KLONING GEN RHOPTRY 1 - Universitas Udayana · 2017-06-06 · Terapi toksoplasmosis dengan obat...
26
1 KLONING GEN PENYANDI PROTEIN RHOPTRY 1 (ROP1) TAKIZOIT Toxoplasma gondii ISOLAT LOKAL I Wayan Surudarma ABSRAK INTISARI Protein Rhopry 1 (ROP1) takizoit Toxoplasma gondii merupakan salah satu protein yang berperan penting pada proses invasi parasit ini ke dalam sel hospes. Protein tersebut diperkirakan sebagai molekul penetration-enhanching factor (PEF). Kloning gen penyandi ROP1 takizoit Toxoplasma gondii isolat RH untuk vaksinasi DNA pada mencit telah diteliti. Imunisasi DNA tersebut dapat meningkatkan respon imun seluler dan humoral. Kloning gen penyandi ROP1 takizoit Toxoplasma gondii isolat lokal belum pernah dilakukan. Tujuan penelitian ini ialah untuk memperoleh klon yang membawa gen penyandi ROP1 takizoit isolat lokal melalui teknik DNA rekombinan. Takizoit Toxoplasma gondii dikultivasi secara in vivo pada mencit strain Balb/C. Isolasi DNA dilakukan setelah beberapa kali pasase untuk diperoleh jumlah takizoit yang cukup (minimal 1x10 8 takizoit/ml) dan selanjutnya DNA diamplifikasi menggunakan PuRe Taq RTG-PCR Beads (Amersham Bioscience) dengan primer spesifik (Cybergene AB) dan produknya diligasi pada pGEM-T ® Easy (Promega). Plasmid rekombinan ditransformasi ke dalam Escherichia coli XL-1 Blue dengan teknik heat shock dan transforman ditanam pada plate agar LB yang mengandung ampisilin, X-gal, dan IPTG. Koloni putih yang menunjukkan koloni rekombinan ditumbuhkan dalam media LB yang telah ditambahkan ampisilin pada suhu 37 o C selama semalam. Plasmid DNA rekombinan diisolasi dengan metode lisis alkali dan dielektroforesis pada gel agarose 1% (SeaKem). Analisis plasmid rekombinan dilakukan dengan cara digesti menggunakan endonuklease restriksi EcoRI dan selanjutnya disekuensing untuk mengetahui urutan basanya (ABI 3130 Genetic Analyzer). Hasil penelitian menunjukkan bahwa kloning gen penyandi ROP1 Toxoplasma gondii isolat lokal menghasilkan klon yang membawa DNA insert dengan ukuran 1441 bp. DNA insert tersebut mempunyai homologi sebesar 99% dengan gen penyandi ROP1 Toxoplasma gondii isolat RH. Kata kunci: ROP1, Toxoplasma gondii, takizoit, DNA rekombinan
Transcript of KLONING GEN RHOPTRY 1 - Universitas Udayana · 2017-06-06 · Terapi toksoplasmosis dengan obat...

1
KLONING GEN PENYANDI PROTEIN RHOPTRY 1 (ROP1) TAKIZOIT Toxoplasma gondii ISOLAT LOKAL
I Wayan Surudarma
ABSRAK INTISARI
Protein Rhopry 1 (ROP1) takizoit Toxoplasma gondii merupakan salah satu protein yang berperan penting pada proses invasi parasit ini ke dalam sel hospes. Protein tersebut diperkirakan sebagai molekul penetration-enhanching factor (PEF). Kloning gen penyandi ROP1 takizoit Toxoplasma gondii isolat RH untuk vaksinasi DNA pada mencit telah diteliti. Imunisasi DNA tersebut dapat meningkatkan respon imun seluler dan humoral. Kloning gen penyandi ROP1 takizoit Toxoplasma gondii isolat lokal belum pernah dilakukan. Tujuan penelitian ini ialah untuk memperoleh klon yang membawa gen penyandi ROP1 takizoit isolat lokal melalui teknik DNA rekombinan.
Takizoit Toxoplasma gondii dikultivasi secara in vivo pada mencit strain Balb/C. Isolasi DNA dilakukan setelah beberapa kali pasase untuk diperoleh jumlah takizoit yang cukup (minimal 1x108 takizoit/ml) dan selanjutnya DNA diamplifikasi menggunakan PuRe Taq RTG-PCR Beads (Amersham Bioscience) dengan primer spesifik (Cybergene AB) dan produknya diligasi pada pGEM-T® Easy (Promega). Plasmid rekombinan ditransformasi ke dalam Escherichia coli XL-1 Blue dengan teknik heat shock dan transforman ditanam pada plate agar LB yang mengandung ampisilin, X-gal, dan IPTG. Koloni putih yang menunjukkan koloni rekombinan ditumbuhkan dalam media LB yang telah ditambahkan ampisilin pada suhu 37oC selama semalam. Plasmid DNA rekombinan diisolasi dengan metode lisis alkali dan dielektroforesis pada gel agarose 1% (SeaKem). Analisis plasmid rekombinan dilakukan dengan cara digesti menggunakan endonuklease restriksi EcoRI dan selanjutnya disekuensing untuk mengetahui urutan basanya (ABI 3130 Genetic Analyzer). Hasil penelitian menunjukkan bahwa kloning gen penyandi ROP1 Toxoplasma gondii isolat lokal menghasilkan klon yang membawa DNA insert dengan ukuran 1441 bp. DNA insert tersebut mempunyai homologi sebesar 99% dengan gen penyandi ROP1 Toxoplasma gondii isolat RH. Kata kunci: ROP1, Toxoplasma gondii, takizoit, DNA rekombinan

2
PENDAHULUAN
Toxoplasma gondii merupakan parasit obligat intraseluler yang mampu
menginfeksi sel berinti semua vertabrata berdarah panas termasuk manusia
(Khan et al., 2006). Infeksi parasit ini disebut toksoplasmosis dan telah
menjangkiti hampir sepertiga populasi dunia (Mital et all, 2005). Prevalensi
toksoplasmosis berkisar antara 5 – 95% tergantung dari lokasi geografisnya
(Yanesa et al., 1994). Di Indonesia, prevalensi zat anti Toxoplasma gondii pada
manusia berkisar antara 2% sampai 63% (Gandahusada, 1998). Toksoplasmosis
pada hewan juga mempunyai arti penting dalam bidang kesehatan dan
perekonomian karena dapat menyebabkan abortus dan kematian neonatus pada
ternak. Kista jaringan dalam daging hewan terinfeksi dapat merupakan sumber
infeksi yang penting pada manusia (Ismael et all, 2003).
Toxoplasma gondii yang menginfeksi sel hospes akan bereplikasi dalam
vakuola parasitoporus dan menyebabkan lisis sel yang terinfeksi. Proses invasi
dan lisis sel yang berulang-ulang menyebabkan munculnya gejala toksoplasmosis
(Remington et all., 1995). Antigen yang berkaitan langsung dengan
imunopatogenesis dan invasi ke dalam sel hospes adalah antigen yang berasal dari
protein permukaan (SAG/surface antigen) dan antigen beredar (ESA/excretory-
secretory antigen). Antigen permukaan sangat berkaitan dengan proses perlekatan
(attachment) awal pada permukaan sel target, sedangkan ESA lebih berperan pada
proses penetrasi dan modifikasi vakuola parasitoporus dalam sel target
(Grimwood et al., 1996; Ajioka et al., 2001; Binder et al., 2004).

3
Protein ESA dilepaskan oleh tiga organela sekretoris apikal, yaitu
micronemes, rhoptries dan dense granules (Carruthers and Sibley, 1997).
Protein micronemes (MIC) dan rhoptries (ROP) dilepaskan pada saat parasit
menarik dirinya ke permukaan sel hospes dengan menggunakan myosin-based
motor complex (Sahoo et al., 2006; Meissner et all., 2002). Mikronema
mengandung berbagai adesin yang membantu pengikatan ligan ke permukaan sel
hospes (Sahoo et al., 2006; Tomley and Soldati, 2001). Protein rhoptry berperan
dalam penetrasi dan pembentukan membran vakuola parasitoporus (Sinai and
Joiner, 2001). Protein rhoptry 1 merupakan ROP dengan berat molekul 66 kDa
dan berfungsi sebagai penetration-enhanching factor (PEF) (Ossorrio et al., 1992;
Guanjin et al., 2001). Protein dense granules (GRA) dilepaskan bila parasit telah
berada dalam sel hospes, dan berperan dalam modifikasi kompartemen
intraseluler, tempat parasit bertahan hidup (Carruthers and Sibley, 1997).
Diagnosis laboratorium untuk infeksi akut Toxoplasma gondii didasarkan
pada tiga metode, yaitu: Isolasi takizoit dari darah atau cairan tubuh melalui kultur
sel, deteksi antigen atau antibodi spesifik secara serologis, dan pemeriksaan
histologis sampel jaringan untuk menemukan takizoit atau kista. Masing-masing
metode tersebut masih memiliki kelemahan (Potasman et al., 1988). Isolasi kultur
mempunyai kesulitan dalam menentukan waktu yang tepat dalam pengumpulan
sampel dan mempertahankan viabilitas parasit dalam spesimen (Israelski, 1988).
Metode serologis mempunyai keterbatasan karena antibodi dan antigen spesifik
Toxoplasma gondii mungkin tidak muncul pada awal infeksi, dan Ig M tidak
meningkat saat reaktivasi bentuk kista parasit (Holliman, 1990). Biopsi jaringan

4
merupakan prosedur invasif sehingga kurang aman dan nyaman untuk diterapkan.
Deteksi Toxoplasma gondii menggunakan PCR dapat mengurangi masalah ketiga
metode tersebut, namun metode PCR tidak dapat digunakan untuk membedakan
antara infeksi akut atau laten (Dupon et al., 1995).
Terapi toksoplasmosis dengan obat biasanya hanya dapat membunuh parasit
dalam stadium takizoit. Terapi ini cukup efektif untuk infeksi akut, namun tidak
efektif untuk infeksi laten karena kista dalam jaringan dapat menjadi aktif kembali
bila kondisi lingkungan memungkinkan (Gandahusada, 1998). Vaksin sangat
penting untuk mencegah reaktivasi pada individu yang immunocompromised,
mencegah infeksi janin pada kehamilan, dan mengontrol multiplikasi takizoit pada
infeksi primer akut (Denkers et al., 1998; Sibley et al., 1996). Vaksin takizoit
Toxoplasma gondii yang dilemahkan (live attenuated) dilaporkan berhasil baik
untuk mencegah aborsi pada hewan, dan telah digunakan secara komersial untuk
ternak domba, namun vaksin ini tidak cukup aman digunakan pada manusia
(Prigione et al., 2000; Alexander et al., 1996). Pendekatan vaksin toksoplasmosis
untuk manusia idealnya didasarkan pada penggunaan antigen rekombinan atau
peptida sintetik yang dapat memberikan perlindungan terhadap seluruh siklus
hidup Toxoplasma gondii (Prigione et al., 2000). Berdasarkan hal-hal tersebut
maka penting sekali dilakukan pengembangan perangkat diagnosis dan vaksin
secara biologi molekuler.
Penelitian untuk mencari antigen Toxoplasma gondii yang bersifat
imunogenik protektif banyak difokuskan pada antigen permukaan yang
diekspresikan pada stadium takizoit (Prigione et al., 2000). Penelitian mengenai

5
vaksin DNA untuk melawan infeksi Toxoplasma gondii secara eksperimental
pada tikus juga telah banyak dilakukan. Antigen yang diuji meliputi antigen
permukaan SAG (SAG1), GRA (GRA1, GRA4, GRA7) dan ROP
(ROP1 dan ROP2) (Scorza et al., 2003; Prigione et al., 2000). Protein MIC3 juga
dilaporkan sebagai kandidat vaksin yang menjanjikan karena merupakan adesin
yang poten dan diekspresikan pada semua stadium infeksius (Ismael et al., 2003).
Plasmid rekombinan pcDNA3-ROP1 (pcROP1) yang diimunisasi secara
intramuskular pada mencit dapat meningkatkan aktivitas sel NK, proliferasi
limfosit T dan sel T CD8+, serta titer Ig G (Guanjin et al., 2001). Penelitian
lainnya adalah peningkatan efikasi pcROP1 dengan menggunakan suatu plasmid
rekombinan pcIFN-α, hasilnya menunjukkan pcIFN-α dapat bekerja terkoordinasi
dengan pcROP1 untuk meningkatkan respon imun seluler (Hong et al., 1999).
Penyediaan asam nukleat dan protein dalam jumlah yang memadai sangat
diperlukan untuk pengembangan vaksin, perangkat diagnostik dan terapi imun.
Teknologi DNA rekombinan memungkinkan pemecahan masalah penyediaan
asam nukleat dan protein tersebut. Pendekatan yang dapat dilakukan adalah
melalui kloning gen penyandi ROP1 hasil amplifikasi DNA takizoit
Toxoplasma gondii. Amplifikasi dilakukan dengan menggunakan primer spesifik
untuk gen penyandi ROP1. Hasil amplifikasi selanjutnya diligasikan ke vektor
pGEM-T® Easy dan ditransformasikan ke dalam hospes E. coli.

6
MATERI DAN METODE
Bahan
Bahan untuk kultivasi secara in vivo pada penelitian ini adalah takizoit
T. gondii isolat lokal dan mencit strain Balb/C. Isolasi DNA takizoit
menggunakan bahan-bahan seperti proteinase-K, larutan NTE (5M NaCl, 1M
Tris-Cl, 0,5M EDTA), SDS 0,5%, fenol, kloroform, isoamil alkohol, Na-asetat,
dan etanol. Amplifikasi DNA menggunakan primer R1F1 dan R1R2 (Cybergene
AB), dan PuRe Taq Ready to Go PCR Beads (Amersham Biosciences).
Vektor kloning yang digunakan adalah pGEM-T® Easy (Promega). Bakteri yang
digunakan adalah E. coli XL-1 Blue. Bahan yang digunakan untuk plating bakteri
adalah 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-gal) (Sigma),
isopropyl β-D-thio-galacto-pyranoside (IPTG) (Sigma) dan ampisilin (Sigma).
Isolasi DNA plasmid rekombinan memerlukan bahan-bahan: lysing solution I (2M
glukosa, 0,5M EDTA, 1M Tris), lysing solution II (0,2 N NaOH, 1% SDS),
lysing solution III (5 M kalium asetat, asam asetat), etanol absolut, etanol 70%,
dan TE. Digesti plasmid menggunakan enzim restriksi endonuklease EcoRI
(Fermentas). Marker DNA yang dipergunakan adalah DirectLoadTM Wide Range
DNA Marker (Sigma). Sekuensing DNA menggunakan BigDye® Terminator v3.1
Cycle Sequensing Kit dan AutoSEQ G-50 Kit (Amersham).
Jalannya Penelitian
Kutivasi parasit in vivo

7
Tiga ekor mencit dewasa disuntik dengan takizoit T. gondii isolat lokal secara
intraperitoneal dengan dosis 1 x 107 takizoit. Mencit menunjukkan gejala sakit
dengan ditandai bulu berdiri, lemah, tidak ada nafsu makan dan minum, frekuensi
pernafasan menurun, dan denyut jantung cepat setelah 72-96 jam. Mencit
kemudian dibunuh dan dilakukan pencucian rongga perut dengan cairan NaCl
fisiologis untuk mendapatkan takizoit. Pencucian dilakukan tiga kali,
masing-masing sebanyak 5–10 ml. Takizoit selanjutnya diinfeksikan lagi pada
30–40 ekor mencit dewasa dengan dosis 1 x 107 untuk mendapatkan jumlah
takizoit yang lebih banyak dengan cara yang sama. Hasil cucian rongga perut
mencit, kemudian disentrifugasi 3.000 rpm (Beckman) pada suhu 4oC selama 10
menit. Pelet yang didapatkan dicuci tiga kali dengan penambahan PBS dan
disentrifugasi 3.000 rpm pada suhu 4oC selama 10 menit dan selanjutnya pelet
siap digunakan untuk isolasi DNA.
Isolasi DNA Sel takizoit yang telah dicuci dengan PBS diresuspensi dengan larutan NTE,
ditambahkan proteinase-K (10 mg/ml) dan SDS 0,5% sehingga konsentrasi akhir
menjadi 100 μg/ml, dan diinkubasi semalam dalam waterbath 37oC (Buchii).
Campuran reaksi tersebut ditambahkan fenol 1:1 dan di-shaking 60 rpm selama
20 menit, kemudian disentrifugasi 3000 rpm selama 15 menit pada suhu kamar
dengan J-6B centrifuge (Beckman). Fase atas yang terbentuk dipindahkan
ke tabung baru dan ditambahkan 1x volume kloroform:isoamil alkohol (24:1) dan
disentrifugasi 3000 rpm selama 10 menit. Langkah ini diulangi sampai bersih
(tidak ada interfase). Fase atas ditambahkan 1/10 volume 3 M Na-asetat dan

8
2x volume etanol absolut (95%) dingin dan dibiarkan 10-15 menit pada suhu
-20oC. Selanjutnya disentrifugasi pada kecepatan maksimum selama 5 menit,
kemudian pelet dibilas dengan alkohol 70% dan dikering-anginkan. Pelet DNA
yang sudah kering kemudian dilarutkan dengan TE dan ditentukan
konsentrasinya.
Elektroforesis DNA
Elektroforesis DNA dilakukan dengan menggunakan gel agarose 1% yang
dibuat dengan cara menimbang agarose 0,3 gram (SeaKem) dan ditambahkan
30 ml TAE 1x, kemudian dilarutkan di dalam microwave (Hitachi) dan dalam
keadaan hangat-hangat kuku ditambahkan 1 µl ethidium bromide (C21H20N3Br;
2,7- diamino-10-ethyl-9-phenyl- henanthridinium bromide; homidium bromide).
Selanjutnya dipindahkan ke gel tray yang telah dipasang sisiran untuk membuat
sumuran. Gel yang terbentuk dimasukkan ke dalam tangki elektroforesis (Biorad),
kemudian dituangkan TAE 1x sampai gel terendam. Selanjutnya sampel DNA
sebanyak 5 µl dicampurkan dengan 1 µl DNA loading buffer di atas parafilm,
kemudian dimasukkan ke dalam sumuran gel dengan hati-hati, kemudian
elektroforesis dilakukan sampai migrasi DNA hampir mencapai bagian ujung gel.
Hasil elektroforesis dapat dilihat dengan UV transilluminator.
Amplifikasi DNA
Primer spesifik yang digunakan untuk mengamplifikasi gen penyandi protein
ROP1 adalah primer foward R1F1 (5’ CGTGACATATACTGCACTGAC 3’)

9
dan primer reverse R1R2 (5’ CATCGTCAAACTCGATCAC 3’). Primer
diencerkan sehingga konsentrasi kedua primer tersebut menjadi 10 pmol/μl
sebelum digunakan. Campuran reaksi untuk proses amplifikasi dibuat dengan
penambahan 2 μl template, 2 μl primer R1F1, dan 2 μl primer R1R2 serta
19 μl dH2O filter ke dalam puRe Taq RTG-PCR sehingga volume total menjadi
25 μl. Tabung berisi campuran reaksi tersebut dimasukkan ke dalam thermocycler
(Gene Cycler, BioRad) dan dijalankan dengan program sebagai berikut:
(1) denaturasi awal dengan suhu 94oC selama 5 menit, (2) denaturasi dengan suhu
94oC selama 1 menit, (3) annealing primer dengan suhu 60oC selama 1 menit,
(4) polimerisasi dengan suhu 72oC selama 1 menit, (5) siklus diulang sehingga
total siklus 35 kali, dan (6) diakhiri dengan polimerisasi tambahan pada suhu 72oC
selama 5 menit. Proses amplifikasi dikontrol dengan menggunakan kontrol positif
dan kontrol negatif. Kontrol positif menggunakan campuran reaksi dengan
template DNA isolat RH, sedangkan kontrol negatif dengan tanpa menggunakan
template. Hasil amplifikasi dicek dengan dielektroforesis pada gel agarose 1%.
Purifikasi produk amplifikasi DNA
Produk PCR diencerkan menjadi 100 µl dengan akuades steril dan dicampur
sampai homogen, kemudian ditambahkan dengan fenol:CIAA (96:4) dengan
volume yang sama, divorteks beberapa detik dan disentrifugasi pada kecepatan
12.000 rpm selama 10 menit (microfugeTM11, Beckman). Supernatan pada lapisan
bagian atas diambil dan ditempatkan pada tabung baru. DNA produk PCR
diendapkan dengan 2x volume etanol absolut dan 1/10 volume natrium asetat 3M
(pH 4,8), kemudian diinkubasi pada suhu -700C selama 1 jam untuk mempercepat

10
pengendapan. Hasil pengendapan disentrifugasi 12.000 rpm selama 10 menit,
selanjutnya pelet dicuci dengan etanol 70% dan dikering-anginkan. Pelet hasil
purifikasi diresuspensi dengan bufer TE (pH 8) dan siap untuk diligasikan ke
vektor.
Ligasi produk amplifikasi DNA dengan vektor kloning
Sebanyak 1µl pGEM-T® Easy (50 ng/µl) ditambah dengan 3 µl produk PCR
yang telah dipurifikasi, 5 µl 2 X buffer T4 DNA ligase dan 1 µl T4 DNA ligase
sehingga volume akhir menjadi 10 µl. Campuran reaksi diinkubasi pada suhu 4oC
selama semalam. Hasil reaksi disimpan pada suhu –20oC dan siap untuk
ditransformasikan pada sel hospes.
Transformasi
Transformasi adalah proses memasukkan vektor ke dalam sel hospes.
Transformasi pada penelitian ini menggunakan teknik heat shock, dan preparasi
sel kompeten menggunakan metode TSS. Hasil transformasi ditanam plate agar
LB yang ditambahkan dengan X-gal, IPTG dan ampisilin.
Preparasi sel kompeten Koloni tunggal E. coli XL1-Blue dibiakkan dalam 5 ml media LB dan
diinkubasi selama semalam pada suhu 37oC dengan kecepatan agitasi 200 rpm.
Bakteri kemudian ditumbuhkan kembali dalam 25 ml LB dan diinkubasi selama
2 – 3 jam sampai didapatkan OD600 antara 0,5 – 0,6 dengan spectrophotometer

11
(Beckman DU-65). Sebanyak 20 ml LB medium yang mengandung bakteri
kemudian dipanen dengan cara disentrifugasi dengan kecepatan 3.000 rpm selama
15 menit pada temperatur 40C (J-6B centrifuge, Beckman). Supernatan dibuang
dan pelet ditambahkan dengan 2 ml TSS 2X. Pelet kemudian diresuspensi dan
dibagi-bagi dalam volume 200 µl. Sel kompeten sudah siap digunakan atau
disimpan pada suhu -700 C.
Transformasi dengan teknik heat shock Sel kompeten bakteri E. coli XL1-Blue (volume 200 μl) ditambah dengan
3 μl plasmid pGEM-T® Easy yang sudah diligasikan dengan produk PCR,
kemudian dicampur dengan baik. Kontrol transforman terdiri atas kontol positif
(E. coli XL1-Blue dengan plasmid pGEM-T® Easy yang diligasikan dengan
control insert DNA), kontrol background (E. coli XL1-Blue dengan plasmid
pGEM-T® Easy saja) dan kontrol negatif (E. coli XL1-Blue tanpa plasmid).
Campuran bahan transformasi kemudian diinkubasi dalam es selama 30 menit dan
kemudian di heat shock dalam water bath 42oC selama 90 detik. Campuran bahan
transformasi yang telah di heat shock dimasukkan ke dalam es selama 1- 2 menit
dan kemudian ditambah 500 μl TSS 1X dan difliking. Bakteri E. coli XL1-Blue
hasil transformasi kemudian diinkubasi dalam water bath 37oC selama 1 jam
sambil digoyang-goyang tiap 5 menit. Selanjutnya sampel siap untuk ditanam
pada plate agar.
Penanaman pada plate agar

12
Sampel hasil transformasi ditumbuhkan pada plate agar LB yang sudah
ditambahkan dengan X-gal, IPTG dan ampisilin. Khusus untuk kontrol sterilitas
tidak ditambah dengan zat-zat tersebut di atas. Sampel ditanam dalam jumlah
yang bertingkat mulai dari 100 μl, 200 μl, dan 300 μl untuk menghasilkan koloni
yang terbaik. Penanaman dilakukan secara merata di seluruh permukaan plate
agar dan diinkubasi pada suhu 37oC selama semalam. Hasil pertumbuhan bakteri
berupa koloni biru dan putih. Koloni rekombinan yang berwarna putih kemudian
dianalisis.
Analisis plasmid rekombinan
Koloni rekombinan dibiakkan dalam 5 ml media LB yang mengandung
ampisilin. Plasmid rekombinan diisolasi dengan metode lisis alkali,
dan selanjutnya dianalisis dengan cara digesti menggunakan endonuklease
restriksi EcoRI.
Penumbuhan bakteri rekombinan
Koloni tunggal bakteri rekombinan dibiakkan dalam 5 ml media LB yang
mengandung ampisilin, dan diinkubasi pada suhu 37oC selama semalam dengan
agitasi 200 rpm.
Isolasi plasmid rekombinan dengan metode lisis alkali Bakteri rekombinan hasil biakan dalam medium LB dituangkan ke tabung
micro tube sampai hampir penuh (1,5 ml), kemudian disentrifugasi 12.000 rpm
selama 5 detik (microfugeTM11, Beckman) dan supernatan dibuang. Pelet yang

13
didapat diresuspensi dengan 100 μl Lising Solution I (LS I), kemudian divorteks
sampai homogen dan diinkubasi dalam es selama 5 menit. Selanjutnya
ditambahkan 200 μl Lising Solution II (LS II), kemudian dicampur dengan
membolak-balikkan tabung micro tube kira-kira sebanyak 5 kali. Kemudian
ditambahkan lagi dengan 150 μl Lising Solution III (LS III), divorteks dan ditaruh
dalam es selama 5 menit. Larutan sampel selanjutnya disentrifugasi 12.000 rpm
selama 5 menit dan diambil supernatannya, lalu dimasukkan ke dalam micro tube
baru dan ditambahkan 250 μl fenol dan 250 μl CIAA kemudian divorteks.
Campuran ini disentrifugasi 12.000 rpm selama 3 menit, dan akan terbentuk tiga
fase larutan. Fase atas larutan diambil dan dimasukkan ke dalam micro tube baru,
kemudian ditambah etanol absolut dingin 2x volume dan selanjutnya diikubasi
pada suhu -70oC selama 1 jam. Larutan yang telah diinkubasi kemudian
disentrifugasi 12.000 rpm selama 5 menit. Supernatan dibuang dan pelet dibilas
dengan etanol dingin 70% (500μl) dan dikering-anginkan. Pelet yang sudah
kering diresuspensi dengan TE (volume disesuaikan dengan banyaknya pelet).
DNA plasmid selanjutnya dielektroforesis pada agarose gel 1% dan diamati
dibawah sinar ultraviolet.
Digesti plasmid rekombinan
Plasmid rekombinan didigesti dengan endonuklease restriksi EcoRI.
Lima mikroliter plasmid rekombinan (konsentrasi 1 µg/µl) ditambah dengan 2 µl
bufer EcoRI, 2 µl EcoRI dan nuclease free water sehingga volume total menjadi
20 µl dan dicampur dengan baik. Campuran reaksi kemudian diinkubasi pada

14
suhu 37oC selama 2 jam. Hasil digesti selanjutnya dielektroforesis pada gel
agarose 1% dan diamati dibawah sinar ultraviolet.
Sekuensing DNA plasmid rekombinan Sekuensing DNA dikerjakan dengan menggunakan ABI 3130 Genetic
Analyzer. Cycle sequensing terhadap plasmid rekombinan dilakukan dengan
menggunakan BigDye® Terminator v3.1 Cycle Sequensing Kit dengan primer
SP6, T7 dan R1R2. Reaksi cycle sequensing sebagai berikut: (1) denaturasi
pertama pada suhu 96oC selama 2 menit, (2) denaturasi 96oC selama 10 detik,
(3) annealing primer 50oC selama 5 detik, (4) polimerisasi 60oC selama 4 menit,
(5) siklus diulang sehingga total siklus 25 kali. Hasil cycle sequensing kemudian
dipurifikasi dengan AutoSEQ G-50 Kit (Amersham). Plasmid rekombinan yang
telah di-cycle sequensing ini akan menghasilkan fragmen DNA dengan panjang
yang berbeda-beda dan memiliki label fluoresensi pada ujungnya. Selanjutnya
fragmen-fragmen tersebut dipisahkan dengan elektroforesis gel poliakrilamid dan
sekuenser DNA akan mendeteksi fluoresen untuk mengidentifikasi A, T, C dan G.
BigDye Terminator dilabel dengan dichlororhodamine (dRhodamine) acceptor
dyes; terminator A dilabel dengan dR6G, terminator C dilabel dengan dROX,
terminator G dilabel dengan dR110, dan terminator T dilabel dengan dTAMRA.
Fragmen DNA yang berlabel fluoresen bermigrasi melalui gel dan melepaskan
sinar laser pada dasar gel. Masing-masing dye akan memancarkan cahaya pada
panjang gelombang yang berbeda-beda dan cahaya ini difokuskan oleh lensa ke
spektrograf. Warna pada electropherogram adalah hijau untuk A, biru untuk C,
hitam untuk G, dan merah untuk T.

15
Sekuen DNA yang didapatkan kemudian dianalisis dengan menggunakan
program BLAST yang diakses melalui NCBI untuk melihat alignment hasil
sekuensing.
HASIL PENELITIAN DAN PEMBAHASAN
Kultivasi Takizoit Toxoplasma gondii Isolat Lokal
Takizoit T. gondii dikultivasi secara in vivo pada 30 mencit Balb/C supaya
mendapatkan jumlah takizoit yang cukup untuk isolasi DNA. Jumlah takizoit
yang diperoleh adalah sebanyak 6,8x108 takizoit per ml.
Isolasi DNA Takizoit Toxoplasma gondii Isolat Lokal
Konsentrasi DNA yang diperoleh dalam penelitian adalah 827,5 ng/μl
dengan kemurnian 1,82. Hasil isolasi DNA dielektroforesis pada gel agarose 1%
dan hasilnya dapat dilihat pada Gambar 4. DNA ini selanjutnya akan digunakan
sebagai template dalam proses amplifikasi.
Amplifikasi DNA dengan Menggunakan Primer Spesifik
Primer spesifik yang digunakan untuk mengamplifikasi gen penyandi
protein ROP1 pada penelitian ini adalah primer foward R1F1
(5’ CGTGACATATACTGCACTGAC 3’) dan primer reverse R1R2
(5’ CATCGTCAAACTCGATCAC 3’). Berdasarkan sekuen gen penyandi ROP1
(kode: M71274.1) genbank NCBI, primer forward akan menempel pada basa ke-
171 sampai 191 dan primer reverse akan menempel pada basa ke-1515 sampai
16
kb
10
4
2 1,5
1 0,75
0,5
M 1 2 3
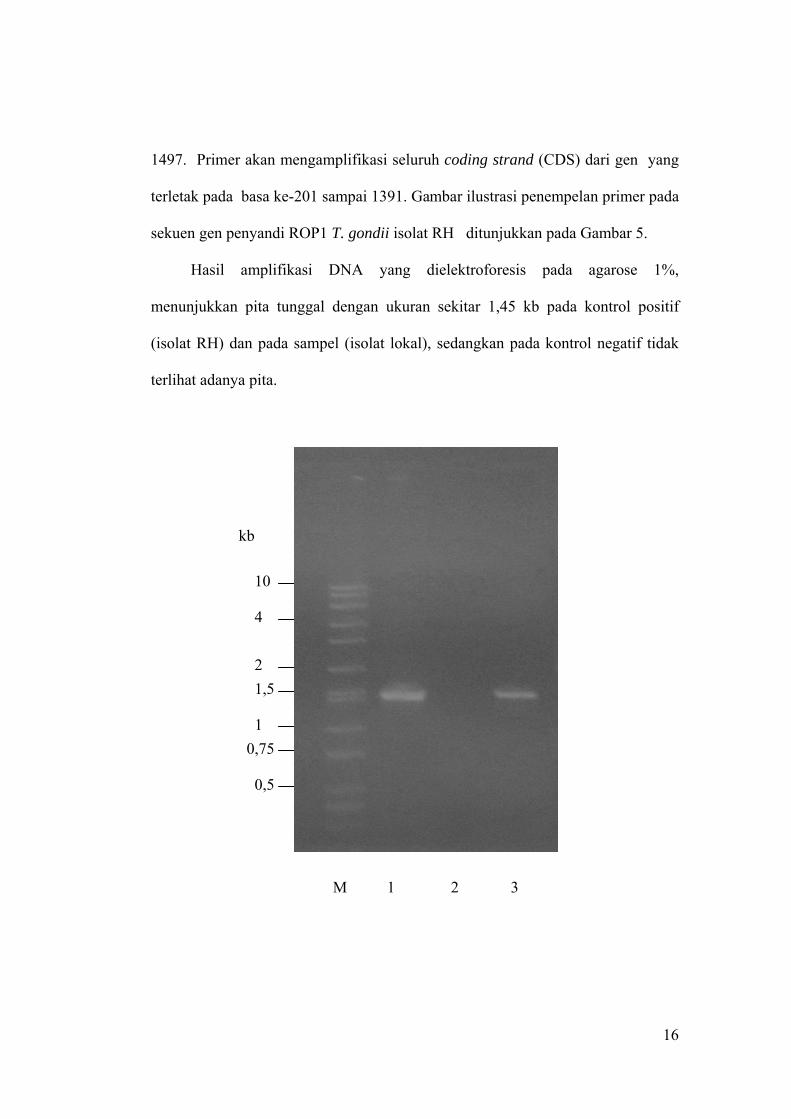
1497. Primer akan mengamplifikasi seluruh coding strand (CDS) dari gen yang
terletak pada basa ke-201 sampai 1391. Gambar ilustrasi penempelan primer pada
sekuen gen penyandi ROP1 T. gondii isolat RH ditunjukkan pada Gambar 5.
Hasil amplifikasi DNA yang dielektroforesis pada agarose 1%,
menunjukkan pita tunggal dengan ukuran sekitar 1,45 kb pada kontrol positif
(isolat RH) dan pada sampel (isolat lokal), sedangkan pada kontrol negatif tidak
terlihat adanya pita.

17
Kloning DNA Produk Amplifikasi
Hasil transformasi kemudian ditanam pada plate agar yang mengandung
ampisilin, IPTG dan X-gal selama semalam pada suhu 37oC. Hasil
penanaman membentuk 1 koloni putih dan 6 koloni biru.
Hasil transformasi pGEM-T Easy pada E. coli XL1-Blue dengan metode
heat shock. Keterangan: B. koloni biru; P. koloni putih. Identifikasi rekombinan pada kloning dengan vektor pGEM-T® Easy meliputi
seleksi resistensi transforman terhadap antibiotika ampisilin, dan kemudian
diikuti dengan skrining aktivitas β-galaktosidase untuk membedakan
sel rekombinan. Sel yang mengandung plasmid normal akan menjadi resisten
terhadap ampisilin dan mampu mensintesis β-galaktosidase, sedangkan sel
rekombinan juga akan resisten terhadap ampisilin tetapi tidak mampu mensintesis
β-galaktosidase.
P
B

18
M 1 2 3 4 5
Skrining ada atau tidaknya β-galaktosidase dilakukan dengan pengujian
menggunakan X-gal yang merupakan analog dari laktose. X-gal akan dipecah oleh
β-galaktosidase menjadi produk yang berwarna biru. Jika X-gal ditambahkan pada
media agar bersama IPTG dan ampisilin, maka koloni transforman bukan
rekombinan akan berwarna biru karena sel-selnya mensintesis β-galaktosidase,
sedangkan koloni transforman rekombinan akan membentuk berwarna putih
akibat tidak mampu mensintesis β-galaktosidase karena rusaknya gen lacZ’.
Analisis Plasmid Rekombinan
Koloni tunggal bakteri rekombinan (koloni putih) dibiakkan dalam 5 ml LB
medium yang mengandung ampisilin. Hasil pembiakan kemudian diisolasi
plasmidnya dan didigesti dengan endonuklease restriksi EcoRI. Hasil digesti
kemudian dielektroforesis pada gel agarose 1 %.
kb
10
4 3 2 1,5
0,75 0,5

19
Hasil eletroforesis plasmid yang didigesti dengan menggunakan endonuklease restriksi EcoRI pada gel agarose 1 %. Keterangan: M. marker; 1. plasmid koloni biru; 2. plasmid koloni biru yang didigesti dengan EcoR I; 3. plasmid koloni putih (rekombinan); 4. plasmid rekombinan yang didigesti dengan EcoR I; 5. produk PCR gen penyandi ROP1.
Digesti plasmid koloni biru menghasilkan pita tunggal dengan ukuran sekitar
3 kb, sesuai dengan ukuran plasmid pGEM-T® Easy. Digesti plasmid rekombinan
menghasilkan dua pita yang masing-masing berukuran sekitar 3 kb dan 1,45 kb,
sesuai dengan ukuran plasmid pGEM-T® Easy dan produk PCR. Digesti dengan
endonuklease restriksi EcoR I akan memotong vektor pGEM-T® Easy pada dua
tempat yang mengapit multiple cloning region, sehingga DNA insert akan
terlepas dari vektor. Plasmid rekombinan tersebut selanjutnya disebut pWTA-R1.
Sekuensing DNA Plasmid pWTA-R1
Plasmid WTA-R1 disekuensing dengan menggunakan primer sekuensing SP6,
T7 dan primer R1R2. Sekuensing dengan menggunakan primer T7 mendapatkan
sekuen nukleotida DNA insert sepanjang 590 bp (basa ke-1 sampai 590), dan
sekuensing menggunakan primer SP6 diperoleh sekuen nukleotida sepanjang 638
bp (basa ke-803 sampai 1441). Sekuensing dengan menggunakan primer R1R2
digunakan untuk melengkapi hasil sekuensing agar seluruh sekuen nukleotida
DNA insert dapat diketahui. Sekuensing dengan menggunakan primer R1R2
mendapatkan sekuen nukleotida sepanjang 982 bp (basa ke-407 sampai 1399).
Penggabungan hasil sekuensing dengan ketiga primer mendapatkan seluruh
sekuen nukleotida DNA insert, yaitu sebesar 1,441 kb.

20
1 cgt gac ata tac tgc act gac ttc gac acc atg gag caa agg ctg cca att att cta ctt
61 gtt ctc tct gtg ttc ttc agt tca acc cca agc gcc gcc ctt tcg agc cac aat gga gtc
121 ccc gct tat cca tcg tat gca cag gta tcg ctc tct tcc aac ggc gag cca cgg cac agg
181 ggc ata cgc ggc agc ttc ctc atg tcc gta aag cca cac gca aac gct gat gac ttc gcc
241 tcc gac gac aac tac gaa ccg ctg ccg agt ttc gtg gaa gct cct gtc aga ggc ccg gac
301 caa gtc cct gcc aga gga gaa gct gct ctt gtc aca gag gag act cca gcg caa cag ccg
361 gcg gtg gct cta ggc agt gca gaa ggg gag ggg acc tcc act act gaa tcc gcc tcc gaa
421 aat tct gaa gat gat gac acg ttt cac gat gcc ctc caa gag ctt cca gag gat ggc ctc
481 gaa gtg cgc cca cca aat gca cag gag ctg ccc cca cca aat gta cag gag ctg ccc cca
541 cca aat gta cag gag ctg ccc cca cca act gaa cag gag ctg ccc cca cca act gaa cag
601 gag ctg ccc cca cca act gaa cag gag ctg ccc cca cca act gaa cag gag cta ccc cca
661 tca act gaa cag gag ctg ccc cca cca gtg ggc gaa ggt caa cgt ctg caa gtc cct ggg
721 gaa cat ggg cca cag ggg ccc cca tac gat gat cag cag ctg ctt tta gag cct acg gaa
781 gag caa cag gag ggc cct cag gag ccg ctg cca ccg ccg ccg ccc ccg act cgg ggc gaa
841 caa ccc gaa gga cag cag ccg cag gga cca gtt cgt caa aat ttt ttt cgt cgg gcg ttg
901 ggg gcc gca aga agc cga ttc gga ggt gca cga cgc cat gtc agt ggg gtg ttc cga aga
961 gtc aga ggt ggt ttg aac cgt ata gta ggt gga gtg agg agt ggt ttc agg cgt gca aga
1021 gaa ggt gtc gtt ggg gga gtc cgt cgt tta aca agt ggt gcc agt ctg ggt ctc cgt cgt
1081 gta gga gaa ggt tta cgt agg agt ttc tat cgt gta aga gga gct gtc agt agc ggt cgt
1141 agg cgt gca gca gat ggt gcc agc aat gta aga gaa aga ttc gtt gcc gca ggc ggg aga
1201 gtc aga gac gct ttc ggc gcg gga ttg acg cgc ctc cgc agg cgc ggc aga act aat ggc
1261 gag gag ggc agg ccc cta ctg ggc gaa gga aga gag cag gat gat gga tcg caa taa tac
1321 ggg cag cat gct gct gga ttc ggc gaa gac gac cgt ttc tcg taa acg agg cag cgg ggt
1381 cct ccg aag tta aga aac ccg gta aac gtg tgt gcc gta acg gtg atc gag ttt gca gat
1441 g
Sekuen DNA insert pWTA-R1.
Hasil sekuensing pWTA-R1 selanjutnya dianalisis dengan program BLAST
untuk mengidentifikasi gen yang diklon. Sekuen DNA insert menunjukkan
alignment yang signifikan dengan sekuen parsial gen penyandi ROP1 T. gondii
isolat RH (kode: AF350261). Alignment hasil sekuensing DNA insert pWTA-
R1 dengan seluruh sekuen parsial gen penyandi ROP1 T. gondii isolat RH tersebut
menunjukkan homologi sebesar 99%. Basa yang tidak sama hanya terdapat pada
urutan ke-1162, yaitu guanin (G) menjadi adenin (A). Perbedaan basa tersebut
menyebabkan perubahan kodon GGC (glisin) menjadi AGC (serin).

21
gi|13560798|gb|AF350261.1|AF350261 T. gondii ROP1 gene, partial sequence. Length=1249 Score = 2426 bits (1224), Expect = 0.0 Identities = 1248/1249 (99%), Gaps = 0/1249 (0%) Strand=Plus/Plus Query 99 CCTTTCGAGCCACAATGGAGTCCCCGCTTATCCATCGTATGCACAGGTATCGCTCTCTTC 158 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1 CCTTTCGAGCCACAATGGAGTCCCCGCTTATCCATCGTATGCACAGGTATCGCTCTCTTC 60 Query 159 CAACGGCGAGCCACGGCACAGGGGCATACGCGGCAGCTTCCTCATGTCCGTAAAGCCACA 218 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 61 CAACGGCGAGCCACGGCACAGGGGCATACGCGGCAGCTTCCTCATGTCCGTAAAGCCACA 120 Query 219 CGCAAACGCTGATGACTTCGCCTCCGACGACAACTACGAACCGCTGCCGAGTTTCGTGGA 278 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 121 CGCAAACGCTGATGACTTCGCCTCCGACGACAACTACGAACCGCTGCCGAGTTTCGTGGA 180 Query 279 AGCTCCTGTCAGAGGCCCGGACCAAGTCCCTGCCAGAGGAGAAGCTGCTCTTGTCACAGA 338 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 181 AGCTCCTGTCAGAGGCCCGGACCAAGTCCCTGCCAGAGGAGAAGCTGCTCTTGTCACAGA 240 Query 339 GGAGACTCCAGCGCAACAGCCGGCGGTGGCTCTAGGCAGTGCAGAAGGGGAGGGGACCTC 398 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 241 GGAGACTCCAGCGCAACAGCCGGCGGTGGCTCTAGGCAGTGCAGAAGGGGAGGGGACCTC 300 Query 399 CACTACTGAATCCGCCTCCGAAAATTCTGAAGATGATGACACGTTTCACGATGCCCTCCA 458 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 301 CACTACTGAATCCGCCTCCGAAAATTCTGAAGATGATGACACGTTTCACGATGCCCTCCA 360 Query 459 AGAGCTTCCAGAGGATGGCCTCGAAGTGCGCCCACCAAATGCACAGGAGCTGCCCCCACC 518 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 361 AGAGCTTCCAGAGGATGGCCTCGAAGTGCGCCCACCAAATGCACAGGAGCTGCCCCCACC 420 Query 519 AAATGTACAGGAGCTGCCCCCACCAAATGTACAGGAGCTGCCCCCACCAACTGAACAGGA 578 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 421 AAATGTACAGGAGCTGCCCCCACCAAATGTACAGGAGCTGCCCCCACCAACTGAACAGGA 480 Query 579 GCTGCCCCCACCAACTGAACAGGAGCTGCCCCCACCAACTGAACAGGAGCTGCCCCCACC 638 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 481 GCTGCCCCCACCAACTGAACAGGAGCTGCCCCCACCAACTGAACAGGAGCTGCCCCCACC 540 Query 639 AACTGAACAGGAGCTACCCCCATCAACTGAACAGGAGCTGCCCCCACCAGTGGGCGAAGG 698 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 541 AACTGAACAGGAGCTACCCCCATCAACTGAACAGGAGCTGCCCCCACCAGTGGGCGAAGG 600 Query 699 TCAACGTCTGCAAGTCCCTGGGGAACATGGGCCACAGGGGCCCCCATACGATGATCAGCA 758 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 601 TCAACGTCTGCAAGTCCCTGGGGAACATGGGCCACAGGGGCCCCCATACGATGATCAGCA 660 Query 759 GCTGCTTTTAGAGCCTACGGAAGAGCAACAGGAGGGCCCTCAGGAGCCGCTGCCACCGCC 818 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 661 GCTGCTTTTAGAGCCTACGGAAGAGCAACAGGAGGGCCCTCAGGAGCCGCTGCCACCGCC 720 Query 819 GCCGCCCCCGACTCGGGGCGAACAACCCGAAGGACAGCAGCCGCAGGGACCAGTTCGTCA 878 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 721 GCCGCCCCCGACTCGGGGCGAACAACCCGAAGGACAGCAGCCGCAGGGACCAGTTCGTCA 780 Query 879 AAAtttttttCGTCGGGCGTTGGGGGCCGCAAGAAGCCGATTCGGAGGTGCACGACGCCA 938 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 781 AAATTTTTTTCGTCGGGCGTTGGGGGCCGCAAGAAGCCGATTCGGAGGTGCACGACGCCA 840 Query 939 TGTCAGTGGGGTGTTCCGAAGAGTCAGAGGTGGTTTGAACCGTATAGTAGGTGGAGTGAG 998

22
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 841 TGTCAGTGGGGTGTTCCGAAGAGTCAGAGGTGGTTTGAACCGTATAGTAGGTGGAGTGAG 900 Query 999 GAGTGGTTTCAGGCGTGCAAGAGAAGGTGTCGTTGGGGGAGTCCGTCGTTTAACAAGTGG 1058 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 901 GAGTGGTTTCAGGCGTGCAAGAGAAGGTGTCGTTGGGGGAGTCCGTCGTTTAACAAGTGG 960 Query 1059 TGCCAGTCTGGGTCTCCGTCGTGTAGGAGAAGGTTTACGTAGGAGTTTCTATCGTGTAAG 1118 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 961 TGCCAGTCTGGGTCTCCGTCGTGTAGGAGAAGGTTTACGTAGGAGTTTCTATCGTGTAAG 1020 Query 1119 AGGAGCTGTCAGTAGCGGTCGTAGGCGTGCAGCAGATGGTGCCAGCAATGTAAGAGAAAG 1178 ||||||||||||||||||||||||||||||||||||||||||| |||||||||||||||| Sbjct 1021 AGGAGCTGTCAGTAGCGGTCGTAGGCGTGCAGCAGATGGTGCCGGCAATGTAAGAGAAAG 1080 Query 1179 ATTCGTTGCCGCAGGCGGGAGAGTCAGAGACGCTTTCGGCGCGGGATTGACGCGCCTCCG 1238 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1081 ATTCGTTGCCGCAGGCGGGAGAGTCAGAGACGCTTTCGGCGCGGGATTGACGCGCCTCCG 1140 Query 1239 CAGGCGCGGCAGAACTAATGGCGAGGAGGGCAGGCCCCTACTGGGCGAAGGAAGAGAGCA 1298 |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1141 CAGGCGCGGCAGAACTAATGGCGAGGAGGGCAGGCCCCTACTGGGCGAAGGAAGAGAGCA 1200 Query 1299 GGATGATGGATCGCAATAATACGGGCAGCATGCTGCTGGATTCGGCGAA 1347 ||||||||||||||||||||||||||||||||||||||||||||||||| Sbjct 1201 GGATGATGGATCGCAATAATACGGGCAGCATGCTGCTGGATTCGGCGAA 1249
Alignment DNA insert pWTA-R1 dengan sekuen parsial gen penyandi
ROP1 isolat RH (kode: AF350261). KESIMPULAN DAN SARAN Penelitian ini berhasil mendapatkan klon yang membawa gen penyandi
ROP1 T. gondii isolat lokal dengan ukuran 1441 bp. Gen penyandi ROP1 T.
gondii isolat lokal tersebut mempunyai homologi 99% dengan gen penyandi
ROP1 T. gondii isolat RH.
Gen penyandi ROP1 takizoit T. gondii isolat lokal yang sudah dikloning
diharapkan dapat diekspresikan untuk menghasilkan protein rekombinan. Protein
tersebut selanjutnya dipelajari imunogenitasnya agar dapat digunakan untuk
pengembangan vaksin, perangkat diagnostik dan antibodi monoklonal.

23
DAFTAR PUSTAKA
1. Ajioka, J.W., Fitzpatrick, J.M. and Reitter, C.P. 2001. Toxoplasma gondii Genomics: Shedding Light on Pathogenesis and Chemotherapy. http://www-ermm.cbcu.cam.ac.uk.
2. Alexander, J., Jebbari, H., Bluethmann, H., Satoskar, A. and Roberts, C.W. 1996. Immunological Control of Toxoplasma gondii and Appropriate Vaccine Design. In: Current Topics in Microbiology and Immunology. Gross, U. (ed.). Springer-Verlag, Berlin, Heidelberg. 183-195.
3. Ausubel and Frederick, M. 1995. Short Protocols in Molecular Biology. A Compendium of Methodes from Current Protocols in Molecular Biology. 3rd edition. John Wiley & Sons, Inc. Canada.
4. Baxter, A. 2001. What is Toxoplasma gondii. Copyright by Page Wise. Inc. http://www.Toxoplasma gondii.htm.
5. Beaman, M.H. 1995. Toxoplasma gondii. In Principles and Practice of Infectious Diseases (Mandell, G.L., Bennett, J.E. and Dolin, R., eds), pp. 2455-2475, Churchill Livingston, New York, NY, USA.
6. Becker and Jeffrey, M. 1996. Biotechnology : a Laboratory Course. 2nd edition. Academic Press, Inc. California.
7. Binder, E.M. and Kim, K. 2004. Location, Location, Location: Trafficking and Function of Secreted Proteases of Toxoplasma and Plasmodium. Traffic 5:914-924.
8. Birge, E.A. 1994, Bacterial and Bacteriophage Genetic. Third Edition. Springer-Verlag , NewYork.
9. Black, M.W. and Boothroyd, J.C. 2000. Lytic Cycle of Toxoplasma gondii. Microbiol. Mol. Biol. Rev. 64(3):607-623.
10. Brown, T.A. 1999. Genomes. Oxford. BIOS Scientific Publisher Ltd. 20-21. 11. Carruthers, V. B., and L. D. Sibley. 1997. Sequential protein secretion from
three distinct organelles of Toxoplasma gondii accompanies invasion of human fibroblasts. Eur. J. Cell Biol. 73:114–123.
12. Cerede, O., Dubremetz, J.F., Soete, M., Deslee, D., Vial, H., Bout, D. and Lebrun, M. 2005. Synergistic Role of Micronemal Proteins in Toxoplasma gondii Virulence. J. Exp. Med. 201(3):453-463.
13. Che, A. 2002. Controlling Fluorescence in E. coli. J. Mol. Microbiol. Biotec. 2 (4): 513 – 519.
14. Coppens, I. and Joiner, K. A. 2001. Parasite–host cell interactions in toxoplasmosis: new avenues for intervention? http://www-ermm.cbcu.cam.ac.uk.
15. Denkers, E.Y. and Gazzinelli, R.T., 1998. Regulation and Function of T-Cell-Mediated Immunity during Toxoplasma gondii Infection. Clin. Microbiol. Rev. 11(4):569-588.
16. Dobrowolski, J. M. and Sibley, D. L. (1996). Toxoplasma invasion mammalian cells is powered by the actin cytoskeleton of the parasite. Cell 84, 933-939.

24
17. Dupon, M., Cazenave, J., Pellegrin, J.L. Detection of Toxoplasma gondii by PCR and tissue culture in cerebrospinal fluid and blood of human immunodeficiency virus-seropositive patients. J Clin. Microbiol. 1995; 33(9):2421-2426.
18. Gandahusada, S., 1998. Toxoplasma gondii. Dalam: Parasitologi Kedokteran. Edisi ketiga., Gandahusada, S., Ilahude, H.H.D., Pribadi, W., (eds). Balai Penerbit FK, UI, Jakarta. Hal.153-161.
19. Grimwood, J., Mineo, J.R. and Kasper, L.H. 1996. Attachment of Toxoplasma gondii to Host Cells is Host Cell Cycle Dependent. Infect. Immun. 64(10):4099-4104.
20. Guanjin, C., Hong, G., Fangli, L.U. and Huaqin, Z. 2001. Contruction of A Recombinant Plasmid Harbouring The Rhoptry Protein Gene of Toxoplasma gondii and Preliminary Observations on DNA Immunity. J. Chin. Med. 114 (8) : 837 – 840, China.
21. Holliman, R.E. The diagnosis of toxoplasmosis. Serodiag. Immunother. Infect Dis. 1990; 4:83-93.
22. Hong, G., Guanjin, C., Huanqin, AZ., Yongan, Z. And Fangli, L.U. 1999. Immune Responses in Mice Vaccinated With Recombinat plasmid pcDNA3 Containing ROP1 Gene From Tooplasma gondii. Chin. J. Parasitol. Parasit. Dis., Chinese.
23. Hoyen, D.O., and Joss, A.W.L. 1992. Human Toxoplasmosis. Oxford University Press. New York.
24. Israelski, D. M. In Sande MA, Volberding PA. eds. The Medical Management of AIDS. Philadelphia, Pa: W.B. Saunders; 1988:193.
25. Jones, T.C., Yeh, S. and Hirsch, J.G. 1972 The interaction between Toxoplasma gondii and mammalian cells. I. Mechanism of entry and intracellular fate of the parasite. J. Exp. Med. 136,1157-1172, PubMed ID: 73030933.
26. Khan, A., Jordan, C., Muccioli, C., Vallochi, A. L., Rizzo L. V., Jr, R. B., Vitor, R. W.A., Silveira, C., and Sibley, L. D. 2006. Genetic Divergence of Toxoplasma gondii Strains Associated with Ocular Toxoplasmosis, Brazil. Emerging Infectious Diseases. www.cdc.gov/eid. 12, No. 6.
27. Martin, V., Cespedes, G., Santilar, G., Pszenny, V., Guarnesa, E., Garberi, J.C. and Angel, S.O. 2000. Antigenicity of Recombinant ROP2 Protein of T. gondii Expressed in Escherichia coli. Departentode Parasitologia, Argentina.
28. Meissner, M., Schluter, D. and Soldati, D. 2002. Role of Toxoplasma gondii myosin A in powering parasite gliding and host cell invasion. Science 298: 837–840.
29. Mital J., Meissner, M., Soldati, D. and Ward, G. E. 2005. Conditional Expression of Toxoplasma gondii Apical Membrane Antigen-1 (TgAMA1) Demonstrates That TgAMA1 Plays a Critical Role in Host Cell Invasion. Mol. Biol. Cell 16, 4341–4349.
30. Ossorrio, P.N., Schwartzman, J.D., and Boothroyd A. Toxoplasma gondii rhoptry protein associated with host cell penetration has unusual charge asymmetry. Mol Biochem. Parasitol.1992; 50:1-16.

25
31. Potasman, I., Resnick, L., Luft B.J., and Remington, J.S. Intrathecal production of antibodies against Toxoplasma gondii in patients with toxoplasmic encephalitis and the acquired immunodeficiency syndrome (AIDS). Ann Intern Med. 1988; 108:49-51.
32. Prigione, I., Facchetti, P., Lecordier, L., Deslee, D., Chiesa, S., Cesbron-Delauw, M.F. and Pistoia, V. 2000. T Cell Clones raised from Chronically Infected Healthy Humans by Stimulation with Toxoplasma gondii Excretory-Secretory Antigens Cross-React with Live Takizoits: Characterization of the Fine Antigenic Specificity of the Clones and Implications for Vaccine Development. J. Immunol. 164:3741-3748.
33. Remington, J. S., McLeod, R. and Desmonts, G. 1995. Toxoplasmosis, p. 140–267. In J. S. Remington and J. O. Klein (ed.), Infectious diseases of the fetus and the newborn infant, 4th ed. W. B. Saunders Company, Philadelphia.
34. Sahoo, N., Beatty, W., Heuser, J., Sept, D. and Sibley, L. D. 2006. Unusual Kinetic and Structural Properties Control Rapid Assembly and Turnover of Actin in the Parasite Toxoplasma gondii. Mol. Biol. Cell 7, Issue 2, 895-906.
35. Sambrook, J., Frich, E. F., and Maniatis, T. 1989. Molecular Cloning, A Laboratory Manual. 2nd ed. Cold Spring Harbor Laboratory Press, New York.
36. Scorza, T., Souza, D., Laloup, M., Dewit, J., Braekeleer, J.D., Verschueren, H., Vercammen, M., Huygen, K. and Jongert, E. 2003. A GRA1 DNA Vaccine Primes Cytolytic CD8+ T Cells to Control Acute Toxoplasma gondii Infection. Infect. Immun. 71(1):309-316.
37. Sinai, A. P., and K. A. Joiner. 2001. The Toxoplasma gondii protein ROP2 mediates host organelle association with the parasitophorous vacuole membrane. J. Cell. Biol. 154:95–108.
38. Strachan, T. and Read, A.P. 1999. Human Molecular Genetic 2. Second Edition. BIOS Scientific Publisher Ltd, USA.
39. Tomley, F., and Soldati, D. 2001. Mix and match modules: structure and function of microneme proteins in apicomplexan parasites. Trends Parasitol. 17:81–88.
40. Vercammen, M., Scorza, T., Huygen, K., Braekeleer, J.D., Diet, R., Jacobs, D. Saman, E. and Verschueren, H. 2000. DNA Vaccination with Gene Encoding Toxoplasma gondii Antigens GRA1, GRA7, and ROP2 Induces Partially Protective Immunity Against Lethal Challenge in Mice. Infect. Immun. 68(1):38-45.
41. Weaver, R. F. 1999. Molecular Biology. WCB. McGraw-Hill, USA. 42. Weiss, L.M. and Kim, K. 2000. The Development and Biology of
Bradyzoites of Toxoplasma gondii. Albert Einstein College of Medicine, New York.
43. Wong, S. Y. and Remington, J. S. 1993. Biology of Toxoplasma gondii. AIDS 7:299-316.

26
44. Yanesa, A., Path, F.R.C. and Kumari. 1994. Prevalence of Toxoplasma Antibodies in Blood Donors in Al-Hassa. Departement of Microbiology Al-Hassa, Saudi Arabia.