







Bahasa
Halaman
Hukum


ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2014 Magnolia Press
Zootaxa 3815 (2): 215–232
www.mapress.com/zootaxa/Article
http://dx.doi.org/10.11646/zootaxa.3815.2.3
http://zoobank.org/urn:lsid:zoobank.org:pub:2A5EC5EA-EBC9-4E79-B8F0-1D17F1E97ECD
Taxonomic notes on some Brazilian species of Synalpheus Spence Bate, 1888,
with new records and description of a new species (Decapoda, Alpheidae)
ARTHUR ANKER1, 3 & PAULO P.G. PACHELLE2
1Tropical Marine Science Institute, National University of Singapore, 18 Kent Ridge Road, Singapore 119227, Republic of Singapore2Laboratório de Invertebrados Marinhos do Ceará, Departamento de Biologia, Universidade Federal do Ceará, Fortaleza, Ceará,
Brazil3Corresponding author. E-mail: [email protected]
Abstract
The present report deals with several western Atlantic species of the snapping shrimp genus Synalpheus Spence Bate, 1888
occurring in Brazilian territorial waters. Two species, Synalpheus cf. ruetzleri Macdonald & Duffy, 2006 and Synalpheus
yano (Ríos & Duffy, 2007), are recorded from the southwestern Atlantic Ocean and Brazil (Ceará) for the first time; S.
yano is also newly reported from the eastern Gulf of Mexico and USA (Florida). Synalpheus androsi Coutière, 1909, a
distinctive species previously known in Brazil only from a single unconfirmed record, is confirmed to be present in Bra-
zilian waters based on material from Bahia and Espírito Santo. Synalpheus tenuispina Coutière, 1909, a poorly known
species from southern Brazil, is redescribed. Synalpheus ul (Ríos & Duffy, 2007), previously known from Bahia and
Alagoas, is reported from Pernambuco based on older “Calypso” material misidentified as Synalpheus longicarpus (Her-
rick, 1891). A new species from the Synalpheus paraneptunus Coutière, 1909 complex is described as Synalpheus maxil-
lispinus sp. nov. based on three specimens dredged from moderate depths (15–55 m) off southern Bahia and Espírito
Santo, eastern Brazil. Finally, an incomplete specimen that appears to belong to a species presently unknown in Brazil is
preliminarily reported as Synalpheus sp. gambarelloides group. Illustrations are provided for all reported taxa except for
S. ul.
Key words: Atlantic Ocean, Brazil, Decapoda, snapping shrimp, Synalpheus, sponge, new record, new species, redescrip-
tion
Introduction
The speciose and taxonomically challenging snapping shrimp genus Synalpheus Spence Bate, 1888 is represented
in the Atlantic Ocean by over 60 species (De Grave & Fransen 2011; Anker et al. 2012). Most of them occur in the
tropical regions of the northwestern Atlantic, mainly in the Caribbean Sea, where most of the recent taxonomic
research on the genus has been done (e.g., Ríos & Duffy 2007; Anker & Tóth 2008; Macdonald et al. 2009;
Hultgren et al. 2010, 2011). In contrast, the synalpheid fauna of the southwestern Atlantic has not studied
exhaustively since Christoffersen (1979) and remains comparatively less known. The only noteworthy recent
studies dealing with the Brazilian material of Synalpheus are Coelho et al. (2006), Bezerra & Coelho (2006) and
Coelho Filho (2006), all simply listing several species, some of them as new records for Brazil; Almeida et al.
(2007, 2012) with an illustrated record of Synalpheus ul (Ríos & Duffy, 2007); and Anker et al. (2012), with
additional records (some new) for Brazil and illustrations of several species based on material collected in Atol das
Rocas. Anker et al. (2012) also commented on some morphologically variable taxa requiring taxonomic revisions,
including Synalpheus minus (Say, 1818), Synalpheus brevicarpus (Herrick, 1891), and Synalpheus apioceros
Coutière, 1909, all three of which occur in Brazil. These and other examples (Hultgren & Duffy 2012) show that
the taxonomy of the American species of Synalpheus is far from being settled.
In September–November 2013, the authors were able to study the unsorted caridean material deposited in the
crustacean collection of Museu de Zoologia, Universidade de São Paulo, São Paulo, Brazil (MZUSP). This
material contained numerous samples of Synalpheus, often in mixed lots together with other alpheid shrimps.
Accepted by J. W. Goy: 28 Apr. 2014; published: 13 Jun. 2014 215

Additional specimens examined for this study came from a small collection made by marine zoology students of
the Universidade Federal do Ceará (UFC), material from sponges reported by Bezerra & Coelho (2006) deposited
partly in UFC, samples from a short survey of São Sebastião harbour in 2009 (all later deposited in MZUSP), as
well as the crustacean collections of the Museu de Oceanografia Petronio Alves Coelho, Universidade Federal de
Pernambuco, Recife, Brazil (MOUFPE), and Florida Museum of Natural History, University of Florida,
Gainesville, U.S.A. (FLMNH).
In the present study, two species of Synalpheus are reported for the first time from the southwestern Atlantic
and Brazil (one of them also from USA); one species with a single uncertain Brazilian record is confirmed from
Brazil based on new material; one poorly known species is redescribed; one species is reported as new state record,
based on previously misidentified material; and one species from the Synalpheus paraneptunus Coutière, 1909
species complex (Anker & Tóth 2008) is described as new. Most species reported in this study are abundantly
illustrated. Carapace length (cl, in mm) was measured along the mid-dorsal line from the tip of the rostrum to the
posterior margin of the carapace. The type material of the new species is deposited in the carcinological collection
of MZUSP.
Taxonomy
Alpheidae Rafinesque, 1815
Synalpheus Spence Bate, 1888
Synalpheus androsi Coutière, 1909
(Fig. 1)
Synalpheus androsi Coutière 1909: 82, fig. 50; Chace 1972: 86; Dardeau 1984: 18; Coelho et al. 2006: 52; Macdonald et al.
2006: 169; Macdonald et al. 2009: 7, fig. 2, pl. 1C; Hultgren & Duffy 2011: 357, fig. 2 (in cladogram); Almeida et al.
2012: 28; Anker et al. 2012: 81 (table), 88 (key).
Zuzalpheus androsi—Ríos & Duffy 2007: 9, pl. 1.
Material examined. 1 male (cl 3.4 mm), Brazil, off Espírito Santo, REVIZEE—Comissão Central: 2, R/V Astro
Garoupa, station 21C, 20º42’21’’S 35º27’25’’W, depth: 55 m, 08.XI.1997 [MZUSP 31138]; 1 male (cl 3.0 mm),
same collection data as for previous specimen [MZUSP 31139]; 1 male (cl 3.0 mm, major P1 missing), 1 male (cl
2.6 mm, most pereiopods detached), 1 female (cl 2.6 mm, most pereiopods detached), same collection data as for
previous specimens [MZUSP 31140]; 1 specimen (not sexed), Brazil, Bahia, off Salvador, Saldanha Leste I
Expedition, station AS 1981, 13°40’S 38°48’W, depth: 49 m, sand and gravel, leg. N. Oc. Almirante Saldanha, det.
E. Silva, 23.IX.1968 [MOUFPE 9212] (material examined by J. Fidelis).
Description. See Coutière (1909) for original description and illustrations and Ríos & Duffy (2007, under
Zuzalpheus androsi) for diagnosis; comparative illustrations of the Brazilian material are provided in Fig. 1.
Distribution. Western Atlantic: Bahamas (Andros Island), Belize (Carrie Bow Cay), Jamaica, Barbados
(Coutière 1909; Dardeau 1984; Ríos & Duffy 2007; Macdonald et al. 2009; Hultgren et al. 2011), Brazil (off Bahia
and Espírito Santo: Vitória-Trindade seamounts) (Coelho et al. 2006; present study).
Ecology. In Belize, Synalpheus androsi inhabits large demosponges, Pseudoceratina crassa (Hyatt) and
Hyattella intestinalis (Lamarck), at a depth range of 14–15 m (Ríos & Duffy 2007). The present material was
dredged from much deeper water (49–55 m) and the collection records give no indication on association with
sponges.
Remarks. The present material represents the first confirmed and illustrated record of Synalpheus androsi in
Brazilian waters, closely matching the description and illustrations in Coutière (1909). Almeida et al. (2012)
pointed out that the first record of S. androsi from Brazil by Coelho et al. (2006) required confirmation. This record
was merely a listing of S. androsi in a species table, without a specific locality other than Bahia, and without
illustrations. The corresponding material was deposited in the collections of Museu de Oceanografia Petronio
Alves Coelho in Recife (MOUFPE) under the accession number DOCEAN-9212. At our request, Jesser Fidelis
examined Coelho’s material, a single dry specimen that had to be rehydrated, and confirmed it as S. androsi (see
material examined for collection details).
ANKER & PACHELLE216 · Zootaxa 3815 (2) © 2014 Magnolia Press
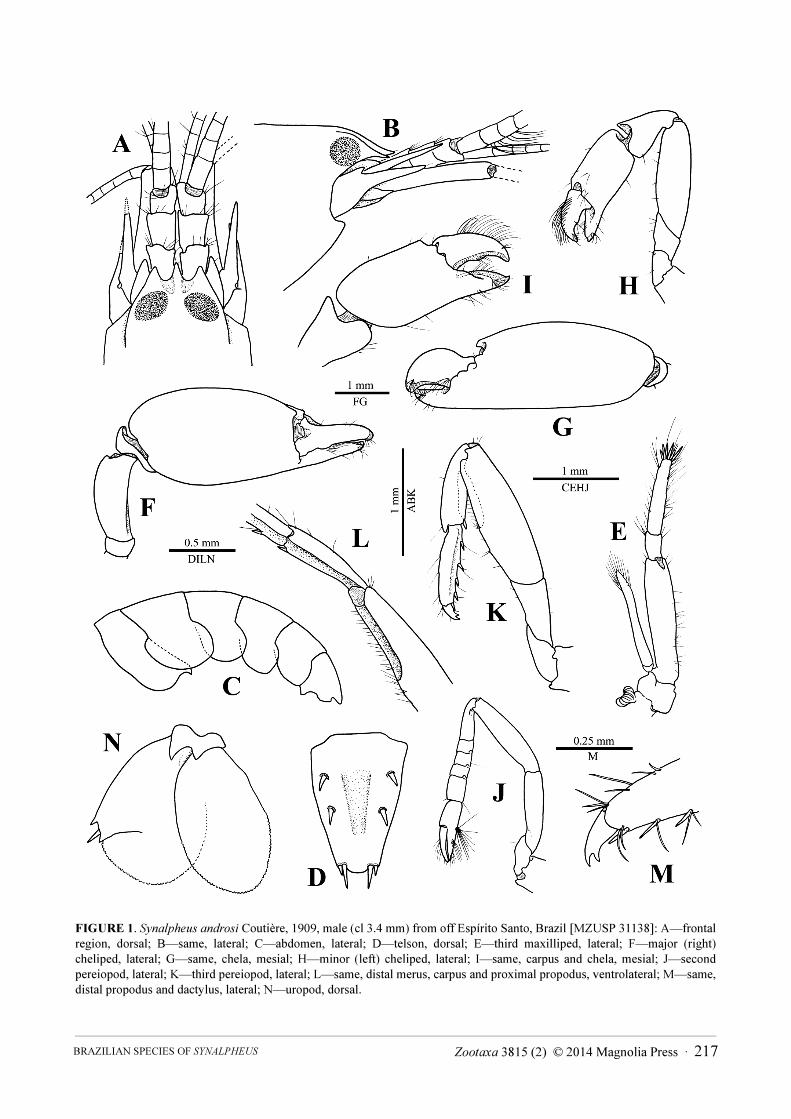
FIGURE 1. Synalpheus androsi Coutière, 1909, male (cl 3.4 mm) from off Espírito Santo, Brazil [MZUSP 31138]: A—frontal
region, dorsal; B—same, lateral; C—abdomen, lateral; D—telson, dorsal; E—third maxilliped, lateral; F—major (right)
cheliped, lateral; G—same, chela, mesial; H—minor (left) cheliped, lateral; I—same, carpus and chela, mesial; J—second
pereiopod, lateral; K—third pereiopod, lateral; L—same, distal merus, carpus and proximal propodus, ventrolateral; M—same,
distal propodus and dactylus, lateral; N—uropod, dorsal.
Zootaxa 3815 (2) © 2014 Magnolia Press · 217BRAZILIAN SPECIES OF SYNALPHEUS

Synalpheus androsi is unique within the Synalpheus gambarelloides (Nardo, 1847) species group in having a
deep excavation on the ventral surface of the third and fourth pereiopods, forming very pronounced lateral flanges.
On the third pereiopod, this excavation extends from the distal half of the merus to the proximal half of the
propodus, gradually flattening towards the dactylus, and is the deepest on the merus and the opposed surface of the
carpus (Fig. 1K, L). Thus the third pereiopods can be folded with the lateral and mesial flanges of the carpus fitting
into and mesial to, respectively, the excavation of the merus (Fig. 1K). This probably helps the shrimp to squat
more effectively and gain additional space in the most narrow sponge canals.
Noteworthy and in direct relationship to its morphological distinctiveness, S. androsi appears to have no close
relatives in the western Atlantic Ocean, forming a moderately supported clade with the eastern Atlantic S.
gambarelloides (Hultgren & Duffy 2011).
Synalpheus maxillispinus sp. nov.
(Figs. 2, 3)
Material examined. Holotype: female (cl 3.0 mm), Brazil, Abrolhos Bank off southern Bahia and northern
Espírito Santo, REVIZEE—Comissão Central: 2, R/V Astro Garoupa, station 17C, 18°34’00”S 38°04’00”W,
depth: 55 m, 22.XI.1997 [MZUSP 30093]. Paratypes: 1 male (cl 2.6 mm), same collection data as for holotype
[MZUSP 30094]; 1 male (cl 3.5 mm), Brazil, Espírito Santo, off Rio Doce, TAAF MD-55/Brasil 1987, station 51/
DC–87, 19°39’S 39°42’W, depth: 15 m, 29.V.1987 [MZUSP 30095].
Description. Rostrum slightly lower than orbital hoods, triangular, distinctly longer than wide at base,
subequal or slightly longer than orbital teeth, subacute distally; orbital teeth triangular, subacute, longer than wide
at base; frontal margin between orbital teeth and rostrum deeply concave, forming with rostrum V- to U-shaped
notches, adjacent rostro-orbital area slightly depressed (Fig. 2A, B, L).
Pleuron of first abdominal somite rounded posteroventrally, without hook; pleura of third to fifth somites also
rounded, fifth slightly more angular (Fig. 2C). Telson moderately broad, about 1.5 times as long as anterior width,
tapering distally; dorsal surface with broad, longitudinal, median depression and two pairs of strong spiniform
setae inserted at about 0.4 and 0.7 length of telson, respectively; posterior margin rounded, less than half-length of
anterior width, with two stout spiniform setae, mesial much longer than lateral (Fig. 2D).
Antennular peduncle with second article 1.3–1.6 times as long as wide; stylocerite falling short of distal margin
of first article, with blunt or subacute tip (Fig. 2A, L). Antennal basicerite with blunt, non-projecting distodorsal
tooth, and strong distolateral tooth reaching at least to middle of second article of antennular peduncle;
scaphocerite with strong broad distolateral tooth reaching beyond end of antennular peduncle; blade always
present, slender, not reaching mid-length of scaphocerite, slightly overreaching distolateral tooth of basicerite,
fringed with short setae (Fig. 2A, B, L).
Mouthparts typical for genus. Third maxilliped with ultimate article bearing one dorsal row of six short
spiniform setae, one ventral row of three longer spiniform setae and crown of seven or so stouter spiniform setae on
apex (Fig. 2E, F).
Major cheliped with palm 2.5–2.8 as long as dactylus; distodorsal tooth on palm not greatly protruding, ending
blunt distally, without acute point; dactylus reaching distinctly beyond pollex (Fig. 3A–E). Minor cheliped with
palm about 1.3 times as long as fingers, latter with cutting surfaces excavated and bidentate tips; dactylus with very
few tufts of setae not organised in rows (Fig. 3F, G).
Second pereiopod with five-articulated carpus, first carpal article about four times length of second, about as
long as sum of second to fifth articles (Fig. 2G). Third pereiopod with merus about three times as long as wide;
propodus with six spiniform setae on ventral margin and one pair of spiniform setae on distoventral margin;
dactylus biunguiculate, moderately stout, secondary unguis subparallel to and shorter than terminal unguis (Fig.
2H, I). Fifth pereiopod with propodus armed with five spiniform setae on ventral margin; grooming brush well
developed (Fig. 2J).
Uropodal exopod with distolateral margin bearing two strong teeth (including lateral tooth of diaeresis) and
one or two spiniform seta(e) (Fig. 2K, M).
Colour in life unknown.
Type locality. Brazil: Abrolhos Bank off southern Bahia / northern Espírito Santo (holotype and first paratype)
and off Rio Doce, Espírito Santo (second paratype).
ANKER & PACHELLE218 · Zootaxa 3815 (2) © 2014 Magnolia Press
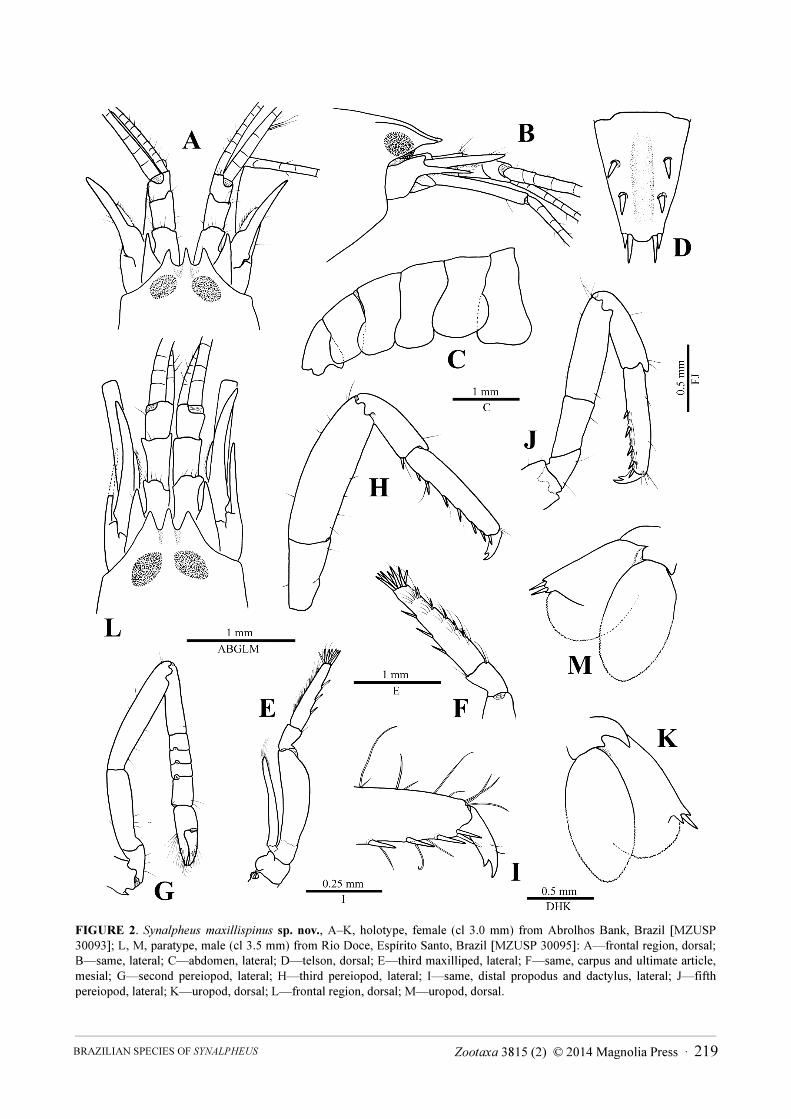
FIGURE 2. Synalpheus maxillispinus sp. nov., A–K, holotype, female (cl 3.0 mm) from Abrolhos Bank, Brazil [MZUSP
30093]; L, M, paratype, male (cl 3.5 mm) from Rio Doce, Espírito Santo, Brazil [MZUSP 30095]: A—frontal region, dorsal;
B—same, lateral; C—abdomen, lateral; D—telson, dorsal; E—third maxilliped, lateral; F—same, carpus and ultimate article,
mesial; G—second pereiopod, lateral; H—third pereiopod, lateral; I—same, distal propodus and dactylus, lateral; J—fifth
pereiopod, lateral; K—uropod, dorsal; L—frontal region, dorsal; M—uropod, dorsal.
Zootaxa 3815 (2) © 2014 Magnolia Press · 219BRAZILIAN SPECIES OF SYNALPHEUS
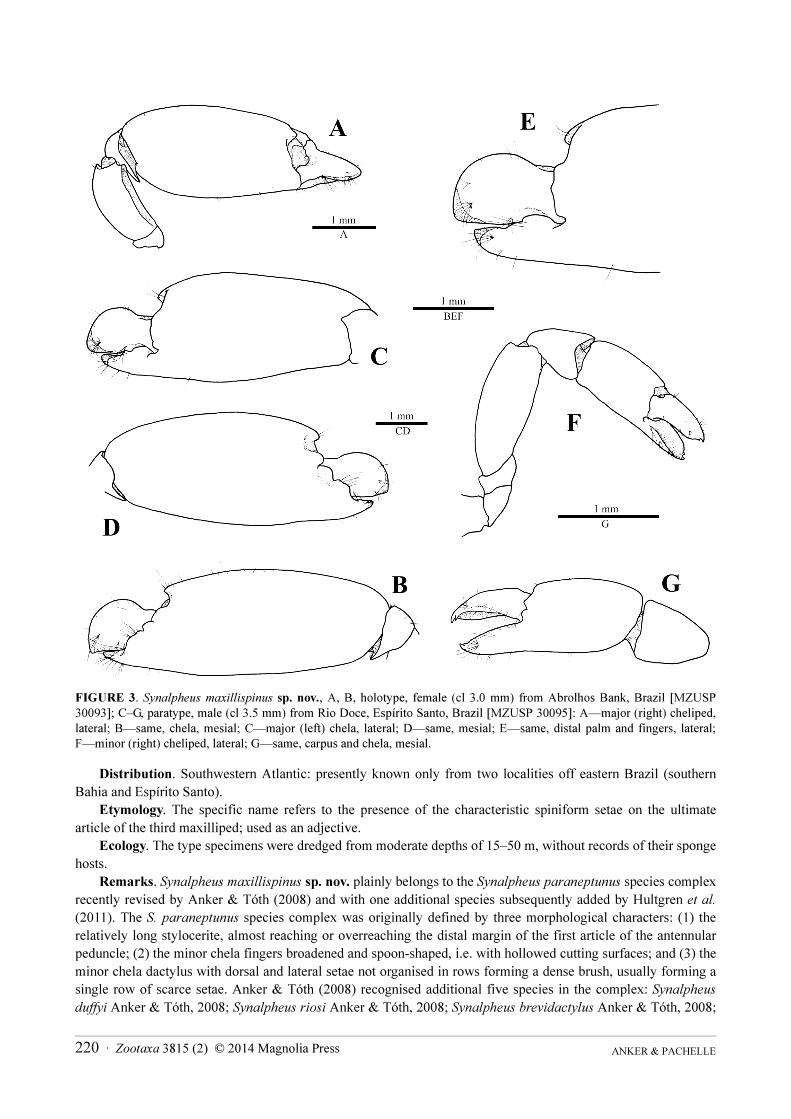
FIGURE 3. Synalpheus maxillispinus sp. nov., A, B, holotype, female (cl 3.0 mm) from Abrolhos Bank, Brazil [MZUSP
30093]; C–G, paratype, male (cl 3.5 mm) from Rio Doce, Espírito Santo, Brazil [MZUSP 30095]: A—major (right) cheliped,
lateral; B—same, chela, mesial; C—major (left) chela, lateral; D—same, mesial; E—same, distal palm and fingers, lateral;
F—minor (right) cheliped, lateral; G—same, carpus and chela, mesial.
Distribution. Southwestern Atlantic: presently known only from two localities off eastern Brazil (southern
Bahia and Espírito Santo).
Etymology. The specific name refers to the presence of the characteristic spiniform setae on the ultimate
article of the third maxilliped; used as an adjective.
Ecology. The type specimens were dredged from moderate depths of 15–50 m, without records of their sponge
hosts.
Remarks. Synalpheus maxillispinus sp. nov. plainly belongs to the Synalpheus paraneptunus species complex
recently revised by Anker & Tóth (2008) and with one additional species subsequently added by Hultgren et al.
(2011). The S. paraneptunus species complex was originally defined by three morphological characters: (1) the
relatively long stylocerite, almost reaching or overreaching the distal margin of the first article of the antennular
peduncle; (2) the minor chela fingers broadened and spoon-shaped, i.e. with hollowed cutting surfaces; and (3) the
minor chela dactylus with dorsal and lateral setae not organised in rows forming a dense brush, usually forming a
single row of scarce setae. Anker & Tóth (2008) recognised additional five species in the complex: Synalpheus
duffyi Anker & Tóth, 2008; Synalpheus riosi Anker & Tóth, 2008; Synalpheus brevidactylus Anker & Tóth, 2008;
ANKER & PACHELLE220 · Zootaxa 3815 (2) © 2014 Magnolia Press

Synalpheus bocas Anker & Tóth, 2008; and Synalpheus belizensis Anker & Tóth, 2008, all from the Caribbean Sea
(Panama, Belize, Dominica). Hultgren et al. (2011) added a seventh species, Synalpheus microneptunus Hultgren,
Macdonald & Duffy, 2011, from Barbados. The S. paraneptunus complex was shown to form a well-supported
clade within the S. gambarelloides group in the combined molecular-morphological analysis of Hultgren & Duffy
(2011). However, and interestingly, this clade also contained a morphologically more distinctive Synalpheus
kensleyi (Ríos & Duffy, 2007), which unlike other species of the S. paraneptunus complex is characterised by a
rather well-developed brush of gambarelloid setae and a strong acute tooth projecting from the dorsal margin of the
antennal basicerite.
Synalpheus maxillispinus sp. nov. is morphologically closest to S. bocas and S. belizensis, sharing with them
the presence of two teeth on the distolateral margin of the uropodal exopod, adjacent to spiniform seta. The new
species can be separated from S. bocas and S. belizensis by the shorter stylocerite, not reaching (vs. overreaching)
distal margin of the first article of the antennular peduncle; the major chela with a short, bluntly ending (vs. more
projecting and sharply pointed) distodorsal tooth; specifically from S. bocas by the scaphocerite having (vs. not
having) a blade1; and the shorter dorsal spiniform setae on the telson; and specifically from S. belizensis by the
more slender third pereiopod, with merus about three (vs. 2.5) times as long as wide (cf. Anker & Tóth 2008).
The presence of two teeth on the distolateral margin of the uropodal exopod separates Synalpheus
maxillispinus sp. nov. from S. paraneptunus, S. duffyi, S. riosi, S. brevidactylus, S. microneptunus and S. kensleyi,
which all have three teeth. In addition, S. maxillispinus sp. nov. can be separated from S. microneptunus by the
presence of five (vs. four) articles in the carpus of the second pereiopod; from S. kensleyi by the minor chela
without setae organised in rows (vs. with distinct gambarelloid setae) and the antennal basicerite without (vs. with)
acutely projecting dorsal tooth; from S. duffyi and S. brevidactylus by the moderately slender, distally blunter (vs.
more slender and acute) orbital teeth and the major chela palm with a bluntly ending (vs. sharply pointed)
distodorsal tooth; from S. riosi by the major chela palm with a dorsally non-swollen, bluntly ending (vs. dorsally
swollen and with a small sharp point) distodorsal tooth and the minor chela without setae grouped in rows (vs. with
setae organised in a single dorsal row) (cf. Ríos & Duffy 2007; Anker & Tóth 2008; Hultgren et al. 2011).
The presence of three spiniform setae on the ventral surface of the ultimate article of the third maxilliped
appears to be another important and possibly diagnostic character of Synalpheus maxillispinus sp. nov. Anker &
Tóth (2008) did not illustrate these setae in S. paraneptunus, S. duffyi, S. riosi and S. belizensis, although some
stout setae in the same position can be seen in their figure 4C of S. duffyi. In addition, Hultgren et al. (2011)
illustrated slender spiniform setae (clearly thicker than adjacent serrulate setae) on the third maxilliped of S.
microneptunus. Therefore, these spiniform setae may also be present, although less developed, in other species of
the S. paraneptunus complex.
All species of the Synalpheus paraneptunus complex are associated with sponges (see Table 1 in Anker et al.
2012 for sponge records) and at least three of them (S. duffyi, S. riosi and S. microneptunus) live in social colonies
(Anker & Tóth 2008; Hultgren et al. 2011; Anker et al. 2012). The sponge hosts of S. paraneptunus, S. riosi and S.
maxillispinus sp. nov. remain unknown; all sponges listed as hosts for S. paraneptunus in Table 1 of Anker et al.
(2012) are in fact sponge records for S. paraneptunus sensu lato and not for S. paraneptunus sensu stricto as
redefined by Anker & Tóth (2008). Whether S. maxillispinus sp. nov. is a social or pair-living species presently
remains unknown.
Synalpheus cf. ruetzleri Macdonald & Duffy, 2006
(Fig. 4)
Synalpheus ruetzleri Macdonald & Duffy 2006: 8, figs. 8–13; Anker et al. 2012: 85 (table), 88 (key).
Zuzalpheus ruetzleri—Ríos & Duffy 2007: 61, pl. 4.
1. �Anker & Tóth (2008, p. 21) obviously made a type-error in the diagnosis of Synalpheus belizensis by stating that the
antennal scaphocerite has no blade. Their illustration of the frontal region of the holotype of S. belizensis (fig. 12A)
clearly shows both scaphocerites having a narrow blade and the presence / absence of a scaphocerite blade is also
used to distinguish S. belizensis from S. bocas under remarks (p. 24) and in the key (p. 27). However, an aberrant
specimen of S. belizensis in Anker & Tóth's fig. 13H has no blade on the left scaphocerite and a reduced blade on the
right scaphocerite.
Zootaxa 3815 (2) © 2014 Magnolia Press · 221BRAZILIAN SPECIES OF SYNALPHEUS
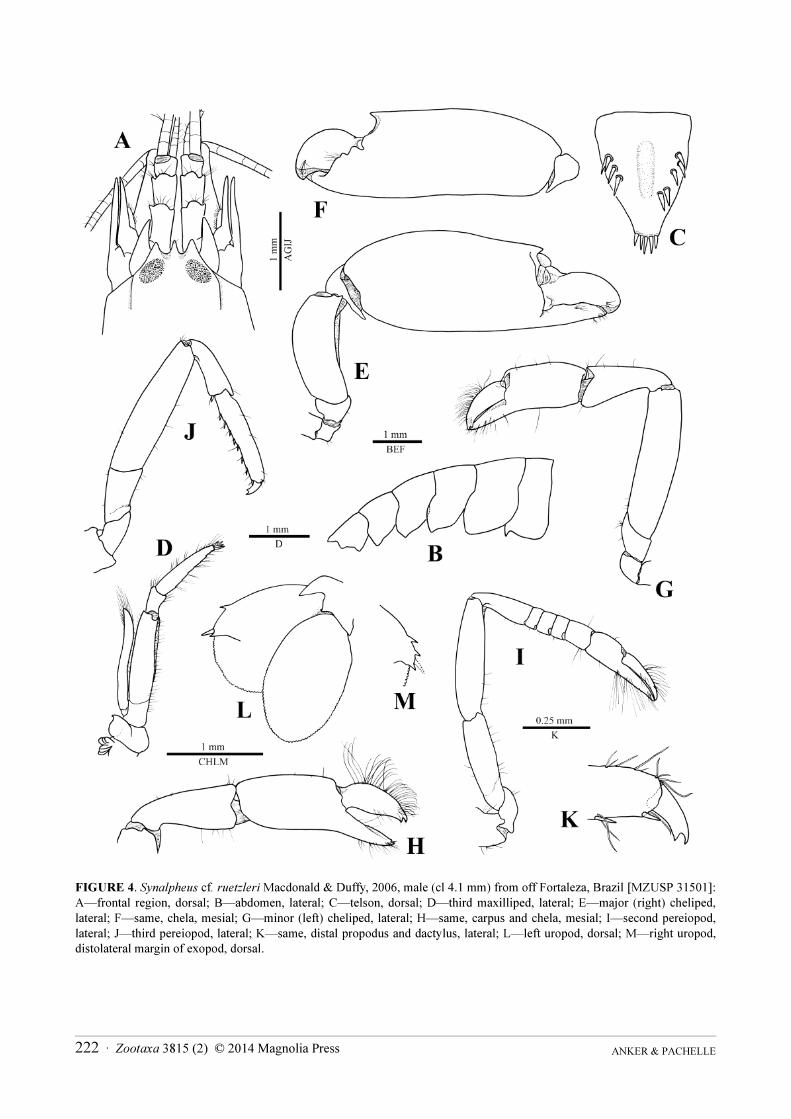
FIGURE 4. Synalpheus cf. ruetzleri Macdonald & Duffy, 2006, male (cl 4.1 mm) from off Fortaleza, Brazil [MZUSP 31501]:
A—frontal region, dorsal; B—abdomen, lateral; C—telson, dorsal; D—third maxilliped, lateral; E—major (right) cheliped,
lateral; F—same, chela, mesial; G—minor (left) cheliped, lateral; H—same, carpus and chela, mesial; I—second pereiopod,
lateral; J—third pereiopod, lateral; K—same, distal propodus and dactylus, lateral; L—left uropod, dorsal; M—right uropod,
distolateral margin of exopod, dorsal.
ANKER & PACHELLE222 · Zootaxa 3815 (2) © 2014 Magnolia Press

Material examined. 1 male (cl 4.1 mm), Brazil, Ceará, off Fortaleza, Parque Estadual Marinho da Pedra da Risca
do Meio, station Dive II, in Agelas dispar, leg. L.E.A. Bezerra, 14.VII.2004 [MZUSP 31501].
Description. See Macdonald & Duffy (2006) for description and illustrations and Ríos & Duffy (2007, under
Zuzalpheus ruetzleri) for diagnosis and colour photograph; comparative illustrations of the Brazilian specimen are
provided in Fig. 4.
Distribution. Western Atlantic: Caribbean Sea (Panama: San Blas Islands, Belize: Carrie Bow Cay); possibly
Brazil (Ceará) (Macdonald & Duffy 2006; Ríos & Duffy 2007; present study).
Ecology. In Belize, Synalpheus ruetzleri was found exclusively in the demosponge Hymeniacidon cf. caerulea
Pulitzer-Finali, in very shallow water, 1–2 m (Ríos & Duffy 2007). In contrast, the Ceará material was found inside
a different sponge, Agelas dispar Duchassaing & Michelotti, collected by scuba diving in less than 20 m, whereas
in Panama, S. ruetzleri appears to associate with various sponges, including H. cf. caerulea (Macdonald & Duffy
2006).
Remarks. The present material, if confirmed to be conspecific with the Caribbean Synalpheus ruetzleri, may
represent the first record of this species in the southwestern Atlantic and Brazilian waters. Like most other species
of the taxonomically difficult Synalpheus brooksi Coutière, 1909 species complex, S. ruetzleri appears to be
morphologically highly variable, as exemplified by differences in the shape of the orbital teeth (Macdonald &
Duffy 2006: figs. 8A, 13A) and distodorsal protuberance of the major chela (idem., figs. 10A, D, G, 13C). The
presence of a small scaphocerite blade separates this species from the otherwise almost identical Synalpheus
bousfieldi Chace, 1972 and Synalpheus idios (Ríos & Duffy, 2007). It also differs from the supposedly variable S.
brooksi, for instance, by the shape and direction of the distodorsal protuberance of the major chela (more pointed
and directed somewhat upward in S. brooksi) in addition to the absence of a scaphocerite blade (Macdonald &
Duffy 2006).
The single male specimen from Ceará matches only partly the description and illustrations provided by
Macdonald & Duffy (2006). The shape of the frontal margin of the carapace is rather similar as are most features of
the antennular and antennal peduncles (cf. Fig. 4A and Macdonald & Duffy 2006: fig. 8A, D). The telson of the
Ceará male is rather different from those of the male holotype and female allotype from Belize, especially in the
narrower posterior half (cf. Fig. 4C and Macdonald & Duffy 2006: fig. 8B, G). However, in the number (4 + 5) and
position of dorsal spiniform setae, the telson of the Ceará specimen is clearly aberrant (Fig. 4C). Another important
difference with the type material is the shape of the distodorsal protuberance of the major chela, which is relatively
short and straight in the Brazilian specimen compared to the dorsally slightly swollen and ending in a small,
downward directed point in both the holotype and allotype (cf. Fig. 4E, F and Macdonald & Duffy 2006: fig. 10A,
D, F, G). However, in this last feature, the Ceará specimen approaches the non-type male of S. ruetzleri from
Panama (cf. Fig. 4E, F and Macdonald & Duffy 2006: fig. 13C). Noteworthy, the armature of the distolateral
margin of the uropodal exopod is variable (one or two teeth) in the Ceará specimen (Fig. 4L, M).
The paucity of the Brazilian material (presently one single male specimen) does not allow to evaluate the
importance of these morphological differences. Sequencing of the barcoding gene segment (COI) of the Ceará
specimen is highly desirable to test the correctness of the present identification.
Synalpheus tenuispina Coutière, 1909
(Figs. 5, 6)
Synalpheus latastei tenuispina Coutière 1909: 26, fig. 8; Chace 1972: 93; Coelho & Ramos 1972: 150; Christoffersen 1980:
171; Christoffersen 1998: 362.
(?) Synalpheus latastei tenuispina—Abele 1976: 273; Martínez-Iglesias et al. 1993: 12; Martínez-Iglesias et al. 1996: 35.
Synalpheus tenuispina—Crosnier & Forest 1965: 364; Ríos & Duffy 2007: 78 (key); Anker et al. 2012: 9, 86 (table), 90 (key).
Material examined. 1 male (cl 7.1 mm), Brazil, São Paulo, São Sebastião, Pier da Petrobras, 12.XII.2009
[MZUSP 31503]; 1 male (cl 6.9 mm, major P1 missing), 1 heavily damaged specimen (only major P1, P2-P5 and
sternal portion of cephalothorax), Brazil, Rio de Janeiro, Niteroi, Baia de Guanabara, Praia de Boa Viagem, scuba
diving, fauna associated with Polychaeta, coll. J.B. Mendonça Jr., 04.X.2006 [MZUSP 22133].
Description. Rostrum with base clearly delimited from anteromesial margin of orbital hoods, much longer
than wide at base, arrow-shaped in dorsal view, longer than orbital teeth, acute distally, tip overreaching mid-length
Zootaxa 3815 (2) © 2014 Magnolia Press · 223BRAZILIAN SPECIES OF SYNALPHEUS

of first article of antennular peduncle; orbital teeth large, triangular, acute distally, about as long as wide at base;
anteromesial margin between orbital teeth and rostrum abruptly sloping ventrally, forming deep V-shaped notch in
dorsal view; rostro-orbital process well developed (Fig. 5A, B).
Pleuron of first abdominal somite pointed posteroventrally, forming feeble hook; pleura of third to fifth
somites also rounded, fifth slightly more angular (Fig. 5C). Telson broad, about 1.3 times as long as anterior width,
slightly tapering distally; dorsal surface with shallow, longitudinal, median depression and two pairs of stout
spiniform setae inserted at about 0.45 and 0.75 length of telson, respectively; posterior margin broadly rounded,
about 0.6 length of anterior width, with two stout spiniform setae, mesial much longer than lateral (Fig. 5D).
Antennular peduncle with second article 1.5 times as long as wide; stylocerite very strong, reaching far beyond
distal margin of first article, with acute tip (Fig. 5A). Antennal basicerite with acutely projecting distodorsal tooth
and much stronger distolateral tooth, latter reaching well into first third of second article of antennular peduncle
and almost equalling tip of stylocerite; scaphocerite with very strong distolateral tooth reaching beyond end of
antennular peduncle; blade well-developed, moderately broad, reaching to about 0.7 length of scaphocerite, fringed
with long setae; carpocerite reaching slightly beyond end of antennular peduncle but not beyond distolateral tooth
of scaphocerite (Fig. 5A, B).
Mouthparts typical for genus. Third maxilliped with very short penultimate article; ultimate article with crown
of about six stout spiniform setae on apex (Fig. 5E, F).
Major cheliped with merus about 2.3 times as long as wide; palm ovoid, 2.3–2.4 as long as dactylus;
distodorsal margin of palm slightly protruding, forming low blunt protuberance, without acute tooth; dactylus
subequal to pollex (Fig. 6A, B). Minor cheliped with merus about 2.3 times as long as greatest width, with convex
dorsal and ventral margins, distodorsal margin projecting as sharp tooth; carpus cup-shaped; palm about 1.2 times
as long as fingers; dactylus and pollex with sharp cutting edges, laterodorsal surface of pollex slightly depressed
(Fig. 6C, D).
Second pereiopod with five-articulated carpus, first carpal article about four times length of second, equal to
sum of second to fifth articles (Fig. 5G). Third pereiopod with merus almost four times as long as wide; carpus with
stout spiniform seta distoventrally; propodus with eight spiniform setae on ventral margin and one pair of
spiniform setae on distoventral margin; dactylus biunguiculate, moderately stout, secondary unguis subparallel to
and shorter than terminal unguis (Fig. 5H, I). Fifth pereiopod with propodus armed with four spiniform setae on
ventral margin and one pair of spiniform setae on distoventral margin, and with well-developed grooming brush
laterally, consisting of at least six rows of serrulate setae (Fig. 5J, K).
Uropod with protopod ending in large subacute tooth laterally; distolateral margin of exopod with two strong
teeth (including lateral tooth of diaeresis) and one stout spiniform seta (Fig. 5L).
Colour in life unknown.
Distribution. Southwestern Atlantic: Brazil (Rio de Janeiro, São Paulo, Santa Catarina). Records of S.
tenuispina from the Caribbean Sea (Abele 1976; Martínez-Iglesias et al. 1993, 1996) most likely refer to a species
currently known as Synalpheus cf. africanus Crosnier & Forest, 1965, and not to S. tenuispina (see below).
Ecology. Synalpheus tenuispina appears to be a non-symbiotic species, dwelling in pier growth or in crevices
of polychaete reefs.
Remarks. Coutière's (1909) original description of Synalpheus latastei tenuispina was based on a single large
female (tl 30 mm) from Desterro, now Florianópolis, Santa Catarina, Brazil. The description is actually a short
paragraph with comparisons of proportions and lengths between the new form and the supposedly closely related,
eastern Pacific Synalpheus latastei Coutière, 1909 (now junior synonym of Synalpheus spinifrons (H. Milne
Edwards, 1837)) and Synalpheus lockingtoni Coutière, 1909, accompanied by a few figures. The holotype of S.
latastei tenuispina was deposited in the Muséum National d'Histoire Naturelle in Paris, France (MNHN), where,
however, it was missing already in the 1960s (Crosnier & Forest 1965) and was also not found during a more recent
search in 2007 (A. Anker, pers. obs.). Based on the morphological differences between S. latastei tenuispina and S.
spinifrons in Coutière's (1909) account, Crosnier & Forest (1965) elevated the Brazilian form to full species, as S.
tenuispina.
The most complete specimen of the present material is an adult male (cl 7.1 mm, tl ~25 mm) from São
Sebastião (Figs. 5, 6). This male agrees well with figures of the missing female holotype in Coutière (1909), except
for the slightly broader rostrum and stouter merus of the third pereiopod (cf. Fig. 5A, H and Coutière 1909: fig. 8a,
m). Obviously more fresh material is needed to understand the range of morphological variation of S. tenuispina
and also to document the colour pattern of the species.
ANKER & PACHELLE224 · Zootaxa 3815 (2) © 2014 Magnolia Press
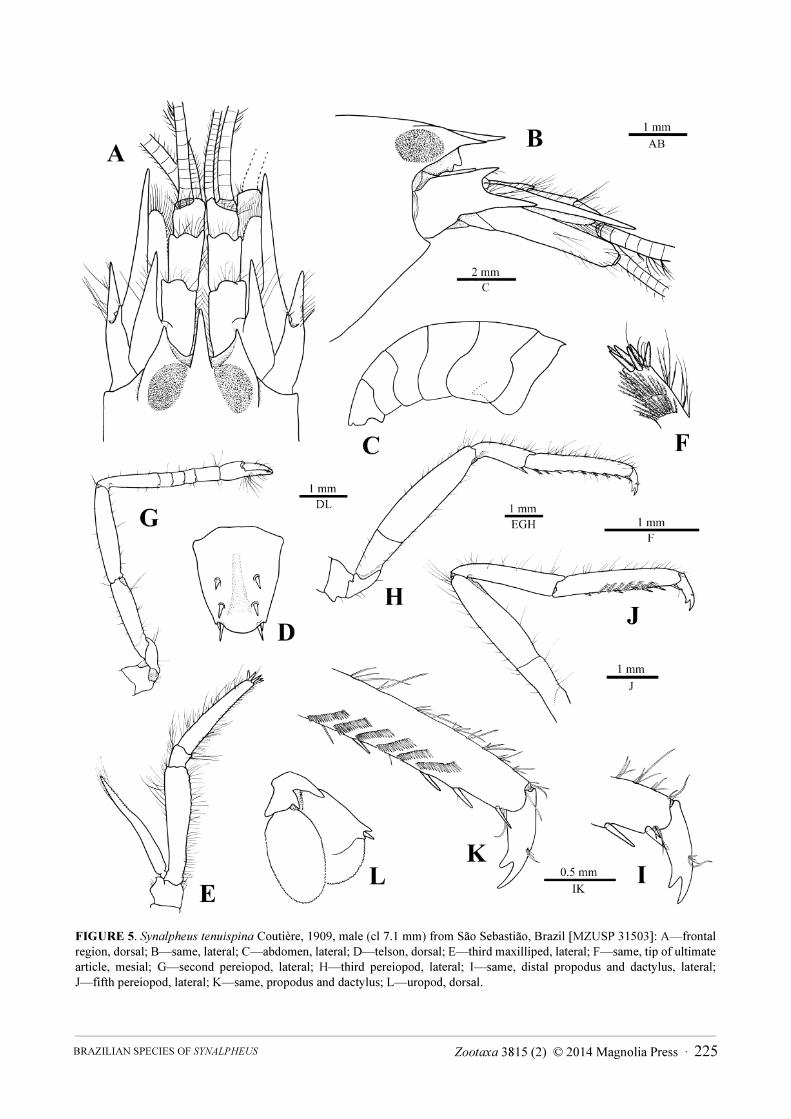
FIGURE 5. Synalpheus tenuispina Coutière, 1909, male (cl 7.1 mm) from São Sebastião, Brazil [MZUSP 31503]: A—frontal
region, dorsal; B—same, lateral; C—abdomen, lateral; D—telson, dorsal; E—third maxilliped, lateral; F—same, tip of ultimate
article, mesial; G—second pereiopod, lateral; H—third pereiopod, lateral; I—same, distal propodus and dactylus, lateral;
J—fifth pereiopod, lateral; K—same, propodus and dactylus; L—uropod, dorsal.
Zootaxa 3815 (2) © 2014 Magnolia Press · 225BRAZILIAN SPECIES OF SYNALPHEUS
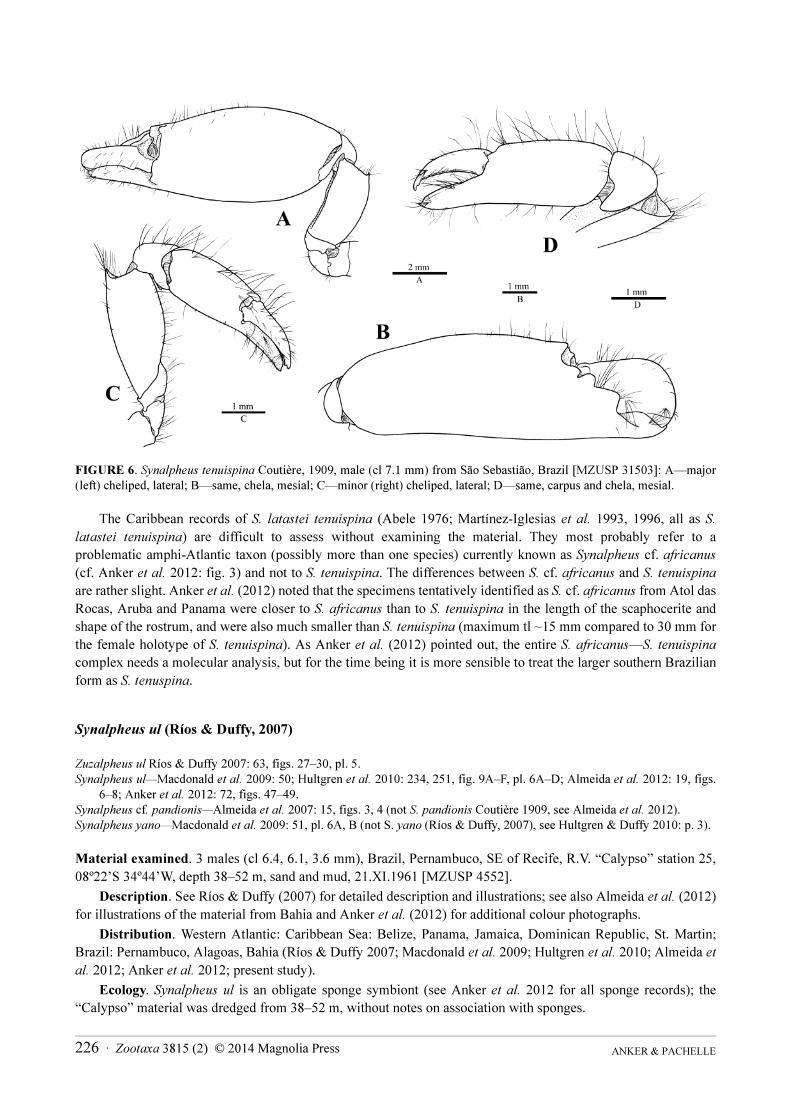
FIGURE 6. Synalpheus tenuispina Coutière, 1909, male (cl 7.1 mm) from São Sebastião, Brazil [MZUSP 31503]: A—major
(left) cheliped, lateral; B—same, chela, mesial; C—minor (right) cheliped, lateral; D—same, carpus and chela, mesial.
The Caribbean records of S. latastei tenuispina (Abele 1976; Martínez-Iglesias et al. 1993, 1996, all as S.
latastei tenuispina) are difficult to assess without examining the material. They most probably refer to a
problematic amphi-Atlantic taxon (possibly more than one species) currently known as Synalpheus cf. africanus
(cf. Anker et al. 2012: fig. 3) and not to S. tenuispina. The differences between S. cf. africanus and S. tenuispina
are rather slight. Anker et al. (2012) noted that the specimens tentatively identified as S. cf. africanus from Atol das
Rocas, Aruba and Panama were closer to S. africanus than to S. tenuispina in the length of the scaphocerite and
shape of the rostrum, and were also much smaller than S. tenuispina (maximum tl ~15 mm compared to 30 mm for
the female holotype of S. tenuispina). As Anker et al. (2012) pointed out, the entire S. africanus—S. tenuispina
complex needs a molecular analysis, but for the time being it is more sensible to treat the larger southern Brazilian
form as S. tenuspina.
Synalpheus ul (Ríos & Duffy, 2007)
Zuzalpheus ul Ríos & Duffy 2007: 63, figs. 27–30, pl. 5.
Synalpheus ul—Macdonald et al. 2009: 50; Hultgren et al. 2010: 234, 251, fig. 9A–F, pl. 6A–D; Almeida et al. 2012: 19, figs.
6–8; Anker et al. 2012: 72, figs. 47–49.
Synalpheus cf. pandionis—Almeida et al. 2007: 15, figs. 3, 4 (not S. pandionis Coutière 1909, see Almeida et al. 2012).
Synalpheus yano—Macdonald et al. 2009: 51, pl. 6A, B (not S. yano (Ríos & Duffy, 2007), see Hultgren & Duffy 2010: p. 3).
Material examined. 3 males (cl 6.4, 6.1, 3.6 mm), Brazil, Pernambuco, SE of Recife, R.V. “Calypso” station 25,
08º22’S 34º44’W, depth 38–52 m, sand and mud, 21.XI.1961 [MZUSP 4552].
Description. See Ríos & Duffy (2007) for detailed description and illustrations; see also Almeida et al. (2012)
for illustrations of the material from Bahia and Anker et al. (2012) for additional colour photographs.
Distribution. Western Atlantic: Caribbean Sea: Belize, Panama, Jamaica, Dominican Republic, St. Martin;
Brazil: Pernambuco, Alagoas, Bahia (Ríos & Duffy 2007; Macdonald et al. 2009; Hultgren et al. 2010; Almeida et
al. 2012; Anker et al. 2012; present study).
Ecology. Synalpheus ul is an obligate sponge symbiont (see Anker et al. 2012 for all sponge records); the
“Calypso” material was dredged from 38–52 m, without notes on association with sponges.
ANKER & PACHELLE226 · Zootaxa 3815 (2) © 2014 Magnolia Press

Remarks. The present specimens from Pernambuco, collected by the French research vessel “Calypso” in
1961, were misidentified as Synalpheus longicarpus (Herrick, 1891) and included by Christoffersen (1979) as part
of his extensive material of S. longicarpus. It is also likely that several other records of S. longicarpus in
Christoffersen (1979) and perhaps in other studies are based on misidentified material. Therefore, the entire
Brazilian material of S. longicarpus, including all the remaining material in Christoffersen (1979), must be re-
examined. Synalpheus ul was previously reported from Brazil, from Camamu Bay, Bahia (Almeida et al. 2007,
2012) and Maceió, Alagoas (Anker et al. 2012); this is the first record of this species from Pernambuco.
Synalpheus yano (Ríos & Duffy, 2007)
(Figs. 7, 8)
Zuzalpheus yano Ríos & Duffy 2007: 69, figs 31–34, pl. 5.
Synalpheus yano—Hernáez et al. 2010: 692, fig. 2; Hultgren et al. 2010: 234; Anker et al. 2012: 76, fig. 50.
Not Synalpheus yano—Macdonald et al. 2009: 51, pl. 6A, B (= S. ul, see Hultgren & Duffy 2010: p. 3).
Material examined. 2 ov. females (cl 4.3 mm, 3.5 mm), Brazil, Ceará, ~15 km N-NE of Fortaleza, scuba diving, in
coral rubble, coll. C. Cerqueira & M. Soares, 21.IV.2013 [MZUSP 31142]; 1 male (cl 3.9 mm), same collection
data as for previous specimens [MZUSP 31141]; 1 ov. female (cl ~4.5 mm), USA, Florida, N-NW of St.
Petersburg, south of Big Bend area, station FI02012-01, 28.55667 N -84.2745 W, dredge, hard bottom, rocks and
coral rubble, depth: 26–30 m, coll. J. Slapcinsky, 24.V.2012 [FLMNH UF 31631].
Description. See Ríos & Duffy (2007) for description and illustrations and Anker et al. (2012) for colour
photographs; comparative illustrations of the Brazilian material and an additional colour photograph of a specimen
from western Florida are provided in Figs. 7, 8.
Distribution. Western Atlantic: Gulf of Mexico (Mexico: Alacranes Reef, USA: western Florida); Caribbean
Sea (Belize, Panama, Jamaica); Brazil (Ceará) (Ríos & Duffy 2007; Anker et al. 2012; present study).
Ecology. Synalpheus yano is an obligate sponge symbiont (see Anker et al. 2012 for all sponge records); all
specimens from Ceará and the single specimen from Florida were extracted from coral rubble that likely contained
cryptic host sponges.
Remarks. The present material represents the first record of Synalpheus yano in Brazil and the southwestern
Atlantic (MZUSP material), as well as USA and the eastern Gulf of Mexico (FLMNH UF material). This species is
closely related to Synalpheus pandionis Coutière, 1909 and Synalpheus ul, which both also occur in Brazil
(Almeida et al. 2012; Anker et al. 2012; see also above), but can be separated from them by the absence of a
scaphocerite blade, and from S. pandionis also by the absence of a rounded protuberance on the mesial face of the
major chela.
Synalpheus sp. gambarelloides group
(Fig. 9)
Material examined. 1 ov. female (cl 5.0 mm, missing posterior part of abdomen and tail fan), Brazil, Ceará, off
Fortaleza, Parque Estadual Marinho da Pedra da Risca do Meio, station Dive II, in Agelas dispar, leg. L.E.A.
Bezerra, 14.VII.2004 [MZUSP 31502].
Remarks. Unfortunately, this adult female from Ceará is damaged, missing the two posterior somites of the
abdomen, telson and uropods. The short stylocerite (Fig. 9A) and the minor chela with well-developed
gambarelloid setae (Fig. 9E) indicate clearly that this specimen belongs to the Synalpheus gambarelloides species
group. However, the combination of the frontal region of the carapace with a short rostral carina extending
posteriorly to about mid-length of corneas and flanked by shallow rostro-orbital depressions (Fig. 9A); the antennal
scaphocerite without a blade and equalling the distolateral tooth of the basicerite (Fig. 9A); the major chela with
distodorsal protuberance poorly developed and bluntly rounded (Fig. 9C, D); and the five-articulated carpus of the
second pereiopod (Fig. 9F) do not allow to assign it unambiguously to any of the species reported so far from
Brazil or the western Atlantic. With a single damaged specimen on hand it is impossible to say whether it belongs
to an undescribed or a morphologically variable species.
Zootaxa 3815 (2) © 2014 Magnolia Press · 227BRAZILIAN SPECIES OF SYNALPHEUS
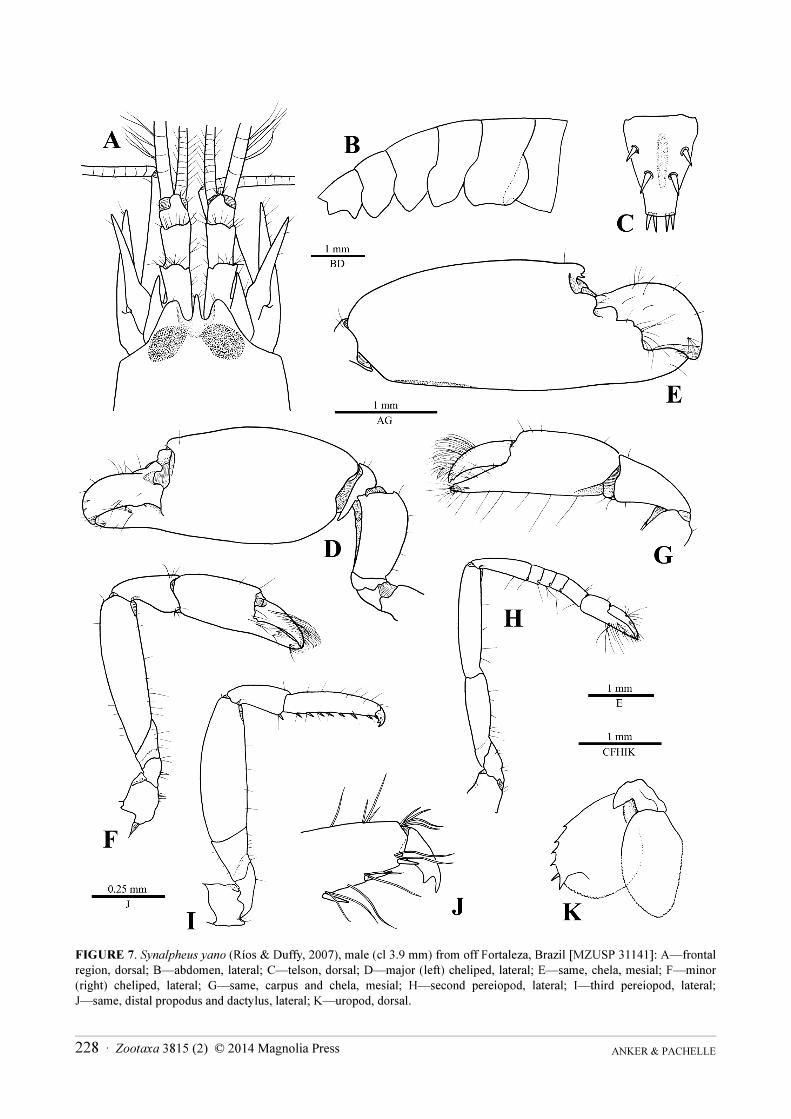
FIGURE 7. Synalpheus yano (Ríos & Duffy, 2007), male (cl 3.9 mm) from off Fortaleza, Brazil [MZUSP 31141]: A—frontal
region, dorsal; B—abdomen, lateral; C—telson, dorsal; D—major (left) cheliped, lateral; E—same, chela, mesial; F—minor
(right) cheliped, lateral; G—same, carpus and chela, mesial; H—second pereiopod, lateral; I—third pereiopod, lateral;
J—same, distal propodus and dactylus, lateral; K—uropod, dorsal.
ANKER & PACHELLE228 · Zootaxa 3815 (2) © 2014 Magnolia Press

FIGURE 8. Synalpheus yano (Ríos & Duffy, 2007): A—ovigerous female (cl 4.3 mm) from Ceará, Brazil [MZUSP 31142],
lateral view; B—ovigerous female (cl ~4.5 mm) from St. Petersburg area, western Florida, USA [FLMNH UF 31631]:
dorsolateral view. Photographs courtesy of F. Monteiro (A) and J. Slapcinsky (B).
Zootaxa 3815 (2) © 2014 Magnolia Press · 229BRAZILIAN SPECIES OF SYNALPHEUS
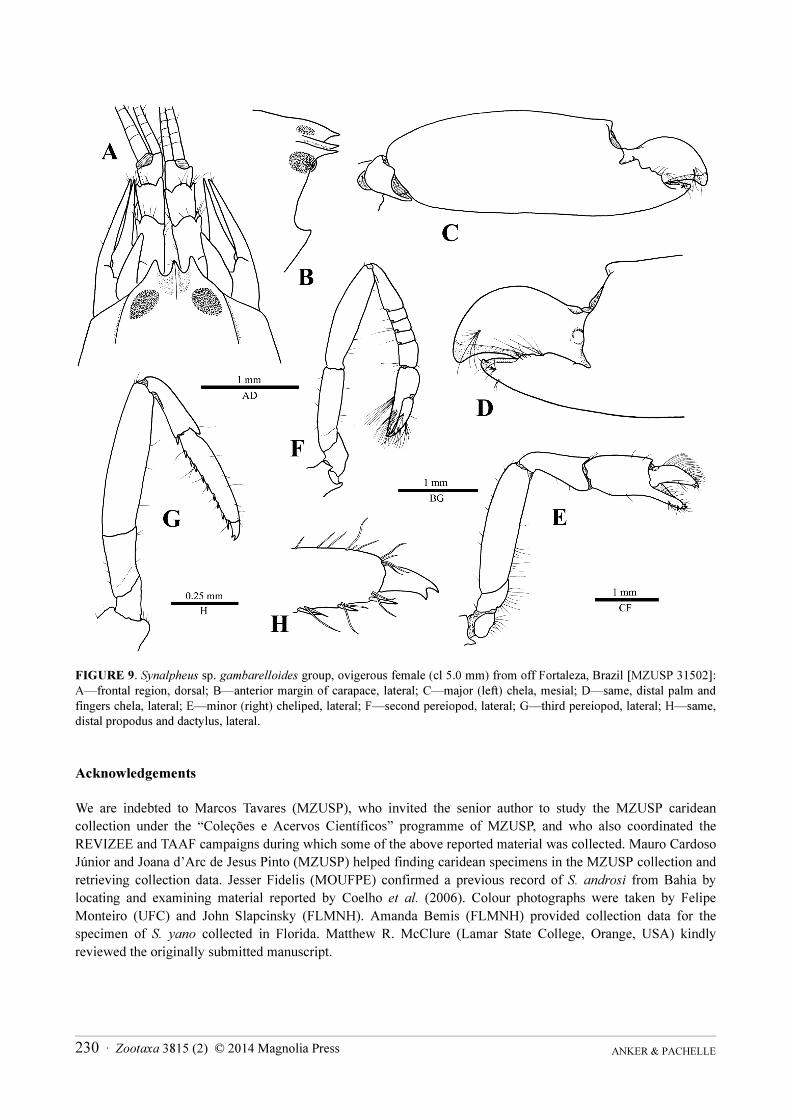
FIGURE 9. Synalpheus sp. gambarelloides group, ovigerous female (cl 5.0 mm) from off Fortaleza, Brazil [MZUSP 31502]:
A—frontal region, dorsal; B—anterior margin of carapace, lateral; C—major (left) chela, mesial; D—same, distal palm and
fingers chela, lateral; E—minor (right) cheliped, lateral; F—second pereiopod, lateral; G—third pereiopod, lateral; H—same,
distal propodus and dactylus, lateral.
Acknowledgements
We are indebted to Marcos Tavares (MZUSP), who invited the senior author to study the MZUSP caridean
collection under the “Coleções e Acervos Científicos” programme of MZUSP, and who also coordinated the
REVIZEE and TAAF campaigns during which some of the above reported material was collected. Mauro Cardoso
Júnior and Joana d’Arc de Jesus Pinto (MZUSP) helped finding caridean specimens in the MZUSP collection and
retrieving collection data. Jesser Fidelis (MOUFPE) confirmed a previous record of S. androsi from Bahia by
locating and examining material reported by Coelho et al. (2006). Colour photographs were taken by Felipe
Monteiro (UFC) and John Slapcinsky (FLMNH). Amanda Bemis (FLMNH) provided collection data for the
specimen of S. yano collected in Florida. Matthew R. McClure (Lamar State College, Orange, USA) kindly
reviewed the originally submitted manuscript.
ANKER & PACHELLE230 · Zootaxa 3815 (2) © 2014 Magnolia Press

Literature
Abele, L.G. (1976) Comparative species composition and relative abundance of decapod crustaceans in marine habitats of
Panamá. Marine Biology, 38, 263–278.
http://dx.doi.org/10.1007/bf00388939
Almeida, A.O., Boehs, G., Araújo-Silva, C.L. & Bezerra, L.E.A. (2012) Shallow-water caridean shrimps from southern Bahia,
Brazil, including the first record of Synalpheus ul (Ríos & Duffy, 2007) (Alpheidae) in the southwestern Atlantic Ocean.
Zootaxa, 3347, 1–35.
Almeida, A.O., Guerrazzi, M.C. & Coelho, P.A. (2007) Stomatopod and decapod crustaceans from Camamu Bay, state of
Bahia, Brazil. Zootaxa, 1553, 1–45.
Anker, A., Pachelle, P.P.G., De Grave, S. & Hultgren, K.M. (2012) Taxonomic and biological notes on some Atlantic species of
the snapping shrimp genus Synalpheus Spence Bate, 1888 (Decapoda, Alpheidae). Zootaxa, 3598, 1–96.
Anker, A. & Tóth, E. (2008) A preliminary revision of the Synalpheus paraneptunus Coutière, 1909 species complex
(Crustacea: Decapoda: Alpheidae). Zootaxa, 1915, 1–28.
Bezerra, L.E.A. & Coelho, P.A. (2006) Crustáceos decápodos associados a esponjas no litoral do Estado do Ceará, Brasil.
Revista Brasileira de Zoologia, 23, 699–702.
http://dx.doi.org/10.1590/s0101-81752006000300012
Chace, F.A. Jr. (1972) The shrimps of the Smithsonian-Bredin Caribbean expeditions with a summary of West Indian shallow-
water species (Crustacea: Decapoda: Natantia). Smithsonian Contributions to Zoology, 98, 1–179.
http://dx.doi.org/10.5479/si.00810282.98
Christoffersen, M.L. (1979) Campagne de la Calypso au large des côtes Atlantiques de l’Amérique du Sud (1961-1962). I
Decapod Crustacea: Alpheoida. Annales de l’Institut Océanographique, 55, 297–377.
Christoffersen, M.L. (1980) Taxonomia e distribuição geográfica dos Alpheoidea (Crustacea, Decapoda, Natantia) do Brasil,
Uruguai e norte da Argentina, incluindo considerações sobre a divisão do sul do continente em províncias biogeográficas
marinhas. Ph.D. Thesis, University of São Paulo, 467 pp.
Christoffersen, M.L (1998) Malacostraca. Eucarida. Caridea, Crangonoidea and Alpheoidea (except Glyphocrangonidae and
Crangonidae). In: Young, P.S. (Ed.), Catalogue of Crustacea of Brazil. Museu Nacional, Rio de Janeiro. pp. 351–372.
Coelho, P.A., Almeida, A.O. de, Souza-Filho, J.F. de, Arruda Bezerra, L.E., & Giraldes, B.W. (2006) Diversity and distribution
of the marine and estuarine shrimps (Dendrobranchiata, Stenopodidea and Caridea) from North and Northeast Brazil.
Zootaxa, 1221, 41–62.
Coelho, P.A. & Ramos, M.A. (1972) A constituição e a distribuição da fauna de decápodos do litoral leste da América do Sul
entre as latitudes de 5°N e 39°S. Trabalhos Oceanograficos da Universidade Federal de Pernambuco, Recife, 13,
133–236.
Coelho Filho, P.A. (2006) Checklist of the decapods (Crustacea) from the outer continental shelf and seamounts from Northeast
of Brazil—REVIZEE Program (NE III). Zootaxa, 1184, 1–27.
Coutière, H. (1909) The American species of snapping shrimps of the genus Synalpheus. Proceedings of the United States
National Museum, 36, 1–93.
http://dx.doi.org/10.5479/si.00963801.36-1659.1
Crosnier, A. & Forest, J. (1965) Remarques sur quelques espèces ouest-africaines d’Alpheidae (Decapoda Macrura).
Description d’Alpheus blachei sp. nov. Bulletin du Muséum National d’Histoire Naturelle, Paris, 36, 355–367.
Dardeau, M.R. (1984) Synalpheus shrimps (Crustacea: Decapoda: Alpheidae). I. The Gambarelloides group, with a description
of a new species. Memoirs of the Hourglass Cruises, 7, 1–125.
De Grave, S. & Fransen, C.H.J.M. (2011) Carideorum catalogus: the recent species of the dendrobranchiate, stenopodidean,
procarididean and caridean shrimps (Crustacea: Decapoda). Zoologische Mededelingen, 89, 195–589.
Hernáez, P., Martínez-Guerrero, B., Anker, A. & Wehrtmann, I.S. (2010) Fecundity and effects of bopyrid infestation on egg
production in the Caribbean sponge-dwelling snapping shrimp Synalpheus yano (Decapoda: Alpheidae). Journal of the
Marine Biological Association of the United Kingdom, 90, 691–698.
http://dx.doi.org/10.1017/s0025315409991093
Herrick, F.H. (1891) Alpheus: a study in the development of Crustacea. In: Brooks, W.K. & Herrick, F.H. (Eds.), The
embryology and metamorphosis of the Macrura. Memories of the National Academy of Sciences, 5, pp. 370–463.
Hultgren, K.M. & Duffy, J.E. (2010) Sponge host characteristics shape the community structure of their shrimp associates.
Marine Ecology Progress Series, 407, 1–12.
http://dx.doi.org/10.3354/meps08609
Hultgren, K.M. & Duffy, J.E. (2011) Multi-locus phylogeny of sponge-dwelling snapping shrimp (Caridea: Alpheidae:
Synalpheus) supports morphology-based species concepts. Journal of Crustacean Biology, 31, 352–360.
http://dx.doi.org/10.1651/10-3382.1
Hultgren, K.M. & Duffy, J.E. (2012) Phylogenetic community ecology and the role of social dominance in sponge-dwelling
shrimp. Ecology Letters, 15, 704–713.
http://dx.doi.org/10.1111/j.1461-0248.2012.01788.x
Hultgren, K.M., Macdonald, K.S. III & Duffy, J.E. (2010) Sponge-dwelling snapping shrimps of Curaçao, with descriptions of
three new species. Zootaxa, 2372, 221–262.
Zootaxa 3815 (2) © 2014 Magnolia Press · 231BRAZILIAN SPECIES OF SYNALPHEUS

Hultgren, K.M., Macdonald, K.S. III & Duffy, J.E. (2011) Sponge-dwelling snapping shrimps (Alpheidae: Synalpheus) of
Barbados, West Indies, with a description of a new eusocial species. Zootaxa, 2834, 1–16.
MacDonald, K.S. III & Duffy, J.E. (2006) Two new species of sponge-dwelling snapping shrimp from the Belizean Barrier
Reef, with a synopsis of the Synalpheus brooksi species complex. American Museum Novitates, 3543, 1–22.
http://dx.doi.org/10.1206/0003-0082(2006)3543[1:tnsoss]2.0.co;2
MacDonald, K.S. III, Hultgren, K.M. & Duffy, J.E. (2009) The sponge-dwelling snapping shrimps of Discovery Bay, Jamaica,
with descriptions of four new species. Zootaxa, 2199, 1–57.
MacDonald, K.S. III, Rios, R. & Duffy, J.E. (2006) Biodiversity, host specificity, and dominance by eusocial species among
sponge-dwelling alpheid shrimp on the Belize Barrier Reef. Diversity and Distributions, 12, 165–178.
http://dx.doi.org/10.1111/j.1366-9516.2005.00213.x
Martínez-Iglesias, J.C., Carvacho, A. & Ríos, R. (1996) Catálogo de los carídeos marinos (Crustacea, Decapoda, Caridea) de
las aguas someras de Cuba. Avicennia, 4/5, 27–40.
Martínez-Iglesias, J.C., Gómez, O., Carvacho, A. & Ríos, R. (1993) Nuevos registros de crustáceos decápodos (Crustacea:
Decapoda) en la plataforma marina de Cuba. Avicennia, 0, 9–13.
Milne Edwards, H. (1834–1840) Histoire naturelle des crustacés, comprenant l’anatomie, la physiologie et la classification de
ces animaux, Librairie encyclopédique de Roret, Paris, 468 + 532 + 638 + 32 pp., 42 pls.
Nardo, G.D. (1847) Prospetto della fauna marina volgare del Veneto estuario con cenni sulle principali specie commestibile
dell'Adriatico, ecc. Estratto dall opera, Venezue e le sue lagune, Venezia, 1–44.
Rafinesque, C.S. (1815) Analyse de la nature ou tableau de l’univers et des corps organisés. Palermo, 224 pp.
Ríos, R. & Duffy, J.E. (2007) A review of the sponge-dwelling snapping shrimp from Carrie Bow Cay, Belize, with description
of Zuzalpheus, new genus, and six new species (Crustacea: Decapoda: Alpheidae). Zootaxa, 1602, 3–89.
Say, T. (1818) An account of the Crustacea of the United States. Journal of the Academy of Natural Sciences of Philadelphia, 1,
235–253.
Spence Bate, C. (1888) Report on the Crustacea Macrura collected by the Challenger during the years 1873–76. Report on the
Scientific Results of the Voyage of H.M.S. ”Challenger” during the years 1873–76, 24, i-xc, 1-942, pls. 1–157.
ANKER & PACHELLE232 · Zootaxa 3815 (2) © 2014 Magnolia Press
Top Related
Copyright © 2022 FDOKUMEN

