






![Exposure to benzo[a]pyrene of Hepatic Cytochrome P450 Reductase Null (HRN) and P450 Reductase Conditional Null (RCN) mice: Detection of benzo[a]pyrene diol epoxide-DNA adducts by immunohistochemistry](https://static.fdokumen.com/doc/165x107/63259f17c9c7f5721c022d3b/exposure-to-benzoapyrene-of-hepatic-cytochrome-p450-reductase-null-hrn-and-p450.jpg)
Bahasa
Halaman
Hukum


(This is a sample cover image for this issue. The actual cover is not yet available at this time.)
This article appeared in a journal published by Elsevier. The attached
copy is furnished to the author for internal non-commercial research
and education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling or
licensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of the
article (e.g. in Word or Tex form) to their personal website or
institutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies are
encouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Toxicology Letters 213 (2012) 381– 391
Contents lists available at SciVerse ScienceDirect
Toxicology Letters
jou rn al h om epage: www.elsev ier .com/ locate / tox le t
Unexpected contribution of cytochrome P450 enzymes CYP11B2 and CYP21,
as well as CYP3A4 in xenobiotic androgen elimination – Insights from
metandienone metabolism
Maria Kristina Parr a,b,∗, Andy Zöllner c,d, Gregor Fußhöllerb, Georg Opfermannb, Nils Schlörer e,Mirela Zorio f, Matthias Bureik c, Wilhelm Schänzerb
a Freie Universität Berlin, Institute of Pharmacy, Pharmaceutical Chemistry, Berlin, Germanyb Center for Preventive Doping Research, Institute of Biochemistry, German Sport University Cologne, Cologne, Germanyc PomBioTech GmbH, D-66125 Saarbrücken, Germanyd Dunn Labortechnik GmbH, Asbach, Germanye Institute of Organic Chemistry, University of Cologne, Germanyf Doping Control Laboratory, National Anti-Doping Agency, Bucharest, Romania
h i g h l i g h t s
◮ Investigation of metandienone hydroxylation by cytochrome P450 enzymes.◮ CYP21 and CYP3A4 catalyze hydroxylation of normetandienone (NorMD).◮ CYP21, CYP3A4, and CYP11B2 take part in generation of 20�OH-NorMD from metandienone.◮ Unexpectedly, non-liver CYPs are involved in metandienone long-term metabolism.
a r t i c l e i n f o
Article history:
Received 4 May 2012
Received in revised form 23 July 2012
Accepted 24 July 2012
Available online xxx
Keywords:
Metandienone
Metabolic pathways
Cytochrome P450
Doping control
GC–MS/MS
NMR
a b s t r a c t
The metabolism of a variety of anabolic steroids frequently misused for doping purposes has been
investigated in the last years. This research mainly focused on main and long-term metabolites suit-
able for detection, but detailed clearance mechanisms have rarely been elucidated. Recent studies on
metandienone focused on the identification of 17�-hydroxymethyl-17�-methyl-18-norandrosta-1,4,13-
trien-3-one (20�OH-NorMD) as long-term metabolite, however, the metabolic pathway of its generation
remained unclear.
Metandienone and its Wagner–Meerwein rearrangement product 17,17-dimethyl-18-norandrosta-
1,4,13-trien-3-one (NorMD) were hydroxylated by different human cytochrome P450 enzymes (CYPs).
Some of their hydroxylation products were chemically synthesized and characterized by mass spectrom-
etry to allow for their trace detection in urine samples. Following oral administration of metandienone
or NorMD in one human volunteer each the post administration urines were checked for the presence of
those hydroxylated metabolites using GC–MS/MS analysis.
The human mitochondrial steroid hydroxylating enzymes CYP11B1 and CYP11B2 were capable
to metabolize metandienone leading to the formation of 11�-hydroxymetandienone and 18-
hydroxymetandienone. Following Wagner–Meerwein rearrangement, the resulting products could be
assigned to 20�OH-NorMD and 11�OH-NorMD. The contribution of CYP11B1 and CYP11B2 in human
metabolism of metandienone was confirmed by analysis of post-administration samples of metandienone
and NorMD. Combined with the results from a previous study, enzymatic pathways were identified that
involve CYP21 and CYP3A4 in the hydroxylation of NorMD, while CYP21, CYP3A4 and CYP11B2 take part
in 20�OH-NorMD generation from MD.
The current study represents a valuable contribution to the elucidation of clearance mechanisms of
anabolic steroids and also indicates that mainly non-liver CYPs seem to be involved in these processes
© 2012 Elsevier Ireland Ltd. All rights reserved.
∗ Corresponding author at: Institute of Pharmacy, Pharmaceutical Chemistry, Freie Universität Berlin, Königin-Luise-Str. 2+4, 14195 Berlin, Germany.
Tel.: +49 30 838 57686; fax: +49 30 838 56906.
E-mail address: [email protected] (M.K. Parr).
0378-4274/$ – see front matter © 2012 Elsevier Ireland Ltd. All rights reserved.
http://dx.doi.org/10.1016/j.toxlet.2012.07.020

Author's personal copy
382 M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391
1. Introduction
Cytochromes P450 (CYPs) form a remarkably versatile enzyme
superfamily that has been found in all kingdoms of life and already
includes more than 12,000 named members (Nelson, 2011). The
human CYP superfamily comprises 57 members that by sequence
homology are divided into 18 families and 43 subfamilies. How-
ever, they may also be categorized according to their subcellular
localization, their electron transport chains or their biological func-
tions (Guengerich, 2005; Hannemann et al., 2006). With respect to
the latter aspect, a distinction has often been made between those
CYPs that have physiological substrates and those that do not, but
collectively act as a protective system against foreign compounds.
However, it is becoming increasingly clear that this differentia-
tion is not a strict one (Guengerich et al., 2011). A prominent
group of CYPs that are known to fulfill important physiologi-
cal functions by acting on endogenous substrates are the steroid
hydroxylases. In humans, there are six of these enzymes (Miller
and Auchus, 2011): CYP11A1 (cytochrome P450scc, side chain
cleavage enzyme), CYP17A1 (CYP17, cytochrome P45017�, steroid
17�-hydroxylase and 17, 20-lyase), CYP21A2 (CYP21, cytochrome
P450c21, steroid 21-hydroxylase), CYP11B1 (cytochrome P45011�,
steroid 11�-hydroxylase), CYP11B2 (cytochrome P450aldo, aldos-
terone synthase) and CYP19A1 (cytochrome P450aro, aromatase).
It is consistent with the view expressed above that more and more
xenobiotics are found to be their substrates, with synthetic steroids
being natural candidates.
Metandienone (MD [1]) is a synthetic anabolic steroid that is
prohibited in sports by the World Anti-Doping Agency (WADA)
(World Anti-Doping Agency, 2010a). Even if known since years
it has gained attention of anti-doping researchers in the last
couple of years since an increasing number of professional
athletes were convicted of using this substance for doping
purposes (World Anti-Doping Agency, 2010b). However, also
recreational and junior athletes and even the broad public are
attracted by anabolic steroids, especially MD (Basaria, 2010).
Commonly used methods to detect its abuse are based on the
identification of different urinary MD metabolites via liquid chro-
matography tandem mass spectrometry (LC–MS/MS) and gas
chromatography (tandem) mass spectrometry (GC–MS(/MS)) anal-
ysis (Schänzer et al., 1991, 1992a, 1995, 1996, 1999, 2006). Since
MD is a 17�-hydroxy-17�-methyl steroid, the analysis of its
metabolism is complicated by the fact that metabolites are not
only the products of enzymatic activity but may also result from
Wagner–Meerwein rearrangement or 17-epimerization: upon
17�-sulfatation, the decomposition of the 17�-sulfate can lead
to six new configurations (four different dehydration products
with 17,17-dimethyl-18-norandrosta-1,4,13-trien-3-one (NorMD
[2]) being the most abundant, a 13-hydroxy-17,17-dimethyl
steroid, and a 17-epimer, respectively) (Schänzer et al., 1992a).
Recently, we could demonstrate that both CYP3A4 and CYP21 are
able to catalyze the regio- and stereospecific hydroxylation of
NorMD [2] to 17�-hydroxymethyl-17�-methyl-18-norandrosta-
1,4,13-trien-3-one (20�OH-NorMD [3]) (Zöllner et al., 2010). By
the name of “nightwatch”, this compound was introduced to
the anti-doping community by Schänzer et al. (2006) as a long-
term metabolite of MD. While CYP3A4 is known to metabolize
a number of both endogenous and exogenous steroids includ-
ing MD (Rendic et al., 1999), the participation of CYP21 in
this context was new as only two other substrates (proges-
terone and 17�-hydroxyprogesterone) were previously known
for this enzyme (Wu et al., 1991). Interestingly, each of the
two CYPs produced a second NorMD metabolite that was not
formed by the other enzyme: CYP3A4 catalyzed the forma-
tion of 17�-hydroxymethyl-17�-methyl-18-norandrosta-1,4,13-
triene-3-one (20�OH-NorMD [4]), a compound that has been
previously detected in urine samples (Parr et al., 2007; Schänzer
et al., 2006); CYP21 produced 16�-hydroxy-17,17-dimethyl-
18-norandrosta-1,4,13-trien-3-one (16�OH-NorMD [5]), a new
metabolite whose in vivo occurrence was not yet clear. It was the
aim of this study to further investigate the metabolic pathways
that lead from MD [1] to 20�OH-NorMD [3] and to identify the
enzymes that participate in it. For this purpose, biotransforma-
tion of MD with recombinant human steroid hydroxylases was
performed and revealed the production of new MD metabolites.
Subsequent analysis of metabolite formation in administration
studies was conducted to test for their presence in vivo. Based on
these data, we suggest an expanded MD metabolization scheme
that explains the formation of the newly observed metabolites by a
combination of steroid hydroxylations and/or Wagner–Meerwein
rearrangement.
2. Experimental procedures
2.1. Chemicals and reagents
Metandienone (MD, 17�-hydroxy-17�-methylandrosta-1,4-dien-3-one [1]),
methyltestosterone (17�-hydroxy-17�-methylandrost-4-en-3-one), androst-
4-ene-3,17-dione, 16�-hydroxyandrost-4-ene-3,17-dione, 11�-hydroxyandrost-
4-ene-3,17-dione, pyrrolidine, methyl magnesium bromide (3 M in diethyl ether),
lithium aluminum hydride and 2,3-dichloro-5,6-dicyano-p-benzoquinone (DDQ)
were purchased from Sigma–Aldrich GmbH (Steinheim, Germany). Epimetendiol
(EMD, 17�-methyl-5�-androst-1-en-3-3�, 17�-diol), 18-norepimetenediol
(NorEMD, 17,17-dimethyl-18-nor-5�-androsta-1,13-dien-3�-ol), 5�-tetrahydro-
methyltestosterone (5�-THMT, 17�-methyl-5�-androstane-3�,17�-diol),
5�-tetrahydromethyltestosterone (5�-THMT, 17�-methyl-5�-androstane-
3�,17�-diol) and epi-metandienone (epiMD, 17�-hydroxy-17�-methylandrosta-
1,4-dien-3-one) as well as d3-testosterone, d3-epitestosterone, d4-etiocholanolone,
and d5-androsterone-glucuronide were chemically synthesized in our labora-
tory (Schänzer et al., 1991, 1992a, 1996; Schänzer and Donike, 1993, 1995),
17�-hydroxymethyl-17�-methyl-18-norandrosta-1,4,13-trien-3-one (20�OH-
NorMD [3]), and 16�-hydroxy-17,17-dimethyl-18-norandrosta-1,4,13-trien-3-one
(16�OH-NorMD [5]) were generated as described by Zöllner et al. (2010).
�-Glucuronidase from Escherichia coli (>140 U/mL) was obtained from Roche Diag-
nostics (Mannheim, Germany) and N-methyl-N-trimethylsilyl-trifluoroacetamide
(MSTFA) from Chem. Fabrik Karl Bucher (Waldstetten, Germany). All other reagents
and solvents were of analytical grade and obtained from VWR International
(Darmstadt, Germany). Recombinantly expressed and purified bovine adrenodoxin
reductase (AdR (Sagara et al., 1993)) and adrenodoxin (Adx (Uhlmann et al., 1992))
as well as human CYP11B1 (Zöllner et al., 2008) and CYP11B2 for initial in vitro
studies were kindly provided by Rita Bernhardt (Dept of. Biochemistry, Saarland
University, Saarbrücken, Germany).
2.2. Chemical synthesis of reference steroids
2.2.1. 17,17-Dimethyl-18-norandrosta-1,4,13-trien-3-one [2]
17,17-Dimethyl-18-norandrosta-1,4,13-trien-3-one (NorMD [2]) was chemi-
cally synthesized as displayed in Scheme 1: 1 g of MD [1] was dissolved in 40 mL of a
mixture of methanol: 1 N hydrochloric acid (1:1) and refluxed for 16 h. After cooling
the product was extracted three times with 50 mL of n-pentane from the solution.
2.2.2. 17˛-Hydroxymethyl-17ˇ-methyl-18-norandrosta-1,4,13-trien-3-one [4]
The synthesis of 17�-hydroxymethyl-17�-methyl-18-norandrosta-1,4,13-
trien-3-one (20�OH-NorMD [4]) was performed as displayed in Scheme 2 starting
with 10 g of androst-4-ene-3,17-dione [I]. The formation of androstenedione
cyanohydrine [IIa and IIb] was achieved by reaction with 16 g of sodium cyanide
in 80 mL of methanol and 10 mL of glacial acetic acid as described by Nitta et al.
(1985). As they reported 17�-cyano-17�-hydroxyandrost-4-en-3-one [IIb] precip-
itates from the mixture under the conditions applied. After hydrolysis in 100 mL of
refluxing hydrochloric acid (6 M) within 24 h the by-products were extracted three
times at pH 12 with 200 mL of t-butyl methyl ether (TBME), and the purified product
was then extracted from the remainder three times with 200 mL of TBME at pH 1.
The obtained 17�-hydroxy-3-oxoandrost-4-ene-17�-carboxylic acid [III] (4.5 g)
O
OH
CH3
1N HC l
MeOH (1:1)
O
CH3
CH3
[2][1]
Scheme 1. Wagner–Meerwein rearrangement of metandienone [1], yielding
NorMD [2].

Author's personal copy
M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391 383
NaCN
AcOH/MeOH
O
CN
OH
6N HCl
O
COOH
OH
LiAlH4
EtOEt
HO
CH2OH
OH
DDQ
dioxa ne
O
CH2OH
OH
1N HCl
MeOH (1 :1)
O
CH3
CH2OH
O
O
O
OH
CN
[IIb ][IIa]
[4]
[I]
]VI[]III[
[7]
Scheme 2. Chemical synthesis of 20�OH-NorMD [4].
was reduced with 5.6 g of lithium aluminum hydride in 100 mL of absolute diethyl
ether resulting in two isomers of 17�-hydroxymethyl-androst-4-ene-3�,17�-diol
[IV]. Dehydrogenation with 3.7 g of DDQ in 60 mL of dioxane under argon atmo-
sphere resulted in 17�-hydroxy-17�-hydroxymethyl-androsta-1,4-dien-3-one
(20-hydroxymetandienone, 20OH-epiMD [7]) within 4 h. After Wagner–Meerwein
rearrangement by overnight refluxing in 130 mL of a mixture of methanol: 1 N
hydrochloric acid (1:1) 880 mg of raw product were obtained. Purification by column
chromatography on silica gel with n-hexane: ethyl acetate (7:3) and crystallization
from n-hexane: ethyl acetate (2:1) yielded 87 mg of 20�OH-NorMD [4].
2.2.3. 11ˇ-Hydroxymetandienone [8]
As displayed in Scheme 3, 11�-hydroxyandrost-4-ene-3,17-dione ([V], 30 mg)
was dissolved in 260 �L of methanol under reflux and 14 �L of pyrrolidine were
added. As described by Gould et al. (1957) after 5 min of refluxing crystals
of 11�-hydroxy-3-pyrrolidino-3,5-androsta-3,5-dien-17-one [VI] were collected,
washed with 200 �L of cold methanol and dried in a desiccator over night.
Their solution in tetrahydrofurane (THF) was added to 3 mL of methyl magne-
sium bromide (3 M in diethyl ether) and stirred for 3 h at room temperature.
After addition of 3 mL of aqueous hydrochloric acid (3.3 M) the raw 11�-
hydroxymethyltestosterone (11�,17�-dihydroxy-17�-methylandrost-4-en-3-one,
[VII]) was extracted from the mixture with TBME. 1,2-Dehydrogenation was
achieved overnight with 13 mg of DDQ in 0.5 mL of boiling dioxane resulting in 11�-
hydroxymetandienone (11�,17�-dihydroxy-17�-methylandrosta-1,4-dien-3-one,
11�OH-MD [8]). Wagner–Meerwein rearrangement (in refluxing methanol: 1 N
hydrochloric acid (1:1)) yielded 11�-hydroxy-17,17-dimethyl-18-norandrosta-
1,4,13-trien-3-one (11�OH-NorMD [6]).
2.2.4. 16˛-Hydroxy-(epi-)metandienone [9] and [10]
In analogy to 11�OH-MD [8], for the synthesis of 16�-hydroxy-(epi-
)metandienone (16�,17�-dihydroxy-17�-methylandrosta-1,4-dien-3-one,
16�OH-epiMD [9] and 16�,17�-dihydroxy-17�-methylandrosta-1,4-dien-3-one,
16�OH-MD [10], Scheme 3) 23.4 mg of 16�-hydroxyandrost-4-ene-3,17-dione
[VIII] were dissolved in 200 �L of boiling methanol and 11 �L of pyrrolidine
were added. After 10 min of refluxing crystals of 16�-hydroxy-3-pyrrolidino-3,5-
androsta-3,5-dien-17-one [IX] appeared. They were collected and washed with
200 �L of cold methanol. After drying in a desiccator they were dissolved in 2 mL of
THF and added to 3 mL of a solution of methyl magnesium bromide (3 M in diethyl
ether). After 3 h at ambient temperature 2 mL of water and 1 mL of concentrated
aqueous hydrochloric acid were carefully added and the mixture was extracted four
times with 5 mL of TBME each. The combined extracts were evaporated to dryness.
The raw products [X] and [XI] were dissolved in 0.5 mL of dioxane and refluxed
for 16 h with 35 mg of DDQ under argon atmosphere to yield 16�OH-epiMD [9]
and 16�OH-MD [10]. By Wagner–Meerwein rearrangement as described above
16�-hydroxy-17,17-dimethyl-18-norandrosta-1,4,13-trien-3-one (16�OH-NorMD
[14]) is obtained.
2.2.5. Recombinant fission yeast strains and whole-cell biotransformations
In this study, the fission yeast strains CAD67 (co-expresses the human
cytochrome P450 reductase (CPR) and CYP3A4 (Dragan et al., 2011)), CAD71
(co-expresses CPR and CYP17 (Zöllner et al., 2010)), CAD75 (co-expresses CPR and
CYP21 (Zöllner et al., 2010)), and MB164 (expresses CYP11B2 (Bureik et al., 2002b))
were used. Recombinant fission yeast strains were cultivated at 30 ◦C in Edinburgh
minimal medium (EMM). Approximately 2 × 108 cells/mL were used per assay.
Incubation was carried out either in 96-well microtiter plates or in 50 mL reaction
tubes at a shaking velocity of 150 rpm for 24 h. The assay volume was either 1 mL
(microtiter plates) or 10 mL (50 mL reaction tubes). The substrate concentration
was always 250 �M. After the incubation, the steroids were extracted by addition
of equal volumes of ethyl acetate, dried under nitrogen flow and subsequently
analyzed by GC–MS as described below.
2.3. Administration studies
Post administration (p.a.) samples of MD [1] and NorMD [2] were ana-
lyzed with special respect to the intermediates and by-products of metabolic
hydroxylation. An administration study of MD ([1], 25 mg, p.o.) was performed in
one healthy male volunteer (54 years, 75 kg) and urine samples were collected for
four weeks. Additionally 2 mg of NorMD [2] were taken by a healthy male volun-
teer (58 years, 78 kg) and urine samples were collected for two weeks. Approval
was obtained from local ethical committees. The samples were prepared accord-
ing to the routine steroid screening procedure used in our laboratory (Geyer et al.,
1998). In brief, after addition of 40 �l of the internal standard solution contain-
ing methyltestosterone, d3-testosterone, d3-epitestosterone, d4-etiocholanolone
and d5-androsterone-glucuronide 2 mL of urine were incubated at pH 7 with �-
glucuronidase from E. coli at 50 ◦C for 1 h. The steroids were extracted with 5 mL
of TBME at pH 9.6, the organic layer was evaporated to dryness and analyzed by
GC–MS/MS. The urinary elimination of the metabolites of interest was investigated
and the kinetics was monitored.
2.4. Instrumentation
2.4.1. NMR spectroscopy
The nuclear magnetic resonance (NMR) analyses were performed at 600 MHz
(1H NMR) and 150 MHz (13C NMR) at 298K on a Bruker (Rheinstetten, Germany)
Avance II 600 instrument equipped with a 5 mm inverse probe head with actively
shielded z-gradient coil. Chemical shifts are reported in ı values (ppm) relative to
tetramethylsilane. Solutions of about 5 mg of each compound in deuterated chlo-
roform were used for conducting 1H; H,H COSY; APT; H,C HMQC; H,C HMBC and
NOESY experiments.
2.4.2. GC–MS(/MS) analyses
For GC–MS(/MS) analyses the final residues of sample preparation were deriva-
tized with TMIS reagent (MSTFA/NH4I/ethanethiol, 1000:2:3, v:w:v) by heating for
20 min at 60 ◦C as introduced by Donike (1969) and Geyer et al. (1998).
GC–MS was performed on an Agilent (Agilent Technologies, Waldbronn,
Germany) 6890 N gas chromatograph coupled to an Agilent 5973 mass selective
detector (MSD) applying the following parameters: column: Agilent HP5MS (17 m;
0.25 mm i.d.; 0.25 �m film thickness), carrier gas: helium, head pressure: 1 bar,
oven temperature program: 0 min 100 ◦C, +40 ◦C/min, 0 min 190 ◦C, +5 ◦C/min, 0 min
245 ◦C, +40 ◦C/min, 3 min 320 ◦C, injection volume: 2 �L, splitless, injection temper-
ature: 300 ◦C, ionization: 70 eV, EI, full scan mode, 40–800 Da.
The GC–MS/MS analyses were either performed on a trace GC ultra gas chro-
matograph (Thermo Fisher, Bremen, Germany) coupled to a TSQ Quantum GC triple
quadrupole mass spectrometer (Thermo Fisher, Bremen, Germany) or on an Agi-
lent 7890A gas chromatograph coupled to an Agilent 7000B triple quadrupole MSD

Author's personal copy
384 M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391
O
O
R1=OH, R2= H : [V] R1= H , R2=OH : [V III]
R1
R2
N
O
R1
R2pyrr olidi ne
MeOH
R1=OH, R2= H : [VI] R1= H , R2=OH : [IX]
Me-Mg-Br
THF/EtO Et
R1=OH, R2= H : [VII ]
R1= H , R2=OH : [XI]+[X] *
O
OH
R1
R2 DDQ
dioxane
R1=OH, R2= H : [8]
R1= H , R2=OH : [10]+ [9] *
O
OH
R1
R2
Scheme 3. Chemical synthesis of the hydroxylated metandienones 11�OH-MD [8] and 16�OH-MD [10] * In case of 16�-hydroxy both 17-epimers [9] and [10], or [X] and
[XI], respectively, were obtained.
applying the following parameters: column: Agilent Ultra-1 (17 m; 0.20 mm i.d.;
0.1 �m film thickness), carrier gas: helium, constant pressure: 1.14 bar, oven tem-
perature program: 0 min 183 ◦C, +3 ◦C/min, 0 min 232 ◦C, +40 ◦C/min, 2 min 310 ◦C,
injection volume: 1.5 �L, split 1:16, injection temperature: 300 ◦C, ionization: 70 eV,
EI, collision gas: argon, data acquisition: MRM mode. Retention times, molecular ions
and ion transitions of the per-TMS derivatives of the relevant compounds are listed
in Table 1.
3. Results
3.1. Chemical syntheses and characterization of reference
material
NorMD [2] was generated from MD [1] by Wagner–Meerwein
rearrangement (Scheme 1). Structure confirmation of NorMD [2]
was performed by NMR (Table 2). The mass spectra of the per-TMS
derivatives of both compounds, [1] and [2], are displayed in Fig. 1.
20OH-epiMD [7] was synthesized in a four step reaction
from androst-4-ene-3,17-dione [I] (Scheme 2, mass spectrum
in Fig. 2A). Stereospecificity was obtained by selective precip-
itation of the 17�-cyano intermediate [IIb] from the mixture.
Table 1
Retention times (RT), molecular ions (M+) and MRM ion transitions of the per-TMS
derivatives of the steroids monitored (with methyltestosterone as 3,5-dienol-bis-
TMS at RT = 15.04 min).
Analyte RT [min] M+ Ion transition used
for quantitation
MD [1] 14.73 444 206 → 191
NorMD [2] 7.72 354 339 → 133
20�OH-NorMD [3] 12.91 442 236 → 133
20�OH-NorMD [4] 12.48 442 236 → 133
16�OH-NorMD [5] 12.71 442 442 → 337
11�OH-NorMD [6] 10.97 442 442 → 337
20OH-epiMD [7] 17.29 532 532 → 339
11�OH-MD [8] 17.33 532 532 → 206
16�OH-epiMD [9] 17.17 532 532 → 206
16�OH-MD [10] 17.72 532 532 → 206
16�OH-MD [11] 17.86 532 532 → 206
16�OH-epiMD [12] 16.74 532 532 → 206
18OH-MD [13] 17.43 532 532 → 206
16�OH-NorMD [14] 10.75 442 442 → 337
EMD 9.26 448 358 → 301
NorEMD 5.92 358 253 → 185
5�-THMT 12.18 450 270 → 213
5�-THMT 12.40 450 270 → 213
EpiMD 12.84 444 206 → 191
Methyltestosterone (ISTD) 15.04 446 301 → 169
Wagner–Meerwein rearrangement of [7] yielded 20�OH-NorMD
[4]. It was unambiguously identified by NMR (Table 2). NOESY
experiments confirmed the stereochemistry (further details in
supporting information). The mass spectrum of its bis-TMS deriva-
tive is displayed in Fig. 3B.
11�OH-MD [8] was obtained from 11�-hydroxyandrost-4-
ene-3,17-dione [V] by stereoselective Grignard methylation at
C-17 after protection of the oxo-group at C-3, and subsequent
1,2-dehydrogenation (Scheme 3, mass spectrum of its bis-TMS
derivative in Fig. 2B). During the analogues reaction starting from
16�-hydroxyandrost-4-ene-3,17-dione [VIII] both the 17�-methyl
product (16�OH-epiMD, [9]) and the 17�-methyl (16�OH-MD
[10], ∼6% of [9], calculated from the peak area in the chro-
matogram) were obtained (Scheme 3). Structure confirmation was
achieved by NMR analyses ([9]: 1H NMR ı: 1.87 (m, 1H, H-14), 4.11
(dd, 1H, H-16), 0.80 (s, 3H, H-18), 1.21 (s, 3H, H-20), 13C NMR ı:
47.3 (C-13), 47.2 (C-14), 77.2 (C-16), 80.5 (C-17), 16.0 (C-18), 18.9
(C-20), [10]: 1H NMR ı: 1.51 (m, 1H, H-14), 4.17 (dd, 1H, H-16),
0.85 (s, 3H, H-18), 1.24 (s, 3H, H-20), 13C NMR ı: 45.8 (C-13), 47.4
(C-14), 77.1 (C-16), 80.3 (C-17), 15.8 (C-18), 20.5 (C-20), stereo-
chemistry assigned by NOESY). Wagner–Meerwein rearrangement
of both products resulted in the same compound (16�OH-NorMD
[14]). The mass spectra of the per-TMS derivatives of [9], [10] and
[14] are displayed in Figs. 2C and D and 4A. The mass spectra of the
intermediates of syntheses are available as supporting information.
3.2. In vitro biotransformation of MD by human steroid
hydroxylases
3.2.1. MD hydroxylation
Initial experiments with recombinantly expressed enzymes
indicated no activity of CYP17 and CYP21 toward MD [1] (data
not shown), while CYP11B1 and CYP11B2 catalyzed the formation
of two metabolites (Parr et al., 2011): one of these was specific
for CYP11B2 while the other was produced by both enzymes.
To produce larger amounts of both metabolites, whole-cell bio-
transformation with the CYP11B2 expressing recombinant fission
yeast strain MB164 was performed. From the knowledge of the
enzymatic properties and on the basis of the mass spectra of the
tris-TMS derivatives (Fig. 2B and F) the assignment as 11�OH-MD
[8] and 18-hydroxymetandienone (17�,18-dihydroxy-17�-
methylandrosta-1,4-dien-3-one, 18OH-MD [13]) was possible.
Comparison of retention times and ion transitions with those of

Author's personal copy
M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391 385
Table 21H and 13C NMR spectral data of NorMD [2], 20�OH-NorMD [3], 20�OH-NorMD [4], and 16�OH-NorMD [5].
[2] [3] [4] [5]
ıC ıH ıC ıH ıC ıH ıC ıH
1 155.8 7.16 d (10) 155.5 7.16 d (10) 155.5 7.16 d (10) 155.5 7.15 d (10)
2 127.6 6.27 dd (10/2) 127.7 6.29 dd (10/2) 127.7 6.28 dd (10/2) 127.7 6.29 dd (10/2)
3 186.3 – 186.3 – 186.3 – 186.3 –
4 124.3 6.10 d (2) 124.4 6.12 d (2) 124.4 6.11 d (2) 124.4 6.11 d (2)
5 168.9 – 168.6 – 168.7 – 168.6 –
6 33.3 ˛: 2.42 ddd (13/5/5) 33.2 ˛: 2.43 m 33.2 ˛: 2.44 m 33.2 ˛: 2.43 m
ˇ: 2.55 ddd (13/13/5) ˇ: 2.56 ddd (13/13/2) ˇ: 2.58 ddd (13/13/5) ˇ: 2.53 m
7 32.6 ˛: 1.15 dddd (13/13/13/4) 32.5 ˛: 1.19 m 32.7 ˛: 1.17 m 32.4 ˛: 1.17 m
ˇ: 2.19 m ˇ: 2.21 m ˇ: 2.22 m ˇ: 2.17 m
8 36.6 ˇ: 2.36 m 36.6 ˇ: 2.45 m 36.9 ˇ: 2.42 m 36.2 ˇ: 2.33 m
9 49.5 ˛: 1.29 ddd (11/11/2) 49.4 ˛: 1.33 ddd (11/11/2) 49.4 ˛: 1.33 ddd (11/11/2) 49.3 ˛: 1.33 m
10 43.6 – 43.2 – 43.2 – 43.3 –
11 24.0 ˛: 2.06 m 23.9 ˛: 2.09 m 24.0 ˛: 2.08 m 23.7 ˛: 2.07 m
ˇ: 1.55 dddd (12/12/12/5) ˇ: 1.60 m ˇ: 1.57 m ˇ: 1.55 m
12 22.0 ˛: 1.86 m 22.4 ˛: 1.92 m 22.4 ˛: 1.88 m 22.3 ˛: 1.93 m
ˇ: 2.07 m ˇ: 2.07 m ˇ: 2.12 m ˇ: 2.07 m
13 142.2 – 137.5 – 137.1 – 140.7 –
14 134.2 – 138.9 – 139.1 – 131.5 –
15 30.0 ˛: 2.11 m 30.7 ˛: 2.20 m 30.8 ˛: 2.19 m 39.1 ˛: 2.17 m
ˇ: 2.26 m ˇ: 2.35 m ˇ: 2.34 m ˇ: 2.55 m
16 39.4 ˛: 2.19 m 33.8 ˛: 2.02 m 33.9 ˛: 1.98 m 80.3 ˛: 3.95 dd
ˇ: 1.65 m ˇ: 1.59 m ˇ: 1.61 m –
17 45.6 – 51.6 – 51.6 – 47.9 –
18 – – – – – – – –
19 18.5 1.23 s 18.5 1.24 s 18.5 1.24 s 18.5 1.17 s
20 CH3 26.7 0.95 s 21.7 0.95 s 21.7 1.01 s 24.6 0.99 s
26.4 0.99 s 18.8 0.91 s
20 CH2OH – – 68.9 3.37 d (10) 68.9 3.32 d (10) – –
3.51 d (10) 3.40 d (10)
the chemically synthesized reference further identified 11�OH-
MD [8]. By Wagner–Meerwein rearrangement, these newly found
products could be converted to 20�OH-NorMD [3] (from 18OH-MD
[13]) and 11�OH-NorMD [6] (from 11�OH-MD [8]), respectively
(mass spectra in Fig. 3A and D).
3.2.2. Hydroxylation of NorMD
Biotransformation of NorMD [2] with a recombinant fission
yeast strain that co-expresses human CPR and CYP21 (CAD75)
resulted in four different products. The two main products (mass
spectra of their bis-TMS derivatives in Fig. 3A and C) were purified
from the remaining substrate and identified as 20�OH-NorMD [3]
and 16�OH-NorMD [5] as already described by Zöllner et al. (2009,
2010). The NMR signals (Table 2) could be assigned by combining
one and two dimensional experiments and NOE measurement
(details as supporting information). In the present study, two more
minor products were detected. On the basis of the mass spectra
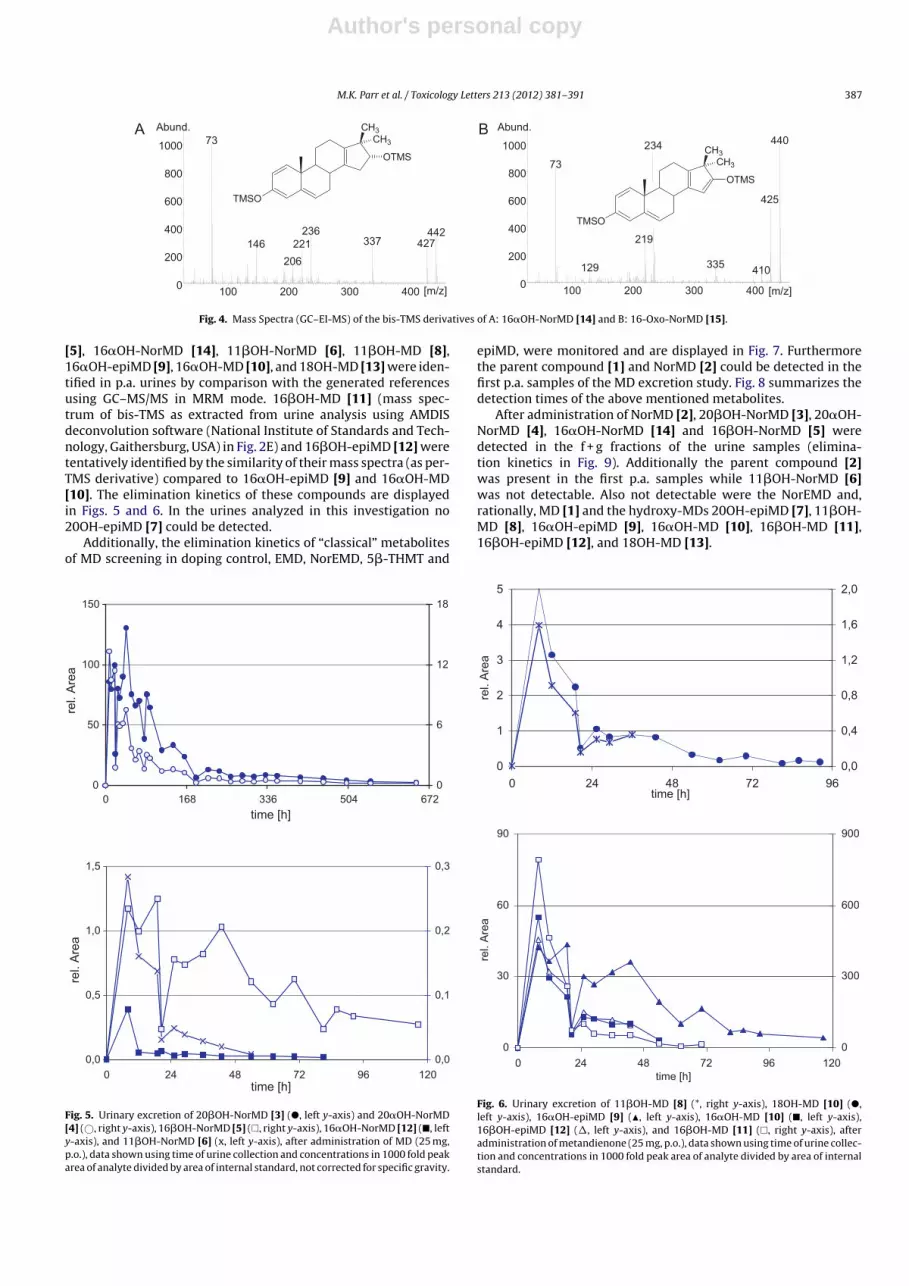
of their per-TMS derivatives (Fig. 4), they are proposed to be
16�-hydroxy-17,17-dimethyl-18-norandrosta-1,4,13-trien-3-one
(16�OH-NorMD [14]) and 17,17-dimethyl-18-norandrosta-1,4,13-
triene-3,16-dione (16-Oxo-NorMD [15]), respectively. As expected
for epimeric steroids, the bis-TMS derivative of 16�OH-NorMD [14]
revealed a very similar mass spectrum compared to 16�OH-NorMD
[5], giving additional evidence for the structure proposal. Compar-
ison of the retention time and mass spectra with the chemically
derived 16�OH-NorMD [14] further confirmed this proposal.
As reported before (Zöllner et al., 2009, 2010), biotransforma-
tion of NorMD [2] with the recombinant fission yeast strain CAD67
(which co-expresses human CPR and CYP3A4) yields the two 17-
epimers 20�OH-NorMD [4] and 20�OH-NorMD [3], respectively. In
this study, we observed that 20�OH-NorMD was more efficiently
produced than 20�OH-NorMD. Moreover, we definitely confirmed
the structure of 20�OH-NorMD [4] by GC–MS comparison with the
chemically synthesized reference. The mass spectra of the bis-TMS
derivatives are displayed in Fig. 3A and B.
3.3. Administration studies
Following the oral administration of MD [1] the combined
unconjugated and glucuronidated metabolite (f + g) fractions were
analyzed as per-TMS derivatives as used in routine sports dop-
ing control. 20�OH-NorMD [3], 20�OH-NorMD [4], 16�OH-NorMD
A
50 10 0 15 0 20 0 25 0 30 0 35 0 4000
200
400
600
800
1000
[m/z]
Abund.
73
143
191
206
229
283
299
339
354
429
444
TMSO
OTM S
CH3
B
40 80 12 0 16 0 20 0 24 0 28 0 3200
200
400
600
800
1000
[m/z]
Abund.
73
91
133
148
161 194
206
229 283 324
339
354
TMSO
Fig. 1. Mass spectra (GC–EI-MS) of the per-TMS derivatives of A: MD [1], and B: NorMD [2].

Author's personal copy
386 M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391
A
50 10 0 15 0 20 0 25 0 30 0 35 0 40 0 45 00
200
400
600
800
1000
[m/z]
Abund.
206532
33773
429147 231
TMSO
CH2OTMS
OTMSB
100 20 0 30 0 40 0 50 00
200
400
600
800
1000
[m/z ]
Abund.
73
532
337
143206
297
243427 517
OTMS
TMS O
CH3TMSO
C
50 100 150 200 250 300 350 400 4500
200
400
600
800
1000
[m/z ]
Abund.
73
206
532
147 337 42 7117 231
299
TMS O
CH3
OTMS
OTMS
D
50 10 0 15 0 20 0 25 0 30 0 35 0 40 0 45 00
200
400
600
800
[m/z ]
Abund . 73
532
206
147
337299117 231 42745
1000
OTMS
TMSO
CH3
OTMS
E
100 15 0 20 0 25 0 30 0 35 0 40 0 45 0
200
400
600
800
1000
[m/z]
Abund73
231206 532
117147
253 299 337 495458427
0
TMS O
OTM S
CH3
OTMS
F
100 20 0 30 0 40 0 50 00
200
400
600
800
1000
[m/z]
Abund .
73
206
532
337143
243103281
442
OTMS
TMS O
CH3
TMSO
339
Fig. 2. Mass spectra (GC–EI-MS) of the tris-TMS derivatives of A: 20OH-epiMD [7], B: 11�OH-MD [8], C: 16�OH-EpiMD [9], D: 16�OH-MD [10], E: 16�OH-MD [11], and F:
18OHMD [13].
A
50 10 0 15 0 20 0 25 0 30 0 35 0 4000
200
400
600
800
1000
[m/z]
Abund.
45
73
91103
133
159
193
206
243
339
427
442
CH2OTMS
TMSO
CH3
B
50 10 0 15 0 20 0 25 0 30 0 35 0 4000
200
400
600
800
1000
[m/z]
Abund.
73
133
193
219
236243 337
339
427
442
TMSO
CH3
CH2OTMS
C
50 10 0 15 0 20 0 25 0 30 0 35 0 4000
200
400
600
800
1000
[m/z]
Abund.
73
146
194206
236
337 427
442
CH3
TMSO
CH3
OTMS
221
D
40 80 12 0 16 0 20 0 24 0 28 0 32 0 36 0 4000
200
400
600
800
1000
[m/z]
Abund.
33773
442194
281
221
179
385
TMSO
TMSO
Fig. 3. Mass spectra (GC–EI-MS) of the bis-TMS derivatives of the hydroxylation products of NorMD [2]: A: 20�OH-NorMD [3], B: 20�OH-NorMD [4], C: 16�OH-NorMD [5],
and D: 11�OH-NorMD [6].
Author's personal copy
M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391 387
A
100 20 0 30 0 4000
200
400
600
800
1000
[m/z]
Abund.
73
236 442337146
206
427221
TMSO
CH3
CH3
OTMS
B
100 20 0 30 0 4000
200
400
600
800
1000
[m/z]
Abund.
440234
73
335129 410
425
219
TMSO
CH3
CH3
OTMS
Fig. 4. Mass Spectra (GC–EI-MS) of the bis-TMS derivatives of A: 16�OH-NorMD [14] and B: 16-Oxo-NorMD [15].
[5], 16�OH-NorMD [14], 11�OH-NorMD [6], 11�OH-MD [8],
16�OH-epiMD [9], 16�OH-MD [10], and 18OH-MD [13] were iden-
tified in p.a. urines by comparison with the generated references
using GC–MS/MS in MRM mode. 16�OH-MD [11] (mass spec-
trum of bis-TMS as extracted from urine analysis using AMDIS
deconvolution software (National Institute of Standards and Tech-
nology, Gaithersburg, USA) in Fig. 2E) and 16�OH-epiMD [12] were
tentatively identified by the similarity of their mass spectra (as per-
TMS derivative) compared to 16�OH-epiMD [9] and 16�OH-MD
[10]. The elimination kinetics of these compounds are displayed
in Figs. 5 and 6. In the urines analyzed in this investigation no
20OH-epiMD [7] could be detected.
Additionally, the elimination kinetics of “classical” metabolites
of MD screening in doping control, EMD, NorEMD, 5�-THMT and
0
50
100
150
6725043361680
time [h ]
rel. A
rea
0
6
12
18
0,0
0,5
1,0
1,5
120967248240time [h ]
rel. A
rea
0,0
0,1
0,2
0,3
Fig. 5. Urinary excretion of 20�OH-NorMD [3] (�, left y-axis) and 20�OH-NorMD
[4] (©, right y-axis), 16�OH-NorMD [5] (�, right y-axis), 16�OH-NorMD [12] (�, left
y-axis), and 11�OH-NorMD [6] (x, left y-axis), after administration of MD (25 mg,
p.o.), data shown using time of urine collection and concentrations in 1000 fold peak
area of analyte divided by area of internal standard, not corrected for specific gravity.
epiMD, were monitored and are displayed in Fig. 7. Furthermore
the parent compound [1] and NorMD [2] could be detected in the
first p.a. samples of the MD excretion study. Fig. 8 summarizes the
detection times of the above mentioned metabolites.
After administration of NorMD [2], 20�OH-NorMD [3], 20�OH-
NorMD [4], 16�OH-NorMD [14] and 16�OH-NorMD [5] were
detected in the f + g fractions of the urine samples (elimina-
tion kinetics in Fig. 9). Additionally the parent compound [2]
was present in the first p.a. samples while 11�OH-NorMD [6]
was not detectable. Also not detectable were the NorEMD and,
rationally, MD [1] and the hydroxy-MDs 20OH-epiMD [7], 11�OH-
MD [8], 16�OH-epiMD [9], 16�OH-MD [10], 16�OH-MD [11],
16�OH-epiMD [12], and 18OH-MD [13].
0
1
2
3
4
5
967248240time [h]
rel. A
rea
0,0
0,4
0,8
1,2
1,6
2,0
0
30
60
90
120967248240
time [h]
rel. A
rea
0
300
600
900
Fig. 6. Urinary excretion of 11�OH-MD [8] (*, right y-axis), 18OH-MD [10] (�,
left y-axis), 16�OH-epiMD [9] (�, left y-axis), 16�OH-MD [10] (�, left y-axis),
16�OH-epiMD [12] (�, left y-axis), and 16�OH-MD [11] (�, right y-axis), after
administration of metandienone (25 mg, p.o.), data shown using time of urine collec-
tion and concentrations in 1000 fold peak area of analyte divided by area of internal
standard.

Author's personal copy
388 M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391
0
30
60
90
120
24019214496480
time [h]
rel. A
rea
0
6
12
18
24
0
2000
4000
6000
120967248240
time [h]
rel. A
rea
0
20
40
60
Fig. 7. Urinary excretion of EMD (�, left y-axis), NorEMD (�, right y-axis), 5�-THMT
(×, right y-axis), and epiMD (∗, left y-axis) after administration of MD (25 mg, p.o.),
data shown using time of urine collection and concentrations in 1000 fold peak area
of analyte divided by area of internal standard.
4. Discussion
4.1. Chemical syntheses and characterization of reference
material
In accordance with the synthesis of 17-methyl steroids as
described in the literature, during the synthesis of 11�OH-MD
[8] the inserted methyl group is 17�-oriented, due to the steri-
cal hindrance by the methyl group at C-13, which is in �-position.
0 168 33 6 504
epi-MD
5b-THMT
NorEMD
EMD
[13] 18OH-MD
[12] 16bOH-epiMD
[11] 16bOH-MD
[10] 16aOH-MD
[9] 16aOH-epiMD
[8] 11bOH-MD
[14] 16aOH-NorMD
[6] 11bOH-NorMD
[5] 16bOH-NorMD
[4] 20aOH-NorMD
[3] 20bOH-NorMD
detection times [h]
Fig. 8. Detection times of the urinary metabolites of MD (25 mg, p.o).
0
300
600
900
288216144720
time [h]
rel. A
rea
0
100
200
300
0
25
50
75
100
967248240
time [h]
rel. A
rea
Fig. 9. Urinary elimination of NorMD (2 mg, p.o.): Kinetics of 20�OH-NorMD [3] (�,
left y-axis), 20�OH-NorMD [4] (©, right y-axis), 16�OH-NorMD [5] (�, right y-axis),
and 16�OH-NorMD [12] (�, left y-axis), data shown using time of urine collection
and concentrations in 1000 fold peak area of analyte divided by area of internal
standard.
During the analogous reaction starting from 16�-hydroxyandrost-
4-ene-3,17-dione [VIII] both the 17�-methyl (16�OH-MD, [10])
and the 17�-methyl product (16�OH-epiMD, [9]) were obtained.
Similar findings were also reported by Schänzer et al. (1990) for the
Grignard methylation in the synthesis of 16�-hydroxystanozolol.
In accordance to this report the earlier eluting product was
obtained in higher yield and could be assigned to 16�OH-epiMD
[9]. This was further confirmed by the formation of an acetonide
from [9], while [10] did not react. As common for 17-epimeric
steroids the mass spectra of their per-TMS derivatives are very
similar.
The mass spectra of the tris-TMS derivatives of the hydroxy-
lated metandienones [8], [9], [10], and [13] (Fig. 2) all show ions
at m/z 517 (M+−15), m/z 442 (M+−90), m/z 427 (M+−15–90), and
m/z 337 (M+−15–2 × 90), representing the fragments generated
by the losses of •CH3 and/or TMS-OH from the molecule. While
20OH-epiMD [7], 16�OH-epiMD [9], 16�OH-MD [10], and 18OH-
MD [13] show a very abundant m/z 206, this fragment, representing
the enolized A/B-ring of androsta-1,4-diene-3-ones, is less abun-
dant in the spectrum of 11�OH-MD [8]. The typical fragment of
17-methyl steroids m/z 143 (carbon atoms C-15 to C-17) is present
in the 11�OH-MD [8] spectrum, while it is incremented by 88 Da to
m/z 231 due to the additional TMS-derivatized OH-group in the D-
ring of 20OH-epiMD [7], 16�OH-epiMD [9], and 16�OH-MD [10].
The loss of 103 Da (TMS-O-CH2•) from the molecule yielding m/z
429 followed by a loss of TMS-OH (90 Da) yielding m/z 339 can be
allocated to the silylated primary hydroxyl group in 20OH-epiMD
[7] and 18OH-MD [13], and is consequently not detected in the
spectra of the other hydroxylated metandienones [8], [9], and [10].
Also, an exclusive fragment with m/z 297 appears in the spectrum
of 11�OH-MD [8]. This can be assigned to the M+-TMS-OH after an
additional loss of the D-ring (−145 Da).

Author's personal copy
M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391 389
Also in the spectra of the TMS-derivatized Wagner–Meerwein
products NorMD [2], 20�OH-NorMD [3], 20�OH-NorMD [4],
11�OH-NorMD [6], and 16�OH-NorMD [14] (Figs. 1B and 3) m/z
206 can be assigned to the fragment of the enolized A/B-ring. Inter-
estingly, in the spectrum of 11�OH-NorMD [6] m/z 194 and m/z
179, that can also be allocated to the A/B-ring, are even more abun-
dant. Most likely the fragment m/z 193 in the spectrum of NorMD
[2], 20�OH-NorMD [3], 20�OH-NorMD [4], 16�OH-NorMD [5], and
16�OH-NorMD [14] is shifted towards m/z 281 (+88 Da) in case of
11�OH-NorMD [6]. However, a structure of this fragment cannot
be proposed from the data available at the moment. As described
by Schänzer et al. (2006) the fragment m/z 236 represents the
hydroxylated C/D-ring (abundant in 20�OH-NorMD [3] and [4], and
16�OH-NorMD [14]) and m/z 133 is generated from that by the
loss of •CH2-OTMS (103 Da) in case of 20�OH-NorMD ([3] and [4]),
while the loss of TMS-OH (90 Da) leads to m/z 146 in case of 16�OH-
NorMD [5] and 16�OH-NorMD [14]. The analogous fragments in
the spectrum of the mono-TMS derivative of NorMD [2], show m/z
148 and m/z 133. The above mentioned loss of •CH2-OTMS from the
molecule identifies 20�OH-NorMD [3] and [4] by the fragment m/z
339. Definitive proof of the structures suggested for [3], [4], and [5]
could be established from NMR data.
4.1.1. Metandienone hydroxylation
As mentioned above, there are six human steroid hydroxy-
lases that catalyze key steps in the biosynthesis of glucocorticoids,
mineralocorticoids and sex hormones (Miller and Auchus, 2011).
Due to the nature of the reactions they catalyze, two of these
enzymes were omitted in the present MD [1] biotransformation
study: CYP11A1 acts on the side chain of the endogenous substrate
cholesterol (which is missing in MD), and CYP19A1 substrates need
to be enolized in the A-ring yielding 2-enol intermediates (Brodie
et al., 1969; Cole and Robinson, 1990; Korzekwa et al., 1991). Due
to the C1–2 double bond this is impossible for MD [1]. Out of the
four remaining enzymes, CYP17 and CYP21 did not convert MD
[1], while CYP11B1 and CYP11B2 could be shown to hydroxylate
this substrate. The adrenal enzymes CYP11B1 and CYP11B2 are the
two sole members of the human CYP11B family and catalyze the
final steps in the biosynthesis of cortisol and aldosterone, respec-
tively (Bureik et al., 2002a; Kawamoto et al., 1990a, 1990b; Mornet
et al., 1989). More specifically, CYP11B1 is known as a steroid
11�-hydroxylase that catalyzes the conversion of 11-deoxycortisol
to cortisol, whereas CYP11B2 displays three different activities,
as it catalyzes the 11�-hydroxylation of 11-deoxycorticosterone
to corticosterone, the 18-hydroxylation of corticosterone and the
18-oxidation of 18-hydroxy corticosterone to aldosterone. Both
CYP11B1 and CYP11B2 have very narrow substrate spectra in com-
parison to the typical xenobiotic converting liver P450s like CYP3A4
or CYP2D6. In the present study we observed that CYP11B1 also cat-
alyzed the 11�-hydroxylation of MD [1] to 11�OH-MD [8], while
CYP11B2 produced both 11�OH-MD [8] and 18OH-MD [10]. Inter-
estingly, both enzymes could therefore accept a substrate that
differs from their physiological target molecules in several respects,
as it contains an additional �1 double bond in ring A, and a short-
ened side-chain in 17�-position instead of 17�- position.
Following 17�-sulfatation, these newly found products can be
converted to 20�OH-NorMD [3] (from 18OH-MD [10]) and 11�OH-
NorMD [6] (from 11�OH-MD [8]), respectively (mass spectra in
Fig. 3A and D) by Wagner–Meerwein rearrangement (Schänzer
et al., 1992b). Since Schizosaccharomyces pombe does not contain a
sulfotransferase enzyme (Wood et al., 2002), these reactions were
not observed during the CYP11B2-dependent biotransformations
of MD in fission yeast.
As already described by Zöllner et al. (2010), other tested CYP
expressing fission yeast strains, especially CYP11B1 and CYP11B2,
were not capable to catalyze hydroxylations when NorMD [2]
is used as substrate, which is also consistent with the find-
ings from the excretion study. The results obtained in this study
suggest enzymatic pathways that involve CYP21 and CYP3A4
in the hydroxylation of NorMD [2], while CYP21, CYP3A4 and
CYP11B2 were shown to take part in 20�OH-NorMD [3] gen-
eration from MD [1] (Scheme 4). The conversion can either
take place by Wagner–Meerwein rearrangement followed by
hydroxylation or in opposite order. Comparison of the relative
intensities of 16�OH-NorMD [5] and 16�OH-NorMD [14] as dis-
played in Fig. 10 suggests that 16-hydroxylation of MD occurs
prior to its conversion to 16�OH-NorMD rather than opposite
order. While 16�OH-NorMD [16] is the dominant product of the
16-hydroxylation of NorMD, the 16-hydroxylation of MD [1] pref-
erentially occurs 16�-oriented. This is also conspicuous in the
excretion of the 16�-hydroxymetandienones (Fig. 6). Also in case of
11�-hydroxylation the data suggest MD hydroxylation before the
rearrangement, as NorMD [2] is not converted to 11�OH-NorMD
[6]. From the data presented the pathway from MD [1] to 20�OH-
NorMD [4] appears to involve Wagner–Meerwein rearrangement
to NorMD [2] first, which is then hydroxylated by CYP21. The oppo-
site order would presuppose the generation of 20OH-epiMD [7],
which could not be detected in the samples analyzed. The classical
O
OH
CH3
O
OH
CH3
O
OH
CH3
O
OH
CH3
OH
HO
HO
O
O
CH2OH
CH3
O
CH3
CH3
O
O
CH3
CH2OH
CYP3 A4
HO
?
CYP21
WM rea rr.WM re arr. WM re arr.
CYP11B2
OH
WM re arr.
[1]
[2]
[9]- ]31[]21[
[5]+ ]3[]41[
[8]
[6]
[4]
WM rearr. ≡ Wagner- Meer wei n rearran geme nt
Scheme 4. Proposed metabolic pathways of MD [1] and NorMD [2] leading to 20�OH-NorMD [3] * In case of 16�OH-MD two 17-epimers each ([9]–[12]) were obtained.

Author's personal copy
390 M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391
0
2
4
6
0 24 48 72
time [h]
rel. A
rea
Fig. 10. Ratio of urinary 16�OH/16�-NorMD [12]/[5] in the p.a. urines with MD [1]
(�) and NorMD [2] (©).
MD metabolite NorEMD appears to be formed from EMD as inter-
mediate rather than from NorMD, as it was not detected after the
administration of NorMD [2].
Conflict of interest
None declared.
Acknowledgments
We thank Gabriele Schultze, Cologne, and Maria Widjaja,
PomBioTech, for technical support and Dr. Calin-Aurel Dragan,
PomBioTech, for helpful discussions. We also thank Prof. Rita
Bernhardt, Saarland University, for purified enzymes (AdR, Adx,
CYP11B1, and CYP11B2, resp.) that were used for the first prelimi-
nary experiments.
The Manfred Donike Institut für Doping Analytik e.V., Cologne,
and the Federal Ministry for the Interior, Berlin, are acknowledged
for financial support of the study.
Appendix A. Supplementary data
Supplementary data associated with this article can be
found, in the online version, at http://dx.doi.org/10.1016/j.toxlet.
2012.07.020.
References
Basaria, S., 2010. Androgen abuse in athletes: detection and consequences. Journalof Clinical Endocrinology and Metabolism 95, 1533–1543.
Brodie, H.J., Kripalan, Kj, Possanza, G., 1969. Studies on mechanism of estrogenbiosynthesis. 6. Stereochemistry of hydrogen elimination at C-2 during aroma-tization. The Journal of the American Chemical Society 91, 1241.
Bureik, M., Lisurek, M., Bernhardt, R., 2002a. The human steroid hydroxylasesCYP1B1 and CYP11B2. Biological Chemistry 383, 1537–1551.
Bureik, M., Schiffler, B., Hiraoka, Y., Vogel, F., Bernhardt, R., 2002b. Functional expres-sion of human mitochondrial CYP11B2 in fission yeast and identification of a newinternal electron transfer protein, etp1. Biochemistry 41, 2311–2321.
Cole, P.A., Robinson, C.H., 1990. Mechanism and inhibition of cytochrome P-450aromatase. Journal of Medicinal Chemistry 33, 2933–2942.
Donike, M., 1969. N-methyl-N-trimethylsilyl-trifluoroacetamide a new silylatingagent from series of silylated amides. Journal of Chromatography 42, 103–104.
Dragan, C.A., Peters, F.T., Bour, P., Schwaninger, A.E., Schaan, S.M., Neunzig, I.,Widjaja, M., Zapp, J., Kraemer, T., Maurer, H.H., Bureik, M., 2011. Convenientgram-scale metabolite synthesis by engineered fission yeast strains expressingfunctional human P450 systems. Applied Biochemistry and Biotechnology 163,965–980.
Geyer, H., Schänzer, W., Mareck-Engelke, U., Nolteernsting, E., Opfermann, G., 1998.Screening procedure for anabolic steroids-control of hydrolysis with deuteratedandrosterone glucuronide and studies with direct hydrolysis. In: Schänzer, W.,Geyer, H., Gotzmann, A., Mareck-Engelke, U. (Eds.), Recent Advances in DopingAnalysis, vol. 5. Sport und Buch Strauss, Köln, pp. 99–102.
Gould, D.H., Herzog, H.L., Hershberg, E.B., 1957. In: Office, U.S.P. (Ed.), New Proges-tational Agents. Schering Corp., United States.
Guengerich, F.P., 2005. Human cytochrome P450 enzymes. In: Ortiz de Montellano,P.R. (Ed.), Cytochrome P450—Structure, Mechanism, and Biochemistry. KluwerAcademic/Plenum Publishers, New York, pp. 377–530.
Guengerich, F.P., Tang, Z., Cheng, Q., Salamanca-Pinzon, S.G., 2011. Approaches todeorphanization of human and microbial cytochrome P450 enzymes. Biochim-ica et Biophysica Acta 1814, 139–145.
Hannemann, F., Bichet, A., Ewen, K.M., Bernhardt, R., 2006. Cytochrome P450 systems– biological variations of electron transport chains. Biochimica et BiophysicaActa 1770, 330–344.
Kawamoto, T., Mitsuuchi, Y., Ohnishi, T., Ichikawa, Y., Yokoyama, Y., Sumimoto, H.,Toda, K., Miyahara, K., Kuribayashi, I., Nakao, K., Hosoda, K., Yamamoto, Y., Imura,H., Shizuta, Y., 1990a. Cloning and expression of a cDNA for human cytochromeP450ALDO as related to primary aldosteronism. Biochemical and BiophysicalResearch Communications 173, 309–316.
Kawamoto, T., Mitsuuchi, Y., Toda, K., Miyahara, K., Yokoyama, Y., Nakao, K., Hosoda,K., Yamamoto, Y., Imura, H., Shizuta, Y., 1990b. Cloning of cDNA and genomicDNA for human cytochrome P-45011 beta. FEBS Letters 269, 345–349.
Korzekwa, K.R., Trager, W.F., Smith, S.J., Osawa, Y., Gillette, J.R., 1991. Theoreticalstudies on the mechanism of conversion of androgens to estrogens by aromatase.Biochemistry 30, 6155–6162.
Miller, W.L., Auchus, R.J., 2011. The molecular biology, biochemistry, and phys-iology of human steroidogenesis and its disorders. Endocrine Reviews 32,81–151.
Mornet, E., Dupont, J., Vitek, A., White, P.C., 1989. Characterization of 2 genes encod-ing human steroid 11-beta-hydroxylase (P-45011-BETA). Journal of BiologicalChemistry 264, 20961–20967.
Nelson, D.R., 2011. Progress in tracing the evolutionary paths of cytochrome P450.Biochimica et Biophysica Acta 1814, 14–18.
Nitta, I., Fujimori, S., Ueno, H., 1985. The syntheses of the corticoid side-chain. 1. Animproved method for the preparation of 17-alpha-hydroxyprogesterone fromandrost-4-ene-3,17-dione. Bulletin of the Chemical Society of the Japan 58,978–980.
Parr, M.K., Opfermann, G., Piper, T., Schlörer, N., Fußhöller, G., Thomas, A., Thevis, M.,Schänzer, W., 2007. Characterisation of steroid metabolites recently detected indoping control analyses. In: Schänzer, W., Geyer, H., Gotzmann, A., Mareck, U.(Eds.), Recent Advances in Doping Analysis, vol. 15. Sport und Buch Strauß, Köln,pp. 143–152.
Parr, M.K., Zoellner, A., Fußhöller, G., Opfermann, G., Hobler, A., Bernhardt, R., Bureik,M., Schänzer, W., 2011. New players in the nightwatch research. In: Schänzer,W., Geyer, H., Gotzmann, A., Mareck, U. (Eds.), Recent Advances in Dope Analysis,vol. 19. Sport und Buch Strauß, Cologne, pp. 60–62.
Rendic, S., Nolteernsting, E., Schänzer, W., 1999. Metabolism of anabolic steroidsby recombinant human cytochrome P450 enzymes: gas chromatographic–massspectrometric determination of metabolites. Journal of Chromatography B 735,73–83.
Sagara, Y., Wada, A., Takata, Y., Waterman, M.R., Sekimizu, K., Horiuchi, T., 1993.Direct expression of adrenodoxin reductase in Escherichia coli and the functionalcharacterization. Biological and Pharmaceutical Bulletin 16, 627–630.
Schänzer, W., Opfermann, G., Donike, M., 1990. Metabolism ofstanozolol—identification and synthesis of urinary metabolites. Journal ofSteroid Biochemistry and Molecular Biology 36, 153–174.
Schänzer, W., Geyer, H., Donike, M., 1991. Metabolism of metandienone inman—identification and synthesis of conjugated excreted urinary metabolites,determination of excretion rates and gas-chromatographic mass-spectrometricidentification of bis-hydroxylated metabolites. Journal of Steroid Biochemistryand Molecular Biology 38, 441–464.
Schänzer, W., Opfermann, G., Donike, M., 1992a. 17-Epimerization of 17-alpha-methyl anabolic-steroids in humans—metabolism and synthesis of17-alpha-hydroxy-17-beta-methyl steroids. Steroids 57, 537–550.
Schänzer, W., Opfermann, G., Donike, M., 1992b. 17-Epimerization of 17�-methylanabolic steroids in humans: metabolism and synthesis of 17�-hydroxy-17�-methyl steroids. Steroids 57, 537–550.
Schänzer, W., Donike, M., 1993. Metabolism of anabolic-steroids in man—synthesisand use of reference substances for identification of anabolic-steroid metabo-lites. Analytica Chimica Acta 275, 23–48.
Schänzer, W., Donike, M., 1995. Synthesis of deuterated steroids for GC/MS quantifi-cation of endogenous steorids. In: Donike, M., Geyer, H., Gotzmann, A., Mareck,U. (Eds.), Recent Advances in Doping Analysis, vol. 2. Sport und Buch Strauß,Köln, pp. 93–112.
Schänzer, W., Horning, S., Donike, M., 1995. Metabolism of anabolic-steroids inhumans—synthesis of 6-beta-hydroxy metabolites of 4-chloro-1,2-dehydro-17-alpha-methyltestosterone, fluoxymesterone, and metandienone. Steroids 60,353–366.
Schänzer, W., Delahaut, P., Geyer, H., Machnik, M., Horning, S., 1996. Long-termdetection and identification of metandienone and stanozolol abuse in athletesby gas chromatography high-resolution mass spectrometry. Journal of Chro-matography B 687, 93–108.
Schänzer, W., Geyer, H., Horning, S., 1999. 17,17-Dimethyl-18-nor-5�-androst-1,13-dien-3�-ol (18-normetenol) in longterm detection and confirmation of positivemetandienone cases. In: Schänzer, W., Geyer, H., Gotzmann, A., Mareck-Engelke,U. (Eds.), Recent Advances in Doping Analysis, vol. 6. Sport und Buch Strauß, Köln,pp. 37–52.
Schänzer, W., Geyer, H., Fußhöller, G., Halatcheva, N., Kohler, M., Parr, M.K.,Guddat, S., Thomas, A., Thevis, M., 2006. Mass spectrometric identification and

Author's personal copy
M.K. Parr et al. / Toxicology Letters 213 (2012) 381– 391 391
characterization of a new long-term metabolite of metandienone in humanurine. Rapid Communications in Mass Spectrometry 20, 2252–2258.
Uhlmann, H., Beckert, V., Schwarz, D., Bernhardt, R., 1992. Expression of bovineadrenodoxin in E. coli and site-directed mutagenesis of/2Fe-2S/cluster ligands.Biochemical and Biophysical Research Communications 188, 1131–1138.
Wood, V., Gwilliam, R., Rajandream, M.A., Lyne, M., Lyne, R., Stewart, A., Sgouros,J., Peat, N., Hayles, J., Baker, S., Basham, D., Bowman, S., Brooks, K., Brown, D.,Brown, S., Chillingworth, T., Churcher, C., Collins, M., Connor, R., Cronin, A., Davis,P., Feltwell, T., Fraser, A., Gentles, S., Goble, A., Hamlin, N., Harris, D., Hidalgo, J.,Hodgson, G., Holroyd, S., Hornsby, T., Howarth, S., Huckle, E.J., Hunt, S., Jagels,K., James, K., Jones, L., Jones, M., Leather, S., McDonald, S., McLean, J., Mooney,P., Moule, S., Mungall, K., Murphy, L., Niblett, D., Odell, C., Oliver, K., O’Neil, S.,Pearson, D., Quail, M.A., Rabbinowitsch, E., Rutherford, K., Rutter, S., Saunders, D.,Seeger, K., Sharp, S., Skelton, J., Simmonds, M., Squares, R., Squares, S., Stevens, K.,Taylor, K., Taylor, R.G., Tivey, A., Walsh, S., Warren, T., Whitehead, S., Woodward,J., Volckaert, G., Aert, R., Robben, J., Grymonprez, B., Weltjens, I., Vanstreels, E.,Rieger, M., Schafer, M., Muller-Auer, S., Gabel, C., Fuchs, M., Fritzc, C., Holzer, E.,Moestl, D., Hilbert, H., Borzym, K., Langer, I., Beck, A., Lehrach, H., Reinhardt, R.,Pohl, T.M., Eger, P., Zimmermann, W., Wedler, H., Wambutt, R., Purnelle, B., Gof-feau, A., Cadieu, E., Dreano, S., Gloux, S., Lelaure, V., Mottier, S., Galibert, F., Aves,S.J., Xiang, Z., Hunt, C., Moore, K., Hurst, S.M., Lucas, M., Rochet, M., Gaillardin, C.,Tallada, V.A., Garzon, A., Thode, G., Daga, R.R., Cruzado, L., Jimenez, J., Sanchez,M., del Rey, F., Benito, J., Dominguez, A., Revuelta, J.L., Moreno, S., Armstrong,J., Forsburg, S.L., Cerrutti, L., Lowe, T., McCombie, W.R., Paulsen, I., Potashkin, J.,
Shpakovski, G.V., Ussery, D., Barrell, B.G., Nurse, P., 2002. The genome sequenceof Schizosaccharomyces pombe. Nature 415, 871–880.
World Anti-Doping Agency, 2010. Adverse analytical findings reported by accreditedlaboratories. Online at http://www.wada-ama.org/en/Science-Medicine/Anti-Doping-Laboratories/Laboratory-Statistics/, 27.04.2010.
World Anti-Doping Agency, 2010. The 2011 Prohibited List. Online athttp://www.wada-ama.org/Documents/World Anti-Doping Program/WADP-Prohibited-list/To be effective/WADA Prohibited List 2011 EN.pdf,21.03.2011.
Wu, D.A., Hu, M.C., Chung, B.C., 1991. Expression and functional study of wild-typeand mutant human cytochrome P450c21 in Saccharomyces cerevisiae. DNA andCell Biology 10, 201–209.
Zöllner, A., Kagawa, N., Waterman, M.R., Nonaka, Y., Takio, K., Shiro, Y., Hannemann,F., Bernhardt, R., 2008. Purification and functional characterization of human 11beta hydroxylase expressed in Escherichia coli. FEBS Journal 275, 799–810.
Zöllner, A., Parr, M.K., Dragan, C.A., Peters, F., Maurer, H., Schanzer, W., Bureik, M.,2009. CYP21 catalyzed production of 18-nor-17 beta-hydroxymethyl, 17 alpha-methylandrosta-1,4,13-trien-3-one: a new player in the fight against doping.Drug Metabolism Reviews 41, 80.
Zöllner, A., Parr, M.K., Dragan, C.-A., Dras, S., Schlörer, N., Peters, F.T., Maurer, H.H.,Schänzer, W., Bureik, M., 2010. CYP21-catalyzed production of the long-termurinary metandienone metabolite 17beta-hydroxymethyl-17 alpha-methyl-18-norandrosta-1,4,13-trien-3-one: a contribution to the fight against doping.Biological Chemistry 391, 119–127.
Top Related
Copyright © 2022 FDOKUMEN

