
ENZIMOLOGI
19
TUGAS ENZIMOLOGI “PERBANDINGAN KECEPATAN AKTIVITAS ENZIM AMILASE PADA Ragi Tape (Saccharomyces cereviceae.), Pati Kecambah Kacang Hijau, dan Pati Kacang Kedelai TERHADAP STRUKTUR KANJI TERGELATINISASI” DISUSUN OLEH : NAMA : RIA AGUSTINA NIM : 11017006 PRODI : BIOLOGI INDUSTRI FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS AHMAD DAHLAN YOGYAKARTA 2013
-
Upload
a-biologist-uad -
Category
Documents
-
view
108 -
download
0
Transcript of ENZIMOLOGI

TUGAS ENZIMOLOGI
“PERBANDINGAN KECEPATAN AKTIVITAS ENZIM AMILASE PADA Ragi Tape
(Saccharomyces cereviceae.), Pati Kecambah Kacang Hijau, dan Pati Kacang Kedelai
TERHADAP STRUKTUR KANJI TERGELATINISASI”
DISUSUN OLEH :
NAMA : RIA AGUSTINA
NIM : 11017006
PRODI : BIOLOGI INDUSTRI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS AHMAD DAHLAN
YOGYAKARTA
2013

BAB I.
PENDAHULUAN
Saat ini, pemanfaatan enzim banyak diaplikasikan secara luas terutama dalam proses
pengolahan pangan komersial. Dewasa ini sebagian besar kebutuhan enzim masih dipenuhi
dengan jalan impor. Hal tersebut tidak menguntungkan dari segi devisa dan pengembangan
bioteknologi di Indonesia. Sumber enzim dapat diperoleh dari tanaman, hewan dan
mikroorganisme. Salah satu enzim pemecah pati adalah enzim α-amilase (α-1,4-glukan-
glukanodidrolase; EC.3.2.1.1.), enzim ini sangat berperan dalam industri pembuatan roti dan
sirup. Enzim α-amilase banyak terdapat pada kacang-kacangan, kecambah kacang-kacangan dan
beberapa jenis mikroogranisme.
Saccharomyces cerevisiae menghasilkan enzim zimase dan invertase. Enzim zimase
berfungsi sebagai pemecah sukrosa menjadi monosakarida (glukosa dan fruktosa (Judoamidjojo
dkk, 1992). Kacang hijau dalam bentuk kecambah mengandung enzim α-amilase (Winarno,
1983). Pada biji kedelai seperti halnya dalam tubuh makhluk hidup lainnnya ditemukan pula
sejumlah enzim. Salah satu enzim tersebut adalah enzim amylase, suatu enzim yang
mengkatalisa pemecahan (hidrolisis) senyawa pati menjadi komponen-komponen yang lebih
kecil seperti glukosa, maltose dan limit dekstrin. Pati tersusun atas dua bentuk yaitu amilosa dan
amilopektin. Pemecahan pati ini dilakukan oleh dua macam enzim amylase yaitu α-amilase dan
β-amilase (Abidin, 1984).
Pada percobaan ini akan dilakukan pemecahan dengan emzim amylase yang bersal dari
Saccharomyces cereviceae, pati kedelai dan pati kecambah kacang hijau. Pemecahan tersebut
akan diamati pada struktur morfologi kanji yang tergelatinisasi. Diamati bagaimana
retrodegradasi gelatin dari kanji oleh enzim amylase, dimana akan terlihat bahwa kanji
tergelatinisasi yang tadinya struktur morfologinya sangat kenyal menjadi rusak struktur
kekenyalannya.
Pada pengamatan, akan dilihat perubahan struktur morfologi kanji tergelatinisasi yang
akan dibandingkan dengan lama waktu perubahan struktur tersebut (kanji tergelatinisasi menjadi
lembek.

BAB II.
TINJAUAN PUSTAKA
Sumber enzim dapat diperoleh dari tanaman, hewan dan mikroorganisme. Salah satu
enzim pemecah pati adalah enzim α-amilase (α-1,4-glukan- glukanodidrolase; EC.3.2.1.1.),
enzim ini sangat berperan dalam industri pembuatan roti dan sirup. Enzim alfa-amilase banyak
terdapat pada kecambah kacang- kacangan (Setyono, 1982).
Ragi untuk tape merupakan campuran untuk populasi, dimana terdapat spesies-spesies
dari Aspergilus, Saccharomyce, Candida, Hansenula, sedang bakteri Acetobacter biasanya tidak
ketinggalan. Genus-genus tersebut hidup bersama secara sinergik; Aspergilus dapat
menyederhanakan amilum, sedang Saccharomyces, Candida, dan Hansenula menguraikan gula
menjadi alcohol dan bermacam-macam zat organik lainnya. Acetobacter menumpang untuk
mengubah alcohol menjadi asam cuka (Dwijoseputro, 2005).
Saccharomyces cerevisiae menghasilkan enzim zimase dan invertase. Enzim zimase
berfungsi sebagai pemecah sukrosa menjadi monosakarida (glukosa dan fruktosa (Judoamidjojo
dkk, 1992).
Kacang hijau dalam bentuk kecambah mengandung enzim α-amilase. Pada umumnya
perkecambahan toge berlangsung selama lima hari, aktivitas enzim α-amilase dapat ditentukan
dengan mengukur hasil degradasi pati yang biasanya diukur dengan penurunan kadar pati yang
larut atau dari kadar maltosa yang dihasilkan. Enzim α-amilase dapat memecah pati secara acak
dari tengah atau dari bagian dalam molekul, oleh karena itu disebut endoamilase (Winarno,
1983).
Umur kecambah sangat berpengaruh pada panjang hipokotil toge. Toge hanya tahan
disimpan selama 12 jam pada suhu ruang, setelah itu mulai layu ditandai dengan perubahan
warna pada akar menjadi kecoklatan. Seperti halnya umur perkecambahan dari biji gandum
berpengaruh terhadap aktivitas enzim α-amilase, dimana aktivitasnya terus bertambah sampai
hari keempat perkecambahan. Selanjutnya mengalami penurunan pada hari kelima umur
kecambah (Marie, 1993).
Pada biji kedelai seperti halnya dalam tubuh makhluk hidup lainnnya ditemukan pula
sejumlah enzim. Salah satu enzim tersebut adalah enzim amilase, suatu enzim yang mengkatalisa
pemecahan (hidrolisis) senyawa pati menjadi komponen-komponen yang lebih kecil seperti

glukosa, maltose dan limit dekstrin. Pati tersusun atas dua bentuk yaitu amilosa dan amilopektin.
Pemecahan pati ini dilakukan oleh dua macam enzim amilase yaitu α-amilase dan β-amilase
(Abidin, 1984).
Kamil (1982) menyatakan enzim α-amilase tidak atau belum terdapat (non-pre-exist)
pada biji kering tetapi baru dibuat kemudian pada waktu permulaan perkecambahan biji (early
stage of germination) oleh asam giberelat (GA), sedangkan enzim β-amilase sudah ada sejak
semula (pre-exist) didalam skutelum dan lapisan aleuron pada biji yang masih kering.
Kedua enzim ini menurut Kamil (1982) berbeda yaitu bahwa enzim α-amilase akan
merombak amilose dan amilopektin menjadi maltose dan glukosa, disamping itu juga akan
merombak dekstrin menjadi maltose dan glukosa, dengan adanya maltase kemudian maltose
akan dirubah menjadi glukosa, sedangkan enzim β-amilase pada waktu mulai perkecambahan
akan masuk kedalam endosperm untuk merombak amilosa menjadi glukosa yang bersifat terlarut
(water soluble) dan bisa diangkut (translocated). Enzim β-amilase akan merombak amilopektin
menjadi dekstrin yang bersifat tidak bisa diangkut (non translocated). Pasokan gula
monosakarida ke embrio menyebabkan ukuran koleoriza dan radikula bertambah besar dan
mampu menembus selaput putih.
PATI DAN TAPIOKA
Amilum adalah polimer karbohidrat dengan rumus (C6H12O6)n. Karbohidrat golongan
polisakarida ini banyak terdapat di alam. Terutama pada sebagian besar tumbuhan. Amilum
disebut juga pati yang terdapat pada umbi, daun, batang, dan biji. Amilum merupakan kelompok
terbesar karbohidrat cadangan yang dimiliki oleh tumbuhan sesudah selulosa. Butir-butir pati

apabila diamati dengan mikroskop ternyata berbeda-beda bentuk dan ukurannya, tergantung dari
tumbuhan apa pati tersebut diperoleh (Poedjadi, 2008).
Pati mengandung dua jenis polimer glukosa, α-amilase dan amilopektin. α-amilase terdiri
dari rantai-rantai unit D-glukosa yang panjang, dan tidak bercabang, digabungkan oleh ikatan
α1,4. Rantai ini juga beragam dalam berat molekulnya, dan beberapa ribu hingga mencapai
500.000. Amilopektin juga memiliki berat molekul yang tinggi dan strukturnya barcabang tinggi.
Ikatan glikosidik menggabungkan residu glukosa yang berdekatan di dalam rantai amilopektin
adalah ikatan α1,4, tetapi titik percabangan amilopektin merupakan ikatan α1,6. Glikogen
merupakan sumber utama polisakarida pada sel hewan. Seperti amilopektin, glikogen merupakan
polisakarida bercabang dari D-glukosa dalam ikatan α1,4 (Lehninger, 1982)
Gambar 3. Struktur Amilosa, Amilopektin dan Polisakarida dari Pati. (a) Amilosa, (b)
Amilopektin, dan (c) Polisakarida dari Pati (Lehninger, 1982)
Tepung tapioka (di pasaran sering dikenal dengan nama tepung kanji) adalah tepung yang
terbuat dari ubi kayu/singkong. Pembuatan dilakukan dengan cara diparut, diperas, dicuci,
diendapkan, diambil sari patinya, lalu dijemur/keringkan. Sifat tepung kanji, apabila dicampur
dengan air panas akan menjadi liat/seperti lem. Tepung tapioka disebut juga tepung kanji atau
tepung sagu (sagu singkong). Tepung tapioka akan memiliki perlakuan berbeda untuk setiap
jenis kue karena sifat yang dimiliki tepung tersebut.
Adapun sifat fisikimia pati tapioka adalah sebagai berikut :

Rasio amilosa dan amilopektin adalah 17% amilosan dan 83% amilopektin.
Bentuk granula semi bulat dengan salah satu bagian ujungnya mengerucut.
Ukuran 5-35µm.
Suhu gelatinisasi berkisar antara 52-64oC.
Kristalisasi 38%.
Kekuatan pembengkakan sebesar 42µm.
Kelarutan 31% (Aulia, 2011).
GELATINISASI PATI
Pomeranz (1991) menyatakan bahwa gelatinisasi merupakan proses pembengkakan
granula pati ketika dipanaskan dalam media air. Granula pati tidak larut dalam air dingin, tetapi
granula pati dapat mengembang dalam air panas. Naiknya suhu pemanasan akan meningkatkan
pembengkakan granula pati. Pembengkakan granula pati menyebabkan terjadinya penekanan
antara granula pati dengan lainnya. Mula-mula pembengkakan granula pati bersifat reversible
(dapat kembali ke bentuk awal), tetapi ketika suhu tertentu sudah terlewati, pembengkakan
granula pati menjadi irreversible (tidak dapat kembali). Kondisi pembengkakan granula pati yang
bersifat irreversible ini disebut dengan gelatinisasi, sedangkan suhu terjadinya peristiwa ini
disebut dengan suhu gelatinisasi.

Menurut Winarno (2002) dan Pomeranz (1991), suhu gelatinisasi tepung tapioka berada
pada kisaran 52-64°C. Menurut Swinkels (1985), suhu gelatinisasi tepung tapioka berkisar antara
65-70°C. Moorthy (2004) menyatakan bahwa gelatinisasi merupakan fenomena kompleks yang
bergantung dari ukuran granula, persentase amilosa, bobot molekul, dan derajat kristalisasi dari
molekul pati di dalaM granula. Pada umumnya granula yang kecil membentuk gel lebih lambat
sehingga mempunyai suhu gelatinisasi yang lebih tinggi daripada granula yang besar. Makin
besar bobot molekul dan derajat kristalisasi dari granula pati, pembentukkan gel semakin lambat.
Menurut Pomeranz (1991), tidak semua granula pati tergelatinisasi pada titik yang sama, tetapi
terjadi pada suatu kisaran suhu tertentu. Menurut Olkku dan Rha (1978) dalam Pomeranz (1991),
proses gelatinisasi melibatkan peristiwa-peristiwa sebagai berikut: (1) hidrasi dan swelling
(pengembangan) granula; (2) hilangnya sifat birefringent; (3) peningkatan kejernihan; (4)
peningkatan konsistensi dan pencapaian viskositas puncak; (5) pemutusan molekul-molekul
linier dan penyebarannya dari granula yang telah pecah.
Granula pati utuh tidak larut dalam air dingin. Granula pati dapat menyerap air dan
membengkak, tetapi tidak dapat kembali seperti semula (retrogradasi). Air yang terserap dalam
molekul menyebabkan granula mengembang. Pada proses gelatinisasi terjadi pengrusakan ikatan
hidrogen intramolekuler. Ikatan hydrogen berperan mempertahankan struktur integritas granula.
Terdapat- nya gugus hidroksil bebas akan menyerap air, sehingga terjadi pem- bengkakan
granula pati. Dengan demikian, semakin banyak jumlah gugus hidroksil dari molekul pati
semakin tinggi kemampuannya menyerap air.Oleh karena itu, absorbsi air sangat berpengaruh
terhadap viskositas (Aulia, 2011).
Pati dengan amilosa tinggi menyebabkan suhu gelatinisasi lebih tinggi. Suhu gelatinisasi
pati bahan baku juga berpengaruh terhadap efisiensi produksi. Semakin rendah suhu gelatinisasi
semakin singkat waktu gelatinisasi, yaitu 20 menit untuk tapioka dan 22 menit untuk pati jagung.
Suhu puncak granula pecah pati jagung adalah 95oC dan tapioka 80
oC, dengan waktu yang
dibutuhkan berturut-turut 30 dan 21 menit. Sifat ini berkaitan dengan energi dan biaya yang
dibutuhkan dalam proses produksi. Pati akan terhidrolisis bila telah melewati suhu gelatinisasi.
Kadar amilopektin yang tinggi (99%) akan meningkatkan suhu awal (70,8oC), maupun suhu
puncak gelatinisasi, yang diikuti oleh peningkatan energy (Scribd, 2012).

Pati dalam jaringan tanaman mempunyai bentuk granula (butir) yang berbeda – beda.
Bila pati mentah dimasukkan ke dalam air dingin, granula patinya akan menyerap air dan
membengkak. Namun demikian jumlah air yang terserap dan pembengkakannya terbatas. Air
yang terserap tersebut hanya dapat mencapai kadar 30 %.Granula pati dapat dibuat membengkak
luar biasa, tetapi bersifat tidak dapat kembali lagi pada kondisi semula. Perubahan tersebut
disebut gelatinasi. Suhu pada saat granula pati pecah disebut suhu gelatinasi yang dapat
dilakukan dengan penambahan air panas. Air dapat ditambahkan dari luar seperti halnya
pembuatan kanji dan puding, atau air yang ada dalam bahan makanan tersebut (Winarno, 1992).
Suhu dari gelatinasi tergantung pada konsentrasi pati. Makin kental larutan, suhu tersebut
makin lambat tercapai, sampai suhu tertentu kekentalan tidak bertambah, bahkan kadang-kadang
turun. Semakin tinggi konsentrasi pati maka gel yang terbentuk semakin kurang kental dan
setelah beberapa waktu viskositas akan turun. Suhu gelatinasi berbeda-beda bagi tiap jenis pati,
misalnya pada jagung 62-70oC, beras 68-78
oC, gandum 54,5-64
oC, dan tapioka 52-64
oC.
Garanula pati memiliki sifat merefleksikan cahaya terpolarisasi sehingga dibawah mikroskop
terlihat kristal hitam putih. Sifat ini disebut sebagai birefrigent. Pada waktu granula mulai pecah,
sifat birefringent ini akan menghilang (Winarno, 1992).
Selama gelatinasi granula-granula pati sangatlah mengembang. Jadi 1 % granula pati
dalam air dingin mempunyai viskositas yang rendah, namun bila dipanaskan sebuah pasta kental
diproduksi dimana semua air yang ada memasuki granula-granula. Hal tersebut menyebabkan
pati mengembang dan mendorong dengan kuat satu sama lain dalam kondisi yang menyerupai
sarang madu. Granula-granula yang sangat mengembang dapat dengan mudah dipecah dan
terintegrasi dengan pengadukan yang tidak terlalu cepat dan berakibat turunnya viskositas
pastanya. Gelatinasi pati, viskositas larutan-larutan pati dan karakteristik gel-gel pati tergantung
tidak hanya pada suhu, namun juga pada jenis dan jumlah zat penyusun lainnya yang ada. Pada
intinya bahan penyusun makanan yang mengikat air secara kuat menghambat gelatinasi pati
dengan mengikat air berebut dengan pati (Herschdoefer, 1986).
Beberapa contoh pati, yaitu jagung, tapioka, gandum, kentang, sagu, dan beras. Pada
suhu tinggi, molekul-molekul pati akan bergerak lebih cepat dan tidak terkontrol, sehingga
berakibat pada terbukanya ikatan intermolekular dan ikatan hidrogennya akan menyerap lebih
banyak air. Munculnya uap pertama kali pada pemanasan lanjut, disebut dengan poin gelatinasi

atau suhu gelatinasi. Poin gelatinasi, biasanya muncul pada kisaran temperatur yang saling
berdekatan, diawali dengan granula-granula berukuran besar membentuk gelatin terlebih dulu,
kemudian disusul dengan granula-granula lebih kecil. Selama proses gelatinasi, granula-granula
tersebut akan membengkak tidak terkira. Melalui pengadukan, granula-granula yang
membengkak tersebut dapat dengan mudah dipecahkan dan mengalami integrasi. Hal ini akan
berakibat pada viskositas tepung yang menurun drastis. Pada tahap awal gelatinasi, terjadi
tekanan antar granula, terutama saat air memasuki molekul-molekul pati. Tekanan ini
menyebabkan molekul pati sekitar granula ditarik secara tangensial di sepanjang permukaan
molekul (Fennema, 1985).
Proses kristalisasi kembali pati yang telah mengalami gelatinisasi tersebut, disebut
retrogradasi. Pati yang didiamkan beberapa saat, akan membentuk endapan di dasar wadahnya.
Jika pati yang dipanaskan sudah dingin, air mengadakan ikatan yang erat dengan molekul–
molekul pati, pada permukaan butir–butir pati yang membengkak. Gel yang kehilangan cairan
(keluarnya atau merembesnya cairan dari suatu gel dari pati), akan mengalami perubahan
struktur dan mengkerut, yang dinamakan sineresis. Proses ini dapat terjadi jika gel dibiarkan
dalam waktu yang lama (Winarno, 1992).
Butiran pati sama sekali tidak larut dalam air dingin, namun pada saat pemanasan butiran
– butiran tiba – tiba menggembung pada suhu yang disebut suhu penggelatinan. Umumnya, pati
dalam butir besar menggembung pada suhu lebih rendah daripada pati berbutir kecil; pati
kentang menggembung pada 59 – 67oC dan pati jagung pada 64 – 72
oC. Suhu penggembungan
ini dipengaruhi berbagai faktor, termasuk pH, pra perlakuan, laju pemanasan, dan adanya garam
dan gula. Pemanasan lebih lanjut diatas suhu penggelatinan mengakibatkan penggembungan
butiran lebih lanjut, dan campuran menjadi kental dan bening. Jika pasta diaduk, struktur butiran
rusak dan kekentalan sangat menurun. Jika pasta pati yang digodog didinginkan, pasta dapat
membentuk gel atau jika pendinginan dilakukan secara perlahan – lahan, komponen rantai lurus
dapat membentuk endapan sferokristal. Gejala ini disebut retrogradasi, sangat bergantung pada
ukuran molekul rantai lurus. Retrogradasi pasta pati sangat dipercepat dengan pembekuan.
Setelah pembekuan pasta pati yang dibekukan, terbentuk massa yang menyerupai spons yang
mudah dilepaskan sebagian besar airnya dengan sedikit tekanan. Penggembungan dihambat oleh
asam lemak karena pembentukan senyawa kompleks yang tidak larut dengan fraksi rantai linier.

Molekul pati jagung yang lebih kecil, mempunyai sekitar 400 satuan glukosa, menunjukkan
kecenderungan yang lebih besar untuk asosiasi. Hidrolisis rantai sampai sekitar 20 – 30 satuan
menghilangkan sama sekali kecenderungan asosiasi dan pengendapan (DeMan, 1997).
Apabila suspensi pati dalam air dipanaskan, air akan menembus lapisan luar granula dan
granula ini mulai menggelembung. Hal ini terjadi pada saat temperatur meningkat dari 60oC -
85oC, granula-granula dapat menggelembung hingga volumenya 5 kali lipat volume semula.
Ketika ukuran granula pati membesar campurannya menjadi kental. Pada suhu kira-kira 85oC,
granula pati pecah dan isinya terdispersi merata ke seluruh air disekelilingnya. Molekul berantai
panjang mulai membuka atau terurai dan campuran pati dan air menjadi makin kental
membentuk sol. Pada pendinginan, apabila perbandingan pati dan air cukup besar maka molekul
pati akan membentuk jaringan dengan molekul air terkurung didalamnya sehingga terbentuk gel.
Keseluruhan proses ini disebut gelatinasi. Gelatinasi pati sangat penting dalam proses
pengolahan seperti pada pengentalan saos, sup dan kuah daging (gravies) dengan penambahan
tepung dan tepung jagung. (Gaman & Sherrington, 1994).
Pati tidak larut dalam air dan jika dilarutkan dalam air dingin, pati dapat menyerap sedikit
air. Sampai suhu 60-70oC, pembengkakan masih dapat kembali ke bentuk semula, kemampuan
kembali ke bentuk semula tergantung pada jenis pati. Dengan temperatur yang lebih tinggi,
terjadi pembengkakan yang tidak dapat kembali ke bentuk semula yang disebut gelatinasi. Pati
mulai tergelatinasi pada suhu antara 60 – 70oC, suhu yang tepat tergantung pada jenis pati
tersebut. Sebagai contoh, pati yang berbeda mempunyai kerapatan yang berbeda pula, yang
menyebabkan perbedaan kemampuan granula menyerap air. Karena kehilangan
kemampuan birefringence terjadi pada saat gelatinasi terjadi secara cepat (pembengkakan
granula), kehilangan kemampuan birefringence adalah indikator yang baik untuk menentukkan
suhu gelatinasi. Granula yang besar yang biasanya tidak padat, mulai membengkak pertama.
Ketika, gelatinasi pada pati terjadi secara optimum, agitasi yang tidak diinginkan dapat memecah
pati yang telah membengkak dan menyebabkan pengurangan pasta (Sutanto, 2001).
Bila suspensi pati dalam air dipanaskan, beberapa perubahan selama terjadinya
gelatinisasi dapat diamati. Mula – mula suspensi pati yang keruh seperti susu tiba – tiba mulai
menjadi jernih pada suhu tertentu, tergantung jenis pati yang digunakan. Terjadinya translusi
larutan pati tersebut biasanya diikuti pembengkakan granula. Bila energi kinetik molekul –

molekul air menjadi lebih kuat daripada daya tarik menarik antarmolekul pati di dalam granula,
air dapat masuk ke dalam butir – butir pati. Hal inilah yang menyebabkan bengkaknya granula
tersebut. Karena jumlah gugus hidroksil dalam molekul pati sangat besar, maka kemampuan
menyerap air sangat besar. Terjadinya peningkatan viskositas disebabkan air yang dulunya
berada di luar granula dan bebas bergerak sebelum suspensi dipanaskan, kini sudah berada dalam
butir – butir pati dan tidak dapat bergerak dengan bebas lagi. Pati yang telah mengalami
gelatinasi dapat dikeringkan, tetapi molekul – molekul tersebut tidak dapat kembali lagi ke sifat
– sifatnya sebelum gelatinasi. Bahan yang telah kering tersebut masih mampu menyerap air
kembali dalam jumlah yang besar (Winarno, 1992).
ENZIM AMILASE
Enzim adalah katalisator sejati. Molekul ini meningkatkan kecepatan reaksi kimia
spesifik, yang tanpa enzim akan berlangsung amat lambat. Enzim tidak dapat mengubah titik
kesetimbangan reaksi yang dikatalisisnya dan enzim juga tidak akan habis dipakai atau diubah
secara permanen. (Lehninger, 1982).
Amilase adalah kelompok enzim yang memiliki kemampuan memutuskan ikatan
glikosida yang terdapat pada senyawa polimer karbohidrat. Hasil molekul amilum ini akan
menjadi monomer-monomer yang lebih sederhana, seperti maltosa, dekstrin dan terutama
molekul glukosa sebagai unit terkecil. Amilase dihasilkan oleh berbagai jenis organisme hidup,
mulai dari tumbuhan, hewan, manusia bahkan pada mikroorganisme seperti bakteri dan fungi.
Kelompok enzim ini memiliki banyak variasi dalam aktivitasnya, sangat spesifik, tergantung
pada sumber organismenya dan tempatnya bekerja (Dessy, 2008).
Pemanfaatan enzim dalam bidang industri harus memperhatikan factor penting yang
sangat mempengaruhi efisiensi dan efektivitas kerja enzim yang digunakan. Faktor yang
mempengaruhi reaksi enzim antara lain konsentrasi enzim, suhu, pH, dan spesifitas enzim
(Hartati et al., 2002: 68-77).
Amilase dapat dikelompokkan menjadi 3 golongan enzim (Winarno, 1986):
1. α-amilase (1,4-α-D-glukan-glukanohidrolase)

Alfa-amilase merupakan enzim ekstraseluler yang menghidrolisis ikatan 1,4-α-D-
glukanohidrolase. Alfa-amilase dibentuk oleh berbagai bakteri dan fungi. Aktifitas α-amilase
ditentukan dengan mengukur hasil degradasi pati, biasanya dari penurunan kadar pati yang larut
atau kadar dekstrinnya dengan menggunakan substrat jenuh. Hilangnya substrat dapat diukur
dengan pengurangan derajat pewarnaan iodium (Winarno, 1986).
Pati yang mengandung amilosa bereaksi dengan iodium menghasilkan warna biru,
sedangkan dekstrin bila bereaksi dengan iodium berwarna coklat. Keaktifan α-amilase juga
dinyatakan dengan pengukuran viskositas dan jumlah produksi yang terbentuk. Laju hidrolisis
akan meningkat bila tingkat polimerisasi menurun dan laju hidrolisis akan lebih cepat pada rantai
lurus (Winarno, 1986).
2. β-amilase (1,4-α-D-glukan maltohidrolase)
Beta-amilase merupakan exoenzim yang memotong amilum menjadi gugus-gugus
maltose. Enzim ini ditemukan pada tanaman tingkat tinggi dan mikroorganisme (Siti, 1995: 7).
Enzim β-amilase memecah ikatan glukosida α-1,4 pada pati dan glikogen yang terjadi secara
bertahap dari arah luar atau ujung rantai gula yang bukan pereduksi, karena pemotongannya dari
arah luar maka enzim ini disebut eksoamilase (Winarno, 1986).
3. γ-amilase (Glukoamilase)
Glukoamilase merupakan enzim yang memotong rantai pati secara acak menjadi
molekul-molekul glukosa. Hasil reaksinya hanya glukosa, sehingga dapat dibedakan dengan α
dan β amilase. Dengan pengaruh enzim glukoamilase posisi glukosa α dapat diubah menjadi β,
pH optimal 4-5 dan suhu optimal 50-60 0C (Winarno, 1986).
Amilase dapat dibedakan menjadi 3 golongan enzim :
• α- Amilase yaitu enzim yang memecah pati secara acak dari tengah atau bagian dalam molekul.
• β- Amilase yaitu enzim yang memecah unit-unit gula dari molekul pati.
• Glukoamilase yaitu Enzim yang dapat memisahkan glukosa dari terminal gulanon pereduksi
substrat.

Enzim α-amilase adalah salah satu enzim pemecah pati, Enzim α-amilase menghidrolisis
ikatan alpha 1,4 glikosida baik pada amilosa maupun amilopektin secara acak. Karena pengaruh
aktifitasnya, pati terputus-putus menjadi dekstrin dengan rantai sepanjang 6-10 unit glukosa. Jika
waktu reaksi diperpanjang, dekstrin tersebut dapat dipotong-potong lagi menjadi campuran
antara glukosa, maltosa, dan ikatan lain yang lebih panjang. Hidrolisis amilosa oleh α-amilase
terjadi melalui dua tahap. Tahap pertama adalah degradasi amilosa menjadi maltosa dan
maltotriosa yang terjadi secara acak, sangat cepat dan diikuti dengan penurunan viskositas.
Tahap kedua merupakan proses degradasi yang relatif lebih lambat yaitu pembentukan glukosa
dan maltosa sebagai hasil akhir, dimulai dari ujung pereduksi secara teratur (Winarno ,1983).
Penggunaan α-amilase dalam proses hidrolisa pati sering juga disebut likuifikasi, karena
adanya penurunan viskositas dengan cepat, dan kecepatannya dapat bervariasi untuk berbagai
substrat. Dalam hidrolisa pati, α-amilase menghasilkan dekstrin yang merupakan substrat untuk
tahap selanjutnya, yaitu bagi enzim glukoamilase sehingga dengan mudah enzim ini
mengkatalisis hidrolisa untuk menghasilkan glukosa (Winarno ,1983).
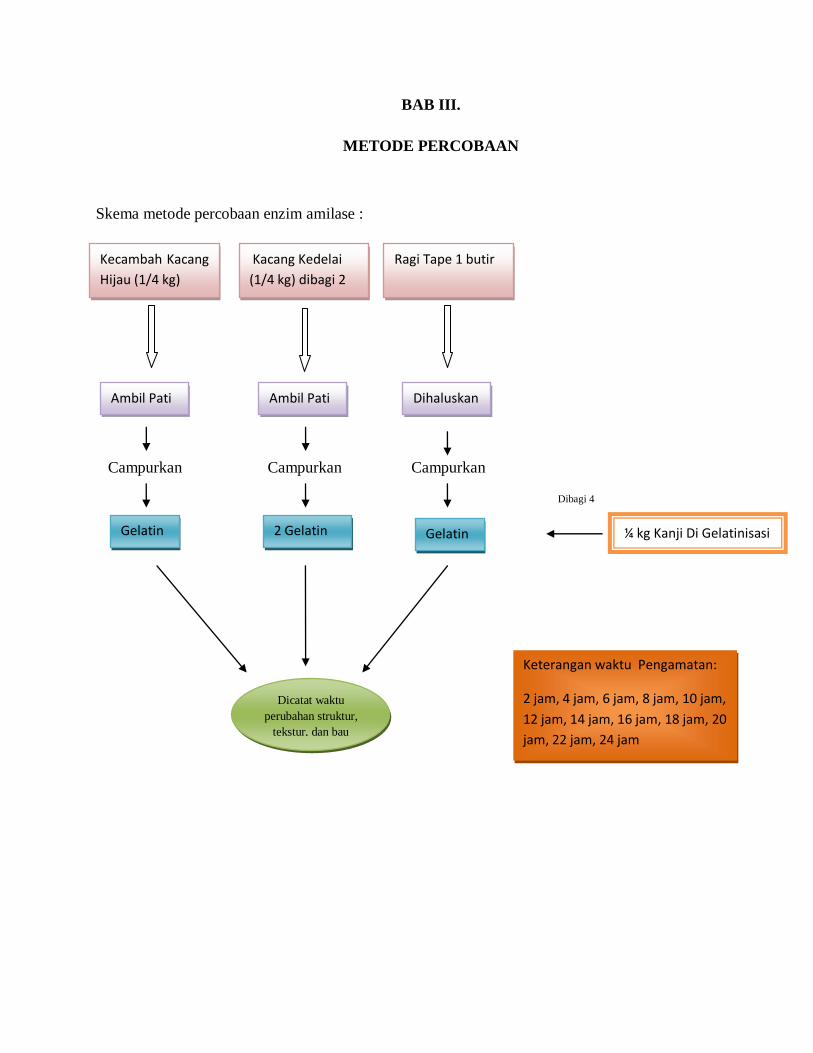
BAB III.
METODE PERCOBAAN
Skema metode percobaan enzim amilase :
Campurkan Campurkan Campurkan
Dibagi 4
Kecambah Kacang
Hijau (1/4 kg)
Kacang Kedelai
(1/4 kg) dibagi 2
Ragi Tape 1 butir
Ambil Pati Ambil Pati Dihaluskan
¼ kg Kanji Di Gelatinisasi Gelatin
n
2 Gelatin Gelatin
n
Dicatat waktu
perubahan struktur,
tekstur, dan bau
gelatin
Keterangan waktu Pengamatan:
2 jam, 4 jam, 6 jam, 8 jam, 10 jam,
12 jam, 14 jam, 16 jam, 18 jam, 20
jam, 22 jam, 24 jam
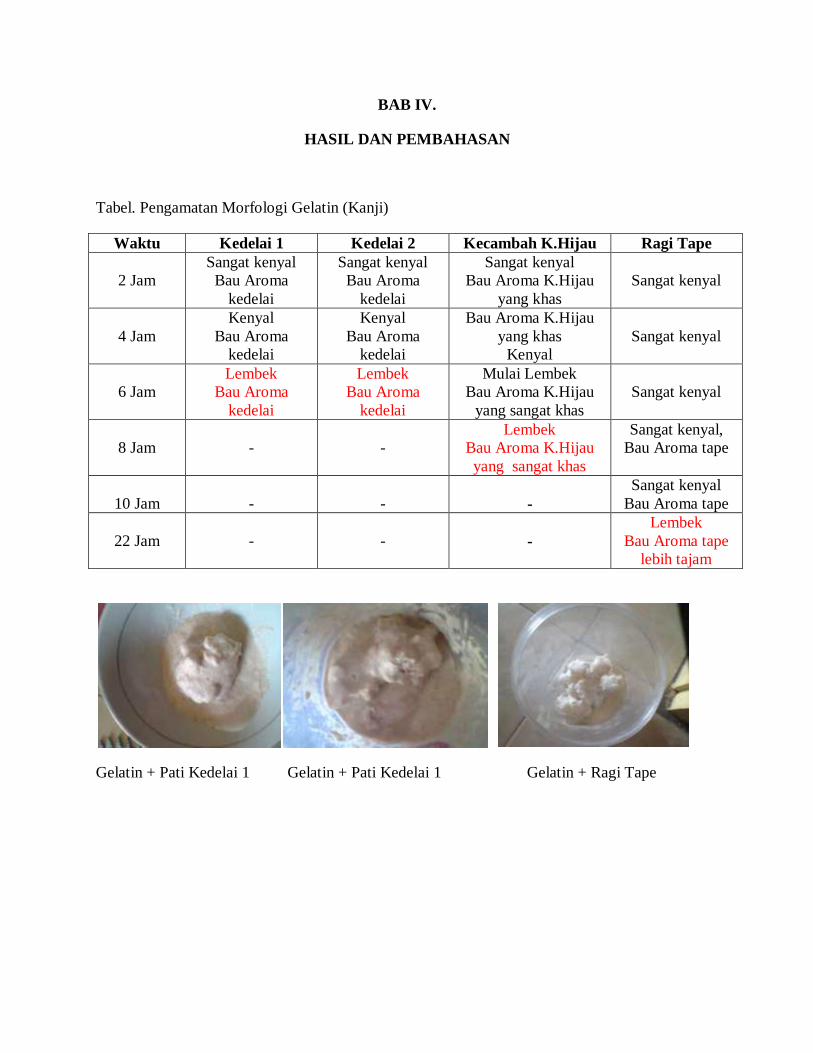
BAB IV.
HASIL DAN PEMBAHASAN
Tabel. Pengamatan Morfologi Gelatin (Kanji)
Waktu Kedelai 1 Kedelai 2 Kecambah K.Hijau Ragi Tape
2 Jam
Sangat kenyal
Bau Aroma
kedelai
Sangat kenyal
Bau Aroma
kedelai
Sangat kenyal
Bau Aroma K.Hijau
yang khas
Sangat kenyal
4 Jam
Kenyal
Bau Aroma
kedelai
Kenyal
Bau Aroma
kedelai
Bau Aroma K.Hijau
yang khas
Kenyal
Sangat kenyal
6 Jam
Lembek
Bau Aroma
kedelai
Lembek
Bau Aroma
kedelai
Mulai Lembek
Bau Aroma K.Hijau
yang sangat khas
Sangat kenyal
8 Jam
-
-
Lembek
Bau Aroma K.Hijau
yang sangat khas
Sangat kenyal,
Bau Aroma tape
10 Jam
-
-
-
Sangat kenyal
Bau Aroma tape
22 Jam
-
-
-
Lembek
Bau Aroma tape
lebih tajam
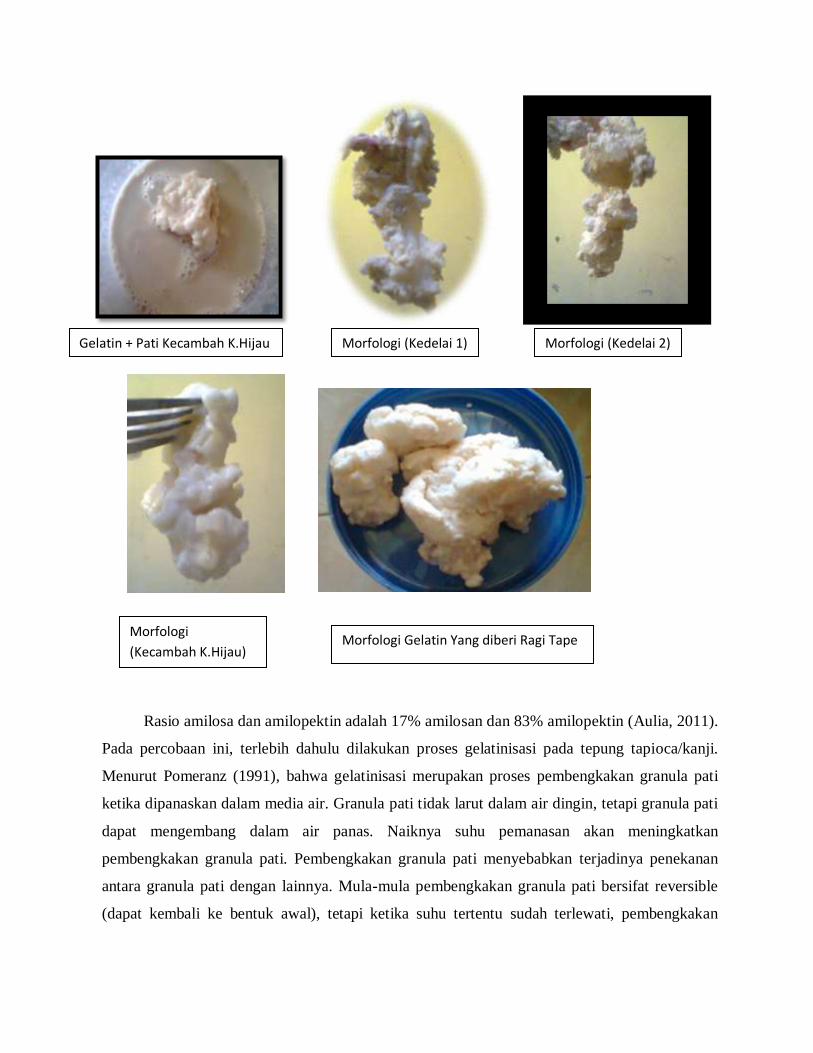
Gelatin + Pati Kedelai 1 Gelatin + Pati Kedelai 1 Gelatin + Ragi Tape
Rasio amilosa dan amilopektin adalah 17% amilosan dan 83% amilopektin (Aulia, 2011).
Pada percobaan ini, terlebih dahulu dilakukan proses gelatinisasi pada tepung tapioca/kanji.
Menurut Pomeranz (1991), bahwa gelatinisasi merupakan proses pembengkakan granula pati
ketika dipanaskan dalam media air. Granula pati tidak larut dalam air dingin, tetapi granula pati
dapat mengembang dalam air panas. Naiknya suhu pemanasan akan meningkatkan
pembengkakan granula pati. Pembengkakan granula pati menyebabkan terjadinya penekanan
antara granula pati dengan lainnya. Mula-mula pembengkakan granula pati bersifat reversible
(dapat kembali ke bentuk awal), tetapi ketika suhu tertentu sudah terlewati, pembengkakan
Gelatin + Pati Kecambah K.Hijau Morfologi (Kedelai 1) Morfologi (Kedelai 2)
Morfologi
(Kecambah K.Hijau) Morfologi Gelatin Yang diberi Ragi Tape

granula pati menjadi irreversible (tidak dapat kembali). Kondisi pembengkakan granula pati
yang bersifat irreversible ini disebut dengan gelatinisasi.
Kanji yang tergelatinisasi tersebut akan diberi 3 perlakuan, yaitu diberi ragi roti, diberi
pati kedelai dan diberi pati kecambah kacang hijau. Sehingga dapat diamati perubahan struktur
morfologi kanji yang tergelatinisasi tersebut.
Hasil yang diperoleh bahwa kanji yang tergelatinisasi yang diberikan pati kacang kedelai
lebih cepat melembeknya dari pada yang diberikan pati kecambah kacang hijau. Pada pati
kacang kedelai, lembeknya kanji yang tergelatinisasi pada jam ke-enam dan pada pati kecambah
kacang hijau, lembeknya kanji yang tergelatinisasi pada jam ke-delapan.
Menurut Kamil (1982), bahwa enzim β-amilase sudah ada sejak semula (pre-exist)
didalam skutelum dan lapisan aleuron pada biji yang masih kering. Enzim β-amilase pada waktu
mulai perkecambahan akan masuk kedalam endosperm untuk merombak amilosa menjadi
glukosa yang bersifat terlarut (water soluble) dan bisa diangkut (translocated). Enzim β-
amilase akan merombak amilopektin menjadi dekstrin yang bersifat tidak bisa diangkut (non
translocated).
Pada biji kacang kedelai, enzim yang berperan adalah enzim β-amilase yang merombak
amilosa menjadi glukosa yang bersifat terlarut (water soluble). Sedangkan pada kacang hijau
dalam bentuk kecambah mengandung enzim α-amilase. Enzim α-amilase dapat memecah pati
secara acak dari tengah atau dari bagian dalam molekul, oleh karena itu disebut endoamilase
(Winarno, 1983). Belum didapatkan teori yang menyatakan apakah kecepatan enzim β-amilase
untuk merombak pati lebih tinggi daripada enzim α-amilase. Namun, percobaan ini
menunjukkan bahwa pada pati kacang kedelai, perombakan struktur amilum kanji tergelatinisasi
lebih cepat daripada pati kecambah kacang hijau. Kedua-dua enzim amilase tersebut, sama-
sama berfungsi untuk merombak pati. Beberapa pustaka mengatakan bahwa, isolasi enzim
amilase banyak dilakukan pada kecambah, yaitu tepatnya mengisolasi enzim α-amilase. Hal ini
dapat didukung oleh pendapat Winarno (1986), yang mengatakan bahwa β- Amilase yaitu
enzim yang memecah unit-unit gula dari molekul pati. Sehingga pada kacang kedelai yang
mengandung enzim β- amilase lebih cepat medegradasi kanji tergelatinisasi.

Pada ragi tape mengandung Saccaharomyces cereviceae. Menurut Judoamidjojo dkk,
(1992), bahwa Saccharomyces cerevisiae menghasilkan enzim zimase dan invertase. Enzim
zimase berfungsi sebagai pemecah sukrosa menjadi monosakarida (glukosa dan fruktosa). Hasil
percobaan menunjukkan bahwa perubahan struktur kanji tergelatinisasi menjadi lembek pada
jam ke-22. Hal ini tentunya berbeda dari enzim yang dihasilkan oleh pati kacang kedelai dan
pati kecambah kacang hijau. Saccaharomyces cereviceae terlebih dahulu melakukan
katabolisme tingkat substrat, kemudian katabolisme tingkat sel sehingga pada jam ke-22, baru
dapat merombak struktur kanji tergelatinisasi.











