







Bahasa
Halaman
Hukum


TMEM16F Forms a Ca2+-Activated CationChannel Required for Lipid Scramblingin Platelets during Blood CoagulationHuanghe Yang,1,3 Andrew Kim,1,3 Tovo David,2 Daniel Palmer,2 Taihao Jin,1 Jason Tien,1 Fen Huang,1 Tong Cheng,1
Shaun R. Coughlin,2 Yuh Nung Jan,1 and Lily Yeh Jan1,*1Departments of Physiology, Biochemistry, and Biophysics, Howard Hughes Medical Institute2Cardiovascular Research Institute
University of California, San Francisco, San Francisco, CA 94143, USA3These authors contributed equally to this work
*Correspondence: [email protected]
http://dx.doi.org/10.1016/j.cell.2012.07.036
SUMMARY
Collapse of membrane lipid asymmetry is a hallmarkof blood coagulation. TMEM16F of the TMEM16family that includes TMEM16A/B Ca2+-activated Cl�
channels (CaCCs) is linked to Scott syndrome withdeficient Ca2+-dependent lipid scrambling. Wegenerated TMEM16F knockout mice that exhibitbleeding defects and protection in an arterial throm-bosis model associated with platelet deficiency inCa2+-dependent phosphatidylserine exposure andprocoagulant activity and lack a Ca2+-activatedcation current in the platelet precursor megakaryo-cytes. Heterologous expression of TMEM16F gener-ates a small-conductance Ca2+-activated nonselec-tive cation (SCAN) current with subpicosiemenssingle-channel conductance rather than a CaCC.TMEM16F-SCAN channels permeate both monova-lent and divalent cations, including Ca2+, and exhibitsynergistic gating by Ca2+ and voltage. We furtherpinpointed a residue in the putative pore regionimportant for the cation versus anion selectivity ofTMEM16F-SCAN and TMEM16A-CaCC channels.This study thus identifies a Ca2+-activated channelpermeable to Ca2+ and critical for Ca2+-dependentscramblase activity during blood coagulation.
INTRODUCTION
Phospholipids in the plasma membrane are asymmetric with
phosphatidylcholine (PC) and sphingomyelin (SPH) predomi-
nantly in the outer leaflet, whereas phosphatidylethanolamine
(PE) and phosphatidylserine (PS) are enriched in the inner
leaflet. This asymmetry is maintained by substrate- and direc-
tion-specific transporters known as flippases and floppases
(Contreras et al., 2010; Pomorski and Menon, 2006; van Meer,
2011). A third and the least understood category of lipid trans-
porters is called scramblases, which can rapidly collapse the
lipid asymmetry in a Ca2+-dependent but energy-independent
fashion, resulting in the exposure of PS to the cell surface (Bev-
ers and Williamson, 2010; Sanyal and Menon, 2009; van Meer,
2011). This scramblase-mediated externalization of PS has
important biological consequences in platelets, highly special-
ized hematopoietic cells that play a vital role in hemostasis and
thrombosis. Platelets are accumulated at sites of vascular injury
and are activated by exposed subendothelial factors such as
collagen or by thrombin generated from the coagulation cascade
pathway (Furie and Furie, 2008). Subsequent rise in the cyto-
plasmic Ca2+ level triggers Ca2+-activated lipid scrambling,
thus exposing PS, which prompts the recruitment and activation
of blood clotting factors—including factor V, factor X, and pro-
thrombin—on the platelet surface (Bevers et al., 1983; Rosing
et al., 1985).
Scott syndrome is a rare bleeding disorder with a defect in the
Ca2+-dependent phospholipid scrambling in the platelets, as
well as erythrocytes and B lymphocyte cell lines derived from
the patient (Zwaal et al., 2004). The first Scott syndrome patient
reported in 1979 had frequent bouts of bleeding episodes asso-
ciated with trauma or surgery, such as tooth extraction and
tonsillectomy, and childbirth that subsequently required blood
transfusion (Weiss et al., 1979). The patient displayed normal
platelet counts and structure, platelet adhesion, platelet secre-
tion, and aggregation but had a marked defect in platelet pro-
coagulant activity due to reduction in the Ca2+-dependent PS
exposure, leading to reduced thrombin generation and defective
coagulation. Recently, Suzuki and colleagues reported that the
transmembrane protein with unknown function 16F (TMEM16F)
is essential for the Ca2+-dependent scramblase activity. Knock-
down of TMEM16F in a B cell line results in a decrease in the rate
of Ca2+ ionophore-induced PS and PE exposure, whereas
TMEM16F overexpression strongly enhances PS exposure (Su-
zuki et al., 2010). DNA sequence analysis further demonstrates
that Scott syndrome patients carry loss-of-function mutations
in both Tmem16F alleles (Castoldi et al., 2011; Suzuki et al.,
2010).
In the mammalian TMEM16 family (Figure 1A), TMEM16A
and TMEM16B are Ca2+-activated chloride channels (CaCCs)
Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc. 111

Figure 1. TMEM16F Knockout Mice Exhibit Deficiencies in Ca2+-Dependent Scramblase Activity in Platelets, Defects in Blood Coagulation,
and Protection against FeCl3-Induced Carotid Thrombosis
(A) Phylogenetic tree of the human TMEM16 family generated by ClustalW2 with default settings. Percentage of amino acid identity with TMEM16A is indicated in
parentheses. Scale bar, 0.1 nucleotide substitutions per site.
(B) Immunoblot analysis of TMEM16F expression in the platelets of wild-type (WT) and TMEM16F KO mice.
(C) Flow cytometry analysis of platelets from WT and TMEM16F KO mice treated with 0, 1, and 10 mM A23187 and FITC-Annexin-V for 30 min.
(D) Tissue-factor-induced thrombin generation curves for WT and TMEM16F KO mice.
(E) Endogenous thrombin potential (ETP, the area under the thrombin generation curve) of platelets from WT and TMEM16F KO mice induced by tissue factor.
*p < 0.05. Error bar represents SEM.
(F) Thrombin generation peak height from the thrombin generation curves induced by tissue factor. *p < 0.05. Error bar represents SEM.
(G) Bleeding time measured by a hemostatic challenge assay. **p < 0.005. n.s., not significant. Error bar represents SEM.
(H) Carotid artery thrombosis by the 4% FeCl3 injury model. Flow versus time after FeCl3 application is shown. p = 0.0002 log rank test.
See also Figure S1.
112 Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc.

(Caputo et al., 2008; Schroeder et al., 2008; Yang et al., 2008)
that are important for cellular functions ranging from fluid
secretion and neuronal excitability to cancer progression (Ayoub
et al., 2010; Huang et al., 2012; Stanich et al., 2011; Yang et al.,
2008). The unexpected association of TMEM16F with Scott
syndrome raises important questions about the function of
TMEM16F, including: (1) whether it forms an ion channel and, if
so, (2) what type of ion channel; (3) whether TMEM16F itself
can scramble membrane lipids; (4) whether knocking out the
mouse Tmem16F gene will cause defective blood coagulation;
and (5) whether TMEM16F function in vivo is also important for
thrombosis.
We have generated TMEM16F knockout mice exhibiting
impaired hemostasis and decreased PS exposure on the surface
of activated platelets, erythrocytes, and splenic B cells, resem-
bling the major defects of Scott syndrome. Additionally,
TMEM16F knockout mice showed a significant protection in
an in vivo thrombosis assay, highlighting the importance of
TMEM16F in the procoagulant state of platelets during thrombus
formation. Surprisingly, loss of TMEM16F function resulted in the
elimination of a Ca2+-dependent cationic current in the platelet
precursor megakaryocytes. Moreover, TMEM16F expressed in
HEK293 cells and Xenopus as well as Axolotl oocytes gave
rise to a small-conductance Ca2+-activated nonselective cation
(SCAN) channel more permeable to Ca2+ than other physio-
logical cations. Importantly, mutagenesis of TMEM16F resulted
in alterations of multiple properties, including ion selectivity
and Ca2+ sensitivity, further supporting our conclusion that
TMEM16F forms the SCAN channel. Thus, the discovery of
TMEM16F as a Ca2+-activated and Ca2+-permeable nonselec-
tive cation channel crucial for Ca2+-dependent lipid scrambling
provides a new mechanistic insight into the blood coagulation
pathway and a potential target for antithrombotic therapy.
RESULTS
TMEM16F Knockout Mice Are Defective inCa2+-Dependent Scramblase Activity and DisplayImpaired HemostasisTo explore the physiological functions of TMEM16F, we gener-
ated TMEM16F knockout (KO) mice by deleting the second
exon, leading to a frame-shift mutation of the Tmem16F gene
(Figures S1A–S1C available online) and the absence of
TMEM16F protein in their platelets (Figure 1B). Homozygotes
are viable, fertile, and without gross differences in morphology
in comparison with wild-type (WT) mice.
To examine the impact of TMEM16F on the Ca2+-dependent
scramblase activity, we performed flow cytometry using the
FITC-conjugated Annexin-V, a specific probe for PS exposure.
The Ca2+ ionophore A23187 induced PS exposure from plate-
lets, red blood cells (RBCs), and CD19+ splenic B cells but sig-
nificantly less PS exposure for those cells without TMEM16F
(Figures 1C and S1D–S1G). Similar to Scott syndrome patients,
TMEM16F KOmice also showed impaired platelet procoagulant
activity as measured by thrombin generation in platelet-rich
plasma after addition of tissue factor (Figures 1D–1F) or factor
XIIa (Figures S1H–S1J), whereas no significant difference was
detected in platelet-poor plasma (Figures S1K to S1M). More-
over, TMEM16F KO mice exhibited prolonged bleeding time
(167 ± 18 s) as compared with that of WT mice (97 ± 10 s,
p < 0.005) (Figure 1G). The bleeding time of heterozygotes was
normal, in agreement with the absence of disease manifestation
of the children of a Scott syndrome patient (Bevers et al., 1983).
To evaluate the effect of TMEM16F on thrombogenesis in vivo,
we used 4% ferric chloride (FeCl3) to induce occlusive thrombus
formation in the carotid artery. Whereas seven out of eight WT
mice showed cessation of flow within 20 min, all nine TMEM16F
KO mice were fully protected from occlusion (Figure 1H). These
findings underscore the important role of TMEM16F in mediating
PS exposure for procoagulant activity leading to both hemo-
stasis and thrombosis—a critical event in the arterial diseases
associated with myocardial infarction and stroke as well as
venous thromboembolic disorders (Furie and Furie, 2008).
A Ca2+- and Voltage-Sensitive Outwardly RectifyingCurrent from Megakaryocytes of WT Mice, but NotTMEM16F KO MiceFor electrophysiological studies of the endogenous TMEM16F
channel, bone marrow megakaryocytes proved to be an ideal
cell type because: (1) megakaryocytes are the immediate pre-
cursors of platelets and share many molecular similarities and
(2) megakaryocytes are easily recognizable among other bone
marrow cells by their large size, distinct morphology, and ability
to bind the megakaryocyte/platelet-specific cell surface marker
CD41 (Figure 2A). The current was recorded with 140 mM
NaCl on both sides of the inside-out membrane patch in order
to minimize contaminations from Ca2+-activated K+ channels
that are impermeable to both Cl� and Na+. We found that
membrane patches from megakaryocytes of WT mice, but not
TMEM16F KO mice, exhibited a Ca2+-dependent outwardly
rectifying current (Figures 2B–2E).
Heterologous Expression of TMEM16F Confirms thatTMEM16F Gives Rise to a Ca2+- and Voltage-ActivatedIon ChannelTo test whether TMEM16F forms a functional ion channel, we ex-
pressed the murine TMEM16F (mTMEM16F) in Axolotl oocytes
(Figure 3) and HEK293 cells (Figure S2). Without endogenous
CaCC (Figure 3B, left), Axolotl oocytes provide an expression
system that enabled expression cloning of CaCC and character-
ization of TMEM16A-CaCC and TMEM16B-CaCC (Schroeder
et al., 2008). The fluorescence signal indicative of surface ex-
pression of mTMEM16F fused with green fluorescence protein
(TMEM16F-eGFP) peaked around 20 days after oocyte injec-
tion with cRNA (Figure 3A). Raising the internal Ca2+ induced
time-dependent currents that were much larger at depolarized
membrane potentials in recordings from inside-out patches
from oocytes (Figure 3B and 3C) and mTMEM16F-transfected
HEK293 cells (Figures S2B and S2C). Unlike TMEM16A-CaCC,
which exhibits outward rectification at low internal Ca2+ concen-
trations but has a roughly linear current-voltage (I-V) relationship
at higher internal Ca2+ concentrations (Figure S3A)—a character-
istic of endogenous CaCC in many cell types (Hartzell et al.,
2005), the TMEM16F currents showed prominent outward recti-
fication even at high internal Ca2+ concentrations (Figures 2, 3
and S2). We have further verified that TMEM16F forms this ion
Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc. 113

Figure 2. A Ca2+-Activated, Outwardly Rec-
tifying Current Present in Megakaryocytes
of WT Mice Was Eliminated from TMEM16F
KO Mice
(A) Megakaryocytes are CD41+ and morphologi-
cally distinct from the other cell types in the mouse
bone marrow.
(B) Representative macroscopic current traces of
inside-out patches excised from megakaryocytes
of WTmice (top) and TMEM16F KOmice (bottom).
The patches were exposed to 100 mM [Ca2+]i with
symmetrical 140 mM NaCl. The currents were
elicited by voltage ramps from�80 to +80 mVwith
a rate (dV/dt) = 0.36 V/s.
(C) A Ca2+-sensitive, outwardly rectifying current
was observed in megakaryocytes of WT mice, but
not TMEM16F KO mice (p < 0.0001). I+80mV and
jI�80mVj are the absolute values of current ampli-
tudes at +80 and �80 mV, respectively. n equals
15 for WT and 14 for TMEM16F KO. Error bar
represent SEM.
(D) Representative macroscopic current traces of
inside-out patches excised from megakaryocytes
of WT (left) and TMEM16F KO (right) mice in the
absence or presence of 100 mM [Ca2+]i. Ca2+-
induced currents that were derived by subtracting the currents recorded with 0 [Ca2+]i from the currents recorded with 100 mM [Ca2+]i are shown at the bottom.
Testing potentials were from �80 mV to +100 mV with 20 mV increments. Both holding and repolarizing potentials were �80 mV. Dotted lines indicate the zero
current level. Both sides of the membrane patch were exposed to 140 mM NaCl solutions.
(E) I-V relationship of the subtracted peak current as shown in (D).
channel by performing mutagenesis studies reported in the later
part of this study.
TheTMEM16FChannel Is aCa2+-ActivatedNonselectiveCation Channel Rather Than a CaCCUnexpectedly, we found that the TMEM16F channel is not
a CaCC. By progressively reducing the NaCl concentration on
the cytoplasmic side of the inside-out oocyte membrane patch
while maintaining the external NaCl concentration at 140 mM,
we found a drastic difference between mTMEM16A-CaCC and
the mTMEM16F-induced current (Figures 4A, S3A, and S3B).
Whereas it caused a leftward shift of the reversal potential for
mTMEM16A-CaCC (Figure S3A) indicative of a chloride channel
with a permeability ratio PNa/PCl of�0.14 (Figure 4A), the reversal
potential for the mTMEM16F-induced current shifted to the right
instead (Figure S3B), revealing a permeability ratio PNa/PCl of�7
(Figure 4A).
To further validate that mTMEM16F gives rise to a Ca2+-acti-
vated cation channel, we recorded from membrane patches
excised from mTMEM16F-expressing Axolotl oocytes and
megakaryocytes of WT mice. The Ca2+- and voltage-activated,
time-dependent current persisted even with isotonic 140 mM
sodiummethanesulfonate (MES) on both sides of the membrane
(Figures 4B and 4C). Because the large anion MES is poorly
permeable through chloride channels (Frings et al., 2000), this
current is most likely carried by Na+ ions. The cation permeability
was confirmed by monitoring the reversal potential when the
intracellular solution was switched from 140 mM NaCl to
140 mM NaMES for TMEM16F-expressing Axolotl oocytes (Fig-
ure S3C) and for megakaryocytes with endogenous TMEM16F
channels (Figure S3D).
114 Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc.
To assess the permeability to other cations, we measured the
reversal potential under bi-ionic conditions for TMEM16F ex-
pressed in heterologous systems (Figures S3E, S3F, 4D, and
4E) and endogenous TMEM16F in megakaryocytes (Figures
S3Dand4D). TheTMEM16Fchannel is almost equally permeable
to Li+, Na+, K+, Rb+, and Cs+ but much more permeable to Ca2+
andBa2+ (Figure 4E) andhas a largepore (�6 A radius) permeable
to cations as large as N-methyl-D-glucamine (NMDG, Figures
4E, S3D, and S3F) and tetraethylammonium (TEA, Figure 4E).
Indeed, the outwardly rectifying currents persisted in solutions
containing NMDG or TEA as the cation and MES as the anion
on both sides of the membrane (Figures S3G and S3H) or when
both sides of the membrane patch were exposed to isotonic
70 mM Ca(MES)2 solutions (Figure 4F). Thus, TMEM16F expres-
sion yielded a Ca2+-activated nonselective cation (CAN) channel
that is more permeable to Ca2+ than monovalent cations.
We further expressed mTMEM16F in Xenopus oocytes, which
normally display CaCC, but not CAN, currents (Figure S4A), and
replaced Cl� with MES to eliminate the CaCC currents. The
mTMEM16F channels were readily expressed on the Xenopus
oocyte surface (Figure S4B), and the resulting current (Figures
S4C–S4H) resembled every aspect of the current in Axolotl
oocytes and HEK293 cells expressing TMEM16F as well as the
endogenous TMEM16F current in megakaryocytes: Ca2+ and
voltage activated and nonselectively permeable to large cations
and Ca2+.
TMEM16F Is a Small-Conductance Ca2+-ActivatedNonselective Cation ChannelA hallmark of CAN channels is a single-channel conductance of
15–35 pS (Teulon, 2000). Using noise analysis, we estimated the

Figure 3. Functional Expression of TMEM16F in Axolotl Oocytes
Elicits a Ca2+- and Voltage-Dependent Current that Resembles the
Endogenous TMEM16F Current in Megakaryocytes of WT Mice
(A) Surface expression of murine TMEM16F (mTMEM16F)-eGFP in an Axolotl
oocyte. The eGFP signal (green) in the representative confocal image is in
close proximity to the signal for Alexa-633-conjugated Concanavalin A (blue),
which selectively binds to glycoproteins in the vitelline membrane of the
Axolotl oocyte.
(B) Representative macroscopic current traces of inside-out patches excised
from uninjected and mTMEM16F-expressing Axolotl oocytes in the absence
or presence of 100 mM [Ca2+]i. Ca2+-induced currents that were derived by
subtracting the currents recorded with 0 [Ca2+]i from the currents recorded
with 100 mM [Ca2+]i are shown at the bottom. Testing potentials were from
�80 mV to +100 mV with 10 mV increments. Both holding and repolarizing
potentials were �80 mV. Dotted lines indicate the zero current level. Both
sides of the membrane patch were exposed to 140 mM NaCl solutions.
(C) I-V relationship of the subtracted peak current shown in (B).
See also Figure S2.
single channel conductance of TMEM16F channels (Figures 5A
and5B) to beg=0.45± 0.07 pS (n = 15). The small single-channel
conductance, as well as the high Ca2+ selectivity and the
synergistic gating by Ca2+ and voltage, distinguishes TMEM16F
from most of the known CAN channels. We therefore named
TMEM16F channels as ‘‘SCAN channels,’’ for ‘‘small-conduc-
tance Ca2+-activated nonselective cation channels.’’
The TMEM16F-SCAN channel is also distinct from the known
CAN andCaCC channels in its pharmacological profile. Whereas
cytoplasmic application of 300 mM niflumic acid (NFA) readily
blocked the TMEM16A-CaCC current, it had no effect on
TMEM16F-SCAN current in Axolotl oocytes, Xenopus oocytes,
or HEK293 cells (Figures 5C, 5D, S5A, S5C, and S5D). Inclusion
of NFA in the external solution also failed to block TMEM16F-
SCAN channels (Figure S5A) even though this same treatment
readily blocked the endogenous CaCC in Xenopus oocytes
(data not shown). Likewise, two other common blockers for
both CaCC and CAN channels (Hartzell et al., 2005; Teulon,
2000; Ullrich et al., 2005), flufenamic acid (FFA) and 5-nitro-2-
(3-phenylpropylamino)-benzoate (NPPB), blocked the endoge-
nous CaCC in Xenopus oocytes, but not the TMEM16F-SCAN
channels expressed in Axolotl or Xenopus oocytes (Figures
S5B and S5C). We also tested the effects of nucleotides, which
inhibit a subset of the CAN channels (Nilius et al., 2004; Teulon,
2000; Ullrich et al., 2005). Cytoplasmic application of 2 mM
AMP or ATP did not inhibit the TMEM16F-SCAN current (Fig-
ure 5E). We further tested the effects of some nonselective
cation channel blockers (Pena and Ordaz, 2008) and found
that cytoplasmic application of 100 mM Ruthenium red (Ru-
Red), 2-Aminoethyl diphenylborinate (2APB), Cd2+, or Gd3+ in-
hibited the mTMEM16F channels, whereas N-Phenylanthranilic
acid (NPC) and SKF96365 had no effect (Figure S5D). Thus,
the TMEM16F-SCAN channel can be blocked by some non-
specific cation channel blockers such as Gd3+, Cd2+, 2APB,
and Ruthenium red but is pharmacologically distinct from the
CaCC channels and known CAN channels that are sensitive to
NFA, FFA, NPPB, DPC, or nucleotides.
Identification of a Residue Important for the Cationversus Anion Selectivity of TMEM16F-SCAN andTMEM16A-CaCC ChannelsIn a search for residues that are different in the putative pore
region of TMEM16F-SCAN and TMEM16A-CaCC channels,
we found a position in the transmembrane segment TM5 that
is occupied by lysine in all known anion channels, namely
TMEM16A-CaCC and TMEM16B-CaCC. In contrast, this posi-
tion is occupied by glutamine in the cation channel (Q559 in
mTMEM16F-SCAN) (Figure 6A). With lysine substitution for
Q559, the Q559K mutant mTMEM16F channel exhibited much
slower activation kinetics (Figures 6B and 6C) and was less
selective to Na+ (PNa/PCl = 2.2 compared to WT control of PNa/
PCl = 6.8) (Figures 6D, 6F, and 6G). In contrast, the K584Qmutant
mTMEM16A channel was less permeable to Cl� (PCl/PNa = 4.1
compared toWT control of PCl/PNa = 7.0) (Figures 6E–6G). These
mutagenesis experiments have therefore identified a key residue
near the N-terminal end of TM5 that is important for the ion selec-
tivity of two closely related TMEM16 channel family members
that form channels with preference for ions of opposite charge.
Characterization of the Ca2+- and Voltage-DependentActivation of TMEM16F-SCAN ChannelsWe found that increasing internal Ca2+ accelerated channel
activation and increased channel open probability (Figure 7A)
and caused a shift of the G-V curve for the voltage dependence
of the mTMEM16F channels expressed in HEK293 cells to
more negative voltages (Figures 7B and S6F and Table S1).
The apparent Ca2+ dissociation constant (KD) dropped from
10.8 mM to 3.4 mM (an increase in Ca2+ sensitivity) (p < 0.01)
when the membrane potential was depolarized from +60
to +180 mV (Figure 7C). This synergistic gating of mTMEM16F
channels by internal Ca2+ and voltage bears some resemblance
to the gating of the BK-type Ca2+-activated K+ channels that
belong to the family of voltage-gated K+ channels (Magleby,
2003), except for the apparent requirement for internal Ca2+
to activate TMEM16F channels—a protein with eight trans-
membrane segments without any canonical voltage-sensing
domains. The Hill coefficients for Ca2+ activation of TMEM16F
channels are between 2 and 3, suggesting that multiple Ca2+
ions may bind to the channel and they work cooperatively to
activate the channel.
Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc. 115
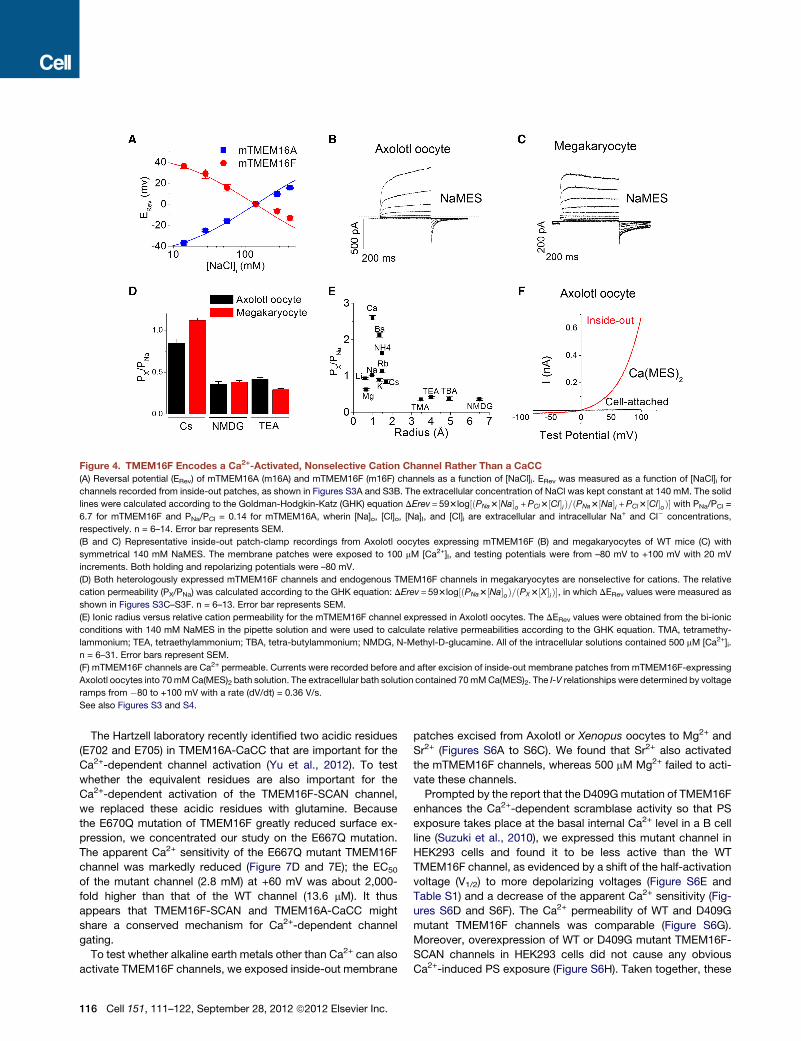
Figure 4. TMEM16F Encodes a Ca2+-Activated, Nonselective Cation Channel Rather Than a CaCC
(A) Reversal potential (ERev) of mTMEM16A (m16A) and mTMEM16F (m16F) channels as a function of [NaCl]i. ERev was measured as a function of [NaCl]i for
channels recorded from inside-out patches, as shown in Figures S3A and S3B. The extracellular concentration of NaCl was kept constant at 140 mM. The solid
lines were calculated according to the Goldman-Hodgkin-Katz (GHK) equation DErev = 593log½ðPNa3½Na�o +PCl3½Cl�iÞ=ðPNa3½Na�i +PCl3½Cl�oÞ� with PNa/PCl =
6.7 for mTMEM16F and PNa/PCl = 0.14 for mTMEM16A, wherin [Na]o, [Cl]o, [Na]I, and [Cl]i are extracellular and intracellular Na+ and Cl� concentrations,
respectively. n = 6–14. Error bar represents SEM.
(B and C) Representative inside-out patch-clamp recordings from Axolotl oocytes expressing mTMEM16F (B) and megakaryocytes of WT mice (C) with
symmetrical 140 mM NaMES. The membrane patches were exposed to 100 mM [Ca2+]I, and testing potentials were from –80 mV to +100 mV with 20 mV
increments. Both holding and repolarizing potentials were –80 mV.
(D) Both heterologously expressed mTMEM16F channels and endogenous TMEM16F channels in megakaryocytes are nonselective for cations. The relative
cation permeability (PX/PNa) was calculated according to the GHK equation: DErev = 593log½ðPNa3½Na�oÞ=ðPX3½X�iÞ�, in which DERev values were measured as
shown in Figures S3C–S3F. n = 6–13. Error bar represents SEM.
(E) Ionic radius versus relative cation permeability for the mTMEM16F channel expressed in Axolotl oocytes. The DERev values were obtained from the bi-ionic
conditions with 140 mM NaMES in the pipette solution and were used to calculate relative permeabilities according to the GHK equation. TMA, tetramethy-
lammonium; TEA, tetraethylammonium; TBA, tetra-butylammonium; NMDG, N-Methyl-D-glucamine. All of the intracellular solutions contained 500 mM [Ca2+]i.
n = 6–31. Error bars represent SEM.
(F) mTMEM16F channels are Ca2+ permeable. Currents were recorded before and after excision of inside-out membrane patches from mTMEM16F-expressing
Axolotl oocytes into 70mMCa(MES)2 bath solution. The extracellular bath solution contained 70mMCa(MES)2. The I-V relationships were determined by voltage
ramps from �80 to +100 mV with a rate (dV/dt) = 0.36 V/s.
See also Figures S3 and S4.
The Hartzell laboratory recently identified two acidic residues
(E702 and E705) in TMEM16A-CaCC that are important for the
Ca2+-dependent channel activation (Yu et al., 2012). To test
whether the equivalent residues are also important for the
Ca2+-dependent activation of the TMEM16F-SCAN channel,
we replaced these acidic residues with glutamine. Because
the E670Q mutation of TMEM16F greatly reduced surface ex-
pression, we concentrated our study on the E667Q mutation.
The apparent Ca2+ sensitivity of the E667Q mutant TMEM16F
channel was markedly reduced (Figure 7D and 7E); the EC50
of the mutant channel (2.8 mM) at +60 mV was about 2,000-
fold higher than that of the WT channel (13.6 mM). It thus
appears that TMEM16F-SCAN and TMEM16A-CaCC might
share a conserved mechanism for Ca2+-dependent channel
gating.
To test whether alkaline earth metals other than Ca2+ can also
activate TMEM16F channels, we exposed inside-out membrane
116 Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc.
patches excised from Axolotl or Xenopus oocytes to Mg2+ and
Sr2+ (Figures S6A to S6C). We found that Sr2+ also activated
the mTMEM16F channels, whereas 500 mM Mg2+ failed to acti-
vate these channels.
Prompted by the report that the D409Gmutation of TMEM16F
enhances the Ca2+-dependent scramblase activity so that PS
exposure takes place at the basal internal Ca2+ level in a B cell
line (Suzuki et al., 2010), we expressed this mutant channel in
HEK293 cells and found it to be less active than the WT
TMEM16F channel, as evidenced by a shift of the half-activation
voltage (V1/2) to more depolarizing voltages (Figure S6E and
Table S1) and a decrease of the apparent Ca2+ sensitivity (Fig-
ures S6D and S6F). The Ca2+ permeability of WT and D409G
mutant TMEM16F channels was comparable (Figure S6G).
Moreover, overexpression of WT or D409G mutant TMEM16F-
SCAN channels in HEK293 cells did not cause any obvious
Ca2+-induced PS exposure (Figure S6H). Taken together, these

Figure 5. mTMEM16F Gives Rise to Small-Conductance Ca2+-Acti-
vated, Nonselective Cation Channels with Distinct PharmacologicalProperties
(A) Noise analysis of mTMEM16F currents expressed in HEK293 cells. (i)
Representative current traces recorded in the presence of various [Ca2+]iat +80 mV. (ii) 2 Hz high-pass-filtered current traces shown in (i). (iii) Enlarged
high-pass-filtered current traces with 2 s duration under each [Ca2+]i.
(B) The variance of the current traces shown in (A) was plotted against the
current amplitudes and fitted with s2 = IVmg� I2=N. The obtained fitting
parameters were g = 0.47 pS and N = 67,006. The averaged channel
conductance obtained from 15 different patches was g = 0.45 ± 0.07 pS.
(C) 300 mM intracellular niflumic acid (NFA) blocked most of the mTMEM16A
Cl- currents, but not the mTMEM16F Na+ currents. Inside-out patches from
Axolotl oocytes were exposed to symmetrical 140 mM NaCl and 100 mM
[Ca2+]i for mTMEM16A and symmetrical 140 mM NaMES and 100 mM [Ca2+]ifor mTMEM16F. Currents were elicited by voltage pulses stepped from �80
to +80 mV.
(D) Summary of the NFA effects shown in (C). INFA and IO are the maximum
peak current amplitudes in the presence and absence of NFA, respectively.
n = 7–8. ***p < 0.0001. Error bars represent SEM.
(E) 2 mM free [AMP]i and [ATP]i had little effect on the mTMEM16F currents.
Inside-out patches from Axolotl oocytes were exposed to symmetrical
140 mM NaMES, 100 mM [Ca2+]I, and AMP or ATP. IO is the maximum cur-
rent amplitude in the absence of the nucleotides. n = 6–7. Error bar repre-
sents SEM.
See also Figure S5.
observations are compatible with the scenario that some
factors in blood cells act together with TMEM16F to mediate
the Ca2+-dependent scramblase activity.
Interestingly, the apparent Ca2+ sensitivity of the endogenous
TMEM16F channels in megakaryocytes is greater than that of
heterologously expressed mTMEM16F channels at certain volt-
ages, as evidenced by the leftward shift of the Ca2+ dose-
response curve (Figure 7F and 7G). The greater Ca2+ sensitivity
of the endogenous TMEM16F channels further suggests that
there may be regulatory factors in native cells that make the
endogenous TMEM16F channels more active.
DISCUSSION
In this study, we report that TMEM16F, which is linked to Scott
syndrome and belongs to the TMEM16 family that includes
TMEM16A and TMEM16B CaCCs, forms a SCAN channel with
considerable Ca2+ selectivity—an unprecedented finding of a
cation channel as a closely related family member of anion chan-
nels.We have further generated TMEM16FKOmice to show that
they recapitulate the defective Ca2+-activated scramblase
activity in platelets, B cells, and red blood cells that are charac-
teristic of Scott syndrome. Not only is the reduction of PS expo-
sure associated with prolonged bleeding, the loss of TMEM16F
function greatly reduced the propensity to thrombosis.
TMEM16F Forms a SCAN ChannelOur conclusion that TMEM16F gives rise to SCAN channels is
supported by the following evidence: (1) SCAN channels are
generated by expressing TMEM16F in three different heterolo-
gous expression systems; (2) mutation of a residue in the
transmembrane domain of TMEM16F alters SCAN channel
properties, including ion selectivity; (3) mutation of an acidic
residue of TMEM16F that corresponds to a putative Ca2+ binding
residue in TMEM16A-CaCC reduces the Ca2+ sensitivity of
SCAN channels; and (4) an endogenous Ca2+-activated cation
channel with similar properties is present in megakaryocytes
from WT, but not TMEM16F KO mice.
The residue that is important for the cation selectivity of
TMEM16F-SCAN and the anion selectivity of TMEM16A-CaCC
resides near the N-terminal end of the fifth transmembrane
segment (TM5) in the putative pore region; swapping this residue
rendered the Q559K-TMEM16F channel more permeable to Cl�
and the K584Q-TMEM16A channel more permeable to Na+.
Unlike themutagenesis of this key residue identified in our study,
mutations reported to alter ion selectivity in previous studies
target residues that are identical in TMEM16A and TMEM16F
(Yang et al., 2008), a finding that has not been replicated in a
recent study (Yu et al., 2012).
Features that distinguish TMEM16F-SCAN channels from
most of the Ca2+-activated nonselective cation (CAN) channels
(Teulon, 2000) include the ability to permeate Ca2+ ions (PCa/
PNa = 2.6), the small single-channel conductance (�0.45 pS),
the synergistic gating by Ca2+ and voltage, and the distinct phar-
macological characteristics. Although TRPM4 and TRPM5, two
recently identified CAN channels (Hofmann et al., 2003; Launay
et al., 2002; Prawitt et al., 2003), also show some voltage depen-
dence, they are clearly different from the TMEM16F-SCAN
channels in other key aspects: TRPM4-CAN and TRPM5-CAN
channels are impermeable to divalent cations, have much larger
single-channel conductance (15–25 pS), and are sensitive to
Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc. 117

Figure 6. A Residue in the Transmembrane Segment 5 Is Important for the Ion Selectivity of mTMEM16F-SCAN and mTMEM16A-CaCC
Channels(A) Sequence alignment of the transmembrane segment 5 (TM5) of human TMEM16A (h16A), murine TMEM16A (m16A), Xenopus TMEM16A (x16A), murine
TMEM16B (m16B), and murine TMEM16F (m16F).
(B) Representative macroscopic current traces of WT and Q559Kmutant mTMEM16F channels expressed in HEK293 cell. The inside-out patches were exposed
to 100 mM [Ca2+]i and symmetrical 140 mM NaCl. Testing potentials were from –80 mV to +180 mV with 20 mV increments. Both holding and repolarizing
potentials were –80 mV.
(C) Semi-log plots of mean activation time constants (t) as a function of voltage for the WT and Q559K-mTMEM16F channels. t was derived from the single
exponential fitting of the current traces shown in (B). n = 5–7. Error bar represents SEM.
(D and E) Representative I-V relationships of the Q559K-mTMEM16F (D) and K584Q-mTMEM16A (E) mutant channels recorded from inside-out patches exposed
to 14 and 140 mM [NaCl]i and 500 mM [Ca2+]i.
(F andG) Summary of the reversal potential shift (DERev) (F) and relative ion permeability (G) for theWT andmutant channelswhen [NaCl]i switched from 140mM to
14 mM. n = 8–19. ***p < 0.0001. Error bar represents SEM.
All of these experiments were recordings from inside-out membrane patches from HEK293 cells.
intracellular adenosine nucleotides and FFA (Hofmann et al.,
2003; Prawitt et al., 2003; Ullrich et al., 2005).
Conflicting conclusions regarding the biophysical properties
of TMEM16F have been drawn in recent studies. Three papers
from the same laboratory identified TMEM16F as a volume-regu-
lated anion channel (Almaca et al., 2009), a Ca2+-activated chlo-
ride channel (Schreiber et al., 2010), and an outwardly rectifying
chloride channel (Martins et al., 2011). These three types of
channels are different from one another, though they are all
anion channels. In contrast, our studies of TMEM16F channels
in megakaryocytes and three different expression systems
clearly reveal that TMEM16F forms a small-conductance Ca2+-
activated nonselective cation channel with minimal anion per-
meability (PNa/PCl is �7). Another study reports an inability of
118 Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc.
TMEM16F to target to the HEK293 cell membrane and a lack
of current detected using whole-cell patch clamp (Duran et al.,
2012), which may be accounted for by the lower intracellular
Ca2+ concentration used in that study. The identification in our
study of a residue that is important for the ion selectivity and
kinetic properties of the TMEM16F-SCAN channel further sup-
ports the notion that TMEM16F is a bona fide ion channel that
prefers permeating cations, including Ca2+.
TMEM16F Is Important for Ca2+Dependent PS Exposurefor Coagulation and ThrombosisFollowing studies of Scott syndrome patients and immortalized
blood cell lines that led to the identification of TMEM16F as the
culprit (Castoldi et al., 2011; Suzuki et al., 2010), we have

Figure 7. Gating of Heterologously Expressed mTMEM16F and
Endogenous TMEM16F Channels in Megakaryocytes Is Ca2+ and
Voltage Dependent
(A) Representative macroscopic current traces of an inside-out patch excised
from a mTMEM16F-expressing HEK293 cell exposed to 1, 2, 7, and 100 mM
[Ca2+]i. Testing potentials were from �80 mV to +180 mV with 20 mV incre-
ments. Both holding and repolarizing potentials were �80 mV.
(B) Mean G-V relations of the mTMEM16F channels under different [Ca2+]i.
Relative conductance was determined by measuring the amplitude of tail
currents 400 ms after repolarization to a fixed membrane potential (�80 mV).
The smooth curves represent Boltzmann fits I=IMax = 1=ð1+ expð�zeðV�V1=2Þ=kTÞÞ (see Table S1). IMax, tail current amplitude in response to deploar-
izaton to +180 mV in 100 mM [Ca2+]i. Error bar represents SEM.
(C) Ca2+ dose-response of the mTMEM16F channel at +60, +120,
and +180mV. I/IMax values were from (B). The smooth curves represent the fits
to the Hill equation: I=IMax =Amp=ð1+ ðKD=½Ca�ÞHÞ, wherein KD is the apparent
dissociation constant, H is the Hill coefficient, and Amp is the maximum value
of I/IMax at certain voltage. +60 mV: (KD = 10.8 ± 4.6 mM, H = 2.8); +120 mV:
(KD = 5.4 ± 1.6 mM, H = 1.5); +180 mV: (KD = 3.4 ± 0.5 mM, H = 1.8). n = 5–21.
Error bar represents SEM.
(D) Representative inside-out patch-clamp recordings of WT (top) and E667Q
mutant (bottom) mTMEM16F channels expressed in HEK293 cells. The
generated TMEM16F KO mice to demonstrate that TMEM16F is
important for the Ca2+-dependent scramblase activity in blood
cells and for thrombin generation. Not only do TMEM16F KO
mice show impaired hemostasis as evidenced by the prolonged
bleeding time, our study of carotid artery thrombus formation
has revealed that TMEM16F is also crucial for thrombosis. These
TMEM16F KOmice thus provide a suitable mouse model to help
understand the physiological and pathophysiological mecha-
nism of hemostasis and thrombosis.
How might the TMEM16F-SCAN channel contribute to the
Ca2+-dependent scramblase activity? There are conceivably
three possible scenarios. First, the influx of Ca2+ through
TMEM16F-SCAN channels may directly activate a yet undefined
scramblase that may be associated with TMEM16F-SCAN
channels. Ca2+ influx through Ca2+-permeable channels, such
as store-operated Ca2+ channels (Varga-Szabo et al., 2011)
and nucleotide-activated P2X1 channels (Mahaut-Smith et al.,
2011) that can promote lipid scrambling in platelets, could
increase the local Ca2+ concentrations to activate TMEM16F-
SCAN channels, which permeate Ca2+, thereby triggering a
positive feedback to facilitate Ca2+ entry, and their very small
single-channel conductance could limit the extent of Ca2+ influx,
thereby reducing energy expenditure and the risk of toxicity.
Second, TMEM16F-SCAN channels may act as a Ca2+-sensitive
regulator of an undefined scramblase; in this scenario, removal
of TMEM16F genetically could impair the efficiency of scram-
blase activation or compromise scramblase maturation or traffic
to the cell membrane. Third, the TMEM16F-SCAN channel may
be a multitasking or multifunctional moonlighting protein such
as opsin (Menon et al., 2011) and can carry out two distinct tasks:
transporting ions and scrambling phospholipids. Conformational
changes induced by Ca2+ binding to the TMEM16F-SCAN
channel might facilitate the formation of transient membrane
defects such as water pores in the lipid-channel interface. These
transient membrane defects could facilitate fast translocation of
the various charged headgroups of phospholipids across the
membrane, thereby disrupting the membrane lipid asymmetry
(Gurtovenko et al., 2010; Sanyal and Menon, 2009). Our study
thus far is consistent with the hypothesis that TMEM16F is
membrane patches were voltage clamped at +60 mV and exposed to different
[Ca2+]i.
(E) Comparison of Ca2+ sensitivity between the WT and E667Q mutant
TMEM16F channels at +60 mV. The smooth curves represent the fits to the Hill
equation. WT: KD = 13.6 ± 1.8 mM, H = 2.2 ± 0.5; E667Q: KD = 2.8 ± 0.3 mM,
H = 1.6 ± 0.3. n = 11–17. Error bar represents SEM.
(F) Representative inside-out patch-clamp recordings from the mTMEM16F-
expressing HEK293 cells (top) and megakaryocytes of WT mice (bottom). The
membrane patches were voltage clamped at +60 mV and exposed to different
[Ca2+]i.
(G) Comparison of the Ca2+ sensitivity of the endogenous TMEM16F-SCAN
current in megakaryocytes with that of the heterologously expressed
TMEM16F channels in HEK293 cells at +60 mV. The smooth curves represent
the fits to the Hill equation. For mTMEM16F currents in HEK293 cells, KD =
13.9 ± 2.9 mM, H = 1.6; for endogenous TMEM16F currents in megakaryo-
cytes, KD = 5.1 ± 2.0 mM, H = 2.5. n = 6–12. Error bar represents SEM.
All experiments were recordings from inside-out membrane patches in sym-
metrical 140 mM NaMES solutions except for the experiments shown in (D)
and (E), which were done in symmetrical 140 mM NaCl solutions.
See also Figure S6 and Table S1.
Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc. 119

important, but not sufficient, for the Ca2+-dependent scramblase
activity.
Given the uncertainty of molecular identity of scramblase and
the complexity of the lipid scrambling machinery in blood cells
(Bevers and Williamson, 2010), it is difficult to pinpoint the exact
role of the TMEM16F-SCAN channel in lipid scrambling. Never-
theless, a few insights can be gained from our current study: (1)
The platelets of TMEM16F KOmice exhibit significantly reduced
but still substantial Ca2+-induced PS exposure when treated
with high concentrations of the Ca2+ ionophore A23187 (Fig-
ures 1C, S1E, and S1F). This implies that either the Ca2+ influx
through TMEM16F-SCAN channels or conformational changes
during channel activation directly activates an unknown scram-
blase; or alternatively, if TMEM16F itself is one of the scram-
blases, there might be other unknown scramblase(s) mediating
the remaining Ca2+-induced PS exposure in platelets without
TMEM16F. (2) In contrast to the enhanced scramblase activity
when TMEM16F is expressed in Ba/F3 cells (Suzuki et al.,
2010) or lymphoma cells (Segawa et al., 2011), overexpression
ofWT or D409Gmutant TMEM16F in HEK293 cells did not cause
Ca2+-induced PS exposure (Figure S6H), though it remains
possible that cell-type-specific regulation renders TMEM16F
capable of translocating phospholipids in blood cells. (3) Al-
though the D409G mutant TMEM16F drastically increases the
Ca2+ sensitivity of lipid scrambling when expressed in a B cell
line (Suzuki et al., 2010), the Ca2+ sensitivity and Ca2+ per-
meability of D409G mutant TMEM16F channels expressed in
HEK293 cells are not enhanced (Figure S6). It seems more likely
that the D409G mutation facilitates PS exposure by altering
TMEM16F interaction with other components of a putative
scramblase complex or by enhancing the efficiency of TMEM16F
to scramble phospholipids.
Consistent with our studies of HEK293 cells, many nonhema-
topoietic cell lines, including CHO, BHK, 3T3, MCF-7, and HeLa,
display no Ca2+-activated scramblase activity (Williamson et al.,
2007). Because TMEM16F is expressed in many different mouse
tissues (Schreiber et al., 2010), it remains to be examined
whether nonhematopoietic primary cells that express TMEM16F
exhibit Ca2+-induced PS exposure and whether other functions
unrelated to lipid scrambling are altered in TMEM16F KO mice.
In conclusion, our finding of TMEM16F as a SCAN channel
with considerable Ca2+ permeability, the presence of functional
TMEM16F-SCAN channels in megakaryocytes, as well as the
generation of TMEM16F KO mice with reduced thrombosis
and blood coagulation deficits that are characteristic of Scott
syndrome will greatly facilitate future studies to elucidate the
molecular mechanism for phospholipid scrambling in blood
cells. The TMEM16F-SCAN channels discovered in this study
hence might serve as a new pharmaceutical target to treat
human hemostatic and thrombotic disorders, such as stroke
and heart attack.
EXPERIMENTAL PROCEDURES
Additional information regarding plasmids, TMEM16F KO mouse generation,
and detailed procedures for flow cytometry, electrophysiology, and in vivo
experiments are described in the Supplemental Information. All animal
procedures were approved by the UCSF Institutional Animal Care and Use
Committee and were performed according to the guidelines provided.
120 Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc.
Generation of TMEM16F Knockout Mice
Using recombineering technology, loxP site was inserted in the 30 intronicregion of exon 2 and a frt-neo-frt-loxP cassette was inserted in the 50 intronicregion of exon 2. Linearized targeting vector was electroporated into embry-
onic stem cells. G418 and Gancyclovir were used in the ES cell media to select
for ES cell clones with proper homologous recombination, which were
confirmed by Southern blot analysis. Correctly targeted ES cells were then
injected into blastocysts, and germline transmission was confirmed by PCR.
The resulting mouse was bred with b-Actin-Flp mouse to delete neomycin
cassette. Male b-Actin-Cre deleter mice were then bredwith female TMEM16F
flox/+ neomycin-deleted mice to generate founding TMEM16F heterozygous
mice. TMEM16F heterozygous mice (het) were crossed with each other to
generate TMEM16F KO mice. PCR genotyping was performed on tail DNA
extracted from such offsprings. All experimental mice were backcrossed
between two and five generations to C57Bl6.
Immunoblotting of Washed Mouse Platelet Lysates
Blood was collected from the vena cava of anesthetized mice in ACD (2.5 g
Trisodium citrate, 1.37 g citric acid monohydrate, 2.0 g dextrose in 100 ml
of ddH2O), mixed with 20 mM PIPES in saline (pH 6.5), and centrifuged at
1003 g in room temperature. The platelet-containing upper phase was mixed
with low-pH washing buffer (140 mM NaCl, 10 mM NaHCO3, 2.5 mM KCl,
0.5 mM Na2HPO4, 1 mM MgCl2, 22 mM Trisodium Citrate, 0.1% dextrose,
and 0.35% BSA [pH 6.5]) with 3 U/ml apyrase (Sigma) and then centrifuged
at 500 3 g at room temperature. Pelleted platelets were resuspended in
Ca2+-free Tyrode’s HEPES (CFTH) buffer (134 mM NaCl, 12 mM NaHCO3,
2.9 mM KCl, 0.34 mM Na2HPO4, 1.0 mM MgCl2, 10 mM HEPES, 0.9% wt/vol
dextrose, and 0.35% wt/vol BSA [pH 7.4]) then lysed using RIPA buffer.
Western blot analysis was performed using a rabbit polyclonal antibody raised
against a peptide corresponding to the N terminus of mouse TMEM16F pro-
tein. The b-actin antibody was used as loading control.
Flow Cytometric Analysis of PS Exposure
To assess the A23187 Ca2+ ionophore-induced PS exposure on the platelet
surface, 1 3 106 platelets were resuspended in 24 ml total volume of CFTH
buffer with 2mMCa2+ with or without A23187 (Sigma-Aldrich). This was imme-
diately followed by addition of 1 ml FITC-conjugated Annexin-V (BD Biosci-
ences) and incubation in 37�C water bath for 30 min. The incubation was
quenched using 500 ml of chilled CFTH buffer, and platelets were analyzed
using FACSCalibur (Becton Dickinson). Red blood cells were collected from
the lower phase following whole-blood centrifugation and were similarly
used for flow cytometric analysis.
For B cell analysis, mouse spleens from WT and KO mice were gently
mashed, filtered, and washed once using PBS. The cells (13 106) were resus-
pended in 1X Annexin-V binding buffer (BD Biosciences) and were incubated
with A23187 in 37�C water bath for 15 min. Cells were subsequently stained
using Annexin-V FITC and CD19 APC on ice for 15 min followed by 500 ml of
annexin-V binding buffer containing 7-AAD (1 mg/ml final).
Thrombin Generation Measured by a Thrombinoscope
Thrombin generation wasmeasured using a fluorogenic thrombin substrate on
a multiwell automated fluorescent plate reader (ThrombinoSCOPE, Maas-
tricht, The Netherlands).
Tail-Bleeding Assay
A small tip (1–2 mm) of the tail was transected and immersed in 37�C saline
water. The time for the bleeding to stop was measured up to 5 min. For
mice bleeding more than 5 min, pressure was applied near the wound to
stop the bleeding. The mice were genotyped after the experiment. Statistical
significance was determined with the Student’s t test.
FeCl3-Induced Carotid Thrombosis Model
Eight- to fourteen-week-old mice were anesthetized. The carotid was dis-
sected free, a flow probe was placed around the left common carotid artery
proximal to the bifurcation, and the artery was exposed to filter papers
soaked in FeCl3 for 3 min. Arterial flow was measured continuously until clot-
ting occurred (defined as no flow for R 2 min). If no stoppage occured,

measurements were ceased at 20 min. Percentage of arteries without occlu-
sion (i.e., with continued flow) at the end of the assay was determined.
Kaplan-Meier-type data were compared by log rank test.
Molecular Biology and Channel Expression
cRNA (5–100 ng) was injected into defolliculated oocytes. HEK293 cells were
transfected with corresponding plasmids using FuGENE6 (Roche) and were
cultured for 1–3 days before recording.
Confocal Imaging
Axolotl or Xenopus oocytes were stained with 50 mg/ml Alexa-633-conjugated
Concanavalin A for 15 min at room temperature. TMEM16F-eGFP proteins in
HEK293 cells were detected using Alexa-488-conjugated anti-rabbit antibody.
All images were acquired using a Leica SP5 laser confocal microscope.
Electrophysiology and Data Analysis
Macroscopic currents were recorded from inside-out patches. Data were
acquired using an Axopatch 200-B patch-clamp amplifier and pClamp9 soft-
ware (Molecular Devices). All experiments were performed at room tempera-
ture (22�C –24�C).To determine the reversal potentials while the NaCl concentration of the
internal solution (without buffer) was varied, the osmolality of each solution
was adjusted to about 330 mOsm/kg by addition of sucrose, except for the
280 mM and 420 mM NaCl solutions, which had higher osmolalities and con-
tained no sucrose. To minimize leak currents, patches with seal resistance
lower than 3 GU were discarded. Similar reversal potential values were ob-
tained by two different protocols (mTMEM16F in Figure 4A with the ramp
protocol and Figure 6F with the voltage-step protocol).
For noise analysis, the currents were recorded at +80 mV using a gap-free
protocol. The data were filtered at 1 KHz with an eight-pole low-pass Bessel
filter were and sampled at 5 KHz.
For megakaryocyte recording, we used a protocol adopted from that of
Mahaut-Smith (Mahaut-Smith, 2004).
Clampfit10 (Molecular Devices) and Origin (OriginLab) were used for data
analysis.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Extended Experimental Procedures, six
figures, and one table and can be found with this article online at http://dx.
doi.org/10.1016/j.cell.2012.07.036.
ACKNOWLEDGMENTS
We thank David Gorczyca for help with confocal imaging; Xiuming Wong for
help with cell culture; and Jianmin Cui, Ehud Isacoff, Min Li, Shi-Bing Yang,
James Berg, and Christian Peters for suggestions and comments on the
manuscript. We also thank Kevin Shannon and Ethan Weiss for sharing their
CBC machine, Thrombinoscope, and other equipment. This study was sup-
ported by the NIH grant HL65185 to S.R.C. and MH65334 to L.Y.J. Y.N.J.
and L.Y.J. are Howard Hughes Medical Institute investigators.
Received: February 27, 2012
Revised: June 12, 2012
Accepted: July 5, 2012
Published: September 27, 2012
REFERENCES
Almaca, J., Tian, Y., Aldehni, F., Ousingsawat, J., Kongsuphol, P., Rock, J.R.,
Harfe, B.D., Schreiber, R., and Kunzelmann, K. (2009). TMEM16 proteins
produce volume-regulated chloride currents that are reduced in mice lacking
TMEM16A. J. Biol. Chem. 284, 28571–28578.
Ayoub, C., Wasylyk, C., Li, Y., Thomas, E., Marisa, L., Robe, A., Roux,M., Abe-
cassis, J., de Reynies, A., and Wasylyk, B. (2010). ANO1 amplification and
expression in HNSCC with a high propensity for future distant metastasis
and its functions in HNSCC cell lines. Br. J. Cancer 103, 715–726.
Bevers, E.M., and Williamson, P.L. (2010). Phospholipid scramblase: an
update. FEBS Lett. 584, 2724–2730.
Bevers, E.M., Comfurius, P., and Zwaal, R.F. (1983). Changes in membrane
phospholipid distribution during platelet activation. Biochim. Biophys. Acta
736, 57–66.
Caputo, A., Caci, E., Ferrera, L., Pedemonte, N., Barsanti, C., Sondo, E.,
Pfeffer, U., Ravazzolo, R., Zegarra-Moran, O., and Galietta, L.J. (2008).
TMEM16A, a membrane protein associated with calcium-dependent chloride
channel activity. Science 322, 590–594.
Castoldi, E., Collins, P.W., Williamson, P.L., and Bevers, E.M. (2011).
Compound heterozygosity for 2 novel TMEM16F mutations in a patient with
Scott syndrome. Blood 117, 4399–4400.
Contreras, F.X., Sanchez-Magraner, L., Alonso, A., and Goni, F.M. (2010).
Transbilayer (flip-flop) lipid motion and lipid scrambling in membranes. FEBS
Lett. 584, 1779–1786.
Duran, C., Qu, Z., Osunkoya, A.O., Cui, Y., and Hartzell, H.C. (2012). ANOs 3-7
in the anoctamin/Tmem16 Cl- channel family are intracellular proteins. Am. J.
Physiol. Cell Physiol. 302, C482–C493.
Frings, S., Reuter, D., and Kleene, S.J. (2000). Neuronal Ca2+ -activated Cl-
channels—homing in on an elusive channel species. Prog. Neurobiol. 60,
247–289.
Furie, B., and Furie, B.C. (2008). Mechanisms of thrombus formation. N. Engl.
J. Med. 359, 938–949.
Gurtovenko, A.A., Anwar, J., and Vattulainen, I. (2010). Defect-mediated traf-
ficking across cell membranes: insights from in silico modeling. Chem. Rev.
110, 6077–6103.
Hartzell, C., Putzier, I., and Arreola, J. (2005). Calcium-activated chloride chan-
nels. Annu. Rev. Physiol. 67, 719–758.
Hofmann, T., Chubanov, V., Gudermann, T., and Montell, C. (2003). TRPM5
is a voltage-modulated and Ca(2+)-activated monovalent selective cation
channel. Curr. Biol. 13, 1153–1158.
Huang, W.C., Xiao, S., Huang, F., Harfe, B.D., Jan, Y.N., and Jan, L.Y. (2012).
Calcium-activated chloride channels (CaCCs) regulate action potential and
synaptic response in hippocampal neurons. Neuron 74, 179–192.
Launay, P., Fleig, A., Perraud, A.L., Scharenberg, A.M., Penner, R., and Kinet,
J.P. (2002). TRPM4 is a Ca2+-activated nonselective cation channel mediating
cell membrane depolarization. Cell 109, 397–407.
Magleby, K.L. (2003). Gating mechanism of BK (Slo1) channels: so near, yet so
far. J. Gen. Physiol. 121, 81–96.
Mahaut-Smith, M.P. (2004). Patch-clamp recordings of electrophysiological
events in the platelet and megakaryocyte. Methods Mol. Biol. 273, 277–300.
Mahaut-Smith, M.P., Jones, S., and Evans, R.J. (2011). The P2X1 receptor and
platelet function. Purinergic Signal. 7, 341–356.
Martins, J.R., Faria, D., Kongsuphol, P., Reisch, B., Schreiber, R., and Kunzel-
mann, K. (2011). Anoctamin 6 is an essential component of the outwardly recti-
fying chloride channel. Proc. Natl. Acad. Sci. USA 108, 18168–18172.
Menon, I., Huber, T., Sanyal, S., Banerjee, S., Barre, P., Canis, S., Warren, J.D.,
Hwa, J., Sakmar, T.P., and Menon, A.K. (2011). Opsin is a phospholipid flip-
pase. Curr. Biol. 21, 149–153.
Nilius, B., Prenen, J., Voets, T., and Droogmans, G. (2004). Intracellular nucle-
otides and polyamines inhibit the Ca2+-activated cation channel TRPM4b.
Pflugers Arch. 448, 70–75.
Pena, F., and Ordaz, B. (2008). Non-selective cation channel blockers: poten-
tial use in nervous system basic research and therapeutics. Mini Rev. Med.
Chem. 8, 812–819.
Pomorski, T., andMenon, A.K. (2006). Lipid flippases and their biological func-
tions. Cell. Mol. Life Sci. 63, 2908–2921.
Prawitt, D., Monteilh-Zoller, M.K., Brixel, L., Spangenberg, C., Zabel, B., Fleig,
A., and Penner, R. (2003). TRPM5 is a transient Ca2+-activated cation channel
Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc. 121

responding to rapid changes in [Ca2+]i. Proc. Natl. Acad. Sci. USA 100, 15166–
15171.
Rosing, J., van Rijn, J.L., Bevers, E.M., van Dieijen, G., Comfurius, P., and
Zwaal, R.F. (1985). The role of activated human platelets in prothrombin and
factor X activation. Blood 65, 319–332.
Sanyal, S., and Menon, A.K. (2009). Flipping lipids: why an’ what’s the reason
for? ACS Chem. Biol. 4, 895–909.
Schreiber, R., Uliyakina, I., Kongsuphol, P., Warth, R., Mirza, M., Martins, J.R.,
and Kunzelmann, K. (2010). Expression and function of epithelial anoctamins.
J. Biol. Chem. 285, 7838–7845.
Schroeder, B.C., Cheng, T., Jan, Y.N., and Jan, L.Y. (2008). Expression cloning
of TMEM16A as a calcium-activated chloride channel subunit. Cell 134, 1019–
1029.
Segawa, K., Suzuki, J., and Nagata, S. (2011). Constitutive exposure of phos-
phatidylserine on viable cells. Proc. Natl. Acad. Sci. USA 108, 19246–19251.
Stanich, J.E., Gibbons, S.J., Eisenman, S.T., Bardsley,M.R., Rock, J.R., Harfe,
B.D., Ordog, T., and Farrugia, G. (2011). Ano1 as a regulator of proliferation.
Am. J. Physiol. Gastrointest. Liver Physiol. 301, G1044–G1051.
Suzuki, J., Umeda, M., Sims, P.J., and Nagata, S. (2010). Calcium-dependent
phospholipid scrambling by TMEM16F. Nature 468, 834–838.
Teulon, J. (2000). Ca2+-activated non-selective cation channels. In Pharma-
cology of ionic channel function: activators and inhibitors, M. Endo, Y. Kurachi,
and M. Mishina, eds. (New York: Springer-Verlag), pp. 625–649.
122 Cell 151, 111–122, September 28, 2012 ª2012 Elsevier Inc.
Ullrich, N.D., Voets, T., Prenen, J., Vennekens, R., Talavera, K., Droogmans,
G., and Nilius, B. (2005). Comparison of functional properties of the Ca2+-
activated cation channels TRPM4 and TRPM5 from mice. Cell Calcium 37,
267–278.
van Meer, G. (2011). Dynamic transbilayer lipid asymmetry. Cold Spring Harb.
Perspect. Biol. 3, a004671.
Varga-Szabo, D., Braun, A., and Nieswandt, B. (2011). STIM and Orai in
platelet function. Cell Calcium 50, 270–278.
Weiss, H.J., Vicic, W.J., Lages, B.A., and Rogers, J. (1979). Isolated deficiency
of platelet procoagulant activity. Am. J. Med. 67, 206–213.
Williamson, P., Halleck, M.S., Malowitz, J., Ng, S., Fan, X., Krahling, S., Rema-
ley, A.T., and Schlegel, R.A. (2007). Transbilayer phospholipid movements in
ABCA1-deficient cells. PLoS ONE 2, e729.
Yang, Y.D., Cho, H., Koo, J.Y., Tak, M.H., Cho, Y., Shim, W.S., Park, S.P., Lee,
J., Lee, B., Kim, B.M., et al. (2008). TMEM16A confers receptor-activated
calcium-dependent chloride conductance. Nature 455, 1210–1215.
Yu, K., Duran, C., Qu, Z., Cui, Y.Y., and Hartzell, H.C. (2012). Explaining
calcium-dependent gating of anoctamin-1 chloride channels requires a revised
topology. Circ. Res. 110, 990–999.
Zwaal, R.F., Comfurius, P., and Bevers, E.M. (2004). Scott syndrome, a
bleeding disorder caused by defective scrambling of membrane phospho-
lipids. Biochim. Biophys. Acta 1636, 119–128.
Top Related
Copyright © 2022 FDOKUMEN

