebagai data pelengkap untuk menjadikan metode ini lebih relevan
dan aplikatif sebagai perangkat pemantauan kesehatan badak jawa. Berdasarkan
informasi baseline inilah suatu model pengelolaan populasi badak disusun dengan
tujuan meningkatkan peluang untuk dapat bertahan terhadap penyakit dengan upaya
untuk mengurangi risiko mortalitas serta meningkatkan peluang reproduksi. Selain
menjadi bagian dalam upaya pengelolaan populasi badak jawa di Taman Nasional
Ujung Kulon, perangkat pemantauan kesehatan seperti ini merupakan bagian dari
metoda pemantauan populasi yang juga merupakan suatu kebutuhan pengelolaan
badak jawa terkait pembentukan populasi ke-2 dimana beberapa ekor badak akan
dipindahkan dari habitat yang ada di Taman Nasional Ujung Kulon ke lokasi lain.
Melalui upaya pemindahan badak ke habitat baru, perangkat untuk mengukur tingkat
cekaman, status/profil nutrisi, dan analisis risiko kesehatan pada badak saat adaptasi,
dalam proses, dan pasca pemindahan merupakan suatu keharusan untuk menjamin
kelangsungan hidup populasi badak di habitat barunya.
Tujuan Penelitian
Penelitian ini bertujuan untuk membangun suatu model pengelolaan populasi
badak jawa dengan memperhatikan aspek nutrisi dan cekaman yang dapat
mempengaruhi status kesehatan serta daya tahan spesies ini terhadap penyakit. Untuk
mencapai tujuan tersebut, penelitian ini memerlukan analisis mengenai:

5
1. Pola pergerakan dan lintasan badak di habitatnya (trajektori) sebagai basis dari
pengumpulan informasi daya dukung alam Taman Nasional Ujung Kulon.
2. Palatabilitas, kuantitas, kualitas pakan, kecernaan, risiko toksin lantaden, dan
ketersediaan garam bagi badak jawa jantan di kawasan Taman Nasional Ujung
Kulon.
3. Profil hormon glukokortikoid akibat cekaman defisit nutiren dan ketersediaan air
dengan menggunakan 5-beta-adiol sebagai indikatornya.
4. Potensi penggunaan hewan model (kuda) sebagai pembanding bagi badak jawa
dalam penelitian cekaman akibat defisit pakan dan air.
5. Penyusunan rekomendasi model pengelolaan badak jawa berbasis nutrisi dan
ketersediaan air di kawasan Taman Nasional Ujung Kulon.
Hipotesis Penelitian
1. Kondisi habitat di Taman Nasional Ujung Kulon merupakan daerah dengan daya
dukung (ketersediaan pakan dan air) yang kurang memadai bagi badak jawa.
Kondisi ini mengakibatkan meningkatnya cekaman pada badak jawa.
2. Metabolit hormon asal feses dapat digunakan sebagai indikator status dan profil
glukokortikoid akibat cekaman, mengingat glukokortikoid merupakan indikator
aktifitas korteks adrenal yang disebabkan oleh adanya cekaman.
3. Kuda dapat digunakan sebagai hewan model pembanding yang baik untuk
penelitian terkait cekaman akibat keterbatasan pakan dan air yang mempengaruhi
badak jawa, mengingat kuda memiliki kesamaan genetika, anatomi saluran cerna,
dan perilaku dengan badak.
Novelty / Kebaruan
Kebaruan yang dihasilkan dalam penelitian ini adalah model yang merupakan
bagian dari solusi bagi masalah reproduksi dan mortalitas yang sampai saat ini belum
terpecahkan. Pendekatan konservasi yang mengacu kepada upaya mitigasi cekaman
bagi spesies badak jawa baru kali ini dilakukan dengan harapan untuk meningkatkan
kapasitas reproduksi serta menekan risiko kematian pada badak jawa. Selain itu,
penelitian ini juga menghasilkan suatu mekanisme penelitian badak jawa melalui
hewan model sehingga berbagai penelitian badak jawa dapat dilakukan pada hewan

6
model tertentu untuk menghindari risiko dan kesulitan yang timbul bila menggunakan
badak jawa secara langsung.
Produk nyata yang merupakan novelty atau kebaruan hasil penelitian ini
adalah penerapan hasil-hasil penelitian ini untuk kepentingan pengelolaan badak jawa
di habitat alaminya. Langkah-langkah penerapan hasil dari penelitian ini terdiri dari:
1. Penggunaan hewan model untuk mensimulasi kondisi habitat serta
mempelajari respons yang mungkin terjadi pada badak jawa di habitat
alaminya.
2. Rekomendasi pengelolaan vegetasi potensial untuk kehidupan badak jawa.
3. Penggunaan profil glukokortikoid sebagai indikator cekaman pada badak.
Alur Penelitian
Tahap pertama dalam penelitian ini menguatkan dugaan adanya korelasi
antara ruang jelajah dan kualitas nutrisi di habitat badak di Taman Nasional Ujung
Kulon dan juga identifikasi cekaman yang berasal dari kuantitas, kualitas, dan
kecernaan nutrien dalam tumbuhan pakan; tahap ini dilanjutkan dengan menguatkan
dugaan adanya cekaman yang bersifat individual (bervariasi antar individu yang
berbeda) ataupun yang bersifat musiman (bervariasi antar musim) akibat terbatasnya
ketersediaan air di musim kering. Informasi kualitatif yang didapat dari kajian ini
kemudian diterapkan dalam uji coba menggunakan hewan model untuk mendapatkan
informasi yang lebih kuantitatif dengan perlakuan yang jauh lebih terukur
dibandingkan dengan temuan dan data dari lapangan sehingga dampak dari cekaman
tersebut dapat dipelajari langsung keterkaitan atau korelasinya. Data dan informasi
dari setiap tahap dari penelitian ini kemudian digunakan untuk menyusun suatu model
pengelolaan badak jawa. Model ini terdiri dari rekomendasi prosedur pemantauan
cekaman dan juga rekomendasi untuk membangun suatu sistem pengelolaan habitat
untuk mitigasi cekaman yang terdiri dari pemantauan intensif untuk melihat status
cekaman dan dampak dari implementasi upaya mitigasi cekaman terhadap
perkembangbiakkan badak jawa secara alami. Pemantauan status cekaman dapat
dilakukan secara non-invasif melalui analisis sampel feses yang dilakukan secara
periodik. Secara teoretis, mitigasi cekaman pada populasi badak jawa akan dapat
meningkatkan peluang keberlangsungan hidup spesies ini (meningkatkan kapasitas
reproduksi dan menekan risiko mortalitas). Penelitian ini juga menjajagi

7
kemungkinan pengembangan metoda pemantauan kesehatan yang dapat dilakukan
secara non-invasif terkait tekanan yang dapat terjadi di habitat alami yaitu: akibat
keterbatasan pakan (kadar toksisitas dalam pakan) serta minimnya ketersediaan air,
maka penelitian ini memiliki kerangka acuan sebagaimana ditampilkan pada Gambar
1 di bawah ini.
Gambar 1. Skema alur penelitian serta kontribusi yang dihasilkan terhadap
konservasi badak jawa. Garis putus mewakili aspek konservasi
(Perlindungan dan Pembuatan kubangan) yang tidak secara langsung
difokuskan dalam penelitian ini.
Metode Umum
Penelitian ini terdiri dari tiga tahapan yang dirancang untuk mempelajari
kondisi habitat badak serta profil asupan (nutrien, air, energi) dan toksin; mempelajari
profil cekaman dan dinamikanya antar individu badak dan antar musim; dan
menggunakan informasi dari kedua tahapan ini dalam tahap ketiga yaitu simulasi
cekaman di habitat badak terhadap hewan model untuk mengkuantifikasi reaksi
tanggap yang terjadi pada hewan model sebagai gambaran atas reaksi tanggap yang
mungkin terjadi pada badak jawa. Tiga ekor badak jawa berkelamin jantan dipilih
sebagai contoh dalam penelitian ini.

8
Tahap 1: Habitat Badak dan Profil Nutrisi
Tahap ini bertujuan untuk mempelajari profil asupan pada badak jawa di
habitat aslinya di Taman Nasional Ujung Kulon sebagai bahan informasi yang
digunakan dalam simulasi/induksi cekapam pada tahap tiga. Penguatan analogi antara
badak dengan hewan model dilakukan dengan membandingkan hasil dari tahap ini
(kecernaan) dengan hasil kecernaan hewan model pada tahap tiga. Tahap ini terdiri
dari metoda:
1. Identifikasi ruang jelajah dan ruang pergerakan badak contoh untuk
mempelajari kondisi habitat (keragaman pakan, ketersediaan kubangan/air,
ketersediaan garam) terkait ruang jelajah dari setiap ekor badak.
2. Identifikasi pakan serta komposisi tumbuhan pakan yang di sukai
3. Identifikasi kualitas pakan serta penghitungan kualitas nutrisi tumbuhan
pakan
4. Penghitungan tingkat kecernaan dengan metode AIA
5. Analisis deskriptif untuk mengidentifikasi perbedaan profil asupan dan
kecernaan antar individu badak.
Tahap 2: Habitat Badak dan Profil Cekaman
Tahap ini bertujuan untuk mempelajari profil cekaman yang terjadi pada
badak jawa di Taman Nasional Ujung Kulon. Perbedaan profil cekaman diamati antar
individu dan juga antar musim yang berbeda (musim kering dan penghujan).
Identifikasi sumber cekaman dalam tahap ini digunakan dalam simulasi/induksi
cekaman pada Tahap 3. Informasi mengenai profil cekaman ini dibandingkan dengan
hasil induksi cekaman dalam Tahap 3 sebagai bagian dari penguatan analogi hewan
model dan badak. Tahap ini terdiri dari metode:
1. Pengumpulan sampel feses dari tiga individu badak contoh pada musim
kering dan penghujan.
2. Pemilihan asai hormon yang memadai untuk digunakan pada sampel feses
sebagai cara mengidentifikasi tingkat cekaman. Pemilihan asai hormon
terdiri dari: pemilihan kit hormon glukokortikoid/metabolitnya, serta
validasi dari hormon/metabolit tersebut.
3. Asai hormon asal feses untuk mempelajari perbedaan kandungan hormon
glukokortikoid/metabolitnya serta profil dan dinamikanya.

9
4. Analisisi deskriptif untuk mempelajari profil cekaman antar individu dan
trend keterkaitan antara profil cekaman dengan musim.
Tahap 3: Simulasi Cekaman pada Hewan Model
Tahap ketiga ini bertujuan untuk memilih hewan model yang sesuai untuk
badak jawa dan mensimulasikan cekaman yang teridentifikasi pada Tahap 1 dan 2
serta mengukur reaksi tanggap yang terjadi pada hewan model tersebut. Penguatan
analogi hewan model dengan badak jawa dilakukan dengan membandingkan hasil
data tahap tiga ini dengan hasil dari tahapan-tahapan sebelumnya. Tahap tiga terdiri
dari metode:
1. Pemilihan hewan model berdasarkan kaidah hewan model serta
kesamaan/kedekatan genetika, anatomi, dan perilaku.
2. Simulasi dan induksi cekaman yang terjadi di habitat asli badak jawa di
Taman Nasional Ujung Kulon (hasil dari tahap satu dan tahap dua)
terhadap hewan model.
3. Kuantifikasi reaksi tanggap terhadap induksi cekaman berdasarakan
parameter perilaku dan parameter fisiologis (respirasi, kadar hormon
kortisol, hematologi, respirasi, dan kecernaan).
4. Penguatan analogi antara hewan model dengan badak melalui
perbandingan reaksi tanggap yang terjadi pada hewan model dan reaksi
pada badak berdasarakan pengamatan dan literatur.
Hasil dari ketiga tahapan di atas digunakan dalam merumuskan sebuah model
pengelolaan yang dapat diusulkan pada otoritas pengelola populasi badak jawa untuk
melengkapi informasi yang diperlukan dalam menyusun strategi intervensi
konservasi. Pendekatan baru dari sisi medis konservasi dan hewan model akan
memberikan kontribusi dalam upaya meningkatkan peluang reproduksi sekaligus
menurunkan risiko mortalitas yang dapat terjadi pada populasi badak jawa. Dua hal
ini pada akhirnya akan memberikan kontribusi terhadap upaya peningkatan populasi
badak jawa sebesar 3% setiap tahun.

10
TINJAUAN PUSTAKA
Habitat Badak Jawa di Taman Nasional Ujung Kulon
Dari lima spesies badak di seluruh dunia, badak jawa merupakan satu-satunya
spesies yang ditemukan hanya di Indonesia, tepatnya di Taman Nasional Ujung Kulon
Banten. Pada tahun 1999 survei menyatakan keberadaan badak jawa di Vietnam
dengan jumlah delapan ekor (Polet et al. 1999), namun saat ini populasi tersebut
sudah dinyatakan punah (IRF-WWF 2011). Dengan punahnya populasi badak jawa di
Vietnam pada tahun 2011, maka keberlangsungan hidup spesies ini hanya bergantung
pada populasi yang ada di Ujung Kulon. Ujung Kulon merupakan kawasan lindung
yang ditetapkan sebagai Taman Nasional pada tahun 1990 dan diberi status sebagai
situs warisan dunia (World heritage site) oleh UNESCO di tahun 1992.
Taman Nasional Ujung Kulon memiliki luasan 120,000 hektar yang terdiri
dari kawasan darat dan laut, dan 30,000 hektar dari luasan ini merupakan
semenanjung yang dihuni oleh badak jawa. Habitat badak di semenanjung Ujung
Kulon ini sebagian besar merupakan dataran dengan tingkat kelerengan yang rendah
dan ditumbuhi berbagai jenis vegetasi mulai dari hutan bakau, hutan rawa pantai,
hutan pantai, hutan sekunder, dan hutan primer (Hommel 1987). Perbatasan antara
hutan sekunder dan hutan primer merupakan area yang sangat disukai oleh badak
karena area tersebut biasanya ditumbuhi oleh tumbuhan pakan badak.
Hommel (1987) juga menyebutkan bahwa tipologi tanah di semenanjung
Ujung Kulon ini terdiri dari jenis tanah litosol yang memiliki kemampuan untuk
menampung air dan menjadikan daerah ini banyak dipenuhi oleh genangan-genangan
air yang digunakan sebagai kubangan oleh banyak satwa, termasuk badak jawa.
Sebagai suatu kawasan lindung dengan status Taman Nasional, Ujung Kulon
merupakan tempat perlindungan satwa sekaligus tempat penelitian yang bersifat
ilmiah maupun edukasi. Kegiatan wisata yang bertanggung jawab (eko-wisata)
merupakan salah satu kegiatan yang diterapkan di Taman Nasional Ujung Kulon
untuk memberikan manfaat ekonomis maupun edukasi kepada masyarakat luas.
Untuk memastikan bahwa konservasi badak jawa dapat berjalan selaras dengan
penelitian dan wisata, maka pengelolaan Taman Nasional Ujung Kulon dibagi
menjadi tiga seksi konservasi dengan pembagian zonasi (zona inti, rimba,

11
pemanfaatan tradisional, dan zona khusus) dengan pengelolaan berbasis resort agar
dapat disesuaikan dengan kebutuhan di masing-masing seksi dan zona tersebut.
Informasi Umum tentang Badak Jawa
Badak jawa pertama kali dikaji secara ilmiah oleh Desmarest di tahun 1822
dan dikategorikan sebagai spesies Rhinoceros sondaicus (Corbett & Hill 1992) dan
merupakan merupakan salah satu spesies langka yang dikategorikan sebagai spesies
yang terancam punah. Menurut Lekagul & McNeely (1977) taksonomi badak Jawa
diklasifikasikan sebagai berikut:
Kerajaan : Animalia
Filum : Chordata
Sub-filum : Vertebrata
Super kelas : Gnathostomata
Kelas : Mamalia
Super Ordo : Mesaxonia
Ordo : Perissodactyla
Super famili : Rhinocerotidae
Famili : Rhinocerotidae
Genus : Rhinoceros Linnaeus, 1758
Spesies : Rhinoceros sondaicus Desmarest 1822
Hoogerwerf (1970) menyebutkan bahwa badak Jawa dewasa memiliki ukuran
tinggi (dari telapak hingga bahu): 169-175 cm dan panjang badan (dari moncong
hingga ujung ekor): 392 cm, dan berat tubuh pada kisaran 2.280 kg. Dibandingkan
dengan badak hitam afrika (Diceros bicornis), badak putih afrika (Ceratoterium
simum) dan badak india (Rhinoceros unicornis), badak jawa merupakan badak yang
tergolong berukuran kecil namun masih lebih besar bila dibandingkan dengan badak
sumatera (Dicerorhinus sumatrensis). Badak jawa memiliki tampilan sebagaimana
disajikan dalam Gambar 2 di bawah ini.

12
Gambar 2. Badak jawa jantan yang tertangkap oleh kamera otomatis (kamera jebak)
di Taman Nasional Ujung Kulon, Banten. Foto: WWF Indonesia – Balai
Taman Nasional Ujung Kulon (2003).
Di masa lampau badak jawa menghadapi tekanan berupa perburuan dan
gangguan langsung dari masyarakat karena waktu itu mereka dianggap sebagai
“hama” yang mengganggu lahan perkebunan masyarakat. Hal ini terjadi karena
badak jawa memiliki preferensi makanan yang merupakan tumbuhan semak dan
perdu yang banyak ditemukan di lahan pertanian masyarakat. Badak jawa yang
dahulu tersebar di pulau Jawa, Sumatera, bahkan sampai ke Indocina populasinya
makin terdesak dan badak jawa terakhir di pulau Sumatera ditembak mati di
Palembang sekitar tahun 1920an dan badak jawa terakhir di luar Ujung Kulon
ditembak di daerah Garut pada tahun 1930an. Sejak itu, badak jawa hanya terdapat di
Taman Nasional Ujung Kulon, Banten dan di Taman Nasional Cat Tien, Vietnam.
Namun, populasi badak jawa di Vietnam telah dinyatakan punah pada pertengahan
tahun 2010 sehingga nasib keberlangsungan spesies ini hanya ada pada populasi di
Indonesia. Walaupun perburuan sudah tidak ada lagi, saat ini populasi badak Jawa
masih menghadapi tantangan yang dapat membahayakan kehidupan mereka.
Sebagaimana dihadapi oleh berbagai spesies badak di seluruh dunia, badak
Jawa juga menghadapi risiko infeksi penyakit dan/atau gangguan kesehatan baik yang
diakibatkan oleh agen infeksius maupun non-infeksius. Fisiologi dan kesehatan pada
spesies badak ini adalah aspek yang belum banyak dipelajari sampai saat ini.
Walaupun telah ada beberapa individu badak yang pernah dipindahkan dari alam dan
ditempatkan di kebun binatang sekitar tahun 1800an (Newton 1874; Rookmaaker
1982; Rookmaaker 2005, Reynolds 1961), belum pernah ada catatan mengenai

13
kerentanan satwa ini terhadap cekaman dan risiko kematian akibat tekanan dan/atau
proses pemindahannya dari habitat alami. Ilmu biomedis hewan diharapkan dapat
digunakan sebagai acuan untuk mempelajari aspek fisiologis, kesehatan, dan juga
kemungkinan perlakuan untuk mencegah gangguan kesehatan pada spesies langka ini.
Penelitian dititik beratkan pada kajian tingkat stres, toksisitas, dan analisis risiko
akibat asupan nutrisi dan konsumsi jenis pakan alami tertentu bagi badak jawa yang
tersedia di habitatnya. Sebagai hewan yang hidup liar, badak jawa sangat bergantung
pada ketersediaan pakan di habitatnya, oleh karena itu, disamping perburuan, faktor
lingkungan seperti perubahan iklim dan ketersediaan pakan merupakan penyebab
kepunahan satwa ini dari berbagai habitat historisnya seperti di Borneo (Cranbrook &
Piper 2007), Kamboja (Poole & Duckworth 2005), Malaysia (Kloss 1927), dan juga
di Pulau Jawa (Scheltema 1912; Walcott 1914).
Sejak tahun 60an pemantauan badak sudah banyak dilakukan oleh beberapa
pihak dengan menggunakan metode yang sederhana seperti penghitungan jejak,
penggunaan kamera dan video otomatis, sampai metode yang lebih rumit yaitu telaah
genetika, jenis, dan komposisi pakan dari feses. Identifikasi individu badak dengan
kamera otomatis juga memungkinkan penghitungan estimasi jumlah individu dengan
metode mark-recapture, analisis komposisi populasi, nisbah kelamin, dan juga
perhitungan untuk menduga pertumbuhan populasi dengan adanya kelahiran individu-
individu baru (Hariyadi et al. 2008). Secara holistik, interaksi badak dengan
komponen lainnya dalam ekosistem juga telah dipelajari. Berdasarkan salah satu
kajian yang pernah dilakukan diperoleh data bahwa ada persaingan ruang antara
banteng (Bos javanicus) dan badak yang mengakibatkan keterbatasan akses badak ke
daerah-daerah tertentu (YMR 2002). Kajian palatabilitas tumbuhan pakan yang
dilakukan oleh berbagai peneliti yang menginventarisir sekitar 94 jenis tumbuhan
yang menjadi konsumsi harian badak jawa, namun kajian lain menunjukkan bahwa
pertumbuhan dan dominasi sejenis palma (Arenga obtusifolia) mengancam
ketersediaan jenis-jenis tumbuhan pakan yang dibutuhkan oleh badak tersebut (Putro
1997; YMR 2004). Jika antara hasil pengamatan dan penelitian terdahulu
dibandingkan dengan keberhasilan badak jawa untuk bertahan hidup sampai saat ini,
badak jawa dikenal sebagai satwa yang tangguh dan dapat mempertahankan
kelangsungan hidupnya dalam jumlah populasi yang sangat sedikit. Diet utama yang
terdiri dari berbagai jenis tumbuhan semak memberi nutrisi yang memadai untuk
bertahan dari berbagai tekanan, termasuk juga tekanan dari penyakit. Berdasarkan

14
penggalian informasi dari masyarakat di sekitar zona penyangga Taman Nasional
Ujung Kulon, berbagai jenis tumbuhan pakan badak telah dimanfaatkan oleh
masyarakat sebagai sumber obat tradisional yang memiliki khasiat sebagai anti
inflamasi, antipiretika, antiseptik, dan juga sebagai obat untuk memperlancar proses
persalinan.
Jenis-jenis Cekaman
Berdasarkan rentang waktu pemaparannya, cekaman dapat dikategorikan ke
dalam dua jenis yaitu: cekaman akut yang terjadi dalam waktu yang relatif singkat
dalam hitungan detik sampai sekitar 60 menit (Figueiredo et al. 2003) dan cekaman
kronis yang berulang setiap hari selama 5 hari (Melia & Duman 1991) sampai waktu
yang lebih lama (mingguan, bulanan, tahunan) seperti yang dialami oleh badak di
kebun binatang yang mengalami cekaman kronis akibat pengandangan dan kunjungan
wisatawan (Carlstead & Brown 2005). Kedua jenis cekaman ini memberikan respons
berupa peningkatan aktifitas adrenal korteks yang ditunjukkan dengan adanya
peningkatak sekeresi hormon glukokortikoid (Barja et al. 2007) pada berbagai hewan
termasuk badak (Turner Jr et al. 2002; Menargues et al. 2008). Selain kadar hormon
glukokortikoid, cekaman akut dan kronis menimbulkan reaksi yang berbeda pada
metabolisme dan sistem pertahanan tubuh hewan. Selain profil hormon dari kelas
glukokortikoid, Davis et al. (2008) menyatakan bahwa profil hematologi khususnya
netrofil dan limfosit merupakan indikator akan adanya cekaman pada hewan-hewan
vertebrata.
Cekaman Akut
Cekaman akut menimbulkan respons yang spesifik berupa aktifitas pada aksis
HPA (Hipotalamus-pituitari-adrenal korteks) yang berujung pada sekresi hormon
glukokortikoid (Figuireido et al. 2003). Cekaman akut juga memicu sistem kekebalan
tubuh untuk bekerja lebih baik dengan cara mendistribusikan lekosit dari darah ke
kulit seperti yang ditunjukkan Dhabhar (2000). Mekanisme ini merupakan kerja
hormon cekaman serupa dengan mekanisme bertahan atau lari (fight or flight) yang
terjadi saat terjadi cekaman akut dan berpotensi untuk meningkatkan daya tahan tubuh
terhadap penyakit, terutama yang masuk melalui jaringan perifer seperti kulit. Selain
distribusi lekosit, reaksi sistem pertahanan tubuh terhadap cekaman akut juga
termasuk distribusi sel Natural Killer (NK) yang senyawa kimianya berfungsi sebagai

15
pembunuh bagi agen-agen asing yang masuk ke dalam tubuh. Sel-sel NK ini
berkurang di dalam darah dan paru-paru, namun jumlahnya tidak berkurang di limpa
(Kanemi et al. 2005).
Cekaman Kronis
Dhabhar (2000) menunjukkan bahwa cekaman kronis memberikan respons
berupa pengurangan distribusi lekosit dari kulit kembali ke dalam darah, hal ini
merupakan kebalikan dari respons yang ditunjukkan oleh adanya cekaman akut.
Figuireido et al. (2003) menunjukkan bahwa cekaman kronis juga menghasilkan
respons berupa sensitisasi aksis HPA terhadap sumber cekaman (biasanya terjadi pada
cekaman yang sama dan berulang). Sensitisasi seperti ini merupakan contoh bahwa
cekaman kronis dapat memicu dan mempertahankan perubahan pada beberapa proses
biokimia yang berujung pada implikasi klinis (Melia & Duman 1991). Perubahan
parameter fungsi-fungsi hormonal, sistem pertahanan tubuh, metabolisme, dan sistem
kardio vaskular akibat adanya cekaman kronis dapat dirumuskan menjadi sebuah
indeks yang dikenal dengan indeks beban allostatic yang dapat menunjukkan
kemungkinan cekaman kronis tersebut mendorong terjadinya penyakit (Juster et al.
2009).
Penyakit-penyakit pada Badak
Selain suspect antraks pada kasus kematian badak jawa di tahun 1980an,
informasi mengenai agen infeksius yang mengancam kehidupan badak jawa belum
dapat diketahui dengan pasti. Salah satu telaah yang pernah dilakukan pada badak
jawa adalah telaah endoparasit yang menemukan berbagai parasit cacing
(Strongyloides spp, Bunostomum spp, Trichostrongylus spp, Fasciola spp,
Schistosoma spp) dan protozoa (Balantidium spp, Entamoeba spp, Cryptosporidium
spp, Cycloposthium spp) dalam feses badak (Tiuria et al. 2006). Sebagai
pembanding, beberapa penyakit yang mungkin menyerang badak di populasi
alaminya terdiri dari: penyakit darah (parasit protozoa Theileria sp dan Trypanosoma
sp), penyakit infeksius (bakteri, fungi, virus), penyakit parasitik (helminth), penyakit
reproduksi (brucellosis, vibriosis), luka, leptospirosis (Jessup et al. 1992), infeksi
Cowdria sp (Kock et al. 1992), dan defisiensi nutrisi dalam kasus
translokasi/pemindahan badak (Clausen 1981; Jonyo 2003).

16
Penyakit yang ditemukan pada badak dalam penangkaran/kebun binatang
lebih beragam dibandingkan dengan penyakit pada populasi alami dan ini
mengakibatkan kerugian material bagi pengelola kebun binatang serta kerugian
ekologis berupa berkurangnya jumlah badak akibat kematian. Hal ini disebabkan oleh
stress dan kondisi habitat buatan manusia yang berbeda dengan habitat alami badak.
Penyakit yang umum ditemukan pada badak dalam penangkaran/kebun binatang
antara lain: lingual abscess (Hatt et al. 2004), retak pada kuku, laminitis, chronic foot
disease (Jacobsen 2005), lesio pada mata, katarak (Sanborn 1908), aneurism pada
arteri koroner (Kock et al. 1991), hemolitik anemia (Jessup et al. 1992),
degenerasi/nekrosa dengan fibrosis pada purkinje myokardial (Kock 1996), ulcer pada
kornea (Gandolf et al. 1999), dermatitis eksudatif (Völlm et al. 2000), dan leukimia
limfoblastik akut (Paglia & Radcliffe 2000). Dari beberapa catatan yang ada, dapat
dilihat bahwa penyakit pada badak yang hidup liar berbeda dengan penyakit pada
badak di kebun binatang. Hal ini menunjukkan bahwa ada perbedaan pada agen
penyebab penyakit, lingkungan, dan juga kondisi fisiologis badak itu sendiri.
Dalam tiga dasawarsa terakhir, banyak muncul jenis-jenis penyakit baru yang
menginfeksi manusia maupun hewan termasuk satwa liar. Beberapa contoh dari
patogen baru yang muncul adalah: Virus Hendra, Virus Nipah, dan Virus West Nile
(Daszak et al. 2004). Bidang medis konservasi ini menjadi bagian penting dalam
proses antisipasi penyakit yang dapat muncul dan menginfeksi populasi satwa liar.
Infeksi penyakit pada satwa liar akan menambah rumit upaya konservasi beberapa
spesies, karena infeksi penyakit ini dapat menyebabkan kematian dan bahkan dapat
meningkatkan risiko kepunahan pada spesies-spesies tertentu. Berdasarkan informasi
yang disampaikan oleh Deem et al. (2001), Wildlife Conservation Society (WCS)
telah mengidentifikasi pentingnya bidang medis konservasi ini dan telah mempelopori
prosedur pemantauan kesehatan pada beberapa spesies satwa liar seperti: Llama
guanaco (Lama guanicoe) dan orangutan (Pongo pygmaeus).
Pengaruh Perubahan Iklim
Iklim pada skala mikro memberikan pengaruh pada fisiologi hewan seperti
dipelajari oleh Suprayogi et al. (2006) yang menunjukkan bahwa pola respirasi pada
kambing sangat dipengaruhi oleh tingkat kelembaban udara. Pada skala global,
perubahan iklim akan menyebabkan perubahan pada dinamika vegetasi akibat

17
meningkatnya kadar CO2 di atmosfir dan juga berubahnya keseimbangan air di dalam
tanah (Huxman & Scott 2007). Korelasi antara perubahan iklim dan dinamika
vegetasi ditunjukkan pula pada studi yang dilakukan oleh Williams et al. (2002) yang
mempelajari perubahan vegetasi dan kondisi atmosferik pada zaman es. Perubahan
struktur vegetasi ini dapat menimbulkan berbagai konsekuensi bagi badak jawa, satwa
herbivora yang menggantungkan sumber pakan sepenuhnya dari vegetasi yang ada.
Perubahan iklim menuju kekeringan seperti ini dapat mengakibatkan perubahan pada
jenis-jenis tumbuhan dominan yang kemudian berpotensi pula untuk mengancam
keberadaan jenis tumbuhan yang dibutuhkan oleh badak. Apabila jenis tumbuhan
pakan badak berkurang akibat perubahan iklim seperti ini, maka defisit pakan akan
dialami oleh badak sebagai konsekuensinya. Salah satu contoh kepunahan
megaherbivora akibat perubahan iklim adalah kepunahan mammoth di Pulau Wrangle
Siberia akibat hilangnya padang rumput serta berbagai tumbuhan yang menjadi bahan
makanan mammoth di lokasi tersebut (Martin & Stuart, 1995).
Pemodelan yang dilakukan oleh Permadi (2008) menunjukkan bahwa
perubahan iklim berpotensi untuk merubah semenanjung Ujung Kulon menjadi
daerah yang lebih kering dengan ketersediaan air yang terbatas. Pemodelan ini
menunjukkan bahwa ada risiko kekeringan yang dapat melanda habitat badak di
semenanjung Taman Nasional Ujung Kulon akibat perubahan iklim. Risiko
kekeringan merupakan hal yang perlu diperhatikan mengingat bahwa Cameron &
Perdue (2005) menunjukkan cekaman kronik yang terjadi pada hewan coba dapat
timbul akibat kekeringan atau kelangkaan air (water avoidance stress), dengan
demikian perlu dikaji lebih lanjut apakah ketersediaan air yang lebih sedikit pada
musim kemarau akan memberikan suatu tekanan pada badak jawa.
Tanah dan lumpur juga merupakan komponen penting di dalam ekosistem
habitat badak karena dari komponen ini badak kerap memperoleh mineral untuk
memenuhi kebutuhan metabolisme sehari-hari. Seperti halnya ingesti pakan dan air,
badak jawa memiliki kebiasaan untuk “memakan” lumpur (salt licking) sebelum
dan/atau setelah berkubang sebagaimana yang diamati pada beberapa sesi pemantauan
perilaku yang dilakukan di Taman Nasional Ujung Kulon (Data Taman Nasional
Ujung Kulon & WWF 2010). Kualitas mineral serta kontaminasi yang ada pada
tanah/lumpur juga berpotensi untuk memberikan dampak terhadap mekanisme peny
rapan nutrien pada badak jawa.

18
Mekanisme fisiologis terhadap cekaman berupa kekeringan
Ketersediaan air dan aktifitas berkubang pada badak jawa (Rhinoceros
sondaicus) merupakan beberapa syarat penting untuk keberlangsungan hidup badak
jawa di habitat alaminya di semenanjung Taman Nasional Ujung Kulon. Banyak
peneliti badak terdahulu (Schenkel, Hoogerwerf, Van Strien, Sajudin, dan Setiawan)
mencatat bahwa badak berkubang lebih dari satu kali dalam sehari dan ditambah
dengan aktifitas berendam di sungai maupun di laut (pesisir pantai).
Termoregulasi
Aktifitas berkubang ini erat kaitannya dengan upaya termoregulasi yang
dilakukan badak untuk menjaga keseimbangan (homeostasis) tubuh mereka.
Termoregulasi merupakan suatu proses yang terkait dengan perilaku hewan untuk
berusaha mempertahankan suhu tubuh yang optimal. Suhu yang terlalu tinggi akibat
proses metabolisme ataupun suhu tubuh yang terlalu rendah akan mengkibatkan
ketimpangan dalam berbagai proses fisiologi tubuh hewan (fungsi dan kinerja
berbagai protein) yang akhirnya mendorong kondisi yang tidak seimbang dalam
fisiologi hewan tersebut. Badak memiliki kulit berlipat dengan lapisan subkutan yang
mengandung banyak pembuluh darah. Lipatan kulit dan pembuluh darah tersebut
berfungsi dalam proses termoregulasi pada badak (Endo et al. 2009).
Kondisi kulit
Kelembaban kulit pada badak menjamin kecukupan kadar air yang diperlukan
untuk menjaga kulit agar dapat mempertahankan struktur dan berfungsi secara
optimal. Struktur kulit badak (badak afrika) disajikan pada Gambar 3, sementara
gambaran kulit yang tidak normal (mengalami lesio) disajikan pada Gambar 4.
Kekurangan kadar air pada kulit dapat menyebabkan terjadinya lesio berupa
retak/celah pada kulit akibat berkurangnya kelenturan kulit saat kelembaban tidak
mencukupi. Baron (2006) menjelaskan bahwa lesio ini memberikan stimulasi pada
reseptor syaraf pada permukaan kulit yang kemudian mengirimkan sinyal rasa nyeri
akibat terjadinya perubahan molekuler pada neuron yang mengakibatkan
meningkatnya sensitifitas neuron tersebut. Sensitifitas neuron ini kemudian
menghantarkan sinyal rasa nyeri ke bagian hipotalamus pada otak.

19
Gambar 3. Struktur kulit badak afrika dalam keadaan normal yang terdiri dari
lapisan keratin pada bagian terluar. Pada gambar ini terlihat lapisan
keratin mengelupas dan digantikan dengan lapisan yang baru. Sumber:
Munson et al. (1998).

20
Gambar 4. Histopatologi pada epidermis badak (badak afrika) yang menunjukkan
adanya degenerasi hidroskopik, dan pembentukan celah/retakan pada
kulit. Tidak ada inflamasi ditemukan pada lesio ini. Pewarnaan HE, bar
= 100 mikron. Sumber: Munson et al. (1998).
Kekurangan kadar air pada kulit mengakibatkan kondisi kulit kurang
fleksibel/elastis sebagai stimulus bagi sekumpulan sel-sel syaraf yang dikenal dengan
istilah neuron pada permukaan kulit yang berfungsi sebagai reseptor mekanis. Woolf
& Mannion (1999) menjelaskan bahwa stimulus seperti ini mengakibatkan buka tutup
kanal ion yang menyebabkan perubahan kadar ion Na++ dan Cl--pada lingkungan
ekstrasel dan intrasel karena adanya aliran ion masuk ataupun keluar. Lebih lanjut
Stuart et al. (1997) menjelasakan bahwa perbedaan ion seperti ini (depolarisasi)
mengakibatkan terjadinya perubahan resting potential (-70 mV) menjadi action
potential yang menjalar sepanjang neuron. Karena adanya insulasi myelin bagian
akson pada sel neuron, maka “lompatan” elektrik action potential ini terjadi pada
node of Ranvier yang tidak terlapisi oleh myelin. Node of Ranvier merupakan satu
bagian dari jaringan syaraf vertebrata berbentuk nodus (simpul) yang tidak terlindungi
oleh lapisan insulasi myelin sehingga memungkinkan bagi aliran ‘listrik’ ringan untuk
melompat dari satu nodus ke nodus berikutnya. Lompatan elektrik seperti ini
memungkinkan action potential untuk menjalar sepanjang serabut syaraf (axon)
menuju ujung neuron yang dikenal sebagai bagian pre synaptic. Bagian ini

21
mengandung rongga di dalam sel yang disebut vesikula berisi zat-zat kimia yang
dikenal dengan istilah neurotransmitter yang terbentuk dari senyawa asam amino
ataupun mono-amina. Sinapsis yang merupakan jembatan antar serabut syaraf dari sel
syaraf perifer seperti ini terdapat di bagian dorsal horn pada tulang belakang atau di
dalam sistem susunan syaraf pusat. Action potential yang mencapai bagian ini
kemudian memicu pergerakan vesikula ke arah membran sel, peleburan vesikula pada
membran yang mengakibatkan terlepasnya neurotransmitter ke celah synapse
(synaptic cleft) untuk mempengaruhi kerja neuron berikut (neuron post-synaptic).
Nyeri pada kulit akibat kekeringan berbeda dengan rasa nyeri akut yang
memberikan stimulus dalam intensitas tinggi (seperti tusukan dan panas) yang
ditanggapi oleh neuron A-delta, nyeri pada kulit akibat kekeringan terjadi pada neuron
tipe C dalam intensitas rendah yang mengakibatkan perasaan tidak nyaman pada
badak, namun tidak cukup kuat untuk memicu refleks otot. Proses komunikasi antar
neuron pada synapse tidak menimbulkan reaksi adrenergik yang mengakibatkan
kontraksi otot, tapi menghantarkan signal nyeri dari saraf perifer ke susunan syaraf
pusat dengan bantuan neurotransmitter (Woolf & Mannion 1999). Neurotransmitter
asetilkolin pada neuron post-synapse melekat pada reseptor asetilkolin yang ada pada
membran sel. Melekatnya asetilkolin menyebabkan terbukanya kanal ion Na++ yang
mengakibatkan depolarisasi pada neuron pada susunan syaraf pusat yang
mengakibatkan terjadinya action potential yang menjalar pada susunan syaraf pusat
menuju sistem korteks pada otak (untuk persepsi) dan berlanjut ke sistem limbik
(untuk respons).
Pada sistem korteks, impulse dari syaraf pusat yang menghantarkan sinyal
nyeri dari syaraf perifer bekerja pada bagian sebelum sambungan (pre-synapse)
dengan mengeluarkan neurotransmitter (serotonin) yang berfungsi untuk merekam
(proses memori) rasa ketidaknyamanan akibat kekeringan yang terjadi pada bagian
kulit. Dalam kondisi seperti ini, bila badak menemukan kubangan dan/atau sumber
air untuk melembabkan kulitnya maka stimulus pada reseptor mekanis pada kulit akan
berkurang. Sel reseptor akan kembali ke resting potential dan berhenti mengirimkan
sinyal nyeri/ketidaknyamanan ke otak. Di otak, serotonin akan mempelajari asosiasi
antara kegiatan berkubang atau berendam dengan hilangnya rasa ketidaknyamanan.
Hal ini kemudian membentuk pola perilaku terkait dengan pengurangan rasa nyeri
dan ketidaknyamanan pada kulit dengan berkubang dan berendam. Selain serotonin,
dalam proses interaksi yang rumit neurotransmitter dopamin bekerja dan

22
menimbulkan rasa nyaman sebagai bagian dari “imbalan” kegiatan berkubang dan
berendam. Perilaku berkubang merupakan suatu dampak reflektif dari adanya nyeri
(Bennet 1999) dan merupakan suatu tindakan mempertahankan diri (self preservation)
yang difasilitasi oleh neurotransmitter dopamin dan serotonin pada otak.
Dinamika yang terjadi pada ekosistem habitat badak tidak selalu
memungkinkan badak untuk dapat segera menghilangkan rasa ketidaknyamanan
akibat kekeringan pada kulit. Selama musim kemarau curah hujan jauh berkurang
dan menyebabkan berkurangnya sumber air untuk berkubang dan berendam. Dengan
berkurangnya sumber air seperti ini, kemungkinan besar badak harus bertahan lebih
lama dengan kondisi tidak nyaman akibat kekeringan pada kulit untuk waktu yang
relatif lebih lama dibanding pada musim hujan. Rasa tidak nyaman ditambah dengan
intensitas kerusakan pada kulit yang makin tinggi menyebabkan impuls nyeri juga
makin meningkat intensitasnya disertai meningkatnya durasi dari stimulasi yang
terjadi pada reseptor nyeri pada kulit. Kondisi seperti ini menyebabkan otak untuk
bekerja menanggapi kondisi yang terjadi dengan aktifasi sistem noradrenergik pada
otak yang memicu sekresi hormon norepinefrin (Morilak et al. 2005).
Berbeda dengan penghantaran rasa nyeri dari reseptor pada kulit melalui
syaraf perifer dan syaraf pusat menuju ke otak yang terjadi dalam waktu yang cepat
(hitungan detik), maka dalam menanggapi intensitas serta durasi ketidaknyamanan
kekeringan pada kulit stimulasi yang terjadi melewati suatu ambang batas yang
kemudian memicu sel-sel neuron pada otak untuk mengeluarkan suatu respons yang
berjalan lambat (dalam hitungan jam atau hari) melalui perantara peptida berupa
hormon. Dalam kondisi seperti ini sel-sel otak terpicu untuk memproduksi hormon
ACTH (Adrenocortocotrophic Hormone). Serotonin yang dikeluarkan oleh neuron
pada sistem korteks menginduksi sekresi ACTH dengan melekat pada reseptor
serotonergik pada membran sel (Bruni et al. 1982) serta mendorong aktifasi caraka
Inositol triosfosfat (IP3) yang dihasilkan dari hidrolisis fosfatidil inositol pada
membran sel. Aktifasi caraka IP3 mengakibatkan mobilisasi ion Ca++ dari retikulum
endoplasmik serta badan golgi sebagai depo ion Ca di dalam sel yang bila digabung
dengan kalmodulin akan mengaktifasi berbagai protein kinase yang akhirnya
mengaktifasi berbagai fungsi enzim dan protein dengan fosforilasi (Berridge 1984).
Kalmodulin dan ion Ca++ mendorong aktifasi kalcineurin, sebuah protein fosfatase
yang bergantung pada kalmodulin dan ion Ca++ dan mengatur aktifitas kanal ion,
neurotransmitter dan juga transkripsi hormon ACTH (Yakel et al. 1997). Sekresi

23
ACTH didorong juga dengan adanya hormon Corticotropin Releasing Factor (CRF)
yang melekat pada reseptor pada membran sel neuron pada bagian otak yang
memproduksi ACTH.
ACTH yang beredar di dalam darah mencapai target organnya yaitu kelenjar
adrenal yang berada di daerah ginjal. Sebagai kelenjar endokrin, adrenal memiliki
sel-sel yang bertugas untuk mentranskripsi hormon berupa protein adrenalin
(epinephrin) dan juga glukokortokoid kortisol yang terkait erat dengan kondisi
cekaman (stres). ACTH melekat pada reseptor pada membran sel adrenal dan
berfungsi sebagai agonis yang memicu hidrolisis fosfatidil inositol menjadi IP3 (Jolles
et al. 1980). IP3 kemudian memicu serangkaian reaksi yang terdiri dari: mobilisasi
ion Ca++ (dari retikulum endoplasmik dan golgi) yang selanjutnya bersama
calmodulin mengaktifasi protein calcineurin dengan fosforilasi yang berujung pada
transkripsi, translasi dan sekresi hormon-hormon cekaman dari sel-sel kelenjar
adrenal. Kortisol yang muncul akibat adanya cekaman, dan hormon tiroksin yang
senatiasa dibutuhkan dalam kehidupan sel bekerja secara sinergis dalam memberikan
keluaran berupa peningkatan metabolisme lipid (trigliserida) di hati untuk kemudian
digunakan dalam proses glukoneogenesis yang terfasilitasi oleh meningkatnya
aktifitas (ekspresi) berbagai enzim. Lipid yang terakumulasi kemudian mengalami
lipolisis dan terurai menjadi asam lemak sebagai bahan baku dalam proses
glukoneogenesis.
Pada hierarki terkecil keseimbangan dalam sistem tubuh badak jawa berjalan
pada tingkat seluler dengan terjaganya metabolisme sel untuk mempertahankan
berbagai fungsi sel sebagai bagian dari jaringan, sistem, dan tingkat individu. Kondisi
“lapar sel” merupakan kondisi dimana kebutuhan metabolisme sel tidak dapat
terpenuhi dalam periode waktu yang cukup lama dan melampaui kemampuan sel
untuk bertahan. Energi yang digunakan oleh sel untuk menjalankan fungsi normal
berasal dari ATP yang disintesis dengan bantuan senyawa berenergi tingi NADH hasil
reaksi kimia dalam siklus Kreb yang terjadi di mitokondria. Siklus Kreb memerlukan
asupan substrat berupa gula yang berasal dari karbohidrat, protein, maupun lemak
yang memasok reaksi-reaksi tertentu dalam siklus tersebut untuk memastikan siklus
berjalan dan menghasilkan NADH dan air sebagai produknya. Kondisi “lapar” pada
sel didefinisikan sebagai defisit berbagai senyawa yang dibutuhkan dalam
metabolisme sel (glukosa) dan juga defisit dari berbagai growth factor, caraka yang

24
diperlukan dalam proses signalling untuk mendorong berbagai fungsi dan respons
selular.
Penelitian yang dilakukan Soto-Gamboa et al. (2009) menunjukkan adanya
korelasi antara kadar ACTH dan kadar hormon kortisol di dalam darah. Selain itu,
penelitian yang sama juga menunjukkan adanya korelasi antara kadar hormon kortisol
dalam darah dengan kadar hormon kortisol beserta metabolitnya dalam feses. Wasser
et al. (2000) menunjukkan bahwa metabolit hormon kortisol yang dapat ditemukan
pada feses terdiri dari: kortisol, kortikosteron, 11-deoksikortisol, deoksikortikosteron,
kortison, prednisolon, progesteron, dan prednison. Penelitian yang sama
menunjukkan bahwa kortikosteron merupakan metabolit yang lazim dijumpai pada
berbagai jenis spesies.
Defisit Glukosa
Monosakarida glukosa merupakan nutrien penting dalam metabolisme karena
glukosa merupakan gula sederhana hasil dari pemecahan karbohidrat dalam
metabolisme hewan dan merupakan sumber utama produksi NADH yang digunakan
dalam sintesa ATP, kebutuhan energi di dalam sel. Penyerapan karbohidrat di dalam
usus halus terjadi berkat bantuan protein khusus SGLT1 yang berfungsi sebagai co-
transporter yang memerlukan ion Na+ dan glukosa untuk dapat menjadi pintu gerbang
terserapnya karbohidrat di dalam usus yang berada pada dinding usus (Dyer et al.
2002). Lebih lanjut Mace et al. (2007) menjelaskan bahwa glukosa masuk ke dalam
sel dari lingkungan ekstraseluler melalui protein transport khusus GLUT2 pada
membran sel yang berfungsi sebagai kanal masuknya molekul glukosa ke dalam sel
dengan bantuan Na. Tanpa adanya protein kanal ini, molekul glukosa yang tidak larut
dalam lemak akan mengalami kesulitan untuk menembus membran sel dan masuk ke
dalam lingkungan intrasel. Protein transpor ini bekerja secara sinergis dengan resptor
insulin yang juga berada pada membran sel untuk membantu penyerapan molekul
glukosa ke dalam sel. Hormon insulin sebagai caraka yang disekresikan oleh sel
insula (sel β pada pankreas) berfungsi sebagai pemicu terbukanya GLUT2 untuk
memfasilitasi masuknya glukosa ke dalam sel.
Pada sel paru glukosa merupakan substrat yang dapat mencegah kematian sel
dengan meningkatkan produksi ATP (Allen & White 1998). Sel yang mengalami
defisit glukosa ditandai dengan meningkatnya aktifitas dan produksi enzim Laktat
Dehidrogenase (LDH) di dalam sitosol seperti yang dinyatakan oleh Allen & White

25
(1998). Peningkatan aktifitas dan produksi enzim ini merupakan mekanisme internal
sel untuk meningkatkan laju konversi piruvat menjadi laktat (atau sebaliknya) dan
memastikan proses kimia dalam siklus Kreb dapat terus berjalan untuk memenuhi
kebutuhan sel. Namun demikian, tanpa adanya glukosa, sel harus menggunakan
substrat lain yaitu lemak yang dirubah menjadi asam lemak dan Asetil-CoA, serta
protein yang diurai menjadi asam amino yang dapat digunakan di dalam siklus Kreb.
Defisit glukosa mengakibatkan meningkatnya AMP di dalam sel yang
kemudian mendorong fosforilasi beberapa enzim dan mengaktifasi sejenis AMP
kinase. AMP Kinase ini kemudian mendorong terjadinya pemisahan antar sel (sel
detachments) yang ditandai dengan adanya protein G-aktin. Aktifitas AMP Kinase ini
menghambat fosforilasi Focal Adhesion Kinase (FAK) yang berperan dalam
perlekatan antar sel dan juga menghambat fosforilasi Protein Kinase C (Suzuki et al.
2003).
Kondisi lapar sel (defisit glukosa di dalam sel) mengakibatkan menurunnya
rasio ATP:AMP di dalam sel yang kemudian mendorong mekanisme metabolisme
tubuh dan memobilisasi sel-sel lemak untuk digunakan sebagai sumber energi
alternatif. Mobilisasi dan penguraian lemak mengakibatkan kadar asam lemak bebas
menjadi tinggi di dalam darah dan ekstrasel, dan asam lemak ini kemudian berfungsi
sebagai caraka yang mengaktifasi reseptor α dengan bantuan peroxisome proliferator.
Aktifasi reseptor ini kemudian mendorong ekspresi isoform dari piruvat
dehidrogenase kinase (PDK4) yang kemudian menghambat aktifitas enzim kompleks
piruvat dehidrogenase yang berakibat pada penumpukan asam laktat dan alanin
sebagai bahan baku glukoneogenesis (Wu et al. 2001). Mekanisme lainnya terjadi
dengan perlekatan antagonis Basilen Blue yang melekat pada reseptor P2 yang
berperan sebagai reseptor bagi messenger yang memicu ekspresi dan keluarnya
sitokrom c dari mitokondria dalam kondisi “lapar” glukosa. Sitokrom c ini
mendorong peningkatan ekspresi Heat Shock Protein (HSP), enzim kaspase-2, GRP-
75 dan GRP-78 yang diatur oleh kadar glukosa. Dengan melekatnya antagonis pada
reseptor P2, maka sitokrom c tidak keluar dari mitokondria dan peningkatan ekspresi
protein dan enzim lainnya tidak terjadi (Cavaliere et al. 2001).
Dalam kondisi lapar (defisit glukosa), sel dapat berhenti untuk berkembang
(replikasi) dan bahkan dapat terdorong untuk mengalami apoptosis/kematian sel.
Rasio ATP:AMP serta kadar glukosa yang rendah di dalam sel menyebabkan aktifasi
reseptor “kematian sel” yang selanjutnya mengaktifasi enzim kaspase 8 dan

26
pengiriman signal untuk mematikan sel. Aktifasi reseptor “kematian sel” dihambat
oleh aktifasi suatu kelas protein kinase ARK5 yang ekspresinya didorong oleh aktifasi
protein Akt. Growth factors dan Fosfatidil inositol 3 kinase merupakan caraka yang
diterima oleh protein Akt tersebut dan merupakan bagian dari mekanisme pertahanan
hidup sel (Suzuki et al. 2003).
Lapar Serum
Selain glukosa, sel juga membutuhkan berbagai pembawa pesan (caraka)
untuk memastikan keberlangsungan hidup dan fungsi sel secara normal. Caraka ini
terdiri dari berbagai senyawa seperti epidermal growth factor (EGF) maupun faktor-
faktor pertumbuhan lainnya. Secara in vitro, kondisi lapar serum dapat diciptakan
dengan menggunakan media pertumbuhan yang mengandung Fetal Calf Serum (FCS)
atau Fetal Bovine Serum (FBS) sebagai nutrisi sel yang kandungannya dapat
dikurangi untuk menciptakan kondisi lapar tersebut. Kondisi lapar serum pada sel
dapat menimbulkan hambatan pada pertumbuhan bahkan kematian sel melalui
berbagai mekanisme. Yamada et al. (1996) menunjukkan bahwa pengurangan nutrisi
pada media pertumbuhan sel (medium Eagle termodifikasi) mengakibatkan aktifasi
suatu reseptor yang dikenal sebagai AT2 (Angiotensin receptor) pada membran sel
yang kemudian mendorong ekspresi dari protein angiotensin II yang mendorong
defosforilasi pada kinase MAP (Mitogen-activated). Dengan defosforilasi seperti ini,
enzim MAP kinase menjadi inaktif dan sel tidak dapat melakukan pembelahan
(replikasi), dan dengan demikian kondisi lapar serum ini mengakibatkan hambatan
pada pertumbuhan yang dapat berakhir dengan terjadinya apoptosis sel tersebut.
Salah satu penyebab terjadinya hambatan pertumbuhan sel akibat lapar serum
adalah tidak adanya faktor-faktor pertumbuhan (EGF dan Insulin-like Growth Factor)
yang merupakan caraka yang diterima oleh reseptor EGF pada membran sel.
Melekatnya EGF pada membran sel memicu serangkaian reaksi yang merupakan
fosforilasi (aktifasi) berbagai enzim tertentu termasuk MAP kinase yang mendorong
faktor transkripsi dan aktifasi protein adaptor Nck yang menghubungkan reseptor
Tirosin kinase dengan Pak-1 yaitu suatu kinase serin-threonin yang berperan dalam
pembentukan aktin dari sitoskeleton (Galisteo et al. 1996). Tanpa adanya faktor
pertumbuhan (EGF) yang melekat pada reseptor yang ada di membran sel, maka
proses pertumbuhan sel tidak dapat berjalan. Peranan reseptor tirosin kinase terlihat
juga dari hasil penelitian Zeng et al. (2001) pada sel endothel HUVEC (Human

27
Umbilical Vein Endothel) yang berinteraksi dengan vascular endothelial growth
factor (VEGF). VEGF sebagai caraka melekat dan mengaktifasi dua jenis reseptor
tirosin kinase pada membran sel yaitu: Flt-1 dan KDR yang memiliki mekanisme
respons yang berbeda. Aktifasi reseptor KDR mendorong proliferasi sel, sementara
aktifasi Flt-1 mendorong respons Fosfatidil Inositol yang bersifat sebagai inhibitor
terhadap proliferasi yang dipicu oleh aktifasi reseptor KDR. Aktifasi KDR sendiri
berujung pada mekanisme mobilisasi Ca+ intrasel, syarat bagi berbagai fungsi sel
seperti pertumbuhan, transkripsi, dan juga kehidupan sel secara umum. Tidak adanya
growth factor yang melekat pada reseptor dalam kondisi lapar serum mengakibatkan
tidak adanya signal yang mendorong proliferasi sel dan/atau ekspresi protein-protein
tertentu yang esensial dalam menjalankan fungsi sel yang dapat berujung pada
kematian sel (Vaux & Korsmeyer 1999).
Ketiadaan caraka seperti growth factor pada kondisi lapar serum tidak hanya
mematikan beberapa proses seluler namun juga mengaktifasi ekspresi beberapa gen
seperti gen sdr (starvation deprivation response) yang ekspresinya meningkat dalam
kondisi lapar serum, namun menurun dengan adanya Epidermal growth factor-EGF
(Gustincich & Schneider 1993) dan protein SRBC yang memiliki 43% kesamaan
dengan sdr (Izumi et al. 1997). Protein SRBC dapat berikatan dengan protein kinase
C dan memodifikasi (menghambat) fungsi PKC dalam perkembangan sel. Kondisi
lapar serum juga dapat menstimulasi ekspresi ADP ribose sintetase yang
menyebabkan tingginya kadar ADP-ribosa di dalam sel yang berujung pada
fragmentasi DNA (Wielckens et al. 1983).
Ambang Batas dan Proses Perbaikan dalam Kondisi Lapar Sel
Lapar sel mengakibatkan kondisi sel yang dapat berakibat pada kematian sel
berupa apoptosis ataupun nekrosa yang didorong oleh berbagai caraka seperti Fas
Ligand, Tumor Necrosing Factor (TnF) dan lain-lain yang memicu rangkaian reaksi
yang berujung pada kematian sel. Namun demikian, sel memiliki mekanisme
pertahanan hidup yang memungkinkan sel untuk menghambat reaksi berantai tadi dan
mencegah kematian sel. Mekanisme pertahanan hidup sel ini dijelaskan oleh Suzuki
et al. (2003) dan disajikan dalam Gambar 5. Berdasarkan diagram ini, maka ambang
batas terjadinya kematian sel sangat ditentukan oleh kemampuan aktifitas enzim
caspase-3 untuk memulai proses kematian sel. Aktifitas enzim ini juga dipengaruhi
oleh aktifitas protein ARK5 yang menunjukkan bahwa peranan konsentrasi caraka

28
kematian sel dan juga caraka growth factor pemicu aktifitas ARK5 merupakan faktor
penentu ambang batas kemampuan sel untuk bertahan hidup (mencegah kematian
sel). Pemisahan sel (cell detachments) dan kematian sel didorong oleh melekatnya
caraka kematian sel berupa TNF dan Fas Ligand. Ekspresi protein caspase yang
merupakan komponen penting pemicu kematian sel dapat dihambat oleh aktifitas
protein ARK5 yang diaktifasi melalui serangkaian reaksi yang diawali dengan
melekatnya caraka berupa growh factor dan enzim kinase Fosfatidil Inositol 3 (PI3).
Proses perbaikan sangat bergantung pada proses masuknya gula yang
dibutuhkan oleh metabolisme sel ke dalam lingkungan intrasel. Proses ini dapat
terjadi berkat adanya aktifitas protein GLUT2 dan SGLT 1 yang berperan dalam
transport glukosa dengan bantuan ion Na+, serta protein GLUT5 yang berperan dalam
transport fruktosa ke dalam sel (Harmon & Macleod 2001). Setelah molekul
monosakarida masuk ke dalam lingkungan intrasel, maka aktifitas enzim heksokinase
diperlukan untuk mengubah glukosa menjadi glukosa-6-fosfat melalui proses
fosforilasi. Pada hewan herbivora monosakarida yang didapat dari penguraian
polisakarida dari tumbuhan adalah fruktosa, oleh karena itu peranan protein transport
GLUT5, GLUT2, dan berbagai enzim sangat diperlukan dalam proses transpor
fruktosa ke dalam lingkungan intrasel serta merubah dan mengurai molekul fruktosa
menjadi piruvat yang merupakan salah satu bahan baku yang diperlukan dalam siklus
Krebs. Selain interaksi ion-ion dan protein tertentu, dalam proses metabolisme
lainnya interaksi antar hormon merupakan kunci dari keseimbangan metabolisme
karena hormon-hormon tersebut berinteraksi dan berfungsi sebagai pembawa pesan
untuk memastikan proses perbaikan ataupun proses perkembangan sel dapat terus
berlangsung. Hal ini terjadi dalam proses respons terhadap cekaman di mana hormon
kortisol memainkan peran yang penting bagi hewan untuk bertahan dalam kondisi
tersebut.

29
Peranan dan Interaksi Hormon Kortisol dan Tiroksin sebagai Caraka
Komunikasi antar sel dalam konteks lapar sel, kebutuhan hidup, dan
kebutuhan sel untuk bertahan hidup difasilitasi oleh berbagai caraka berupa hormon
seperti tiroksin (hormon tiroid – T4) dan hormon yang berhubungan dengan kondisi
cekaman (adrenalin, kortisol). Hormon tiroid merupakan hormon yang berperan
dalam berbagai proses metabolisme terutama yang berhubungan dengan
pertumbuhan. Hormon tiroid larut di dalam lemak sehingga dapat dengan mudah
menembus membran sel dan masuk ke dalam nukleus untuk mendorong transkripsi
enzim-enzim yang bersifat oksidatif fosforilatif. Enzim-enzim ini merupakan
komponen penting dalam pembentukan ATP, modal utama kehidupan sel dan
menempatkan peranan hormon tiroksin sebagai suatu hormon yang esensial untuk
kehidupan sel.
Gambar 5. Rangkaian reaksi yang dipicu oleh kondisi lapar sel (menurunnya kadar gula
dalam sel) yang berujung pada pemisahan sel (cell detachments), dan kematian
sel. Sumber: Suzuki et al. (2003), Wu et al. (2001), dan Cavaliere et al.
(2001).
Sementara itu kortisol merupakan hormon yang sekresinya diatur oleh
corticotropin releasing hormone (CRH) dari hipothalamus dan merupakan suatu
respons terhadap cekaman metabolik seperti dalam kondisi lapar sel. CRH

30
merupakan caraka yang melekat pada membran sel kelenjar adrenal memicu ekspresi
dan sekresi hormon kortisol. Kortisol kemudian memainkan peranan penting dalam
metabolisme lipid, suatu kondisi yang terpicu oleh kondisi lapar sel. Berbeda dengan
tiroksin, hormon kortisol bukan merupakan kompenen esensial dalam kehidupan sel,
melainkan bagian dari mekanisme yang diperlukan oleh sel untuk bertahan hidup
dalam kondisi cekaman metabolik seperti kelaparan, keberadaan zat hepatotoksin, dan
cekaman lainnya (Helen & Brindley 1978). Penelitian yang dilakukan oleh Garg &
Chander (1997) menunjukkan tingginya kadar hormon kortisol pada plasma darah
kerbau yang mengalami cekaman berupa torsio uteri. Tingginya kadar hormon
kortisol diiringi dengan rendahnya kadar hormon tiroksin yang disebabkan oleh
adanya efek inhibitori kortisol terhadap sekresi Thyroid Stimulating Hormone (TSH).
Keberadaan kortisol ini dikaitkan juga dengan adanya kondisi lapar akibat torsio uteri
yang menyebabkan anoreksia pada hewan tersebut.
Dalam penelitian yang dilakukan pada hati tikus, Helen & Brindley (1978)
menunjukkan bahwa kortisol mengakibatkan peningkatan pada biomasa liver, dan
peningkatan trigliserida pada plasma darah. Peningkatan kadar trigliserida ini terjadi
akibat peranan hormon kortisol dalam meningkatkan proses lipolisis dan menurunkan
esterifikasi asam lemak. Dalam penelitian yang sama hormon tiroksin juga berperan
dalam peningkatan kadar trigliserida dalam darah dengan meningkatkan sintesa
trigliserida dari gliserol fosfat. Kortisol juga memiliki peranan langsung maupun
tidak langsung dalam meningkatkan aktifitas enzim fosfatida fosfohidrolase, enzim
yang berperan dalam sintesa dan akumulasi trigliserida pada hati.
Hormon kortisol –seperti yang ditunjukkan oleh Nessmith et al. (1983)-
memilki peranan dalam meningkatkan aktifitas enzim glukosa-6-fosfat dehidrogenase
yang berperan dalam produksi NADPH suatu senyawa berenergi tinggi yang biasanya
dihasilkan siklus Krebs. Kerja sinergi kortisol dan tiroksin menghasilkan keluaran
berupa peningkatan aktifitas enzim yang disertai dengan peningkatan lipid pada hati.
Bila bekerja sendirian tiroksin menghasilkan peningkatan lipid pada hati dan hanya
menunjukkan peningkatan kecil pada aktifitas enzim, sementara kortisol sendiri hanya
memberikan keluaran berupa peningkatan aktifitas enzim. Selain meningkatkan
metabolisme lipid dalam kondisi lapar sel, kortisol dan tiroksin juga berperan dalam
mendorong penguraian otot menjadi asam amino alanin, senyawa yang diperlukan
untuk mempertahankan proses dalam siklus krebs saat kadar monosakarida menjadi

31
rendah dalam kondisi lapar sel. Sebaliknya, kedua hormon ini memiliki fungsi dalam
meredam pelepasan asam amino glutamin dari sel otot tersebut (Karl et al. 1976).
Interaksi antara sistem endokrin dengan sistem tanggap kebal telah dipelajari
pada sistem endokrin hormon glukokortikoid (termasuk kortisol) yang memberikan
pengaruh pada sistem tanggap kebal dengan berfungsi sebagai caraka untuk memicu
perubahan pada fungsi sel-sel limfoid seperti sel T (Kelley 1988). Dalam keadaan
lain hormon glukokortikoid memiliki kemampuan untuk memicu apoptosis pada sel
leukimia yang berasal dari sel T dengan cara berperan dalam proses fragmentasi DNA
(Bansal et al. 1991).
Berbagai ancaman dan gangguan pada keseimbangan sel dapat terjadi akibat
dorongan berbagai aspek seperti toksisitas, nutrisi, dan juga faktor cekaman yang
kemudian memicu berbagai reaksi hormonal sebagai respon dan upaya sel untuk
bertahan hidup. Salah satu ancaman yang mengancam kehidupan herbivora seperti
badak adalah konsumsi antinutrisi yang bersifat toksin yang berasal dari tumbuhan
jenis Lantana camara. Tumbuhan ini adalah salah satu jenis tumbuhan pakan badak
yang mengandung toksin namun juga disukai oleh badak sebagai makanan sehari-hari
dengan palatabilitas yang cukup tinggi.
Lantana camara sebagai Sumber Bahan Toksin/Antinutrisi
Sebagai satwa liar yang sepenuhnya tergantung pada ketersediaan pakan di
alam, badak jawa memakan sekitar 27 famili tumbuhan pakan yang ada di kawasan
Taman Nasional Ujung Kulon (Hoogerwerf 1970). Dengan ketersediaan pakan yang
ada saat ini, dan ditunjang dengan pengamatan visual menggunakan kamera otomatis
pada kondisi fisik badak jawa yang dapat dilihat dalam kondisi normal (tidak kurus),
tidak ada indikasi bahwa satwa ini untuk mengalami malnutrisi. Namun demikian,
keracunan akibat konsumsi jenis pakan dan anti nutrisi tertentu merupakan salah satu
ancaman yang mungkin terjadi pada satwa liar seperti badak jawa. Pengamatan yang
dilakukan di lapangan selama lima tahun terakhir menunjukkan adanya jenis
tumbuhan yang dapat mendominasi tumbuhan pakan yang tersedia bagi badak jawa.
Jenis tumbuhan Lantana camara merupakan salah satu tumbuhan dengan kandungan
zat lantaden yang dikenal telah mengakibatkan keracunan pada berbagai hewan ternak
(Mandial 2006; Sharma 2006). Secara kualitatif berdasarkan frekuensi ragutan yang
ditemukan dalam sensus badak di Taman Nasional Ujung Kulon, tumbuhan ini
merupakan salah satu jenis tumbuhan yang banyak dimakan oleh badak (Sriyanto et

32
al. 1995). Tumbuhan Lantana camara memiliki ciri khas berupa bunga berwarna
cerah sebagaimana ditampilkan dalam Gambar 6.
Gambar 6. Tumbuhan perdu Lantana camara di habitat badak di Taman Nasional
Ujung Kulon memiliki bunga berwarna cerah. Foto: Ahariyadi – WWF
Indonesia (2004)
Tumbuhan ini merupakan tumbuhan invasif yang memiliki daya tahan lebih
tinggi dibandingkan dengan berbagai jenis tumbuhan lainnya, dan berpotensi untuk
mengakibatkan ketidakseimbangan di dalam ekosistem (Sharma et al. 2005). Sharma
(2006) menyebutkan bahwa ekstraksi dan identifikasi Lantaden A (C32H44O5) dan
Lantaden B (C33H48O5) sebagai komponen hepatotoksin yang ditemukan dalam
tumbuhan Lantana camara telah dilakukan sejak tahun 1948. Lantaden diserap
melalui saluran pencernaan, namun penyerapan paling optimal terjadi di usus halus.
Keracunan lantaden terjadi dalam dua tahap yaitu tahap saluran pencernaan saat
toksin keluar setelah tumbuhan dicerna, dan toksin tersebut diserap masuk ke
peredaran darah, diikuti dengan tahap hepatik, toksin masuk ke hati dan disertai
timbulnya gejala-gejala klinis seperti anoreksia, jaundice, dan fotosensitifitas
(Mandial 2006). Keracunan lantaden disebabkan oleh konsumsi tumbuhan Lantana
camara tanpa memerlukan metabolisme perantara, dan mengakibatkan kerusakan
pada hati (Pass et al. 1979). Konsumsi daun Lantana kering sebanyak 4 g/kg berat
badan setiap hari pada ruminansia kecil (kambing) selama 6 hari berturut-turut
menyebabkan stasis pada rumen yang merupakan akibat dari menurunnya impuls
syaraf karena kerusakan pada hati (Pass et al. 1979). Selain lantaden, tumbuhan

33
Lantana cammara juga mengandung zat-zat antinutrisi dari golongan triterpenoid dan
flavonoid (Nayak et al. 2008).
Hewan-hewan Model dan Pembanding untuk Badak Jawa
Dalam kajian-kajian biomedis, penggunaan hewan model lazim dilakukan
untuk menganalogikan respons biologis pada manusia. Dalam bidang biomedis
hewan, konsep pemilihan hewan model dapat diterapkan untuk melakukan analogi
sistem biologis pada satwa langka tanpa harus mengorbankan dan/atau mengancam
kehidupan satwa langka tersebut. Kaidah-kaidah pemilihan hewan model meliputi:
ketersediaan, kemudahan-kemudahan (perawatan, transportasi, perizinan), kemiripan
fisiologis, efisiensi biaya, dan yang terpenting kemiripan respons hewan model dan
badak jawa terhadap parameter-parameter nutrisi, dan hormon cekaman.
Badak Sumatera (Dicerorhinus bicornis)
Secara kekerabatan, badak sumatera dan badak jawa berada dalam famili yang
sama yaitu Rhinocerotidae. Hal ini memberikan potensi bagi badak sumatra untuk
menjadi analog yang tepat untuk badak jawa. Badak sumatera hidup secara soliter di
ekosistem hutan hujan tropis (Strien 1986) dengan pakan alami yang terdiri dari 31
genera tumbuhan dari 13 famili (Strien 1974). Sama halnya dengan badak jawa,
badak sumatera juga merupakan browser yang meragut tumbuhan pakan
menggunakan bibir bagian atas. Secara anatomis, semua spesies badak merupakan
hewan monogastrik dengan sistem pencernaan yang mengandalkan fermentasi dan
penguraian selulosa pada sekum (Pough 1989). Penyakit-penyakit seperti: dermatitis
dan kecacingan merupakan kasus-kasus yang pernah terjadi pada badak sumatra baik
di alam maupun di penangkaran. Penyakit dan/atau gejala klinis seperti ini merupakan
hal yang dapat terjadi pada semua spesies badak yang hidup liar, dan berpotensi pula
untuk terjadi pada badak jawa. Berdasarkan kekerabatan yang dekat, kesamaan
tipologi habitat, kemiripan komposisi pakan, dan juga kemiripan kerentanan terhadap
penyakit dan/atau gejala klinis yang bersumber dari nutrisi dan cekaman, maka
penggunaan badak sumatra sebagai model untuk badak jawa kemungkinan besar akan
memberikan hasil yang akurat.

34
Badak India (Rhinoceros unicornis)
Berdasarkan filogram pada gambar 7 dapat dilihat bahwa salah satu kerabat
dekat dari badak jawa adalah badak India yang dikenal juga sebagai Greater One-
Horned Rhinoceros. Studi yang dilakukan oleh Clauss et al. (2005) telah
menghasilkan suatu data fisiologi pencernaan tentang tingkat retensi dan penyerapan
air serta nutrisi yang terjadi pada spesies badak yang berada di penangkaran. Atas
dasar kekerabatan yang dekat, dan juga banyaknya informasi yang telah digali dari
spesies badak ini, maka badak india merupakan hewan model yang cukup tepat bagi
badak jawa untuk mempelajari dan membandingkan beberapa aspek fisiologi dan
perilaku.
Kuda (Equus caballus)
Berdasarkan telaah genetika dengan membandingkan urutan gen r12S RNA
pada DNA mitokondria, tapir dan kuda termasuk kerabat dekat badak (Fernando et al.
2005) sebagaimana disajikan dalam Gambar 7. Sebagai herbivora monogastrik,
anatomi sistem pencernaan kuda dan badak memiliki kesamaan dengan adanya sekum
(Fowler & Miller 2003) yang berfungsi sebagai reservoir tempat bakteri yang
membantu penguraian selulosa (komponen utama tumbuhan) sebagai sumber
polisakarida bagi herbivora monogastrik (Raven & Johnson 1989). Demikian pula
dengan kebutuhan makronutrien dan mineral, kuda dan badak memiliki kesamaan
dengan perbedaan pada kebutuhan vitamin yang larut dalam lemak (Fowler & Miller
2003). Penelitian digesti yang dilakukan oleh Clauss et al. (2005) pada badak india
(Rhinoceros unicornis) menajamkan kemiripan fisiologi pencernaan badak dan kuda
pada tingkat retensi dan absorpsi nutrisi. Kuda bukan merupakan satwa dilindungi
dan sudah dimanfaatkan oleh manusia untuk berbagai keperluan, oleh karena itu,
induksi stres dan toksisitas dapat dilakukan pada kuda tanpa menimbulkan implikasi
dan ancaman bagi kelestarian satwa yang dilindungi. Penggunaan kuda sebagai
hewan model untuk badak memenuhi kaidah pemilihan hewan model untuk hewan
(orphan). Salah satu hal penting yang perlu dicermati dalam perbandingan ini adalah
fakta bahwa palatabilitas pakan badak tidak sama dengan kuda, oleh karena itu
penggunaan kuda sebagai model pencernaan untuk badak memerlukan pembuatan
formulasi pakan khusus (ransum) untuk menjamin palatabilitas tinggi dan kualitas
nutrisi yang memadai untuk kuda.

35
Gambar 7. Filogram yang menunjukkan kekerabatan berbagai spesies berdasarkan variasi
pada DNA 12S RNA. Angka menunjukkan kedekatan kekerabatan berdasarkan
perhitungan parsimoni maksimum. Sumber: Fernando et al. (2005)
Kuda dan badak diklasifikasikan dalam ordo yang sama yaitu Perissodactyla
(hewan berkuku ganjil) dengan berbagai persamaan anatomis dan fisiologis. Seperti
halnya badak, kuda merupakan hewan herbivora yang monogastrik. Komposisi darah
kuda dan badak mengandung asam amino tirosin dalam bentuk bebas di dalam sel
darah merah, serta memiliki antioksidan yang tinggi. Kedua hal ini menyebabkan
kuda dan badak memiliki kemampuan tinggi untuk melakukan aktifitas, bertahan
terhadap penyakit, serta untuk bertahan hidup di lingkungan yang berat (Harley et al.
2004). Uji coba dengan tikus menunjukkan bahwa tyrosin berperan sebagai peredam
stres dengan mempengaruhi proses biokimia dalam hippokampus (Hao et al. 2001).

36
Analogi Perbandingan
Pemilihan hewan model sebagai pembanding untuk badak jawa dilakukan
berdasarkan kesamaan-kesamaan fisiologis, anatomis, genetika, dan juga perilaku.
Badak merupakan hewan monogastrik yang mengandalkan penguraian selulosa dari
makanannya di sekum yang berada di bagian belakang (hindgut fermenter), dan oleh
karena itu hewan model yang digunakan dalam kajian pakan harus juga merupakan
hewan yang juga merupakan hindgut fermenter seperti badak. Clauss et al. (2005)
menunjukkan bahwa kuda dan badak memiliki kemiripan dalam sistem pencernaan,
termasuk karakteristik penyerapan nutrien.
Kesamaan antar hewan model dipertegas dengan fakta bahwa hewan-hewan
dalam ordo Perissodactyla (kuda dan badak) kerap menunjukkan gejala klinis yang
serupa. Salah satu contoh kemiripan ini adalah gejala klinis yang dikenal dengan
istilah kolik yang disebabkan oleh etiologi yang beragam (Chaffin & Cohen 1999),
sementara kolik juga ditemukan pada kasus kematian badak di tahun 1982 (Strien
1982; WWF/IUCN 1982) dan kasus kematian di tahun 2003 di Taman Nasional
Ujung Kulon. Hal ini menunjukkan bahwa secara klinis kedua spesies hewan ini
memiliki persamaan, walaupun penyebab kolik pada kuda lebih banyak disebabkan
karena faktor pemeliharaan, sementara risiko dan penyebab kolik pada badak yang
hidup secara alami belum diketahui secara pasti sampai saat ini.
Faktor komposisi pakan dan asupan antinutrisi pada hewan-hewan
Perissodactyla merupakan aspek yang penting dan berpotensi untuk mengakibatkan
perubahan fisiologis. Glade et al. (1984) dalam suatu eksperimen mencatat bahwa
perubahan diet berupa komposisi kandungan energi dan protein mengakibatkan
meningkatnya kadar kortisol dalam serum yang merupakan karakteristik yang sama
dengan respons hewan terhadap cekaman. Dalam aspek antinutrisi Sharma (2006)
menyebutkan bahwa hepatotoksin lantaden dari Lantana camara dalam tingkat
asupan tertentu dapat menyebabkan keracunan dengan gejala klinis berupa perubahan
perilaku pada hewan ruminansia dan non-ruminansia termasuk kuda.
Contoh-contoh di atas menunjukkan bahwa kuda merupakan hewan model
responsif terhadap jenis-jenis cekaman yang dihadapi oleh badak di habitat alaminya
terkait keterbatasan pakan dan air. Pemilihan kuda sebagai hewan model pembanding
badak jawa untuk telaah diet dan dinamika hormon kortisol merupakan langkah yang
logis dan dapat digunakan untuk memastikan bahwa keterbatasan pakan dan air
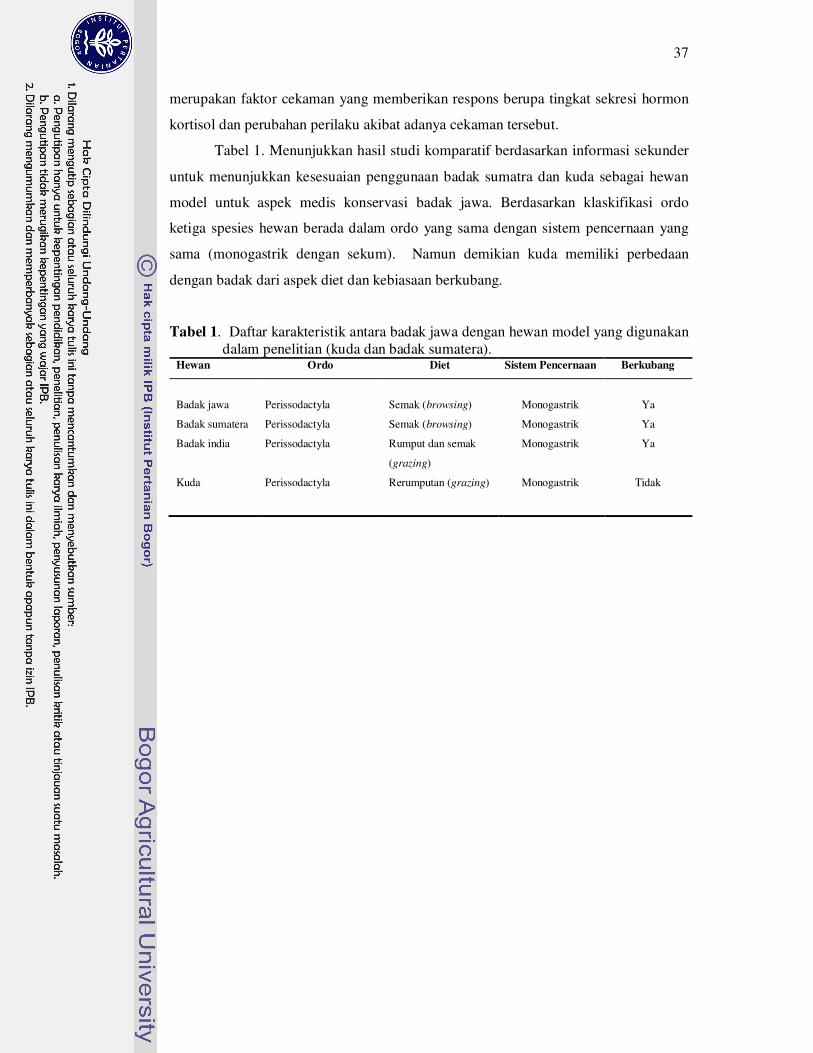
37
merupakan faktor cekaman yang memberikan respons berupa tingkat sekresi hormon
kortisol dan perubahan perilaku akibat adanya cekaman tersebut.
Tabel 1. Menunjukkan hasil studi komparatif berdasarkan informasi sekunder
untuk menunjukkan kesesuaian penggunaan badak sumatra dan kuda sebagai hewan
model untuk aspek medis konservasi badak jawa. Berdasarkan klaskifikasi ordo
ketiga spesies hewan berada dalam ordo yang sama dengan sistem pencernaan yang
sama (monogastrik dengan sekum). Namun demikian kuda memiliki perbedaan
dengan badak dari aspek diet dan kebiasaan berkubang.
Tabel 1. Daftar karakteristik antara badak jawa dengan hewan model yang digunakan
dalam penelitian (kuda dan badak sumatera). Hewan Ordo Diet Sistem Pencernaan Berkubang
Badak jawa Perissodactyla Semak (browsing) Monogastrik Ya
Badak sumatera
Badak india
Perissodactyla
Perissodactyla
Semak (browsing)
Rumput dan semak
(grazing)
Monogastrik
Monogastrik
Ya
Ya
Kuda Perissodactyla Rerumputan (grazing) Monogastrik Tidak

38
Daftar Pustaka
Allen CW, White CB,. 1998. Glucose modulates cell death due to normobaric
hyproxic by maintaining cellular ATP. Am J Physiol Lung Cell Mol
Physiol 274:159-164
Bansal N, Houle A, Melnykovych G,. 1991. Apoptosis: mode of cell death
induced in T Cell leukimia lines by dexamethasone and other agents.
FASEB J. 5:211-216
Barja I, et al. 2007. Stress physiological response to tourist pressure in the wild
population of European pine marten. The journal of steroid biochemistry
and molecular biology. (104)3-5:136-142
Baron R. 2006. Mechanism of disease: neuropathic pain-a clinical perspective.
Nature clinical practice neurology. 2:95-106
Bennet RM,. 1999. Emerging Concepts in the Neurobiology of Chronic Pain:
Evidence of Abnormal Sensory Processing in Fibromyalgia. Mayo Clin
Proc;74:385-398
Berridge MJ,. 1984. Inositol Trisphosphate and Diacylglycerol as Second
messengers. Biochem J 220: 345-360
Bruni JF, Hawkins RL, Yen SS,. 1982. Serotonergic Mechanism in the Control of
Beta-endorphin and ACTH Release in Male Rats. Life Sci. 30(15): 1247-
1254
Cameron HL, Perdue MH. 2005. Stres Impairs Murine Intestinal Barrier Function:
Improvement by Glucagon-Like Peptide-2. The journal of pharmacology
and experimental therapeutics Vol. 314, No. 1
Carlstead K, Brown JL,. 2005. Relationships between patterns of fecal corticoid
excretion and behaviour, reproduction, and environemental factors in
captive black (Diceros bicornis) and white (Ceratohterium simum)
Rhinoceros. Zoo biology 24: 215-232
Cavaliere F, et al. 2001. Hypoglycaemia-induced cell death: features of
neuroprotection by the P2 receptor antagonist basilen blue. Neurochemistry
International Volume 38, Issue 3: 199-207
Chaffin MK, Cohen ND,. 1999. Diagnostic Assessment of Foals with Colic.
Proceedings of the Annual Convention of the AAEP vol 45
Clausen B,. 1981. Survey For Trypanosomes in Black Rhinoceros (Diceros
bicornis). Journal of Wildlife Diseases Vol. 17, No. 4, October, 1981

39
Clauss M, et al. 2005. Studies on digestive physiology and feed digestibilities in
captive Indian rhinoceros (Rhinoceros unicornis). Journal of Animal
Physiology and Animal Nutrition 89 (2005) 229–237
Corbett GB, Hill JE. 1992. The mammals of Indomalayan region: a systematic
review. Natural History Museum Publications. Oxford University Press.
Cranbrook E, Piper PJ,. 2007. Short Communications: Javan rhinoceros
Rhinoceros sondaicus in Borneo. The Raffles Bulletin of Zoology 2007
55(1): 217-220
Daszak P, Cunningham AA, Hyatt AD,. 2000. Emerging infectious diseases of
wildlife –threats to biodiversity and humant health. Science 287: 443-449
Daszak P,. et al. 2004. Conservation Medicine and New Agenda for Newly
Emerging Diseases. Ann. N.Y. Acad. Sci. 1026: 1–11
Deem SL, et al. 2001. Conservation Medicine. Annals New York Academy of
Sciences pp: 370-377
Dhabhar FS,. 2000. Acute stress enhances while chronic stress supresses skin
immunity: the roles of stress hormones and leukocyte trafficking. Annals of
the New York academy of Sciences vol 917: Neuroimmunomodulation
perspectives at the new millenium:876-893
Dyer J, et al. 2002. Molecular characterisation of carbohydrate digestion and
absorption in equine small intestines. Equine veterinary journal 34(4):349-
358
Endo H, et al. 2009. The morphological basis of the armor-like folded skin of the
greater Indian rhinoceros as a thermoregulator. Mammal study 34: 195-200
Fernando P, Polet G, Foead N. Ng L. Melnick DJM. 2004. Mitochondrial DNA
analysis of the critically endangered Javan Rhinoceros. PHKA-WWF-
Columbia University
Figueiredo HF, et al. 2003. Stress integration after acute and chronic predator
stress: Differential activation of central stress circuitry and sensitization of
th ehypothalamo-pituitary-adrenocortical axis. Endocrinology 144(12):
5249-5258
Firdaus Y,. 2008. Laporan Climate Modelling. Kajian Dampak Perubahan Iklim
Terhadap Kerentanan Badak Jawa. WWF Indonesia – Program Iklim dan
Energi
Fowler ME, Miller RE,. 2003. Zoo and Wild Animal Medicine 5th
edition.
Saunder Publishing, St Louis Missouri. p: 559.

40
Galisteo ML, et al. 1996. The Adaptor protein Nck Links receptor tyrosine
kinases with Serine-Threonin Kinase Pak1. The Journal of Biological
Chemistry 271(35): 20997-2100
Gandolf AR, Willis AM, Blumer ES, Atkinson MW,. 1999. Management of a
Melting Corneal Ulcer in a Greater One-horned Rhinoceros (Rhinoceros
unicornis). Proceedings American Associations of Zoo Veterinarians
Garg SL, Chander S,. 1997. Plasma Cortisol and thyroid hormone Concentrations
in Buffaloes with uterine Torsions. Buffalo Bullettin Vol 16(4): 75-76
Glade MJ, Gupta S, Reimers TJ,. 1984. Hormonal Responses to High and Low
Planes of Nutrition in Weanling Thoroughbreds. J Anim Sci 1984. 59:658-
665
Gustincich S, Schenider C,. 1993. Serum Deprivation Response Gene Is Induced
by Serum Starvation but not by contact Inhibition. Cell Growth and
Differentiation vol 4: 753-760
Hao S, Avraham Y, Bonne O, Berry EM,. 2001. Separation-induced body weight
loss, impairment in alternation behavior, and autonomic tone: effects of
tyrosine. Pharmacology Biochemistry and Behavior Volume 68, Issue 2
Hariyadi ARS, Santoso A, Setiawan R, Priambudi A,. 2008. Automatic Camera
Survey for Monitoring Reproductive Pattern and Behaviour of Javan
Rhinoceros (Rhinoceros sondaicus) in Ujung Kulon National Park,
Indonesia. Proceeding of the 3rd
International Meeting on Asian
Zoo/Wildlife Medicine and Conservation (AZWMC)
Harley EH, Matshikiza M, Robson P, Weber B,. 2004. Red blood cell metabolism
shows major anomalies in Rhinocerotidae and Equidae, suggesting a novel
role in general antioxidant metabolism. Animals and Environments.
Proceedings of the Third International Conference of Comparative
Physiology and Biochemistry. Pp 334-340
Harmon DL, MacLeod KR,. 2001. Glucose Uptake and Regulation By Intestinal
Tissues: Implications and whole-body Energetics. J. Anim. Sci. 79(E.
Suppl.):E59–E72
Harvell CD, et al. 2002. Climate warming and disease risks for terrestrial and
marine biota. Science 296(5576):2158-2162
Hatt JM, Wenker C, Castell J, Clauss M,. 2004. Dietary dan Veterinary
Management of a Lingual Abscess in a Geriatric Captive Black Rhino
(Diceros bicornis) With Iron storage Disease. European Association of Zoo
and Wildlife Veterinarians (EAZWV) 5th
Scientific Meeting. Ebeltoft-
Denmark

41
Helen PG, Brindley DN,. 1978. The Effects of Cortisol, Corticotropin, and
thyroxine on the synthesis of Glycerolipids and on Phospatidae
Phophohydrolase activity in Rat Liver. Biochem J. 176: 777-784
Hommel PWFM, 1987. Landscape ecology of Ujung Kulon (West java). Disertasi
universitas Wageningen, Belanda.
Hoogerwerf A. 1970. Udjung Kulon the land of the last javan rhinoceros. E.J.
brills. Leiden, Netherlands
Huxman TE, Scott RL., 2007. Climate Change, Water Dynamics, and Landscape
Water Balance. Southwest Hydrology. January-February
IRF-WWF. 2011. Vietnam’s javan rhino now extinct. WWW.panda.org. [9 Juni
2012]
Izumi Y, et al. 1997. A protein Kinase Cδ binding Protein SRBC whose
expression is Induced by Serum Starvation. The Journal of Biological
Chemistry Vol 272(11): 7381-7389
Jacobsen J,. 2005. A Review of Rhino Foot Problem. Fort Worth Zoo
Jessup DA, et al. 1992. Retrospective Evaluation of Leptospirosis In Free-ranging
and Captive Black Rhinoceros (Diceros bicornis) by Microscopic
Agglutination Titers dan Fluorescence Antibody Testing. Journal of Zoo
and Wildlife Medicine 23(4): 401-408
Juster RP, McEwen BS, Lupien SJ. 2009. Allostatic load biomarkers of chronic
stress and impact on helath and cognition. Neuroscience and
biobehavioural reviews: 1-15
Jolles J, et al. 1980. Modulation of brain Phosphoinotiside metabolism by ACTH-
sensitive Protein Phosporylation. Nature vol 289: 623-625
Jonyo JF,. 2005. Doctoring Rhinos: Diseases seen in Kenya
Kanemi O, et al. 2005. Acute stress reduces intraprenchymal lung natural killer
cells via beta-adrenergic stimulation. Clinical experimental immunology
139(1): 25-34
Karl IE, Garber AJ, Kipnis DM,. 1976. Alanine and Glutamine Sythesis and
Release from Skeletal Muscle. The Journal of biological Chemistry 251(3):
844-850
Kelley KW,. 1998. Cross-talk between the Immune and Endocrine Systems, J
Anim Sci. 66:2095-2108
Kloss CB,. 1927. Further records of the one-horned rhinoceros in the Malay
States. Journ. Fed. Malay States Mus. XIII, 1927

42
Kock N, et al. 1991. Coronary Artery Aneurism In Two Black Rhinoceroses
(Diceros bicornis) in Zimbabwe. Journal of Zoo and Wildlife Medicine
22(3): 355-358
Kock ND, et al. 1992. Serological Evidence for Cowdria Ruminantium Infection
in Free-ranging Black (Diceros bicornis) dan White (Ceratotherium simum)
Rhinoceroses in Zimbabwe. Journal of Zoo and Wildlife Medicine 23(4):
409-413
Kock ND,. 1996. Myocardial Purkinje Degeneration and Necrosis with Fibrosis in
Free-Ranging Black Rhinoceroses (Diceros bicornis) in Zimbabwe.
Journal of Wildlife Diseases, :32(2), 1996, pp :367-369
Lekagul , McNeely. 1977. Mammals of Thailand. Sahakarnbhath Co. Bangkok
Mace OJ, Afflek J, Patel N, Kellet GL,. 2007. Sweet taste recpetors in rats small
intestines stimulate glucose absorption through apical GLUT2. Journal of
Physiology 582.1:379-392
Mandial RK, 2006. An Insight into the Toxicological and Medicinal Properties of
Lantana camara plant. College of Veterinary and Animal Science
Palampur Himachal Pradesh
Martin PS, Stuart AJ,. 1995. Mammoth extinction: two continents and Wrangel
island. Radiocarbon Volume 37 no.1: 7-10.
McSweeney CS, Pass MA,. 1983. The mechanism of ruminal stasis in Lantana-
poisoned sheep. Quarterly journal of experimental physiology 68: 301-313
Melia KL, Duman RS,. 1991. Involvement of corticotropin –releasing factor in
chronic stress regulation of the brain noradrenergic system. Proc. Natl.
Acad. Sci. USA. Vol 88: 8382-8386 Neurobiology
Menargues A, Urios V, Mauri M,. 2008. Welfare assessments of captive asian
elephants (Elephas maximus) and Indian rhinoceros (Rhinoceros unicornis)
using salivary cortisol measurements. Animal Welfare (17): 305-312
Morilak DA, et al. 2005. Role of brain norepinephrine to behavioural response to
stress. Progress in neuro-psychopharmacology & biological psychiatry
29:1214-1224.
Munson L, Koehler JW, Wikonson JE, Miller RE,. 1998. Vesicular and Ulcerative
Dermatopathy resembling Superficial Necrotic Dermatitis in Captive Black
Rhinoceroses (Diceros bicornis). Vet Pathol 35: 31-42
Nayak BS, Raju SS, Rasumbhag A. 2008. Investigation of wound healing activity
of Lantana Camara L. In Sprague dawley Rats using burn wound model.
International Journal of Applied Research in Natural Products Vol 1(1): 15-
19

43
Nessminth S, Baltzell J, Bardanier CD,. 1983. Interaction of Glucocorticoid and
thyroxine in the responses of rats starvation-refeeding J. Nutr. 113: 2260-
2265
Newton FRS,. March 17 1874. The Secretary on Addition to the menagerie.
PHKA. 2007. Strategi dan rencana aksi konservasi badak Indonesia. Direktorat
jendral Perlindungan hutan dan kelestarian Alam, kementerian kehutanan
Republik Indonesia.
Paglia DE, Radcliffe RW,. 2000. Anthracycline Cardiotoxicity in a Black
Rhinoceros (Diceros bicornis): Evidence for Impaired Antioxidant
Capacity Compounded by Iron Overload. Vet Pathol 37:86–88
Pass MA, Seawright AA, Lamberton JA, Heath TJ,. 1979. Lantaden A toxicity in
sheep. A model for cholestasis. Pathology. 1979 Jan;11(1):89-94
Permadi YF,. 2008. Kajian dampak perubahan iklim terhadap kerentanan badak
Jawa. Laporan proyek WWF Indonesia.
Polet G, et al. 1999. The javan rhinos, Rhinoceros sondaicus annamiticus, of Cat
Tien National Park, Vietnam: Current status and management implications.
Pachyderm 27:34-48
Poole CM, Duckworth JW,. 2005. A documented 20th
century record of Javan
rhino Rhinoceros sondaicus from Cambodia. Mammalia 69(3-4): 443-444
Pough FH, Heisser JB, McFarland WN,. 1989. Vertebrate Life – 3rd
edition.
MacMillan Publishing Company, New York p: 744
Putro HR,. 1997. Heterogenitas habitat Badak Jawa (Rhinoceros sondaicus Desm.
1822) Di Taman Nasional Ujung Kulon. Media Konservasi Edisi Khusus,
1997 : Hal.17-40
Raven PH, Johnson GB,. 1989. Biology – Second Edition. Times Mirror/Mosby
College Publishing. St Louis Missouri. p: 981
Reynolds RJ, 1961. Javan Rhino in The Berlin zoo. International Zoo news 8(3):
June –July 1961
Rookmaaker K,. 2005. A Javan Rhinoceros, Rhinoceros sondaicus,in Bali in
1839. Zool Garten N. F. 75 (2005) 2, 5. 129-131
Rookmaaker LC, 1982. The type locality of the Javan rhinoceros (Rhinoceros
sondaicus Desmarest, 1822). Sonderdrudre aus Zeitschrifl f.
Siugetierkunde Bd. 47 (1982), H. 6, S. 381-382
Sanborn ER,. 1908. Interesting Animal Surgery. Zoological Society Bullettin no
30. 1908

44
Scheltema JF,. 1912. Peeps at Many lands – Java. Adam & Charles Black. London
Schoeman JP, Goddard A, Herrtage ME,. 2007. Serum cortisol and thyroxine
concentrations as predictors of death in critically ill puppies with parvoral
diarrhea. Journal of the American veterinary medical association (231)10:
1534-1539
Sharma S, Sharma O, Singh B, Bhat TK,. 2000. Biotransformation of lantadenes,
the pentacyclic triterpenoid hepatotoxins of lantana plant, in guinea pig.
Toxicon 38(9): 1191-1202
Sharma GP, Ragubhanshi AS, Singh JS,. 2005. Lantana invasion: an overview.
Weed biology and management (5)4: 157-165
Sharma OP, 2006. An Overview of the Research on the Hepatotoxic Plant
Lantana Camara.
Soto-Gamboa M, Gonzalez S, Hayes LD, Ebensperger LA,. 2009. Validation of a
radioimmunoassay for measuring fecal cortisol metabolites in the
Hystricomorph rodent, Octodon degus. Journal of experimental zoology
311A:496-503
Sriyanto A, et al. 1995. A Current status of the Javan Rhino population in Ujung
Kulon National Park. Javan Rhino Colloquium
Strien NJ,. 1974. Dicerorhinus sumatrensis (Fisher). The Sumatran or two-horned
Asiatic rhinoceros: a study of literature. Medelebingen
Landbouwwhogeschool, Wageningen, Netherland
Strien NJ, 1982. Report on short mission to Ujung Kulon National Park, Java
Indonesia inconnectionwith the reported deaths of the five Javan rhinos.
Laporan kegiatan WWF.
Strien NJ,. 1986. The Sumatran rhinoceros – Dicerorhinus sumatrensis (Fischer,
1814) – in the Gunung Leuser National Park, Sumatra, Indonesia: its
distribution, ecology, and conservation. Verlag Paul Parey, Hamburg
Stuart G, Nelson S, Sakmann B, Hausser M,.1997. Action potential initiation and
backpropagation in neurons of the mammalian CNS. Trends in
neuroscience 20:125-131
Suprayogi A, Astuti DA, Satrija F, Suprianto. 2006. Physiological Status of Sheep
Reared Indoor System Under the Tropical Rain Forest Climatic Zone.
Supporting papers Proceedings of the 4th
ISTAP “Animal Production and
Sustainable Agriculture in the Tropic”. Faculty of Animal Science, Gajah
Mada University.

45
Suzuki A, et al. 2003. Induction of cell–cell detachment during glucose starvation
through F-actin conversion by SNARK, the fourth member of the AMP-
activated protein kinase catalytic subunit family. Biochemical and
Biophysical Research Communications Volume 311(1): 156-161
Suzuki A, et al. 2003. ARK5 suppresses the cell death induced by nutrient
starvation and death receptors via inhibition of caspase 8 activation, but not
by chemotherapeutic agents or UV irradiation. Oncogene 22: 6177–6182
Tiuria R, et al. 2006. Identification of endoparasites from faeces of Javan rhino
(Rhinoceros sondaicus) in Ujung Kulon National Park, Indonesia.
Proceedings of AZWMP 2006 Chulalongkorn Uni. Fac. Of Vet. Sc.,
Bangkok, Thailand
Turner Jr JW, Tolson P, Hamad M,. 2002. Remote assessment of stress in white
rhinoceros (Ceratotherium simum) and black rhinoceros (Diceros bicornis)
by measurements of adrenal steroids in feces. Journal of zoo and wildlife
medicine 33(3): 214-221
Vaux DL, Korsmeyer SJ,. 1999. Cell Death in Development. Cell vol 96: 245-254
Völlm J, et al. 2000. A case of extended exsudative dermatitis in an Indian rhino.
European Association of zoo and wildlife veterinarians (EAZWV) Paris,
France: 205-208
Walcott AS,. 1914. Java and Her Neighbours. A Traveler’s note in Java, Celebes,
the Molluccas, and Sumatra. GP Putnams and Sons. The knickerbockers
press. New York & London
Wasser SK, et al. 2000. A generalized fecal glucocorticoid assay for use in diverse
array of nondomestic mammalian and avian species. General and
comparative endocrinology 120:260-275
Wielckens K, et al. 1983. Stimulation of Poly(ADP-ribosyl)ation during Erlich
Ascites Tumor Cell “Starvation” and Suppression of Concomitant DNA
Fragmentation by Benzamide. The Journal of Biological Chemistry Vol
258(7): 4098-4104
Williams JW, et al. 2002. Rapid and widespread vegetation responses to past
climate change in the North Atlantic region. Geology vol 30; no. 11; p.
971–974
Woolf CJ, Mannion RJ,. 1999. Neuropathic pain: aetiology, symptoms,
mechanism, and management. Lancet 353:1959-64
Wu P, Peters JM, Harris RA,. 2001. Adaptive Increase in Pyruvate
Dehydrogenase Kinase 4 During Starvation is mediated by Peroxisome
Proliferator-activated Receptor α. Biochemical and Biophysical Research
Communication. 287(2): 391-396

46
WWF-IUCN. 1982. Mystery of Dead Javan rhinos. The Environmentalist vol. 2
no. 3
Yamada T, Horiuchi M, Dzau VJ,. 1996. Angiotensi type II receptor mediates
programmed cell death. Proc Acad Sci. USA. Cell Biology Vol 93: 156-160
Yakel JL, et al. 1997. Calcineurin regulation of synaptic function: from ion
channels to transmitter release and gene transcription. Trends in
Pharmacological Sciences. Volume 18, Issue 4: 124-134
YMR [Yayasan Mitra Rhino]. 2004. Implementasi monitoring habitat badak jawa
dengan menekan laju invasi langkap (Arenga obtusifolia) di Taman
Nasional Ujung Kulon. PHKA-WWF-YMR
Zeng H, Dvorak HF, Mukhopadhyay D,. 2001. Vascular Permeability Factor
(VPF)/Vascular Endothelial Growth Factor (VEGF) Receptor-1 Down-
modulates VPF/VEGF Receptor-2-mediated Endothelial Cell Proliferation,
but Not Migration, through Phosphatidylinositol 3-Kinase-dependent
Pathways. The Journal of Biological Chemistry 270(29): 26969-26979

STUDI POLA PERGERAKAN, PALATABILITAS, KUALITAS PAKAN, DAN
KECERNAAN NUTRIEN SERTA RISIKO TOKSISITAS LANTADEN PADA
BADAK JAWA DI TAMAN NASIONAL UJUNG KULON
Abstrak
Badak jawa (Rhinoceros sondaicus) merupakan satwa langka yang hanya terdapat di
Taman Nasional Ujung Kulon, Banten Indonesia. Kondisi habitat yang ada saat ini
diduga mengalami perubahan perlahan akibat suksesi alami yang berakibat pada
berubahnya struktur vegetasi yang ada. Sebagai hewan herbivora yang sangat
bergantung pada tumbuhan sebagai sumber makanannya, maka kualitas nutrisi
tumbuhan pakan serta kualitas asupan pakan menjadi bagian yang penting untuk
dipantau dalam populasi saat ini. Penelitian ini bertujuan untuk dapat memetakan
pola pergerakan badak di habitatnya dan mengukur palatabilitas, kualitas pakan,
kecernaan, serta asupan nutrien dan lantaden (toksin) dari badak yang diikuti.
Parameter yang dicatat terdiri dari: jalur lintasan badak (trajektori)beserta korelasinya
dengan keragaman pakan, dan jumlah kubangan. Palatabilitas, kualitas pakan, risiko
toksisitas dari konsumsi Lantana camara, kecernaan nutrien, dan juga ketersediaan
garam. Palatabilitas ditentukan dengan memilih lima jenis pakan dengan jumlah
ragutan terbanyak dari setiap badak. Analisis proksimat digunakan untuk mengukur
kualitas nutrien pakan, dan jumlah asupan nutrien.Toksin lantaden dihitung dengan
mengalikan hasil perhitungan jumlah ragutan (dalam gram) dengan kadar lantaden
dari studi literatur. Kandungan garam diukur dari tanah kubangan dengan metode
titrimetri,sementaraAcid insoluble ash (AIA) digunakan untuk mengukur tingkat
kecernaan bahan kering (BK). Hasil pengamatan menunjukkan korelasi yang kuat
antara ruang jelajah dengan keragaman jenis pakan serta jumlah kubangan. Hasil
pengamatan palatabilitas dan analisis proksimat menunjukkan keragaman jenis pakan,
identifikasi jenis pakan dengan kualitas nutrien tinggi, dan indikasi rendahnya kadar
air, nutrien, energi dari pakan pada bulan-bulan dengan jumlah kejadian hujan yang
rendah (kering) di bulan Oktober 2009 dan Maret 2010. Kadar asupan lantaden berada
pada tingkat yang relatif rendah (23.63 mg/ekor/hari). Upaya untuk memperbaiki
kondisi ini dapat dilakukan dengan pengkayaan jumlah tumbuhan pakan yang
mengandung kualitas nutrien tinggi yang antara lain terdiri dari: Moringa citrifolia,
Callicarpa longifolia, Chisocheton microcarphus, (protein tinggi); Alstonia
angustiloba, Callicarpa longifolia, Macaranga spp, (lemak tinggi); Derris
thyorsifolia, Pterospermum javanicum, Percampyulus glances, (energy tinggi);
Paederia scandens, Alstonia scholaris, Costus speciosus (kandungan air
tinggi).Tumbuhan-tumbuhan ini idealnya ditanam dalam kerapatan 15 tumbuhan per
hektar (untuk jenis pohon) sampai kerapatan 5,406 tumbuhan per hektar (untuk jenis
semai) untuk memastikan ketersediaan yang seimbang.
Kata kunci: badak jawa, palatabilitas, nutrien, kecernaan

48
Study of movement, palatability, feed quality , nutrient digestibility, and risk of
lantadene toxicity in Javan rhinoceros in Ujung Kulon National Park
Abstract
Javan rhinoceros (Rhinoceros sondaicus) is a critically endangered species
living in Ujung Kulon National Park, Banten Indonesia. The condition of the current
habitat is predicted to undergo gradual changes due to natural succession that affects
the structures of vegetations. This species is a large herbivore that relies on the feed
plant type as the source of food, so the quality of these plants, as well as the nutrient
intake become a crucial factor determining the well being of the animal and nutrition
status needs to be included as part of the population monitoring. This research is
designed for mapping the movement patterns (trajectory) and their correlation with
feed diversity and the abundance of the wallow holes. This research also measured
feed palatability, feed quality, nutrient digestibility, the risk of toxin intake from
Lantana camara consumption, as well as NaCl availability. Palatability was
determined by selecting the five feed plants with the highest amount of consumption
from each rhino based on the estimation of browse marks. Proximate analysis was
done to determine the nutrient quality form each feed plant. Lantadene toxin intake
was calculated by multiplying the calculated feed intake (in gram) with toxin contents
based on references. NaCl content was determined using tirimetric method done on
mud samples from wallow sites, while acid insolubel ash (AIA) is used as indicator in
measuring the digestibility. The result showed that feed plants with high palatability
(most liked), identification of feed plants with high nutritional values, and there was
an indication of low water, nutrien, and energy intake at certain times during months
with low rain occurences (dry) in October 2009 and March 2010. Lantaden toxin
intake was relatively small to cause clinical implications to the rhino (23.63 mg/day).
The possible intervention to overcome the situation is to enhance the feed availability
as well as to improve feed quality by replenishing high quality feed plants consisting
of: Moringa citrifolia, Callicarpa longifolia, Chisocheton microcarphus, (high
protein); Alstonia angustiloba, Callicarpa longifolia, Macaranga spp, (high fat);
Derris thyorsifolia, Pterospermum javanicum, Percampyulus glances, (high energy);
Paederia scandens, Alstonia scholaris, Costus speciosus (high water). These plants
should ideally be planted in the density of: 15 plants per hectare (tree forms) to 5,406
plants per hectara (seedling forms) to ensure availability of these plants for the
rhinoceros.
Key words: Javan rhinoceros, palatability, nutrients, digestibility

49
Pendahuluan
Dari seluruh habitat badak yang ada di dunia, Taman Nasional Ujung Kulon
merupakan satu-satunya habitat yang dihuni oleh badak jawa. Walaupun kondisi
ekosistem di Taman Nasional ini bukanlah kondisi yang paling ideal bagi badak jawa
karena kondisi vegetasi yang berubah (suksesi) dan juga kondisi ketersediaan air yang
sangat terbatas di musim kering,dengan demikian, faktor nutrisi merupakan faktor
penting yang perlu dipelajari dari badak jawa. Nutrisi merupakan faktor penentu dari
kemampuan hewan untuk bertahan hidup, beradaptasi, serta bertahan terhadap
cekaman yang muncul dalam kehidupan sehari-hari.
Pengamatan pada badak sumatera (Dicerorhinus sumatrensis) yang dilakukan
oleh Mundiany et al.(2005) menunjukkan bahwa konsumsi berat kering harian setiap
ekor badak berada pada angka 3-4% dari berat badannya dengan kecernaan bahan
kering mencapai 57.49% sampai 80.45%.Badak sumatera menyukai 37 jenis pakan
yang tersedia secara alami di hutan yang antara lain terdiri dari: cakar elang
(Gardenia tubifera), soka merah (Psychotria angulata), sulangkar (Leea sambucina),
waru (Hibiscus tiliaceus), dan kasapan (Croton caudatus). Tumbuhan sulangkar dan
waru merupakan jenis tumbuhan yang juga disukai oleh badak jawa.
Dengan jumlah populasi yang kecil yaitu dalam kisaran 32-47 ekor (Hariyadi
et al. 2011), kemampuan badak jawa untuk bertahan hidup, beradaptasi, mentolerir
cekaman, serta pada akhirnya kemampuan untuk bertahan terhadap penyakit menjadi
faktor yang sangat penting dalam menentukan kemampuan spesies ini untuk terus
bertahan hidup dan bertahan dari kepunahan. Terlebih lagi, faktor nutrisi merupakan
faktor yang sangat penting dalam proses perkembang biakan yang kemampuannya
ditentukan oleh kemampuan sistem tubuh untuk mensintesis hormon reproduksi
(hormon steroid) yang bahan dasarnya diperoleh dari asupan nutrien seperti glukosa
dan asam lemak (Koolman & Röhm, 2001).
Permasalahan yang dihadapai dalam pengelolaan badak jawa adalah
kurangnya data dan informasi mengenai kondisi habitat serta implikasinya pada status
nutrisi pada badak jawa. Rangkaian kegiatan yang dilakukan dalam kajian nutrisi dan
kecernaan ini memberikan tambahan informasi yang antara lain terdiri dari: pola
pergerakan badak yang dicatat oleh pengamat yang mengikuti pergerakan badak dari
hari ke hari, keragaman pakan dari setiap ekor badak, dan persentase asupan nutrien
dari setiap ekor badak. Informasi ini merupakan basis yang digunakan sebagai

50
landasan dan acuan dalam merancang kajian-kajian dan analisis dalam bab-bab
selanjutnya. Palatabilitias, pendugaan jumlah asupan, dan identifikasi antinutrisi
(toksin) merupakan informasi kunci yang dihasilkan dalam kajian penelitian ini.
Tujuan Penelitian
Status nutrisi merupakan kunci keberlangsungan hidup badak jawa di Taman
Nasional Ujung Kulon, namun ironisnya perangkat dan protokol pemantauan yang
memungkinkan pihak pengelola untuk mendapatkan informasi terkait status nutrisi ini
belum tersedia. Mempertimbangkan adanya kebutuhan untuk mengembangkan suatu
protokol pemantauan status nutrisi secara intensif yang dapat diterapkan untuk satwa
langka seperti badak jawa, maka penelitian ini bertujuan untuk menyusun protokol
pemantauan yang memberikan informasi mengenai:
1. Pola pergerakan, ruang jelajah, dan lintasan badak di habitatnya
(trajektori) sebagai basis dari pengamatan asupan pakan yang terjadi di
alam.
2. Keragaman pakan yang digemari (palatabilitas) badak obyek penelitian ini
3. Kualitas dan kuantitas asupan nutrien yang didapat badak jawa
4. Kecernaan bahan kering pakan
5. Ketersediaan garam di lokasi kubangan
6. Risiko toksisitas tumbuhan Lantana camara
Penelitian ini dirancang untuk menguji hipotesis bahwa kondisi habitat di Taman
Nasional Ujung Kulon merupakan daerah dengan daya dukung yang kurang memadai
bagi badak jawa.

51
Bahan dan Metode
Penelitian di Taman Nasional Ujung Kulon dilakukan untuk mempelajari:
komposisi pakan, palatabilitas, konsumsi, dan kecernaan tiga individu badak jawa
(Individu kode: 12, 13, dan 18) berkelamin jantan yang diikuti secara intensif selama
6 hari setiap bulannya dalam periode pengamatan bulan Oktober 2009 sampai April
2010. Penelitian ini menggunakan badak jantan sebagai sampel dengan pertimbangan
sebagai berikut:
1. Badak jantan memiliki pola ruang jelajah yang relatif konsisten sepanjang
waktu dibandingkan betina karena tidak dipengaruhi oleh masa
kebuntingan dan pengasuhan anak (White et al. 2007),maka pola
pergerakan dan profil diet setiap ekor badak yang diamati merupakan satu
hal yang dapat mewakili kondisi normal.
2. Badak jantan memiliki jumlah lebih banyak dibanding badak betina
dengan rasio 3:2 yaitu 16 jantan dan 11 betina (Hariyadi et al. 2011),
sehingga peluang untuk menemukan badak jantan akan lebih tinggi
dibandingkan dengan peluang untuk menemukan badak betina.
3. Badak yang dipilih dalam penelitian ini memiliki ruang jelajah yang datar
dan memungkinkan pengumpulan feses untuk analisis AIA.
Ruang Jelajah Badak Jawa
Tiga individu badak jawa jantan dewasa yang dipilih sebagai contoh dalam
penelitian ini sudah tercatat melalui kegiatan video trap WWF Indonesia periode
April 2008 – Juni 2009 dan setiap individu mewakili daerah konsentrasi yang berbeda
di semenanjung Taman Nasional Ujung Kulon. Berdasarkan penelitian yang
dilakukan oleh Sriyanto et al. (1995), individu 12 mewakili daerah konsentrasi 3
dengan kepadatan tinggi di area selatan, individu 18 mewakili daerah konsentrasi 1
dengan kepadatan sedang di area barat, sementara individu 13 mewakili daerah
konsentrasi 4 dengan kepadatan rendah di area utara. Ketiga daerah ini memiliki
struktur vegetasi dengan kepadatan tumbuhan yang berbeda (7.000 – 17.000
individu/ha untuk tumbuhan bawah; 5.200 – 8.400 individu/ha tumbuhan semai; 650-
1.200 individu/ha tumbuhan pancang; 80-180 individu/ha untuk tumbuhan tiang; dan

52
70-50 individu/ha untuk tumbuhan pohon) seperti ditampilkan pada Tabel 2 yang
disusun berdasarkan analisis vegetasi yang dilakukan oleh Rahmat et al.(2007).
Ketiga individu badak jantan yang digunakan sebagai contoh dalam penelitian ini
ditampilkan dalam Gambar 8. Rahmat et al. (2007) mencatat 231 jenis tumbuhan di
ruang jelajah badak, dan 184 diantaranya (80%) merupakan tumbuhan makanan
badak jawa.
Tabel 2. Jenis vegetasi tumbuhan paling dominan di lokasi ruang jelajah badak jawa.
Badak Vegetasi dominan
Bawah Semai Pancang Tiang Pohon
12 Donax canaeformis Leea sambucina Arenga obtusifolia Arenga obtusifolia Hibiscus tiliaceus
13 D melanochaetis Leea sambucina E polyantha Ardisia humilis B arborescens
18 D melanochaetis E polycntha Ardisia humilis Cerbera manghas Vitex pubescens
Sumber: Rahmat et al. (2007)
Informasi lintasan badak didapatkan dengan cara mengikuti setiap badak
secara langsung di jalur lintasan masing-masing yang telah diidentifikasi sebelumnya.
Untuk menghindari cekaman terhadap badak akibat keberadaan pengamat yang terlalu
dekat dengan badak, maka pengamat mengikuti pergerakan badak dalam jarak yang
cukup jauh yaitu sekitar 24-30 jam di belakang badak yang diikuti. Jarak ini
menempatkan pengamat dalam jarak yang relatif aman dan tidak dapat terdeteksi oleh
badak baik melalui penciuman, pendengaran, maupun penglihatan. Lokasi terjadinya
kontak visual pertama dengan badak dan verifikasi atas identitas badak (kode
individu: 12, 3, dan 18) didefinisikan sebagai titik awal. Pengambilan data
pengamatan dilakukan mulai dari titik awal ini yang kemudian dilanjutkan dengan
penelusuran jejak selama 1-3 bulan (minimal enam hari intensif setiap bulannya)
mengikuti ruang jelajah masing-masing badak. Titik akhir pengamatan didefinisikan
sebagai lokasi yang sama yaitu sampai badak kembali ke titik awal sehingga saat
dipetakan lintasan badak tersebut membentuk suatu lingkaran yang realatif tertutup
(temu gelang).

53
A
B
C
Gambar 8. Tiga ekor badak jantan yang dipilih sebagai obyek penelitian: badak
nomor 12 (A), nomor 13(B), dan nomor 18(C). Foto: Balai Taman
Nasional Ujung Kulon-WWF Indonesia (2010)
Informasi koordinat temuan jejak dan trajektori badak direkam dalam alat GPS
(Global Positioning system) Garmin 76CSX dan kemudian diterjemahkan ke dalam
peta digital untuk menggambarkan ruang jelajah badak secara visual, serta
memastikan bahwa badak telah diikuti secara penuh sampai kembali ke titik awal
(temu gelang) selama 7 bulan pengamatan dari bulan Oktober 2009 sampai bulan
April 2010. Luas ruang jelajah dan jarak tempuh yang ditunjukkan oleh ketiga ekor
badak sampel dibandingkan dengan pola asupan serta nutrisi untuk mendapatkan
korelasi antara keempat parameter di atas.

54
Palatabilitas
Palatabilitas ditentukan dengan memilih jenis tumbuhan pakan dengan
kuantitas terbanyak berdasarkan estimasi ragutan di lokasi tempat makan badak
(rumpang). Lima jenis tumbuhan pakan dengan jumlah ragutan terbanyak dari setiap
ekor badak dikategorikan sebagai tumbuhan dengan palatabilitas tinggi, dan
penggabungan dari tumbuhan palatabilitas tinggi dari setiap ekor badak digabungkan
untuk mewakili palatabilitas dari sampel populasi badak yang diamati dalam
penelitian ini.
Data dikumpulkan dengan cara mengikuti lintasan, mengidentifikasi rumpang
(lahan pakan), mengidentifikasi bekas ragutan untuk memastikan aktifitas makan,
mencatat, dan menimbang bekas makan (feeding marks). Pengamatan seperti ini
mengikuti metode yang pernah dilakukan Birkett & Stevens-Wood (2005) untuk
memberikan informasi tentang komposisi pakan kesukaan dari tiap individu. Gambar
9 menunjukkan kondisi lahan pakan badak yang ada di ekosistem Taman Nasional
Ujung Kulon.
A
B
Gambar 9. Kondisi lahan pakan badak (rumpang) dengan perbandingan tinggi
tumbuhan pakan dan manusia (A) serta kondisi rumpang yang relatif
terbuka dibandingkan areal pepohonan di sekitarnya (B). Foto:
RSetiawan – WWF Indonesia (2009).

55
Tiga kelompok pengamat yang masing-masing terdiri dari lima orang
mencatat jenis tumbuhan yang dimakan oleh badak dan mengukur (dengan
pendugaan) banyaknya tumbuhan pakan yang dikonsumsi oleh badak-badak tersebut.
Pendugaan ini dilakukan dengan memperkirakan banyaknya tumbuhan yang dimakan
badak dan menimbang tumbuhan tersebut untuk menduga berat pakan yang dimakan
oleh masing-masing badak dalam periode pengamatan. Bekas ragutan badak dapat
dibedakan dengan melakukan langkah-langkah sebagai berikut:
1. Mengidentifikasi hewan besar selain badak yang memiliki perilaku makan
mirip dengan badak Jawa. Salah satu hewan ini adalah banteng (Bos
javanicus) yang juga menjelajahi daerah badak dan memakan beberapa
jenis tumbuhan yang sama dengan badak. Membedakan bekas ragutan
badak dengan bekas ragutan hewan lain. Bekas ragutan badak memiliki
karakteristik berupa potongan dahan dan ranting yang mirip seperti
pemotongan dengan menggunakan gunting tumpul. Hal ini terjadi karena
karakteristik anatomi rahang dan perilaku makan badak yang
menggunakan bibir atas (prehensil) untuk menggenggam dahan dan
ranting serta memangkas dahan dan ranting tersebut dengan menggunakan
gigi seri yang berada di rahang bawahnya.
2. Mengikuti pergerakan badak berdasarkan lokasi pakan dan bekas ragutan
untuk mendapatkan informasi mengenai ruang jelajah yang digunakan
setiap ekor badak.
3. Identifikasi dan pendugaan jumlah tumbuhan yang diragut dilakukan
dengan mengamati bekas ragutan, mengambil sebagian tumbuhan dari
ranting lain (sampel) untuk menyamai dengan bekas ragutan, dan
menimbang sampel tersebut dalam keadaan segar, kering jemur, dan
kering akhir.
Kualitas Pakan
Sampel tumbuhan pakan yang telah diidentifikasi dan dipilih kemudiandikirim
ke laboratorium Balai Penelitian Ternak (Balitnak) di Ciawi, Bogor untuk menjalani
analisis proksimat untuk mendapatkan data kandungan air, protein dan lemak dari
setiap jenis tumbuhan pakan. Sampel tumbuhan pakan juga dianalisis dengan bom
kalorimeter untuk mengukur energi yang terkandung.

56
Asupan nutrien dihitung berdasarkan konsumsi kering (KBK) yang dikalikan
dengan jumlah nutrien (protein dan lemak) dan energi hasil dari analisis
proksimat.Asupan nutrien ini disajikan dalam satuan g/hari bagi setiap individu,
sementara asupan energi disajikan dalam satuan kilokalori (kkal) per hari untuk setiap
ekor badak. Informasi tersebut merupakan basis yang digunakan dalam analisis
nutrien dan perbandingan dengan hewan model pada penelitian berikutnya.
Penghitungan kadar air dari tumbuhan pakan (dalam persen) dilakukan dengan
menghitung selisih berat segar dengan berat kering jemur serta menghitung persen
penyusutan yang terjadi untuk menentukan kadar air. Pengukuran berat kering akhir
sampel tumbuhan pakan dilakukan di laboratorium Balitnak Ciawi dan berat kering
akhir ini didapatkan setelah pengeringan dalam oven dengan suhu 105o C sampai
mencapai berat konstan.
Total kadar air tumbuhan pakan kemudian dihitung berdasarkan penjumlahan
dari kadar air antara berat segar dengan berat kering jemur dengan kadar air yang
terukur dari berat kering akhir hasil analisis laboratorium menggunakan metode
gravimetri.
Konsumsi Pakan
Pengamatan dilakukan dengan mengidentifikasi jenis tumbuhan pakan serta
memastikan adanya bekas ragutan. Badak jawa meragut tumbuhan pakan dengan cara
memegang ranting tumbuhan tersebut dengan bibir atas (prehensil) dan kemudian
memotong/memangkas bagian ranting tersebut dengan gigi insisor bawah yang
memiliki bagian tajam. Cara pemangkasan tumbuhan seperti ini memberikan tanda
bekas ragutan yang memiliki ciri sebagai berikut:
1. Tumbuhan menunjukkan adanya bagian yang lebih pendek dibanding
dengan bagian tumbuhan lainnya
2. Dahan atau ranting yang dipangkas menunjukkan adanya bekas potongan
yang halus seperti digunting.
Pendugaan jumlah dan jenistumbuhan yang dikonsumsi oleh badak dilakukan
dengan cara memotong bagian dahan ataupun ranting dari tumbuhan yang sama
sampai kira-kira ke titik bekas ragutan berada. Bagian tumbuhan ini kemudian
ditimbang untuk mendapatkan berat segar dari bagian dahan dan ranting tersebut.
Prosedur ini dilakukan pada bagian tumbuhan lainnya yang menunjukkan adanya

57
bekas ragutan. Total jumlah dahan dan ranting yang ditimbang ini merupakan
estimasi dari jumlah asupan pakan badak dari satu lokasi rumpang (areal pakan).
Pengamat memperhatikan dan memperkirakan usia bekas ragutan yang ada
pada tumbuhan tersebut untuk memastikan bahwa ragutan tersebut merupakan bekas
aktifitas makan yang dilakukan dalam periode pengamatan (maksimum 30 jam yang
lalu) dan bukan merupakan bekas aktifitas makan yang dilakukan oleh badak di luar
periode waktu ini.
Kuantitas asupan berupa konsumsi bahan kering (KBK) ditentukan dengan
menggunakan referensi literatur. Dengan mengacu kepada informasi konsumsi berat
kering pada badak india menurut Clauss et al. (2005) yang berada pada rentang 0.8-
1.3% dari berat badan per hari, maka konsumsi berat kering (KBK) harian pada
badak jawa diasumsikan berada pada tingkat 1% dari berat badan yang merupakan
nilai tengah dalam rentang tersebut. Berat badan badak dalam penelitian ini
diestimasi berdasarkan ukuran jejak yang dibandingkan dengan estimasi usia dan
berat berdasarkan tabel regresi yang disusun oleh Purchase (2007) dan ditentukan
sebagai berikut:
• Badak 12 (dewasa tua>65-70 bulan): 1,000 kg
• Badak 13 (dewasa sekitar >50 tahun): 1,000 kg
• Badak 18 (muda>25 bulan): 700 kg
Langkah-langkah ini menghasilkan informasi mengenai komposisi jenis
tumbuhan yang dikonsumsi, pendugaan jumlah pakan segar dan bahan kering yang
dikonsumsi (dalam kg). Berikut ini metode uji yang digunakan dalam analisis
proksimat untuk mengukur: kadar air (gravimetrik), nutrien protein (Kjeldahl) dan
lemak (ekstraksi dengan sokslet), energi, serat kasar, kalsium, fosfor, AIA, dan tannin
dari sampel pakan yang telah dikeringkan.

58
Metode Uji
Air : IKM 01 (gravimetri)
Protein : IKM 02 (Kjelhdahl destruksi – autoanalisis)
Lemak : IKM 03 (gravimetri dan ekstraksi dengan soxlet)
Energi : IKM 04 (bomb kalorimeter )
Serat Kasar : IKM 05 (ekstraksi asam dan basa)
Abu : IKM 06 (gravimetri)
Ca : IKM 09 (AAS )
P : IKM 10 (spektrofotometri )
AIA : (gravimetri )
Tanin : (Spektrofotometri )
Kecernaan
Pengukuran kecernaan dilakukan dengan metode Van Keulen dan Young
(1977) penghitungan koefisien penyerapan (persen kecernaan) dilakukan dengan
mengukur selisih kandungan AIA (acid insoluble ash) pada asupan pakan dengan
feses. Hal ini sangat cocok untuk diterapkan dalam penelitian satwa liar seperti badak
jawa yang hanya dapat dipelajari di habitat alaminya sehingga metode penghitungan
koefisien kecernaan berdasarkan koleksi feses total (total fecal collection) hampir
tidak mungkin untuk dilakukan.
Pengambilan feses dilakukan dengan tata cara pengambilan yang menjamin
tersedianya sampel feses segar lengkap dengan informasi pendukung yang diperlukan
sebagaimana yang telah dilakukan oleh Fernando et al. (2004). Prosedur
pengumpulan sampel terdiri dari kegiatan-kegiatan di bawah ini:
1. Mencari dan menemukan feses badak jawa yang terbaru atau sesegar
mungkin, dengan cara menyusuri jejak badak terbaru (usia jejak di bawah
tiga hari). Usia jejak dikenali dengan memperhatikan keutuhan jejak,
kesegaran tanah, adanya sisa-sisa daun, dan keberadaan jamur yang tumbuh
di dalam jejak. Usia jejak yang segar biasanya memiliki bentuk yang utuh
sebagaimana ditampilkan dalam Gambar 10 dan tidak terdapat sisa daun
(serasah) ataupun jamur.
2. Memperhatikan letak feses serta memastikan bahwa sampel feses tersebut
tidak berada di aliran air sungai/terapung dikubangan. Perlu dipastikan juga
bahwa gumpalan feses (boli) yang ditemukan berada dalam kondisi utuh
tidak rusak akibat hujan.

59
3. Pengambilan sampel feses dilakukan dengan mengambil beberapa boli feses
dan menyimpannya dalam kantong plastik bersegel (plastik klip)
4. Peneliti mengisi lembar data informasi mengenai temuan contoh feses yang
meliputi; tanggal, waktu, koordinat posisi, lokasi, tipe vegetasi dominan,
temuan tapak dan arahnya
5. Peneliti mencatat informasi detil tentang sampel feses tersebut yang terdiri
dari: kondisi feses (letaknya ternaung atau daerah terbuka, apakah dekat
aliran sungai, keadaan kering atau basah, prakiraan usia, jumlah boli dalam
satu tumpukan, apakah tersebar atau mengumpul, adanya dekomposer, bau
dan rata-rata diameter boli) juga perlu diketahui prakiraan jarak dari feses
sebelumnya, dari aliran sungai, dari kubangan dan dari transek.
6. Peneliti juga mencatat informasi lain seperti: tapak satwa lainnya, adanya
tumpukan feses yang berdekatan tetapi berbeda ukuran, dan lain sebagainya
Gambar 10. Contoh jejak badak yang relatif segar. Badak berada di lokasi ini 12
jam sebelum jejak ditemukan. Foto: Ahariyadi-WWF Indonesia (2007)
Feses badak ditemukan dalam bentuk tumpukan yang terdiri dari beberapa
gumpalan feses berbentuk bola (boli), dan kesegaran feses tersebut dapat ditentukan
dari warna, kelembaban, serta keberadaan serangga yang mengelilinginya. Feses
segar biasanya berwarna coklat kehijauan dengan kelembaban permukaan yang khas
karena masih mengandung lapisan lendir (tidak lebih dari satu jam sejak ekskresi),
serta masih banyak dikelilingi serangga lalat. Dua buah boli dari setiap tumpukan

60
segar diambil sebagai sampel untuk analisis. Sampel ini kemudian ditimbang untuk
mendapatkan berat feses segar.
Keseluruhan sampel feses yang diekskresikan setiap hari oleh individu badak
dalam penelitian ini dikumpulkan dan ditimbang dalam keadaan basah untuk
mendapatkan berat feses segar, dan kemudian dikeringkan dengan menjemurnya di
udara terbuka sampai sampel dapat dilihat lebih kering. Sampel feses tersebut
kemudian ditimbang kembali untuk mendapatkan berat feses kering udara.
Sampel feses untuk mengukur pengeringan diambil dengan cara memilih
tumpukan boli yang relatif paling segar dan utuh dengan memperoleh kualitas sampel
feses yang baik. Maksimal 10% dari jumlah feses yang ada dalam tumpukan tersebut
diambil secara manual, dan sisa tumpukan feses dibiarkan berada di lokasi tersebut
agar tidak mempengaruhi fungsi tumpukan feses dalam penandaaan ruang jelajah
yang dilakukan oleh badak. Penghitungan penyusutan berat kering sampel feses dan
tumbuhan pakan dilakukan dengan menimbang berat basah di lapangan, berat kering
udara setelah mengalami penjemuran di bawah sinar matahari sampai berat menjadi
konstan, dan berat kering akhir setelah mengalami pengeringan dengan oven bersuhu
105o sampai mencapai berat konstan. Dalam kondisi ini sampel siap dianalisis lebih
lanjut di laboratorium untuk pengukuran AIA dari feses dalam kondisi kering akhir.
Hal yang sama juga dilakukan terhadap sampel tumbuhan pakan yang diragut
oleh badak untuk mendapatkan pengukuran AIA dari masing-masing jenis tumbuhan
pakan tersebut.
Metode AIA (Acid Insoluble Ash) Menurut Van Keulen & Young (1977)
Pengukuran AIA dilakukan dengan menggunakan 2 g sampel (pakan dan/atau
feses) yang diabukan pada suhu 600oC. Lalu abu dimasukkan dalam piala gelas dan
ditambah 25 ml HCl 2N dan dididihkan hingga volume awal menjadi kira-kira
setengahnya. Abu disaring ke dalam Crucible (yang sudah diketahui bobotnya).
Endapan dicuci dengan air suling panas (85-100oC) sampai bebas asam. Hasil
saringan diabukan lagi dan kemudian berat abu yang tidak larut dalam asam hasil
proses ini ditimbang dan dihitung dengan menggunakan rumus yang berlaku untuk
feses dan pakan sebagai berikut:
Persen AIA = berat abu yang tidak larut dalam asam x 100%
Berat sampel

61
Sementara dengan banyaknya AIA pada fese, maka penghitungan kecernaan berat
kering dilakukan dengan menggunakan rumus:
AIA feses – AIA pakan yang dikonsumsi x 100%
AIA feses
Analisis AIA ini dikoreksi dengan menggunakan faktor koreksi sebesar 10%
seperti yang digunakan dalam beberapa analisis kecernaan menggunakan metode AIA
(Mainka et al. 1989; Sims et al. 2007). Faktor koreksi ini perlu digunakan dalam
menghitung kecernaan berat kering mengingat adanya variasai akibat proses dehidrasi
saat konversi dari asupan segar menjadi asupan kering (Sims et al. 2007). Mainka et
al. (1989) menggunakan angka 10% sebagai faktor koreksi untuk memperhitungkan
dehidrasi.
Risiko Toksisitas dari Konsumsi Lantana camara
Kandungan antinutrisi lantaden dalam setiap gram kering tumbuhan Lantana
camara dihitung berdasarakan informasi yang disampaikan oleh Sharma et al.(2000)
yang mengidentifikasi adanya lantaden dari berat kering tumbuhan Lantana camara.
Kandungan lantaden dalam tumbuhan Lantana kering didefinisikan sebagai jumlah
lantaden yang telah diukur dalam studi laboratorium yaitu: 1,930.4 mg/100g Lantana
kering. Penghitungan asupan lantaden dilakukan dengan mengukur berat kering
konsumsi harian Lantana camara pada setiap ekor badak dan mengalikan jumlah
tersebut dengan kandungan Lantaden. Perhitungan tersebut dikompilasi dalam rumus
sebagai berikut:
Asupan lantanden = konsumsi bahan kering x kandungan lantaden
(mg/ekor/hari) (gram kering/ekor/hari) (mg/100g)
100
Ketersediaan Garam
Empat kubangan di wilayah jelajah ketiga ekor badak dipilih berdasarkan
status kubangan yang merupakan kubangan aktif (masih digunakan oleh badak paling
tidak dalam 1 bulan terakhir). Untuk memastikan ketersediaan kandungan mineral
NaCl dalam tanah di sekitar kubangan, maka sampel tanah dari beberapa kubangan di
wilayah jelajah ketiga ekor badak tersebut dikumpulkan dengan menggunakan pipa

62
PVC berukuran diameter 1.75 cm (0.5 inci) yang ditancapkan sedalam 10 sentimeter
ke dalam lumpur di tepi kubangan. Lumpur yang mengisi pipa tersebut didorong
keluar dan dikumpulkan dalam kantong plastik untuk dianalisis lebih lanjut dengan
metode titrimetrik di laboratorium SUCOFINDO Cilegon Banten.
Analisis Data
Data disajikan secara deskriptif dan korelasi antara jarak tempuh dengan keragaman
pakan yang tersedia dianalisis menggunakan koefisien korelasi Pearson.

63
Hasil dan Pembahasan
Karakteristik Ruang Jelajah Badak Jawa
Penyusuran jejak dengan mengikuti jalur lintasan badak (trajektori)
menunjukkan bahwa ketiga badak jantan yang dipilih sebagai obyek penelitian
memiliki jarak dan ruang jelajah yang berbeda. Perbedaan ini disebabkan adanya
perbedaan ketersediaan pakan, air, dan kubangan. Bentuk ruang jelajah dari masing-
masing badak disajikan dalam Gambar 11. Hubungan antara luas ruang jelajah
dengan kondisi habitat ini dipertegas dengan adanya korelasi positif yang kuat (R=
0.9971) antara luas ruang jelajah badak dalam hektar dengan keragaman asupan pakan
(jumlah pakan yang dimakan) serta korelasi positif yang kuat pula (R = 0.9998) antara
luas ruang jelajah tersebut dengan jumlah kubangan yang digunakan (15 kubangan di
wilayah badak nomor 12; 25 kubangan di wilayah badak nomor 18; 33 kubangan di
wilayah badak nomor 13). Dari data yang ada dapat dilihat bahwa individu 12 yang
berada pada daerah yang memiliki kepadatan populasi tinggi di daerah selatan
semenanjung memiliki ruang jelajah yang lebih sempit (169 ha), sementara individu
13 dan 18 yang masing-masing berada di daerah dengan kepadatan populasi rendah
dan sedang memiliki ruang jelajah yang lebih luas (974 ha dan 631 ha).
Pengamatan di lapangan menunjukkan bahwa pada area dengan konsentrasi
kepadatan tinggi di area selatan (individu no 12) badak cenderung memiliki ruang
jelajah yang kecil, sementara pada area dengan konsentrasi kepadatan rendah di utara
(individu 13) badak menunjukkan ruang jelajah yang relatif besar. Berdasarkan
catatan ini dapat dilihat adanya indikasi bahwa tingkat kepadatan populasi (kualitatif)
memiliki korelasi negatif dengan luasnya ruang jelajah.
Dinerstein (2003) mencatat dari spesies badak di India dan Nepal (Rhinoceros
unicornis) bahwa badak jantan cenderung untuk memiliki ruang jelajah yang tidak
saling bersinggungan dengan pejantan lainnya. Sebaliknya, badak betina
menunjukkan persinggungan yang cukup besar dengan badak jantan. Dinerstein
(2003) juga mencatat bahwa ruang jelajah badak cenderung berubah menjadi lebih
besar pada musim kering dan berubah menjadi lebih sempit di musim hujan.
Berdasarkan informasi ini, maka pada kondisi badak jawa dalam penelitian ini ada
dua faktor yang mempengaruhi ruang jelajah badak yaitu faktor teritorial dan faktor
ketersediaan sumber daya pakan, dan kemungkinan situasi yang terjadi pada badak
jawa:
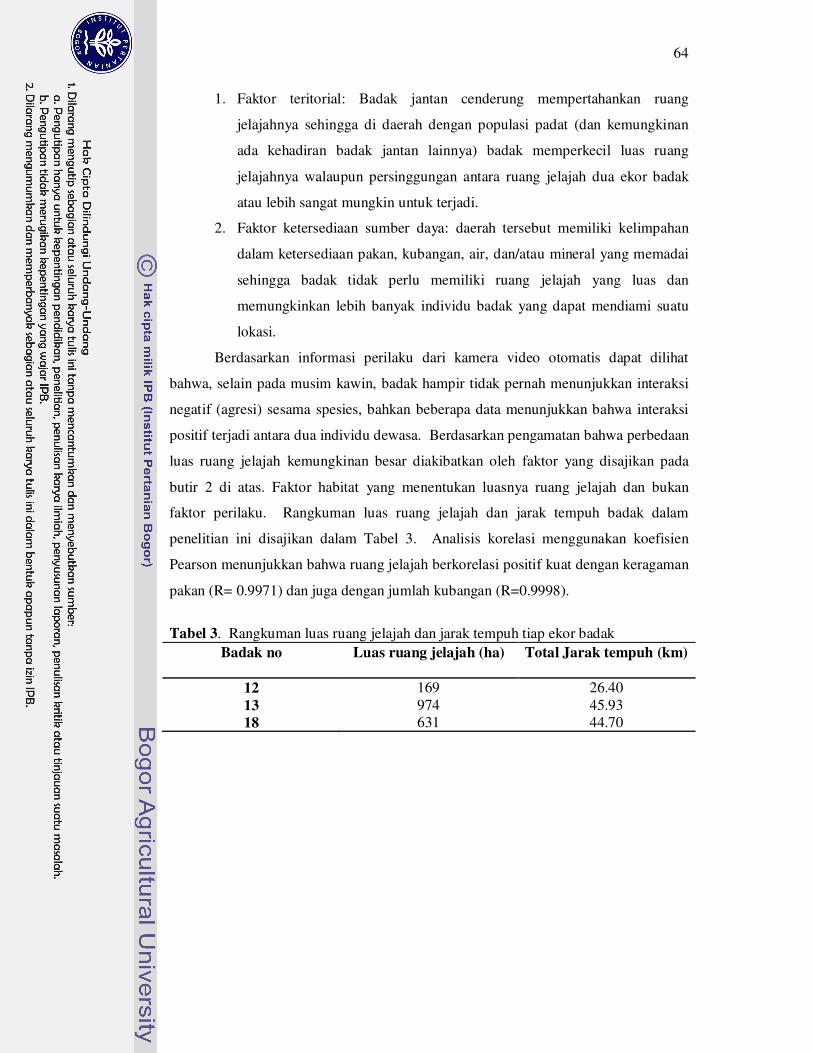
64
1. Faktor teritorial: Badak jantan cenderung mempertahankan ruang
jelajahnya sehingga di daerah dengan populasi padat (dan kemungkinan
ada kehadiran badak jantan lainnya) badak memperkecil luas ruang
jelajahnya walaupun persinggungan antara ruang jelajah dua ekor badak
atau lebih sangat mungkin untuk terjadi.
2. Faktor ketersediaan sumber daya: daerah tersebut memiliki kelimpahan
dalam ketersediaan pakan, kubangan, air, dan/atau mineral yang memadai
sehingga badak tidak perlu memiliki ruang jelajah yang luas dan
memungkinkan lebih banyak individu badak yang dapat mendiami suatu
lokasi.
Berdasarkan informasi perilaku dari kamera video otomatis dapat dilihat
bahwa, selain pada musim kawin, badak hampir tidak pernah menunjukkan interaksi
negatif (agresi) sesama spesies, bahkan beberapa data menunjukkan bahwa interaksi
positif terjadi antara dua individu dewasa. Berdasarkan pengamatan bahwa perbedaan
luas ruang jelajah kemungkinan besar diakibatkan oleh faktor yang disajikan pada
butir 2 di atas. Faktor habitat yang menentukan luasnya ruang jelajah dan bukan
faktor perilaku. Rangkuman luas ruang jelajah dan jarak tempuh badak dalam
penelitian ini disajikan dalam Tabel 3. Analisis korelasi menggunakan koefisien
Pearson menunjukkan bahwa ruang jelajah berkorelasi positif kuat dengan keragaman
pakan (R= 0.9971) dan juga dengan jumlah kubangan (R=0.9998).
Tabel 3. Rangkuman luas ruang jelajah dan jarak tempuh tiap ekor badak
Badak no Luas ruang jelajah (ha) Total Jarak tempuh (km)
12 169 26.40
13 974 45.93
18 631 44.70
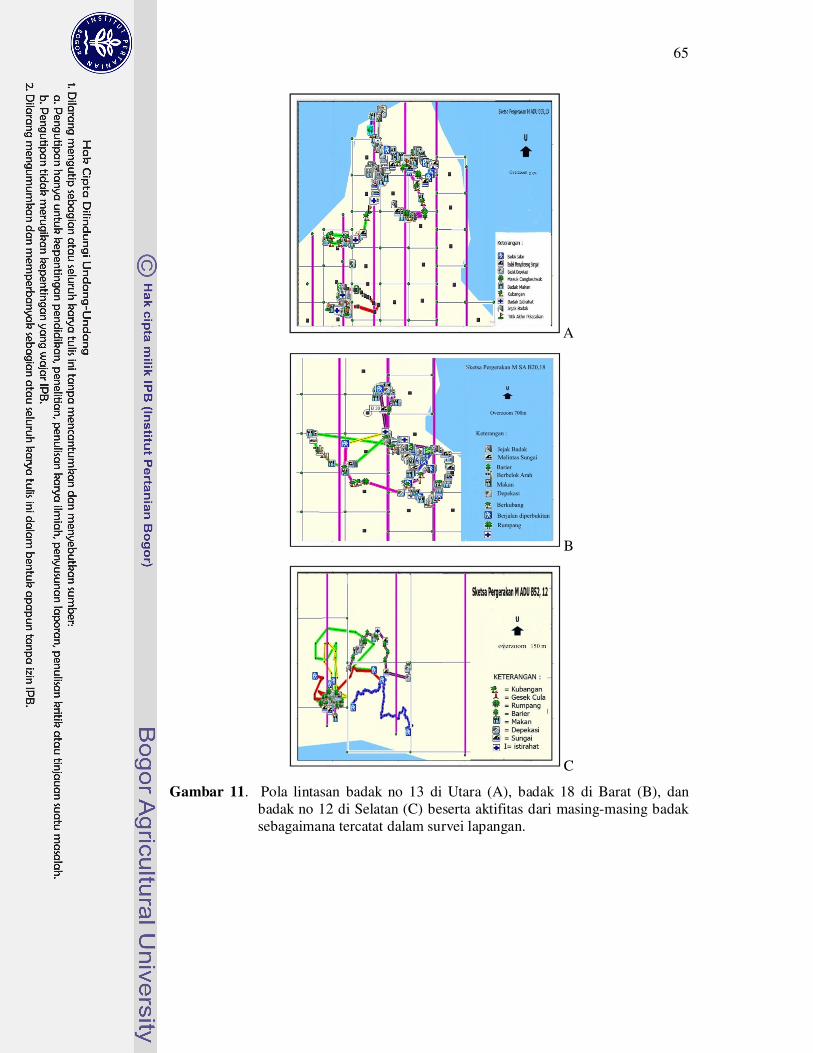
65
A
B
C
Gambar 11. Pola lintasan badak no 13 di Utara (A), badak 18 di Barat (B), dan
badak no 12 di Selatan (C) beserta aktifitas dari masing-masing badak
sebagaimana tercatat dalam survei lapangan.

66
Palatabilitas dan Keragaman Pakan
Beberapa contoh tumbuhan pakan badak jawa ditampilkan dalam Gambar 12
untuk menunjukkan jenis tumbuhan pakan berupa tumbuhan semak dengan daun
lebar.Kompilasi catatan tumbuhan pakan yang dikomsumsi oleh badak selama masa
pengamatan terdiri dari 108 jenis tumbuhan, dan 94 dari 108 jenis tumbuhan ini
dipilih sebagai perwakilan dari jenis tumbuhan pakan tersebut untuk menjalani
analisis proksimat di Balai Penelitian Ternak (Balitnak) Ciawi, Bogor. Hasil analisis
proksimat tumbuhan pakan ini disajikan dalam Lampiran 2. Selain ruang jelajah yang
berbeda, setiap badak menunjukkan variasi palatabilitas jenis pakan yang berbeda-
beda pula sebagaimana ditunjukkan dalam Tabel 4. Perbandingan jenis tumbuhan
palatabilitas tinggi dengan data kelimpahan pakan yang dikumpulkan oleh Rahmat et
al. (2007) menunjukkan adanya indikasi keterkaitan antara tingkat palatabilitas
tumbuhan pakan dengan kelimpahannya di lokasi tersebut. Tumbuhan dengan
palatabilitas tinggi biasanya secara proporsional memiliki kelimpahan yang tinggi di
lokasi tersebut.
A B C
Gambar 12. Beberapa contoh tumbuhan pakan badak Jawa yang tercatat dalam
pengamatan di lapangan. Foto di atas menunjukkan jenis tumbuhan
tepus: Amomum sp (A), Rotan: Calamus sp (B), dan Mara: Mallotus
floribundus (C). Foto: Rsetiawan – WWF Indonesia (2009)
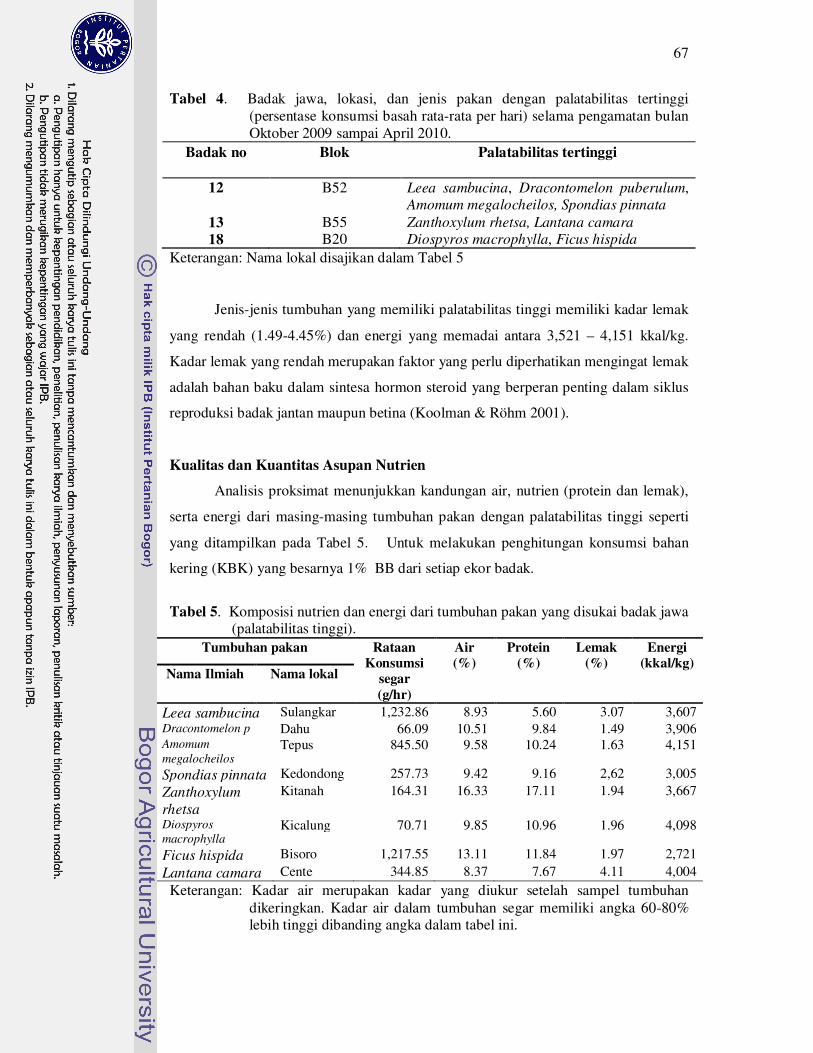
67
Tabel 4. Badak jawa, lokasi, dan jenis pakan dengan palatabilitas tertinggi
(persentase konsumsi basah rata-rata per hari) selama pengamatan bulan
Oktober 2009 sampai April 2010.
Badak no Blok Palatabilitas tertinggi
12 B52 Leea sambucina, Dracontomelon puberulum,
Amomum megalocheilos, Spondias pinnata
13 B55 Zanthoxylum rhetsa, Lantana camara
18 B20 Diospyros macrophylla, Ficus hispida
Keterangan: Nama lokal disajikan dalam Tabel 5
Jenis-jenis tumbuhan yang memiliki palatabilitas tinggi memiliki kadar lemak
yang rendah (1.49-4.45%) dan energi yang memadai antara 3,521 – 4,151 kkal/kg.
Kadar lemak yang rendah merupakan faktor yang perlu diperhatikan mengingat lemak
adalah bahan baku dalam sintesa hormon steroid yang berperan penting dalam siklus
reproduksi badak jantan maupun betina (Koolman & Röhm 2001).
Kualitas dan Kuantitas Asupan Nutrien
Analisis proksimat menunjukkan kandungan air, nutrien (protein dan lemak),
serta energi dari masing-masing tumbuhan pakan dengan palatabilitas tinggi seperti
yang ditampilkan pada Tabel 5. Untuk melakukan penghitungan konsumsi bahan
kering (KBK) yang besarnya 1% BB dari setiap ekor badak.
Tabel 5. Komposisi nutrien dan energi dari tumbuhan pakan yang disukai badak jawa
(palatabilitas tinggi).
Tumbuhan pakan Rataan
Konsumsi
segar
(g/hr)
Air
(%)
Protein
(%)
Lemak
(%)
Energi
(kkal/kg) Nama Ilmiah Nama lokal
Leea sambucina Sulangkar 1,232.86 8.93 5.60 3.07 3,607
Dracontomelon p Dahu 66.09 10.51 9.84 1.49 3,906 Amomum
megalocheilos Tepus 845.50 9.58 10.24 1.63 4,151
Spondias pinnata Kedondong 257.73 9.42 9.16 2,62 3,005
Zanthoxylum
rhetsa
Kitanah 164.31 16.33 17.11 1.94 3,667
Diospyros
macrophylla Kicalung 70.71 9.85 10.96 1.96 4,098
Ficus hispida Bisoro 1,217.55 13.11 11.84 1.97 2,721
Lantana camara Cente 344.85 8.37 7.67 4.11 4,004
Keterangan: Kadar air merupakan kadar yang diukur setelah sampel tumbuhan
dikeringkan. Kadar air dalam tumbuhan segar memiliki angka 60-80%
lebih tinggi dibanding angka dalam tabel ini.
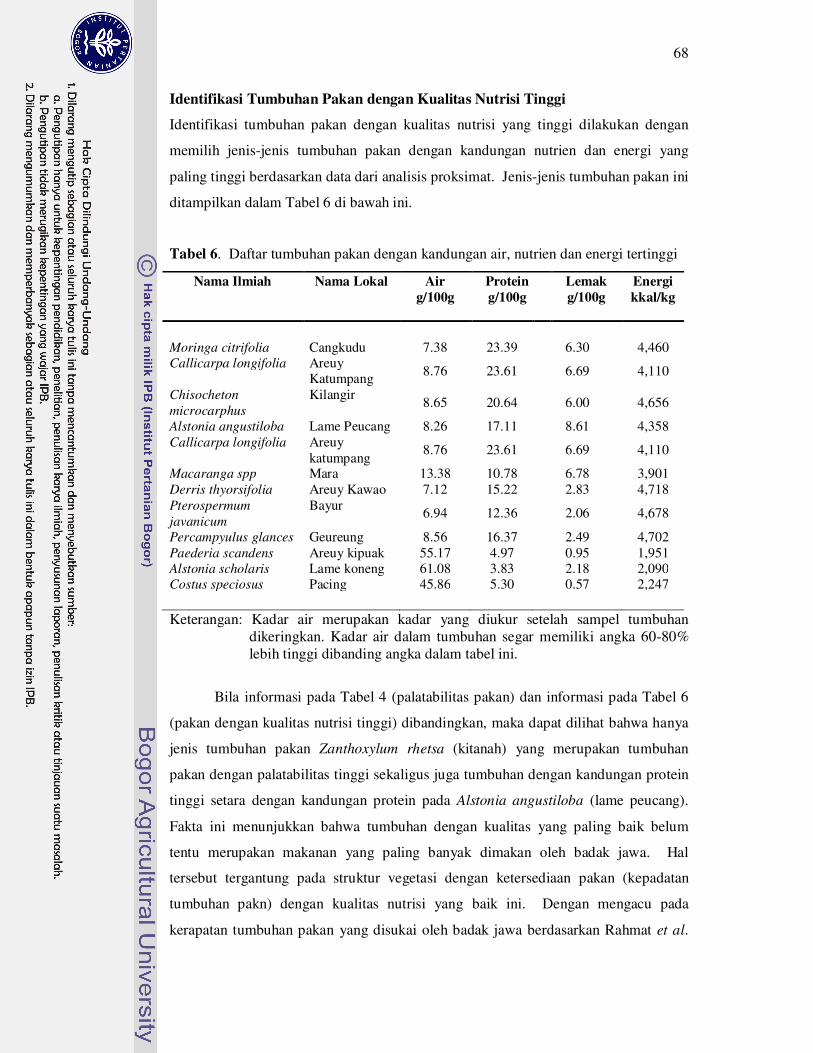
68
Identifikasi Tumbuhan Pakan dengan Kualitas Nutrisi Tinggi
Identifikasi tumbuhan pakan dengan kualitas nutrisi yang tinggi dilakukan dengan
memilih jenis-jenis tumbuhan pakan dengan kandungan nutrien dan energi yang
paling tinggi berdasarkan data dari analisis proksimat. Jenis-jenis tumbuhan pakan ini
ditampilkan dalam Tabel 6 di bawah ini.
Tabel 6. Daftar tumbuhan pakan dengan kandungan air, nutrien dan energi tertinggi
Nama Ilmiah Nama Lokal Air
g/100g
Protein
g/100g
Lemak
g/100g
Energi
kkal/kg
Moringa citrifolia Cangkudu 7.38 23.39 6.30 4,460
Callicarpa longifolia Areuy
Katumpang 8.76 23.61 6.69 4,110
Chisocheton
microcarphus
Kilangir 8.65 20.64 6.00 4,656
Alstonia angustiloba Lame Peucang 8.26 17.11 8.61 4,358
Callicarpa longifolia Areuy
katumpang 8.76 23.61 6.69 4,110
Macaranga spp Mara 13.38 10.78 6.78 3,901
Derris thyorsifolia Areuy Kawao 7.12 15.22 2.83 4,718
Pterospermum
javanicum
Bayur 6.94 12.36 2.06 4,678
Percampyulus glances Geureung 8.56 16.37 2.49 4,702
Paederia scandens Areuy kipuak 55.17 4.97 0.95 1,951
Alstonia scholaris Lame koneng 61.08 3.83 2.18 2,090
Costus speciosus Pacing 45.86 5.30 0.57 2,247
Keterangan: Kadar air merupakan kadar yang diukur setelah sampel tumbuhan
dikeringkan. Kadar air dalam tumbuhan segar memiliki angka 60-80%
lebih tinggi dibanding angka dalam tabel ini.
Bila informasi pada Tabel 4 (palatabilitas pakan) dan informasi pada Tabel 6
(pakan dengan kualitas nutrisi tinggi) dibandingkan, maka dapat dilihat bahwa hanya
jenis tumbuhan pakan Zanthoxylum rhetsa (kitanah) yang merupakan tumbuhan
pakan dengan palatabilitas tinggi sekaligus juga tumbuhan dengan kandungan protein
tinggi setara dengan kandungan protein pada Alstonia angustiloba (lame peucang).
Fakta ini menunjukkan bahwa tumbuhan dengan kualitas yang paling baik belum
tentu merupakan makanan yang paling banyak dimakan oleh badak jawa. Hal
tersebut tergantung pada struktur vegetasi dengan ketersediaan pakan (kepadatan
tumbuhan pakn) dengan kualitas nutrisi yang baik ini. Dengan mengacu pada
kerapatan tumbuhan pakan yang disukai oleh badak jawa berdasarkan Rahmat et al.

69
(2007), maka kerapatan masing-masing tumbuhan pakan berkualitas baik dalam Tabel
6 idealnya tersedia dalam kerapatan: 5,406 individu/ha (jenis tumbuhan bawah); 2,222
individu/ha untuk jenis semai ; 268 individu/ha untuk jenis pancang ; 32 individu/ha
untuk jenis tiang ; dan 15 individu per hektar untuk jenis pohon.
Konsumsi bahan kering berbanding lurus dengan asupan air, nutrien (lemak
dan protein), serta energi. Data asupan harian dari komponen-komponen yang terdiri
dari air, nutrien (protein & lemak), serta energi disajikan dalam Tabel 7. Data dari
Tabel ini menunjukkan bahwa ada fluktuasi asupan harian yang menyebabkan asupan
air, nutrient, dan energi tidak selalu tersedia dalam jumlah yang besar sepanjang
tahun.
Asupan air dari pakan berada pada tingkat yang rendah pada bulan November
(setelah musim kering di bulan Oktober); asupan protein menunjukkan titik terendah
di bulan Oktober (musim kering); asupan lemak menunjukkan titik terendah pada
bulan-bulan Oktober, November, Februari, Maret; dan asupan energi dapat dilihat ada
pada tingkat terendah pada bulan Desember. Fakta ini menunjukkan bahwa populasi
badak jawa menghadapi waktu-waktu tertentu saat terjadi keterbatasan air, nutrien,
dan energi.
Kompilasi perhitungan total konsumsi air, nutrien, serta energi dari setiap
badak sampel penelitian ini disajikan dalam Tabel 8. Analisis deskriptif pada rataan
asupan bahan kering harian pada ketiga ekor badak menunjukkan adanya perbedaan
antara badak 18 dengan kedua ekor badak lainnya (badak 12 dan 13). Hal ini
disebabkan karena perbedaan bobot tubuh antara badak dewasa (badak 12 dan badak
13) dengan badak muda (badak 18) yang memiliki bobot badan lebih kecil.
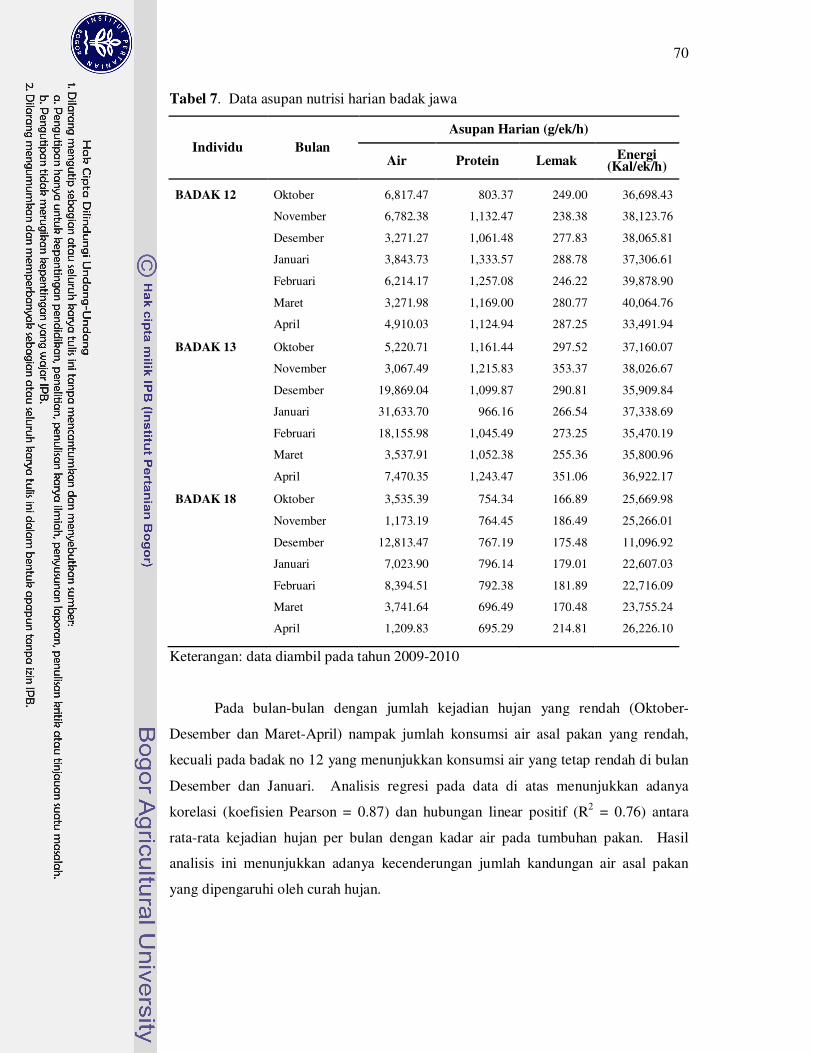
70
Tabel 7. Data asupan nutrisi harian badak jawa
Individu Bulan
Asupan Harian (g/ek/h)
Air Protein Lemak Energi (Kal/ek/h)
BADAK 12 Oktober
6,817.47
803.37
249.00
36,698.43
November
6,782.38
1,132.47
238.38
38,123.76
Desember
3,271.27
1,061.48
277.83
38,065.81
Januari
3,843.73
1,333.57
288.78
37,306.61
Februari
6,214.17
1,257.08
246.22
39,878.90
Maret
3,271.98
1,169.00
280.77
40,064.76
April
4,910.03
1,124.94
287.25
33,491.94
BADAK 13 Oktober
5,220.71
1,161.44
297.52
37,160.07
November
3,067.49
1,215.83
353.37
38,026.67
Desember
19,869.04
1,099.87
290.81
35,909.84
Januari
31,633.70
966.16
266.54
37,338.69
Februari
18,155.98
1,045.49
273.25
35,470.19
Maret
3,537.91
1,052.38
255.36
35,800.96
April
7,470.35
1,243.47
351.06
36,922.17
BADAK 18 Oktober
3,535.39
754.34
166.89
25,669.98
November
1,173.19
764.45
186.49
25,266.01
Desember
12,813.47
767.19
175.48
11,096.92
Januari
7,023.90
796.14
179.01
22,607.03
Februari
8,394.51
792.38
181.89
22,716.09
Maret
3,741.64
696.49
170.48
23,755.24
April
1,209.83
695.29
214.81
26,226.10
Keterangan: data diambil pada tahun 2009-2010
Pada bulan-bulan dengan jumlah kejadian hujan yang rendah (Oktober-
Desember dan Maret-April) nampak jumlah konsumsi air asal pakan yang rendah,
kecuali pada badak no 12 yang menunjukkan konsumsi air yang tetap rendah di bulan
Desember dan Januari. Analisis regresi pada data di atas menunjukkan adanya
korelasi (koefisien Pearson = 0.87) dan hubungan linear positif (R2 = 0.76) antara
rata-rata kejadian hujan per bulan dengan kadar air pada tumbuhan pakan. Hasil
analisis ini menunjukkan adanya kecenderungan jumlah kandungan air asal pakan
yang dipengaruhi oleh curah hujan.
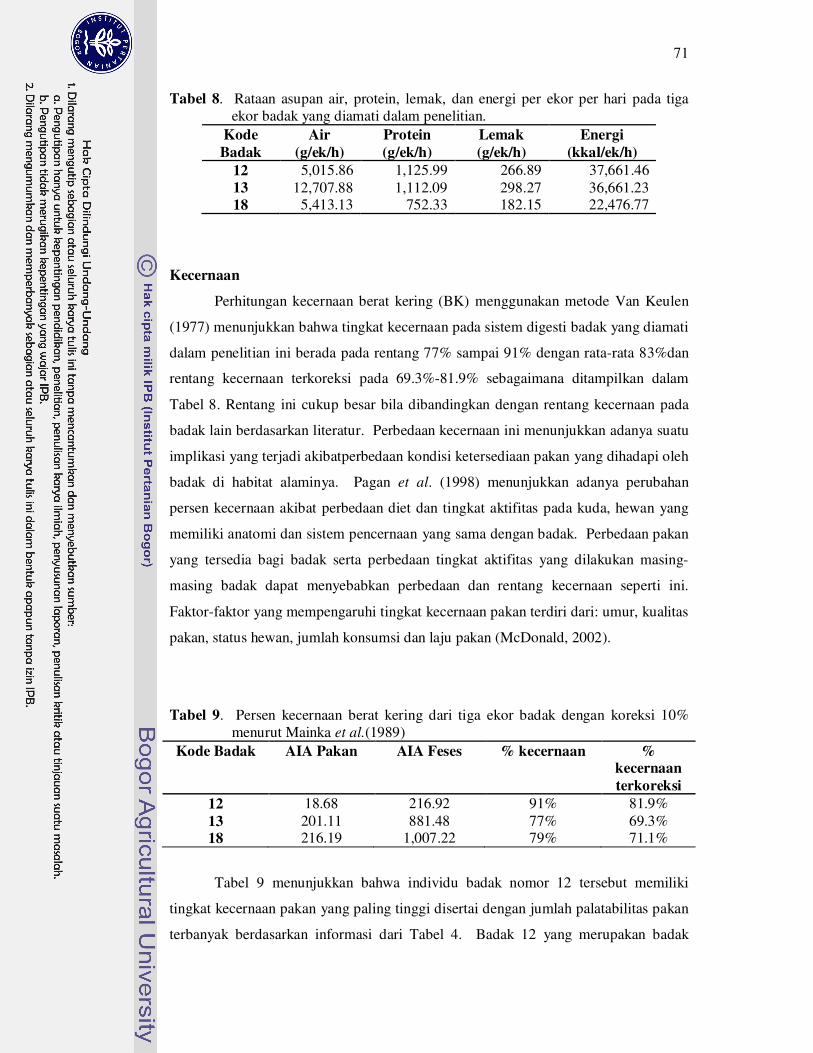
71
Tabel 8. Rataan asupan air, protein, lemak, dan energi per ekor per hari pada tiga
ekor badak yang diamati dalam penelitian.
Kode
Badak
Air
(g/ek/h)
Protein
(g/ek/h)
Lemak
(g/ek/h)
Energi
(kkal/ek/h)
12 5,015.86 1,125.99 266.89 37,661.46
13 12,707.88 1,112.09 298.27 36,661.23
18 5,413.13 752.33 182.15 22,476.77
Kecernaan
Perhitungan kecernaan berat kering (BK) menggunakan metode Van Keulen
(1977) menunjukkan bahwa tingkat kecernaan pada sistem digesti badak yang diamati
dalam penelitian ini berada pada rentang 77% sampai 91% dengan rata-rata 83%dan
rentang kecernaan terkoreksi pada 69.3%-81.9% sebagaimana ditampilkan dalam
Tabel 8. Rentang ini cukup besar bila dibandingkan dengan rentang kecernaan pada
badak lain berdasarkan literatur. Perbedaan kecernaan ini menunjukkan adanya suatu
implikasi yang terjadi akibatperbedaan kondisi ketersediaan pakan yang dihadapi oleh
badak di habitat alaminya. Pagan et al. (1998) menunjukkan adanya perubahan
persen kecernaan akibat perbedaan diet dan tingkat aktifitas pada kuda, hewan yang
memiliki anatomi dan sistem pencernaan yang sama dengan badak. Perbedaan pakan
yang tersedia bagi badak serta perbedaan tingkat aktifitas yang dilakukan masing-
masing badak dapat menyebabkan perbedaan dan rentang kecernaan seperti ini.
Faktor-faktor yang mempengaruhi tingkat kecernaan pakan terdiri dari: umur, kualitas
pakan, status hewan, jumlah konsumsi dan laju pakan (McDonald, 2002).
Tabel 9. Persen kecernaan berat kering dari tiga ekor badak dengan koreksi 10%
menurut Mainka et al.(1989)
Kode Badak
AIA Pakan AIA Feses % kecernaan %
kecernaan
terkoreksi
12 18.68 216.92 91% 81.9%
13 201.11 881.48 77% 69.3%
18 216.19 1,007.22 79% 71.1%
Tabel 9 menunjukkan bahwa individu badak nomor 12 tersebut memiliki
tingkat kecernaan pakan yang paling tinggi disertai dengan jumlah palatabilitas pakan
terbanyak berdasarkan informasi dari Tabel 4. Badak 12 yang merupakan badak

72
dewasa tua dengan ruang jelajah terkecil memiliki palatabilitas jenis pakan yang lebih
beragam dibandingkan dengan badak 13 dan badak 18. Selain faktor kondisi habitat,
kecernaan juga dipengaruhi faktor usia, fisiologi, dan juga genetika dari hewan yang
diamati.
Kecernaan terkoreksi yang dihitung dengan metode AIA dalam penelitian ini
(69.3%-81.9%) dapat dibandingkan dengan kecernaan pada badak sumatra (Koleksi
total): 57.49% - 80.45% (Mundiany et al. 2005), dan perbandingan ini menunjukkan
bahwa ada kemiripan dalam hal kecernaan berdasarkan kedua metode ini yaitu pada
pada rentang kecernaan berat kering 69.3%-80.45%. Informasi ini menunjukkan
kemungkinan adanya kompatibilitas antara kedua metode (AIA dan koleksi total)
dalam menghitung kecernaan pada badak. Kecernaan pada badak jawa berada pada
rentang yang sebanding dengan perhitungan kecernaan pada badak Sumatra
menggunakan koleksi total. Fakta ini menunjukkan bahwa perhitungan kecernaan
menggunakan metode AIA adalah kompatibel dengan perhitungan kecernaan
menggunakan metode koleksi total.Pengukuran dengan metode AIA biasanya
mempunyai nilai lebih tinggi dibandingkan dengan metode koleksi total, hal ini
disebabkan saat pengambilan sampel feses secara kualitatif.
Kajian RisikoToksisitas Lantaden
Berdasarkan jumlah asupan Lantana camara yang tercatat di lapangan,
estimasi asupan lantaden perhari pada ketiga ekor badak dalam penelitian ini disajikan
dalam Tabel 10 di bawah ini. Rataan asupan Lantana camara dari Tabel 10 ini
menunjukkan bahwa badak mengkonsumsi lantana kering: 122.43 g/ekor/hari atau
setara dengan asupan lantaden sebesar rataan 23.63 g/ek/hari. McSweeney & Pass
(1983) menunjukkan bahwa jumlah asupan Lantana camara dengan dosis 4g/kg berat
badan yang setara dengan konsumsi Lantana kering sebanyak 4,000 g/ek/hari pada
badak berbobot 1 ton (dosis lantanden 772.16 g/ek/hari) akan menyebabkan
kerusakan pada hati yang ditunjukkan dengan gejala klinis berupa ruminal stasis
seperti yang terjadi pada domba. Asupan yang terjadi pada badak masih lebih kecil
dibandingkan dengan dosis ini.
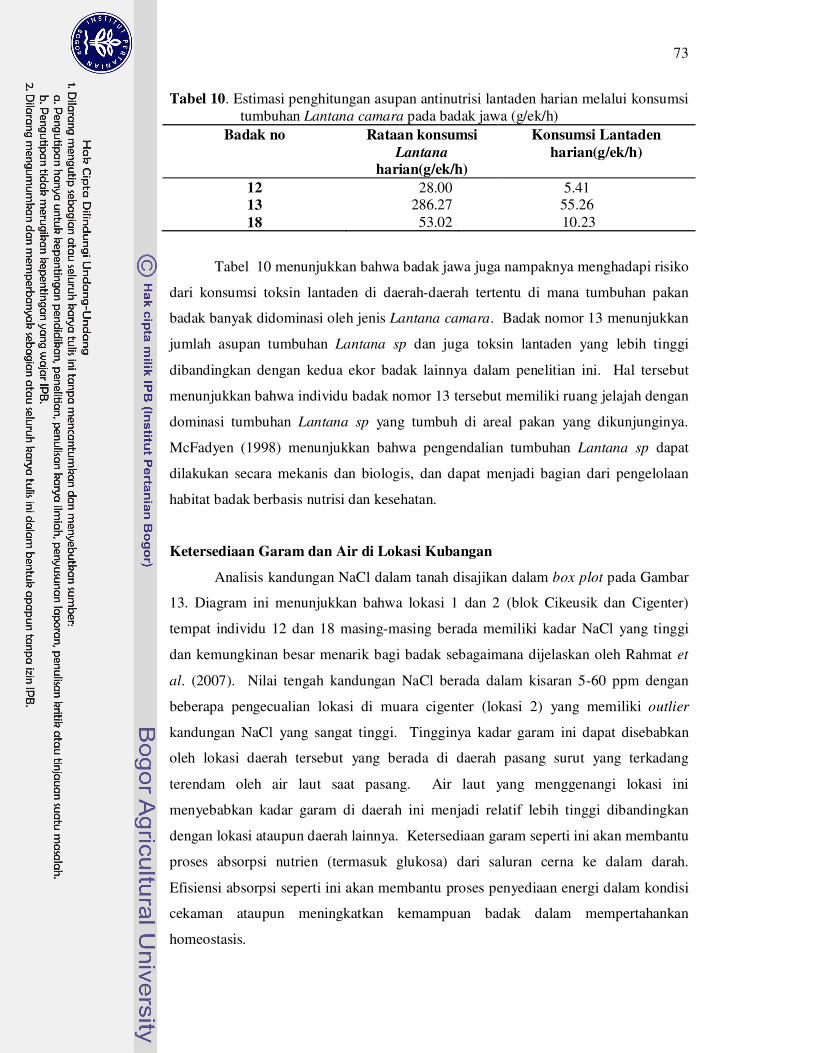
73
Tabel 10. Estimasi penghitungan asupan antinutrisi lantaden harian melalui konsumsi
tumbuhan Lantana camara pada badak jawa (g/ek/h)
Badak no Rataan konsumsi
Lantana
harian(g/ek/h)
Konsumsi Lantaden
harian(g/ek/h)
12 28.00 5.41
13 286.27 55.26
18 53.02 10.23
Tabel 10 menunjukkan bahwa badak jawa juga nampaknya menghadapi risiko
dari konsumsi toksin lantaden di daerah-daerah tertentu di mana tumbuhan pakan
badak banyak didominasi oleh jenis Lantana camara. Badak nomor 13 menunjukkan
jumlah asupan tumbuhan Lantana sp dan juga toksin lantaden yang lebih tinggi
dibandingkan dengan kedua ekor badak lainnya dalam penelitian ini. Hal tersebut
menunjukkan bahwa individu badak nomor 13 tersebut memiliki ruang jelajah dengan
dominasi tumbuhan Lantana sp yang tumbuh di areal pakan yang dikunjunginya.
McFadyen (1998) menunjukkan bahwa pengendalian tumbuhan Lantana sp dapat
dilakukan secara mekanis dan biologis, dan dapat menjadi bagian dari pengelolaan
habitat badak berbasis nutrisi dan kesehatan.
Ketersediaan Garam dan Air di Lokasi Kubangan
Analisis kandungan NaCl dalam tanah disajikan dalam box plot pada Gambar
13. Diagram ini menunjukkan bahwa lokasi 1 dan 2 (blok Cikeusik dan Cigenter)
tempat individu 12 dan 18 masing-masing berada memiliki kadar NaCl yang tinggi
dan kemungkinan besar menarik bagi badak sebagaimana dijelaskan oleh Rahmat et
al. (2007). Nilai tengah kandungan NaCl berada dalam kisaran 5-60 ppm dengan
beberapa pengecualian lokasi di muara cigenter (lokasi 2) yang memiliki outlier
kandungan NaCl yang sangat tinggi. Tingginya kadar garam ini dapat disebabkan
oleh lokasi daerah tersebut yang berada di daerah pasang surut yang terkadang
terendam oleh air laut saat pasang. Air laut yang menggenangi lokasi ini
menyebabkan kadar garam di daerah ini menjadi relatif lebih tinggi dibandingkan
dengan lokasi ataupun daerah lainnya. Ketersediaan garam seperti ini akan membantu
proses absorpsi nutrien (termasuk glukosa) dari saluran cerna ke dalam darah.
Efisiensi absorpsi seperti ini akan membantu proses penyediaan energi dalam kondisi
cekaman ataupun meningkatkan kemampuan badak dalam mempertahankan
homeostasis.
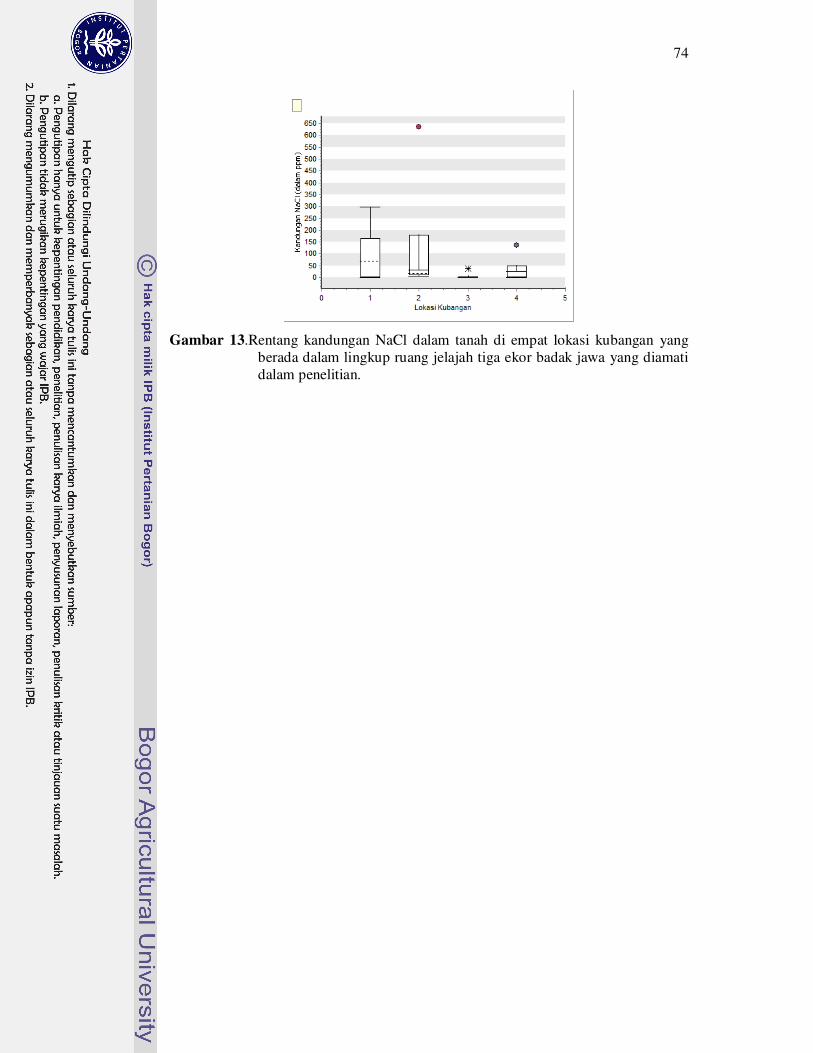
74
Gambar 13.Rentang kandungan NaCl dalam tanah di empat lokasi kubangan yang
berada dalam lingkup ruang jelajah tiga ekor badak jawa yang diamati
dalam penelitian.

75
Simpulan dan Saran
Simpulan
1. Metode penyusuran lintasan (trajektori) merupakan suatu cara yang memiliki
peluang keberhasilan tinggi dan berpotensi untuk diterapkan sebagai prosedur
pemantauan kualitas nutrisi yang dapat diterapkan untuk pemantauan
kecukupan nutrisi badak di habitat alaminya.
2. Hasil analisis komposisi pakan di lapangan menunjukkan bahwa pakan yang
memiliki palatabilitas tinggi terdiri dari: Leea sambucina (Sulangkar),
Dracontomelon puberulum (Dahu), Amomum megalocheilos (Tepus),
Spondias pinnata (Tepus), Zanthoxylum rhetsa (Kitanah), Diospyros
macrophylla (Kicalung), dan Ficus hispida (Bisoro).
3. Badak 12 menghuni daerah dengan keragaman pakan yang cukup tinggi
dengan kualitas pakan yang relatif tinggi protein dan energi. Hal ini
menunjukkan struktur vegetasi yang memadai di ruang jelajah badak 12,
sehingga hewan ini tidak memerlukan ruang jelajah yang besar.
4. Jenis pakan ideal yang kaya nutrisi (protein dan lemak) serta energi bagi badak
jawa terdiri dari tumbuhan: Moringa citrifolia, Callicarpa longifolia,
Chisocheton microcarphus, (protein tinggi); Alstonia angustiloba, Callicarpa
longifolia, Macaranga spp, (lemak tinggi); Derris thyorsifolia, Pterospermum
javanicum, Percampyulus glances, (energi tinggi); Paederia scandens,
Alstonia scholaris, Costus speciosus (kandungan air tinggi).
5. Ada indikasi kualitas air, nutrien, dan energi yang kurang memadai pada
waktu-waktu tertentu (periode kering Oktober-November 2009 dan Februari-
Maret 2010), terutama kandungan protein dan lemak yang rendah pada
tumbuhan dengan palatabilitas tinggi. Defisit nutrien lemak akan
mempengaruhi proses sintesa hormon steroid yang berfungsi dalam siklus
reproduksi. Secara keseluruhan konsumsi nutrien badak jawa masih memenuhi
kebutuhan untuk bertahan hidup dengan tingkat kecernaan yang tinggi.
6. Perbandingan hasil berdasarkan metode AIA dan koleksi total ini
menunjukkan bahwa metode AIA sedikit lebih tinggi bila dibandingkan
dengan metode koleksi total dan dapat digunakan dalam menghitung
kecernaan pada satwa liar dimana metode koleksi total tidak dapat dilakukan.

76
7. Asupan toksin lantaden melalui konsumsi Lantana camara yang terjadi pada
badak jawa tidak akan menimbulkan dampak pada kesehatan karena
jumlahnya yang sangat kecil (23.63 mg/ek/hari) atau 2.36 x 10-7
% dari berat
badan badak.
8. Kebiasaan menjilat lumpur (salt licking) merupakan upaya untuk memenuhi
kebutuhan garam, karena ada kandungan garam yang memadai pada lumpur
dan tanah di sekitar kubangan.
Saran
1. Perlu pengkajian potensi persemaiantumbuhan pakan bernutrisi tinggi untuk
dikembangkan sebagai bagian dari upaya pengkayaan habitat badak jawa.
2. Dengan berpedoman pada hasil analisis vegetasi yang dilakukan oleh Rahmat
et al. (2007), optimalisasi habitat badak di Taman Nasional Ujung Kulon perlu
dilakukan dengan memperluas sebaran dan memperbanyak ketersediaan
tumbuhan (sebutkan 5 terbaik) dengan formulasi kerapatan: 5,406 individu/ha
(jenis tumbuhan bawah) ; 2,222 individu/ha untuk jenis semai ; 268
individu/ha untuk jenis pancang ; 32 individu/ha untuk jenis tiang ; dan 15
individu per hektar untuk jenis pohon.

77
Daftar Pustaka
Birkett A, Stevens-Wood B,. 2005. Effect of low rainfall and browsing by large
herbivores on an enclosed savannah habitat in Kenya. African journal of
ecology 43: 123-130
Clauss M, et al. 2005. Studies on digestive physiology and feed digestibilities in
captive Indian rhinoceros (Rhinoceros unicornis). Journal of Animal
Physiology and Animal Nutrition 89 (2005) 229–237
Dinerstein E, 2003. The return of the unicorns. The natural history and conservation
of the greater one-horned rhinoceros. Columbia Universtiy Press, New york
Fernando P, Polet G, Foead N. Ng L. Melnick DJM. 2004. Mitochondrial DNA
analysis of the critically endangered Javan Rhinoceros. PHKA-WWF-Columbia
University
Hariyadi ARS, et al. 2011. Estimating the population structure of the Javan rhino
(Rhinoceros sondaicus) in Ujung Kulon National Park using the mark-recapture
method based on camera and video trap identification. Pachyderm no 49: 90-99
Koolman J, Rohm KH, Wanandi SI[ed], Sadikin M [ed]. 2001. Atlas berwarna dan
teks Biokimia. Cetakan I. Penerbit Hipokrates, Jakarta
Mainka SA, Zhao GL, Li M, 1989. Utilization of a bamboo, sugar cane, and gruel diet
by two juvenile giant pandas (Ailuropoda melanoleuca). Journal of zoo and
wildlife medicine 20:39-44
McFadyen REC,. 1998. Biological control of weeds. Annual review of entomology
43:369-393
McSweeney CS, Pass MA,. 1983. The mechanism of ruminal stasis in Lantana-
poisoned sheep. Quarterly journal of experimental physiology 68: 301-313
Mundiany L, Agil M, Astuti DA,. 2005. Studi Kasus: Estimasi Gambaran Nutrien
pada Badak Sumatera (Dicerorhinus sumatrensis) Jantan di Suaka Rhino
Sumatera Taman Nasional Way Kambas. [Skripsi] Institut Pertanian Bogor
Pagan JD, et al. 1998. Exercise affects digestibility and rate of passage of all-forage
and mixed diets in throroughbred horses. American society for nutritional
sciences. J. Nutr. 128: 2704S-2707S
Purchase D,. 2007. Using spoor to determine the age and weight of subadult black
rhinoceroses (Diceros bicornis L). South African Journal of Wildlife Research
37(1): 96-100

78
Rahmat UM, Santosa Y, Kartono AP,. 2007. Analisis Tipologi Habitat Preferensial
bagi Badak Jawa (Rhinoceros sondaicus, Desmarest 1822) di Taman Nasional
Ujung Kulon. Institut Pertanian Bogor
Sharma OP, Singh A, Sharma S,. 2000. Levels of lantadene, bioactive pentacyclic
triterpenoids, in young and mature leaves of Lantana camara var acuelata.
Fitoterapia 71(5):487-491
Sims JA, et al. 2007. Determination of bamboo-diet digestibility and fecal output by
giant pandas. Ursus 18(1): 38-45
Sriyanto A, et al. 1995. A Current status of the Javan Rhino population in Ujung
Kulon National Park. Javan Rhino Colloquium
VanKeulen J, Young, BA. 1977. Evaluation of Acid Insoluble Ash as a Natural
Marker in Ruminant digestibility Studies. Journal of Animal Science vol 44 no:
2. 282-287
White AM, Swaisgood RR, Czekala N,. 2007. Ranging patterns in white rhinoceros,
Ceratotherium simum simum: implications for mating strategies. Animal
behaviour (74): 349-356

PENENTUAN ASAI HORMON GLUKOKORTIKOID UNTUK MEMANTAU
INDIKATOR CEKAMAN PADA BADAK JAWA JANTAN.
Abstrak
Salah satu aspek penting dalam pengelolaan populasi badak jawa (Rhinoceros
sondaicus) di Taman Nasional Ujung Kulon Banten adalah upaya untuk mempelajari
dan memantau tingkat cekaman yang dihadapi oleh badak jawa di habitatnya. 3α,11β-
dihydroxy-CM dari feses merupakan metabolit glukokortikoid yang diduga dapat
digunakan untuk analisis tingkat cekaman pada badak jawa. Kajian cekaman dilakukan
pada tiga individu (badak 12, 13, dan 18) dengan tingkat asupan pakan dan nutrien
yang berbeda dan juga antar musim untuk mempelajari variasi kadar glukokortikoid di
musim kering dan musim hujan. Hasil yang diperoleh menunjukkan adanya fluktuasi
pada profil 3α,11β-dihydroxy-CM pada individu badak 12 dan 18 selama musim
kering, dan individu 18 (individu muda) memiliki fluktuasi 3α,11β-dihydroxy-CM yang
besar. Hal ini menunjukkan bahwa individu nomor 18 tersebut mengalami cekaman
yang fluktuatif yang tidak hanya disebabkan oleh faktor kekeringan, namun mungkin
pula disebabkan oleh cekaman akibat rendahnya asupan energi asal pakan per berat
badan (%), dan juga disebabkan oleh adanya interaksi sosial dengan badak dewasa.
Musim kering yang memiliki jumlah kejadian hujan yang rendah yaitu rataan 0.2
kejadian hujan setiap harinya cenderung menyebabkan cekaman yang tinggi pada
badak. Penelitian ini menunjukkan bahwa defisit energi asal pakan dan keterbatasan air
merupakan sumber cekaman yang besar bagi badak jawa.
Kata kunci: badak jawa, hormon, cekaman, EIA

80
Analysis of 3α,11β-dihydroxy-CM profile for indicator of stress on male javan
rhinoceros
Abstract
One of the most important aspects in conservation of javan rhinoceros
(Rhinoceros sondaicus) in Ujung Kulon National Park, Banten was to study and to
monitor the levels of stress that the animals faced in their habitat.There was an
indication that 3α,11β-dihydroxy-CM from feces was a glucocorticoid metabolite that
could potentially be used for analyzing the levels of stress in javan rhino. Assessment
was done to study stress variations among the three rhinos (rhino 12,13 and 18) that
had different levels of feed intake, as well as to study variations of glococorticoid levels
in dry and rainy seasons. The result from this study showed that there was a fluctuation
in 3α,11β-dihydroxy-CM levels in rhino number 18 and 12, while rhino number 18 (a
young rhino) showed a large fluctuation in 3α,11β-dihydroxy-CM level. This indicated
that this young rhinoexperienced fluctuative stress that may be caused not only by
energy intake deficit (energy intake per body weight) and water deficit, but also by
social interactions involving adult males in the same home range. Dry season where
the daily rain occassion is low (0.2 rain occurrences per day) tend to cause higher
stress for the rhinos. This research shows that the deficit in energy intake per body
weight (%) and water limitation are among the biggest sources of stress for rhino
population.
Key words: Javan rhinoceros, hormones, stress, EIA

81
Pendahuluan
Mekanisme dan Respons Individu terhadap Cekaman
Seperti halnya hewan mamalia lainnya, badak jawa menghadapi berbagai
macam kondisi yang secara langsung ataupun tidak langsung dapat mengganggu fungsi
fisiologis tubuh. Kondisi yang mengancam keseimbangan fungsi fisiologis ini
didefinisikan sebagai sumber cekaman atau stressor (Morgan & Tromborg 2007).
Dengan adanya cekaman ini, badak jawamelakukan proses tanggap cekaman melalui
mekanisme yang melibatkan berbagai proses pengiriman sinyal melalui sistem saraf,
sistem endokrin, dan juga melibatkan proses adaptasi fisiologis. Proses-proses tersebut
merupakan mekanisme tanggap cekaman yang dilakukan oleh hewan untuk
mempertahankan keseimbangan metabolisme (homeostasis). Salah satu strategi dalam
mempertahankan homeostasis ini adalah dengan produksi hormon-hormon seperti
adrenalin dan kortisol yang dapat membantu penyediaan energi secara cepat. Uji coba
Induksi cekaman pada hewan model kuda dalam salah satu bagian penelitian disertasi
ini telah membuktikan bahwa kadar kortisol dalam darah meningkat secara
signifikan.Hal ini menandakan adanya peningkatan aktifitas korteks adrenal melalui
mekanisme pengaturanporos hipotalamus-pituitari-adrenal. Sekresi hormon-hormon
tersebutkemudian beredar di dalam darah dan mendorong proses glukoneogenesis dan
lipolisis (Koolman & Röhm 2001) sebagai upaya untuk menyediakan energi secara
cepat sebagai upaya mempertahankan keseimbangan fungsi fisiologis tubuh dalam
kondisi cekaman.
Cekaman dari lingkungan yang dirasakan oleh badak diterima langsung oleh
berbagai reseptor yang terdapat pada permukaan tubuh yang mengirimkan sinyal ke
pusat nyeri yang kemudian mengirimkan sinyal cekaman ke hipotalamus (Baron 2006),
adapun cekaman fisiologis akibatperubahan komposisi kimia darah (hipoglisemia,
toksin) juga mengirimkan sinyal cekaman kepada hipotalamus ini. Demikian pula
dengan perubahan pada nutrisi yang dapat dideteksi di dalam saluran pencernaan serta
pusat termoregulasi yang mengirimkan sinyal kepada hipotalamus dalam kondisi
adanya cekaman (Coenen 2005). Sinyal cekaman yang diterima oleh hipotalamus ini
kemudian dilanjutkan dengan reaksi tanggap cekaman yang dikenal dengan aksis HPA
(Hipotalamus-Pituitari-Adrenal) yang berujung pada peningkatan aktifitas korteks
adrenal dan adanya peningkatan sekresi hormon cekaman seperti adrenalin dan kortisol
tergantung dari sifat cekaman tersebut (akut atau kronis).Reaksi tanggap cekaman juga

82
dapat menyebabkan adanya perubahan perilaku yang dimediasi oleh proses neuro-
endokrin yang melibatkan pengiriman sinyal oleh sistem saraf yang kemudian memicu
sekresi hormon. Perubahan ini merupakan mekanisme perilaku stress avoidance yang
memungkinkan hewan tersebut untuk mendeteksi dan menghindari sumber cekaman
(Dickens et al. 2010).
Gambar 14. Reaksi biokimia yang merupakan proses tanggap terhadap cekaman bagi
mamalia (sumber: Coenen 2005).

83
Definisi Cekaman
Dickens et al. (2010) menjelaskan bahwa badak memiliki mekanisme tanggap
cekaman yang bertujuan untuk mempertahankan homeostasis atau keseimbangan
fisiologi di dalam tubuh. Mekanisme ini memiliki proses yang berbeda terhadap
sumber cekaman akut yang berupa tekanan tunggal yang segera ditanggapi dengan
mekanisme fight or flight hasil dari mediasi sistem saraf simpatetik. Bila sumber
cekaman ini berlangsung berulang-ulang atau berlangsung dalam waktu yang cukup
lama, maka cekaman ini menjadi kronis (Wingfield & Romero 2001). Kedua jenis
cekaman ini memberikan stimulasi terhadap hipotalamus dan memberikan respons
berupa peningkatan aktifitas adrenal korteks yang ditunjukkan dengan adanya
peningkatan sekeresi hormon glukokortikoid (Barja et al. 2007) pada berbagai hewan
termasuk badak (Turner Jr et al. 2002; Menargues et al. 2008). Selain kadar hormon
glukokortikoid, cekaman akut dan kronis menimbulkan reaksi yang berbeda pada
metabolisme dan sistem pertahanan tubuh hewan.
Hal ini sangat relevan dengan keadaan yang dihadapi oleh badak jawa di
habitatnya yang mengalami defisit pakan dan juga defisit air pada musim kering.
Individu badak no 12 dipilih sebagai sampel untuk mewakili kondisi defisit pakan
karena individu ini menunjukkan ruang jelajah yang paling kecil dengan jumlah
asupan, keragaman pakan, dan juga kualitas nutrisi yang paling rendah dibandingkan
dengan kedua ekor badak lainnya.
Kondisi defisit air di habitat badak jawa didefinisikan sebagai musim kering
saat ketersediaan sumber air bagi badak di Taman Nasional Ujung Kulon diprediksi
menurun berdasarkan skenario perubahan iklim (Permadi 2008). Musim kering ini
memiliki karakteristik berupa curah hujan yang rendah, dan oleh karena itu, data
hormon untuk menggambarkan kondisi defisit air didefinisikan sebagai kadar hormon
pada bulan-bulan dengan curah hujan yang rendah berdasarkan jumlah kejadian hujan
per hari pada bulan tersebut.
Hormon Terkait Cekaman Beserta Metabolitnya
Kadar hormon kelompok glukokortikoid yang merupakan respons dari aktifitas
Adrenocorticotrophin Hormone (ACTH) telah banyak digunakan sebagai indikator
adanya stress pada berbagai spesies hewan seperti: anjing (Schoeman et al. 2007),
karnivora (Young et al. 2004), Kerbau (Garg & Chander 1997), pengerat (Soto-
Gamboa et al. 2009), dan sapi perah (Morrow et al. 2000). Selain kortisol, indikator

84
hormon stres lainyang dapat digunakan untuk mengukur cekaman pada hewan adalah
kortikosteron (Wasser et al. 2000). Kejadian cekaman dapat juga diukur dengan
indikator metabolit kortisol yang diekskresikan dalam feces dan/atau urin seperti
androstenediol yang meningkat seiring adanya cekaman pada hewan (peningkatan
aktifitas pada korteks adrenal) sebagaimana dijelaskan dalam Barja et al. (2007). Salah
satu metabolit kortisol yang memiliki potensi sebagai indikator adanya aktifitas korteks
adrenal akibat cekaman adalah 3α,11β-dihydroxy-CM (Ghalib et al. 2011).
Kortisol merupakan hormon cekaman yang disekresikan oleh kelenjar adrenal
dan dieksresikan melalui feses ataupun urin seperti yang ditampilkan pada Gambar 14.
Namun demikian, ada kemungkinan bahwa kortisol yang telah diekskresikan melalui
feses ataupun urine sudah mengalami inaktivasi dimana hormon tersebut sudah
mengalami konversi metabolik, konjugasi dan reduksisehingga akan diekskresikan
sebagai metabolit dari kortisol. Oleh karena itu, dalam merancang suatu metoda untuk
memantau tingkat cekaman berdasarkan kadar hormon pada feses, opsi untuk
mendeteksi metabolit kortisol telah dikembangkan dan digunakan untuk memastikan
bahwa tingkat hormon cekaman tetap dapat diukur walaupun hormon kortisol telah
mengalami perubahan karena flukutasi cekaman berkorelasi positif dengan fluktuasi
kortisolatau kortikosteron (Wasser et al. 2000). Pada hewan monitoring stress dapat
dilakukan dengan menganalisa metabolit glukokortikoid 5-beta-androstendiol pada
sampel (Barja et al. 2007) dan pada hewan dengan menganalisa 3α,11β-dihydroxy-CM
pada sampel.
Permasalahan yang dihadapi dalam pengelolaan populasi badak jawa saat ini
adalah tidak adanya perangkat yang dapat digunakan untuk memantau tingkat cekaman
yang dihadapi badak jawa dengan tepat. Kendala ini mengakibatkan kurangnya
informasi yang diperlukan untuk meningkatkan peluang untuk berkembang biak dan
bertahan dari penyakit. Sampai saat ini belum diketahui tentang indikator yang dapat
digunakan untuk memantau tingkat cekaman pada badak jawa dari feses, belum ada
informasi mengenai profil glukokortikoid pada badak, dan belum ada analisis mengenai
kemungkinan adanya perbedaan profil glukokortikoid (sebagai reaksi tanggap terhadap
cekaman) antar individu badak maupun perbedaan profil glukokortikoid antara kondisi
musim (kering dan penghujan).

85
Tujuan Penelitian
Dengan mempertimbangkan potensi dari penggunaan metabolit glukokortikoid
pada sampel feses, dan juga mengingat adanya kebutuhan untuk memantau tingkat
cekaman dan kapasitas reproduksi pada hewan langka seperti badak jawa, maka
penelitian ini dirancang untuk mendapatkan informasi sebagai berikut:
1. Mengidentifikasi indikator yang potensial untuk digunakan dalam analisis profil
glukokortikoid pada feses badak jawa.
2. Mempelajari karekteristik profil glukokortikoid asal feses pada individu badak
yang digunakan pada penelitian.
3. Mempelajari hubungan antara faktor cekaman (defisit air) dengan profil
glukokortikoid asal feses pada badak jawa.
Penelitian ini dirancang untuk menguji hipotesis bahwa metabolit hormon asal feses
badak jawa dapat digunakan sebagai indikator status dan profil glukokortikoid

86
Bahan dan Metode
Penetapan FaktorCekaman di Habitat Badak Jawa
Cekaman di habitat badak jawa di Taman Nasional Ujung Kulon didefinisikan
dalam dua faktor yaitu faktor asupan pakan dan faktor ketersediaan air. Faktor pakan
dibedakan dengan jumlah asupan pakan dan nutrisi setiap ekor badak seperti yang telah
dirangkum dalam Tabel 7 pada penelitian tentang asupan dan nutrisi. Ketersediaan air
diwakili oleh musim dengan membedakan bulan-bulan berdasarkan jumlah kejadian
hujan per hari yang dicatat oleh pengamat selama berada di habitat badak di Taman
Nasional Ujung Kulon.
PenetapanSampel Feses
Badak jawa adalah salah satu spesies hewan yang sudah langka dan sangat sulit
untuk dijumpai di habitat alaminya. Selain itu, sampai saat ini belum ada Badak Jawa
yang berada di penangkaran ataupun di kebun binatang. Oleh karena itu, pengambilan
sampel darah untuk kepentingan analisis hormon hampir tidak mungkin untuk
dilakukan. Salah satu hal yang paling mungkin dilakukan adalah melakukan analisis
hormon berdasarkan sampel feses sebagaimana pernah dilakukan oleh Santymire &
Armstrong (2009) pada spesies anjing liar.
Konsep yang sama dicoba diterapkan pada badak jawa dengan syarat bahwa
sampel feses yang digunakan dalam analisis ini harus berada dalam kondisi segar
(kurang lebih 24-30 jam).Analisis hormon dari feses badak dirancang untuk dapat
mempelajari perbedaan hormon (terkait cekaman dan reproduksi) antar individu dan
juga antar musim. Tahap induksi cekaman pada hewan model dalam rangkaian
penelitian ini menunjukkan bahwa defisit pakan menyebabkan meningkatnya cekaman
(ditunjukkan dengan meningkatnya kadar kortisol dalam darah), dengan demikian,
pengukuran kadar hormon stress dilakukan untuk menguji apakah pola yang sama juga
terjadi pada badak yang mengalami cekaman (defisit pakan dan juga defisit air).

87
Pengumpulan Sampel Feses
Feses badak merupakan tumpukan yang terdiri dari beberapa boli, dan
kesegaran feses tersebut dapat ditentukan dari warna, kelembaban, serta keberadaan
serangga yang mengelilinginya. Feses segar biasanya berwarna coklat kehijauan
dengan kelembaban permukaan yang khas karena masih mengandung lapisan lendir
(tidak lebih dari satu jam sejak ekskresi), serta masih banyak dikelilingi serangga lalat.
Dengan memperhitungkan jarak antara pengamat dengan badak berdasarkan jejak dan
dengan prosedur pengamatan yang mengharuskan pengamat berada 24-30 jam
(maksimum 1.25 hari) di belakang badak yang diamatinya, maka dapat disimpulkan
bahwa feses yang dikumpulkan berdasarkan kriteria di atas berusia tidak lebih dari 30
jam sejak defekasi.Sampel feses dikumpulkan dalam periode waktu yang sama (musim
hujan atau musim kering) untuk dibandingkan dalam musim yang sama.
Lima gram feses segar (diukur berdasarkan jumlah feses yang dapat diambil
dengan menggunakan sendok kayu khusus) diambil untuk analisis hormon. Feses yang
relatif segar dapat menjamin ketersediaan hormon kortisol serta metabolitnya untuk
digunakan dalam analisis hormon. Sampel feses untuk analisis hormon dengan EIA
dipreservasi (diawetkan) dalam 25 ml etanol 90% (teknis) dan disimpan dalam botol
plastik 50 ml untuk kemudian dikirim ke laboratorium Unit Reproduksi dan
Rehabilitasi di Institut Pertanian Bogor untuk disimpan dalam freezer dengan suhu -20o
C.
Pemilihan Indikator Kadar Hormon Cekaman Asal Feses
Indikator kadar hormon cekaman dipilih berdasarkan kemungkinan adanya
hormon cekaman (glukokortikoid) maupun metabolitnya di dalam feses. Kit yang
tersedia untuk asai ini adalah: kortisol (Produksi DRG Jerman), kortikosteron (Produksi
CUSABIO,Cina), dan 3α, 11β-dihydroxyetiocholanolone (3α,11β-dihydroxy-CM) yang
dikembangkan oleh German Primate Centre di Gottingen, Jerman dan asainya
dilakukan di laboratorium Unit Rehabilitasi dan Reproduksi (URR) IPB, Bogor. Setiap
kit/indikator ini digunakan dalam uji paralelisme untuk menentukan kelayakan kit dan
indikator tersebut dalam asai hormon.
Uji paralelisme digunakan sebagai prosedur baku untuk menguji kelayakan
sebuah kit atau antibodi bila digunakan dalam assay hormon yang dimaksud. Kurva
yang dihasilkan dari assay pada sampel harus menunjukkan paralel (tidak ada
kemungkinan persinggungan atau perpotongan) denga kurva baku yang sudah

88
ditentukan dari produsen kit. Kurva baku ini dibuat berdasarkan kalibrasi
perikatanyang spesifik antara antibodi dan antigen yang digunakan dalam kit tersebut
(Francois-Gerard et al. 1988). Schoeneker et al. (2004) menjelaskan bahwa selain
untuk validasi kelayakan penggunaan kit atau antibodi tertentu uji paralelisme juga
diperkuat dengan penghitungan variasi intra-assay and inter-assay. Kretzschmar et al.
(2004) menggunakan assay EIA pada badak putih Afrika dan mencari faktor pencairan
(dilution factor) yang paling optimal. Titik perikatan (binding) 50% dimana setengah
dari penanda dapat melekat pada antibodi merupakan titik ideal untuk menentukan
faktor pencairan. Faktor pencairan di luar rentang tersebut akan mengurangi akurasi
dari asai hormon yang dilakukan.
Faktor lainnya yang sangat mempengaruhi ketepatan asai hormon yang
digunakan adalah cross reactivity atau reaktifitas silang antara penanda / antibodi yang
digunakan untuk asai sebuah hormon yang dapat juga mendeteksi hormon lain (atau
metabolitnya) yang tidak diharapkan, oleh karena itu reaktifitas silang yang tinggi
antara dua hormon atau lebih akan secara signifikan mengurangi ketepatan asai hormon
yang digunakan (Agil et al. 2008). 5-beta-androstenediol sebagai salah satu metabolit
glukokortikoid merupakan antibodi yang tidak memiliki reaktifitas silang dengan
hormon lain dan hanya memiliki tingkat reaktifitas silang dibawah 0.5% dengan
testosteron. 5-beta-androstenediol ini memiliki reakfitas silang 1.49% dengan
androstenedione yang digunakan dalam assay testosteron (Brown et al. 2002). 3α, 11β-
dihydroxy-CM merupakan antibodi yang belum pernah digunakan pada sampel feses
badak jawa dan memiliki kemungkinan terjadi reaktifitas silang dengan androgen
karena antibodi ini mendeteksi metabolit kortisol dan juga metabolit androgen yang
memiliki 19 karbon dalam rangkaian molekulnya.
Persiapan Sampel Feses
Sebagai prosedur baku dalam kajian hormon dari feses, maka penggunaan kit
kortisol, kortikosteron, dan 3α,11β-dihydroxy-CM pada feses harus menjalani uji
validitas menggunakan uji paralelisme. Uji validitas ini merupakan langkah penting
untuk menentukan kekuatan validitas dari EIA untuk kajian kuantitatif (Francois-
Gerard et al. 1988). Kit Kortisol (produksi DRG, Jerman), Kortikosteron (Produksi
CUSABIO, Cina), dan antibodi 3α,11β-dihydroxy-CM digunakan untuk melakukan uji
paralelisme pada sampel feses badak jawa yang telah dikumpulkan.

89
Setiap sampel yang dikumpulkan dari lapangan diproses dengan metode yang
dijelaskan dalam Ziegler & Wittwer (2005). Ekstraksi dilakukan dengan mencairkan
sampel yang berada dalam kondisi beku sampai kondisi sampel yang berada dalam
larutan fiksasi etanol 90% (teknis) kembali menjadi cair dan larutan fiksasi menguap
secara menyeluruh dan menyisakan sampel feses dalam kondisi kering. Ekstraksi 0.1-
0.5g dari sampel feses dilanjutkan dengan mencampur feses tersebut dengan 2.5 ml
etanol 90% dan 2.5 ml akuades. Campuran diaduk di dalam tabung plastik secara
merata dengan mengguncang tabung tersebut. Setelah tercampur, larutan ini kemudian
disentrifugasi selama lima menit untuk memisahkan cairan dengan partikel padat yang
merupakan ekstrak untuk digunakan dalam assay kortisol, kortikosteron, dan 3α,11β-
dihydroxy-CM.
Asai Kortisol
Kit kortisol produksi DRG Jerman bekerja dengan sistem ELISA kompetitif dan
terdiri dari: sumur mikrotiter (96 sumur) yang sudah dilapisi dengan antibodi anti-
kortisol monoclonal, larutan standard dengan pengawet Proclin 0.3%, Konjugat enzim
kortisol-horseradish peroxidase dengan menggunakan pengawet Proclin 0.3%, Larutan
substrat tetrametilbenzidin (TMB), larutan stop yang mengandung 0.5 M H2SO4. Asai
kortisol ini memiliki karakteristik rentang asai 0-800 ng/ml dan konsentrasi untuk kurva
standard pada 800ng/ml, 400ng/ml, 200ng/ml, 100ng/ml, 50 ng/ml, 20 ng/ml, dan 0
ng/ml; reaktifitas silang dengan kortisol (100%), Kortikosteron (45%), Progesteron
(95%), Deoxycortisol dan Dexametason (masing-masing 2%), estron, estriol, dan
testosteron (masing-masing dibawah 0.01%); sensitifitas 2.5 ng/ml (6.9 nmol/l); presisi
intra asai 3.2%-8.1% (20 repetisi); dan presisi inter asai 6.6%-7.7% (20 repetisi).
Prosedur Asai Kortisol
1. Sumur-sumur pada lempeng mikrotiter (plate) ditempatkan sesuai nomor
dan dipastikan tidak bergeser.
2. 20 µl standard, kontrol, dan sampel ditempatkan di sumur yang telah
disediakan dengan menggunakan ujung mikropipet yang baru untuk setiap
sampel.
3. 200 µl konjugat enzim diteteskan di setiap sumur dengan menggunakan
mikro pipet.
4. Semua larutan diaduk dengan sempurna selama 10 detik.

90
5. Inkubasi dilakukan selama 60 menit tanpa menutup sumur pada lempeng
mikrotiter.
6. Cairan yang ada di dalam sumur dikeluarkan dan setiap sumur dibilas
dengan menggunakan larutan pencuci (400 µl setiap sumur). Sisa tetes
larutan dikeringkan dengan bantuan kertas penyerap.
7. 100 µl larutan substrat diteteskan di setiap sumur yang sudah kering
8. Inkubasi dilakukan selama 15 menit pada suhu ruangan
9. Reaksi enzimatik dihentikan dengan menambahkan larutan stop sebanyak
100 µl ke setiap sumur.
10. Lempeng mikrotiter dibaca dengan menggunakan optical density (OD) 450
+/- 10 nm tidak lebih dari 10 menit setelah larutan stop diberikan.
Asai Kortikosteron
Kit kortikosteron produksi CUSABIO Cina terdiri dari: sumur mikrotiter (96
sumur) yang sudah dilapisi dengan antibodi anti-kortikosteron monoclonal, larutan
standard, Konjugat enzim HRP, Larutan substrat A dan B, larutan stop yang
mengandung 0.5 M H2SO4. Asai kortikosteron ini memiliki karakteristik rentang asai
0.6-160 ng/ml dan konsentrasi untuk kurva standard pada 160ng/ml, 40ng/ml, 10ng/ml,
2.5ng/ml, dan 0.63 ng/ml; tidak ada reaktifitas silang; sensitifitas dibawah 0.4 ng/ml;
presisi intra asai di bawah 10% (20 repetisi); dan presisi inter asai dibawah8% (35
repetisi).
Prosedur Asai Kortikosteron
1. Sumur-sumur pada lempeng mikrotiter (plate) ditempatkan sesuai nomor
dan dipastikan tidak bergeser.
2. 50 µl standard, kontrol, dan sampel ditempatkan di sumur yang telah
disediakan dengan menggunakan ujung mikropipet yang baru untuk setiap
sampel.
3. 50 µl konjugat enzim dan 50 µl antibodi diteteskan di setiap sumur dengan
menggunakan mikro pipet.
4. Semua larutan diaduk dengan sempurna selama 10 detik.
5. Inkubasi dilakukan selama 60 menit tanpa menutup sumur pada lempeng
mikrotiter pada suhu 370 C.

91
6. Cairan yang ada di dalam sumur dikeluarkan dan setiap sumur dibilas
dengan menggunakan larutan pencuci (200 µl setiap sumur). Aduk selama
10 detik, buang semua larutan, dan sisa tetes larutan dikeringkan dengan
bantuan kertas penyerap.
7. 50 µl larutan A dan 50 µl larutan Bpewarna diteteskan di setiap sumur yang
sudah kering.
8. Inkubasi dilakukan selama 15 menit pada suhu ruangan
9. Reaksi enzimatik dihentikan dengan menambahkan larutan stop sebanyak
50 µl ke setiap sumur dan aduk selama 1-2 detik sampai warna merata.
10. Lempeng mikrotiter dibaca dengan menggunakan optical density (OD) 450
+/- 10 nm tidak lebih dari 10 menit setelah larutan stop diberikan.
Asai 3α,11β-dihydroxy-CM
Kit 3α,11β-dihydroxy-CM yang terdiri dari: sumur mikrotiter (96 sumur) yang
sudah dilapisi dengan antibodi anti- 3α,11β-dihydroxy-CM monoclonal, larutan
standard, Konjugat enzim 3α,11β-dihydroxy-CM, Larutan substrat tetrametilbenzidin
(TMB), larutan stop yang mengandung 0.5 M H2SO4. Asai 3α,11β-dihydroxy-CM ini
memiliki karakteristik rentang konsentrasi hormon pada standar kurvanya antara 0.6
ng/ml sampai dengan 78 ng/ml dengan konsentrasi untuk kurva standar adalah 78ng/ml,
39ng/ml, 19.5ng/ml, 9.6ng/ml, 4.8 ng/ml, 2.4 ng/ml, 1.2 ng/ml, 0.6 ng/ml,; reaktifitas
silang dengan androgen (testosteron); sensitifitas 2.5 ng/ml (6.9 nmol/l); presisi intra
asai 4.4%-5.4% (18 repetisi masing-masing untuk high dan low); dan presisi inter asai
6.8%-11.2% (6 repetisi masing-masing untuk high dan low).
Prosedur Asai 3α,11β-dihydroxy-CM.
1. Sumur-sumur pada lempeng mikrotiter (plate) ditempatkan sesuai nomor
dan dipastikan tidak bergeser.
2. 50 µl standard, kontrol, dan sampel ditempatkan di sumur yang telah
disediakan dengan menggunakan ujung mikropipet yang baru untuk setiap
sampel.
3. 50 µl konjugat enzim dan 50 µl antibodi diteteskan di setiap sumur dengan
menggunakan mikro pipet.
4. Semua larutan diguncang dengan sempurna selama 10 detik.

92
5. Inkubasi dilakukan selama satu malamdalam lempeng mikrotiter pada suhu
40 C.
6. Cairan yang ada di dalam sumur dikeluarkan dan setiap sumur dibilas empat
kali dengan menggunakan larutan pencuci. Guncang selama 10 detik, buang
semua larutan, dan sisa tetes larutan diketukkan sampai habis pada kertas
penyerap.
7. 150 µl streptavidin-peroksidase (produksi Sigma, Jerman) ditambahkan ke
dalam setiap sumur.
8. Lempeng di inkubasi selama 30 menit di dalam gelap pada suhu ruangan.
9. Cairan yang ada di dalam sumur dikeluarkan dan setiap sumur dibilas empat
kali dengan menggunakan larutan pencuci. Guncang selama 10 detik, buang
semua larutan, dan sisa tetes larutan diketukkan sampai habis pada kertas
penyerap.
10. 150 µl larutan substrat/pewarna (0.025% benzidin dan 0.05% H2O2).
11. Lempeng diinkubasi selama 30-45 menit di dalam gelap pada suhu ruangan
sampai warna kuning muncul.
12. Reaksi enzimatik dihentikan dengan menambahkan larutan stop (H2SO4)
sebanyak 50 µl ke setiap sumur dan aduk selama 1-2 detik sampai warna
merata.
13. Lempeng mikrotiter dibaca dengan menggunakan optical density (OD) 450
nm tidak lebih dari 10 menit setelah larutan stop diberikan.
Perangkat lunak Gen5 (Biotek, Jerman) digunakan untuk menyusun kurva
berdasarkan pembacaan dari EIA reader pada semua jenis asai. Perbandingan kurva
standard sesuai spesifikasi dari masing-masing kit pada uji paralelisme dijadikan
sebagai penentu konsistensi hormon pada feses badak. Bila kurva standar
menunjukkan garis yang paralel (tidak saling memotong), maka uji paralelisme ini
dilanjutkan ke tahap duauntuk melihat adanya perbedaan kadar hormon glukokortikoid
maupun metabolitnya pada beberapa sampel dan kemudian melakukan uji paralelisme
pada beberapa perwakilan sampel yang dipilih. Pemilihan sampel dilakukan dengan
memperkirakan sampel-sampel yang mewakili kondisi cekaman rendah: individu
dengan jumlah asupan dan kualitas pakan yang tinggi hujan (2 sampel), cekaman
tinggi: individu dengan jumlah dan kualitas asupan yang rendah (2 sampel),
sertacekaman sedang: individu dengan jumlah dan asupan nutrisi dalam tingkat

93
sedang/menengah (2 sampel). Semua sampel dipilih untuk mewakili kondisi pada
musing kering dengan jumlah hari hujan yang rendah serta musim hujan dengan jumlah
hari hujan yang tinggi. Semua sampel digunakan untuk mengukur variasi inter-assay
dan intra-assay. Deviasi kurang dari 15% pada variasi intra-assay dan inter-assay ini
menunjukkan kelayakan assay hormon yang dilakukan pada sampel yang dikumpulkan
dari feses badak jawa yang terkumpul dari lapangan.
Analisis Data
Kuantifikasi cekaman dilakukan untuk memastikan adanya perbedaan cekaman
antar individu badak jawa berupa tingginya kadar indikator glukokortikoid.
Perbandingan cekaman antar individu dilakukan dengan membandingkan tingkat
cekaman yang dialami badak dengan konsumsi pakan dengan kandungan air rendah
terhadap badak dengan konsumsi pakan dengan kandungan air tinggi pada musim yang
sama. Kadar glukokortikoid dari sampel feses setiap individu badak diukur dan rata-
rata kuantitas hormon (dalam ng/g feses) digunakan sebagai indikator untuk
membandingkan perbedaan yang ada secara deskriptif. Analisis dilakukan secara
deskriptif mengingat jumlah sampel yang terlalu sedikit untuk digunakan dalam
ANOVA.
Kuantifikasi cekaman dilakukan antar musim yang berbeda dilakukan untuk
mempelajari dinamika cekaman yang dialami semua badak (12, 13, dan 18) dalam
kondisi sumber air yang terbatas di musim kering dan dalam kondisi kecukupan air di
musim hujan. Sampel feses perlu didapat dari ketiga individu di bulan Oktober (musim
kering dengan rata-rata jumlah curah hujan yang rendah) dan di bulan Januari (musim
hujan dengan rata-rata jumlah curah hujan yang tinggi). Kadar hormon dari sampel
feses setiap individu badak diukur dan rata-rata kuantitas hormon (dalam ng/g feses)
digunakan untuk mebandingkan perbedaan yang ada secara deskriptif.

94
Hasil dan Pembahasan
Hasil Uji Paralelisme
Dari uji validitas menggunakan uji paralelisme dari ketiga kit hormon di atas
dapat dilihat bahwa hormon kortisol dan kortikosteron tidak dapat digunakan untuk
menganalisa metabolit glukokortikoid pada feses badak karena kurva standar dari
kedua kit ini tidak paralel dengan kurva sampel pada pengenceran bertingkat.
Berdasarkan uji paralelisme ini kit kortisol dan kortikoseron tidak dilanjutkan
penggunaannya dalam penelitian ini.
Pengukuran metabolit glukokortikoid 3α,11β-dihydroxy-CM sebagai indikator
keberadaan hormon cekaman menunjukkan hasil yang paralel antara kurva standard
dengan kurva sampel dengan tampak adanya kecenderungan yang sama antara profil
kurva standard an kurva sampel.
Uji paralelisme menggunakan 3α,11β-dihydroxy-CM ini menunjukkan 50%
binding pada konsentrasi 3,04 pg dan pengenceran 1:80. Validasi lanjutan juga
menunjukkan bahwa koefisien variasi (CV) intra-assay adalah 5.4% untuk QC low dan
4.4% untuk QC high dengan 18 repetisi. Demikian pula dengan CV inter-assay yang
menunjukkan angka 11.2% QC low dan 6.8% untuk QC high dengan 6 repetisi.
Berdasarkan hasil dari uji paralelisme ini yang didukung dengan CVintra-assay dan
inter-assay yang berada di bawah 15%, maka dapat disimpulkan bahwa, 3α,11β-
dihydroxy-CMmenunjukkan kemungkinan untuk dapat digunakan dan dapat
dilanjutkandengan validasi lebih lanjut sebelum diimplementasikan dalam kajian profil
glukokortikoid dari sampel feses badak jawa.
Faktor Pengenceran untuk Kuantifikasi Metabolit Glukokortikoid (3α,11β-
dihydroxy-CM)
Kurva standar dan kurva sampel pada uji paralelisme ditampilkan pada Gambar
15 dapat digunakan untuk menentukan faktor pengenceran ekstrak sampel feces badak
jawa yang akan dianalisa dengan kit hormon tersebut. Garis vertikal putus-putus pada
Gambar 15 menandai titik tengah pada kurva standard menunjukkan titik dimana terjadi
perikatan (binding) 50% dari antibodi terhadap antigen yang kemudian menunjukkan
faktor pengenceran1:80.
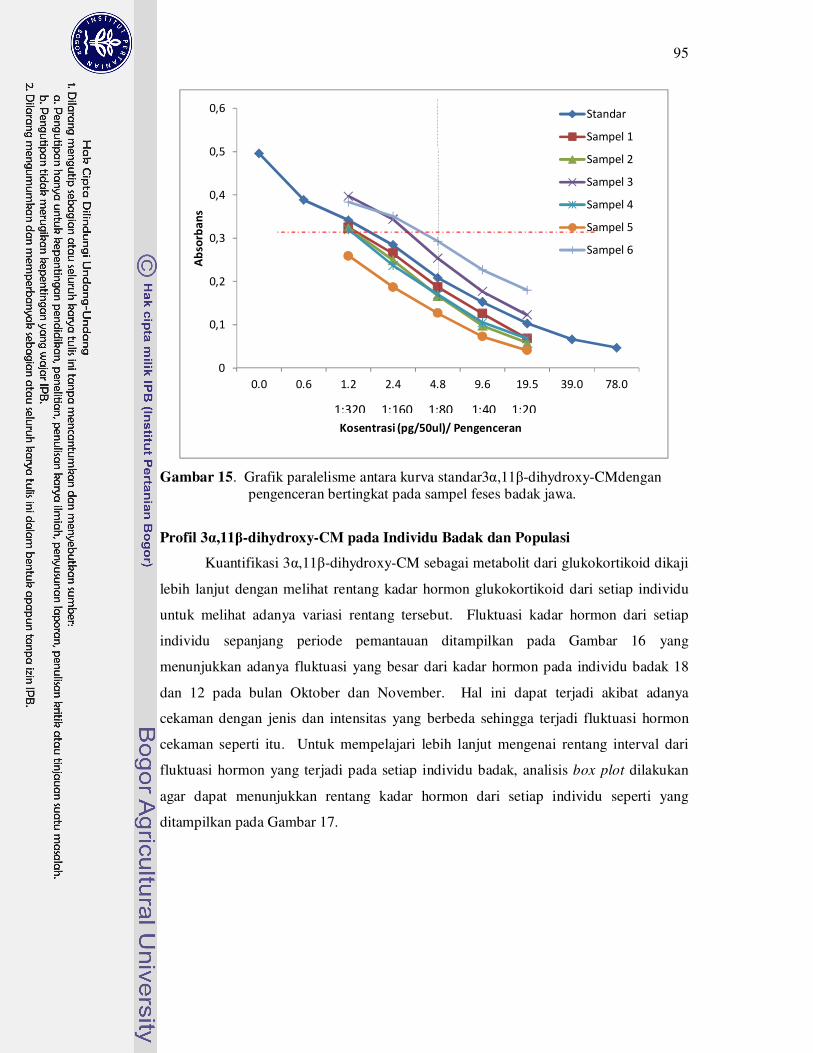
95
Gambar 15. Grafik paralelisme antara kurva standar3α,11β-dihydroxy-CMdengan
pengenceran bertingkat pada sampel feses badak jawa.
Profil 3α,11β-dihydroxy-CM pada Individu Badak dan Populasi
Kuantifikasi 3α,11β-dihydroxy-CM sebagai metabolit dari glukokortikoid dikaji
lebih lanjut dengan melihat rentang kadar hormon glukokortikoid dari setiap individu
untuk melihat adanya variasi rentang tersebut. Fluktuasi kadar hormon dari setiap
individu sepanjang periode pemantauan ditampilkan pada Gambar 16 yang
menunjukkan adanya fluktuasi yang besar dari kadar hormon pada individu badak 18
dan 12 pada bulan Oktober dan November. Hal ini dapat terjadi akibat adanya
cekaman dengan jenis dan intensitas yang berbeda sehingga terjadi fluktuasi hormon
cekaman seperti itu. Untuk mempelajari lebih lanjut mengenai rentang interval dari
fluktuasi hormon yang terjadi pada setiap individu badak, analisis box plot dilakukan
agar dapat menunjukkan rentang kadar hormon dari setiap individu seperti yang
ditampilkan pada Gambar 17.
0
0,1
0,2
0,3
0,4
0,5
0,6
0.0 0.6 1.2 2.4 4.8 9.6 19.5 39.0 78.0
Ab
sorb
an
s
Kosentrasi (pg/50ul)/ Pengenceran
Standar
Sampel 1
Sampel 2
Sampel 3
Sampel 4
Sampel 5
Sampel 6
1:320 1:160 1:80 1:40 1:20
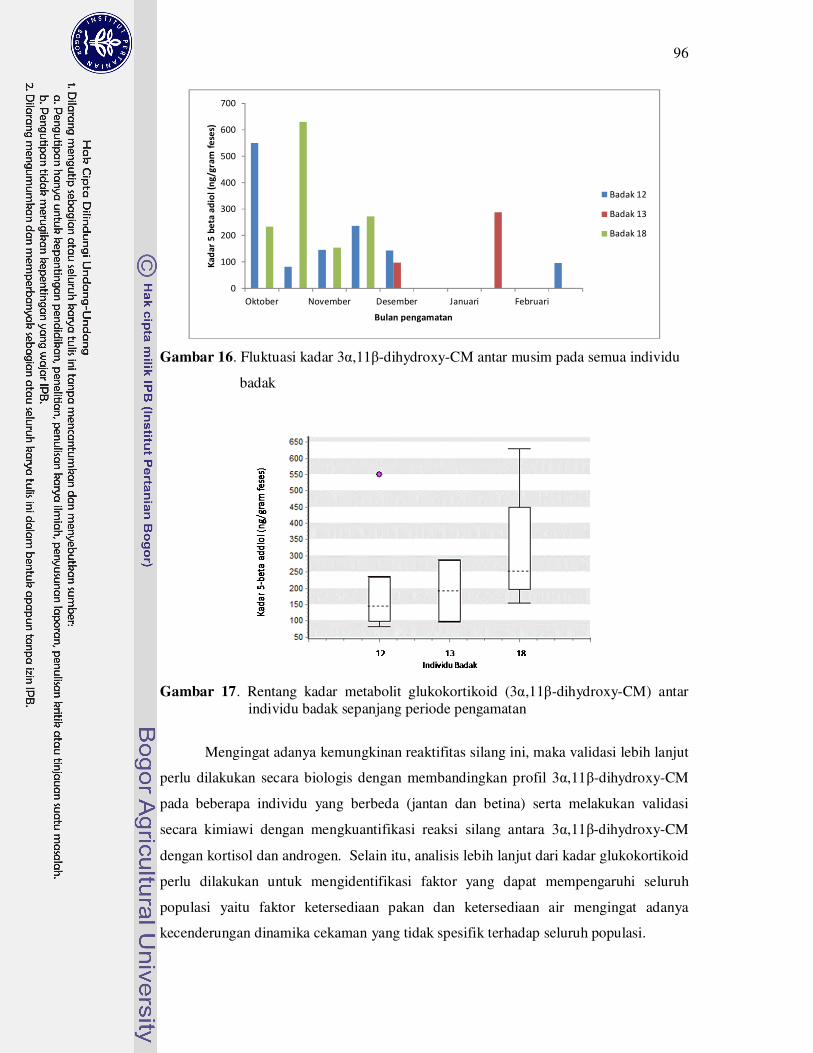
96
Gambar 16. Fluktuasi kadar 3α,11β-dihydroxy-CM antar musim pada semua individu
badak
Gambar 17. Rentang kadar metabolit glukokortikoid (3α,11β-dihydroxy-CM) antar
individu badak sepanjang periode pengamatan
Mengingat adanya kemungkinan reaktifitas silang ini, maka validasi lebih lanjut
perlu dilakukan secara biologis dengan membandingkan profil 3α,11β-dihydroxy-CM
pada beberapa individu yang berbeda (jantan dan betina) serta melakukan validasi
secara kimiawi dengan mengkuantifikasi reaksi silang antara 3α,11β-dihydroxy-CM
dengan kortisol dan androgen. Selain itu, analisis lebih lanjut dari kadar glukokortikoid
perlu dilakukan untuk mengidentifikasi faktor yang dapat mempengaruhi seluruh
populasi yaitu faktor ketersediaan pakan dan ketersediaan air mengingat adanya
kecenderungan dinamika cekaman yang tidak spesifik terhadap seluruh populasi.
0
100
200
300
400
500
600
700
Oktober November Desember Januari Februari
Ka
da
r 5
be
ta a
dio
l (n
g/g
ram
fe
ses)
Bulan pengamatan
Badak 12
Badak 13
Badak 18
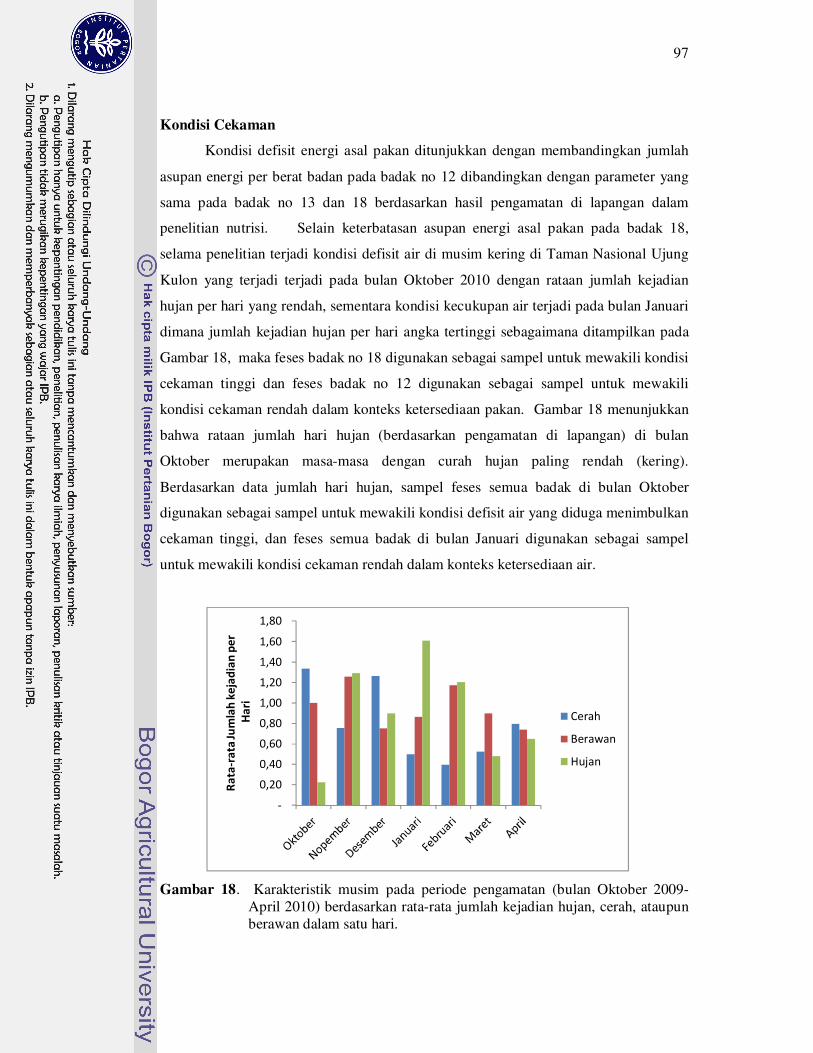
97
Kondisi Cekaman
Kondisi defisit energi asal pakan ditunjukkan dengan membandingkan jumlah
asupan energi per berat badan pada badak no 12 dibandingkan dengan parameter yang
sama pada badak no 13 dan 18 berdasarkan hasil pengamatan di lapangan dalam
penelitian nutrisi. Selain keterbatasan asupan energi asal pakan pada badak 18,
selama penelitian terjadi kondisi defisit air di musim kering di Taman Nasional Ujung
Kulon yang terjadi terjadi pada bulan Oktober 2010 dengan rataan jumlah kejadian
hujan per hari yang rendah, sementara kondisi kecukupan air terjadi pada bulan Januari
dimana jumlah kejadian hujan per hari angka tertinggi sebagaimana ditampilkan pada
Gambar 18, maka feses badak no 18 digunakan sebagai sampel untuk mewakili kondisi
cekaman tinggi dan feses badak no 12 digunakan sebagai sampel untuk mewakili
kondisi cekaman rendah dalam konteks ketersediaan pakan. Gambar 18 menunjukkan
bahwa rataan jumlah hari hujan (berdasarkan pengamatan di lapangan) di bulan
Oktober merupakan masa-masa dengan curah hujan paling rendah (kering).
Berdasarkan data jumlah hari hujan, sampel feses semua badak di bulan Oktober
digunakan sebagai sampel untuk mewakili kondisi defisit air yang diduga menimbulkan
cekaman tinggi, dan feses semua badak di bulan Januari digunakan sebagai sampel
untuk mewakili kondisi cekaman rendah dalam konteks ketersediaan air.
Gambar 18. Karakteristik musim pada periode pengamatan (bulan Oktober 2009-
April 2010) berdasarkan rata-rata jumlah kejadian hujan, cerah, ataupun
berawan dalam satu hari.
-
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
1,80
Ra
ta-r
ata
Ju
mla
hk
eja
dia
n p
er
Ha
ri
Cerah
Berawan
Hujan

98
Faktor Cekaman dan Profil Glukokortikoid
Statistik deskriptif menunjukkan adanya trend perbedaan rataan kadar hormon
antara badak 12, 13, 18 (yang memiliki perbedaan tingkat kecernaan dan konsumsi
energi per berat badan). Dari data ini dapat dilihat bahwa median tertinggi kadar
3α,11β-dihydroxy-CM ada pada individu badak 18, demikian juga dengan rentang
tertinggi dari fluktuasi kadar hormon tersebut ditunjukkan oleh individu 18. Hal ini
menunjukkan bahwa badak 18 menghadapi intensitas cekaman yang tinggi dan
berbeda-beda selama periode pengamatan. Individu 18 adalah seekor badak muda
(sub-adult) yang menunjukkan rataan asupan energi yang relatif lebih rendah
dibandingkan dengan badak lainnya. Selain itu, badak muda seperti ini sering
mendapatkan cekaman sosial dibandingkan dengan badak dewasa, dan cekaman seperti
ini sering terjadi saat badak muda berusaha menentukan ruang jelajahnya sendiri
(Hearne & Swart 1991). Intensitas cekaman juga menguat seiring dengan peningkatan
kadar testosteron pada jantan saat memasuki masa kematangan seksual (siklus
spermatogenesis) yang biasanya jatuh di awal musim hujan (Kretzschmar et al. 2002).
Hal yang perlu dicermati dari temuan tingginya kadar glukokortikoid pada badak 18 di
bulan November ini adalah kemungkinan adanya reaktifitas silang antara 3α,11β-
dihydroxy-CM dengan androgen. Badak 18 (badak jantan muda) kemungkinan
menghadapi masa peningkatan kematangan seksual (masa pubertas) yang ditandai
dengan meningkatnya hormon testosteron. Kemungkinan lain yang perlu diperhatikan
adalah perubahan kadar androgen yang berfluktuasi secara musiman (Paplinska et al.
2007).
Badak dengan rataan konsumsi energi per berat badan (%) yang rendah
cenderung menunjukkan profil rataan kadar 3α,11β-dihydroxy-CM yang tinggi di
dalam feses. Defisit energi sangat mungkin menjadi pemicu aktifitas glukokortikoid,
dimana glukokortikoid berperan dalam glukoneogenesis yang berperan dalam
pengadaan sumber energi tambahan untuk memasok siklus Kreb. Kemungkinan ini
diperkuat dengan adanya kecenderungan tingginya metabolit glukokortikoid pada
tingkat konsumsi energi yang rendah seperti yang ditunjukkan oleh grafik pada Gambar
19.

99
Gambar 19. Perbandingan tingkat konsumsi energi per berat badan (sumbu x) dengan
kadar metabolit glukokortikoid pada feses.
Demikian pula dengan juga rataan kadar hormon di musim dengan curah hujan
yang berbeda(perbedaan antar musim) dimanakadar hormon cenderung lebih tinggi
pada individu badak dan di musim kering di bulan Oktober sebagaimana ditampilkan
pada Gambar 20.
Gambar 20. Perbandingan kadar hormon cekaman antar individu badak dengan asupan
berat kering yang berbeda (A) dan antar musim/curah hujan (B)
Ketersediaan air merupakan salah satu faktor yang diduga sebagai pemicu
cekaman pada badak jawa, dan hal ini dibuktikan dengan analisis data yang
menunjukkan adanya hubungan antara jumlah curah hujan dengan kadar hormon
0
50
100
150
200
250
300
350
3,00% 3,20% 3,40% 3,60% 3,80%K
an
du
ng
an
3α
, 1
1β
-dih
yd
roxy
-CM
d
ala
m f
ese
s
(ng
/g f
ese
s)Konsumsi energi per berat badan (%)
0
50
100
150
200
250
300
350
400
0 0,5 1 1,5 2
Ra
taa
n k
ad
ar
3α
,11
β-d
ihy
dro
xy-C
M(n
g/g
ram
fese
s)
Rataan curah hujan (kejadian hujan per hari)

100
glukokortikoid. Rataan kadar glukokortikoid pada hari-hari dengan curah hujan rendah
(sekitar 0.3 kejadian hujan per hari) menunjukkan angka yang tinggi, sementara rataan
kadar glukokortikoid pada hari-hari dengan curah hujan tinggi (sekitar 1-1.5 kejadian
hujan per hari) menunjukkan angka yang rendah. Fakta seperti ini menunjukkan bahwa
rendahnya curah hujan memiliki implikasi terhadap menurunnya ketersediaan air (Smit
& Grant 2009) dan merupakan faktor pemicu cekaman pada badak jawa yang berujung
pada tingginya sekresi glukokortikoid pada periode tersebut.

101
Simpulan dan Saran
Simpulan
1. Kit komersial hormon kortisol dan kortikosteron tidak dapat digunakan untuk
menganalisa metabolit glukokorticoid dalam sampel feces badak Jawa karena
uji parelelisme menunjukkan bahwa kit tersebut tidak dapat mendeteksi
keberadaan metabolit glukokortikoid yang dieskresikan dalam feces
2. Hasil analisis dengan asai hormone 3α, 11β-dihydroxy-CM menunjukkan bahwa
kurva pengenceran bertingkat dari sampel feses badak jawa paralel dengan
kurva standar. Hal tersebut mengindikasikan kemungkinan asai hormon 3α,
11β-dihydroxy-CM dapat mendeteksi keberadaan metabolit glukokortikoid pada
feses badak jawa.
3. Profil kadar glukokortikoid pada setiap individu menunjukkan adanya indikasi
perbedaan dan kadar hormon tinggi pada badak 18 yang merefleksikan
dinamika cekaman yang lebih bervariasi dibandingkan dengan badak lainnya.
4. Faktor cekaman berupa keterbatasan asupan energi dan ketersediaan air
merupakan faktor yang diduga memicu terjadinya peningkatan kadar
glukokortikoid.
Saran
1. Melakukan validasi lebih lanjut terhadap 3α,11β-dihydroxy-CM untuk
menentukan potensi penggunaannya dalam pemantauan cekaman pada badak
jawa.
2. Melakukan validasi lanjutan untuk menentukan apakah hormone assay tersebut
dapat digunakan untuk menganalisa metabolit glucocorticoid dalam feces badak
jawa. (masih ada beberapa step validasi yang harus dilakukan untuk
memastikan apakah hormone assay tersebut tepat)
3. Mengembangkan protokol studi longitudinal monitoring fisiologi stress secara
non-invasif dengan menganalisa metabolit glukokortikoid dalam feces dengan
asai hormon yang tepat

102
Daftar Pustaka
Barja I, et al. 2007. Stress physiological response to tourist pressure in the wild
population of European pine marten. The journal of steroid biochemistry and
molecular biology. (104)3-5:136-142
Baron R. 2006. Mechanism of disease: neuropathic pain-a clinical perspective. Nature
clinical practice neurology. 2:95-106
Coenen M,. 2005. Exercise and stress: impact on adaptive processes involving water
and electrolytes. Livestock production science 92:131-145
Dickens MJ, Delehanty DJ, Romero LM,. 2010. Stress: an inevitable component of
animal translocation. Biological conservation 143:1329-1341
Francois-Gerard C, Gerard P, Rentier B,. 1988. Elucidation of non-parallel EIA curves.
Journal of immunological methods (111)1:59-65
Garg SL, Chander S,. 1997. Plasma Cortisol and thyroid hormone Concentrations in
Buffaloes with uterine Torsions.Buffalo BullettinVol 16(4): 75-76
Ghalib, Supriatna I, Agil M, Engelhardt A,. 2011. Non-invasive hormone monitoring:
fecal androgen and glucocorticoid in male crested macaques (Macacanigra) in
relation to seasonal and social factors. Thesis. Bogor Agriculture University
Hearne JW, Swart J,. 1991. Optimal translocation strategies for saving the black
rhinoceros. Ecological modelling 59: 279-292
Kretzschmar P, Gansloßer U, Dehnhard M,. 2004. Relationships between androgens,
environmental factors and reproductive behaviours in male white rhinoceros
(Ceratotherium simum simum). Hormones and Behaviour 45: 1-9
Koolman J, Rohm KH, Wanandi SI[ed], Sadikin M [ed]. 2001. Atlas berwarna dan teks
Biokimia. Cetakan I. Penerbit Hipokrates, Jakarta
Menargues A, Urios V, Mauri M,. 2008. Welfare assessments of captive asian
elephants (Elephas maximus) and Indian rhinoceros (Rhinoceros unicornis)
using salivary cortisol measurements. Animal Welfare (17): 305-312
Morgan KN, Tromborg CT. 2007. Sources of stress in captivity. Applied animal
behaviour science 102: 262-302
Morrow CJ, Kolver ES, Verkerk GA, Matthews LR,. 2000. Urinary corticosteroids: an
indicator of stress in dairy cattle. Proceedings of the New Zealand Society of
Animal Production 60: 218-221
Paplinska JZ, et al. 2007. Reproduction in male swamp wallabies (Wallabia bicolor):
puberty and the effects of season. Journal of Anatomy 211:518-533

103
Permadi YF,. 2008. Kajian dampak perubahan iklim terhadap kerentanan badak Jawa.
Laporan proyek WWF Indonesia.
Santymire RM, Armstrong, DM,. 2009. Development of a Field-Friendly Technique for
Fecal Steroid Extraction and Storage Using the African Wild Dog (Lycaon
pictus). Zoo Biology (28): 1 -14
Schoeman JP, Goddard A, Herrtage ME,. 2007. Serum cortisol and thyroxine
concentrations as predictors of death in critically ill puppies with parvoral
diarrhea. Journal of the American veterinary medical association (231)10:
1534-1539
Smit IPJ, Grant CC,. 2009. Managing surface-water in a large semi-arid savanna park:
effects on grazer distribution patterns. Journal for nature conservation 17(2):61-
71
Soto-Gamboa M, Gonzalez S, Hayes LD, Ebensperger LA,. 2009. Validation of a
radioimmunoassay for measuring fecal cortisol metabolites in the
Hystricomorph rodent, Octodon degus. Journal of experimental zoology
311A:496-503
Turner Jr JW, Tolson P, Hamad M,. 2002. Remote assessment of stress in white
rhinoceros (Ceratotherium simum) and black rhinoceros (Diceros bicornis) by
measurements of adrenal steroids in feces. Journal of zoo and wildlife medicine
33(3): 214-221
Wasser SK, et al. 2000. A generalized fecal glucocorticoid assay for use in diverse
array of nondomestic mammalian and avian species. General and comparative
endocrinology 120:260-275
Wingfield JC, Romero LM,. 2001. Adrenocortical responses to stress and their
modulation. Dalam: McEwen BS, Goodman HM (eds), Handbook of physiology
section 7: The endocrine system, coping with the environments: Neural and
endocrine mechanism vol. IV Oxford University Press New York: 211-234
Young KM, et al. 2004. Noninvasive monitoring of adrenocortical activity in
carnivores by fecal glucocorticoids analyses. General and comparative
endocrinology: 1-18
Ziegler TE, Wittwer DJ,. 2005. Fecal Steroid Research In the Field and In Laboratory:
Improved methods for Storage, Transport, Processing, and Analysis. American
Journal of Primatology 67: 159-174

KAJIAN KUANTITATIF MENGGUNAKAN HEWAN MODEL SEBAGAI
PEMBANDING UNTUK BADAK JAWA DALAM PENELITIAN CEKAMAN
AKIBAT DEFISIENSI PAKAN DAN DEFISIENSI AIR
Abstrak
Setelah mempelajari faktor cekaman yang dihadapi oleh badak jawa akibat
defisit pakan dan defisit air, maka penelitian ini dirancang untuk lebih mendalami
respons yang ditunjukkan oleh kuda (sebagai hewan model untuk badak) terhadap
cekaman tersebut. Penelitian ini bertujuan untuk melihat respons perilaku, fisiologis
(hematologi dan respirasi) pada hewan model kuda akibat cekaman defisit pakan dan
air 50% dari kebutuhan normal. Perlakuan yang diberikan adalah pakan kontrol sesuai
kebutuhan (K) dan 50% dari kebutuhan (50%K). Jumlah kuda yang dipakai terdiri dari
dua ekor kuda jantan berusia dewasa dan muda dengan bobot badan masing-masing
104 dan 98 kg. Induksi cekaman dilakukan pada dua ekor kuda model dengan
mengurangi asupan pakan secara bertahap sampai asupan pakan hanya 2.65% dari berat
tubuh kuda (defisit pakan) dan pengurangan air minum sampai 50% dari jumlah ad
libitum yang biasanya tersedia di kandang. Parameter yang diamati terdiri dari
hematologi rutin dan kadar kortisol darah yang diukur selama periode kontrol dan
perlakuan induksi cekaman. Hasil penelitian menunjukkan bahwa kadar kortisol dan
hematologi (rasio netrofil:limfosit) menunjukkan bahwa defisit pakan merupakan
cekaman akut yang dapat diatasi oleh kuda (ditandai dengan penurunan kortisol pada
penghujung perlakuan). Kadar kortisol dan hematologi (rasio netrofil:limfosit)
menunjukkan bahwa defisit air merupakan cekaman yang bersifat kronis dan tidak
dapat diatasi dengan mudah. Dengan defisit pakan dan air, kedua ekor kuda
menunjukkan perubahan-perubahan: perilaku (mengurangi aktifitas berjalan dan
melakukan aktifitas menggigit ember/pagar / crib biting), peningkatan kadar kortisol,
hematologi (perubahan rasio Netrofil/limfosit yang menunjukkan adanya migrasi
limfosit/imunosupresi saat cekaman), dan peningkatan respirasi pada kedua ekor kuda
saat mengalami cekaman.
Kata kunci: badak jawa, hewan model, cekaman, kortisol, perilaku

106
Quantitiative study using animal model for javan rhinoceros in research for stress
due to feed and water deprivations
Abstract
After learning that feed and water limitations are the major factores causing
stress in javan rhinoceros, this research is designed to further study the possible
physiological responses from horse (as a model for the rhinoceros) in the presence of
the stress. This research is designed to study the changes in the behaviour, blood
cortisol level, hematology (neutrophil/limphocyte ratio), and respiration rate.
Induction of stress is done by reducing the ration down to the level of 50% of the
normal supply. Treatments consist of : control for feed (K) and reduction of feed to
50% (50%K) of the normal ad libitum quantity. Two horses are used in this study
consisting of one adult and one juvenile weighing 104 kg and 98 kg respectively.
Induction of stress is done by reduing the feed intake to only 2.65% dry matter intake
of the body weight (feed deprivation), and reducing the water supply to 50% of the
normal supply. Parameters include: behaviour, blood cortisol, hematology, and
respiration rate measured thorughout the control and treatments. The result shows
thatcortisol level and hematology (Neutrophil:lymphocyte ratio) suggests that feed
deprivation is an acute stress that can be overcome (marked with reduction of cortisol
level at the end of treatment). The same parameters show that water deprivation is a
chronic stress that cannot be readily overcome. With water and feed deprivation, both
horses show changes in behaviour such as reduction of walking and crib biting
activities, increase in cortisol level in blood, as well as the changes in neutrophil :
lymphocyte ratio suggesting that there are migration of lymphocytes from the blood
stream, as well as immunosuppression during the stress conditions. The result also
shows increase in respiration rate in both horses under stress.
Keywords: javan rhinoceros, model animal, stress, cortisol, behaviour

107
Pendahuluan
Respons badak terhadap cekaman merupakan salah satu aspek penting yang
perlu dipelajari agar dampak dari cekaman tersebut dapat diantisipasi dan upaya
mitigasi cekaman dapat dilakukan untuk menunjang pelestarian spesies ini. Namun
demikian, induksi cekaman secara langsung pada badak jawa tidak mungkin dilakukan
karena tingkat kesulitan untuk menemukan badak jawa secara langsung dan juga status
spesies tersebut yang sudah terancam punah menyebabkan uji coba yang berpotensi
untuk membahayakan kehidupan badak jawa tidak dapat dilakukan. Oleh karena itu
kebutuhan akan penelitian kuantitatif berdasarkan hasil kajian sumber cekaman secara
kualitatif di habitat alami badak jawa muncul.
Kebutuhan ini melandasi konsep pendekatan baru dengan menggunakan hewan
model sebagai pembanding bagai badak jawa. Sesuai dengan kaidah penggunaan
hewan model pada ilmu biomedis hewan, jenis hewan model dipilih berdasarkan
berbagai kecocokan seperti: kedekatan secara genetis, kemiripan anatomi, dan
kemiripan perilaku. Informasi dari berbagai sumber memberikan informasi mengenai:
genetika badak jawa dan kekerabatan dengan badak sumatera, tapir, dan kuda
(Fernando et al. 2004); anatomi saluran pencernaan badak sebagai hindgut fermenter
yang mengandalkan proses fermentasi pada saluran pencernaan bagian belakang atau
sekum (Clauss et al. 2005; Clauss et al. 2006); dan perilaku badak jawa yang diamati
langsung di lapangan (Sajudin & Djaja 1984) ataupun yang diamati dengan
menggunakan kamera jebak (Hariyadi et al. 2010).
Studi kualitatif maupun kuantitatif untuk membandingkan perilaku hewan
dapatdisusun dalam sebuah ethogram yang kemudian menjadi dasar (baseline) yang
dibandingkan penelitian. Ethogram merupakan perangkat yang kerap digunakan oleh
pengamat perilaku untuk dapat mengklasifikasikan berbagai perilaku hewan serta
memungkinkannya untuk dapat dianalisis lebih lanjut secara kuantitatif sehingga
analisis perilaku dapat memberikan informasi yang obyektif (Colgan 1978).
Penggunaan ethogram pada penelitian ini dimaksudkan untuk dapat mengklasifikasikan
perilaku badak jawa, badak Sumatra, dan kuda serta memungkinkan perbandingan yang
obyektif pada penggolongan tersebut. Pemilihan hewan model dilakukan untuk
membandingkan sistem pencernaan (saluran digesti) dengan fokus pada proses
kecernaan yang terjadi setelah proses pencernaan di lambung. Oleh karena itu,
kesamaan sistem pencernaan pada kuda dan badak seperti yang ditampilkan pada

108
Gambar 21 merupakan salah satu kriteria penting pemilihan kuda sebagai hewan model
untuk aspek kecernaan pada badak jawa.
A B
Gambar 21. Perbandingan anatomi sistem pencernaan kuda (A) dan badak (B) yang
menunjukkan karakteristik hewan monogastrik (lambung satu) dan
keberadaan sekum untuk pencernaan selulosa. Sumber:
http.wren.aps.uo.guelph.ca.
Kriteria pemilihan hewan model dilakukan pula dengan mengidentifikasi
parameter perilaku yang tepat, sehingga perlakuan dan induksi cekaman pada hewan
model akan dapat memberikan gambaran perubahan perilaku serta fisiologis yang
kompatibel dan dapat diterapkan pada badak jawa.
Proses Pemilihan Hewan Model
Hewan model untuk badak jawa dipilih berdasarkan beberapa kriteria yang
terdiri dari: kedekatan genetika, kemiripan anatomi, kemiripan kimia darah, dan
kemiripan perilaku. Perbandingan kriteria di atas dilakukan melalui studi literatur yang
memberikan informasi mengenai genetika, anatomi, kimia darah, dan perilaku dari
masing-masing kandidat hewan model. Kandidat model dalam penelitian ini terdiri
dari dua kelompok hewan yaitu: (1) kelompok hewan model yang lazim digunakan
dalam studi biomedis yang terdiri dari tikus, kambing, dan kuda; serta (2) kelompok
hewan badak: Badak sumatera dan badak india.
1) Kelompok hewan model konvensional
Tikus banyak digunakan dalam kajian mengenai cekaman kronis dan akut
(Figuireido et al. 2003),dampak dari defisit pakan (Nessmithet al. 1983),dan dampak
dari defisit air (Cameron & Perdue 2005), namun tikus tidak memiliki sistim
Sekum

109
pencernaan yang mirip dengan badak sehingga dikhawatirkan tidak dapat memberikan
gambaran yang obyektif terhadap penyerapan nutrisi yang terjadi pada badak.
Kekurangan ini menyebabkan tikus tidak dipilih sebagai hewan model untuk badak
jawa.
Kambing juga banyak digunakan dalam penelitian mengenai ruminal stasis
akibat keracunan (McSweeney & Pass 1983) dan gangguan pencernaan akibat
keracunan (Pass et al. 1979), namun kambing merupakan hewan ruminan yang
memiliki sistem pencernaan yang jauh berbeda dengan badak. Perbedaan anatomi
pencernaan ini menyebabkan kambing tidak dipilih sebagai hewan model bagi badak
jawa.
Kuda telah banyak digunakan dalam penelitian terkait cekaman (Coenen 2005),
kecernaan dan penyerapan karbohidrat (Pagan et al. 1998; Dyer et al. 2002), serta
fluktuasi hormon (Glade et al. 1984). Kuda memiliki kimia darah yang serupa dengan
badak dalam konteks kadar tyrosin bebas di dalam sel darah merah (Harley et al. 2004),
dan juga kemiripan dalam anatomi saluran pencernaan sebagai hindgut fermenter
(Clauss et al. 2005; Clauss et al. 2006). Dari kelompok hewan model konvensional,
kuda memiliki kekerabatan genetik yang paling dekat dengan badak jawa, dan
berdasarkan informasi ini kuda dipilih sebagai hewan model yang paling memadai
untuk badak jawa.
2) Kelompok hewan badak (Rhinoceratidae)
Badak india telah banyak digunakan dalam berbagai penelitian terkait kecernaan
(Clauss et al. 2005; Clauss et al. 2006), hormon cekaman (Carlstead & Brown 2005),
dan perilaku (Hutchin & Kreger 2006). Badak india memiliki kekerabatan yang sangat
dekat dengan badak jawa, namun status spesies badak ini masih ada di dalam kategori
terancam punah sehingga perlakuan intensif ataupun induksi cekaman tidak dapat
dilakukan pada hewan dengan status seperti ini.
Badak sumatera telah digunakan dalam studi perilaku, kecernaan, dan kajian
hormon (Agil et al. 2007; Agil et al. 2008). Badak sumatera memiliki kekerabatan
genetika yang tidak terlalu dekat dengan badak jawa, namun spesies badak ini dapat
dengan mudah diamati di fasilitas Suaka badak sumatera di Way Kambas. Spesies
badak ini berada dalam kategor sangat terancam punah (Critically endangered)
sehingga tidak mungkin untuk dijadikan obyek penelitian intensif dan induksi cekaman.
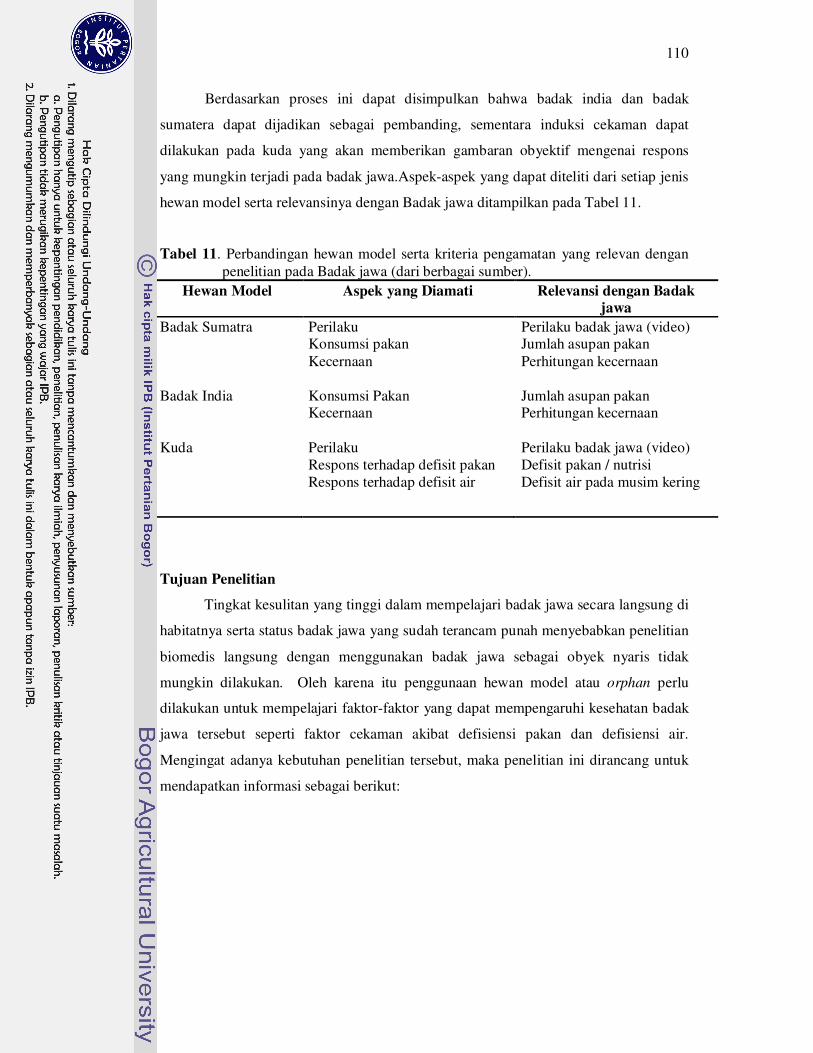
110
Berdasarkan proses ini dapat disimpulkan bahwa badak india dan badak
sumatera dapat dijadikan sebagai pembanding, sementara induksi cekaman dapat
dilakukan pada kuda yang akan memberikan gambaran obyektif mengenai respons
yang mungkin terjadi pada badak jawa.Aspek-aspek yang dapat diteliti dari setiap jenis
hewan model serta relevansinya dengan Badak jawa ditampilkan pada Tabel 11.
Tabel 11. Perbandingan hewan model serta kriteria pengamatan yang relevan dengan
penelitian pada Badak jawa (dari berbagai sumber).
Hewan Model Aspek yang Diamati Relevansi dengan Badak
jawa
Badak Sumatra Perilaku
Konsumsi pakan
Kecernaan
Perilaku badak jawa (video)
Jumlah asupan pakan
Perhitungan kecernaan
Badak India Konsumsi Pakan
Kecernaan
Jumlah asupan pakan
Perhitungan kecernaan
Kuda Perilaku
Respons terhadap defisit pakan
Respons terhadap defisit air
Perilaku badak jawa (video)
Defisit pakan / nutrisi
Defisit air pada musim kering
Tujuan Penelitian
Tingkat kesulitan yang tinggi dalam mempelajari badak jawa secara langsung di
habitatnya serta status badak jawa yang sudah terancam punah menyebabkan penelitian
biomedis langsung dengan menggunakan badak jawa sebagai obyek nyaris tidak
mungkin dilakukan. Oleh karena itu penggunaan hewan model atau orphan perlu
dilakukan untuk mempelajari faktor-faktor yang dapat mempengaruhi kesehatan badak
jawa tersebut seperti faktor cekaman akibat defisiensi pakan dan defisiensi air.
Mengingat adanya kebutuhan penelitian tersebut, maka penelitian ini dirancang untuk
mendapatkan informasi sebagai berikut:

111
1. Kompilasi dan dinamika perilaku kuda pada kondisi cekaman defisit
pakan dan defisit air.
2. Dinamika hormon kortisol/glukokortikoid dan hematologi terhadap
cekaman berupa defisiensi pakan dan air.
3. Dinamika pola respirasi dan kecernaan kuda dalam kondisi cekaman.
Adapun hipotesis yang akan diuji dalam penelitian ini adalah: model untuk
penelitian badak jawa dapat dilakukan dengan menggunakan kuda sebagai pembanding
dalam aspek kecernaan (posterior dari esophagus) dengan adanya respons (sensitifitas)
terhadap perlakuan defisit air dan pakan (profil hormon, kecernaan, dan respirasi).

112
Bahan dan Metode
Perbandingan Perilaku
Perilaku badak dan hewan model pembandingnya (badak sumatera dan kuda)
perlu disusun dalam sebuah ethogram yang kemudian menjadi dasar (baseline) yang
dibandingkan dengan perilaku badak jawa sebagai contoh dalam penelitian ini.
Ethogram merupakan perangkat yang kerap digunakan oleh pengamat perilaku untuk
dapat mengklasifikasikan berbagai perilaku hewan serta memungkinkannya untuk
dapat dianalisis lebih lanjut secara kuantitatif sehingga analisis perilaku dapat
memberikan informasi yang obyektif sesuai dengan penjelasan Colgan (1978).
Penggunaan ethogram pada penelitian ini dimaksudkan untuk dapat mengklasifikasikan
perilaku badak Jawa, badak Sumatra, dan kuda serta memungkinkan perbandingan
yang obyektif pada penggolongan tersebut.
Perilaku badak Jawa dapat dengan mudah diamati menggunakan kamera video
otomatis yang digunakan oleh Balai Taman Nasional Ujung Kulon dan WWF Indonesia
dalam memantau populasi badak jawa di kawasan Taman Nasional Ujung Kulon
(Hariyadi et al. 2010). Melalui pemilihan hewan model serta pemilihan parameter
perilaku yang tepat, perlakuan dan induksi cekaman pada hewan model akan dapat
memberikan gambaran perubahan perilaku serta fisiologis yang kompatibel dan dapat
diterapkan pada badak jawa.
Dua ekor badak sumatera (Dicerorhinus sumatrensis) dan dua ekor kuda sumba
berkelamin jantan dipilih sebagai hewan model yang digunakan sebagai model perilaku
untuk dibandingkan dengan perilaku pada badak jawa yang sudah tercatat dalam
penelitian kamera jebak (Hariyadi et al. 2010). Tabel perilaku badak sumatera dibuat
berdasarkan informasi berdasarkan kompilasi oleh Siswandi(2005) yang mengamati
perilaku badak sumatera jantan Torgamba dan Andalas di Taman Nasional Way
Kambas.Dua ekor kuda jantan (satu ekor dewasa dan satu ekor kuda muda) dengan
berat masing-masing 104 kg dan 98 kg dipilih sebagai hewan model untuk menjalani
perlakuan induksi cekaman dengan mengurangi ransum pakan 50% dari jumlah yang
biasanya tersedia di kandang (defisit pakan) dan mengurangi ransum air 50% dari
jumlah yang biasanya tersedia di kandang (defisit air).
Perilaku kuda dikompilasi dengan melakukan pengamatan dan menyusun tabel
perilaku dua ekor kuda sumba yang menjadi obyek penelitian di Rumah Sakit Hewan

113
Fakultas Kedokteran Hewan IPB Darmaga dengan dasar ijin dari komisi etik no: 05-
2011 RSH-IPB untuk melakukan induksi cekaman.
Gambar 22 menampilkan profil kedua ekor kuda jantan yang digunakan dalam
penelitian ini yang terdiri dari kuda jantan dewasa bernama Garuda dan kuda jantan
muda bernama Elang.Untuk menyesuaikan dengan ketersediaan data perilaku pada
badak jawa dan badak sumatera, perilaku kuda dicatat pada pagi, siang/sore, dan malam
hari baik saat kuda dalam keadaan bebas sebelum penelitian, maupun dalam periode
pencucian atau washing out setelah perlakuan defisit pakan.
A B
Gambar 22. Kuda jantan dewasa bernama Garuda (A) dan kuda jantan dewasa muda
bernama Elang (B) yang digunakan sebagai hewan model dalam
pengamatan di Rumah Sakit Hewan IPB Darmaga
Pengambilan Data Pakan dan Air Minum (Fase Kontrol)
Sebelum perlakuan defisit pakan dan defisit air diberikan kepada kuda yang
menjadi hewan model dalam penelitian ini terlebih dahulu dilakukan pencatatan jadwal
dan kuantitas pakan yang diberikan sehari-hari dalam keadaan normal. Kedua ekor
kuda model ditempatkan dalam kandang untuk memastikan bahwa konsumsi pakan dan
air dapat terpantau dan tercatat secara obyektif.
Kedua ekor kuda model ditempatkan di dalam kandang dan diberi waktu enam
hariyang cukup untuk aklimatisasi (Noble et al. 2007) sebelum pemberian pakan dan
air, dan sebelum pengamatan perilaku dilakukan. Sampel darah diambil dari vena
jugularis kuda model untuk mendapatkan data hematologi rutin, dan juga kadar kortisol
pada darah. Pemeriksaan darah dilakukan di Pusat Studi Satwa Primata (PSSP) IPB.
Pengambilan darah dilakukan pada saat kuda baru dimasukkan ke kandang dan enam
hari kemudian setelah masa aklimatisasi berakhir, sementara pengamatan perilaku
dilakukan pada pagi dan siang hari dalam dua hari selama periode aklimatisasi.

114
Kuda diberi pakan ransum yang terdiri dari pelet (Royal Horse) dan bran
(Bogasari) dengan perbandingan 1:2 (pelet:bran) yang dicampur dengan 800 ml air.
Selain itu, kuda juga diberi rumput segar sebanyak 14.8-14.9 kg/e/h yang diambil dari
lapangan tempat kedua ekor kuda tersebut makan dalam di luar kandang dalam keadaan
normal. Komposisi pakan seperti ini diberikan setiap dua hari pada kedua ekor kuda
model. Air minum diberikan dalam bak minum yang diisi penuh dengan menggunakan
ember takar berukuran delapan liter setiap tiga hari. Pemberian pakan dan air minum
secara normal ini dilakukan dalam periode enam hari.
Induksi Cekaman
1. Perlakuan defisiensi Pakan
Untuk mensimulasikan kondisi defisit pakan yang terjadi pada badak jawa,
maka pengurangan ransum pakan (perlakuan defisit pakan) dilakukan pada kedua ekor
kuda jantan yang digunakan sebagai hewan model dalam penelitian ini. Perlakuan
defisit pakan dilakukan secara bertahap dengan mengurangi jumlah ransum pakan yang
diberikan menjadi 70% dan akhirnya menjadi 50% dari jumlah yang biasanya
diberikan. Dengan berat badan kuda sebesar 104 kg untuk kuda dewasa (Garuda) dan
98 kg untuk kuda muda (Elang), pengurangan yang dilakukan dirancang untuk
menurunkan asupan pakan kering kuda dari 5.34%-5.56% berat badan menjadi 2.65%-
2.76% berat badan.Tingkat asupan ini berbeda dengan asupan kering pakan badak yang
berada pada 1% berat badan. Pengurangan jatah pakan kuda ini tidak dilakukan untuk
menyamai secara persis kondisi yang dihadapi oleh badak dengan mempertimbangkan
etika penggunaan hewan model. Pengurangan pakan dilakukan secara bertahap agar
tidak menimbulkan perubahan yang dapat mengakibatkan kolik (Evans et al. 1990) dan
juga dengan mempertimbangkan agar total asupan pakan kering tidak jatuh di bawah
2.35% dari berat badan kuda. NRC (1989) menyebutkan bahwa kebutuhan minimun
bagi seekor kuda untuk mempertahankan fungsi normalnya adalah dengan asupan
kering 2.35% setiap hari, oleh karena itu, asupan kering di bawah 2.35% dikhawatirkan
akan menimbulkan defisit asupan nutrien yang akan berdampak negatif.Pengurangan
pakan secara bertahap seperti ini dilakukan dengan memperhitungkan faktor
kesejahteraan hewan model (animal welfare).
Pada hari pertama sampai hari ketiga pada perlakuan defisit pakan ini, setiap
ekor kuda model menerima pakan 70% dari biasanya yaitu konsumsi bahan kering
(KBK) sebesar 3,41 g/ek/hari atau 3.28% berat badan kuda dewasa (Garuda) dan 3.48%

115
berat badan kuda muda (Elang) . Kemudian pada hari keempat sampai keenam jumlah
asupan kering dikurangi menjadi 50% dari yang biasa diberikan yaitu konsumsi bahan
kering (KBK) sebesar 2.7 g/ek/hari atau 2.6% berat badan kuda dewasa (Garuda) dan
2.76% berat badak kuda muda (Elang). Dalam setiap perlakuan (kontrol dan defisiensi
pakan) pengamatan perilaku kuda model dilakukan pada pagi, dan siang/sore pada hari
keempat, dan keenam.
Pengambilan sampel darah dilakukan pada harikeempat dan ketujuh dengan
mengambil darah dari vena jugularis. Sampel darah kemudian dikirim ke laboratorium
Pusat Studi Satwa Primata IPB untuk menjalani analisis: hematologi rutin, fungsi ginjal
(ureum, kreatinin), dan hormon kortisol.
2. Perlakuan Defisiensi Air
Untuk mensimulasikan kondisi defisit air yang kerap dialami oleh badak jawa,
maka kedua ekor kuda model mendapatkan perlakuan berupa pengurangan jatah air
minum harian selama enam hari. Baseline data air minum didapatkan dari informasi
yang tercatat selama periode kontrol adalah pemberian rata-rata 16 liter setiap hari dan
pengurangan jumlah ketersediaan air menjadi 50% dilakukan dengan memberikan air
satu kali setiap dua hari (16 liter air pada hari berselang). Jadwal pemberian air selama
periode uji coba defisiensi air disajikan dalam Tabel 12.
Perlakuan dilakukan selama enam hari dan pengamatan perilaku dilakukan pada
pagi, dan siang/sore di hari ketiga dan hari keenam. Pengambilan sampel darah
dilakukan dua kali yaitu pada hari kedua dan ketujuh dengan mengambil darah dari
vena jugularis. Sampel darah kemudian dikirim ke laboratorium Pusat Studi Satwa
Primata IPB untuk menjalani analisis: hematologi rutin, fungsi ginjal (ureum,
kreatinin), dan hormon kortisol.
Tabel 12. Pemberian air minum untuk setiap ekor kuda pada periode perlakuan defisit
air
Kuda 100% air
(l/hari)
50%air
(l/hari)
Garuda 16 8
Elang 16 8

116
Periode “Pencucian”
Periode pencucian bukanlah bagian dari perlakuan, namun proses ini dilakukan
setelah perlakuan defisit pakan dan sebelum perlakuan defisit air untuk memastikan
bahwa kedua ekor kuda model terbebas dari pengaruh perlakuan defisit pakan dan
pengandangan yang dijalani sebelumnya. Hal ini dilakukan dengan melepaskan kuda
model dari kandangnya dan membiarkan mereka beraktifitas dan merumput seperti
biasanya. Periode ini dilakukan selama tiga hari untuk memungkinkan kuda
beradaptasi kembali dengan kondisi dan lingkungannya. Rivera et al.
(2002)menunjukkan bahwa kuda sudah menunjukkan perubahan kadar kortisol ke
tingkat basal pada hari ke-empat, oleh karena itu, masa “pencucian” yang dilakukan
selama tiga hari sudah mencukupi untuk mengembalikan kadar kortisol ke tingkat awal
(aklimatisasi) untuk kemudian dibandingkan dengan kadar kortisol setelah induksi
cekaman.
Pengamatan Pola Respirasi
Respirasi pada kuda dihitung berdasarkan pergerakan pada bagian samping
badan (rusuk dan abdomen) pada saat respirasi berlangsung. Satu siklus respirasi
dihitung sebagai satu proses yang dimulai dari penghirupan udara yang ditandai dengan
mengembangnya rongga dada dan abdomen dan diakhiri dengan penghembusan udara
yang ditandai dengan menyusutnya rongga dada dan abdomen. Penghitungan siklus
respirasi dilakukan dengan mengamati kuda di dalam kandang dalam waktu 30 detik
sambil menghitung siklus respirasi yang terjadi dalam periode pengamatan, data ini
kemudian dikonversikan menjadi jumlah siklus per menit (respirasi per menit) dengan
mengalikan jumlah respirasi dalam 30 detik dengan dua.
Penghitungan Konsumsi dan Kecernaan kuda
Konsumsi dihitung dengan menimbang, mencatat, dan menjumlahkan berat
pakan segar harian yang diberikan kepada setiap ekor kuda di kandang. Konsumsi
pakan kuda terdiri dari rumput, pelet, dan dedak yang diberikan setiap hari dan
ditimbang menggunakan timbangan top loading merk Oxone dengan kapasitas
maksimum 15 kg. Jumlah pemberian pakan dicatat setiap hari dan kemudian rataan
pemberian pakan per ekor per hari dilakukan berdasarkan jumlah hari perlakuan.
Untuk melengkapi informasi mengenai perubahan pada kecernaan yang terjadi
dalam kondisi cekaman, feses kuda dikumpulkan untuk kemudian menjalani proses

117
gravimetri untuk mendapatkan kadar Acid Insoluble Ash (AIA) pada feses sebelum dan
setelah perlakuan induksi cekaman untuk mempelajari adanya perubahan kecernaan
sebagai respons terhadap adanya cekaman defisit pakan maupun air. Penghitungan
kecernaan dilakukan dengan metoda VanKeulen & Young (1977) dengan koreksi 10%
berdasarkan Mainka et al. (1989) dan Sims et al. (2007).
Analisis Data
Analisis perilaku dilakukan dengan menggunakan statistik deskriptif untuk
membandingkan trend perilaku pada kedua ekor kuda model pada fase kontrol dan pada
saat diberikan perlakuan. Statistik deskriptif juga akan dilakukan pada hasil
laboratorium untuk hematologi (rasio netrofil : limfosit), dan cekaman (berdasarkan
kadar kortisol dalam darah).Statistik deskriptif digunakan untuk memperlajari pola
kecenderungan pada respirasi sebelum dan setelah induksi cekaman akibat defisit pakan
dan defisit air. Regresi linear berganda dilakukan untuk mengidentifikasi komponen
yang paling berperan dalam menimbulkan gejala-gejala terkait cekaman.
Penguatan analogi antara kuda dengan badak dilakukan dengan
membandingkan trend peningkatan respirasi dan kecernaan. Data tingkat respirasi dan
kecernaan kuda dalam kondisi normal didapat dari hasil studi literatur, sementara data
tingkat respirasi dan kecernaan kuda dalam kondisi cekaman diperoleh dengan
mengukur respirasi kuda selama masa percobaan dan mengukur kecernaan berdasarkan
metoda AIA agar hasilnya dapat dibandingkan dengan data pada badak. Data tingkat
respirasi badak dalam keadaan normal dan dalam kondisi cekaman diperoleh dari hasil
pengamatan menggunakan rekaman video (Hariyadi et al. 2010).
Penguatan analogi hewan model dengan badak dilakukan dengan
membandingkan hasil penelitian menggunakan hewan model dengan parameter yang
sama pada badak berdasarkan penelitian sebelumnya dan data-data dari studi literatur.
Kemiripan trend perilaku dan parameter lain (respirasi, kecernaan, glukokortikoid)
dijadikan landasan untuk menguatkan analogi antara kuda dengan badak.

118
Hasil dan Pembahasan
Pemilihan Hewan Model
Berdasarkan proses pemilihan hewan model badak sumatera (Dicerorhinus
sumatrensis), badak India (Rhinoceros unicornis) dipilih sebagai pembanding untuk
mendapatkan parameter perilaku dan kecernaan, sementara kuda (Equus caballus)
dipilih sebagai hewan model yang dapat digunakan untuk perlakuan berupa induksi
cekaman mensimulasi kondisi yang dihadapi badak jawa di habitatnya. Kesesuaian
kuda sebagai hewan model untuk badak jawa diperkuat dengan adanya persamaan
perilaku antara kuda dan badak.
Perbandingan Perilaku Hewan Model
Perilaku hewan model (badak sumatera dan kuda) disusun dalam sebuah
ethogram yang kemudian menjadi dasar (baseline) yang dibandingkan dengan perilaku
badak jawa yang dijadikan contoh dalam penelitian ini. Ethogram perilaku ketiga
spesies ini disajikan dalam Tabel 13, sementara kuantitas aktifitas (durasi waktu) pada
badak dan kuda disajikan dalam Gambar 23 dan Gambar 24. Ethogram yang
ditampilkan menunjukkan bahwa ketiga spesies hewan di atas memiliki kesamaan
perilaku berupa: Jalan maju, Bersuara/dengus, garuk/gosok muka, dan makan/minum.
Kuda dan badak jawa memiliki kesamaan perilaku yang terdiri dari: Jalan maju, gerak
kepala, gerak ekor, jalan mundur, gerak satu kaki, bersuara/dengus, gosok muka, dan
makan/minum. Dari 23 perilaku (event behaviour) yang tercatat dalam ethogram
terlihat bahwa badak jawa memiliki 14 (60.87%) kesamaan perilaku dengan kuda dan 9
(39%) kesamaan perilaku dengan badak sumatera. Fakta ini menguatkan analogi antara
kuda dan badak jawa dalam pemilihan kuda sebagai hewan model.
Perbandingan perilaku pada badak jawa dan pada kuda menunjukkan adanya
perbedaan pada durasi aktifitas yang tercatat. Badak jawa menunjukkan bahwa
aktifitas lokomotor (bergerak dan berjalan) merupakan aktifitas yang dominan,
sedangkan pada kuda makan merupakan aktifitas yang dominan dilakukan oleh kuda
dalam kondisi lepas kandang. Di dalam kandang kuda menunjukkan aktifitas berdiri
diam sebagai aktifitas yang dominan, dan aktifitas berdiri diam seperti ini juga banyak
dilakukan oleh badak jawa sebagaimana terekam melalui video yang dipasang di
Taman Nasional Ujung Kulon untuk memantau perilaku badak jawa (Hariyadi et al.
2010).
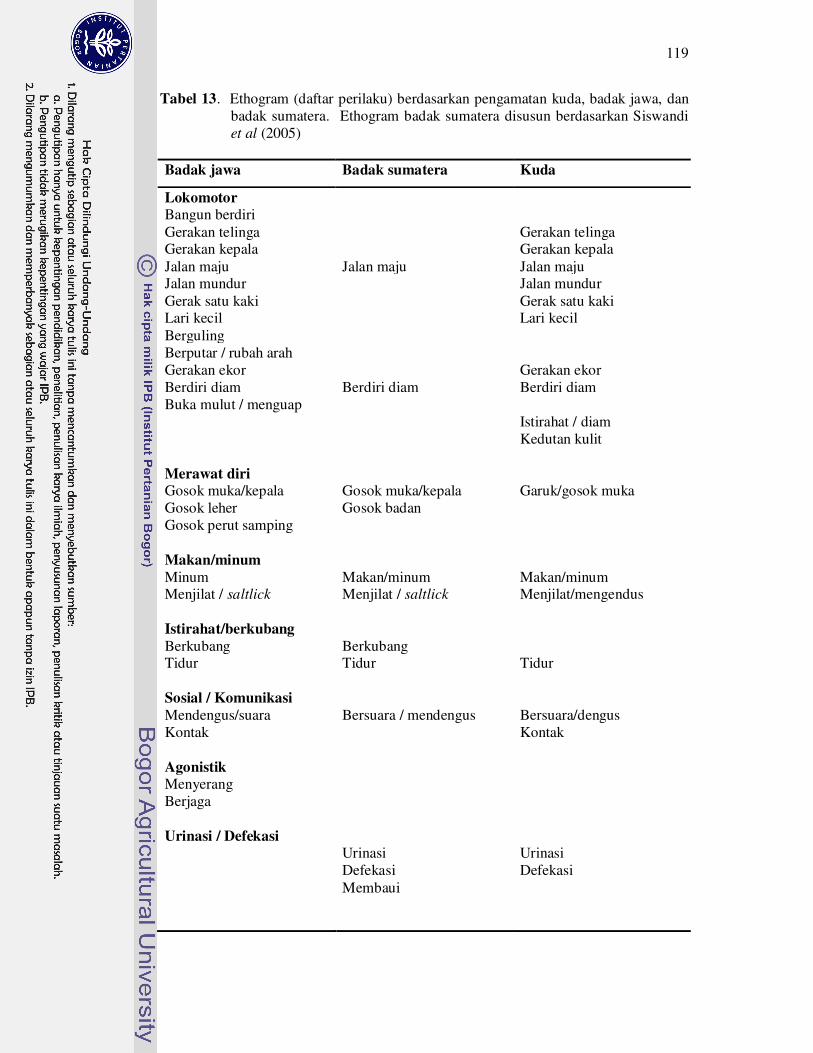
119
Tabel 13. Ethogram (daftar perilaku) berdasarkan pengamatan kuda, badak jawa, dan
badak sumatera. Ethogram badak sumatera disusun berdasarkan Siswandi
et al (2005)
Badak jawa Badak sumatera Kuda
Lokomotor
Bangun berdiri
Gerakan telinga
Gerakan kepala
Jalan maju
Jalan maju
Gerakan telinga
Gerakan kepala
Jalan maju
Jalan mundur Jalan mundur
Gerak satu kaki Gerak satu kaki
Lari kecil
Berguling
Lari kecil
Berputar / rubah arah
Gerakan ekor
Berdiri diam
Berdiri diam
Gerakan ekor
Berdiri diam
Buka mulut / menguap
Merawat diri
Istirahat / diam
Kedutan kulit
Gosok muka/kepala Gosok muka/kepala Garuk/gosok muka
Gosok leher
Gosok perut samping
Makan/minum
Minum
Menjilat / saltlick
Gosok badan
Makan/minum
Menjilat / saltlick
Makan/minum
Menjilat/mengendus
Istirahat/berkubang
Berkubang
Tidur
Sosial / Komunikasi
Mendengus/suara
Kontak
Agonistik
Menyerang
Berkubang
Tidur
Bersuara / mendengus
Tidur
Bersuara/dengus
Kontak
Berjaga
Urinasi / Defekasi
Urinasi Urinasi
Defekasi Defekasi
Membaui
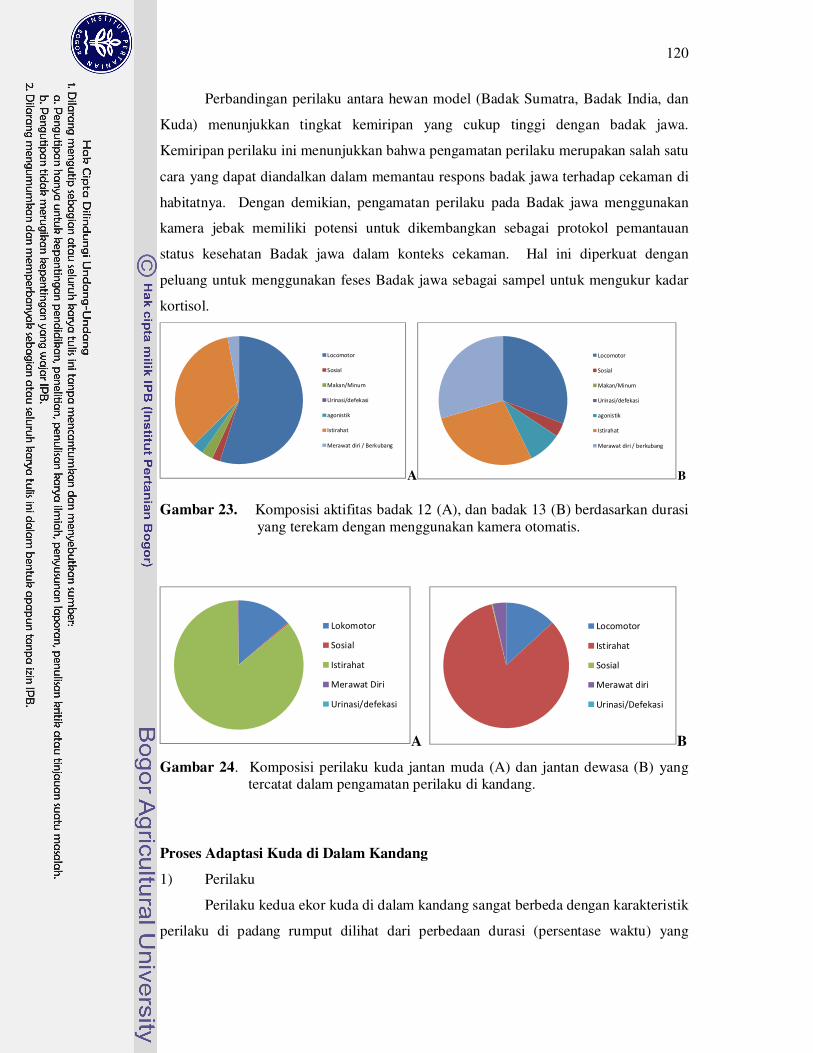
120
Perbandingan perilaku antara hewan model (Badak Sumatra, Badak India, dan
Kuda) menunjukkan tingkat kemiripan yang cukup tinggi dengan badak jawa.
Kemiripan perilaku ini menunjukkan bahwa pengamatan perilaku merupakan salah satu
cara yang dapat diandalkan dalam memantau respons badak jawa terhadap cekaman di
habitatnya. Dengan demikian, pengamatan perilaku pada Badak jawa menggunakan
kamera jebak memiliki potensi untuk dikembangkan sebagai protokol pemantauan
status kesehatan Badak jawa dalam konteks cekaman. Hal ini diperkuat dengan
peluang untuk menggunakan feses Badak jawa sebagai sampel untuk mengukur kadar
kortisol.
A B
Gambar 23. Komposisi aktifitas badak 12 (A), dan badak 13 (B) berdasarkan durasi
yang terekam dengan menggunakan kamera otomatis.
A B
Gambar 24. Komposisi perilaku kuda jantan muda (A) dan jantan dewasa (B) yang
tercatat dalam pengamatan perilaku di kandang.
Proses Adaptasi Kuda di Dalam Kandang
1) Perilaku
Perilaku kedua ekor kuda di dalam kandang sangat berbeda dengan karakteristik
perilaku di padang rumput dilihat dari perbedaan durasi (persentase waktu) yang
Locomotor
Sosial
Makan/Minum
Urinasi/defekasi
agonistik
Istirahat
Merawat diri / Berkubang
Locomotor
Sosial
Makan/Minum
Urinasi/defekasi
agonistik
Istirahat
Merawat diri / berkubang
Lokomotor
Sosial
Istirahat
Merawat Diri
Urinasi/defekasi
Locomotor
Istirahat
Sosial
Merawat diri
Urinasi/Defekasi

121
digunakan oleh hewan. Di padang rumput, sebagian besar aktifitas yang dilakukan
adalah makan atau merumput, sementara di dalam kandang kedua ekor kuda lebih
banyak menghabiskan waktu dengan berdiri diam saat tidak ada makanan di kandang
mereka. Gambar 25 menunjukkan komposisi aktifitas harian kedua ekor kuda selama
periode aklimatisasi di dalam kandang. Perbedaan aktifitas kegiatan kuda yang dilepas
dengan kuda di kandang disampaikan oleh Rivera et al (2002) yang juga menunjukkan
bahwa perubahan fisiologi terkati adaptasi dari kegiatan di lapangan ke dalam kandang
memerlukan waktu 3-4 hari.
2) Tingkat Cekaman
Kadar kortisol dalam periode aklimatisasi ini menunjukkan adanya trend
penurunan pada kedua ekor kuda sebagaimana ditampilkan pada Gambar 26. Hal ini
terjadi akibat tingginya tingkat cekaman yang diterima oleh hewan saat ditarik dari
padang rumput ke dalam kandang yang mengakibatkan meningkatnya kadar kortisol
dalam darah. Setelah menghabiskan waktu beberapa hari di dalam kandang, hewan
mulai beradaptasi dengan situasi kandang dan tidak merasakan cekaman setinggi pada
saat pertama kali dimasukkan ke dalam kandang. Linklater et al (2010) menyebutkan
bahwa penurunan kortisol juga mungkin terjadi akibat adanya mekanisme kontrol
intrinsik yang menekan aktifitas korteks adrenal yang berujung pada penurunan kadar
kortisol. Linklater et al (2010) lebih lanjut menjelaskan bahwa mekanisme kontrol
internal ini biasanya terjadi akibat penggunaan pengekang atau anestesi/sedasi terhadap
hewan tersebut keberadaan mekanisme kontrol internal seperti ini. Kadar kortisol
setelah proses aklimatisasi masih merupakan data yang relevan untuk digunakan
sebagai baseline kadar kortisol pra perlakuan mengingat mekanisme kontrol internal
tersebut bukan merupakan peubah,mengingat tidak ada pengekangan dan penggunaan
anestesi dalam penelitian ini.Sesuai dengan yang disampaikan oleh Rivera et al (2002),
kadar kortisol serta juga perilaku kuda setelah aklimatisasi ini digunakan sebagai basis
dalam analisis perbandingan kadar kortisol danperilaku dalam setiap perlakuan induksi
cekaman.
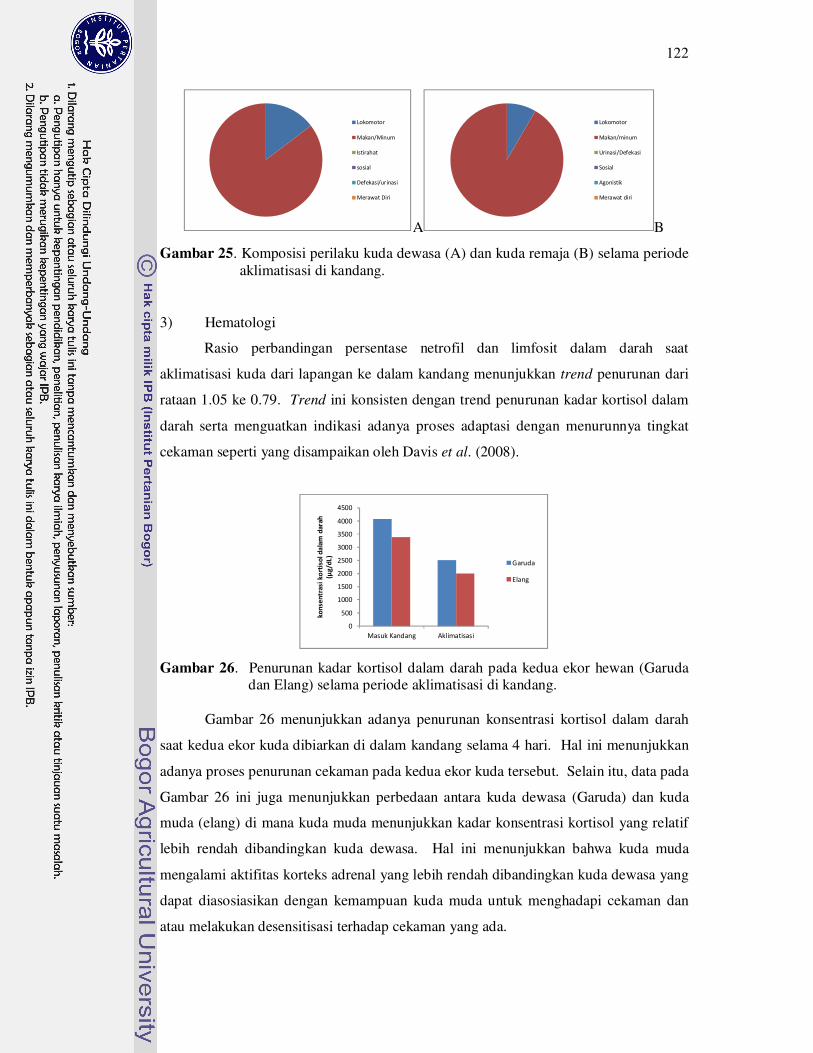
122
A B
Gambar 25. Komposisi perilaku kuda dewasa (A) dan kuda remaja (B) selama periode
aklimatisasi di kandang.
3) Hematologi
Rasio perbandingan persentase netrofil dan limfosit dalam darah saat
aklimatisasi kuda dari lapangan ke dalam kandang menunjukkan trend penurunan dari
rataan 1.05 ke 0.79. Trend ini konsisten dengan trend penurunan kadar kortisol dalam
darah serta menguatkan indikasi adanya proses adaptasi dengan menurunnya tingkat
cekaman seperti yang disampaikan oleh Davis et al. (2008).
Gambar 26. Penurunan kadar kortisol dalam darah pada kedua ekor hewan (Garuda
dan Elang) selama periode aklimatisasi di kandang.
Gambar 26 menunjukkan adanya penurunan konsentrasi kortisol dalam darah
saat kedua ekor kuda dibiarkan di dalam kandang selama 4 hari. Hal ini menunjukkan
adanya proses penurunan cekaman pada kedua ekor kuda tersebut. Selain itu, data pada
Gambar 26 ini juga menunjukkan perbedaan antara kuda dewasa (Garuda) dan kuda
muda (elang) di mana kuda muda menunjukkan kadar konsentrasi kortisol yang relatif
lebih rendah dibandingkan kuda dewasa. Hal ini menunjukkan bahwa kuda muda
mengalami aktifitas korteks adrenal yang lebih rendah dibandingkan kuda dewasa yang
dapat diasosiasikan dengan kemampuan kuda muda untuk menghadapi cekaman dan
atau melakukan desensitisasi terhadap cekaman yang ada.
Lokomotor
Makan/Minum
Istirahat
sosial
Defekasi/urinasi
Merawat Diri
Lokomotor
Makan/minum
Urinasi/Defekasi
Sosial
Agonistik
Merawat diri
0
500
1000
1500
2000
2500
3000
3500
4000
4500
Masuk Kandang Aklimatisasi
ko
nse
ntr
asi
ko
rtis
ol
da
lam
da
rah
(μg
/dL
)
Garuda
Elang
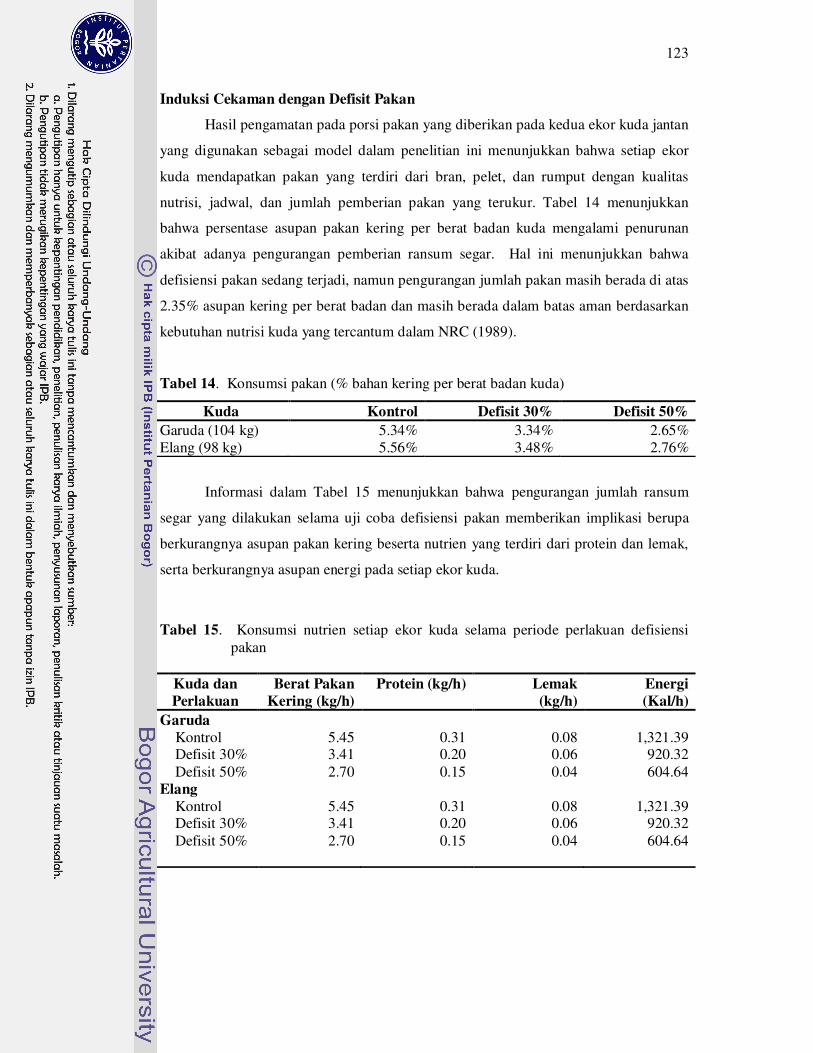
123
Induksi Cekaman dengan Defisit Pakan
Hasil pengamatan pada porsi pakan yang diberikan pada kedua ekor kuda jantan
yang digunakan sebagai model dalam penelitian ini menunjukkan bahwa setiap ekor
kuda mendapatkan pakan yang terdiri dari bran, pelet, dan rumput dengan kualitas
nutrisi, jadwal, dan jumlah pemberian pakan yang terukur. Tabel 14 menunjukkan
bahwa persentase asupan pakan kering per berat badan kuda mengalami penurunan
akibat adanya pengurangan pemberian ransum segar. Hal ini menunjukkan bahwa
defisiensi pakan sedang terjadi, namun pengurangan jumlah pakan masih berada di atas
2.35% asupan kering per berat badan dan masih berada dalam batas aman berdasarkan
kebutuhan nutrisi kuda yang tercantum dalam NRC (1989).
Tabel 14. Konsumsi pakan (% bahan kering per berat badan kuda)
Kuda Kontrol Defisit 30% Defisit 50%
Garuda (104 kg) 5.34% 3.34% 2.65%
Elang (98 kg) 5.56% 3.48% 2.76%
Informasi dalam Tabel 15 menunjukkan bahwa pengurangan jumlah ransum
segar yang dilakukan selama uji coba defisiensi pakan memberikan implikasi berupa
berkurangnya asupan pakan kering beserta nutrien yang terdiri dari protein dan lemak,
serta berkurangnya asupan energi pada setiap ekor kuda.
Tabel 15. Konsumsi nutrien setiap ekor kuda selama periode perlakuan defisiensi
pakan
Kuda dan
Perlakuan
Berat Pakan
Kering (kg/h)
Protein (kg/h) Lemak
(kg/h)
Energi
(Kal/h)
Garuda
Kontrol 5.45 0.31 0.08 1,321.39
Defisit 30% 3.41 0.20 0.06 920.32
Defisit 50% 2.70 0.15 0.04 604.64
Elang
Kontrol 5.45 0.31 0.08 1,321.39
Defisit 30% 3.41 0.20 0.06 920.32
Defisit 50% 2.70 0.15 0.04 604.64

124
Respons terhadap Perlakuan Defisit Pakan
Perlakuan defisit pakan (pengurangan ransum pakan) secara bertahap dilakukan
untuk mensimulasikan kondisi defisit energi dan nutrien sebagaimana dialami oleh
badak di ruang jelajah di habitat alaminya di Taman Nasional Ujung Kulon.
1) Perilaku
Pengamatan dilakukan untuk mengamati perubahan pada durasi dari berbagai
aktifitasyang dilakukan kuda sebagai perilaku yang umum, dan pengamatan juga
dilakukan untuk mengidentifikasi adanya penambahan perilaku yang tidak muncul pada
periode aklimatisasi. Pada perlakuan defisit pakan nampak bahwa durasi aktifitas
berjalan menunjukkan trend penurunan dibandingkan dengan kontrol dan juga
perlakuan defisit air sementara aktifitas yang muncul pada perlakuan defisit pakan
namun tidak muncul pada periode kontrol adalah: mengunyah (dengan mulut kosong)
dan menggigit ember. Tambahan kebiasaan dengan gerakan mulut dan mengigit ember
merupakan respons yang kerap dijumpai pada kuda yang tengah mengalami cekaman
dan perilaku ini terkait dengan adanya peningkatan detak jantung (Bachman et al 2003;
Nagy et al. 2009). Perbandingan komposisi aktifitas perilaku kuda dalam penelitian ini
disajikan pada Gambar 28.
2) Tingkat Cekaman
Tingkat cekaman diamati dari kadar kortisol dalam darah dan Gambar 27
menunjukkan bahwa kadar kortisol selama perlakuan defisit pakan menunjukkan
fluktuasi/lonjakan yang cukup tinggi pada saat perlakuan baru berjalan 3 hari, namun
kadar kortisol ini menurun pada hari-hari berikutnya. Hal ini menunjukkan bahwa kuda
mengalami cekaman yang cukup tinggi saat ransum pakan mulai dikurangi menjadi
70% dari ransum yang biasa diterima. Menurunnya kadar kortisol pada hari-hari
berikutnya dalam perlakuan defisit pakan ini menunjukkan bahwa kuda dapat
beradaptasi dengan kondisi berkurangnya pakan (walaupun jumlah pakan yang diterima
berkurang lagi menjadi hanya 50% dari ransum yang biasa diterima).
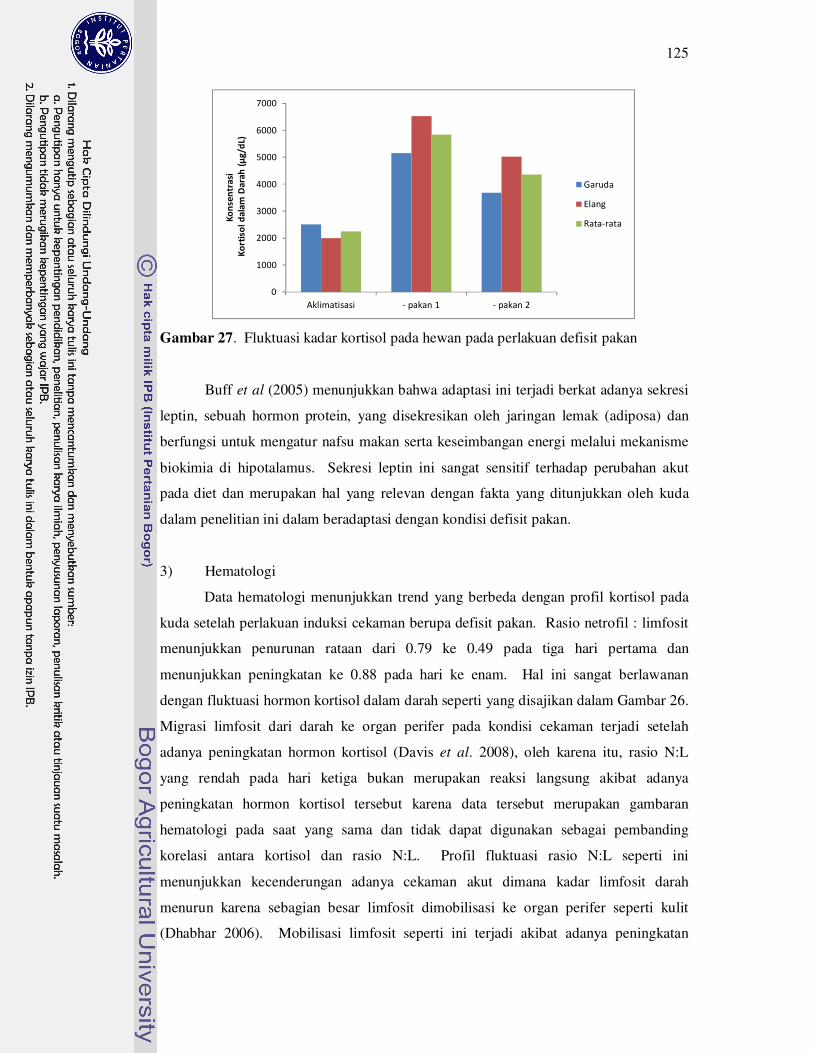
125
Gambar 27. Fluktuasi kadar kortisol pada hewan pada perlakuan defisit pakan
Buff et al (2005) menunjukkan bahwa adaptasi ini terjadi berkat adanya sekresi
leptin, sebuah hormon protein, yang disekresikan oleh jaringan lemak (adiposa) dan
berfungsi untuk mengatur nafsu makan serta keseimbangan energi melalui mekanisme
biokimia di hipotalamus. Sekresi leptin ini sangat sensitif terhadap perubahan akut
pada diet dan merupakan hal yang relevan dengan fakta yang ditunjukkan oleh kuda
dalam penelitian ini dalam beradaptasi dengan kondisi defisit pakan.
3) Hematologi
Data hematologi menunjukkan trend yang berbeda dengan profil kortisol pada
kuda setelah perlakuan induksi cekaman berupa defisit pakan. Rasio netrofil : limfosit
menunjukkan penurunan rataan dari 0.79 ke 0.49 pada tiga hari pertama dan
menunjukkan peningkatan ke 0.88 pada hari ke enam. Hal ini sangat berlawanan
dengan fluktuasi hormon kortisol dalam darah seperti yang disajikan dalam Gambar 26.
Migrasi limfosit dari darah ke organ perifer pada kondisi cekaman terjadi setelah
adanya peningkatan hormon kortisol (Davis et al. 2008), oleh karena itu, rasio N:L
yang rendah pada hari ketiga bukan merupakan reaksi langsung akibat adanya
peningkatan hormon kortisol tersebut karena data tersebut merupakan gambaran
hematologi pada saat yang sama dan tidak dapat digunakan sebagai pembanding
korelasi antara kortisol dan rasio N:L. Profil fluktuasi rasio N:L seperti ini
menunjukkan kecenderungan adanya cekaman akut dimana kadar limfosit darah
menurun karena sebagian besar limfosit dimobilisasi ke organ perifer seperti kulit
(Dhabhar 2006). Mobilisasi limfosit seperti ini terjadi akibat adanya peningkatan
0
1000
2000
3000
4000
5000
6000
7000
Aklimatisasi - pakan 1 - pakan 2
Ko
nse
ntr
asi
Ko
rtis
ol
da
lam
Da
rah
(μ
g/d
L)
Garuda
Elang
Rata-rata

126
ekspresi molekul adesif yang mendorong perlekatan sel-sel monosit, granulosit, dan
limfosit ke organ-organ terentu mulai dari dinding pembuluh darah dan organ-organ
perifer (Goebel & Mills 2000). Dengan fakta seperti ini dapat disimpulkan bahwa
defisit pakan memberikan cekaman yang bersifat akut.
Kekurangan pakan merupakan satu jenis cekaman yang dapat dihadapi dengan
adaptasi sehingga defisit pakan tersebut tidak lagi menjadi cekaman yang tinggi atau
kronis. Sebagai mekanisme untuk mengurangi kebutuhan energi, terlihat bahwa hewan
mengurangi aktifitas berjalan dalam kondisi defisit pakan ini. Hal ini sangat relevan
dengan data dari Badak jawa individu no 12 di Taman Nasional Ujung Kulon yang
menunjukkan tingkat asupan pakan paling rendah dan menunjukkan ruang dan jarak
jelajah yang paling kecil dibandingkan dua ekor Badak lainnya sebagaimana
dipaparkan dalam penelitian mengenai analisi nutrisi dan kecernaan pada Badak jawa.
Respons hormonal terhadap variasi dan perbedaan diet juga telah ditunjukkan oleh
Glade et al (1984) dimana flukutasi hormonal yang tidak seimbang dapat menimbulkan
risiko kesehatan pada kuda, terutama kuda yang masih muda.
Respons terhadap Perlakuan Defisit Air
Perlakuan defisit air merupakan simulasi dari kondisi yang dihadapi oleh badak
jawa di Taman Nasional Ujung Kulon di musim kering di mana banyak sumber air
(anak sungai) yang mengering dan ketersediaan air bagi Badak jawa akan jauh
berkurang. Selain itu prediksi skenario perubahan iklim yang dilakukan oleh Permadi
(2008) menunjukkan adanya kemungkinan terjadinya kekeringan yang berkepanjangan
sepanjang tahun.
1) Perilaku
Pengamatan dilakukan untuk mengamati perubahan pada durasi dari berbagai
aktifitas yang dilakukan kuda sebagai perilaku yang umum, dan pengamatan juga
dilakukan untuk mengidentifikasi adanya penambahan perilaku yang tidak muncul pada
periode aklimatisasi. Pada perlakuan defisit pakan nampak bahwa durasi aktifitas
berjalan menunjukkan trend penurunan dibandingkan dengan kontrol dan juga
perlakuan defisit air sementara aktifitas yang muncul pada perlakuan defisit pakan
namun tidak muncul pada periode kontrol adalah: mengunyah (dengan mulut kosong)
dan aktifitas menggigit ember yang banyak dilakukan oleh kuda jantan muda.
Kebiasaan menggigit seperti ini dijelasakan oleh Bachman et al. (2003) dan Nagy et al.

127
(2009) sebagai mekanisme yang dilakukan oleh kuda dalam kondisi tercekam, dan
perilaku seperti ini memiliki korelasi positif dengan peningkatan detak jantung dan
kadar kortisol dalam plasma darah. Perbandingan komposisi aktifitas perilaku kuda
dalam penelitian ini disajikan pada Gambar 28.
2) Tingkat Cekaman
Tingkat cekaman diamati dari kadar kortisol dalam darah serta tingkat respirasi
pada kuda dibandingkan dengan tingkat respirasi normal atau istirahat. Respirasi
normal/istirahat berdasarkan Evans et al. (1990) adalah 8-16 respirasi per menit.
Fluktuasi kadar kortisol dalam darah menunjukkan kecenderungan untuk meningkat
secara eksponensial (R2=0.948) selama periode perlakuan seperti yang ditampilkan
dalam Gambar 29. Demikian juga halnya dengan tingkat respirasi yang meningkat
hampir secara linear (R2 = 0.912) selama perlakuan defisit air sebagaimana ditampilkan
pada Gambar 28. Peningkatan kadar kortisol nampak berjalan seiring dengan
meningkatnya respirasi pada kuda, namun demikian belum dapat disimpulkan apakah
kedua parameter ini (kortisol dan respirasi) saling berkorelasi ataupun bergantung satu
dengan yang lainnya mengingat pada perlakuan defisit pakan respirasi terus meningkat
walaupun kadar kortisol mulai menurun akibat adanya proses adaptasi.
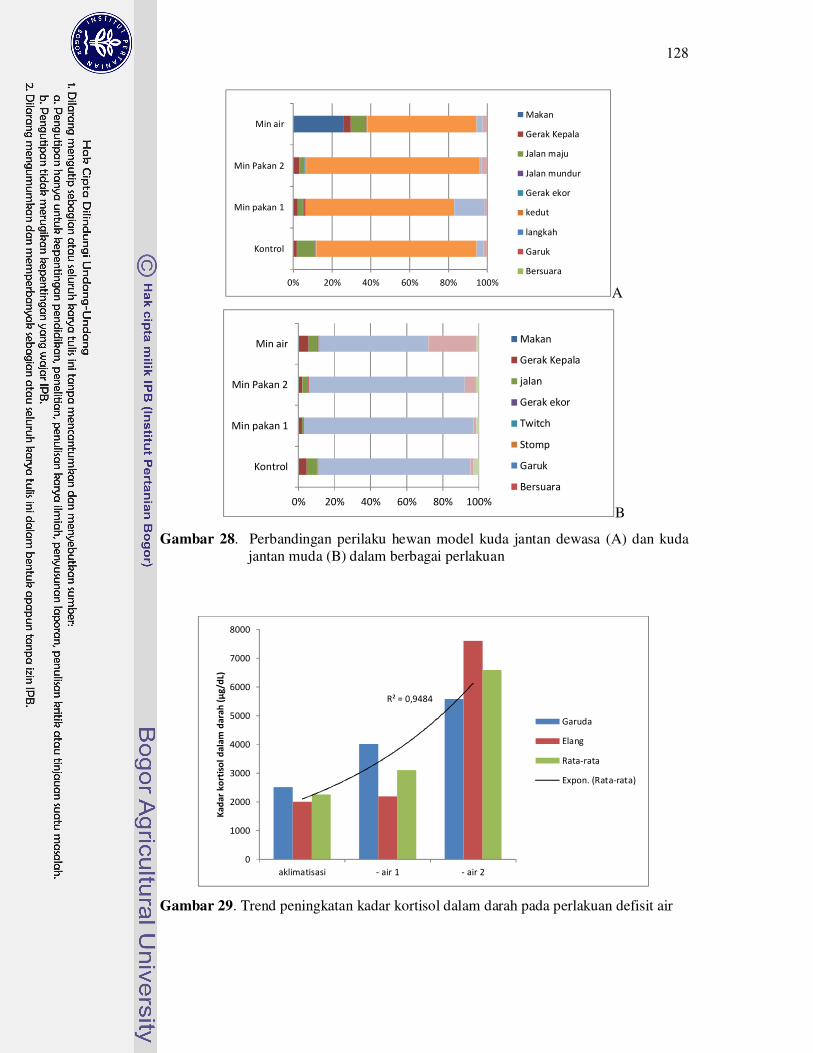
128
A
B
Gambar 28. Perbandingan perilaku hewan model kuda jantan dewasa (A) dan kuda
jantan muda (B) dalam berbagai perlakuan
Gambar 29. Trend peningkatan kadar kortisol dalam darah pada perlakuan defisit air
0% 20% 40% 60% 80% 100%
Kontrol
Min pakan 1
Min Pakan 2
Min airMakan
Gerak Kepala
Jalan maju
Jalan mundur
Gerak ekor
kedut
langkah
Garuk
Bersuara
0% 20% 40% 60% 80% 100%
Kontrol
Min pakan 1
Min Pakan 2
Min air Makan
Gerak Kepala
jalan
Gerak ekor
Twitch
Stomp
Garuk
Bersuara
R² = 0,9484
0
1000
2000
3000
4000
5000
6000
7000
8000
aklimatisasi - air 1 - air 2
Ka
da
r k
ort
iso
l d
ala
m d
ara
h (
μg
/dL)
Garuda
Elang
Rata-rata
Expon. (Rata-rata)

129
3) Hematologi
Data hematologi menunjukkan trend yang berbeda dengan profil kortisol pada
kuda setelah perlakuan induksi cekaman berupa defisit pakan. Rasio netrofil : limfosit
tidak menunjukkan perubahan yang nyata dari rataan dari 0.79 ke 0.81 pada tiga hari
pertama dan menunjukkan penurunan ke rataan 0.63 pada hari ke enam. Hal ini tidak
seiring dengan fluktuasi hormon kortisol dalam darah seperti yang disajikan dalam
Gambar 29. Rasio N:L yang relatif tidak berubah pada hari ketiga bukan merupakan
reaksi langsung akibat adanya peningkatan hormon kortisol tersebut karena data
tersebut merupakan gambaran hematologi pada saat yang sama dan tidak dapat
digunakan sebagai pembanding korelasi antara kortisol dan rasio N:L.
Kecenderungan penurunan rasio N:L pada hari keenam menunjukkan persentase
limfosit yang lebih besar di dalam darah dibandingkan dengan kondisi pada hari ketiga.
Hal ini dapat langsung dikaitkan dengan tidak adanya mobilisasi limfosit dari darah ke
organ perifer dan menunjukkan adanya kecenderungan imunosupresi yang lazim terjadi
pada cekaman yang bersifat kronis (Dhabhar 2006). Dengan kecenderungan seperti ini
dapat disimpulkan bahwa defisit air merupakan cekaman yang bersifat kronis.
Ketersediaan air merupakan faktor yang sangat penting dan perlu diperhatikan
mengingat cekaman akibat menurunnya ketersediaan air merupakan cekaman yang
bersifat kronis dan sulit untuk diatasi sebagaimana dicerminkan dari kecenderungan
peningkatan kadar kortisol dan penurunan rasio N:L. Selain melakukan perilaku
tambahan seperti aktifitas menggigit ember dan menjilati pagar, kuda tidak melakukan
pengurangan aktifitas berjalan atau perubahan aktifitas lainnya untuk beradaptasi
dengan kondisi keterbatasan air. Coenen (2005) juga menunjukkan adanya peningkatan
kortisol dalam darah kuda akibat berkurangnya asupan air sebagaimana disimulasikan
dalam penelitian ini. Cekaman kronis seperti yang terjadi akibat defisit air dapat
menimbulkan implikasi kesehatan yang serius karena mengakibatkan imunosupresi
(Domingues-Gerpe & Rey-Mendez 2001) yang menurunkan daya tahan tubuh hewan
terhadap serangan agen infeksius.

130
Pola Respirasi Dalam Kondisi Cekaman
Pengamatan respirasi serta perbandingan tingkat respirasi pada kuda
dibandingkan dengan tingkat respirasi pada kondisi normal atau istirahat. Respirasi
normal/istirahat yang diambil berdasarkan informasi dalam Evans et al. (1990) adalah
8-16 respirasi per menit. Analisis pada tingkat respirasi saat kondisi defisit pakan dan
defisit airmenunjukkan bahwa kedua ekor kuda menunjukkan tingkat respirasi yang
tinggi dibandingkan dengan respirasi dalam kondisi normal atau istirahat seperti yang
ditampilkan pada Gambar 30. Peningkatan tingkat respirasi terjadi dari 8-16 respirasi
per menit melonjak ke 35-47 respirasi per menit yang merupakan kenaikan sebesar
1.93-3.37 kali lipat dari pernapasan dalam kondisi normal. Pada badak, peningkatan
respirasi akibat cekaman dari adanya interaksi agonistik terjadi dari 16-23 respirasi per
menit (Citino & Bush 2007) menjadi 42.60respirasi per menit (Hariyadi et al. 2010)
atau kenaikan sebesar 0.85-1.66 kali lipat. Hal ini menunjukkan bahwa kuda
mengalami peningkatan respirasi yang relatif sebanding dengan badak jawa seperti
yang ditampilkan pada Gambar 31. Walaupun peningkatan tingkat respirasi tidak
menunjukkan adanya korelasi dengan fluktuasi pada kadar kortisol dalam darah, baik
kuda maupun badak menunjukkan kecenderungan peningkatan respirasi dalam kondisi
cekaman. Fakta ini menunjukkan bahwa peningkatan respirasi seperti ini merupakan
indikator yang cukup sensitif untuk memantau keadaan cekaman secara kualitatif pada
kuda ataupun pada badak jawa.
Tingkat respirasi hewan pada saat menjalani kedua jenis cekaman menunjukkan
kecenderungan peningkatan. Oleh karena itu, pengamatan pada tingkat respirasi dapat
digunakan sebagai data penunjang dalam menganalisis tingkat cekaman yang dihadapi.
Namun demikian tingkat respirasi ini bukan merupakan indikator yang spesifik untuk
cekaman pada badak, karena selain akibat adanya cekaman peningkatan respirasi dapat
pula disebabkan oleh meningkatnya aktifitas hewan (Evans et al. 1990).
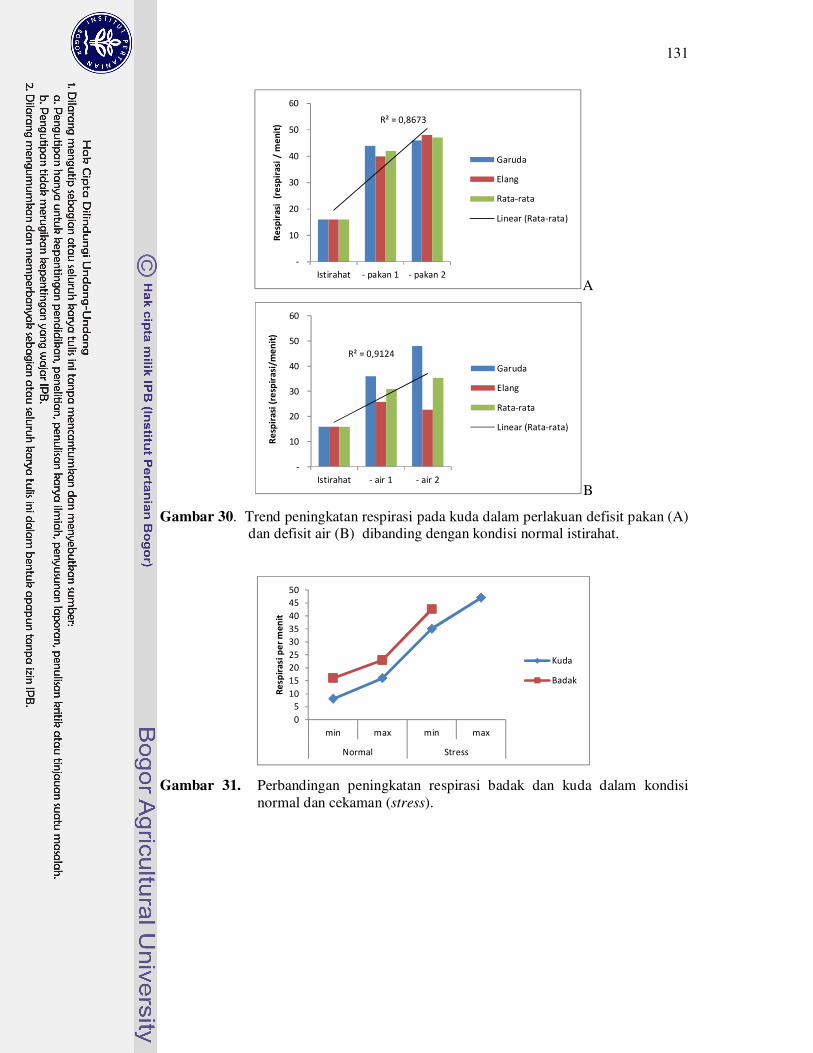
131
A
B
Gambar 30. Trend peningkatan respirasi pada kuda dalam perlakuan defisit pakan (A)
dan defisit air (B) dibanding dengan kondisi normal istirahat.
Gambar 31. Perbandingan peningkatan respirasi badak dan kuda dalam kondisi
normal dan cekaman (stress).
R² = 0,8673
-
10
20
30
40
50
60
Istirahat - pakan 1 - pakan 2R
esp
ira
si
(re
spir
asi
/ m
en
it)
Garuda
Elang
Rata-rata
Linear (Rata-rata)
R² = 0,9124
-
10
20
30
40
50
60
Istirahat - air 1 - air 2
Re
spir
asi
(re
spir
asi
/me
nit
)
Garuda
Elang
Rata-rata
Linear (Rata-rata)
0
5
10
15
20
25
30
35
40
45
50
min max min max
Normal Stress
Re
spir
asi
pe
r m
en
it
Kuda
Badak

132
Identifikasi Komponen Pemicu Cekaman
Dari penelitian ini dapat dilihat bahwa terjadinya cekaman dapat diasosiasikan
dengan meningkatnya kadar kortisol dalam darah dan juga meningkatnya respirasi.
Regresi linear berganda dilakukan untuk mengindentifikasi kontribusi dan korelasi dari
berbagai faktor seperti: asupan protein, asupan air, asupan energi, rasio N/L terhadap
Kadar kortisol dan respirasi. Regresi linear berganda menunjukkan hasil sebagai
berikut:
• Respirasi = 99.3785 – 538.6646 x protein + 0.0803 x Energi – 2.0521 x Air
(L) + 11.2048 x n/l + 0.0006 x kortisol. (R2 =0.8358)
• Kortisol = 11,409.0257 – 60,927.1702 x protein + 11.5232 x Energi – 240.3852
x Air (L) – 2,648.2992 x n/l + 27.8213 x Respirasi. (R2 = 0.6284)
Hasil ini menunjukkan kecenderungan bahwa faktor yang paling menentukan
dalam peningkatan respirasi adalah asupan protein (koefisien = 538.67), diikuti dengan
rasio n/L (koefisien 11.20), dan air (koefisien = 2.05). Demikian pula dengan
peningkatan kadar kortisol dalam darah yang ditentukan oleh asupan protein (koefisien
= 60,927.17), diikuti dengan rasio n/L (koefisien= 2,648.30), dan air (koefisien =
240.3852).
Kecernaan Kuda Dalam Kondisi Cekaman
Pada kondisi kualitas pemeliharaan yang sama, komposisi pakan yang terukur,
dan periode pemberian pakan yang teratur selama perlakuan pada kedua ekor kuda,
maka dapat dipastikan bahwa konsumsi pakan bagi kedua ekor kuda tersebut
merupakan faktor konstan dalam uji coba ini. Oleh karena itu, perubahan fisiologi
pencernaan yang terjadi pada kedua ekor kuda tersebut merupakan pengaruh dari
cekaman yang timbul. AIA pada pakan dan feses kuda serta perhitungan kecernaan
selama perlakuan ditampilkan pada Tabel 16.
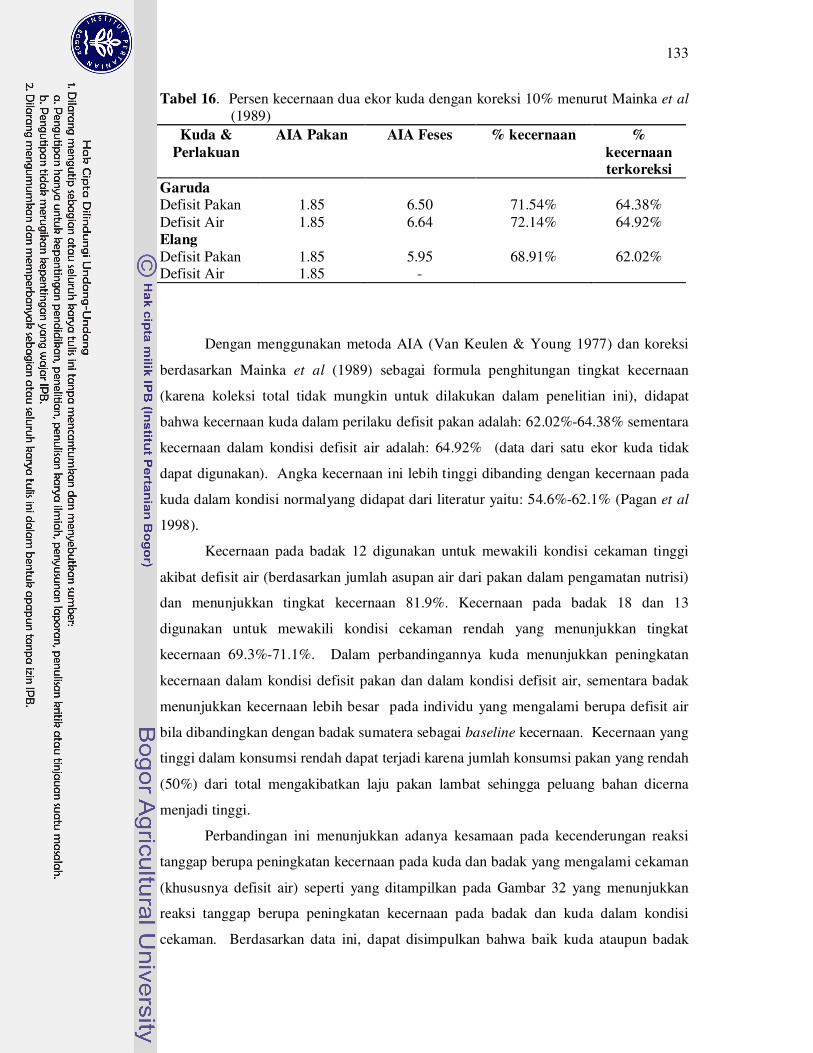
133
Tabel 16. Persen kecernaan dua ekor kuda dengan koreksi 10% menurut Mainka et al
(1989)
Kuda &
Perlakuan
AIA Pakan AIA Feses % kecernaan %
kecernaan
terkoreksi
Garuda
Defisit Pakan 1.85 6.50 71.54% 64.38%
Defisit Air 1.85 6.64 72.14% 64.92%
Elang
Defisit Pakan 1.85 5.95 68.91% 62.02%
Defisit Air 1.85 -
Dengan menggunakan metoda AIA (Van Keulen & Young 1977) dan koreksi
berdasarkan Mainka et al (1989) sebagai formula penghitungan tingkat kecernaan
(karena koleksi total tidak mungkin untuk dilakukan dalam penelitian ini), didapat
bahwa kecernaan kuda dalam perilaku defisit pakan adalah: 62.02%-64.38% sementara
kecernaan dalam kondisi defisit air adalah: 64.92% (data dari satu ekor kuda tidak
dapat digunakan). Angka kecernaan ini lebih tinggi dibanding dengan kecernaan pada
kuda dalam kondisi normalyang didapat dari literatur yaitu: 54.6%-62.1% (Pagan et al
1998).
Kecernaan pada badak 12 digunakan untuk mewakili kondisi cekaman tinggi
akibat defisit air (berdasarkan jumlah asupan air dari pakan dalam pengamatan nutrisi)
dan menunjukkan tingkat kecernaan 81.9%. Kecernaan pada badak 18 dan 13
digunakan untuk mewakili kondisi cekaman rendah yang menunjukkan tingkat
kecernaan 69.3%-71.1%. Dalam perbandingannya kuda menunjukkan peningkatan
kecernaan dalam kondisi defisit pakan dan dalam kondisi defisit air, sementara badak
menunjukkan kecernaan lebih besar pada individu yang mengalami berupa defisit air
bila dibandingkan dengan badak sumatera sebagai baseline kecernaan. Kecernaan yang
tinggi dalam konsumsi rendah dapat terjadi karena jumlah konsumsi pakan yang rendah
(50%) dari total mengakibatkan laju pakan lambat sehingga peluang bahan dicerna
menjadi tinggi.
Perbandingan ini menunjukkan adanya kesamaan pada kecenderungan reaksi
tanggap berupa peningkatan kecernaan pada kuda dan badak yang mengalami cekaman
(khususnya defisit air) seperti yang ditampilkan pada Gambar 32 yang menunjukkan
reaksi tanggap berupa peningkatan kecernaan pada badak dan kuda dalam kondisi
cekaman. Berdasarkan data ini, dapat disimpulkan bahwa baik kuda ataupun badak
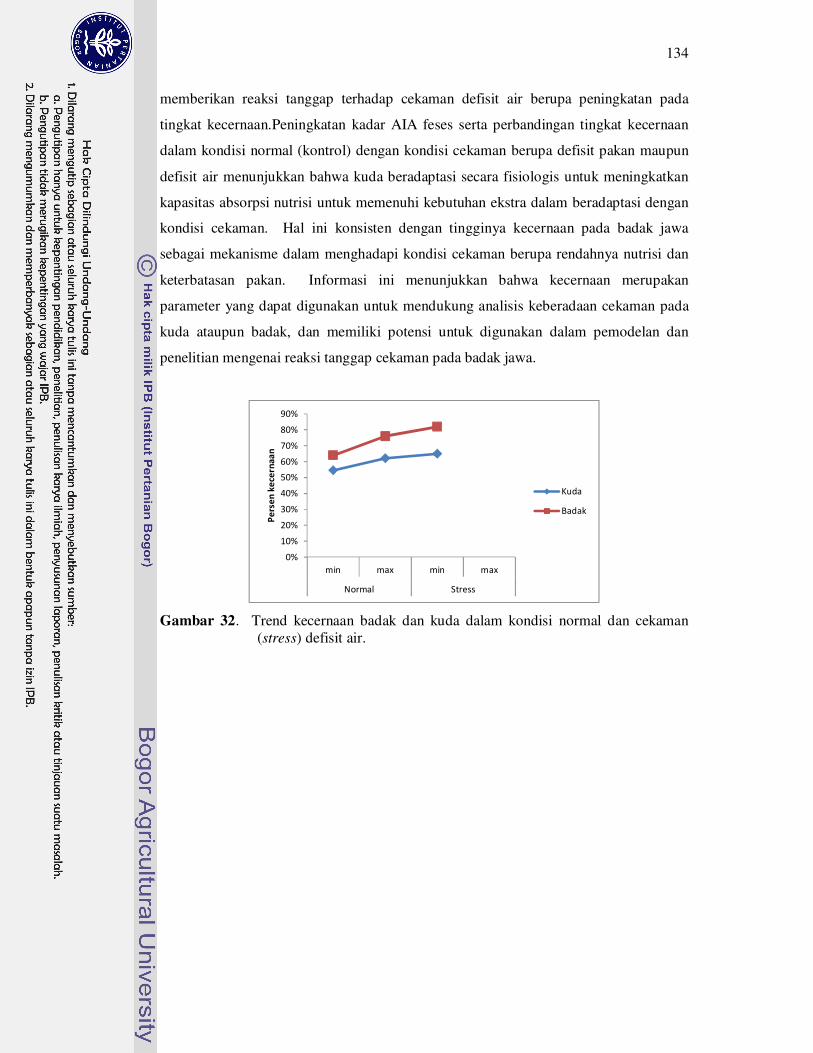
134
memberikan reaksi tanggap terhadap cekaman defisit air berupa peningkatan pada
tingkat kecernaan.Peningkatan kadar AIA feses serta perbandingan tingkat kecernaan
dalam kondisi normal (kontrol) dengan kondisi cekaman berupa defisit pakan maupun
defisit air menunjukkan bahwa kuda beradaptasi secara fisiologis untuk meningkatkan
kapasitas absorpsi nutrisi untuk memenuhi kebutuhan ekstra dalam beradaptasi dengan
kondisi cekaman. Hal ini konsisten dengan tingginya kecernaan pada badak jawa
sebagai mekanisme dalam menghadapi kondisi cekaman berupa rendahnya nutrisi dan
keterbatasan pakan. Informasi ini menunjukkan bahwa kecernaan merupakan
parameter yang dapat digunakan untuk mendukung analisis keberadaan cekaman pada
kuda ataupun badak, dan memiliki potensi untuk digunakan dalam pemodelan dan
penelitian mengenai reaksi tanggap cekaman pada badak jawa.
Gambar 32. Trend kecernaan badak dan kuda dalam kondisi normal dan cekaman
(stress) defisit air.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
min max min max
Normal Stress
Pe
rse
n k
ece
rna
an
Kuda
Badak

135
Adaptasi Fisiologis Terhadap Cekaman
Peningkatan respirasi dan tingkat kecernaan merupakan fenomena yang lazim
ditemukan pada kuda dalam kondisi cekaman (Pagan et al 1998; Coenen 2005). Kedua
perubahan ini merupakan respons fisiologis tubuh untuk menghadapi cekaman yang
datang. Peningkatan respirasi menunjang peningkatan produksi energi dari molekul
ATP yang diurai dalam siklus Krebs yang banyak didorong oleh proses enzimatik dan
oksidatif (Koolman & Rohm 2001). Peningkatan kecernaan juga merupakan salah satu
mekanisme untuk meningkatkan penyerapan nutrien dari saluran pencernaan ke
pembuluh darah dan kemudian didistribusikan ke sel untuk digunakan sebagai sumber
energi yang siap pakai seperti glukosa, ataupun bahan baku lainnya untuk siklus Krebs
seperti protein ataupun lemak. Peningkatan metabolisme seperti ini merupakan langkah
untuk menggalang energi yang digunakan dalam respons fight or flight yang dipicu
oleh hormon cekaman yang disekresikan dari korteks adrenal.

136
Simpulan dan Saran
1. Perlakuan defisit pakan dan defisit air merupakan cara dapat dilakukan untuk
menimbulkan cekaman pada hewan model (kuda).
2. Cekaman akibat defisit pakan mengakibatkan perubahan perilaku berupa
pengurangan aktifitas berjalan dan adanya aktifitas menggigit embar atau pagar
(crib biting).
3. Defisit air merupakan sumber cekaman dengan karakteristik kronis yang ditandai
dengannaiknya kadar kortisol dan turunnya rasio N/L akibat migrasi limfosit dari
organ perifer kembali ke darah (imunosupresi).
4. Defisit pakan merupakan sumber cekaman dengan karakteristik akut yang
ditandai dengan kenaikan sementara kadar kortisol dalam darah yang diikuti
dengan peningkatan rasio N/L akibat migrasi limfosit dari darah ke organ perifer
(imunomodulasi).
5. Dampak lain dari adanya cekaman adalah adanya peningkatan respirasi dan
kecernaan pada kuda yang mengalami cekaman, dan hal ini merupakan adaptasi
fisiologis yang dilakukan untuk menjamin pemenuhan kebutuhan energi dalam
kondisi cekaman tersebut.
6. Kuda menunjukkan trend peningkatan kecernaan dan respirasi yang sebanding
dengan badak, dan ini menunjukkan bahwa kuda merupakan model yang layak
untuk digunakan dalam studi terkait cekaman pada badak.
Saran
1. Melakukan penelitian mengenai korelasi antara tingkat cekaman kronis ataupun
akut dengan kerentanan terhadap penyakit yang dapat mengancam populasi
badak jawa dengan menggunakan hewan model.
2. Menerapakan konsep pemantauan perilaku dan profil hormon cekaman untuk
badak jawa di habitat alaminya dengan referensi reaksi tanggap pada hewan
model.
3. Menerapkan upaya mitigasi cekaman kronis akibat defisit air dengan
meningkatkan akses jalur badak ke titik-titik air untuk minum maupun
berkubang

137
Daftar Pustaka
Agil M, Purwantara B, Alikodra HS, Hodges K, Toelihere MS. 2007. Reproductive
biology of the sumatran rhinoceros Dicerorhinus sumatrensis (Fischer 1814).
Disertasi program Doktor Institut Pertanian Bogor. Sekolah Pasca Sarjana IPB
Agil M, Setiadi DR, Supriatna I, Purwantara B,. 2008. Non-Invasive Endocrine
Monitoring of Reproduction and Stres in the wild animal: Analyzing hormone
metabolites in Urine and Faeces using Enzymeimmunoassay. Proceeding of
AZWMC 2008
Bachman I, et al. 2003. Behavioural and physiological responses to an acute stressor in
crib-biting and control horses. Applied animal behaviour science 82: 297-311
Buff PR, Morrison CD, Ganjam VK, Keisler DH,. 2005. Effects of short-term feed
deprivation and melatonin implants on circadian patterns of leptin in the horse.
Journal of animal science 83(5): 1023-1032
Cameron HL, Perdue MH. 2005. Stres Impairs Murine Intestinal Barrier Function:
Improvement by Glucagon-Like Peptide-2. The journal of pharmacology and
experimental therapeutics Vol. 314, No. 1
Carlstead K, Brown JL,. 2005. Relationships between patterns of fecal corticoid
excretion and behaviour, reproduction, and environmental factors in captive
black (Diceros bicornis) and white (Ceratohterium simum) Rhinoceros. Zoo
biology 24: 215-232
Citino SB, Bush M. 2007. Reference Cardiopulmonary Physiologic Parameters for
Standing, Unrestrained White Rhinoceroses (Ceratotheriumsimum). Journal of
Zoo and Wildlife Medicine 38(3):375–379
Clauss M, et al. 2005. Studies on digestive physiology and feed digestibilities in
captive Indian rhinoceros (Rhinoceros unicornis). Journal of Animal
Physiology and Animal Nutrition 89 (2005) 229–237
Clauss M, et al. 2006. Digestion coefficients achieved by the black rhinoceros (Diceros
bicornis) a large, browsing, hidgut fermenter. Journal of animal physiology and
animal nutrition. 1-10
Coenen M,. 2005. Exercise and stress: impact on adaptive processes involving water
and electrolytes. Livestock production science 92:131-145
Colgan, PW. 1978. Quantitative ethology. Wiley-Interscience publications. John Wiley
and Sons, New York
Davis AK, Maney DL, Maersz JC,. 2008. The use of leucocyte profiles to measure
stress in vertebrates: a review for ecologists. Functional ecology 22(5): 760-772

138
Dhabhar FS,. 2000. Acute stress enhances while chronic stress supresses skin
immunity: the roles of stress hormones and leukocyte trafficking. Annals of the
New York academy of Sciences vol 917: Neuroimmunomodulation perspectives
at the new millenium:876-893
Domingues-Gerpe L, Rey-Mendez M,. 2001. Alterations induced by chronic stress in
lymphocyte subsets of blood and primary and secondary immune organs of
mice. BMC Immunology 2:7. http://www.biomedcentral.com/1471-2172/2. [9
Juni 2012]
Dyer J, et al. 2002. Molecular characterisation of carbohydrate digestion and absorption
in equine small intestines. Equine veterinary journal 34(4):349-358
Evans W, Borton A, Hints HF, Vleck LDV,. 1990. The Horse Second Edition. W.H.
Freeman and Company, New York
Fernando P, Polet G, Foead N. Ng L. Melnick DJM. 2004. Mitochondrial DNA
analysis of the critically endangered Javan Rhinoceros. PHKA-WWF-Columbia
University
Figueiredo HF, et al. 2003. Stress integration after acute and chronic predator stress:
Differential activation of central stress circuitry and sensitization of th
ehypothalamo-pituitary-adrenocortical axis. Endocrinology 144(12): 5249-5258
Firdaus Y,. 2008. Laporan Climate Modelling. Kajian Dampak Perubahan Iklim
Terhadap Kerentanan Badak jawa. WWF Indonesia – Program Iklim dan
Energi
Glade MJ, Gupta S, Reimers TJ,. 1984. Hormonal Responses to High and Low Planes
of Nutrition in Weanling Thoroughbreds. J Anim Sci 1984. 59:658-665
Goebel MU, Mills PJ,. 2000. Acute psychological stress and exercise and changes in
peripheral leukocytes adhesion molecule expression and density. Psychosomatic
medicine 62: 664-670.
Hariyadi ARS, Setiawan R, Daryan, Yayus A, Purnama H,. 2010. Preliminary
behaviour observations of the Javan rhinoceros (Rhinoceros sondaicus) based
on video trap surveys in Ujung Kulon National Park. Pachyderm 47: 93-99
Harley EH, Matshikiza M, Robson P, Weber B,. 2004. Red blood cell metabolism
shows major anomalies in Rhinocerotidae and Equidae, suggesting a novel role
in general antioxidant metabolism. Animals and Environments. Proceedings of
the Third International Conference of Comparative Physiology and
Biochemistry. Pp 334-340
Hutchin M, Kreger MD,. 2006. Rhinoceros behaviour: implications for captive
management and conservation. Int. Zoo. Yb 40:150-173
Koolman J, Rohm KH, Wanandi SI[ed], Sadikin M [ed]. 2001. Atlas berwarna dan teks
Biokimia. Cetakan I. Penerbit Hipokrates, Jakarta

139
Linklater WL, MacDonald EA, Flamand JRB, Czekala NM,. 2010. Declining and low
fecal corticoids are associated with distress, not acclimatization to stress, during
the translocation of African rhinoceros. Animal conservation 13: 104-111.
Mainka SA, Zhao GL, Li M, 1989. Utilization of a bamboo, sugar cane, and gruel diet
by two juvenile giant pandas (Ailuropoda melanoleuca). Journal of zoo and
wildlife medicine 20:39-44
Sims JA, et al. 2007. Determination of bamboo-diet digestibility and fecal output by
giant pandas. Ursus 18(1): 38-45
McSweeney CS, Pass MA,. 1983. The mechanism of ruminal stasis in Lantana-
poisoned sheep. Quarterly journal of experimental physiology 68: 301-313
NRC. 1989. Nutrient requirements of horses, 5th ed. National Academy Press,
Washington DC.
Nagy K, et al. 2009. The effect of a feeding stress-test on the behaviour and the heart
rate variability of control and crib-biting horses (with or without inhibition).
Applied animal behaviour science 121: 140-147
Nessminth S, Baltzell J, Bardanier CD,. 1983. Interaction of Glucocorticoid and
thyroxine in the responses of rats starvation-refeeding J. Nutr. 113: 2260-2265
Noble GK, et al. 2007. Effect of excercise training, circadian rythm, age, and sex, on
insulin-like growth factor-1 in the horse. Journal of animal science 85: 163-171
Pagan JD, et al. 1998. Exercise affects digestibility and rate of passage of all-forage and
mixed diets in throroughbred horses. American society for nutritional sciences.
J. Nutr. 128: 2704S-2707S
Pass MA, Seawright AA, Lamberton JA, Heath TJ,. 1979. Lantadene A toxicity in
sheep. A model for cholestasis. Pathology. 1979 Jan;11(1):89-94
Permadi YF,. 2008. Kajian dampak perubahan iklim terhadap kerentanan badak jawa.
Laporan proyek WWF Indonesia.
Rivera E, Benjamin B, Nielsen J, Shelle J, Zanella AJ,. 2002. Behavioural and
physiological responses of horses to initial training: the comparison between
pastured versus stalled horses. Applied animal behavioural science 78(2-4): 235-
252
Sajudin HR, Djaja B,. 1984. Monitoring populasi badak jawa (Rhinoceros sondaicus
Desmarest 1822) di Semenanjung Ujung Kulon. Laporan akhir penelitian
IUCN/WWF proyok nomor 1960. Fakultas Biologi UNAS
VanKeulen J, Young, BA. 1977. Evaluation of Acid Insoluble Ash as a Natural Marker
in Ruminant digestibility Studies. Journal of Animal Science vol 44 no: 2. 282-
287

PEMBAHASAN UMUM:
MODEL DALAM PENGELOLAAN POPULASI BADAK JAWA
Pendahuluan
Badak Jawa (Rhinoceros sondaicus) merupakan salah satu megaherbivora
yang tidak ikut punah pada saat terjadi perubahan iklim global di satu periode
geologis yang dikenal dengan nama periode Holocene. Pada periode ini banyak
spesies menjadi punah (terutama herbivora besar) akibat perubahan iklim yang
berujung pada perubahan sebaran dan kelimpahan vegetasi (Williams et al. 2002). Hal
ini menunjukkan tingkat ketergantungan yang tinggi pada herbivora besar, termasuk
badak jawa, terhadap struktur vegetasi di lingkungan/habitat tempat tinggalnya;
padahal vegetasi itu sendiri bukanlah komponen habitat yang statis.
Perubahan habitat badak -walaupun tidak se ekstrem perubahan yang terjadi
pada periode holocene- telah teramati oleh beberapa penelitian diantaranya adalah
oleh Muntasib et al. (2002) yang melihat adanya kecenderungan dari sejenis
tumbuhan langkap (Arenga obtusifolia) untuk mendominasi habitat badak di Taman
Nasional Ujung Kulon. Kecenderungan seperti ini mengakibatkan menurunnya
ketersediaan pakan badak serta berpotensi untuk mengakibatkan berubahnya iklim
mikro. Penelitian yang dilakukan oleh Sarma et al. (2009) juga menjelaskan bahwa
suksesi alami mengakibatkan perubahan pada habitat dan perubahan ini
mengakibatkan penurunan dari ketersediaan pakan di padang rumput yang dihuni oleh
badak india (Rhinoceros unicornis) di suaka margastwa Pobitora, Assam, India.
Perubahan pada habitat dapat terjadi secara berangsur-angsur ataupun secara
drastis akibat bencana alam seperti letusan gunung berapi. Letusan Krakatau di tahun
1883 adalah salah satu contoh fenomena alam yang dapat dikategorikan sebagai
bencana klimatik yang mengakibatkan perubahan signifikan pada vegetasi di daerah
sekitarnya (Budyko 1999). Oleh karena itu, berbagai fenomena alam dapat menjadi
risiko yang mengancam kepunahan badak Jawa secara langsung seperti epidemi
penyakit, letusan gunung berapi, dan tsunami; maupun secara tidak langsung seperti
perubahan iklim drastis ataupun perubahan vegetasi akibat suksesi yang terjadi secara
alamiah.
Populasi badak Jawa di semenanjung Taman Nasional Ujung Kulon
menghadapi ancaman risiko penyakit yang dapat mendorong populasi ini menuju
kepunahan. Pada awal tahun 1982, lima ekor badak ditemukan mati di daerah selatan

142
semenanjung Ujung Kulon dengan dugaan adanya penyakit menular (Strien 1982).
Dugaan ini muncul didorong dengan adanya temuan kematian satwa lain di
semenanjung Ujung Kulon dan beberapa kematian pada kerbau di sekitar kawasan
Taman Nasional. Kasus kematian karena penyakit masih ditemukan pada seekor
badak betina di tahun 2003.
Kasus kematian badak masih berlanjut di tahun 2010 dimana 3 tumpukan
tulang belulang yang berasal dari 3 ekor badak ditemukan di semenanjung Ujung
Kulon dan analisis menunjukkan adanya agen infeksius Trypanosoma evansi yang
kemungkinan dapat menulari badak dan menyebabkan kematian (Hariyadi et al.
2011). Berdasarkan kasus kematian dari tahun 2000 sampai 2010, perhitungan angka
mortalitas adalah 1 kematian setiap tahun. Dengan demikian, tiga kematian di tahun
2010 merupakan angka yang secara signifikan berada di atas rata-rata angka kematian
per tahun. Sebaran temuan kasus kematian badak dari tahun 2000 sampai 2010
ditampilkan pada Gambar 33.
Gambar 33. Lokasi temuan kematian badak Jawa dalam periode tahun 2000-2010.
Titik hijau menunjukkan kasus dengan informasi yang relatif lengkap
mengenai penyebab kematian.
Keberadaan ancaman penyakit terhadap badak jawa diperkuat dengan adanya
hasil penelitian yang dilakukan oleh Khairani (2009) yang menunjukkan adanya agen
infeksius berupa parasit darah dan juga cacing yang dapat menular dari ternak kerbau

143
ke badak Jawa dan bahkan kepada manusia (zoonosis). Berdasarkan temuan-temuan
ini, para pakar yang tergabung dalam kelompok spesialis badak Asia: Asian Rhino
Specialist Group (AsRSG) merekomendasikan untuk menyusun sebuah protokol
kajian risiko penyakit serta pemantauan penyakit yang dapat menyerang populasi
badak Jawa. Penyusunan protokol yang dilengkapi dengan pelatihan bagi staf
lapangan merupakan langkah penting yang akan meningkatkan efektifitas patroli serta
meningkatkan peluang untuk menemukan dan mencegah penyakit-penyakit yang
membahayakan populasi badak Jawa.
Tekanan yang Dihadapi oleh Badak Jawa
Populasi badak jawa di Taman Nasional Ujung Kulon telah diketahui sebagai
populasi yang jumlahnya tidak lebih dari 50 individu saja, sementara rata-rata
perkembangan populasinya tidak lebih dari 1% saja setiap tahunnya (Hariyadi et al.
2011). Pemerintah Indonesia telah mentargetkan pertumbuhan populasi badak jawa
untuk bertambah dengan tingkat laju pertumbuhan populasi 3% setiap tahunnya.
Perbandingan kondisi pertumbuhan populasi yang teramati saat ini dengan
target yang telah dicanangkan oleh pemerintah menunjukkan adanya kesenjangan
(gap) pada laju pertumbuhan populasi badak tersebut. Kesenjangan ini diharapkan
dapat dikurangi dengan meminimalisir cekaman yang dihadapi oleh badak, sehingga
mereka dapat melakukan adaptasi secara optimal terhadap perubahan yang terjadi di
habitatnya. Penelitian ini telah mengidentifikasi berbagai cekaman yang dihadapi
badak jawa di habitat aslinya, dan cekaman ini antara lain terdiri dari:
1) Defisit Energi Asal Pakan
Perbandingan jumlah asupan nutrien (protein dan lemak) dalam penelitian ini
dengan asupan nutrien (protein dan lemak) pada badak sumatera yang dilakukan oleh
Dierenfeld et al. (2000) menunjukkan jumlah asupan yang relatif sama. Hal yang
perlu dicermati adalah perlakuan pada badak sumatera dicatat berdasarkan asupan
yang diterima badak sumatera di dalam fasilitias penangkaran yang biasanya memiliki
kualitas asupan lebih rendah dari kualitas asupan yang ada di alam (Dierenfeld et al.
2000). Berdasarkan informasi ini, maka kualitas asupan pakan badak sumatera di
penangkaran berdasarkan literatur ini merupakan tingkat minimum yang diperlukan
oleh badak jawa di habitat alaminya.

144
Perhitungan dengan menggunakan formula Kleiber seperti yang dilakukan
oleh Clauss et al. (2005) pada badak india menunjukkan bahwa kebutuhan energi
ideal untuk mempertahankan homeostasis (maintenance energy) bagi badak jawa
adalah: 20,825-28,051 kkal per hari (badak 12 dan 13) dan 15,937-21-467 kkal per
hari (badak 18). Secara umum, jumlah asupan pakan yang tercatat di lapangan
menunjukkan rataan harian yang memadai untuk memenuhi kebutuhan energi badak
jawa, namun perhatian khusus perlu diberikan pada badak 18 (badak muda) yang
jumlah rataan energi hariannya hanya sedikit di atas kebutuhan minimum. Hal ini
terjadi akibat adanya penurunan asupan energi asal pakan pada bulan Desember.
Fakta ini menunjukkan adanya risiko bahwa kebutuhan energi kemungkinan tidak
dapat dipenuhi pada waktu-waktu tertentu dan harus segera dipenuhi dalam waktu
berikutnya. Jumlah asupan energi asal pakan pada badak jawa perlu dipastikan untuk
menjamin kemampuan badak untuk mempertahankan hidup dan berkembang biak.
Hasil kajian hematologi pada hewan model menunjukkan bahwa kondisi
defisit jumlah konsumsi pakan -bila terjadi- merupakan jenis cekaman bersifat akut
dan merupakan kondisi riil yang harus dihadapi oleh badak Jawa di Taman Nasional
Ujung Kulon. Data dari tabel 4 menunjukkan adanya kemungkinan bahwa pakan
sebagian besar pakan yang dipilih oleh badak adalah jenis-jenis dengan kandungan air
yang cukup tinggi (9-17% air) sehingga berat kering dari tumbuhan pakan menjadi
sangat kecil dibandingkan dengan berat segarnya. Ini merupakan salah satu
mekanisme yang dilakukan oleh badak untuk mencegah terjadinya defisit air yang
merupakan tingkat cekaman yang tinggi sebagaimana ditunjukkan dalam pengamatan
profil glukokortikoid dan juga induksi defisit air pada hewan model.
2) Defisit Air
Analisis kadar glukokortikoid dari feses badak menunjukkan adanya
kecenderungan peningkatan hormon cekaman pada musim kering dengan jumlah
kejadian hujan yang rendah setiap harinya. Perbedaan kadar hormon ini sangat jelas
terlihat (walaupun tidak berbeda nyata secara statistik) di semua badak contoh dalam
penelitian ini terutama badak 12. Badak 12 merupakan badak dengan kandungan air
yang paling kecil dari asupan pakannya sehingga hewan ini memerlukan ketersediaan
air minum yang banyak. Perhitungan kebutuhan air dilakukan berdasarkan informasi

145
dari Galpine (2006) yang menunjukkan kebutuhan air 0.03 liter air per kilogram berat
badan, dan perhitungan ini menunjukkan bahwa badak 12 dengan bobot 1,000 kg
memerlukan 28 L air setiap harinya. Hasil kajian pakan dalam penelitian ini
menunjukkan bahwa badak 12 hanya mendapatkan rataan 5.01 L air dari tumbuhan
pakan dan harus mendapatkan 23 liter dari minum setiap harinya. Kebutuhan akan air
pada badak 12 ini akan meningkat pada musim kering di mana beberapa sumber air
mengering dan mengakibatkan menurunnya ketersediaan air. Hal ini konsisten
dengan temuan tingginya kadar glukokortikoid dari feses badak 12 di musim kering
sekitar bulan Oktober 2010 (0,2 kejadian hujan per hari). Fakta ini diperkuat dengan
kecenderungan yang sama saat cekaman defisit air disimulasikan pada kuda yang
menunjukkan peningkatan kortisol secara eksponensial. Kajian hematologi pada
hewan model menunjukkan bahwa kondisi defisit air merupakan cekaman yang
bersifat kronis.
Model Pengelolaan Badak Jawa
Hasil penelitian ini memberikan beberapa opsi yang potensial untuk
diterapkan sebagai bagian dari pengelolaan badak jawa. Opsi-opsi ini dapat dijadikan
model pengelolaan dengan pendekatan baru (basis pengkayaan nutrisi dan mitigasi
cekaman) yang belum pernah dirancang dan diimplementasikan secara optimal
sebelumnya. Model pengelolaan Badak Jawa berbasis nutrisi dan mitigasi cekaman
ditampilkan pada Gambar 34 dan terdiri dari berbagai komponen yaitu:
1. Komponen pemantauan yang terdiri dari: pemantauan status nutrisi dan
status cekaman berdasarkan profil hormon glukokortikoid dan/atau
metabolitnya.
2. Komponen pengkayaan nutrisi berupa intervensi habitat secara aktif untuk
memastikan ketersediaan tumbuhan pakan dengan kualitas nutrisi yang
tinggi untuk badak di Taman Nasional Ujung Kulon
3. Komponen mitigasi cekaman dengan memastikan ketersediaan air serta
akses menuju air yang tersedia sepanjang tahun.
4. Komponen riset menggunakan hewan model untuk mempelajari kecernaan
dan reaksi tanggap cekaman untuk mengantisipasi dan meminimalisir
dampak negatif dari nutrisi dan kecernaan, dampak negatif dari cekaman,
serta implikasi kesehatan lainnya.

146
Gambar 34. Model pengelolaan populasi dan habitat badak jawa dengan pendekatan
aspek nutrisi, cekaman, dan kesehatan
Komponen 1: Teknik Pemantauan
Pemantauan Status Nutrisi
Keberhasilan pengamat dalam mengikuti pola pergerakan badak (trajektori)
serta identifikasi lokasi-lokasi yang menjadi areal makan badak (rumpang)
menunjukkan potensi pengembangan dari teknik ini menjadi suatu teknik pemantauan
kuantitas dan kualitas nutrisi yang terjadi di habitat alami badak. Kemampuan
pemetaan dan analisis spatial akan mendorong keakuratan dari metode ini ke tingkat
yang lebih tinggi dan dapat memberikan informasi yang sahih bagi pihak pengelola
populasi badak jawa
Pemantauan Cekaman Berdasarkan Profil Hormon
Penelitian ini menunjukkan bahwa badak kemungkinan besar bisa beradaptasi
terhadap kondisi defisit pakan ini sebagaimana ditunjukkan oleh rendahnya kadar
hormon cekaman (kelas glukokortikoid) dari feses badak yang mengalami defisit
pakan (individu 12). Kemampuan badak untuk beradaptasi saat ini ditunjang dengan
kemampuan jelajah badak di areal yang masih cukup banyak ditumbuhi vegetasi
pakan yang disukai badak (palatabilitas tinggi). Literatur juga menyebutkan

147
kemungkinan adanya mekanisme internal dalam fisiologi tubuh badak yang
melakukan supresi terhadap sekresi glukokortikoid (Linklater et al. 2010).
Kecenderungan (trend) kenaikan kadar kortisol dalam darah dan penurunan kadar
kortisol tersebut pada pola cekaman yang muncul pada kuda sebagai hewan model
yang digunakan untuk mempelajari respons akibat kondisi cekaman terkait dengan
defisit pakan. Kemampuan mendeteksi kadar glukokortikoid merupakan hal yang
penting dalam mengembangkan protokol pemantauan yang memungkinkan pengamat
untuk mengambil data tingkat cekaman yang dihadapi oleh populasi badak di habitat
alaminya.
Komponen 2: Pengkayaan Nutrisi
Berkurangnya ketersediaan pakan bagi badak Jawa disebabkan oleh berbagai
hal yang sebagian tidak dapat dikendalikan seperti suksesi alami dan perubahan iklim.
Namun demikian, sebagian dari penyebab berkurangnya ketersediaan pakan masih
dapat dikendalikan seperti: dominasi/invasi vegetasi tertentu (Arenga sp atau Lantana
camara), dan kualitas nutrisi yang kurang memadai. Pengkayaan nutrisi dapat
dilakukan dengan tiga langkah yang terdiri dari: mengendalikan tumbuhan invasif;
meningkatkan ketersediaan tumbuhan dengan kandungan air, nutrien, dan energi
tinggi; serta mengurangi tumbuhan Lantana camara yang memiliki kandungan toksin.
Menekan Laju Invasi dan Dominasi Tumbuhan Langkap (Arenga obtusifolia)
Tumbuhan jenis palma ini memiliki pola penyebaran dengan menggunakan
akar dan biji. Tumbuhan langkap ini memiliki sifat alelopatik yang menghambat
pertumbuhan vegetasi lainnya di areal tempat mereka tumbuh. Risiko invasi vegetasi
dapat ditentukan menggunakan protokol yang dikenal dengan Weed Risk Assessment
(Dawson et al. 2009). Penyebaran tumbuhan langkap ini dapat dihambat dengan
bernagai cara seperti menebang atau menggunakan herbisida sebagaimana telah
dilakukan oleh YMR (2004). Talukdar et al. (2010) menyebutkan bahwa WWF telah
melakukan uji coba penebangan tumbuhan langkap ini dan mencatat bahwa areal uji
coba tersebut ditumbuhi oleh tumbuhan pakan badak tiga bulan setelah tumbuhan
langkap ditebang dari areal ini.
Setelah tumbuhan pakan tumbuh, areal ini kemudian dikunjungi untuk
pertama kalinya oleh badak Jawa (induk dan anak) yang menunjukkan bahwa
intervensi habitat semacam ini memiliki potensi untuk memberikan dampak positif

148
berupa peningkatan akses bagi badak ke areal yang tadinya tidak pernah dikunjungi
(potensi perluasan ruang jelajah). Tumbuhnya vegetasi yang merupakan makanan
badak menunjukkan bahwa pengelolaan semacam ini berpotensi untuk meningkatkan
ketersediaan pakan yang dibutuhkan oleh badak disamping juga berpotensi untuk
membuka ruang jelajah baru yang lebih luas. Analisis nutrisi dan kecernaan
menunjukkan bahwa ruang jelajah yang luas memiliki korelasi yang kuat dengan
keragaman pakan. Oleh karena itu, meningkatnya luas ruang jelajah memiliki potensi
untuk membantu badak menemukan keragaman pakan yang lebih tinggi.
Dengan mempertimbangkan berbagai potensi dari upaya untuk menekan laju
invasi dan dominasi tumbuhan langkap, maka upaya pengelolaan perlu
memperhatikan dan memilih lokasi-lokasi ideal untuk mengimplementasikan upaya
ini agar memberikan hasil yang optimal. Dengan mengetahui sebaran badak di
semenanjung Taman Nasional Ujung Kulon saat ini (Gambar 35), maka lokasi-lokasi
ideal untuk melakukan pengendalian langkap ini ditampilkan dalam gambar 36.
Gambar 35. Distribusi badak jawa di semenanjung Taman Nasional Ujung Kulon
(titik merah) berdasarkan hasil survey 2005-2010. (sumber: WWF
Indonesia & Balai TNUK)

149
Pengkayaan Air, Nutrien, dan Energi dari Tumbuhan Pakan
Selain defisit dari jumlah asupan pakan, referensi silang antara tumbuhan
pakan dengan palatabilitas tinggi dengan kandungan nutrisinya (protein, lemak,
energi) menunjukkan bahwa jenis tumbuhan pakan dengan palatabilitias tinggi tidak
mengandung kualitas nutrisi yang tinggi. Hal ini merupakan masalah yang cukup
besar dalam konteks asupan dan nutrisi bagi badak Jawa di Taman Nasional Ujung
Kulon.
Gambar 36. Lokasi ideal untuk plot pengendalian langkap (Arenga obtusifolia) di
semanjung Ujung Kulon ditandai dengan poligon berwarna merah.
Berdasarkan identifikasi jenis-jenis tumbuhan pakan dalam analisis nutrisi dan
kecernaan, maka tindak lanjut yang paling memungkinkan adalah membuat
persemaian khusus untuk memperbanyak tumbuhan pakan bernutrisi tinggi yang
terdiri dari: Moringa citrifolia, Callicarpa longifolia, Chisocheton microcarphus,
(protein tinggi); Alstonia angustiloba, Callicarpa longifolia, Macaranga spp, (lemak
tinggi); Derris thyorsifolia, Pterospermum javanicum, Percampyulus glances, (energi
tinggi); Paederia scandens, Alstonia scholaris, Costus speciosus (kandungan air
tinggi).

150
Hasil dari persemaian ini dapat kemudian digunakan untuk memperkaya areal
pakan (rumpang) badak yang sudah ada ataupun yang akan dibuat dengan
memperhitungkan kerapatan dari tumbuhan pakan ideal yaitu kerapatan 5,406
individu/ha (jenis tumbuhan bawah) ; 2,222 individu/ha untuk jenis semai ; 268
individu/ha untuk jenis pancang ; 32 individu/ha untuk jenis tiang ; dan 15 individu
per hektar untuk jenis pohon.
Pengendalian Lantana camara
Hasil analisis pakan dan palatabilitias tumbuhan pakan menunjukkan bahwa
individu badak nomor 13 menunjukkan pola asupan antinurisi lantaden yang relatif
lebih tinggi dibandingkan dengan kedua ekor badak lainnya. Hal ini menunjukkan
kemungkinan bahwa daerah pakan di dalam ruang jelajah badak nomor 13 ini
memiliki jumlah tumbuhan Lantana camara yang lebih tinggi dibandingkan daerah
pengamatan lainnya, dan oleh karenanya patut dijadikan prioritas dalam upaya
pemeliharaan habitat badak di Taman Nasional Ujung Kulon. Lokasi ideal bagi
upaya pengendalian Lantana camara ditampilkan dalam Gambar 37.
Pengurangan tumbuhan Lantana camara diharapkan dapat membantu
pertumbuhan tumbuhan pakan lain yang memiliki nutrisi tinggi. Untuk memastikan
ketersediaan pakan dengan nutrisi tinggi yang telah diidentifikasi, upaya pembibitan
dan penanaman tumbuhan-tumbuhan tersebut dapat dilakukan di areal pakan badak
yang telah dikurangi jumlah Lantana nya, ataupun di areal pengendalian langkap
(Arenga obtusifolia). Upaya optimalisasi habitat seperti ini dapat diperkuat dengan
upaya pembuatan akses terhadap kubangan atau air dengan cara membuatkan jalur
lintasan yang memungkinkan badak untuk mencapai sumber air dan/atau kubangan
yang dibutuhkannya. Cara lainnya adalah dengan membuat sumber air, kubangan
buatan, ataupun daerah resapan garam secara manual untuk memastikan ketersediaan
komponen ini demi menunjang kehidupan badak Jawa.

151
Gambar 37. Usulan areal pengendalian tumbuhan Lantana camara di areal pakan
badak ditandai dengan poligon berwarna kuning.
Komponen 3: Mitigasi Cekaman Akibat Defisit Air
Defisit air merupakan satu faktor yang cukup sulit untuk ditangani karena
penyebab dari defisit air ini adalah faktor iklim global (curah hujan). Air berperan
penting dalam kehidupan badak sebagai sumber air minum dan sebagai fungsi
termoregulasi (berkubang).
Cara-cara yang dapat dilakukan oleh pihak pengelola untuk membantu badak
dalam beradaptasi terhadap kondisi defisit air ini adalah dengan membuat akses bagi
badak ke sumber-sumber air permanen yang tidak pernah kering sepanjang tahun.
Akses ini dapat dilakukan dengan membuat areal pakan (rumpang) ataupun membuat
lintasan menuju kantong-kantong air tersebut. Cara lainnya adalah dengan
memperkaya areal pakan (rumpang) badak dengan tumbuhan pakan yang kaya air dan
bila memungkinkan kaya akan nutrisi. Jenis-jenis tumbuhan dengan karakter seperti
ini adalah tumbuhan Areuy kipuak (Paederia scandens), Lame kuning (Alstonia
scholaris), dan Pacing (Costus speciosus).
Langkah pengelolaan lain yang dapat dilakukan adalah dengan membuat
kubangan di tempat-tempat yang telah diperhitungkan secara strategis. Pembuatan

152
kubangan yang biasanya didampingi juga dengan pembuatan salt lick dilakukan untuk
memfasilitasi fungsi termoregulasi sekaligus untuk menjaga integritas kulit yang
dibutuhkan oleh badak. Salt lick berfungsi sebagai reservoir di mana badak
mendapatkan garam yang dibutuhkan untuk proses metabolisme.
Komponen 4: Penggunaan Hewan Model untuk Riset
Hasil penelitian ini menunjukkan bahwa analogi hewan model dengan badak
jawa dapat diperkuat dalam aspek-aspek tertentu. Kaidah penggunaan hewan model
dalam riset biomedis dapat pula diterapkan dalam upaya konservasi badak jawa
terkait aspek kecernaan dan nutrisi, serta mitigasi cekaman dan mitigasi penyakit.
Kecernaan dan Nutrisi
Dalam konteks pengelolaan populasi badak jawa di Taman Nasional Ujung
Kulon, hewan model dapat digunakan untuk mempelajari aspek komposisi diet,
nutrisi, dan kecernaan secara lebih dalam serta mempelajari implikasinya terhadap
aspek kesehatan hewan yang terdiri dari:
1. Kesehatan reproduksi (dapat dipantau melalui profil siklus pada betina atau
kadar hormon androgen pada badak jantan)
2. Kesehatan secara umum (dipantau melalui pemantauan agen penyakit).
Hewan model (kuda) dapat digunakan untuk menemukan komposisi pakan
yang optimal bagi badak jawa. Berbagai komposisi pakan dengan kandungan air,
energi, nutrien, dan mineral yang berbada dapat diuji coba pada kuda untuk
mempelajari tingkat kecernaan, asupan gizi, implikasi terhadap pakan terhadap
kapasitas reproduksi (siklus, profil hormon, sperm count), toleransi terhadap
cekaman, dan toleransi terhadap penyakit ataupun agen infeksius tertentu.
Informasi dari komponen ini berupa perhitungan komposisi pakan yang tepat
dapat digunakan untuk memberi masukan untuk upaya pengkayaan nutrisi badak pada
komponen 2.

153
Pencegahan Cekaman dan Implikasi Kesehatan
Dalam konteks penelitian lanjut terkait pengelolaan populasi badak jawa di
Taman Nasional Ujung Kulon, hewan model (kuda) dapat digunakan untuk
melakukan berbagai uji coba dan kuantifikasi dari sumber cekaman yang ada di
habitat alami serta mengantisipasi reaksi tanggap terhadap sumber cekaman tersebut.
Sumber cekaman yang dapat digunakan dalam uji coba menggunakan hewan model
adalah reaksi tanggap fisiologis terhadap cekaman serta mengukur ambang batas
tingkat cekaman (intensitas dan durasi) yang dapat ditolerir dengan melakukan
ekstrapolasi data uji coba hewan model. Ambang batas toleransi hewan terhadap
cekaman dapat dikembangkan lebih lanjut untuk melakukan kuantifikasi sehingga
para pengelola dapat mengantisipasi tingkat cekaman pada badak jawa dan dapat
mencegah peningkatan intensitas dan durasu cekaman tersebut sehingga implikasi
klinis dan dampak negatif cekaman tersebut terhadap kesehatan pada populasi badak
jawa dapat dicegah.
Faktor-faktor Lain yang Relevan
1) Faktor Habitat
Kelimpahan pakan merupakan kunci pertahanan hidup badak jawa. Oleh
karena itu, selain tersedianya habitat bagi badak Jawa di Taman Nasional Ujung
Kulon, pemerintah Indonesia telah mencanangkan perlunya untuk melakukan
optimasi dan perluasan habitat serta mencari habitat kedua demi membentuk populasi
kedua untuk melengkapi populasi yang sudah ada di Taman Nasional Ujung Kulon.
Untuk mencapai tujuan ini, para pengambil keputusan dan pembuat kebijakan perlu
mempertimbangkan faktor-faktor habitat seperti: ketersediaan air, ketersediaan pakan,
dan juga kualitas pakan yang tersedia mengingat faktor-faktor tersebut merupakan
sumber cekaman bagi badak bila tidak dikelola dengan baik dan optimal.
2) Faktor Populasi
Komposisi dan demografi dalam populasi badak yang ada saat ini merupakan
informasi yang penting dalam pengelolaan badak Jawa saat ini. Daya dukung alam
yang ada saat ini diduga sudah mencapai kapasitas optimal untuk populasi badak
jawa, dan persaingan antar individu badak untuk mendapatkan sumber daya untuk
bertahan hidup (makanan, air, pasangan) akan semakin ketat. Faktor ini perlu
dicermati oleh pengelola populasi agar keberadaan badak jawa di Taman Nasional

154
Ujung Kulon dapat terus berlanjut. Selain populasi di Ujung Kulon, para ahli sudah
menyepakati perlunya pembentukan populasi kedua di luar Taman Nasional Ujung
Kulon. Pembentukan populasi kedua akan menimbulkan dampak negatif bagi
populasi induk dan populasi baru bila komposisi rasio kelamin tidak seimbang dalam
memilih pionir bagi populasi kedua. Selain itu, pemilihan induk perlu
mengidentifikasi banyaknya individu betina produktif yang masih mampu
menghasilkan keturunan. Informasi ini kemudian ditindak lanjuti dengan memilih
induk yang sesuai untuk membentuk populasi baru, namun meninggalkan individu-
individu betina produktif lainnya di Taman Nasional Ujung Kulon untuk memastikan
bahwa populasi Ujung Kulon masih memiliki kapasitas untuk berkembang biak.
Informasi seperti ini bisa didapatkan melalui: identifikasi visual kamera ataupun video
jebak, analisis hormon reproduksi jantan dan betina melalui sampel feses, dan juga
mencatat (track record) kelahiran badak dari tahun ke tahun.
3) Faktor Individu dan Perilaku Hewan
Kesehatan dan kesejahteraan individu badak merupakan faktor yang sangat
penting dalam konsep pelestarian badak terutama dalam konteks perluasan habitat dan
pembentukan populasi baru di habitat kedua. Hariyadi et al. (2011) menunjukkan
secara kualitatif tentang adanya risiko penyakit yang terdiri dari bakteria (E.coli,
Salmonella spp, dan Clostridium sp) serta risiko tertinggi yang berasal dari parasit
Trypanosoma evansi. Oleh karenanya, pengelola populasi badak Jawa idealnya
menerapkan upaya untuk mencegah penyebaran atau mencegah risiko patogenisitas
dari agen-agen infeksius di atas.
Cara yang dapat dilakukan adalah dengan mencegah interaksi antara hewan
ternak masyarakat dengan hewan liar di Taman Nasional Ujung Kulon yang
berpotensi menjadi vektor ataupun reservoir bagi agen infeksius. Hal ini dapat
dilakukan dengan mendorong masyarakat untuk mengandangkan ternak mereka atau
menggembalakan ternak mereka di luar kawasan Taman Nasional Ujung Kulon.
Cara lain yang tidak kalah pentingnya adalah upaya untuk mengurangi risiko
ataupun tingkat cekaman bagi badak Jawa. Minimnya tingkat cekaman akan
memungkinkan badak untuk mengoptimalkan asupan nutrisi dan mineral untuk
meningkatkan daya tahan tubuh mereka serta meningkatkan kekebalan tubuh terhadap
penyakit.

155
SIMPULAN DAN SARAN UMUM
Simpulan Umum
Setelah semua informasi dianalisis dan dirangkum dari setiap bagian dalam disertasi
ini, maka hasil penelitian dalam disertasi ini dapat dirumuskan menjadi berbagai
simpulan dan saran/rekomendasi sebagai berikut:
1. Kondisi habitat berkorelasi kuat dengan keragaman pakan yang juga
mempengaruhi ketersediaan nutrisi, air, dan energi asal pakan untuk badak
jawa. Setidaknya satu wilayah jelajah badak dalam penelitian ini menunjukkan
rataan konsumsi energi dan air yang rendah. Kandungan air asal pakan
mengalami fluktuasi dan pada musim kering terlihat ada kecenderungan
penurunan pada kandungan air asal pakan di musim kering
2. Metabolit hormon kortisol berupa 3α, 11β-dihidroxy-CM diduga memiliki
potensi untuk digunakan sebagai indikator aktifitas korteks adrenal akibat
adanya cekaman. Pada musim kering terjadi kecenderungan peningkatan
cekaman pada badak.
3. Analogi antara badak jawa dan kuda terbangun dan diperkuat dengan
informasi dari berbagai literatur serta dari pengamatan langsung dalam
penelitian ini. Kuda adalah hewan model yang layak digunakan untuk
penelitian badak jawa terkait dengan aspek spesifik terkait perilaku,
kecernaan, dan respons fisiologis terhadap cekaman terutama cekaman yang
bersifat akut.

156
Saran Umum
1. Perlu penyusunan protokol pemantauan reguler dengan menggunakan
parameter yang telah dikaji dalam penelitian ini, serta mengoptimalkan
penggunaan hewan model untuk mensimulasikan berbagai peubah yang dapat
mempengaruhi kesejahteraan dan keberlangsungan hidup badak jawa seperti:
cekaman dan penyakit
2. Pemantauan populasi dengan teknik yang lebih canggih untuk memantau
status nutrisi, cekaman, dan kapasitas reproduksi (siklus pada betina dan kadar
testosteron pada jantan)
3. Pengkayaan habitat dengan mendorong perbanyakan tumbuhan pakan
terutama yang mengandung nutrien tinggi. Pengkayaan habitat seperti ini
dapat dikaitkan dengan upaya pengendalian tumbuhan pengganggu serta
mengurangi jumlah tumbuhan yang mengandung toksin dan/atau antinutrisi.r

DAFTAR PUSTAKA
Agil M, Purwantara B, Alikodra HS, Hodges K, Toelihere MS. 2007. Reproductive
biology of the sumatran rhinoceros Dicerorhinus sumatrensis (Fischer 1814).
Disertasi program Doktor Institut Pertanian Bogor. Sekolah Pasca Sarjana IPB
Agil M, Setiadi DR, Supriatna I, Purwantara B,. 2008. Non-Invasive Endocrine
Monitoring of Reproduction and Stres in the wild animal: Analyzing hormone
metabolites in Urine and Faeces using Enzymeimmunoassay. Proceeding of
AZWMC 2008
Albaugh GP, et al. 1992. Isolation of exfoliated colonic epithelial cells, a novel, non-
invasive approach to the study of cellular markers. Int J Cancer. 1992 Sep
30;52(3):347-50
Allen CW, White CB,. 1998. Glucose modulates cell death due to normobaric
hyproxic by maintaining cellular ATP. Am J Physiol Lung Cell Mol Physiol
274:159-164
Bachman I, et al. 2003. Behavioural and physiological responses to an acute stressor
in crib-biting and control horses. Applied animal behaviour science 82: 297-
311
Balai Taman Nasional Ujung Kulon (BTNUK). 2010. Pemantapan zonasi di Taman
Nasional Ujung Kulon. Direktorat Jendral Perlindungan Hutan dan Konservasi
Alam (PHKA), Kementrian Kehutanan RI.
Bansal N, Houle A, Melnykovych G,. 1991. Apoptosis: mode of cell death induced in
T Cell leukimia lines by dexamethasone and other agents. FASEB J. 5:211-216
Barja I, et al. 2007. Stress physiological response to tourist pressure in the wild
population of European pine marten. The journal of steroid biochemistry and
molecular biology. (104)3-5:136-142
Baron R. 2006. Mechanism of disease: neuropathic pain-a clinical perspective. Nature
clinical practice neurology. 2:95-106
Bennet RM,. 1999. Emerging Concepts in the Neurobiology of Chronic Pain:
Evidence of Abnormal Sensory Processing in Fibromyalgia. Mayo Clin
Proc;74:385-398
Berridge MJ,. 1984. Inositol Trisphosphate and Diacylglycerol as Second
messengers. Biochem J 220: 345-360
Birdane FM, et al. 2004. Renal Cell Carcinoma in a Dog: Pathologic and Cytologic
Findings. Revue Méd. Vét., 155, 3, 212-216

158
Birkett A, Stevens-Wood B,. 2005. Effect of low rainfall and browsing by large
herbivores on an enclosed savannah habitat in Kenya. African journal of
ecology 43: 123-130
Brown GE, et al. 2002. Acute hormonal response to sublingual androstenediol intake
in young men. Journal of applied Physiology 92:142-146
Bruni JF, Hawkins RL, Yen SS,. 1982. Serotonergic Mechanism in the Control of
Beta-endorphin and ACTH Release in Male Rats. Life Sci. 30(15): 1247-1254
Budyko M,. 1999. Climate catastrophes. Global and planetary change 20: 281-288
Buff PR, Morrison CD, Ganjam VK, Keisler DH,. 2005. Effects of short-term feed
deprivation and melatonin implants on circadian patterns of leptin in the horse.
Journal of animal science 83(5): 1023-1032
Cameron HL, Perdue MH. 2005. Stres Impairs Murine Intestinal Barrier Function:
Improvement by Glucagon-Like Peptide-2. The journal of pharmacology and
experimental therapeutics Vol. 314, No. 1
Carlstead K, Brown JL,. 2005. Relationships between patterns of fecal corticoid
excretion and behaviour, reproduction, and environmental factors in captive
black (Diceros bicornis) and white (Ceratohterium simum) Rhinoceros. Zoo
biology 24: 215-232
Cavaliere F, et al. 2001. Hypoglycaemia-induced cell death: features of
neuroprotection by the P2 receptor antagonist basilen blue. Neurochemistry
International Volume 38, Issue 3: 199-207
Chaffin MK, Cohen ND,. 1999. Diagnostic Assessment of Foals with Colic.
Proceedings of the Annual Convention of the AAEP vol 45
Citino SB, Bush M,. 2007. Reference cardiopulmonary physiologic parameters for
standing, unrestrained white rhinoceroses (Ceratotherium simum). Journal of
Zoo and Wildlife Medicine 38(3): 375-379
Clausen B,. 1981. Survey For Trypanosomes in Black Rhinoceros (Diceros bicornis).
Journal of Wildlife Diseases Vol. 17, No. 4, October, 1981
Clauss M, et al. 2005. Studies on digestive physiology and feed digestibilities in
captive Indian rhinoceros (Rhinoceros unicornis). Journal of Animal
Physiology and Animal Nutrition 89 (2005) 229–237
Clauss M, et al. 2005. Energy and mineral nutrition and water intake in the captive
indian rhinoceros (Rhinoceros unicornis). Zoo Biology 24: 1-14
Clauss M, et al. 2006. Digestion coefficients achieved by the black rhinoceros
(Diceros bicornis) a large, browsing, hidgut fermenter. Journal of animal
physiology and animal nutrition. 1-10

159
Coenen M,. 2005. Exercise and stress: impact on adaptive processes involving water
and electrolytes. Livestock production science 92:131-145
Corbett GB, Hill JE. 1992. The mammals of Indomalayan region: a systematic
review. Natural History Museum Publications. Oxford University Press.
Colgan, PW. 1978. Quantitative ethology. Wiley-Interscience publications. John
Wiley and Sons, New York
Cranbrook E, Piper PJ,. 2007. Short Communications: Javan rhinoceros Rhinoceros
sondaicus in Borneo. The Raffles Bulletin of Zoology 2007 55(1): 217-220
Daszak P, Cunningham AA, Hyatt AD,. 2000. Emerging infectious diseases of
wildlife –threats to biodiversity and humant health. Science 287: 443-449
Daszak P,. et al. 2004. Conservation Medicine and New Agenda for Newly Emerging
Diseases. Ann. N.Y. Acad. Sci. 1026: 1–11
Davis AK, Maney DL, Maersz JC,. 2008. The use of leucocyte profiles to measure
stress in vertebrates: a review for ecologists. Functional ecology 22(5): 760-772
Dawson W, Burslem DFRP, Hulme PE,. 2009. The suitabilityof weed risk assessment
as a conservation tool to identify invasive pant threats in East African
rainforests. Biological Conservation (142): 1018-1024
Deem SL, et al. 2001. Conservation Medicine. Annals New York Academy of
Sciences pp: 370-377
DeSouza MJ, et al. 1994. Gonadal hormones and semen quality in male runners a
volume threshold effect of endurance training. International journal of sports
medicine 15(7): 383-391
Dhabhar FS,. 2000. Acute stress enhances while chronic stress supresses skin
immunity: the roles of stress hormones and leukocyte trafficking. Annals of the
New York academy of Sciences vol 917: Neuroimmunomodulation perspectives
at the new millenium:876-893
Dickens MJ, Delehanty DJ, Romero LM,. 2010. Stress: an inevitable component of
animal translocation. Biological conservation 143:1329-1341
Dierenfeld ES, Widman REC, Romo S,. 2000. Feed intake, diet utilization, and
composition browses consumed by the Sumatran rhino (Dicerorhinus
sumatrensis) in a north American zoo. Zoo Biology 19: 169-180
Dinerstein E, 2003. The return of the unicorns. The natural history and conservation
of the greater one-horned rhinoceros. Columbia Universtiy Press, New york
Domingues-Gerpe L, Rey-Mendez M,. 2001. Alterations induced by chronic stress in
lymphocyte subsets of blood and primary and secondary immune organs of

160
mice. BMC Immunology 2:7. http://www.biomedcentral.com/1471-2172/2. [9
Juni 2012]
Dyer J, et al. 2002. Molecular characterisation of carbohydrate digestion and
absorption in equine small intestines. Equine veterinary journal 34(4):349-358
Endo H, et al. 2009. The morphological basis of the armor-like folded skin of the
greater Indian rhinoceros as a thermoregulator. Mammal study 34: 195-200
Evans W, Borton A, Hints HF, Vleck LDV,. 1990. The Horse Second Edition. W.H.
Freeman and Company, New York
Fernando P, Polet G, Foead N. Ng L. Melnick DJM. 2004. Mitochondrial DNA
analysis of the critically endangered Javan Rhinoceros. PHKA-WWF-Columbia
University
Figueiredo HF, et al. 2003. Stress integration after acute and chronic predator stress:
Differential activation of central stress circuitry and sensitization of th
ehypothalamo-pituitary-adrenocortical axis. Endocrinology 144(12): 5249-5258
Fowler ME, Miller RE,. 2003. Zoo and Wild Animal Medicine 5th edition. Saunder
Publishing, St Louis Missouri. p: 559.
Francois-Gerard C, Gerard P, Rentier B,. 1988. Elucidation of non-parallel EIA
curves. Journal of immunological methods (111)1:59-65
Galisteo ML, et al. 1996. The Adaptor protein Nck Links receptor tyrosine kinases
with Serine-Threonin Kinase Pak1. The Journal of Biological Chemistry
271(35): 20997-2100
Galpine, NJ. 2006. Boma management of black and white rhinoceros at Mombo,
Okavango delta – Some lessons. Ecological Journal 7: 55-61
Gandolf AR, Willis AM, Blumer ES, Atkinson MW,. 1999. Management of a Melting
Corneal Ulcer in a Greater One-horned Rhinoceros (Rhinoceros unicornis).
Proceedings American Associations of Zoo Veterinarians
Garg SL, Chander S,. 1997. Plasma Cortisol and thyroid hormone Concentrations in
Buffaloes with uterine Torsions. Buffalo Bullettin Vol 16(4): 75-76
Ghalib, Supriatna I, Agil M, Engelhardt A,. 2011. Non-invasive hormone monitoring:
fecal androgen and glucocorticoid in male crested macaques (Macaca nigra) in
relation to seasonal and social factors. Thesis. Bogor Agriculture University
Glade MJ, Gupta S, Reimers TJ,. 1984. Hormonal Responses to High and Low Planes
of Nutrition in Weanling Thoroughbreds. J Anim Sci 1984. 59:658-665
Goebel MU, Mills PJ,. 2000. Acute psychological stress and exercise and changes in
peripheral leukocytes adhesion molecule expression and density. Psychosomatic
medicine 62: 664-670.

161
Gustincich S, Schenider C,. 1993. Serum Deprivation Response Gene Is Induced by
Serum Starvation but not by contact Inhibition. Cell Growth and Differentiation
vol 4: 753-760
Hao S, Avraham Y, Bonne O, Berry EM,. 2001. Separation-induced body weight loss,
impairment in alternation behavior, and autonomic tone: effects of tyrosine.
Pharmacology Biochemistry and Behavior Volume 68, Issue 2
Hariyadi ARS, Santoso A, Setiawan R, Priambudi A,. 2008. Automatic Camera
Survey for Monitoring Reproductive Pattern and Behaviour of Javan
Rhinoceros (Rhinoceros sondaicus) in Ujung Kulon National Park, Indonesia.
Proceeding of the 3rd
International Meeting on Asian Zoo/Wildlife Medicine and
Conservation (AZWMC)
Hariyadi ARS, Setiawan R, Daryan, Yayus A, Purnama H,. 2010. Preliminary
behaviour observations of the Javan rhinoceros (Rhinoceros sondaicus) based
on video trap surveys in Ujung Kulon National Park. Pachyderm 47: 93-99
Hariyadi ARS, et al. 2011. Estimating the population structure of the Javan rhino
(Rhinoceros sondaicus) in Ujung Kulon National Park using the mark-recapture
method based on camera and video trap identification. Pachyderm no 49: 90-99
Hariyadi ARS, Handayani, Priambudi A, Setiawan R,. 2011. Investigation of the
death of Javan rhinoceros (Rhinoceros sondaicus) in Ujung Kulon National
Park. Proceedings of 5th ASVP (Asian Society of Veterinary Pathology)
conference and congress: 32-34
Harley EH, Matshikiza M, Robson P, Weber B,. 2004. Red blood cell metabolism
shows major anomalies in Rhinocerotidae and Equidae, suggesting a novel role
in general antioxidant metabolism. Animals and Environments. Proceedings of
the Third International Conference of Comparative Physiology and
Biochemistry. Pp 334-340
Harmon DL, MacLeod KR,. 2001. Glucose Uptake and Regulation By Intestinal
Tissues: Implications and whole-body Energetics. J. Anim. Sci. 79(E.
Suppl.):E59–E72
Harvell CD, et al. 2002. Climate warming and disease risks for terrestrial and marine
biota. Science 296(5576):2158-2162
Hatt JM, Wenker C, Castell J, Clauss M,. 2004. Dietary dan Veterinary Management
of a Lingual Abscess in a Geriatric Captive Black Rhino (Diceros bicornis)
With Iron storage Disease. European Association of Zoo and Wildlife
Veterinarians (EAZWV) 5th Scientific Meeting. Ebeltoft-Denmark
Hearne JW, Swart J,. 1991. Optimal translocation strategies for saving the black
rhinoceros. Ecological modelling 59: 279-292

162
Helen PG, Brindley DN,. 1978. The Effects of Cortisol, Corticotropin, and thyroxine
on the synthesis of Glycerolipids and on Phospatidae Phophohydrolase activity
in Rat Liver. Biochem J. 176: 777-784
Hermes R, Hildebrandt TB, Goritz F,. 2004. Reproductive problems directly
attributable to long-term captivity –assymetric reproductive aging. Animal
reproduction science (82-83):49-60
Hermes R, Goritz F, Streich WJ, Hildebrandt TB,. 2007. Assisted reproduction in
female rhinoceros and elephants – Current status and future persepectives.
Reprod. Dom. Anim. 42(suppl 2):33-44
Hommel PWFM, 1987. Landscape ecology of Ujung Kulon (West java). Disertasi
universitas Wageningen, Belanda.
Hoogerwerf A. 1970. Udjung Kulon the land of the last javan rhinoceros. E.J. brills.
Leiden, Netherlands
Hutchin M, Kreger MD,. 2006. Rhinoceros behaviour: implications for captive
management and conservation. Int. Zoo. Yb 40:150-173
Huxman TE, Scott RL., 2007. Climate Change, Water Dynamics, and Landscape
Water Balance. Southwest Hydrology. January-February
IRF-WWF. 2011. Vietnam’s javan rhino now extinct. WWW.panda.org. [9 Juni 2012]
Izumi Y, et al. 1997. A protein Kinase Cδ binding Protein SRBC whose expression is
Induced by Serum Starvation. The Journal of Biological Chemistry Vol
272(11): 7381-7389
Jacobsen J,. 2005. A Review of Rhino Foot Problem. Fort Worth Zoo
Jensen TK, et al. 2002. Inhibin B as a serum marker of spermatogenesis: correlation
to differences in sperm concentration and follicle-stimulating hormone levels. A
study of 349 Danish men. Journal of clinical endocrinology and metabolism
82(12): 4059-4063
Jessup DA, et al. 1992. Retrospective Evaluation of Leptospirosis In Free-ranging and
Captive Black Rhinoceros (Diceros bicornis) by Microscopic Agglutination
Titers dan Fluorescence Antibody Testing. Journal of Zoo and Wildlife
Medicine 23(4): 401-408
Juster RP, McEwen BS, Lupien SJ. 2009. Allostatic load biomarkers of chronic stress
and impact on helath and cognition. Neuroscience and biobehavioural reviews:
1-15
Jolles J, et al. 1980. Modulation of brain Phosphoinotiside metabolism by ACTH-
sensitive Protein Phosporylation. Nature vol 289: 623-625
Jonyo JF,. 2005. Doctoring Rhinos: Diseases seen in Kenya

163
Kanemi O, et al. 2005. Acute stress reduces intraprenchymal lung natural killer cells
via beta-adrenergic stimulation. Clinical experimental immunology 139(1): 25-
34
Karl IE, Garber AJ, Kipnis DM,. 1976. Alanine and Glutamine Sythesis and Release
from Skeletal Muscle. The Journal of biological Chemistry 251(3): 844-850
Khairani KO, 2009. Disease surveillance and epidemiology of water buffalo (Bubalus
bubalis) surrounding Ujung Kulon national park for conservation program of
the Javan rhinoceros (Rhinoceros sondaicus) in Indonesia. Laporan kegiatan
International Rhino Foundation (IRF).
Kelley KW,. 1998. Cross-talk between the Immune and Endocrine Systems, J Anim
Sci. 66:2095-2108
Kloss CB,. 1927. Further records of the one-horned rhinoceros in the Malay States.
Journ. Fed. Malay States Mus. XIII, 1927
Kock N, et al. 1991. Coronary Artery Aneurism In Two Black Rhinoceroses (Diceros
bicornis) in Zimbabwe. Journal of Zoo and Wildlife Medicine 22(3): 355-358
Kock ND, et al. 1992. Serological Evidence for Cowdria Ruminantium Infection in
Free-ranging Black (Diceros bicornis) dan White (Ceratotherium simum)
Rhinoceroses in Zimbabwe. Journal of Zoo and Wildlife Medicine 23(4): 409-
413
Kock ND,. 1996. Myocardial Purkinje Degeneration and Necrosis with Fibrosis in
Free-Ranging Black Rhinoceroses (Diceros bicornis) in Zimbabwe. Journal of
Wildlife Diseases, :32(2), 1996, pp :367-369
Koolman J, Rohm KH, Wanandi SI[ed], Sadikin M [ed]. 2001. Atlas berwarna dan
teks Biokimia. Cetakan I. Penerbit Hipokrates, Jakarta
Kretzschmar P, Gansloßer U, Dehnhard M,. 2004. Relationships between androgens,
environmental factors and reproductive behaviours in male white rhinoceros
(Ceratotherium simum simum). Hormones and Behaviour 45: 1-9
Kudryavtseva NN, Amstislavskaya TG, Kucheryavy S,. 2004. Effects on repeated
aggressive encounters on approach to a female and plasma testosterone in male
mice. Hormones and behaviour 45: 103-107
Lekagul , McNeely. 1977. Mammals of Thailand. Sahakarnbhath Co. Bangkok
Linklater WL, MacDonald EA, Flamand JRB, Czekala NM,. 2010. Declining and
low fecal corticoids are associated with distress, not acclimatization to stress,
during the translocation of African rhinoceros. Animal conservation 13: 104-
111.

164
Mace OJ, Afflek J, Patel N, Kellet GL,. 2007. Sweet taste recpetors in rats small
intestines stimulate glucose absorption through apical GLUT2. Journal of
Physiology 582.1:379-392
Mainka SA, Zhao GL, Li M, 1989. Utilization of a bamboo, sugar cane, and gruel diet
by two juvenile giant pandas (Ailuropoda melanoleuca). Journal of zoo and
wildlife medicine 20:39-44
Mandial RK, 2006. An Insight into the Toxicological and Medicinal Properties of
Lantana camara plant. College of Veterinary and Animal Science Palampur
Himachal Pradesh
Martin PS, Stuart AJ,. 1995. Mammoth extinction: two continents and Wrangel
island. Radiocarbon Volume 37 no.1: 7-10.
McFadyen REC,. 1998. Biological control of weeds. Annual review of entomology
43:369-393
McSweeney CS, Pass MA,. 1983. The mechanism of ruminal stasis in Lantana-
poisoned sheep. Quarterly journal of experimental physiology 68: 301-313
Melia KL, Duman RS,. 1991. Involvement of corticotropin –releasing factor in
chronic stress regulation of the brain noradrenergic system. Proc. Natl. Acad.
Sci. USA. Vol 88: 8382-8386 Neurobiology
Menargues A, Urios V, Mauri M,. 2008. Welfare assessments of captive asian
elephants (Elephas maximus) and Indian rhinoceros (Rhinoceros unicornis)
using salivary cortisol measurements. Animal Welfare (17): 305-312
Moen AN, 1973. Wildlife Ecology An Analytical Approach. Cornell University. WH
Freeman and Company. San Fransisco
Morgan KN, Tromborg CT. 2007. Sources of stress in captivity. Applied animal
behaviour science 102: 262-302
Morilak DA, et al. 2005. Role of brain norepinephrine to behavioural response to
stress. Progress in neuro-psychopharmacology & biological psychiatry
29:1214-1224.
Morrow CJ, Kolver ES, Verkerk GA, Matthews LR,. 2000. Urinary corticosteroids:
an indicator of stress in dairy cattle. Proceedings of the New Zealand Society of
Animal Production 60: 218-221
Mundiany L, Agil M, Astuti DA,. 2005. Studi Kasus: Estimasi Gambaran Nutrien
pada Badak Sumatera (Dicerorhinus sumatrensis) Jantan di Suaka Rhino
Sumatera Taman Nasional Way Kambas. Skripsi Institut Pertanian Bogor
Munson L, Koehler JW, Wikonson JE, Miller RE,. 1998. Vesicular and Ulcerative
Dermatopathy resembling Superficial Necrotic Dermatitis in Captive Black
Rhinoceroses (Diceros bicornis). Vet Pathol 35: 31-42

165
Muntasib EKSH, Suratmo FG, Alikodra HS, Mardiastuti A, Prasetyo LB,. 2002.
Penggunaan ruang habitat oleh badak jawa (Rhinoceros sondaicus, Desm 1822)
di Taman Nasional Ujung Kulon. Disertasi Sekolah Pascasarjana Institut
Pertanian Bogor (IPB).
NRC. 1989. Nutrient requirements of horses, 5th ed. National Academy Press,
Washington DC.
Nagy K, et al. 2009. The effect of a feeding stress-test on the behaviour and the heart
rate variability of control and crib-biting horses (with or without inhibition).
Applied animal behaviour science 121: 140-147
Nayak BS, Raju SS, Rasumbhag A. 2008. Investigation of wound healing activity of
Lantana Camara L. In Sprague dawley Rats using burn wound model.
International Journal of Applied Research in Natural Products Vol 1(1): 15-19
Nessminth S, Baltzell J, Bardanier CD,. 1983. Interaction of Glucocorticoid and
thyroxine in the responses of rats starvation-refeeding J. Nutr. 113: 2260-2265
Newton FRS,. March 17 1874. The Secretary on Addition to the menagerie.
Noble GK, et al. 2007. Effect of excercise training, circadian rythm, age, and sex, on
insulin-like growth factor-1 in the horse. Journal of animal science 85: 163-171
PHKA. 2007. Strategi dan rencana aksi konservasi badak Indonesia. Direktorat
jendral Perlindungan hutan dan kelestarian Alam, kementerian kehutanan
Republik Indonesia.
Pagan JD, et al. 1998. Exercise affects digestibility and rate of passage of all-forage
and mixed diets in throroughbred horses. American society for nutritional
sciences. J. Nutr. 128: 2704S-2707S
Paglia DE, Radcliffe RW,. 2000. Anthracycline Cardiotoxicity in a Black Rhinoceros
(Diceros bicornis): Evidence for Impaired Antioxidant Capacity Compounded
by Iron Overload. Vet Pathol 37:86–88
Paplinska JZ, et al. 2007. Reproduction in male swamp wallabies (Wallabia bicolor):
puberty and the effects of season. Journal of Anatomy 211:518-533
Pass MA, Seawright AA, Lamberton JA, Heath TJ,. 1979. Lantadene A toxicity in
sheep. A model for cholestasis. Pathology. 1979 Jan;11(1):89-94
Permadi YF,. 2008. Kajian dampak perubahan iklim terhadap kerentanan badak
Jawa. Laporan proyek WWF Indonesia.
Plechner AJ,. 2004. Cortisol abnormality as a cause of elevated estrogen and immune
destabilization: insights for human medicine from a veterinary perspective.
Medical hypotheses (6)4:575-581

166
Polet G, et al. 1999. The javan rhinos, Rhinoceros sondaicus annamiticus, of Cat
Tien National Park, Vietnam: Current status and management implications.
Pachyderm 27:34-48
Poole CM, Duckworth JW,. 2005. A documented 20th century record of Javan rhino
Rhinoceros sondaicus from Cambodia. Mammalia 69(3-4): 443-444
Pough FH, Heisser JB, McFarland WN,. 1989. Vertebrate Life – 3rd
edition.
MacMillan Publishing Company, New York p: 744
Prasad AS,. 2005. 3 clinical, endocrinological, and biochemical effects of zinc
deficiency. Clinics in endocrinology and metabolism (14)3:567-589
Purchase D,. 2007. Using spoor to determine the age and weight of subadult black
rhinoceroses (Diceros bicornis L). South African Journal of Wildlife Research
37(1): 96-100
Putro HR,. 1997. Heterogenitas habitat Badak Jawa (Rhinoceros sondaicus Desm.
1822) Di Taman Nasional Ujung Kulon. Media Konservasi Edisi Khusus, 1997
: Hal.17-40
Putz T, et al. 1999. Epidermal Growth Factor (EGF) receptor blocakde inhibits the
action of EGF, Insulin-like Growth Factor I, and a protein kinase A activator on
the mitogen-activated protein kinase pathway in prostate cancer cell lines.
Cancer Research 59: 227-233
Rahmat UM, Santosa Y, Kartono AP,. 2007. Analisis Tipologi Habitat Preferensial
bagi Badak Jawa (Rhinoceros sondaicus, Desmarest 1822) di Taman Nasional
Ujung Kulon. Institut Pertanian Bogor
Raven PH, Johnson GB,. 1989. Biology – Second Edition. Times Mirror/Mosby
College Publishing. St Louis Missouri. p: 981
Reynolds RJ, 1961. Javan Rhino in The Berlin zoo. International Zoo news 8(3): June
–July 1961
Rivera E, Benjamin B, Nielsen J, Shelle J, Zanella AJ,. 2002. Behavioural and
physiological responses of horses to initial training: the comparison between
pastured versus stalled horses. Applied animal behavioural science 78(2-4):
235-252
Rookmaaker K,. 2005. A Javan Rhinoceros, Rhinoceros sondaicus,in Bali in 1839.
Zool Garten N. F. 75 (2005) 2, 5. 129-131
Rookmaaker LC, 1982. The type locality of the Javan rhinoceros (Rhinoceros
sondaicus Desmarest, 1822). Sonderdrudre aus Zeitschrifl f. Siugetierkunde
Bd. 47 (1982), H. 6, S. 381-382

167
Sagar L, Sehgal R, Ojha S,. 2005. Evaluation of antimotility effect of Lantana camara
L. Var. acuelata constituents on neostigmine induced gastrointestinal transit in
mice. BMC complementary and alternative medicine 5(18): 1-6
Sajudin HR, Djaja B,. 1984. Monitoring populasi badak jawa (Rhinoceros sondaicus
Desmarest 1822) di Semenanjung Ujung Kulon. Laporan akhir penelitian
IUCN/WWF proyok nomor 1960. Fakultas Biologi UNAS
Sanborn ER,. 1908. Interesting Animal Surgery. Zoological Society Bullettin no 30.
1908
Santymire RM, Armstrong, DM,. 2009. Development of a Field-Friendly Technique
for Fecal Steroid Extraction and Storage Using the African Wild Dog (Lycaon
pictus). Zoo Biology (28): 1 -14
Sarma PK, Talukdar BK, Sarma K, Barua M,. 2009. Assessment of habitat change
and threats to the grater one-horned rhino (Rhinoceros unicornis) in Pobitora
Wildlife Sanctuary, Assam, using multi-temporal satellite data. Pachyderm no
46. 18-24
Scheltema JF,. 1912. Peeps at Many lands – Java. Adam & Charles Black. London
Schoeman JP, Goddard A, Herrtage ME,. 2007. Serum cortisol and thyroxine
concentrations as predictors of death in critically ill puppies with parvoral
diarrhea. Journal of the American veterinary medical association (231)10:
1534-1539
Schoenecker KA, Lyda RO, Kirkpatrick J,. 2004. Comparison of three fecal steroid
metabolites for pregnancy detection used with single sampling in bighorn sheep
(Ovis canadiensis). Journal of wildlife diseases 40(2): 273-281
Sharma OP, Singh A, Sharma S,. 2000. Levels of lantadene, bioactive pentacyclic
triterpenoids, in young and mature leaves of Lantana camara var acuelata.
Fitoterapia 71(5):487-491
Sharma S, Sharma O, Singh B, Bhat TK,. 2000. Biotransformation of lantadenes, the
pentacyclic triterpenoid hepatotoxins of lantana plant, in guinea pig. Toxicon
38(9): 1191-1202
Sharma GP, Ragubhanshi AS, Singh JS,. 2005. Lantana invasion: an overview. Weed
biology and management (5)4: 157-165
Sharma OP, 2006. An Overview of the Research on the Hepatotoxic Plant Lantana
Camara.
Sims JA, et al. 2007. Determination of bamboo-diet digestibility and fecal output by
giant pandas. Ursus 18(1): 38-45

168
Siswandi R, Agil M, Setijanto H,. 2005. Pola Aktifitas Harian Badak Sumatera
(Dicerorhinus sumatrensis) di Suaka Rhino Sumatera Taman Nasional Way
Kambas. Skripsi Institut Pertanian Bogor
Smit IPJ, Grant CC,. 2009. Managing surface-water in a large semi-arid savanna park:
effects on grazer distribution patterns. Journal for nature conservation
17(2):61-71
Soto-Gamboa M, Gonzalez S, Hayes LD, Ebensperger LA,. 2009. Validation of a
radioimmunoassay for measuring fecal cortisol metabolites in the
Hystricomorph rodent, Octodon degus. Journal of experimental zoology
311A:496-503
Sriyanto A, et al. 1995. A Current status of the Javan Rhino population in Ujung
Kulon National Park. Javan Rhino Colloquium
Strien NJ,. 1974. Dicerorhinus sumatrensis (Fisher). The Sumatran or two-horned
Asiatic rhinoceros: a study of literature. Medelebingen Landbouwwhogeschool,
Wageningen, Netherland
Strien NJ, 1982. Report on short mission to Ujung Kulon National Park, Java
Indonesia inconnectionwith the reported deaths of the five Javan rhinos.
Laporan kegiatan WWF.
Strien NJ,. 1986. The Sumatran rhinoceros – Dicerorhinus sumatrensis (Fischer,
1814) – in the Gunung Leuser National Park, Sumatra, Indonesia: its
distribution, ecology, and conservation. Verlag Paul Parey, Hamburg
Stuart G, Nelson S, Sakmann B, Hausser M,. 1997. Action potential initiation and
backpropagation in neurons of the mammalian CNS. Trends in neuroscience
20:125-131
Suprayogi A, Astuti DA, Satrija F, Suprianto. 2006. Physiological Status of Sheep
Reared Indoor System Under the Tropical Rain Forest Climatic Zone.
Supporting papers Proceedings of the 4th
ISTAP “Animal Production and
Sustainable Agriculture in the Tropic”. Faculty of Animal Science, Gajah Mada
University.
Suzuki A, et al. 2003. Induction of cell–cell detachment during glucose starvation
through F-actin conversion by SNARK, the fourth member of the AMP-
activated protein kinase catalytic subunit family. Biochemical and Biophysical
Research Communications Volume 311(1): 156-161
Suzuki A, et al. 2003. ARK5 suppresses the cell death induced by nutrient starvation
and death receptors via inhibition of caspase 8 activation, but not by
chemotherapeutic agents or UV irradiation. Oncogene 22: 6177–6182
Talukdar, B.K., Sectionov and L.B. Whetham. 2010. Proceeding of the Asian Rhino
Specialist Group meeting held at Kaziranga National Park, India 10-12 February
2010. Guwahati, Assam, India. Page 6-7

169
Tiuria R, et al. 2006. Identification of endoparasites from faeces of Javan rhino
(Rhinoceros sondaicus) in Ujung Kulon National Park, Indonesia. Proceedings
of AZWMP 2006 Chulalongkorn Uni. Fac. Of Vet. Sc., Bangkok, Thailand
Turner Jr JW, Tolson P, Hamad M,. 2002. Remote assessment of stress in white
rhinoceros (Ceratotherium simum) and black rhinoceros (Diceros bicornis) by
measurements of adrenal steroids in feces. Journal of zoo and wildlife medicine
33(3): 214-221
VanKeulen J, Young, BA. 1977. Evaluation of Acid Insoluble Ash as a Natural
Marker in Ruminant digestibility Studies. Journal of Animal Science vol 44 no:
2. 282-287
Vaux DL, Korsmeyer SJ,. 1999. Cell Death in Development. Cell vol 96: 245-254
Völlm J, et al. 2000. A case of extended exsudative dermatitis in an Indian rhino.
European Association of zoo and wildlife veterinarians (EAZWV) Paris,
France: 205-208
Walcott AS,. 1914. Java and Her Neighbours. A Traveler’s note in Java, Celebes, the
Molluccas, and Sumatra. GP Putnams and Sons. The knickerbockers press.
New York & London
Wasser SK, et al. 2000. A generalized fecal glucocorticoid assay for use in diverse
array of nondomestic mammalian and avian species. General and comparative
endocrinology 120:260-275
White AM, Swaisgood RR, Czekala N,. 2007. Ranging patterns in white rhinoceros,
Ceratotherium simum simum: implications for mating strategies. Animal
behaviour (74): 349-356
Wielckens K, et al. 1983. Stimulation of Poly(ADP-ribosyl)ation during Erlich
Ascites Tumor Cell “Starvation” and Suppression of Concomitant DNA
Fragmentation by Benzamide. The Journal of Biological Chemistry Vol 258(7):
4098-4104
Williams JW, et al. 2002. Rapid and widespread vegetation responses to past climate
change in the North Atlantic region. Geology vol 30; no. 11; p. 971–974
Wingfield JC, Romero LM,. 2001. Adrenocortical responses to stress and their
modulation. Dalam: McEwen BS, Goodman HM (eds), Handbook of
physiology section 7: The endocrine system, coping with the environments:
Neural and endocrine mechanism vol. IV Oxford University Press New York:
211-234
Woolf CJ, Mannion RJ,. 1999. Neuropathic pain: aetiology, symptoms, mechanism,
and management. Lancet 353:1959-64

170
Wright EM, Loo, DDF,. 2000. Coupling Between Na+, Sugar, and Water Transport
Across the Intestine. Annals of The New York Academy of Science Volume 915:
Epithelial Transport and Barrier Function: Pathomechanisms in Gastrointestinal
Disorders. 54-66
Wu P, Peters JM, Harris RA,. 2001. Adaptive Increase in Pyruvate Dehydrogenase
Kinase 4 During Starvation is mediated by Peroxisome Proliferator-activated
Receptor α. Biochemical and Biophysical Research Communication. 287(2):
391-396
WWF-IUCN. 1982. Mystery of Dead Javan rhinos. The Environmentalist vol. 2 no. 3
Yamada T, Horiuchi M, Dzau VJ,. 1996. Angiotensi type II receptor mediates
programmed cell death. Proc Acad Sci. USA. Cell Biology Vol 93: 156-160
Yakel JL, et al. 1997. Calcineurin regulation of synaptic function: from ion channels
to transmitter release and gene transcription. Trends in Pharmacological
Sciences. Volume 18, Issue 4: 124-134
Young KM, et al. 2004. Noninvasive monitoring of adrenocortical activity in
carnivores by fecal glucocorticoids analyses. General and comparative
endocrinology: 1-18
YMR [Yayasan Mitra Rhino]. 2004. Implementasi monitoring habitat badak jawa
dengan menekan laju invasi langkap (Arenga obtusifolia) di Taman Nasional
Ujung Kulon. PHKA-WWF-YMR
YMR-WWF. 2002. Studi Persaingan Ekologi antara Badak Jawa (Rhinoceros
sondaicus) dengan Banteng (Bos javanicus). Laporan Kegiatan kerjasama
BTNUK-WWF-YMR
Zeng H, Dvorak HF, Mukhopadhyay D,. 2001. Vascular Permeability Factor
(VPF)/Vascular Endothelial Growth Factor (VEGF) Receptor-1 Down-
modulates VPF/VEGF Receptor-2-mediated Endothelial Cell Proliferation, but
Not Migration, through Phosphatidylinositol 3-Kinase-dependent Pathways. The
Journal of Biological Chemistry 270(29): 26969-26979
Ziegler TE, Wittwer DJ,. 2005. Fecal Steroid Research In the Field and In Laboratory:
Improved methods for Storage, Transport, Processing, and Analysis. American
Journal of Primatology 67: 159-174
Zmuda JM, et al. 1997. Longitudinal relation between endogenous testosterone and
cardiovascular disease risk factors in middle-aged men. American journal of
epidemiology 146(8): 609-617

LAMPIRAN 1
IZIN KOMISI ETIK

172

173

LAMPIRAN 2
DATA NUTRISI BADAK JAWA

176

177
ASUPAN DAN NUTRISI BADAK MSADUB5212 (BADAK 12)
Bulan
Segar Kering Air Protein Lemak Energi
Oktober 3.970,00 1.270,00 180,02 102,03 31,62 4.660,70
November 1.837,50 412,09 44,54 46,67 9,82 1.571,03
Desember 3.020,00 652,21 58,93 69,23 18,12 2.482,70
Januari 1.786,30 392,99 41,54 52,41 11,35 1.466,09
Februari 2.223,33 497,69 51,62 62,56 12,25 1.984,71
Maret 1.538,80 370,56 34,67 43,32 10,40 1.484,64
April 3.700,00 900,00 189,90 101,25 25,85 3.014,28
Rata-rata 2.582,28 642,22 85,89 68,21 17,06 2.380,60
StDev 980,87 334,27 68,17 24,50 8,59 1.159,17
Asupan / berat 0,10% 0,03% 0,01% 0,00% 0,00% 0,12%
(asumsi 1 ton berat)
Estimasi asupan
Lantana 129,11 gram segar per hari
36,15 gram kering per hari
0,004% gram kering per berat badan
Konsumsi (gram) Nutrisi
ASUPAN DAN NUTRISI BADAK MADUB5513 (BADAK 13)
Bulan
Segar Kering Air Protein Lemak Energi
Oktober 2.101,17 768,15 86,38 87,64 22,51 2.852,88
November 1.307,05 287,55 29,57 34,96 10,16 1.093,46
Desember 4.792,50 1.054,35 124,23 115,97 30,66 3.786,15
Januari 7.560,76 1.424,95 136,03 137,67 37,98 5.320,58
Februari 3.633,46 799,36 92,04 83,57 21,84 2.835,35
Maret 3.456,41 760,41 64,02 80,02 19,42 2.722,34
April 2.788,36 613,44 58,01 76,28 21,54 2.264,95
Rata-rata 3.662,82 815,46 84,33 88,02 23,44 2.982,24
StDev 2.052,10 354,40 37,49 32,36 8,79 1.310,34
Asupan / berat 0,21% 0,04% 0,00% 0,00% 0,00% 0,13%
(asumsi 1 ton berat)
Estimasi asupan
Lantana 1.318,61 gram segar per hari
369,21 gram kering per hari
0,037% gram kering per berat badan
Konsumsi (gram) Nutrisi

178
ASUPAN DAN NUTRISI BADAK MSADUB2018 (BADAK 18)
Bulan
Segar Kering Air Protein Lemak Energi
Oktober 4.360,00 1.528,00 153,54 164,66 36,43 5.603,39
November 10.210,49 2.246,31 234,70 245,31 59,84 8.107,89
Desember 15.249,09 1.078,48 103,97 113,96 28,53 4.058,33
Januari 7.265,23 1.058,62 121,16 118,87 27,20 3.428,20
Februari 4.520,89 710,10 78,41 80,38 18,45 2.304,37
Maret 3.383,80 744,44 117,22 74,07 18,13 2.526,32
April 755,18 166,14 14,73 16,50 5,10 622,46
Rata-rata 6.534,95 1.076,01 117,68 116,25 27,67 3.807,28
StDev 4.866,58 664,07 67,48 73,06 17,31 2.448,49
Asupan / berat 0,49% 0,07% 0,01% 0,01% 0,00% 0,24%
(asumsi 1 ton berat)
Estimasi asupan
Lantana 416,93 gram segar per hari
116,74 gram kering per hari
0,012% gram per kg badan
Konsumsi (gram) Nutrisi
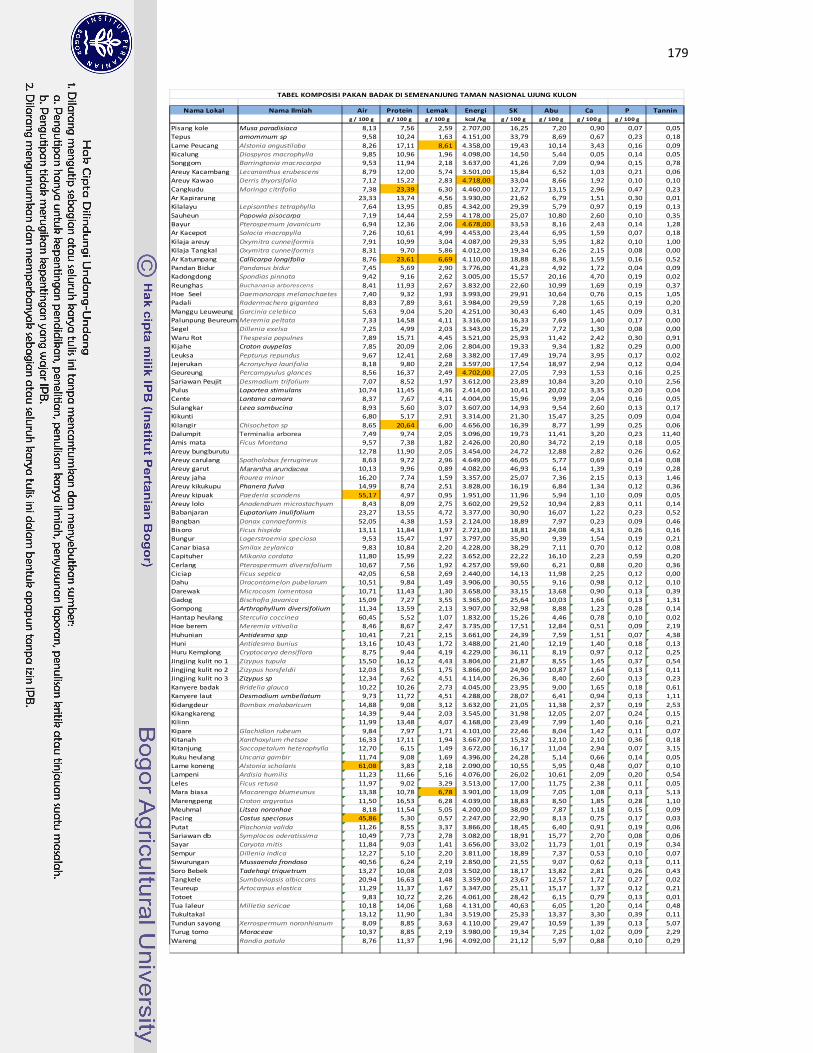
179
Nama Lokal Nama Ilmiah Air Protein Lemak Energi SK Abu Ca P Tannin
g / 100 g g / 100 g g / 100 g kcal /kg g / 100 g g / 100 g g / 100 g g / 100 g
Pisang kole Musa paradisiaca 8,13 7,56 2,59 2.707,00 16,25 7,20 0,90 0,07 0,05
Tepus amommum sp 9,58 10,24 1,63 4.151,00 33,79 8,69 0,67 0,23 0,18
Lame Peucang Alstonia angustiloba 8,26 17,11 8,61 4.358,00 19,43 10,14 3,43 0,16 0,09
Kicalung Diospyros macrophylla 9,85 10,96 1,96 4.098,00 14,50 5,44 0,05 0,14 0,05
Songgom Barringtonia macrocarpa 9,53 11,94 2,18 3.637,00 41,26 7,09 0,94 0,15 0,78
Areuy Kacambang Lecananthus erubescens 8,79 12,00 5,74 3.501,00 15,84 6,52 1,03 0,21 0,06
Areuy Kawao Derris thyorsifolia 7,12 15,22 2,83 4.718,00 33,04 8,66 1,92 0,10 0,10
Cangkudu Moringa citrifolia 7,38 23,39 6,30 4.460,00 12,77 13,15 2,96 0,47 0,23
Ar Kapirarung 23,33 13,74 4,56 3.930,00 21,62 6,79 1,51 0,30 0,01
Kilalayu Lepisanthes tetraphylla 7,64 13,95 0,85 4.342,00 29,39 5,79 0,97 0,19 0,13
Sauheun Popowia pisocarpa 7,19 14,44 2,59 4.178,00 25,07 10,80 2,60 0,10 0,35
Bayur Pterospemum javanicum 6,94 12,36 2,06 4.678,00 33,53 8,16 2,43 0,14 1,28
Ar Kacepot Salacia macropylla 7,26 10,61 4,99 4.453,00 23,44 6,95 1,59 0,07 0,18
Kilaja areuy Oxymitra cunneiformis 7,91 10,99 3,04 4.087,00 29,33 5,95 1,82 0,10 1,00
Kilaja Tangkal Oxymitra cunneiformis 8,31 9,70 5,86 4.012,00 19,34 6,26 2,15 0,08 0,00
Ar Katumpang Callicarpa longifolia 8,76 23,61 6,69 4.110,00 18,88 8,36 1,59 0,16 0,52
Pandan Bidur Pandanus bidur 7,45 5,69 2,90 3.776,00 41,23 4,92 1,72 0,04 0,09
Kadongdong Spondias pinnata 9,42 9,16 2,62 3.005,00 15,57 20,16 4,70 0,19 0,02
Reunghas Buchanania arborescens 8,41 11,93 2,67 3.832,00 22,60 10,99 1,69 0,19 0,37
Hoe Seel Daemonorops melanochaetes 7,40 9,32 1,93 3.993,00 29,91 10,64 0,76 0,15 1,05
Padali Radermachera gigantea 8,83 7,89 3,61 3.984,00 29,59 7,28 1,65 0,19 0,20
Manggu Leuweung Garcinia celebica 5,63 9,04 5,20 4.251,00 30,43 6,40 1,45 0,09 0,31
Palunpung Beureum Meremia peltata 7,33 14,58 4,11 3.316,00 16,33 7,69 1,40 0,17 0,00
Segel Dillenia exelsa 7,25 4,99 2,03 3.343,00 15,29 7,72 1,30 0,08 0,00
Waru Rot Thespesia populnes 7,89 15,71 4,45 3.521,00 25,93 11,42 2,42 0,30 0,91
Kijahe Croton auypelas 7,85 20,09 2,06 2.804,00 19,33 9,34 1,82 0,29 0,00
Leuksa Pepturus repundus 9,67 12,41 2,68 3.382,00 17,49 19,74 3,95 0,17 0,02
Jejerukan Acronychya laurifolia 8,18 9,80 2,28 3.597,00 17,54 18,97 2,94 0,12 0,04
Geureung Percampyulus glances 8,56 16,37 2,49 4.702,00 27,05 7,93 1,53 0,16 0,25
Sariawan Peujit Desmodium trifolium 7,07 8,52 1,97 3.612,00 23,89 10,84 3,20 0,10 2,56
Pulus Laportea stimulans 10,74 11,45 4,36 2.414,00 10,41 20,02 3,35 0,20 0,04
Cente Lantana camara 8,37 7,67 4,11 4.004,00 15,96 9,99 2,04 0,16 0,05
Sulangkar Leea sambucina 8,93 5,60 3,07 3.607,00 14,93 9,54 2,60 0,13 0,17
Kikunti 6,80 5,17 2,91 3.314,00 21,30 15,47 3,25 0,09 0,04
Kilangir Chisocheton sp 8,65 20,64 6,00 4.656,00 16,39 8,77 1,99 0,25 0,06
Dalumpit Terminalia arborea 7,49 9,74 2,05 3.096,00 19,73 11,41 3,20 0,23 11,40
Amis mata Ficus Montana 9,57 7,38 1,82 2.426,00 20,80 34,72 2,19 0,18 0,05
Areuy bungburutu 12,78 11,90 2,05 3.454,00 24,72 12,88 2,82 0,26 0,62
Areuy carulang Spatholobus ferrugineus 8,63 9,72 2,96 4.649,00 46,05 5,77 0,69 0,14 0,08
Areuy garut Marantha arundacea 10,13 9,96 0,89 4.082,00 46,93 6,14 1,39 0,19 0,28
Areuy jaha Rourea minor 16,20 7,74 1,59 3.357,00 25,07 7,36 2,15 0,13 1,46
Areuy kikukupu Phanera fulva 14,99 8,74 2,51 3.828,00 16,19 6,84 1,34 0,12 0,36
Areuy kipuak Paederia scandens 55,17 4,97 0,95 1.951,00 11,96 5,94 1,10 0,09 0,05
Areuy lolo Anadendrum microstachyum 8,43 8,09 2,75 3.602,00 29,52 10,94 2,83 0,11 0,14
Babanjaran Eupatorium inulifolium 23,27 13,55 4,72 3.377,00 30,90 16,07 1,22 0,23 0,52
Bangban Donax cannaeformis 52,05 4,38 1,53 2.124,00 18,89 7,97 0,23 0,09 0,46
Bisoro Ficus hispida 13,11 11,84 1,97 2.721,00 18,81 24,08 4,31 0,26 0,16
Bungur Lagerstroemia speciosa 9,53 15,47 1,97 3.797,00 35,90 9,39 1,54 0,19 0,21
Canar biasa Smilax zeylanica 9,83 10,84 2,20 4.228,00 38,29 7,11 0,70 0,12 0,08
Capituher Mikania cordata 11,80 15,99 2,22 3.652,00 22,22 16,10 2,23 0,59 0,20
Cerlang Pterospermum diversifolium 10,67 7,56 1,92 4.257,00 59,60 6,21 0,88 0,20 0,36
Ciciap Ficus septica 42,05 6,58 2,69 2.440,00 14,13 11,98 2,25 0,12 0,00
Dahu Dracontomelon pubelarum 10,51 9,84 1,49 3.906,00 30,55 9,16 0,98 0,12 0,10
Darewak Microcosm lomentosa 10,71 11,43 1,30 3.658,00 33,15 13,68 0,90 0,13 0,39
Gadog Bischofia javanica 15,09 7,27 3,55 3.365,00 25,64 10,03 1,66 0,13 1,31
Gompong Arthrophyllum diversifolium 11,34 13,59 2,13 3.907,00 32,98 8,88 1,23 0,28 0,14
Hantap heulang Sterculia coccinea 60,45 5,52 1,07 1.832,00 15,26 4,46 0,78 0,10 0,02
Hoe berem Meremia vitivolia 8,46 8,67 2,47 3.735,00 17,51 12,84 0,51 0,09 2,19
Huhunian Antidesma spp 10,41 7,21 2,15 3.661,00 24,39 7,59 1,51 0,07 4,38
Huni Antidesma bunius 13,16 10,43 1,72 3.488,00 21,40 12,19 1,40 0,18 0,13
Huru Kemplong Cryptocarya densiflora 8,75 9,44 4,19 4.229,00 36,11 8,19 0,97 0,12 0,25
Jingjing kulit no 1 Zizypus tupula 15,50 16,12 4,43 3.804,00 21,87 8,55 1,45 0,37 0,54
Jingjing kulit no 2 Zizypus horsfeldii 12,03 8,55 1,75 3.866,00 24,90 10,87 1,64 0,13 0,11
Jingjing kulit no 3 Zizypus sp 12,34 7,62 4,51 4.114,00 26,36 8,40 2,60 0,13 0,23
Kanyere badak Bridelia glauca 10,22 10,26 2,73 4.045,00 23,95 9,00 1,65 0,18 0,61
Kanyere laut Desmodium umbellatum 9,73 11,72 4,51 4.288,00 28,07 6,41 0,94 0,13 1,11
Kidangdeur Bombax malabaricum 14,88 9,08 3,12 3.632,00 21,05 11,38 2,37 0,19 2,53
Kikangkareng 14,39 9,44 2,03 3.545,00 31,98 12,05 2,07 0,24 0,15
Kilinn 11,99 13,48 4,07 4.168,00 23,49 7,99 1,40 0,16 0,21
Kipare Glachidion rubeum 9,84 7,97 1,71 4.101,00 22,46 8,04 1,42 0,11 0,07
Kitanah Xanthoxylum rhetsae 16,33 17,11 1,94 3.667,00 15,32 12,10 2,10 0,36 0,18
Kitanjung Saccopetalum heterophylla 12,70 6,15 1,49 3.672,00 16,17 11,04 2,94 0,07 3,15
Kuku heulang Uncaria gambir 11,74 9,08 1,69 4.396,00 24,28 5,14 0,66 0,14 0,05
Lame koneng Alstonia scholaris 61,08 3,83 2,18 2.090,00 10,55 5,95 0,48 0,07 0,10
Lampeni Ardisia humilis 11,23 11,66 5,16 4.076,00 26,02 10,61 2,09 0,20 0,54
Leles Ficus retusa 11,97 9,02 3,29 3.513,00 17,00 11,75 2,38 0,11 0,05
Mara biasa Macarenga blumeunus 13,38 10,78 6,78 3.901,00 13,09 7,05 1,08 0,13 5,13
Marengpeng Croton argyratus 11,50 16,53 6,28 4.039,00 18,83 8,50 1,85 0,28 1,10
Meuhmal Litsea noronhae 8,18 11,54 5,05 4.200,00 38,09 7,87 1,18 0,15 0,09
Pacing Costus speciosus 45,86 5,30 0,57 2.247,00 22,90 8,13 0,75 0,17 0,03
Putat Plachonia valida 11,26 8,55 3,37 3.866,00 18,45 6,40 0,91 0,19 0,06
Sariawan db Symplocos oderatissima 10,49 7,73 2,78 3.082,00 18,91 15,77 2,70 0,08 0,06
Sayar Caryota mitis 11,84 9,03 1,41 3.656,00 33,02 11,73 1,01 0,19 0,34
Sempur Dillenia indica 12,27 5,10 2,20 3.811,00 18,89 7,37 0,53 0,10 0,07
Siwurungan Mussaenda frondosa 40,56 6,24 2,19 2.850,00 21,55 9,07 0,62 0,13 0,11
Soro Bebek Tadehagi triquetrum 13,27 10,08 2,03 3.502,00 18,17 13,82 2,81 0,26 0,43
Tangkele Sumbaviopsis albiccans 20,94 16,63 1,48 3.359,00 23,67 12,57 1,72 0,27 0,02
Teureup Artocarpus elastica 11,29 11,37 1,67 3.347,00 25,11 15,17 1,37 0,12 0,21
Totoet 9,83 10,72 2,26 4.061,00 28,42 6,15 0,79 0,13 0,01
Tua laleur Milletia sericae 10,18 14,06 1,68 4.131,00 40,63 6,05 1,20 0,14 0,48
Tukultakal 13,12 11,90 1,34 3.519,00 25,33 13,37 3,30 0,39 0,11
Tundun sayong Xerrospermum noronhianum 8,09 8,85 3,63 4.110,00 29,47 10,59 1,39 0,13 5,07
Turug tomo Moraceae 10,37 8,85 2,19 3.980,00 19,34 7,25 1,02 0,09 2,29
Wareng Randia patula 8,76 11,37 1,96 4.092,00 21,12 5,97 0,88 0,10 0,29
TABEL KOMPOSISI PAKAN BADAK DI SEMENANJUNG TAMAN NASIONAL UJUNG KULON

LAMPIRAN 3
DATA HEWAN MODEL

182

183
Jumlah pakan yang diberikan secara rutin kepada kedua ekor kuda model (Garuda dan
Elang) selama periode kontrol.
Kuda Model Air (L/hari) Bran (kg/hari) Pellet (kg/hari) Rumput (kg/hari)
Garuda 8 0.875 0.38 17.375
Elang 8 0.875 0.375 17.25
184
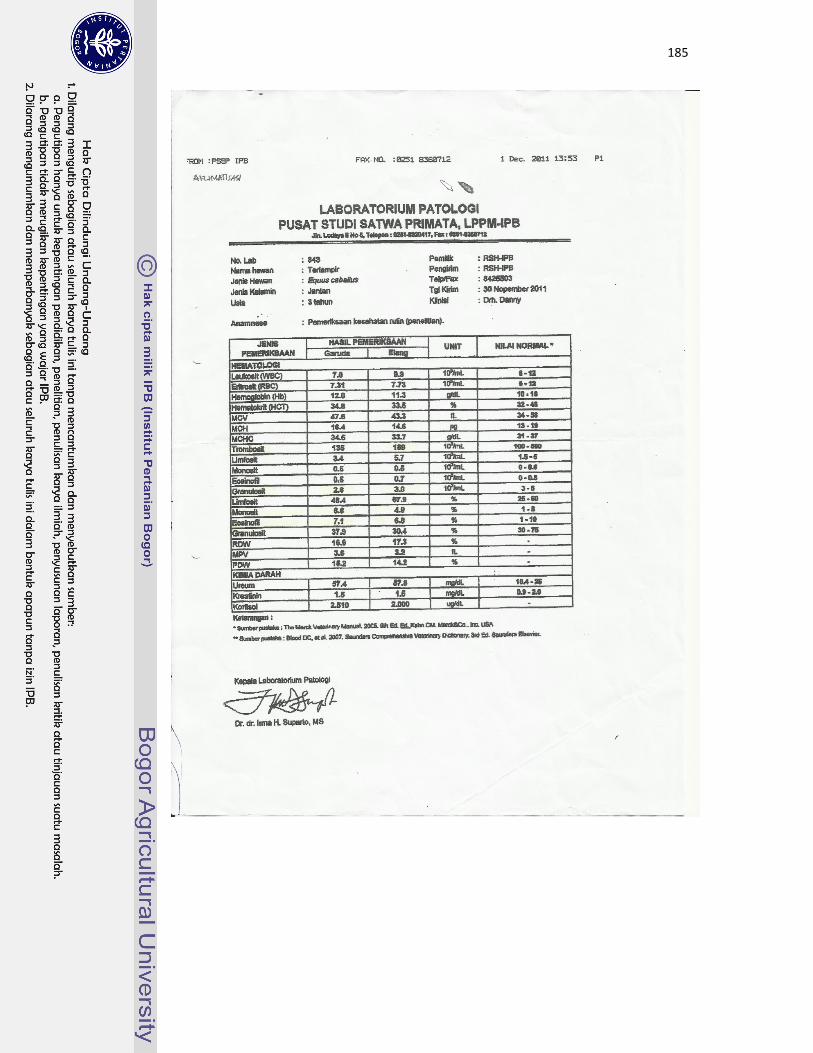
185
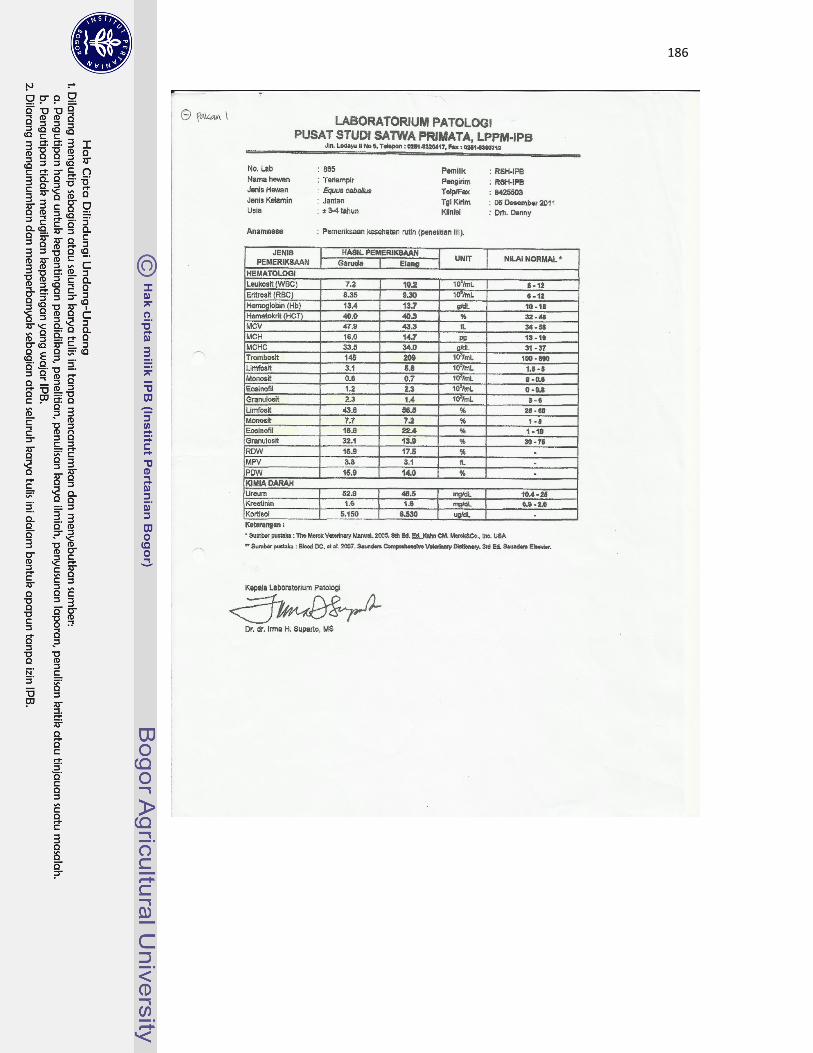
186
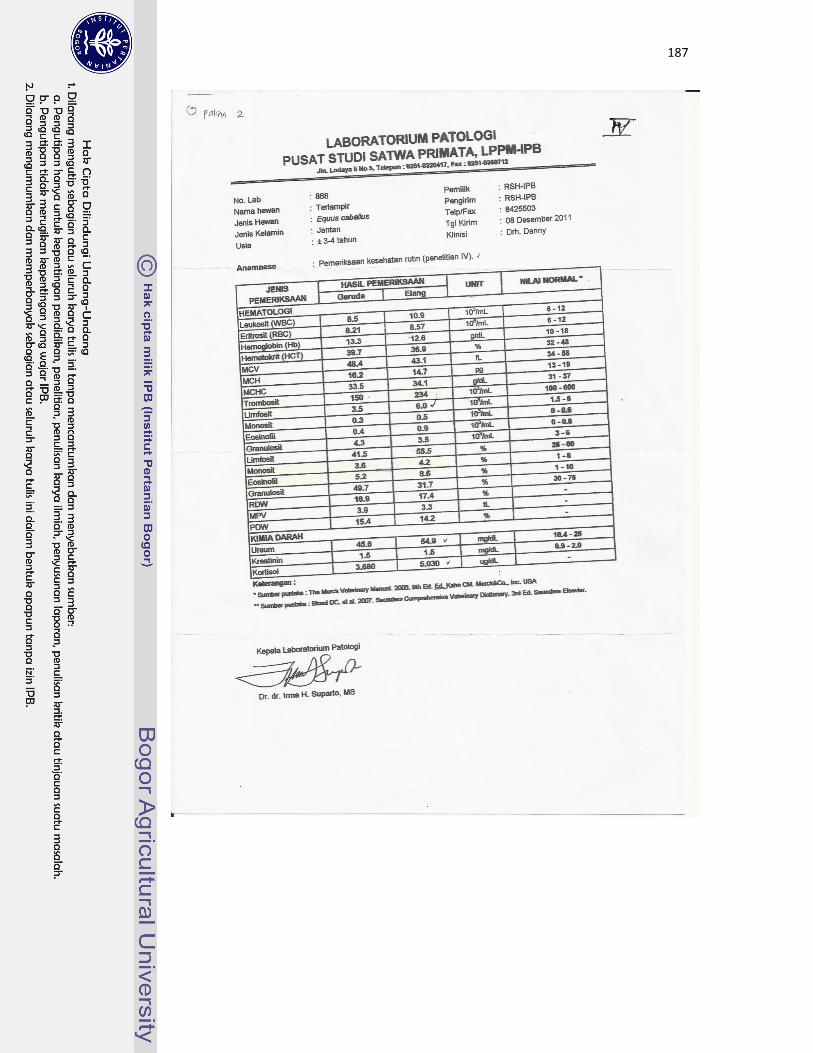
187
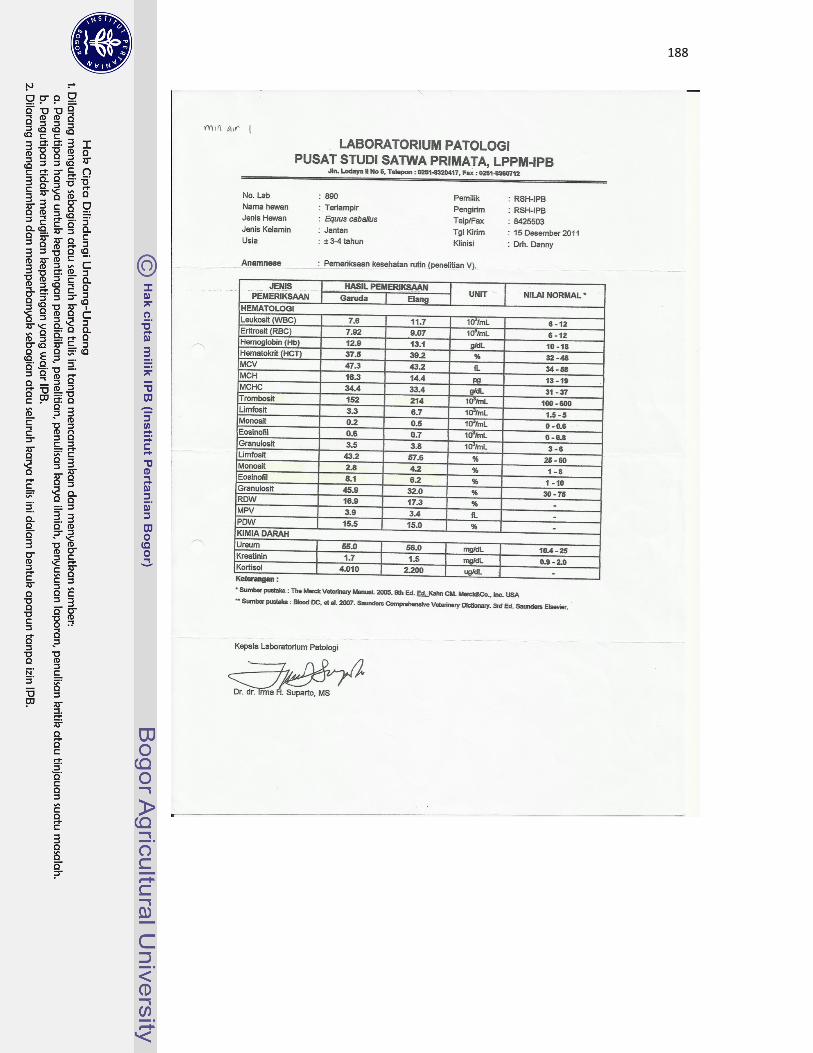
188
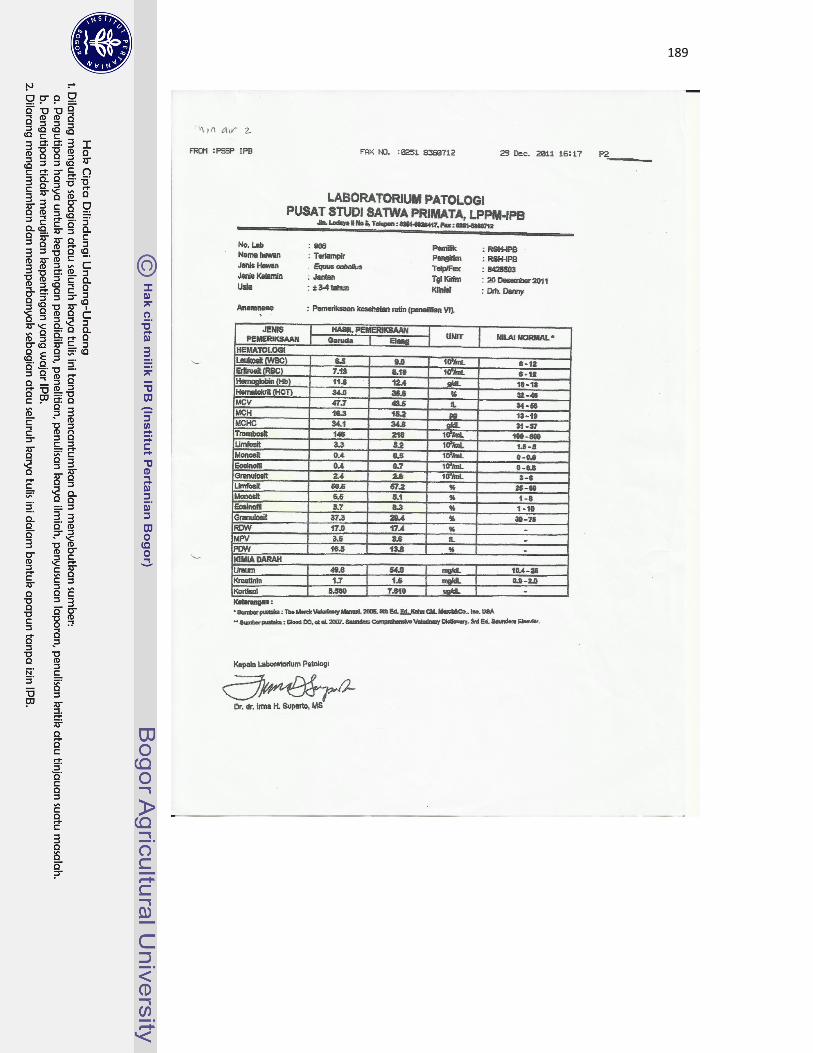
189

LAMPIRAN 4
DATA PARALELISME KIT HORMON

192
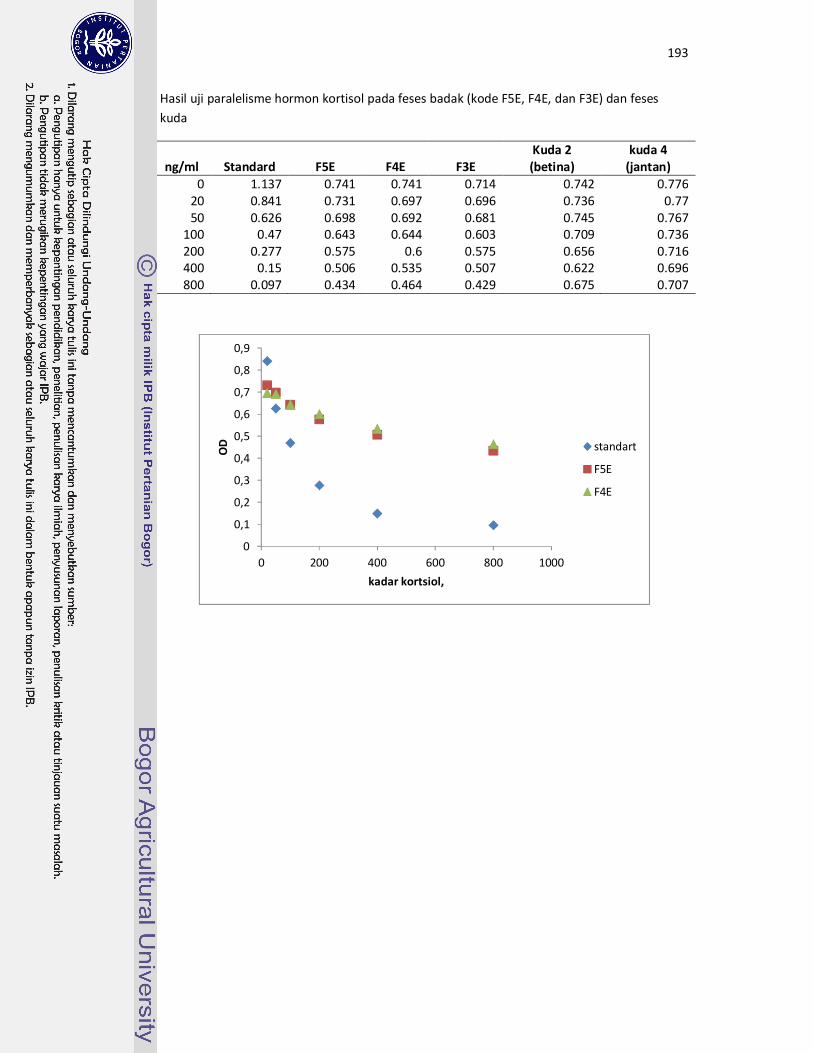
193
Hasil uji paralelisme hormon kortisol pada feses badak (kode F5E, F4E, dan F3E) dan feses
kuda
ng/ml Standard F5E F4E F3E
Kuda 2
(betina)
kuda 4
(jantan)
0 1.137 0.741 0.741 0.714 0.742 0.776
20 0.841 0.731 0.697 0.696 0.736 0.77
50 0.626 0.698 0.692 0.681 0.745 0.767
100 0.47 0.643 0.644 0.603 0.709 0.736
200 0.277 0.575 0.6 0.575 0.656 0.716
400 0.15 0.506 0.535 0.507 0.622 0.696
800 0.097 0.434 0.464 0.429 0.675 0.707
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
0 200 400 600 800 1000
OD
kadar kortsiol,
standart
F5E
F4E
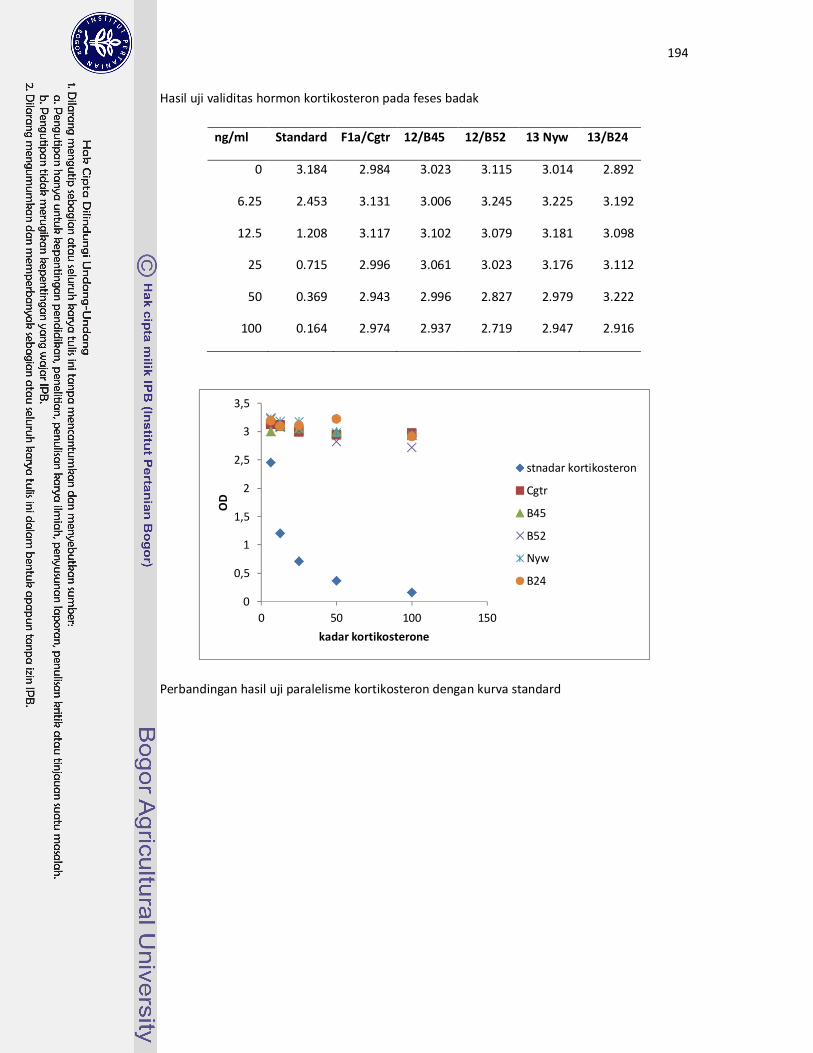
194
Hasil uji validitas hormon kortikosteron pada feses badak
ng/ml Standard F1a/Cgtr 12/B45 12/B52 13 Nyw 13/B24
0 3.184 2.984 3.023 3.115 3.014 2.892
6.25 2.453 3.131 3.006 3.245 3.225 3.192
12.5 1.208 3.117 3.102 3.079 3.181 3.098
25 0.715 2.996 3.061 3.023 3.176 3.112
50 0.369 2.943 2.996 2.827 2.979 3.222
100 0.164 2.974 2.937 2.719 2.947 2.916
Perbandingan hasil uji paralelisme kortikosteron dengan kurva standard
0
0,5
1
1,5
2
2,5
3
3,5
0 50 100 150
OD
kadar kortikosterone
stnadar kortikosteron
Cgtr
B45
B52
Nyw
B24
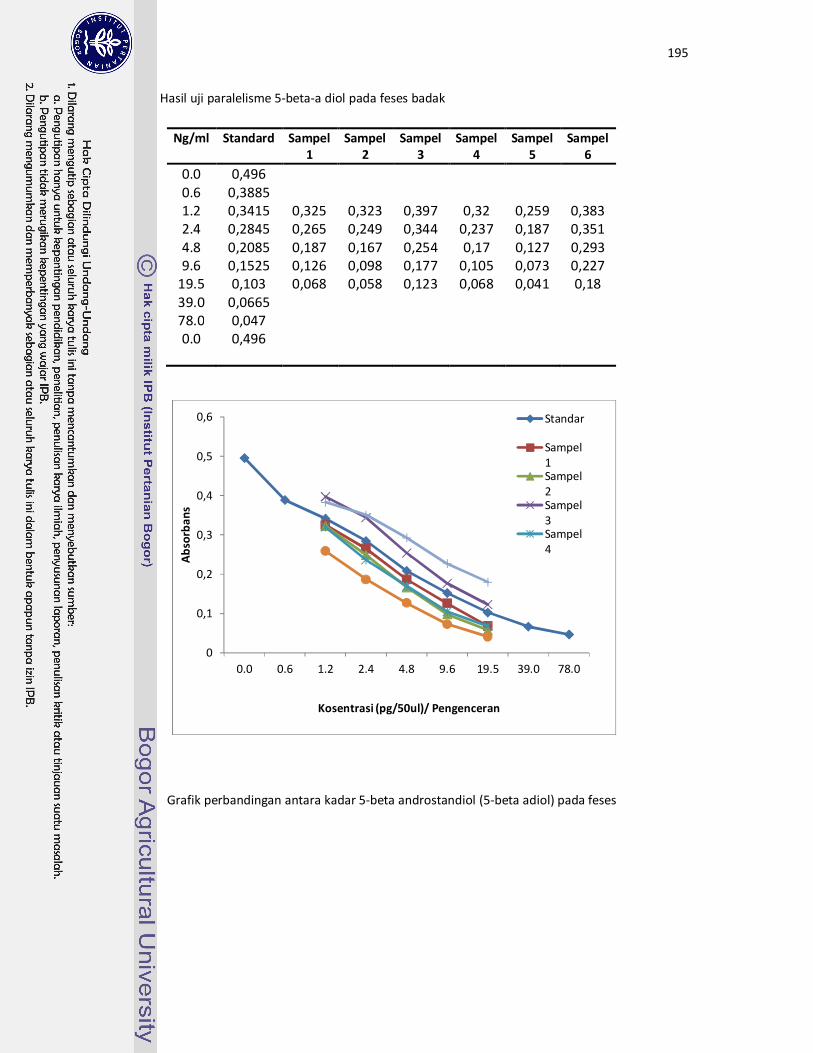
195
Hasil uji paralelisme 5-beta-a diol pada feses badak
Ng/ml Standard Sampel
1
Sampel
2
Sampel
3
Sampel
4
Sampel
5
Sampel
6
0.0 0,496
0.6 0,3885
1.2 0,3415 0,325 0,323 0,397 0,32 0,259 0,383
2.4 0,2845 0,265 0,249 0,344 0,237 0,187 0,351
4.8 0,2085 0,187 0,167 0,254 0,17 0,127 0,293
9.6 0,1525 0,126 0,098 0,177 0,105 0,073 0,227
19.5 0,103 0,068 0,058 0,123 0,068 0,041 0,18
39.0 0,0665
78.0 0,047
0.0 0,496
Grafik perbandingan antara kadar 5-beta androstandiol (5-beta adiol) pada feses
0
0,1
0,2
0,3
0,4
0,5
0,6
0.0 0.6 1.2 2.4 4.8 9.6 19.5 39.0 78.0
Ab
sorb
an
s
Kosentrasi (pg/50ul)/ Pengenceran
Standar
Sampel
1Sampel
2Sampel
3Sampel
4
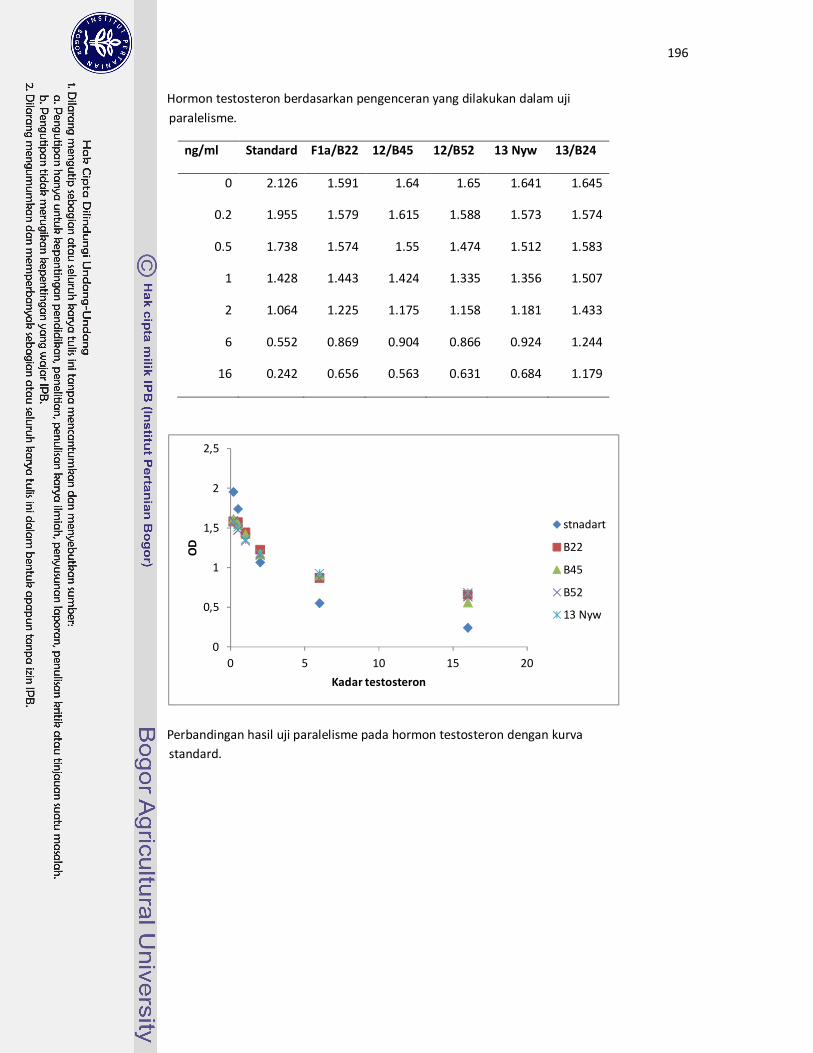
196
Hormon testosteron berdasarkan pengenceran yang dilakukan dalam uji
paralelisme.
ng/ml Standard F1a/B22 12/B45 12/B52 13 Nyw 13/B24
0 2.126 1.591 1.64 1.65 1.641 1.645
0.2 1.955 1.579 1.615 1.588 1.573 1.574
0.5 1.738 1.574 1.55 1.474 1.512 1.583
1 1.428 1.443 1.424 1.335 1.356 1.507
2 1.064 1.225 1.175 1.158 1.181 1.433
6 0.552 0.869 0.904 0.866 0.924 1.244
16 0.242 0.656 0.563 0.631 0.684 1.179
Perbandingan hasil uji paralelisme pada hormon testosteron dengan kurva
standard.
0
0,5
1
1,5
2
2,5
0 5 10 15 20
OD
Kadar testosteron
stnadart
B22
B45
B52
13 Nyw

197
Hasil uji paralelisme
Uji paralelisme dilakukan menggunakan semua kit yang telah dipilih berdasarkan
indikator hormon cekaman. Uji paralelisme kit hormon kortisol, kortikosteron, dan antibodi
terhadap salah satu metabolit glukokortikoid yaitu 5-beta adiol disajikan dalam Lampiran 4.
Dari uji validitas menggunakan uji paralelisme dari keempat kit hormon di atas dapat
dilihat bahwa hormon kortisol pada feses badak masih menunjukkan kemungkinan paralel
dengan kurva standard (walaupun masih terjadi perpotongan antara kurva standard dengan
sampel). Penelitian yang dilakukan oleh Turner et al. (2002) menunjukkan bahwa kadar
kortisol dapat dikuantifikasi dari feses badak putih (Ceratotherium simum). Hal ini
menunjukkan bahwa hormon kortisol memiliki kemungkinan yang cukup besar untuk
dideteksi dan diukur dalam feses badak, namun pengukuran ataupun kuantifikasi dari kadar
hormon ini masih belum dapat dilakukan dengan validitas yang dapat dipertanggung jawabkan
dalam penelitian ini.
Penggunaan kit kortikosteron pada feses tidak menunjukkan hasil yang paralel; hal ini
ditunjukkan dengan adanya perpotongan antara kurva standard dengan sampel yang
kemudian diikuti oleh slope yang samasekali berbeda antara keduanya. Dengan hasil seperti
ini kit kortikosteron tidak dapat digunakan untuk mengukur ataupun melakukan kuantifikasi
kadar hormon cekaman pada sampel feses badak Jawa, walaupun Turner et al. (2002) masih
dapat menggunakan kortikosteron sebagai hormon indikator cekaman pada badak putih. Hal
ini dapat terjadi karena ketidak cocokan kit kortikosteron yang dirancang untuk penggunaan
serum dengan sampel feses.

198
Penggunaan metabolit glukokortikoid yaitu 5-beta-adiol sebagai indikator keberadaan
hormon cekaman menunjukkan hasil yang paralel antara kurva standard dengan kurva sampel
dimana relatif tidak ada perpotongan antara kedua kurva ini dan slope dari kedua kurva ini
menunjukkan kemiringan yang sama sehingga dapat dipastikan bahwa kedua kurva ini berada
dalam kondisi paralel dan tidak saling memotong. Uji paralelisme menggunakan 5-beta adiol
ini menunjukkan 50% binding pada konsentrasi 3,04 pg dan pengenceran 1:80. Hasil uji
paralelisme juga menunjukkan bahwa koefisien variasi intra-assay adalah 5.4% untuk sampel
dengan konsentrasi hormon rendah dan 4.4% untuk sampel konsentrasi hormon tinggi.
Demikian pula dengan koefisien variasi inter-assay yang menunjukkan angka 11.2% untuk
sampel dengan konsentrasi hormon rendah dan 6.8% untuk sampel dengan konsentrasi
hormon yang tinggi. Berdasarkan hasil dari uji paralelisme ini yang didukung dengan
koefiesien variasi intra-assay dan inter-assay yang berada di bawah 15%, maka dapat
disimpulkan bahwa, 5-beta-adiol memiliki potensi untuk digunakan dalam kajian profil
glukokortikoid dari sampel feses badak jawa. Penggunaan secara umum/luas dapat dilakukan
setelah kit ini menjalani validasi biologis dan kimiawi untuk memastikan ketepatan kit tersebut
dalam mendeteksi hormon asal feses.

MODEL PENGELOLAAN POPULASI BADAK
JAWA (Rhinoceros sondaicus) BERDASARKAN
ANALISIS NUTRISI DAN TINGKAT CEKAMAN
SEBAGAI PARAMETER KESEHATAN
ADHI RACHMAT SUDRAJAT HARIYADI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Ilmu Biomedis hewan
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
2012

Penguji Luar Komisi
Penguji Ujian Tertutup : 1. Dr. drh. Setyo Widodo
(Staff pengajar Fakultas Kedokteran Hewan IPB)
2. Prof. Dr. Ir. Sambas Basuni
(Guru Besar Fakultas Kehutanan IPB)
Pelaksanaan : 27 Juli 2012
Penguji Ujian Terbuka : 1. Dr. Iman Santoso
(Kepala Badan Litbang Kementerian Kehutanan RI)
2. Prof. Dr. Ir. Sri Supraptini Mansjoer
(Guru Besar Program Studi Satwa Primata IPB)
Pelaksanaan : 20 September 2012

Judul Penelitian: Model Pengelolaan Populasi Badak Jawa (Rhinoceros sondaicus) berdasarkan
Analisis Nutrisi dan Tingkat Cekaman sebagai Parameter Kesehatan
Nama: Adhi Rachmat Sudrajat Hariyadi
NRP: B361080011
Program Studi: Ilmu Biomedis Hewan
Menyetujui:
Komisi Pembimbing
Prof. Drh. Dondin Sajuthi, MST, PhD
Ketua
Dr. Drh. Hera Maheshwari, MSc Prof. Dr. Ir. Hadi S Alikodra MS
Anggota Anggota
Prof. Dr. Ir. Dewi Apri Astuti, MS
Anggota
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
drh.Agus Setiyono, MS, PhD, ApVet Dr. Ir. Dahrul Syah, M.Sc.Agr
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………

Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………

Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………
Tanggal Ujian : 20 September 2012 Tanggal Lulus : ………………




















