







Bahasa
Halaman
Hukum


Transformation, Localization, and Biomolecular Binding of HgSpecies at Subcellular Level in Methylating and NonmethylatingSulfate-Reducing BacteriaZoyne Pedrero,*,† Romain Bridou,‡ Sandra Mounicou,† Remy Guyoneaud,‡ Mathilde Monperrus,*,†
and David Amouroux†
†Laboratoire de Chimie Analytique Bio-Inorganique et Environnement, Institut Pluridisciplinaire de Recherche sur l'Environnementet les Materiaux UMR 5254, CNRS Universite de Pau et des Pays de l’Adour, 2 Avenue Pierre Angot, 64053 Pau, France‡Equipe Environnement et Microbiologie, Institut Pluridisciplinaire de Recherche sur l'Environnement et les Materiaux, UMR5254,CNRS Universite de Pau et des Pays de l’Adour, Batiment IBEAS, BP1155, 64013 Pau Cedex, France
*S Supporting Information
ABSTRACT: Microbial activity is recognized to play animportant role on Hg methylation in aquatic ecosystems.However, the mechanism at the cellular level is still poorlyunderstood. In this work subcellular partitioning and trans-formation of Hg species in two strains: Desulfovibrio sp.BerOc1 and Desulfovibrio desulfuricans G200 (which exhibitdifferent Hg methylation potential) are studied as an approachto the elucidation of Hg methylation/demethylation processes.The incubation with isotopically labeled Hg species (199Hgiand Me201Hg) not only allows the determination ofmethylation and demethylation rates simultaneously, but alsothe comparison of the localization of the originally added andresulting species of such metabolic processes. A dissimilar Hgspecies distribution was observed. In general terms, monomethylmercury (MeHg) is preferentially localized in the extracellularfraction; meanwhile inorganic mercury (Hgi) is associated to the cells. The investigation of Hg binding biomolecules on thecytoplasmatic and extracellular fractions (size exclusion chromatography coupled to ICP-MS) revealed noticeable differences inthe pattern corresponding to the Hg methylating and nonmethylating strains.
■ INTRODUCTION
Mercury methylation in aquatic ecosystems is a critical step inthe accumulation of this hazardous species in fish, leading tohuman exposure. It is well established that microbialmethylation is one of the major biotic processes that transforminorganic mercury (Hgi) into toxic MeHg. Sulfate-reducingbacteria (SRB) are recognized as the principal Hg methylatorsin anoxic waters and sediments,1,2 which hold up to 98% ofmercury in aquatic ecosystems.3 Despite being predominantlyanaerobic, recent studies have revealed methylation activity inoxygen rich environments,4 such as the peryphiton ofAmazonian macrophytes.5 Due to the direct impact on theecosystem, several studies focused on the investigation of Hgmethylation by SRB, most of them concentrated on theinfluence of environmental parameters6−8 and biogeochemicalfactors.9 However, Hg methylation mechanisms remainunknown and it has been suggested that it could be aprotective mechanism against Hgi toxicity developed during theearly evolution of some microorganisms.10
It should be also considered that some SRB exhibit both Hgimethylation and MeHg demethylation potentials.11 To estimatethe net methylation, the demethylation process should be also
studied. It has been proposed that MeHg demethylation couldbe produced by reductive pathways under oxic conditions,mediated by the mer-operon system and leading to theformation of Hg0 and CH4.
12,13 In contrast, oxidativedemethylation dominates under anoxic conditions, producingHgi and CO2.
12,14,15
One of the main hypothesized mechanisms for SRB suggeststhat Hgi can be methylated through the acetyl-coenzyme A(CoA) pathway,16 which implies the participation of carbonmonoxide dehydrogenase (CODH) to produce the methylmoiety. However, Ekstrom et al.17 demonstrated later that itcannot be extrapolated to incomplete oxidizing (convertingcarbon substrates to acetate rather than to CO2) SRB strains,where Hg methylation seems to be independent of acetyl-CoA.Apparently, Hg methylation is related to the acetyl-CoA incomplete oxidizers, meanwhile in incomplete oxidizers furtherefforts are required to determine the biochemical pathways.
Received: June 15, 2012Revised: September 27, 2012Accepted: October 10, 2012Published: October 10, 2012
Article
pubs.acs.org/est
© 2012 American Chemical Society 11744 dx.doi.org/10.1021/es302412q | Environ. Sci. Technol. 2012, 46, 11744−11751

In addition, the capability of Hg methylation by SRB is straindependent, rather than dependent on genus or species,11,18
which reflects the necessity of performing pure cells culturestudies. Unfortunately, the interpretation and comparison ofmost of the published data is difficult, due to the differences incell culture conditions (Hg concentration, incubation time,etc.) and results expression units.The incubation with multiple stable isotopic tracers
represents a powerful tool to deal with the simultaneousmicrobial Hg methylation and demethylation determina-tion.11,19,20 It has been on real environmental samples such aswater,21 sediments,9 and periphyton successfully applied to thestudy of Hg methylation/demethylation in complex.5 Thisexperimental approach, characterized by its high precision, isconfirmed as a valuable tool for tracing environmental andmetabolic processes.22
The transmembrane uptake of Hgi has been proposed as adecisive step for MeHg production.23 Schaefer and Morel havedemonstrated that Hg methylation in Geobacter sulfureducens isgreatly promoted by Hg speciation.23 However, there aredivergences with respect to the Hgi uptake, meanwhile someauthors consider it as a passive process.24 Among the differentHg ligands investigated, organic ligands enhance uptakepotential, where the highest one corresponds to cysteine(Cys) through formation of Hg−thiol complex.23 These resultssuggest the influence of Hg binding biomolecules and itsspecific role on its uptake.The identification of molecular targets of Hg contributes to
the understanding of metabolic pathways, since it determinestheir toxicity, mobility, and transformation, etc. As a first step,the study of Hg species distribution among the proteinfractions of the bacterial cell will provide useful and originalinformation as, so far, most of the speciation studies are limitedto the quantification of Hgi and MeHg.In this work, the advantage of using multiple isotopic labeled
species is exploited to investigate not only the methylation anddemethylation potential of two pure bacterial strains:Desulfovibrio sp. BerOc1 and Desulfovibrio desulfuricans G200(Hg methylating control), but also the localization and origin ofthe different Hg species at subcellular levels. Cytoplasmic andextracellular fractions were screened for Hg species bindingbiomolecules by HPLC-ICP-MS in combination with GC-ICP-MS.
■ EXPERIMENTAL SECTIONCulture Conditions. All experiments were performed in
autoclaved glass or Teflon flasks that were cleaned byultrasonication in successive baths of 10% HNO3 and HCl of2-hour cycles followed by rinsing three times in 18 MΩdeionized water.Sulfate-reducing bacterial strains were grown in the dark at
30 °C and pH 7.0−7.1 under fumarate respiration with 10 mMpyruvate and fumarate in order to avoid complexation of Hgspecies with sulfides produced under sulfate-reducing con-ditions. Defined mineral base optimal growth media contained( in g L−1 unless otherwise indicated): NaCl, 3; MgCl2·6 H2O,0.40; CaCl2·2 H2O, 0.10; NH4Cl, 0.25; KH2PO4, 0.20; KCl,0.50; SL12 B, 1 mL L−1; selenite tungstate solution, 1 mL L−1;NaHCO3, 30; vitamins solution (V725), 0.25 mL L−1. Inaddition, precise growth and experimental procedure werefollowed as described below.(1) Strain BerOc1, Desulfovibrio sp. an incomplete oxidizer
isolated from brackish lagoon sediments,26 previously tested for
its capacity to methylate Hg,18 was grown using above-described medium and NaCl was added to a final concentrationof 10 g L−1. This halophilic strain is closely related to the Hgmethylating Desulfovibrio desulfuricans strain ND132, recentlyproposed as a model microorganism to study Hg methyl-ation.27,28 Logarithmic phase growing cells were incubated 96 hat 30 °C to an optical density (A600 nm) of 0.3. The resultingculture was diluted 1:10 in 200 mL of anoxic fresh medium andincubated overnight at 30 °C to an optical density of 0.1 in fourdifferent 250-mL flasks. The overnight cultures were thencarefully transferred in 250-mL PTFE centrifuge flasks underanoxic conditions and centrifuged at 8000g, 25 °C for 15 min.Cells pellets were pooled, washed into mineral base mediumthree times sequentially. Washed cells were gently homogen-ized in 45 mL of anoxic fresh medium without carbon andenergy source (e.g., pyruvate), transferred into a 45-mL PTFEflask and spiked with 100 ng of 199HgCl2 per g of culture mediaand 10 ng of Me201HgCl per g of culture media. The startingresting cell density was determined by cell numeration with aMalassez cell counting chamber. Samples were collected after 5h incubation (37 °C) to determine the concentrations andisotopic composition of Hg species in the different incubatingfractions as described later.(2) Strain G200. D. desulfuricans, which is closely related to
strain G20, was isolated from an oil well souring site.29 It is aDesulfovibrio model microorganism that was intensely involvedin the development of genetic studies for this genus over thelast 20 years.30−33 In this study, it was used as a negative Hgmethylating control and was grown in the same medium andconditions as described for strain BerOc1 except that the NaClconcentration was kept at 3 g L−1. Incubation with Hg specieswas performed under the same conditions previously describedfor the methylating strain BerOc1.Experiments were reproducible as similar results concerning
Hg distribution in cell and media have been observed in twoother repetitions of these experiments, with a variation ofapproximately 10%.
Cell Fractionation. After 5 h of incubation, the cell cultureswere centrifuged at 10 000g at 4 °C for 15 min for theseparation of cells (pellets) and extracellular fraction (super-natant).Pellets were resuspended in 20 mL of 200 mM ammonium
acetate buffer, pH 7.5. Cell disruption was performed by actionof French Press (French Press, Thermo spectronic) at anequivalent pressure of 20 Kpsi applied in a 35-mL standard cellunit FA-032 (40K Standard). The bacterial cell lysate wascollected in a 30-mL Teflon PTFE tube maintained in ice toprevent proteolysis and enzymatic subcellular degradation. Itwas immediately distributed in 1.5-mL Eppendorf tubes(Eppendorf LoBind Protein) and centrifuged at 25 000g at 4°C (Eppendorf, 5417R) for 2 hours (adapted from 16) in orderto separate the supernatant (later referred as cytoplasm) andcell debris fractions (Supporting Information).Separation of the cell-associated and extracellular fractions,
cell integrity after centrifugation of the culture, and the celldisruption efficiency of French press were followed bymicroscopic observations (×1000, Olympus BX60), usingeach previous sequential step as reference control.
Reagents and Standards. All solutions were preparedusing ultrapure water (18 MΩ cm, Millipore). For thepreparation of samples, standards, and blanks, trace metalgrade acids (HNO3 and HCl) were purchased from FisherScientific (Illkirch, France).
Environmental Science & Technology Article
dx.doi.org/10.1021/es302412q | Environ. Sci. Technol. 2012, 46, 11744−1175111745
Stock solutions (1000 mg (Hg) L−1) of Hgi and MeHg ofnatural isotopic composition were prepared by dissolvingmercury(II) chloride (Strem Chemicals, USA) in 1% HNO3and MeHg chloride (Strem Chemicals, USA) in methanol(Sigma Aldrich, France), respectively. Working standardsolutions were prepared daily by appropriate dilution of thestock standard solutions in 1% HNO3 and stored at 4 °C untiluse. 199HgCl2 was prepared by dissolving 199HgO (Oak RidgeNational Laboratory, TN, USA) in HCl (12 mol L−1). Me201Hgwas synthesized from methylcobalamin and 201HgO obtainedfrom Oak Ridge National Laboratory according to theprocedure previously described.34 The isotopic abundance ofisotopes 199 and 201 in the 199Hgi and Me201Hg tracers was 98and 92%, respectively. In both cases the abundance of 202Hgwas lower than 1.5%. The exact concentrations of 199Hgi andMe201Hg of the working solutions were determined by reverseisotope dilution analysis. Hgi and MeHg were derivatized byusing NaBPr4 purchased from Merseburger Spezialchemikalien(Germany).Instrumentation. Liquid chromatographic separations were
carried out with an Agilent 1100 liquid chromatograph(Agilent, Wilmington, DE, USA) equipped with a binaryHPLC pump, an autosampler, and a diode array detector. AnAgilent ICP MS 7500ce (Yokogawa Analytical Systems, Tokyo,Japan) was coupled to the liquid chromatographic system forthe detection of the eluting mercury-containing species.A Thermo Electron GC (Focus) coupled to a Thermo
Electron ICP-MS (X7 X series) was used for the determinationof total concentration of each Hg species. The experimentalconditions are described elsewhere.18 For GC-ICP-MS analysis,samples were digested by using an analytical microwave(CEM), following the methodology proposed in previousexperiments.11
GC-ICP-MS Analysis. All the different culture samplesfractions (bulk after incubation, extracellular fraction, pellets,cell debris, and cytoplasm) were digested with 6 N HNO3 in ananalytical microwave and analyzed by GC-ICP-MS as detailedelsewhere.18 Quantification of isotopically enriched Hgi andMeHg was carried out by reverse isotope dilution analysis byadding the adequate amount of natural Hgi and MeHg NISTstandards. Hg species were propylated by using NaBPr4 andextracted in isooctane by manual shaking for 5 minutes.Extracts were stored at −20 °C before analysis by GC-ICP-MS.The experimental data were mathematically processed by
applying isotope pattern deconvolution approaches previouslydeveloped in our laboratory.22 This technique allows thequantification of both Hg species concentrations and trans-formation factors (i.e., methylation and demethylation)affecting the two isotopic tracers during the analyticalprocedure.
Hg-Containing Biomolecules Analysis by SEC-ICP-MS.Hg-containing biomolecules in cytoplasmic and extracellularfractions were analyzed by HLPC-ICP-MS. ICP-MS was dailyoptimized for maximum sensitivity using a tuning solutioncontaining 1 μg·L−1 Y, Tl, Li, Ce. For size exclusionchromatography experiments, a Superdex 75 (10 × 300 mm× 13 μm) (GE Healthcare, Uppsala, Sweden) with a linearmass separation range between 3 and 70 kDa was used. Thecolumn was calibrated with transferring (81 kDa), bovineserum albumin (66 kDa), superoxide dismutase (40 kDa),carbonic anhydrase (29 kDa), and myoglobulin (16.7 kDa).Before sample injection, the column was cleaned by flushingwith the mobile phase containing 20 mM EDTA during 60 minand conditioned (60 min) with the mobile phase. Samples werefractionated by injection of 100 μL and isocratic elution at 0.7mL min−1 with 100 mM ammonium acetate pH 7.4. Columncontamination by Hg species was monitored by injection of 10mM cysteine solution. The reproducibility of the chromato-grams was verified by triplicate injection of the differentextracts. Chromatographic data were processed using MicrosoftExcel software.Size-exclusion chromatographic fractions containing Hg were
individually collected, digested as mentioned above andanalyzed by GC-ICP-MS for mercury speciation.
■ RESULTS AND DISCUSSION
Hg Species: Transformation and Localization. Methyl-ation and Demethylation Yields. The Hg species (Hgi andMeHg) analyses on the bulk, at the initial and final incubationtime, show a quantitative recovery of the initially added speciesfor both cultures (Table 1).Both investigated strains were able to demethylate MeHg:
Desulfovibrio desulfuricans G200 exhibited 25% of demethyla-tion, meanwhile 10% of demethylation was observed forDesulfovibrio sp. BerOc1. The latter strain exhibits both Hgmethylation and demethylation potentials, as reported for otherSRB strains.11
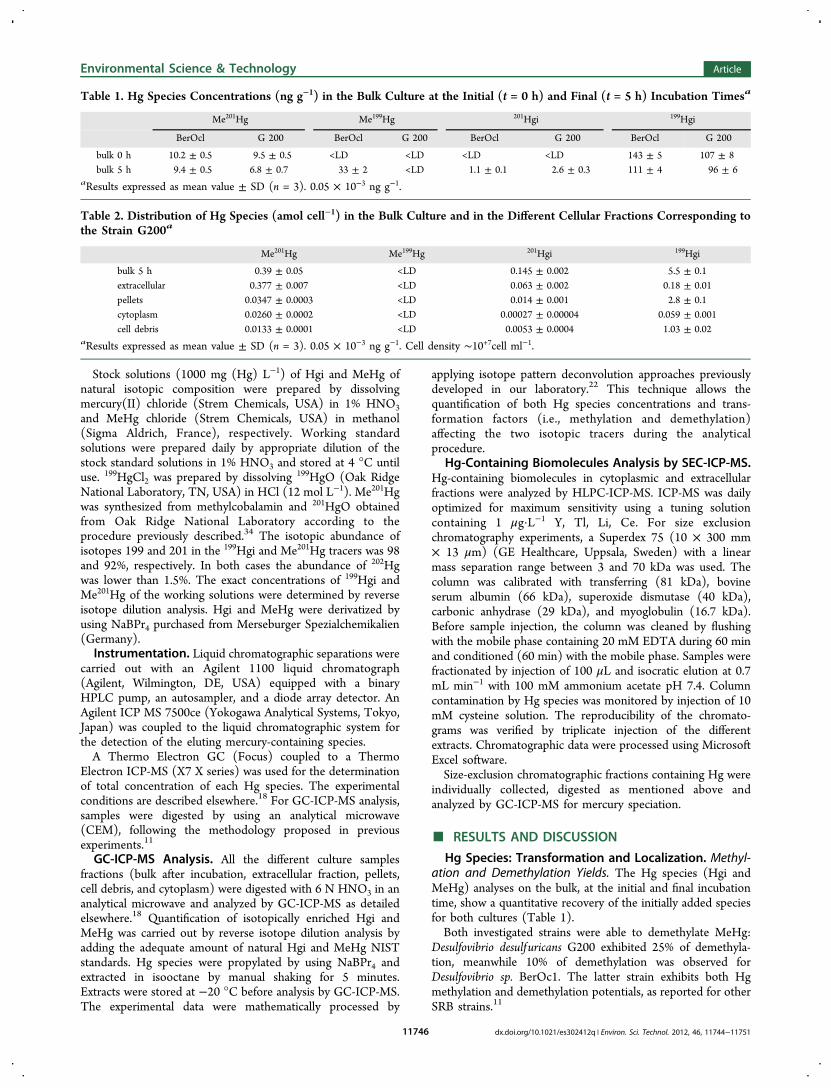
Table 1. Hg Species Concentrations (ng g−1) in the Bulk Culture at the Initial (t = 0 h) and Final (t = 5 h) Incubation Timesa
Me201Hg Me199Hg 201Hgi 199Hgi
BerOcl G 200 BerOcl G 200 BerOcl G 200 BerOcl G 200
bulk 0 h 10.2 ± 0.5 9.5 ± 0.5 <LD <LD <LD <LD 143 ± 5 107 ± 8bulk 5 h 9.4 ± 0.5 6.8 ± 0.7 33 ± 2 <LD 1.1 ± 0.1 2.6 ± 0.3 111 ± 4 96 ± 6
aResults expressed as mean value ± SD (n = 3). 0.05 × 10−3 ng g−1.
Table 2. Distribution of Hg Species (amol cell−1) in the Bulk Culture and in the Different Cellular Fractions Corresponding tothe Strain G200a
Me201Hg Me199Hg 201Hgi 199Hgi
bulk 5 h 0.39 ± 0.05 <LD 0.145 ± 0.002 5.5 ± 0.1extracellular 0.377 ± 0.007 <LD 0.063 ± 0.002 0.18 ± 0.01pellets 0.0347 ± 0.0003 <LD 0.014 ± 0.001 2.8 ± 0.1cytoplasm 0.0260 ± 0.0002 <LD 0.00027 ± 0.00004 0.059 ± 0.001cell debris 0.0133 ± 0.0001 <LD 0.0053 ± 0.0004 1.03 ± 0.02
aResults expressed as mean value ± SD (n = 3). 0.05 × 10−3 ng g−1. Cell density ∼10+7cell ml−1.
Environmental Science & Technology Article
dx.doi.org/10.1021/es302412q | Environ. Sci. Technol. 2012, 46, 11744−1175111746
A 20% methylation rate of Hgi was observed for strainBerOc1, meanwhile it was insignificant for strain G200 underthe same conditions. Despite being phylogenetically relatedwithin the genus Desulfovibrio, important differences regardingHg methylation are observed. This is in good agreement withprevious studies that reveal that methylation capacity is strain-dependent rather than genus/species-dependent.11,18
The differences in Hg methylation and demethylationpotentials on phylogenetically close strains illustrates theimportance of carrying out experiments at strain level tounravel localization and functioning of these antagonisticphysiological mechanisms.Uptake and Localization of Hg Species. In general terms,
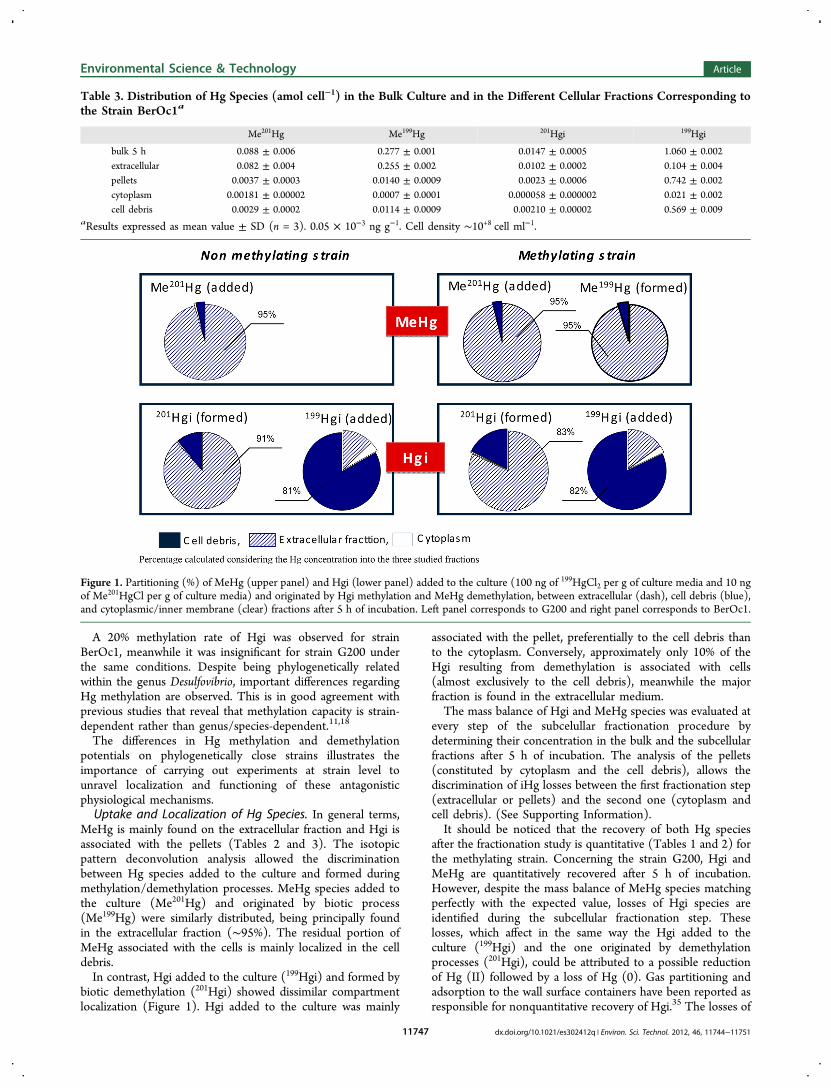
MeHg is mainly found on the extracellular fraction and Hgi isassociated with the pellets (Tables 2 and 3). The isotopicpattern deconvolution analysis allowed the discriminationbetween Hg species added to the culture and formed duringmethylation/demethylation processes. MeHg species added tothe culture (Me201Hg) and originated by biotic process(Me199Hg) were similarly distributed, being principally foundin the extracellular fraction (∼95%). The residual portion ofMeHg associated with the cells is mainly localized in the celldebris.In contrast, Hgi added to the culture (199Hgi) and formed by
biotic demethylation (201Hgi) showed dissimilar compartmentlocalization (Figure 1). Hgi added to the culture was mainly
associated with the pellet, preferentially to the cell debris thanto the cytoplasm. Conversely, approximately only 10% of theHgi resulting from demethylation is associated with cells(almost exclusively to the cell debris), meanwhile the majorfraction is found in the extracellular medium.The mass balance of Hgi and MeHg species was evaluated at
every step of the subcelullar fractionation procedure bydetermining their concentration in the bulk and the subcellularfractions after 5 h of incubation. The analysis of the pellets(constituted by cytoplasm and the cell debris), allows thediscrimination of iHg losses between the first fractionation step(extracellular or pellets) and the second one (cytoplasm andcell debris). (See Supporting Information).It should be noticed that the recovery of both Hg species
after the fractionation study is quantitative (Tables 1 and 2) forthe methylating strain. Concerning the strain G200, Hgi andMeHg are quantitatively recovered after 5 h of incubation.However, despite the mass balance of MeHg species matchingperfectly with the expected value, losses of Hgi species areidentified during the subcellular fractionation step. Theselosses, which affect in the same way the Hgi added to theculture (199Hgi) and the one originated by demethylationprocesses (201Hgi), could be attributed to a possible reductionof Hg (II) followed by a loss of Hg (0). Gas partitioning andadsorption to the wall surface containers have been reported asresponsible for nonquantitative recovery of Hgi.35 The losses of
Table 3. Distribution of Hg Species (amol cell−1) in the Bulk Culture and in the Different Cellular Fractions Corresponding tothe Strain BerOc1a
Me201Hg Me199Hg 201Hgi 199Hgi
bulk 5 h 0.088 ± 0.006 0.277 ± 0.001 0.0147 ± 0.0005 1.060 ± 0.002extracellular 0.082 ± 0.004 0.255 ± 0.002 0.0102 ± 0.0002 0.104 ± 0.004pellets 0.0037 ± 0.0003 0.0140 ± 0.0009 0.0023 ± 0.0006 0.742 ± 0.002cytoplasm 0.00181 ± 0.00002 0.0007 ± 0.0001 0.000058 ± 0.000002 0.021 ± 0.002cell debris 0.0029 ± 0.0002 0.0114 ± 0.0009 0.00210 ± 0.00002 0.569 ± 0.009
aResults expressed as mean value ± SD (n = 3). 0.05 × 10−3 ng g−1. Cell density ∼10+8 cell ml−1.
Figure 1. Partitioning (%) of MeHg (upper panel) and Hgi (lower panel) added to the culture (100 ng of 199HgCl2 per g of culture media and 10 ngof Me201HgCl per g of culture media) and originated by Hgi methylation and MeHg demethylation, between extracellular (dash), cell debris (blue),and cytoplasmic/inner membrane (clear) fractions after 5 h of incubation. Left panel corresponds to G200 and right panel corresponds to BerOc1.
Environmental Science & Technology Article
dx.doi.org/10.1021/es302412q | Environ. Sci. Technol. 2012, 46, 11744−1175111747
Hgi observed during both steps of subcellular fractionation areonly observed for G200, where the final Hgi concentration ismuch higher due to the incapability to methylate Hgi.In this pioneering experiment, bacterial cells were exposed to
Hg concentration higher than natural ones in order to allow theSEC investigations of the subcellular fractions. Although Hgexposure concentration could have affected cell partitioning, theobserved distribution of MeHg in the extracellular fractions isin good agreement with previously reported data.36 Anyway,further experiments should be carried out in the future atenvironmental concentration levels and closer to the naturalHg/thiols ratio. It will imply solving the analytical challengebrought by the expected very low concentrations in collectedfractions after partitioning and SEC chromatography will be thekey feature of this study.Hg Binding Biomolecules in Cytoplasmic and Extrac-
ellular Medium. Considering the difficulty of extracting intactHg−biomolecule complexes from membranes without usingdenaturing reagents, we first screened the Hg distributionamong biomolecules from cytoplasm and extracellular fractionsusing size exclusion chromatography coupled to UV and ICP-MS.Cytoplasm. The UV signal of the cytoplasmic fraction
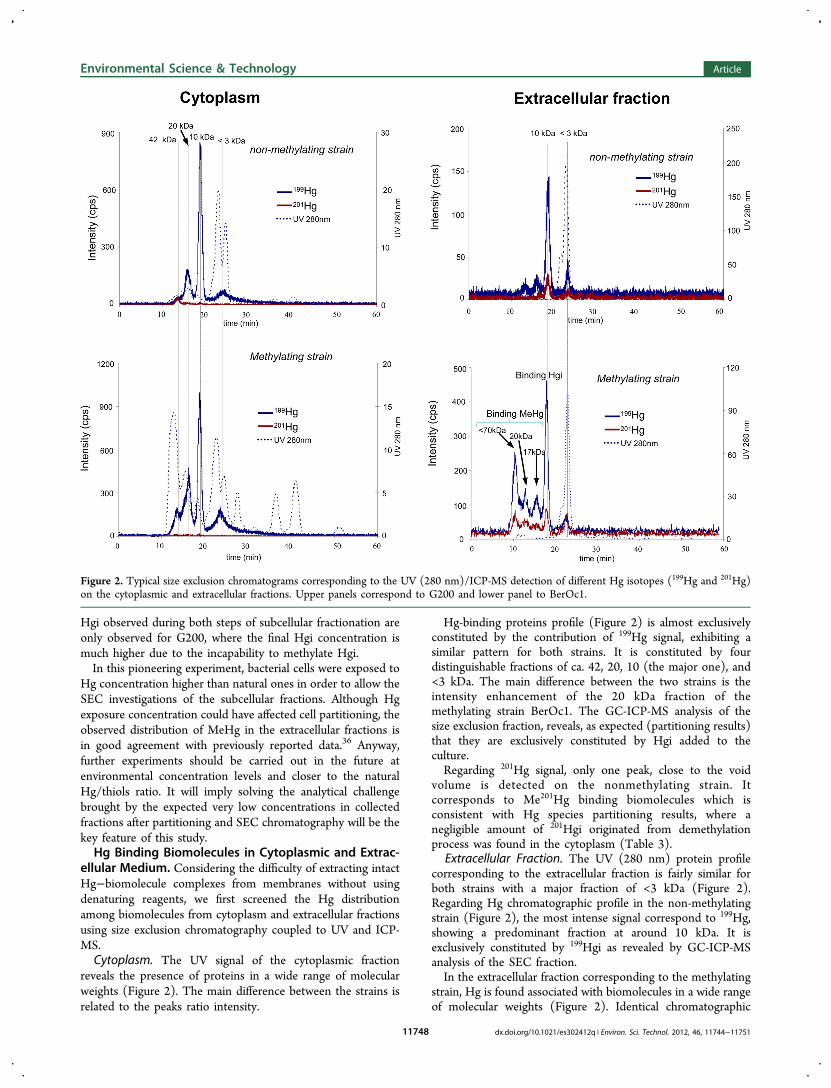
reveals the presence of proteins in a wide range of molecularweights (Figure 2). The main difference between the strains isrelated to the peaks ratio intensity.
Hg-binding proteins profile (Figure 2) is almost exclusivelyconstituted by the contribution of 199Hg signal, exhibiting asimilar pattern for both strains. It is constituted by fourdistinguishable fractions of ca. 42, 20, 10 (the major one), and<3 kDa. The main difference between the two strains is theintensity enhancement of the 20 kDa fraction of themethylating strain BerOc1. The GC-ICP-MS analysis of thesize exclusion fraction, reveals, as expected (partitioning results)that they are exclusively constituted by Hgi added to theculture.Regarding 201Hg signal, only one peak, close to the void
volume is detected on the nonmethylating strain. Itcorresponds to Me201Hg binding biomolecules which isconsistent with Hg species partitioning results, where anegligible amount of 201Hgi originated from demethylationprocess was found in the cytoplasm (Table 3).
Extracellular Fraction. The UV (280 nm) protein profilecorresponding to the extracellular fraction is fairly similar forboth strains with a major fraction of <3 kDa (Figure 2).Regarding Hg chromatographic profile in the non-methylatingstrain (Figure 2), the most intense signal correspond to 199Hg,showing a predominant fraction at around 10 kDa. It isexclusively constituted by 199Hgi as revealed by GC-ICP-MSanalysis of the SEC fraction.In the extracellular fraction corresponding to the methylating
strain, Hg is found associated with biomolecules in a wide rangeof molecular weights (Figure 2). Identical chromatographic
Figure 2. Typical size exclusion chromatograms corresponding to the UV (280 nm)/ICP-MS detection of different Hg isotopes (199Hg and 201Hg)on the cytoplasmic and extracellular fractions. Upper panels correspond to G200 and lower panel to BerOc1.
Environmental Science & Technology Article
dx.doi.org/10.1021/es302412q | Environ. Sci. Technol. 2012, 46, 11744−1175111748

patterns are observed for the two Hg monitored isotopes, withthe 199Hg signal being more intense (as expected frompartitioning results) by a factor of ca. 2.8 than the 201Hgsignal. Besides the dominant fraction detected at around 10kDa consisting of 199Hgi as for the non-methylating strain,three higher molecular weight Hg-containing biomoleculesfractions of ca. > 70, 20, and 17 kDa are found. The GC-ICP-MS analysis of the mentioned SEC fractions revealed theirbinding to MeHg resulting from methylation process(Me199Hg). Regarding 201Hg signal distribution, a trend similarto that of 199Hg is observed; higher molecular weightbiomolecules bind Me201Hg, meanwhile 201Hgi is found withlower molecular weight (10 and <3 kDa) biomolecules.Implications for the Molecular Fate of Hg in SRB Cells.
It is interesting to notice that the uptake and localization of theHg species initially added and formed by biotic process, is quitesimilar for the two Desulfovibrio strains. However, a remarkabledifference on Hg methylation/demethylation potentials isobserved. It indicates that regardless its phylogenetic relation-ship and its similarity concerning the uptake and partitioning ofHg species, differences on induced or constituent enzymes ofthe tested strains can be specific keys in the metabolic pathwaysinvolved in Hg transformations.The preferential localization of MeHg in the extracellular
fraction of the investigated strains is in good agreement withthe recent observations of the release of the organomercurialspecies by Geobacter sulfurreducens23 and Desulfovibrio desulfur-icans36 resulting from Hgi methylation. It has been hypothe-sized that Hgi methylation, followed by the efficient excretionof the organomercurial species may play a role in thedetoxification pathway by microorganisms.10,36,37 It should benoticed that until now, Hg cell partitioning studies exclusivelydescribe the localization of MeHg produced by methylationprocesses. However, isotopic tracers provide evidence that bothMeHg produced by bacteria methylation and MeHg directlyadded to the cell culture are preferentially localized in theextracellular medium. The particular distribution of this organicHg species, observed in both strains (methylating and non-methylating), could be understood as a protective mechanismagainst its toxicity.The opposite partitioning regarding Hgi species added to the
culture (associated with the cells) and formed by MeHgdemethylation (extracellular fraction) question the premise ofcell “permeability” to Hgi species. Despite being recentlyhypothesized that Hgi uptake occurs via active processes,probably by using transport pathways for essential metals, thelocalization of Hgi resulting from MeHg demethylationexclusively in the extracellular medium, reflects that thepartitioning mechanism is much more complex. It can beunderstood as cell saturation by the Hgi initially incorporated,the one directly added to the culture media, but it may be alsointerpreted as part of a detoxification system. The fact thatMeHg and its demethylation product are located in theextracellular medium suggests that MeHg dealkylation couldoccur on the outer cell membrane layer, resulting on the releaseof Hgi from the cell. Nevertheless, it should be considered thatthis study reflects the Hg species distribution after 5 h ofincubation. Therefore, kinetic experiments to determine Hgspecies partitioning at subcellular level constitute the nearfuture perspective of this investigation, as well as the analyseson outer and inner cell membranes.Potential answers to the questions regarding Hg uptake and
its transformation lies on the metalloproteome. A recent critical
publication points out the largely uncharacterized microbialmetalloproteome.38 The Hg-biomolecules investigations havebeen restricted so far to the comparison of Hgi uptake andmethylation rates by addition of amino acids and low molecularweight biomolecules (i.e., Cys, GSH).23,36 However, thepresent work, which represents the first screening ofbiomolecules binding Hg, evidence that the methylating strain,under the tested experimental conditions, releases proteins orbiomolecules of relatively high molecular weight, whichexclusively bind MeHg. Although the role of these proteinscannot be established with the available data, they reveal anunambiguous Hg species specificity and these proteins could beinvolved in the methylation and/or cell release processes bybinding the formed MeHg. The differences in Hg-bindingbiomolecules patterns from the methylating and non-methylating SRB confirm the occurrence of different metabolicprocesses in those phylogenetically close strains. ConcerningHg binding proteins in the cytoplasm, a similar molecularweight distribution pattern is observed for both strains,constituted by four protein fractions binding the Hgi addedto the culture. The noticeable differences in Hg methylationrates of both strains, despite the similarity on the Hg-proteinbinding, suggest that these biomolecules could be probablyinvolved on Hgi transport but not directly on the methylationprocess, which has been recently suggested by Schaefer et al. tooccur in the periplasm.36
These pioneering fractionation results represent a first steptoward understanding the uptake and subcellular localization ofthe species resulting from Hg methylation and demethylationprocesses. The identification of the observed biomoleculesbinding Hg by molecular mass spectrometry techniquesrepresents the next perspectives of this work in view of theelucidation of the Hg methylation metabolism.
■ ASSOCIATED CONTENT*S Supporting InformationAdditional figure describing subcellular partitioning method.This information is available free of charge via the Internet athttp://pubs.acs.org/.
■ AUTHOR INFORMATIONCorresponding Author*E-mail: [email protected] (Z.P.) and [email protected] (M.M.); phone: +33 540175027; fax:+33 559407781).NotesThe authors declare no competing financial interest.
■ ACKNOWLEDGMENTSFinancial support from Agence National de la Recherche (CES2008013) is acknowledged.
■ REFERENCES(1) Compeau, G. C.; Bartha, R. Sulfate-reducing bacteria: Principalmethylators of mercury in anoxic estuarine sediment. Appl. Environ.Microbiol. 1985, 50 (2), 498−502.(2) King, J. K.; Kostka, J. E.; Frischer, M. E.; Saunders, F. M. Sulfate-reducing bacteria methylate mercury at variable rates in pure cultureand in marine sediments. Appl. Environ. Microbiol. 2000, 66 (6), 2430−2437.(3) Stein, E. D.; Cohen, Y.; Winer, A. M. Environmental distributionand transformation of mercury compounds. Crit. Rev. Environ. Sci.Technol. 1996, 26 (1), 1−43.
Environmental Science & Technology Article
dx.doi.org/10.1021/es302412q | Environ. Sci. Technol. 2012, 46, 11744−1175111749

(4) Sigalevich, P.; Cohen, Y. Oxygen-dependent growth of thesulfate-reducing bacterium Desulfovibrio oxyclinae in coculture withMarinobacter sp. strain MB in an aerated sulfate-depleted chemostat.Appl. Environ. Microbiol. 2000, 66 (11), 5019−5023.(5) Acha, D.; Hintelmann, H.; Yee, J. Importance of sulfate reducingbacteria in mercury methylation and demethylation in periphyton fromBolivian Amazon region. Chemosphere 2011, 82 (6), 911−916.(6) Jay, J. A.; Morel, F. M. M.; Hemond, H. F. Mercury speciation inthe presence of polysulfides. Environ. Sci. Technol. 2000, 34 (11),2196−2200.(7) Benoit, J. M.; Gilmour, C. C.; Mason, R. P. The influence ofsulfide on solid-phase mercury bioavailability for methylation by purecultures of Desulfobulbus propionicus (1pr3). Environ. Sci. Technol.2001, 35 (1), 127−132.(8) Kim, M.; Han, S.; Gieskes, J.; Deheyn, D. D. Importance oforganic matter lability for monomethylmercury production in sulfate-rich marine sediments. Sci. Total Environ. 2011, 409 (4), 778−784.(9) Avramescu, M. L.; Yumvihoze, E.; Hintelmann, H.; Ridal, J.;Fortin, D.; Lean, R. S. D., Biogeochemical factors influencing netmercury methylation in contaminated freshwater sediments from theSt. Lawrence River in Cornwall, Ontario, Canada. Sci. Total Environ.2011, 409 (5), 968−978.(10) Clarkson, T. W. Human toxicology of mercury. J. Trace ElementsExp. Med. 1998, 11 (2−3), 303−317.(11) Bridou, R.; Monperrus, M.; Gonzalez, P. R.; Guyoneaud, R.;Amouroux, D. Simultaneous determination of mercury methylationand demethylation capacities of various sulfate-reducing bacteria usingspecies-specific isotopic tracers. Environ. Toxicol. Chem. 2011, 30 (2),337−344.(12) Barkay, T.; Miller, S. M.; Summers, A. O. Bacterial mercuryresistance from atoms to ecosystems. FEMS Microbiol. Rev. 2003, 27(2−3), 355−384.(13) Barkay, T.; Wagner-Dobler, I. Microbial transformations ofmercury: Potentials, challenges, and achievements in controllingmercury toxicity in the environment. Adv. Appl. Microbiol. 2005, 57,1−52.(14) Marvin-Dipasquale, M.; Agee, J.; McGowan, C.; Oremland, R.S.; Thomas, M.; Krabbenhoft, D.; Gilmour, C. C. Methyl-mercurydegradation pathways: A comparison among three mercury impactedecosystems. Environ. Sci. Technol. 2000, 34 (23), 4908−4916.(15) Oremland, R. S.; Culbertson, C. W.; Winfrey, M. R.Methylmercury decomposition in sediments and bacterial cultures:Involvement of methanogens and sulfate reducers in oxidativedemethylation. Appl. Environ. Microbiol. 1991, 57 (1), 130−137.(16) Choi, S. C.; Chase, T., Jr; Bartha, R. Metabolic pathways leadingto mercury methylation in Desulfovibrio desulfuricans LS. Appl. Environ.Microbiol. 1994, 60 (11), 4072−4077.(17) Ekstrom, E. B.; Morel, F. M. M.; Benoit, J. M. Mercurymethylation independent of the acetyl-coenzyme a pathway in sulfate-reducing bacteria. Appl. Environ. Microbiol. 2003, 69 (9), 5414−5422.(18) Ranchou-Peyruse, M.; Monperrus, M.; Bridou, R.; Duran, R.;Amouroux, D.; Salvado, J. C.; Guyoneaud, R. Overview of mercurymethylation capacities among anaerobic bacteria including representa-tives of the sulphate-reducers: Implications for environmental studies.Geomicrobiol. J. 2009, 26 (1), 1−8.(19) Hintelmann, H.; Evans, R. D.; Villeneuve, J. Y. Measurement ofmercury methylation in sediments by using enriched stable mercuryisotopes combined with methylmercury determination by gaschromatography-inductively coupled plasma mass spectrometry. J.Anal. At. Spectrom. 1995, 10 (9), 619−624.(20) Rodríguez Martín-Doimeadios, R. C.; Tessier, E.; Amouroux,D.; Guyoneaud, R.; Duran, R.; Caumette, P.; Donard, O. F. X.Mercury methylation/demethylation and volatilization pathways inestuarine sediment slurries using species-specific enriched stableisotopes. Mar. Chem. 2004, 90 (1−4 SPEC. ISS.), 107−123.(21) Monperrus, M.; Tessier, E.; Amouroux, D.; Leynaert, A.;Huonnic, P.; Donard, O. F. X. Mercury methylation, demethylationand reduction rates in coastal and marine surface waters of theMediterranean Sea. Mar. Chem. 2007, 107 (1), 49−63.
(22) Rodríguez-Gonzalez, P.; Monperrus, M.; García Alonso, J. I.;Amouroux, D.; Donard, O. F. X. Comparison of different numericalapproaches for multiple spiking species-specific isotope dilutionanalysis exemplified by the determination of butyltin species insediments. J. Anal. At. Spectrom. 2007, 22 (11), 1373−1382.(23) Schaefer, J. K.; Morel, F. M. M. High methylation rates ofmercury bound to cysteine by Geobacter sulfurreducens. Nat. Geosci.2009, 2 (2), 123−126.(24) Benoit, J. M.; Gilmour, C. C.; Mason, R. P. Aspects ofbioavailability of mercury for methylation in pure cultures ofDesulfobulbus propionicus (1pr3). Appl. Environ. Microbiol. 2001, 67(1), 51−58.(25) Pfennig, N.; Truper, H. G. The family Chromatiaceae. In TheProkaryotes. A Handbook on the Biology of Bacteria. Ecophysiology,Isolation, Identification, Applications, 2nd ed.; Balows, A., Truper, H. G.,Dworkin, M., Harder, W., Schleifer, K.-H., Eds.; Springer: Berlin,Heidelberg & New York, 1992; pp 3200−3221.(26) Ranchou-Peyruse, A.; Herbert, R.; Caumette, P.; Guyoneaud, R.Comparison of cultivation-dependent and molecular methods forstudying the diversity of anoxygenic purple phototrophs in sedimentsof an eutrophic brackish lagoon. Environ. Microbiol. 2006, 8 (9), 1590−1599.(27) Brown, S. D.; Gilmour, C. C.; Kucken, A. M.; Wall, J. D.; Elias,D. A.; Brandt, C. C.; Podar, M.; Chertkov, O.; Held, B.; Bruce, D. C.;Detter, J. C.; Tapia, R.; Han, C. S.; Goodwin, L. A.; Cheng, J.-F.;Pitluck, S.; Woyke, T.; Mikhailova, N.; Ivanova, N. N.; Han, J.; Lucas,S.; Lapidus, A. L.; Land, M. L.; Hauser, L. J.; Palumbo, A. V. GenomeSequence of the Mercury-Methylating Strain Desulfovibrio desulfuricansND132. J. Bacteriol. 2011, 193 (8), 2078−2079.(28) Gilmour, C. C.; Elias, D. A.; Kucken, A. M.; Brown, S. D.;Palumbo, A. V.; Schadt, C. W.; Wall, J. D. Sulfate-reducing bacteriumDesulfovibrio desulfuricans ND132 as a model for understandingbacterial mercury methylation. Appl. Environ. Microbiol. 2011, 77 (12),3938−3951.(29) Weimer, P. J.; Vankavelaar, M. J.; Michel, C. B.; Ng, T. K. Effectof phosphate on the corrosion of carbon-steel and on the compositionof corrosion products in 2-stage continuous cultures of desulfovibriodesulfuricans. Appl. Environ. Microbiol. 1988, 54 (2), 386−396.(30) Rapp, B. J.; Wall, J. D. Genetic transfer in Desulfovibriodesulfuricans. Proc. Natl. Acad. Sci. 1987, 84 (24), 9128−9130.(31) Rapp-Giles, B. J.; Casalot, L.; English, R. S.; Ringbauer, J. A., Jr.;Dolla, A.; Wall, J. D. Cytochrome c3 mutants of Desulfovibriodesulfuricans. Appl. Environ. Microbiol. 2000, 66 (2), 671−677.(32) Rousset, M.; Casalot, L.; Rapp-Giles, B. J.; Dermoun, Z.; DePhilip, P.; Belaich, J. P.; Wall, J. D. New shuttle vectors for theintroduction of cloned DNA in Desulfovibrio. Plasmid 1998, 39 (2),114−122.(33) Voordouw, G.; Pollock, W. B.; Bruschi, M.; Guerlesquin, F.;Rapp-Giles, B. J.; Wall, J. D. Functional expression of Desulfovibriovulgaris Hildenborough cytochrome c3 in Desulfovibrio desulfuricansG200 after conjugational gene transfer from Escherichia coli. J. Bacteriol.1990, 172 (10), 6122−6126.(34) Rodríguez Martín-Doimeadios, R. C.; Stoichev, T.; Krupp, E.;Amouroux, D.; Holeman, M.; Donard, O. F. X. Micro-scalepreparation and characterization of isotopically enriched monome-thylmercury. Appl. Organomet. Chem. 2002, 16 (10), 610−615.(35) Rodriguez-Gonzalez, P.; Epov, V. N.; Bridou, R.; Tessier, E.;Guyoneaud, R.; Monperrus, M.; Amouroux, D. Species-specific stableisotope fractionation of mercury during Hg(II) methylation by ananaerobic bacteria (Desulfobulbus propionicus) under dark conditions.Environ. Sci. Technol. 2009, 43 (24), 9183−9188.(36) Schaefer, J. K.; Rocks, S. S.; Zheng, W.; Liang, L.; Gu, B.; Morel,F. M. M. Active transport, substrate specificity, and methylation ofHg(II) in anaerobic bacteria. Proc. Natl. Acad. Sci. U.S.A. 2011, 108(21), 8714−8719.(37) Landner, L. Biochemical Model for the Biological Methylationof Mercury suggested from Methylation Studies in vivo withNeurospora crassa. Nature 1971, 230 (5294), 452−454.
Environmental Science & Technology Article
dx.doi.org/10.1021/es302412q | Environ. Sci. Technol. 2012, 46, 11744−1175111750

(38) Cvetkovic, A.; Menon, A. L.; Thorgersen, M. P.; Scott, J. W.;Poole, F. L., II; Jenney, F. E., Jr.; Lancaster, W. A.; Praissman, J. L.;Shanmukh, S.; Vaccaro, B. J.; Trauger, S. A.; Kalisiak, E.; Apon, J. V.;Siuzdak, G.; Yannone, S. M.; Tainer, J. A.; Adams, M. W. W. Microbialmetalloproteomes are largely uncharacterized. Nature 2010, 466(7307), 779−782.
Environmental Science & Technology Article
dx.doi.org/10.1021/es302412q | Environ. Sci. Technol. 2012, 46, 11744−1175111751
Top Related
Copyright © 2022 FDOKUMEN

