12. Reproduksi
29
BAB XII. REPRODUKSI SEKSUAL TUMBUHAN TINGGI Reproduksi dilakukan oleh makhluk hidup, termasuk tumbuhan sebagai usaha mempertahankan kelestarian jenisnya dari kepunahan. Tumbuhan tinggi, melalui biji berkembang biak dan melestarikan jenisnya. Bagaimana proses reproduksi yang dijalani tumbuhan khsusunya tumbuhan berbiji akan diuraikan dalam bab ini. 12.1. BAGIAN-BAGIAN BUNGA Tidak semua bunga memiliki bagian-bagian bunga yang lengkap, seperti kelopak (sepal), mahkota (petal), benang sari (stamen), dan putik (carpel). Bunga yang memiliki keempat organ tersebut disebut bunga lengkap (complete flower), dan yang tidak memiliki salah satunya disebut bunga tak lengkap (uncomplete flower). Bunga sempurna (perfect flower) adalah bunga yang memiliki putik (carpel) dan benang sari (stamen), dan bunga tidak sempurna adalah bunga tak lengkap yang hanya memiliki putik atau benang sari saja.
-
Upload
syarif-hidayat-amrullah -
Category
Documents
-
view
62 -
download
1
Transcript of 12. Reproduksi

BAB XII. REPRODUKSI SEKSUAL TUMBUHAN TINGGI
Reproduksi dilakukan oleh makhluk hidup, termasuk tumbuhan sebagai usaha
mempertahankan kelestarian jenisnya dari kepunahan. Tumbuhan tinggi, melalui biji
berkembang biak dan melestarikan jenisnya. Bagaimana proses reproduksi yang dijalani
tumbuhan khsusunya tumbuhan berbiji akan diuraikan dalam bab ini.
12.1. BAGIAN-BAGIAN BUNGA
Tidak semua bunga memiliki bagian-bagian bunga yang lengkap, seperti kelopak (sepal),
mahkota (petal), benang sari (stamen), dan putik (carpel). Bunga yang memiliki keempat
organ tersebut disebut bunga lengkap (complete flower), dan yang tidak memiliki salah
satunya disebut bunga tak lengkap (uncomplete flower). Bunga sempurna (perfect flower)
adalah bunga yang memiliki putik (carpel) dan benang sari (stamen), dan bunga tidak
sempurna adalah bunga tak lengkap yang hanya memiliki putik atau benang sari saja.
Gambar 12.1. Bagian-bagian bunga
12.2. PEMBENTUKAN GAMET
Proses reproduktif dimulai dengan pembentukan dan perkembangan gamet jantan
dan gamet betina. Di bawah ini tidak akan dibicarakan secara rinci bagaimana gamet jantan
dan betina itu dibentuk dalam bunga, tetapi perlu diketahui bahwa gamet jantan dihasilkan

dalam anthera (kotak sari) sedangkan gamet betina dihasilkan dalam ovarium. Banyak
tumbuhan membentuk baik anthera maupun karpel secara bersama-sama pada setiap
maristem bunga, sehingga menghasilkan bunga yang disebut bunga sempurna atau bunga
hermafrodit atau bunga yang biseksual. Beberapa tumbuhan menghasilkan anthera saja
(bunga staminat) atau hanya karpel saja (monociuos) menghasilkan kedua jenis bunga
tersebut pada satu pohon, sedangkan tumbuhan yang berumah dua (dioeciuos) hanya
menghasilkan satu jenis bunga saja pada satu pohon.
Kenyataan bahwa stamen selalu berkembang lebih dulu dari tengah-tengah
meristem dari pada karpel, diduga akibat perbedaan kadar zat pengatur tumbuh atau
hormon yang diturunkan dari atau diarahkan pada tengah-tengah meristem, sangat penting
dalam mengontrol ekspresi seks dalam bunga. Pemberian auksin baik alami maupun sintetik
yang disemprotkan pada tumbuhan monoecious, telah meningkatkan proporsi bunga betina.
Auksin telah menyebabkan perkembangan bunga betina meskipun primordianya secara
genetik dipersiapkan untuk bunga jantan sedangkan giberelin memberikan pengaruh yang
berlawanan. Sekali perkembangan ovarium telah dimulai, maka perkembangan stamennya
akan dihambat. Pada tumbuhan berumah dua, perubahan seks ini mungkin saja terjadi.
Apabila bunga sudah menghasilkan gamet jantandan gamet betina, maka penyerbukan atau
polinasi dapat terjadi yang selanjutnya hasil fusi gamet jantan dan betina ini akan
menghasilkan zigot.
a. Gamet betina
Di dalam putik berlangsung pembentukan sel telur (ovum). Sel induk (megasporosit)
membelah secara meiosis menghasilkan 4 sel haploid, kemudian 3 sel berdegenerasi dan 1
sel menjadi megaspora. Selanjutnya megaspora membelah secra mtosis 3 kali
menghasilkan 8 sel baru. Kedelapan sel tersebut menjadi bagian dari bakal buah, terdiri
dari: 3 sel menuju kutub kalaza dan membentuk antipoda, 2 sel melebur membentuk
kandung lembaga sekunder, 2 sel berada di kutub mikrofil menjadi sinergid, dan 1 sel
menjadi ovum (lihat gambar 12.2)

b. Gamet jantan
Serbuk saru berasal dari mikrosporosit (sel diplid) yang berkembang di dalam kotak serbuk
sari. Mikrosporosit membelah secara meiosis menghasilkan 4 mikrospora haplid, kemudian
masing-masing membelah scara mitosis menghasilkan butiran serbuk sari, yaitu gametofit
jantan yang belum dewasa yang terdiri dari sel generatif dan sel vegetatif (pembuluh). (lihat
gambar 12.2)
Gambar 12.2. Gametogenesis

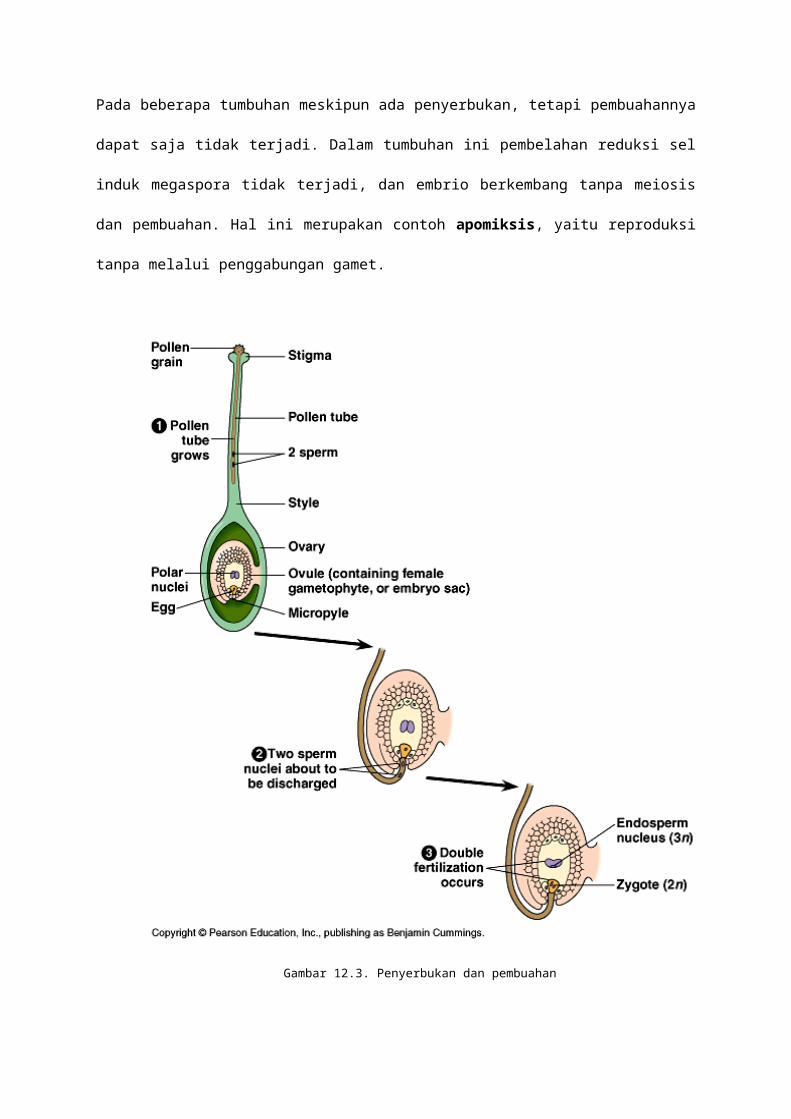
12.3. PENYERBUKAN DAN PEMBUAHAN
Pada tumbuhan, peristiwa pembuahan selalu dilalui oleh peristiwa penyerbukan, walaupun
tidak selalu diikuti oleh peristiwa pembuahan. Pembuahan yang terjadi pada tumbuhan tidak
hanya peleburan sperma dengan ovum, tetapi sperma juga membuahi inti kandung lembaga
sekunder yang menghasilkan endopermae. Dengan demikian pembuahan pada tumbuhan
(angiospermae) bersifat ganda.
Penyerbukan adalah peristiwa jatuhnya serbuk sari ke kepala putik dengan melalui berbagai
bantuan; seperti angin (anemogami), air (hidrogami), burung (ornithogami), serangga
(entomogami), bahkan termasuk oleh manusia (antropogami).
Setiap serbuk sari dapat melakukan penyerbukan terhadap bunga lain. Oleh karena itu ada
beberapa cara penyerbukan: autonogami (serbuk sari berasal dari bunga itu sendiri),
geitonogami (serbuk sari berasal dari bunga lain tapi satu individu), allogami (serbuk ari
bunga individu lain), dan bastar (serbuk sari berasal dari bunga individu lain yang berbeda
jenis).
Bila penyerbukan berhasil, maka akan diiukti dengan proses pembuahan yang berlangsung
di dalam ruang arkegonium (bakal buah) yang membungkus bakal biji (ovule).
Setelah spermatozoa dan ovum matang, maka dilanjutkan dengan proses pembuahan yang
diawali dengan penyerbukan. Keberadaan zat gula pada kepala putik menyebabkan serbuk
sari tumbuh membentuk buluh serbuk sari. Pembuahan berlangsung 2 (dua) kali, pertama
spermatozoa 1 membuahi ovum menghasilkan zigot (2n), kedua spermatozoa 2 membuahi
kandung lembaga sekunder menghasilkan endospermae (3n). Perhatikan gambar 12.3.
Pada beberapa tumbuhan meskipun ada penyerbukan, tetapi pembuahannya dapat saja
tidak terjadi. Dalam tumbuhan ini pembelahan reduksi sel induk megaspora tidak terjadi, dan
embrio berkembang tanpa meiosis dan pembuahan. Hal ini merupakan contoh apomiksis,
yaitu reproduksi tanpa melalui penggabungan gamet.

Gambar 12.3. Penyerbukan dan pembuahan
12.4. PERKEMBANGAN EMBRIO
(1). Kapasitas Tumbuh
Zigot merupakan sel pertama dari generasi sporofit yang memiliki potensi maksimal
untuk melakukan pertumbuhan, karena dia mampu untuk tumbuh menjadi organisme baru
yang utuh. Setelah satu kali pembelahan, setiap sel anak akan berkurang kecenderungan
morfogeniknya, karena setiap sel anak hanya mampu menghasilkan bagian organisme
secara terbatas dan sudah mengarah kepada pembentukan morfologi tertentu. Meskipun

demikian apabila kedua sel anak tadi dipisahkan, setiap sel akan memperoleh kembali
kapasitasnya untuk tumbuh menjadi suatu organisme. Berkurangnya kapasitas kedua sel
anak yang berasal dari zigot tersebut disebabkan setiap sel anak sekarang tidak
terpisahkan, tetapi dibawah pengaruh sel-sel lain yang berdekatan yang berhubungan
dngannya melalui hubungan protoplasma (plasmodesmata). Dengan bertambahnya
kompleksitas organisme, kapasitas yang berkaitan dengan pertumbuhan dari setiap sel akan
berkurang sejalan dengan pengaruh semua sel dan jaringan yang mengitarinya.
(2). Pertumbuhan embrio
Tidak lama setelah terbentuk, zigot mulai tumbuh dengan cepat dan berkembang
menjadi suatu embrio, yang selanjutnya akan beristirahat sampai biji berkecambah.
Perkembangan endosperm mendahului pertumbuhan embrio, dan endosperm ini
merupakan jaringan yang akan memberi makan embrio selama perkembangannya.
Pola pertumbuhan embrio berbeda dari satu tumbuhan ke tumbuhan lain, tetapi
penjelasan secara umum akan sangat membantu kita. Permulaan pembelahan sel zigot
akan menghasilkan dua sel, satu diantaranya akan berkembang menjadi embrio dan yang
lain akan menjadi suspensor. Suspensor mengatur pelekatan dan orientasi embrio serta
mendorongnya ke dalam masa endosperm yang akan memberinya makanan.
Perkembangan embrio melalui tahap-tahap globular, jantan dan torpedo, dikatakan
demikian karena penampilannya memperlihatkan bentuk-bentuk tersebut. Dua lekukan pada
tahap jantung akan berkembang menjadi kotiledon, dan pada tahap kotiledon embrio telah
mengembangkan radikula atau meristem akar dan meristem ”shoot” (tunas pucuk) (lihat
gambar 12.4)
Gambar. 12.4. Tahap pertumbuhan dan perkembangan embrio
Dengan tumbuhnya embrio, maka endosperm akan tercerna karena digunakan untuk
memberi makan embrio. Proses ini berlanjut tanpa istirahat sampai tidak ada endosperm
yang tersisa, dan semua bahan-bahan sisa dipindahkan ke kotiledon seperti halnya pada biji
kacang. Alternatif lain seperti pada jagung, endosperm tetap tinggal dalam biji sampai
perkecambahan terjadi (cermati gambar 12.6).
(3). Pertumbuhan Embrio In Vitro
Pada tahun 1930, P.White seorang ahli fisiologi Amerika telah berhasil
mengembangkan suatu media untuk mengkultur embrio yang berukuran sebesar awal tahap
jantung (heart stage) di luar induknya. Akhir-akhir ini diketahui bahwa endosperm
kemungkinan besar merupakan sumber nutrien yang terbaik untuk pertumbuhan embrio,
dan penambahan air kelapa ke dalam medium telah memungkinkan embrio yang masih
pada tahap awal perkembangannya dikultur dan berhasil. Masalah yang timbul dengan
pemakaian air kelapa dalam medium adalah komposisi air kelapa yang sangat kompleks
dan sebagian besar tidak diketahui. Air kelapa kaya akan nitrogen, gula, berbagai macam
zat tumbuh dan hormon serta banyak lagi senyawa-senyawa lainnya dengan aktivitas
fisiologi yang bervariasi.
(5). Embriogenesis dalam Kultur Sel dan Kultur Jaringan
Pada tahun 1950, laboratorium Steward memproleh kemajuan pesat dalam usaha
mengkultur jaringan dan sel dari beberapa tanaman terutama wortel, dengan menggunakan
medium basal White ditambah dengan air kelapa. Kemudian dengan cara memindahkannya
ke dalam medium yang tepat, baik padat maupun cair, sel-sel tersebut secara normal
berkembang menghasilkan embrio, membentuk akar dan ”shoot” dan akhirnya menjadi
tumbuhan dewasa yang lengkap dan berbunga.
Jelaslah bahwa sel-sel yang berasal dari berbagai sumber pada tanaman, apabila
diperlukan dan diberi kesempatan yang baik dan tepat, dapat mengubah dirinya dari
keterbatasan-keterbatasan dalam lingkungan induknya kepada sifat-sifat zigot yang khas,

dan dapat melaksanakan embriogenesis atau semacamnya, menghasilkan suatu tanaman
yang utuh. Sel-sel yang berasal dari bagian tumbuhan yang dewasa dan terdiferensiasi
(batang, akar dan lain-lain), responnya sangat terbatas dan memerlukan perlakuan yang
khusus. Kapasitasyang dimiliki oleh zigot akan tetap ada dalam sel dan tidak akan hilang
sebagai akibat pembelahan. Dalam perkembangan suatu organisme, kecenderungan
morfogenetik dari sel-sel akan ditutup, dan akan dibuka kembali apabila sel-sel manjadi
bebas dari jaringan dan kultur dalam medium yang tepat. Hormon dan zat tumbuh lainnya
yang seimbang, dan nutrisi yang tepat, perlu mendapat perhatian dalam usaha menginduksi
dan memelihara perkembangan sel secara teratur.
Gambar 12.5 . Pengklonan wortel dalam tabung reaksi. Percobaan ini menunjukkan bahwa tanaman dapat diregenerasi dari sel somatik (sel non reproduktif). Tanaman baru yang dihasilkan merupakan turunan (duplikat) genetik dari tanaman induknya.
(6). Totipotensi Sel Tumbuhan
Sel-sel somatik dari berbagai bagian tumbuhana, apabila diberi kondisi yang baik dan tepat,
dpat berkembagng menjadi suatu tumbuhan baru yang utuh. Hal ini disebut totipoyensi,
karena, karena mereka memiliki potensi penuh untuk berkembang sa ma seperti yang
dimiliki oleh zigot, M eskipun demikian tidak semua sel tumbuhan dapat melakukan
perrnudaan kembali (rejuvenation) melalui cara ini, karena beberapa dari mereka telah

terpatok pada satu pola yang peristiwanya tidak dapat kembali (irreversible), akibat
terjadinya pengendapan bahan-baha dinding sel yang padat dan tidak larut. Meskipun
demikian semua jaringan tumbuhan mengandung sel-sel yang totipoten. Hal di atas pada
dasarnya merupakan perwujudan yang menunjukkan bahwa setiap sel hidup pada
tumbuhan di dalamnya membawa informasi genetik yang lengkap, yang pwnting untuk
menghasilkan satu tumbuhan. Hilangnya kemampuan sel dalam proses perkembangan,
bukan disebabkan hilangnya informasi dari sel tersebut, tetapi berkurangnya kemampuan
untuk memanfaatkan informasi. Jadi setiap sel sebenarnya memiliki kapasitas penuh untuk
tumbuh atau suatu potensi morfogenetik seperti yang dimiliki oleh zigot (perhatikan kembali
gambar 12.6), tetapi dalam proses perkembangannya kapasitas untuk tumbuh harus
dikontrol demi keterturan dan keserasian.
12.5. PEMBENTUKAN BUAH DAN BIJI
Perkembangan buah dan biji dimulai setelah peneyerbukan terjadi. Apabila tidak terjadi
penyerbukan, maka bunga akan cepat tua dan mati. Pada tumbuha apomistik, rangsangan
penyerbukan saja sudah cukup untuk memulai perkembangan embrio, dan diduga bahwa
polen telah menyediakan zat tumbuh atau hormon yan gmerangsang perkembangan
embrio. Menyemprotkan auksin pada bnga yang tidak diserbuk, akan menghasilkan buah
yang partenokarpik (tanpa biji). Pada beberapa tumbuhan, terutama pada spesies yang
berbiji banyak (stony fruit), asam giberelat bertindak sebagai pengganti auksin. Dalam
spesies-spesies ini diketahui bahwa giberelin, dan bukan auksin.
(1) Perkembangan Buah dan Biji
Tahap awal dalam perkembangan buahah dan biji adalah pembelahan sel yang cepat tanpa
banyak mengalami perbesaran. Faktor utamanya karena terlibatnya sitokinin yang banayak
diproduksi oleh endsperm yang triploid (atau pentapolid). Berbagai jaringan dari tumbuhan
seperti ovarium, dasar bunga dan kadang-kadang bagian dari tabung bunga, dapat terlibat
dalam pembentukan buah.
Setelah.tahap pembelahan sel, fase pertumbuhan berikutnya terutama dilakukan dengan
perbesaran sel. Dari beberapa penelitian diudga bahwa hal ini disebakan oleh auksin yang
dihasilkan di dalam biji. Apabila biji-biji tersebut dikeluarkan dari buah yang sedang
berkembang, perkembangan buah tersebut akan terhenti, akan tetapi perkembangan dapat
diteruskan apabila diberi auksin. Pada strawbery dan ketimun, perkembangan tergantung

pada auksin yang berasal dari ovulus, namun beberapa buah-buahn lebih memberikan
respon terhadap giberelin daripadaterhadap auksin.
Struktur Biji
Dalam tahap akir perkembangannya, biji akan mengalami dehidrasi sapai kandungan airnya
hanya sekitar 5% hingga 15% dari bobotnya. Embrio itu akan berhenti berkembang saampai
biji berkecambah. Embrio dikelilingi oleh kotiledonnya yang sudah membesar, oleh
endospermae, atau oleh keduanya. Embrio dan persediaan makanannya terbungkus oleh
suatu selaput biji (seed coat) yang terbentuk dari integumen bakal biji, nenek moyang biji.
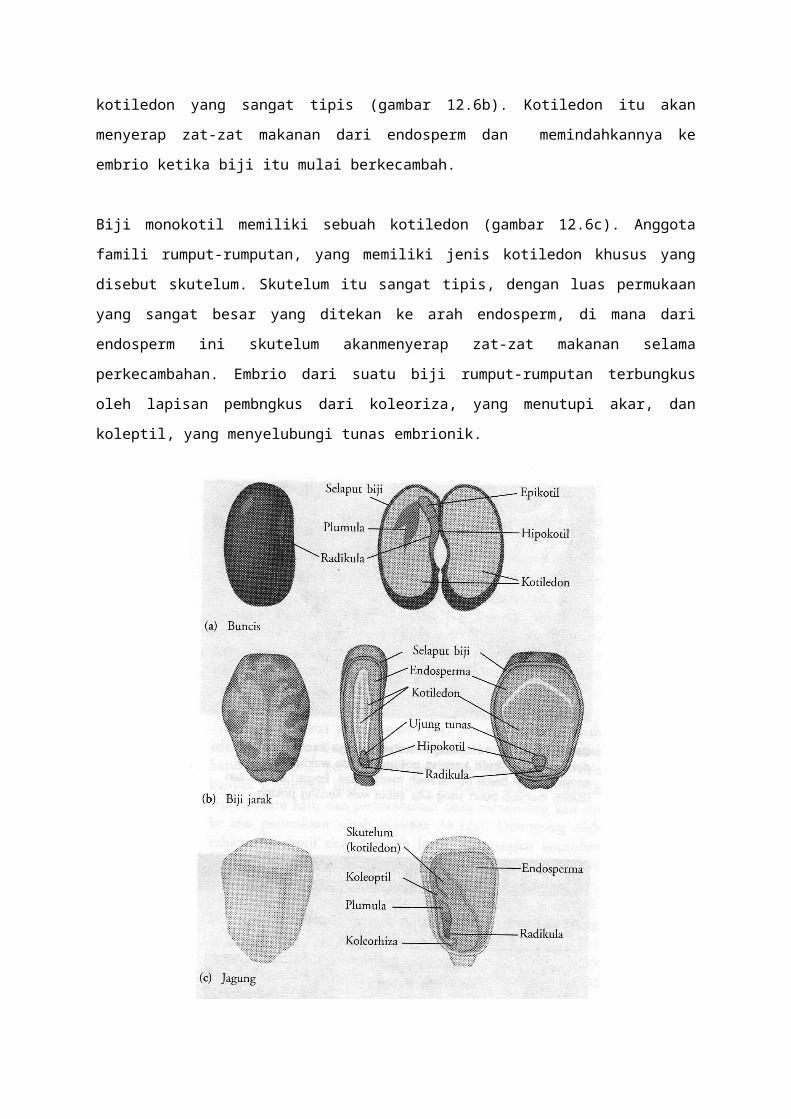
Kita dapat melihat lebih dekat jenis biji-dikotil dengan cara membuka biji kacang (gambar
12.6a). Pada tahapan ini, embrio merupakan suatu struktur memanjang, sumbu
embrioniknya, bertaut pada kotiledon berdaging. Di bawah titik dimana kotiledon bertaut,
sumbu embrionik itu disebut hipokotil. Hipokotil berakhir pada radikula (radicle), atau akar
embrionik. Bagian sumbu embrionik di atas kotiledon adalah hipokotil. Pada ujungnya
terdapat plumula, yang terdiri dari dari ujung tunas dengan sepasang daun miniatur.
Kotiledon kacang berdaging sebelum biji itu berkecambah karena kotiledon menyerap
makanan dari endosperm ketika biji berkembang. Namun demikian, biji beberapa dikotil,
seperti biji jarak mempertahankan persediaan makanannya dalam endosperm dan memiliki
kotiledon yang sangat tipis (gambar 12.6b). Kotiledon itu akan menyerap zat-zat makanan
dari endosperm dan memindahkannya ke embrio ketika biji itu mulai berkecambah.
Biji monokotil memiliki sebuah kotiledon (gambar 12.6c). Anggota famili rumput-rumputan,
yang memiliki jenis kotiledon khusus yang disebut skutelum. Skutelum itu sangat tipis,
dengan luas permukaan yang sangat besar yang ditekan ke arah endosperm, di mana dari
endosperm ini skutelum akanmenyerap zat-zat makanan selama perkecambahan. Embrio
dari suatu biji rumput-rumputan terbungkus oleh lapisan pembngkus dari koleoriza, yang
menutupi akar, dan koleptil, yang menyelubungi tunas embrionik.

Gambar 12.6. struktur biji. (a) Kotiledon berdaging pada buncis, suatu dikotil, menyiman makanan yang
diserap dari endosperm ketika biji itu berkembang. (b) Biji jarak memiliki kotileoden bermembran yang
akan menyerap makanan dari endosperm ketika biji itu berkecambah. (c) Jagung, suatumonokotil, hanya
memiliki satu kotiledon (skul=telum jagung dan rumputan lain). Tunas yang belum sempurna dibungkus
dalam suatu struktur yang disebut koleoptil.
(2) Pematangan Buah
Proses pematangan buah banyak melibatkan perubahan kimia d an fisiologi yang kompleks
yang m enyangkut rasa, ukuran, warna, tekstur dan aroma. Pada proses pematangan buah
dapat terjadi konversi asam dan pati menjadi gula bebas, peningkatan pektinase yang akan
melunakkan dan merusak dinding sel, peningkatan berbagai macam pigmen, seperti
antosianin, serta hilangnya pigmen klorofil. Banyak dari perubahan-perubahan dirangsang
atau diakibatkan oleh etilen yang dihasilkan oleh buah itu sendiri.
Produksi etilen oleh buah telah diketemukan pada saat penyimpanan. Etilen yang
diproduksi oleh setiap buah, telah memberikan efek kumulatif dan merangsang buah-buah

lain untuk matang lebih cepat dan secara tidak sengaja akan memproduksi lebih banyak
etilen. Prose pematangan dalam penyimpanan buah secara massal, akan mengakibatkan
proses “autokatalitik”, yang dapat berdampak merugikan. Penyimpanan ditempat dingin atau
mengalirkan gas inert (nitrogen atau CO2) akan menghambat pematangan buah.
Perubahan fisiologi yang terjadi selama proses pematangan adalah terjadinya klimakterik
dalam respirasi (respiratory climacteric). Banayak buah-buahan melakukan klimakterik
dengan cara menginduksinya dengan penambahan etilen. Klimakterik biasanya diikuti
dengan masa hidup yang pendek dan produksi etilen yang meningkat secara dramatis.
Etilen diperkirakan mempengaruhi proses pematangan buah dengan meliabtkan klimakterik
dalam repsirasi melalui dua cara. Pngaruh etilen terhadap permeabilitas membran
mengakibatkan permeabilitas sel meningkat besar sekali selama proses pematangan. Hal
tersebut di atas mengakibatkan pelunakan buah dan bercampur baurnya metabolit dengan
enzim, sehingga metabolisme respirasi dipercepat.
Selama kliakterik, kandungan protein meningkat diduga etilen telah merangsang sintesis
protein pada saat tersebut. Protein yang dibentuk ini barangkali terlibat dalam proses
pematangan, dan klimakterik marupakan gambaran tentang adanya peningkatan enzim-
enzim respirasi. Dalam beberapa peristiwa, klimakterik merupakan proses yang aerobik
sehingga prosesnya dapat dicegah atau diperlambat dengan cara mengurangi tekanan
oksigen pada penyimapananya.
12.6 PERKECAMBAHAN
Bagi banyak orang perkecambahan sebuahbiji menandakan permulaan kehidupan,
akan tetapi pada kenyatannya biji itu sudah menganudng tumbuhan ukuran miniatur,
lengkap dengan akar dan tunas embrionik. Perkecambahan biji tumbuhan tidak
memulai kehidupan akan tetapi meneruskan pertumbuhan dan perkembangan yang
secara temporer dihentikan ketika biji menjadi dewasa dan embrionya menjadi tidak
aktif (dorman).
Biji akan menjadi dewasa dalam buah. Setelah buah matang dan bijinya dikeluarkan,
biasanya biji dalam keadaan dorman untuk waktu lama atau pendek saja. Hal ini
berarti bahwa meskipun biji tersebut mendapat cukup air dan diberi kondisi yang
baik untk perkecambahan, biji tersebut tetap tidak akan berkecambah. Dormansi
dapat diakibatkan oleh terbentuknya senyawa-senyawa kimia penghambat pada
permukaan biji, kurangnya zat-zat perangsang yang penting atau disebabkan oleh

kulit biji yang keras sehingga air dan oksigen tidak masuk. Dormansi ini dapat hilang
dengan berbagai macam cara seperti dengan melakukan pembekuan,
memperpaanjang perioda pendinginan, memperpanjang pendedahan pada keadaan
dingin, memberikan kelembaban yang tinggi dengan adanaya oksigen, melakukan
pemanasan secaran intensif (dibakar), melalui usus burung atau mamalia, dilakukan
secara mekanik (ampelas, dipecah), atau dibiarkan ditumbuhi jamur.
Apabila kondisi yang diperlukan untuk menghilangkan dormansi ini berjala, embrio
akan mulai membuat giberelin dan sitokinin, yang diperlukan untuk mengungguli
efek kerja penghambat pertumbuhan (growth inhibitor), sehingga pertumbuhan pun
dapat dimulai. Kalau pada saat tersebut air diberikan, maka bij pun akan
berkecambah.
(1) Dormansi Biji
Dorman artinya “tidur”atau berisitirahat. Biji dorman memiliki laju metabolisme yang
sangat lambat dan sedang tidak bertumbuh dan berkembang.
Dormansi pada biji meningkatkan peluang bahwa perkecaambahan akan terjadi
pada waktu dan tempat yang paling menguntungkan bagi pertumbuhan biji.
Pematahan priode dormansi umumnya memerlukan kondisi lingkungan yang
tertentu. Biji tumbuhan gurun, misalnya, hanya berkecambah setelah curah hujan
memadai. Jika mereka harus berkecambah setelah hujan rintik-rintik yang sedang,
tanah mungkin akan terlalu cepat kering sehingga tidak dapat mendukung
pertumbuhan biji. Di tempat di mana kebakaran alamiha biasa terjadi, banyak biji
memerlukan panas yang sangat tinggi untuk mengakhiri dormansi; dengan demikian
pertumbuhan biji menjadi paling berlimpah setelah api menghanguskan vegetasi
yang menjadi saingannya tersebut. Di tempat di mana musim dingin sangat parah,
biji mungkin memerlukan pemaparan terhadap cuaca dingin yang lebih lama; biji
yang disemaikan selama musim panas atau musim gugur tidak akan berkecambah
sampai semi berikutnya. Hal ini akan memastikan musim tumbuh yang panjang
sebelum musim dingin berikutnya. Biji yang sangat kecil, seperti beberapa biji dari
varietas lettuce, memerlukan cahaya untuk perkecambahan dan akan mengakhiri
dormansinya hanya jika ditanam cukup dangkal sehingga kecambah benih bisa
muncul menembus permukaan tanah. Beberapa biji harus dilemahkan dengan
senyawa-senyawa kimia ketika biji-biji tersebut melewati saluran pencernaan hewan

dan akibatnya cenderung akan terbawa hingga jarak yang cukup jauh sebelum
berkecambah.
Lama waktu di mana biji dorman masih hidup dan mampu berkecaambah bervariasi
dari beberapa hari hingga beberapa dekade atau bahkan lebih lama lagi, bergantung
pada spesies dan kondisi lingkungan. Sebagian besar biji sangat tahan lama
sehngga bisa tahan selama satu atau dua tahun sampai kondisi lingkungan
memiungkinkan untuk berkecambah. Dengan demikian, tanah memiliki kumpulan biji
yang belum berkecambah yang kemungkinan telah menumpuk selama beberapa
tahun. Hal ini merupakan salah satu mengapa vegetasi bisa muncul kembali
sedemikian cepatnya setelah kejadian kebakaran, kekeringan, banjir, atau beberapa
bencana alam lainnya.
(2) Proses Perkecambahan Biji
Perkecambahan biji tergantung pada imbibisi, penyerapan air akibat potensial air
yang rendah pada biji yang kering. Air yang berimbibisi mengakibatkan biji
mengembang dan memecahkan kulit pembungkusnya dan juga memicu perubahan
metabolik pada embrio yang menyebabkan biji tersebut melanjutkan pertumbuhan
(gambar 12.7). Enzim-enzim akan mulai mencerna bahan-bahan yang disimpan
pada endosperm atau kotiledon, dan nutrien-nutriennya dipindahkan ke bagian
embrio yang sedang tumbuh. Mobilisasi cadangan makanan ini telah dipelajari
paling banyak pada biji barley dan rumput-rumputan lainnya.
Gambar 12.7. Mobilisasi zat-zat makanan selama perkembahan
Organ pertama yang muncul dari biji yang berkecambah adalah radikula, yaitu akar
embrionik. Berikutnya, ujung tunas harus menembus permukaan tanah. Pada
kacang dan banyak tumbuhan dikotil alinnya, hipokotil akan berbentuk seperti suatu
kait, dan pertumbuhan akan mendorong kait itu ke atas permukaan tanah (gambar
12.8a). Dirangsang oleh cahaya, hipokotil akan tumbuh lurus, mengangkat kotiledon
dan epikotil. Dengan demikian, ujung tunas yang lembut dan kotiledon yang sangat
besar itu ditarik ke atas permukaan tanah yang abrasif. Sekarang epikotil
menyebarkan helai daun pertamanya, yang mengembang, menjadi hijau, dan mulai
membuat makanan melalui fotosintesis. Kotiledon akan layu dan rontok dari biji
karena cadangan makanan telah dihabiskan oleh embrio yang berkecambah itu.
Cahaya merupakan petunjuk utama yang memberitahu benih bahwa ia telah
menembus tanah. Kita dapat menipu biji kacang sehngga biji tersebut bertingkallaku
seolah-olah ia masih tetap terkubur dengan cara mengecambahkan biji dalam
kegelapan.Biji yang tidak diterangi memperpanjang hipokotil yang berlebihan
dengan suatu kait pada ujungnya, dan helai daun tidak mampu berubah menjadi
warna hijau. Setalah biji kehabisan cadangan makanannya, biji yang berbentuk
gelondongan berhenti tumbuh dan kemusian mati.
Kacang polong, meskipun berada dalam famili yang sama dengan buncis, memiliki
gaya perkecambahan yang berbeda (gambar 12.8b). Suatu kait terbentuk pada

epikotil bukan pada hipokotil, dan ujung tunasnya terangkat secara lembut dari tanah
akibat pemanjangan epikotil dan pelurusan kait. Kotiledon kacang polong, berbeda
dengan kotiledon buncis, tetap berada di bawah tanah.
Jagung dan rumput-rumputan lainnya, yang merupakan monokotil, menggunakan
metode yang berbeda untuk menembus tanah ketika mereka berkecambah (12.8c).
Koleptil, yaitu lapisan yang membungkus dan melindungi tunas embrionik,
mendesak naik ke atas melalui tanah menuju udara. Ujung tunasnya kemudian
tumbuh lurus ke atas melalui saluran atau terowongan yang disediakan oleh koleotil
tubuler.
Gambar 12.8. Perkecamabahan biji. Radikula, akar embrio, pertama kali muncul dari biji. Kemudian
tunas akan memecah permukaan tanah melalui salah satu mekanisme berikut ini. (a) Pada buncis,
pelurusan suatu kait pada hipokotil akan menarik tunasan kotiledon dari tanah. (b) Pada polong, kait
berada di atas kotiledon pada epikotil, dan kotiledon itu tetap berada di dalam tanah. (c) pada jagung
dan rumput-rumputan lain, tunas tumbuh ke atas dan lurus melalui salurankoleoptil.
-----


The events of "double fertilization" of the egg and polar nuclei by the two sperm cells. Image from Purves et al., Life: The Science of Biology, 4th Edition, by Sinauer Associates (http://www.sinauer.com/) and WH Freeman (http://www.whfreeman.com/),
STUDENT OBJECTIVES
1. Explain why, although they are very different from each other, both male and female reproductive systems produce gametes.
2. Describe the location, structure, and functions of the scrotum and testes.3. Explain the principal events of spermatogenesis.4. Describe the location, structure, and functions of the seminiferous tubules, straight tubules, rete
testis, ductus epididymis (within the epididymis), ductus (vas) deferens (as a component of the spermatic cord), ejaculatory duct, and urethra as ducts of the male reproductive system.
5. Describe the location, structure, and functions of the accessory sex glands: seminal vesicles, prostate gland, and bulbourethral (Cowper’s) glands and explain the characteristics of semen.
6. Explain the structure and functions of the penis.7. Describe the location, structure, and functions of the ovaries.8. Explain the principal events of oogenesis.9. Explain the location, structure, and functions of the uterine (Fallopian) tubes, uterus and vagina.10. Describe the components of the vulva and explain their functions.11. Explain the structure and functions of the mammary glands.12. Describe the principal events of the ovarian and uterine cycles.13. Compare the various kinds of birth control methods and their effectiveness.14. Describe the development of the male and female reproductive systems.15. Describe the effects of aging on the reproductive systems.

Coconut fruit containing the seed you usually buy at the grocery store. Image from Purves et al., Life: The Science of Biology, 4th Edition, by Sinauer Associates (http://www.sinauer.com/) and WH Freeman (http://www.whfreeman.com/), used with permission.
Fruits
Fruits are a development of the ovary wall and sometimes other flower parts as well. As seeds mature, they release the hormone auxin, which stimulates the wall of the ovary to develop into the fruit. In fact, commercial fruit growers may stimulate fruit development in nonpollinated flowers by applying synthetic auxin to the flower.
Fruits promote the dispersal of their content of seeds in a variety of ways.
Wind. The maple "key" and dandelion parachute are examples.
Water. Many aquatic angiosperms and shore dwellers (e.g., the coconut palm) have floating fruits that are carried by water currents to new locations.
Hitchhikers. The cocklebur and sticktights achieve dispersal of their seeds by sticking to the coat (or clothing) of a passing animal.
Edible fruits. Nuts and berries entice animals to eat them. Buried and forgotten (nuts) or passing through their g.i. tract unharmed (berries), the seeds may end up some distance away from the parent plant.
Mechanical. Some fruits, as they dry, open explosively expelling their seeds. The pods of many legumes (e.g., wisteria) do this.

Structure of two dicot seeds. Image from Purves et al., Life: The Science of Biology, 4th Edition, by Sinauer
Associates (http://www.sinauer.com/) and WH Freeman (http://www.whfreeman.com/








![[PPT]Teknologi Reproduksi - Ekadasa Drestanta Wibisana ... · Web viewTeknologi Reproduksi Pengertian Teknologi Reproduksi Teknologi reproduksi adalah ilmu reproduksi atau ilmu tentang](https://static.fdokumen.com/doc/165x107/5af3b53d7f8b9a4d4d8c8743/pptteknologi-reproduksi-ekadasa-drestanta-wibisana-viewteknologi-reproduksi.jpg)