







Bahasa
Halaman
Hukum


ORIGINAL PAPER
Salt tolerance of the annual halophyte Cakile maritima as affectedby the provenance and the developmental stage
Wided Megdiche Æ Nader Ben Amor Æ Ahmed Debez ÆKamel Hessini Æ Riadh Ksouri Æ Yasmine Zuily-Fodil ÆChedly Abdelly
Received: 19 September 2006 / Revised: 9 January 2007 / Accepted: 11 January 2007 / Published online: 21 March 2007
� Franciszek Gorski Institute of Plant Physiology, Polish Academy of Sciences, Krakow 2007
Abstract Though halophytes are naturally adapted to
salinity, their salt-tolerance limits are greatly influenced by
their provenance and developmental stage. In the present
study, physio-biochemical responses of two Tunisian eco-
types of the oilseed coastal halophyte Cakile maritima
(Brassicaceae) to salinity (0–400 mM NaCl) were moni-
tored during germination and vegetative growth stages.
Tabarka and Jerba seeds were collected from humid or arid
climatic areas, respectively. Plant response to salinity ap-
peared to depend on the ecotype and salinity levels.
Increasing salinity inhibited germination process. Jerba
seeds were found to be more salt tolerant than the Tabarka
ones. At the autotrophic stage of growth and under salt-free
conditions, Jerba was less productive than Tabarka (in
terms of dry matter accumulation), but plant biomass
production and leaf expansion (area and number) of the
former ecotype were progressively improved by 100 mM
NaCl, as compared to the control. In contrast, at the same
salt concentration, these parameters decreased under
increasing salinity in Tabarka (salt sensitive). Leaf
chlorophyll content was reduced at severe salinity, but this
effect was more conspicuous in the sensitive Tabarka
plants. Na+ contents in the Jerba and Tabarka leaves col-
lected from the 400 mM NaCl-treated plants were 17- and
12-fold higher than in the respective controls. This effect
was accompanied by a significant reduction in the leaf K+,
Mg2+ and Ca2+ contents, especially in the salt-treated
Tabarka. A significant accumulation of proline and soluble
carbohydrates in leaves was found during the period of
intensive leaf growth. These organic compounds likely
play a role in leaf osmotic adjustment and in protection of
membrane stability at severe salinity.
Keywords Cakile maritima � Compatible solute �Germination � Ion accumulation � Provenance �Salt tolerance � Vegetative growth stage
Introduction
Salinity is an increasing environmental problem throughout
the world. According the estimation of the United Nations
Environment Program, 20% of cultivated land worldwide
is adversely affected by a high salt concentration, which
inhibits plant growth and yield (Flowers and Yeo 1995).
The deleterious effects of salinity on plant growth are
associated with low osmotic potential of soil solution
(water stress), nutritional imbalance, specific ion effects
(salt stress), or a combination of these factors (Ashraf
1994; Ashraf and Harris 2004; Marshner 1995).
Saline soils occur naturally in both coastal region, where
groundwater is contaminated by seawater, and in areas
subjected to inadequate irrigation and/or draining (De
Souza Filho et al. 2003). One of the most common
approaches suggested for dealing with salinity is the
Communicated by A. Kacperska-Lewak.
W. Megdiche � N. B. Amor � A. Debez � K. Hessini �R. Ksouri (&) � C. Abdelly
Laboratoire d’Adaptation des Plantes aux Stress Abiotiques,
Centre de Biotechnologie a la Technopole de Borj-Cedria
(CBBC), BP 901, 2050 Hammam-lif, Tunisia
e-mail: [email protected]; [email protected]
Present address:
A. Debez
Institut fur Botanik, Universitat Hannover,
Herrenhauser Str. 2, 30419 Hannover, Germany
Y. Zuily-Fodil
Laboratoire d’Ecophysiologie Moleculaire de l’Universite Paris
XII - Val de Marne, Paris, France
123
Acta Physiol Plant (2007) 29:375–384
DOI 10.1007/s11738-007-0047-0

utilization of genetic engineering to increase salt tolerance
of common crop species (Maggio et al. 2000). The out-
come of such an approach, although promising, remains
still non significant, because salt tolerance is a complex
trait. An alternative approach may consist in the utilization
of salt-tolerant species (halophytes), being native of the
region (Lieth et al. 1999). Indeed, halophytic species are
known to represent a potentially important crop (O’Leary
1984; Glenn et al. 1999), in addition to their ecological role
in rehabilitation of damaged ecosystems and/or in land-
scaping (Debez et al. 2004). Salt-tolerant plants, growing
normally in saline coastal ecosystems, evolved a number of
adaptive traits expressed at various levels of organization,
which allow them to grow and achieve their complete cycle
of development under such hostile conditions (Flowers
et al. 1986; Tipirdamaz et al. 2005). Such behaviour im-
plies a complexity of salt tolerance mechanisms, among
which osmotic adjustment (usually accomplished by taking
up inorganic ions), as well as an accumulation of com-
patible solutes plays the most important role (Meloni et al.
2004). Nevertheless, at the earliest stages of the life cycle
(germination, seedling establishment), halophytes are
generally as salt-sensitive as glycophytes (Debez et al.
2001; Khan et al. 2002).
Cakile maritima (sea rocket) is a succulent, annual
halophyte, confined to maritime strandlines on sand or
shingle and associated fore dunes. This species shows
considerable variability, within and between subspecies
(Davy et al. 2006). Genotypic differences play a major role
in adaptation of plants to their specific environments
(Gunasekera et al. 2006). As a part of a national project
aiming at the evaluation of local halophytes with eco-
nomical and/or ecological potentials, the physiology of salt
tolerance of C. maritima was investigated using a single
Tunisian ecotype (Debez et al. 2004). In continuation of
this work, the present study was aimed at monitoring the
salt-induced responses in plants of two Tunisian ecotypes
of C. maritima at different developmental stages, with a
special emphasis put on seed-germination capacity, accu-
mulation of ions and compatible solutes (soluble carbo-
hydrates and proline) in leaves and the role of these
substances in osmotic adjustment. So far, differences in C.
maritima behaviour with respect to its developmental stage
and geographic origin are poorly documented.
Materials and methods
Plant material and culture conditions
Seeds of C. maritima were collected at two Tunisian
littoral sites: at Tabarka (extreme north, humid climatic
area) and at Jerba (south, arid climatic area). After their
disinfection, seeds were germinated in Petri dishes (20
seeds each) containing two layers of filter paper moistened
with salt solutions of different concentrations (in the
0–400 mM NaCl range). Petri dishes (three replicates per
treatment) were incubated in the dark at 20 ± 2�C for
2 weeks. Germination was considered as occurred when
emerging radicule was visible. In addition to the final
germination percentage, a theoretical model (Debez et al.
2004) was used for a more accurate assessment of salt
impact on seed germination. Such a mathematical simula-
tion assumes that germination is composed of two phases: a
latency phase of duration t0, during which the seeds acquire
the aptitude to germinate, and the germination itself. Suc-
ceeding the latency phase, the probability k of germination
per unit of time is equal and constant with time for all
seeds. The model was formulated as:
yðtÞ ¼ ymaxð1þ e�kðt�t0ÞÞ: ð1Þ
With y(t)- the number of germinated seeds at time t and
ymax-the plateau reached by y(t), the number of viable seeds
is determined. The time interval between the end of latency
and reaching the plateau is determined by t0 and k together.
k depends on time needed for germination of 50% of viable
seeds and is calculated as:
t50% ¼ t0 þ lnð2Þ=k: ð2Þ
The values of ymax, k and t0 were determined by fitting
the observed values to the above equation using the non-
linear regression module of the StatisticaTM program.
In the second experiment, seeds were germinated in pots
filled with inert and humid sand. Ten-day-old seedlings,
selected for their uniformity in size and form, were culti-
vated in pots under greenhouse conditions (25 ± 5�C
temperature, 60 ± 10% relative humidity). Three-week-old
plants were then irrigated at alternate days with a nutrient
solution (Hewitt 1966), pH 7.3, containing 0, 100, 200, or
400 mM NaCl. Leaf samples (originating from four plants
per treatment) harvested at 4, 6, 10, 15, and 20 days after
salt addition to the culture medium, were used for moni-
toring the main growth parameters, ion status and organic
solute contents in the studied tissues.
Determination of chlorophyll content
Chlorophyll content (mg g–1 FW) in mature leaves was
determined according to Torrecillas et al. (1984), with a
slight modification (we used a pure acetone rather than
acetone 80%). Five ml of pure acetone was added to fresh
leaf samples (ca. 100 mg each), cut into discs. The
extraction took place in darkness at 4�C for 72 h. Extract
absorbance was measured at 649 and 665 nm and total
376 Acta Physiol Plant (2007) 29:375–384
123

chlorophyll content was calculated according to the fol-
lowing equation:
Total chlorophyll (lg ml�1Þ¼ 6:45 ðA665 nmÞ þ 17:72ðA649 nmÞ: ð3Þ
Leaf growth, water relations, and ion assay
Leaf growth was assessed by determining their total dry
weight (DW). Total leaf area was measured by a leaf area
meter (Portable Areameter LI/3000A LI-COR). Leaf water
content was calculated as ml H2O per g DW. Dry weight of
the samples was estimated after their drying at 60�C for
72 h.
After ion extraction from leaf samples (30 mg DW
each) in 0.5% HNO3, chloride was assayed by coulometry
(Buchler chloridometer), Na+ and K+ by flame emission
photometry (Corning, UK), and Ca2+ and Mg2+ by atomic
absorption spectrophotometry (Perkin Elmer 4000).
Proline and total soluble carbohydrate contents
Proline was determined following the ninhydrin method
described by Bates et al. (1973), using L-proline as a
standard. Leaf samples (100 mg FW) were homogenized in
1.5 ml of 3% (w/v) aqueous sulfosalicylic acid and cen-
trifuged for 30 min at 14,000g. To the supernatant
(500 ll), 2 ml of acid ninhydrin and 2 ml of glacial acetic
acid were added and the mixture was boiled for 1 h. After
extraction with toluene, the free proline was quantified
(k = 520 nm) from the organic phase using an Anthelie
Advanced 2, SECOMAN spectrophotometer. Proline in the
test samples was calculated from a standard curve prepared
against L-Proline (5–30 lg, from MERCK KGaA):
y = 0.059x – 0.014, R2 = 0.99.
The content of total soluble carbohydrates in the studied
samples was determined according to Mc Cready et al.
(1950) and Staub (1963), using glucose as a standard.
Twenty-five milligram (DW) leaf samples was homoge-
nized with 5 ml methanol 80% and boiled while shaking at
70�C for 30 min. The homogenate was centrifuged for
15 min at 6,000g. After decanting, the residue was re-
suspended in 5 ml of the extraction solution and centri-
fuged at 6,000 g for 10 min. The supernatant was decanted
and combined with the original extract. For measurement
of total soluble carbohydrates, anthrone–sulfuric acid assay
was used. An aliquot of 250 ll was added to 5 ml of an-
throne–sulfuric acid solution. The mixture was shaken,
heated in a boiling water-bath for 10 min and cooled at
4�C. The absorption was determined by spectrophotometry
(Anthelie Advanced 2, SECOMAN) at 640 nm. A standard
curve was prepared using different concentration of glu-
cose (0–100 lg, from MERCK KGaA). From the standard
curve, the concentrations of soluble carbohydrates in the
test samples were calculated (y = 0.0095x – 0.0299,
R2 = 0.979).
Experimental design and statistical analysis
The germination data were subjected to a two-way analysis
of variance (ANOVA), using salinity (S) and ecotype (A)
as the factors. The means were compared by Newman–
Keuls post-hoc test at 5% probability. Concerning the plant
culture in a greenhouse, the experimental design was a
completely randomized 2 · 2 factorial with four replicates
per treatment.
Results
Seed germination
Germination capacity was dependent on the plant ecotype
and applied salinity. In a salt-free medium, germination
percentages were significantly higher in Jerba as compared
to Tabarka (100% and ca. 40%, respectively) (Fig. 1a).
Salinity in the 50–300 mM NaCl range decreased signifi-
cantly germination percentages in both ecotypes, although
Jerba was more tolerant. In both ecotypes, germination was
virtually suppressed at 400 mM NaCl (Fig. 1b). On the
other hand, salinity delayed germination process, as indi-
cated by the increase of t0 and t50% (Fig. 1c), with a more
marked effect on Tabarka seedlings (sensitive).
Leaf development, water relations and total chlorophyll
content
In the control treatment (plants grown in the absence of
salt), Tabarka seedlings were more productive than Jerba,
as indicated by dry matter increases (Fig. 2). Yet, the latter
showed a maximal growth potential at a moderate salinity
level, i.e. at 100 mM NaCl, which indicates a typical hal-
ophytic behaviour. In contrast, growth of Tabarka seedlings
markedly decreased at 100 mM NaCl (Fig. 2). Salinity
levels higher than 100 mM NaCl restricted significantly the
plant growth, although to a greater extent in the sensitive
Tabarka plants. This ecotype-dependent salt response was
more pronounced along with the duration to salt exposure.
At 400 mM NaCl, plant biomass production represented 77
and 28% of the control values in Jerba and Tabarka eco-
types, respectively.
Leaf growth was also salt- and ecotype dependent. For
instance, the moderate salinity (100 mM NaCl) increased a
Acta Physiol Plant (2007) 29:375–384 377
123

leaf number and area in Jerba seedlings, comparing to the
control (Fig. 3a, b), but it reduced leaf growth in Tabarka.
However, the 400 mM NaCl-treated plants of both eco-
types showed significantly smaller leaf area, as compared
to other treatments (Fig. 3b).
Irrespective of the ecotype, total chlorophyll content in
the leaves increased during the culture period (Fig. 4). In
Jerba plants treated with 100 mM NaCl, the chlorophyll
content was maintained at the level similar to that in the
control. This parameter was significantly reduced by severe
salinity, and to a higher extent in Tabarka seedlings, as
seen after 20 days of the salt-treatment (by ca. 20 and 30%
in Jerba and Tabarka ecotypes, respectively). Nevertheless,
neither necrosis nor chlorosis symptoms were observed in
leaves of plants submitted to severe salinity.
Changes in leaf water content paralleled those of bio-
mass production. In both ecotypes, water content in seed-
lings subjected to moderate salinity was maintained near to
the control level (Fig. 5). Increasing salinity resulted in
a progressive decrease in leaf hydration, especially in
Tabarka plants (Fig. 5). At 400 mM NaCl, the reduction of
water content in Jerba and Tabarka leaves reached 30 and
51%, as compared to the respective controls.
Ion content and solute accumulation
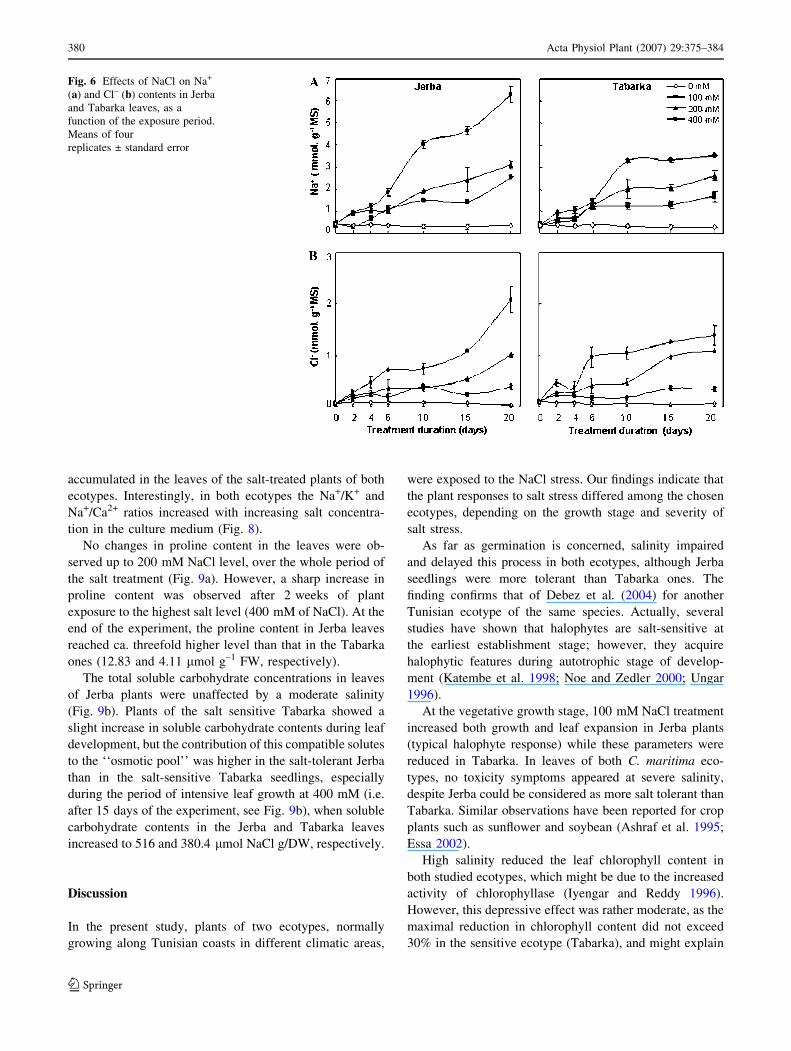
In Jerba leaves, Na+ content increased with salinity along
with the exposure time (Fig. 6a). In Tabarka, Na+ content
reached a plateau 15 days after the start of the exposure. In
plants challenged with 400 mM, NaCl content in Jerba and
Fig. 1 Effects of NaCl on
germination parameters in two
ecotypes of C. maritima.
Observed germination
percentage (a), ymax (the
number of viable seeds, as % of
the sown seeds) (b), the time for
50% germination (t50%), given
by t0 + ln(2)/k (c), the
probability of germination per
time unit, k (d). The values of
ymax, t0, and k were determined
by fitting Eq. 1 to the observed
germination kinetic data. Means
of three replicates ± standard
error. Values followed by at
least one the same letter were
not significantly different at
P < 0.05, according to the
Newman–Keuls post-hoc test
Fig. 2 Effects of NaCl on the
leaf dry weight of Jerba and
Tabarka ecotypes, as a function
of the exposure period. Means
of four replicates ± standard
error
378 Acta Physiol Plant (2007) 29:375–384
123

Tabarka leaves was ca. 17- and 12-fold greater than in the
control values, respectively (Fig. 6a). Sodium accumula-
tion was associated with an increase in chloride uptake
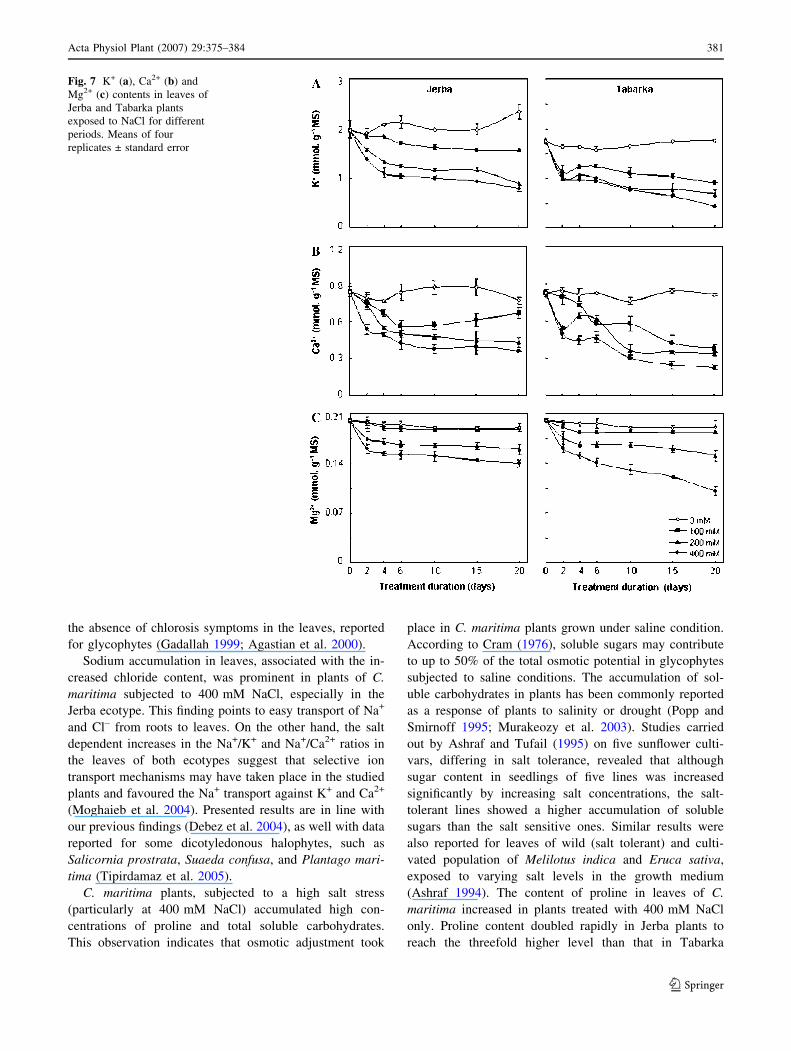
(Fig. 6b), and with a marked decrease in K+, Ca2+ and
Mg2+ contents (Fig. 7). A more pronounced impact of
salinity was observed in the salt-sensitive Tabarka plants,
especially at high salinity levels (>100 mM NaCl). In fact,
both Na+ and Cl– were found to be the principal ions
Fig. 3 Effects of NaCl on leaf
area (a) and leaf number (b) in
Jerba and Tabarka ecotypes, as
a function of the exposure
period. Means of four
replicates ± standard error
Fig. 4 Effects of NaCl on total
chlorophyll content in Jerba and
Tabarka leaves, as a function of
the exposure period. Means of
four replicates ± standard error
Fig. 5 Effects of NaCl on water
content in Jerba and Tabarka
leaves, as a function of the
exposure period. Means of four
replicates ± standard error
Acta Physiol Plant (2007) 29:375–384 379
123
accumulated in the leaves of the salt-treated plants of both
ecotypes. Interestingly, in both ecotypes the Na+/K+ and
Na+/Ca2+ ratios increased with increasing salt concentra-
tion in the culture medium (Fig. 8).
No changes in proline content in the leaves were ob-
served up to 200 mM NaCl level, over the whole period of
the salt treatment (Fig. 9a). However, a sharp increase in
proline content was observed after 2 weeks of plant
exposure to the highest salt level (400 mM of NaCl). At the
end of the experiment, the proline content in Jerba leaves
reached ca. threefold higher level than that in the Tabarka
ones (12.83 and 4.11 lmol g–1 FW, respectively).
The total soluble carbohydrate concentrations in leaves
of Jerba plants were unaffected by a moderate salinity
(Fig. 9b). Plants of the salt sensitive Tabarka showed a
slight increase in soluble carbohydrate contents during leaf
development, but the contribution of this compatible solutes
to the ‘‘osmotic pool’’ was higher in the salt-tolerant Jerba
than in the salt-sensitive Tabarka seedlings, especially
during the period of intensive leaf growth at 400 mM (i.e.
after 15 days of the experiment, see Fig. 9b), when soluble
carbohydrate contents in the Jerba and Tabarka leaves
increased to 516 and 380.4 lmol NaCl g/DW, respectively.
Discussion
In the present study, plants of two ecotypes, normally
growing along Tunisian coasts in different climatic areas,
were exposed to the NaCl stress. Our findings indicate that
the plant responses to salt stress differed among the chosen
ecotypes, depending on the growth stage and severity of
salt stress.
As far as germination is concerned, salinity impaired
and delayed this process in both ecotypes, although Jerba
seedlings were more tolerant than Tabarka ones. The
finding confirms that of Debez et al. (2004) for another
Tunisian ecotype of the same species. Actually, several
studies have shown that halophytes are salt-sensitive at
the earliest establishment stage; however, they acquire
halophytic features during autotrophic stage of develop-
ment (Katembe et al. 1998; Noe and Zedler 2000; Ungar
1996).
At the vegetative growth stage, 100 mM NaCl treatment
increased both growth and leaf expansion in Jerba plants
(typical halophyte response) while these parameters were
reduced in Tabarka. In leaves of both C. maritima eco-
types, no toxicity symptoms appeared at severe salinity,
despite Jerba could be considered as more salt tolerant than
Tabarka. Similar observations have been reported for crop
plants such as sunflower and soybean (Ashraf et al. 1995;
Essa 2002).
High salinity reduced the leaf chlorophyll content in
both studied ecotypes, which might be due to the increased
activity of chlorophyllase (Iyengar and Reddy 1996).
However, this depressive effect was rather moderate, as the
maximal reduction in chlorophyll content did not exceed
30% in the sensitive ecotype (Tabarka), and might explain
Fig. 6 Effects of NaCl on Na+
(a) and Cl– (b) contents in Jerba
and Tabarka leaves, as a
function of the exposure period.
Means of four
replicates ± standard error
380 Acta Physiol Plant (2007) 29:375–384
123
the absence of chlorosis symptoms in the leaves, reported
for glycophytes (Gadallah 1999; Agastian et al. 2000).
Sodium accumulation in leaves, associated with the in-
creased chloride content, was prominent in plants of C.
maritima subjected to 400 mM NaCl, especially in the
Jerba ecotype. This finding points to easy transport of Na+
and Cl– from roots to leaves. On the other hand, the salt
dependent increases in the Na+/K+ and Na+/Ca2+ ratios in
the leaves of both ecotypes suggest that selective ion
transport mechanisms may have taken place in the studied
plants and favoured the Na+ transport against K+ and Ca2+
(Moghaieb et al. 2004). Presented results are in line with
our previous findings (Debez et al. 2004), as well with data
reported for some dicotyledonous halophytes, such as
Salicornia prostrata, Suaeda confusa, and Plantago mari-
tima (Tipirdamaz et al. 2005).
C. maritima plants, subjected to a high salt stress
(particularly at 400 mM NaCl) accumulated high con-
centrations of proline and total soluble carbohydrates.
This observation indicates that osmotic adjustment took
place in C. maritima plants grown under saline condition.
According to Cram (1976), soluble sugars may contribute
to up to 50% of the total osmotic potential in glycophytes
subjected to saline conditions. The accumulation of sol-
uble carbohydrates in plants has been commonly reported
as a response of plants to salinity or drought (Popp and
Smirnoff 1995; Murakeozy et al. 2003). Studies carried
out by Ashraf and Tufail (1995) on five sunflower culti-
vars, differing in salt tolerance, revealed that although
sugar content in seedlings of five lines was increased
significantly by increasing salt concentrations, the salt-
tolerant lines showed a higher accumulation of soluble
sugars than the salt sensitive ones. Similar results were
also reported for leaves of wild (salt tolerant) and culti-
vated population of Melilotus indica and Eruca sativa,
exposed to varying salt levels in the growth medium
(Ashraf 1994). The content of proline in leaves of C.
maritima increased in plants treated with 400 mM NaCl
only. Proline content doubled rapidly in Jerba plants to
reach the threefold higher level than that in Tabarka
Fig. 7 K+ (a), Ca2+ (b) and
Mg2+ (c) contents in leaves of
Jerba and Tabarka plants
exposed to NaCl for different
periods. Means of four
replicates ± standard error
Acta Physiol Plant (2007) 29:375–384 381
123

seedlings. Proline content has been found to be higher in
many salt-tolerant plants than in salt sensitive ones. This
amino acid, which occurs widely in the salt stressed
plants, accumulates in larger amounts than other amino
acids and could be considered as an indicator of salt
tolerance (Ali et al. 1999; Ashraf 1993, 1994). In line
with this suggestion, accumulation of proline was found to
be higher in salt-tolerant alfalfa plants than in the sensi-
tive ones (Fougere et al. 1991; Petrusa and Winicov
1997). Higher proline accumulation was observed also in
salt-tolerant ecotypes of Agrostis stolonifera than in salt
sensitive ones (Ahmad et al. 1981).
Fig. 8 Leaf Na+/K+ (a) and
Na+/Ca2+ (b) ratios in two
ecotypes of C. maritima grown
in nutrient solution containing
0, 100, 200 or 400 mM NaCl, as
a function of the exposure
period. Means of four
replicates ± standard error
Fig. 9 Proline (a), and soluble
carbohydrates (b) in leaves of
Jerba and Tabarka plants
exposed to NaCl for different
periods. Means of four
replicates ± standard error
382 Acta Physiol Plant (2007) 29:375–384
123

In our study, the increase in proline content occurred
when water content in the leaves decreased to 50% in
comparison with the control. Leaf water potential of plants
exposed to 400 mM NaCl was found to be fivefold lower in
Jerba than in Tabarka at the end of the experiment (data not
shown). Therefore, proline accumulation may be associ-
ated with a decrease in water potential of tissues, as re-
ported by Slama et al. (2007) for the salt-treated halophyte
Sesuvium portulacastrum. The difference in ww seemed to
be related to a high accumulation of sodium ions, soluble
carbohydrates and proline in Jerba and thus, may partially
explain its high salt tolerance. According to Ali et al.
(1999), the accumulation of these compatible solutes and
certain ions in response to osmotic stress represents an
important adaptive response to salt and drought stress.
In summary, presented experiments showed marked
differences in salt tolerance of C. maritima plants of two
Tunisian ecotypes. Jerba was more salt tolerant than Ta-
barka, at both germination and vegetative growth stages.
This was related to a greater ability of the former to
maintain osmotic adjustment resulting from its higher
ability to accumulate sodium as well as proline and car-
bohydrates within leaf cells in response to salinity. This
difference between plants of two ecotypes of C. maritima
may reflect the difference in climatic conditions of their
native habitats.
Acknowledgments This work was supported by the Tunisian-
French ‘‘Comite Mixte de Cooperation Universitaire’’ (CMCU)
network # 02FO924.
References
Ahmad I, Wainwright SJ, Stewart GR (1981) The solute and water
relations of Agrostis stolonifera ecotypes differing in their salt
tolerance. New Phytol 87:615–629
Ali G, Srivastava PS, Iqbal M (1999) Proline accumulation, protein
pattern and photosynthesis in regenerants grown under NaCl
stress. Biol Plant 42:89–95
Agastian P, Kingsley SJ, Vivekanandan M (2000) Effect of salinity on
photosynthesis and biochemical characteristics in mulberry
genotypes. Photosynthetica 38:287–290
Ashraf M (1993) Effect of sodium chloride on water relations and
some organic osmotica in arid zone plant species Melilotusindica (L.) All Der Tropen 94:95–102
Ashraf M (1994) Organic substances responsible for salt tolerance in
Eruca sativa. Biol Plant 36:61–71
Ashraf M, Harris PJC (2004) Potential biochemical indicator of
salinity tolerance in plants. Plant Sci 166:3–16
Ashraf M, Tufail M (1995) Variation in salinity tolerance in
sunflower (Helianthus annuus L.). J Agr Soil Sci 174:351–362
Ashraf M, Zafar ZU, O’Learly JW (1995) Genetic variation for salt
tolerance in sunflower (Helianthus annuus L.). Hereditas
123:141–145
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free
proline for water stress studies. Plant Soil 39:205–207
Cram WJ (1976) Negative feedback regulation of transport in cells.
The maintenance of turgor, volume and nutrient supply. In:
Luttge U, Pitman MG (ed.) Encyclopaedia of plant physiology,
vol. 2, New Series. Springer, Berlin, pp 284–316
Davy AJ, Scott R, Cordazzo CV (2006) Biological flora of the British
Isles: Cakile maritima Scop. J Ecol 94:695–711
De Souza Filho GA, Ferreira BS, Dias JM, Queiroz KS, Branco AT,
Bressan-Smith RE, Oliveira JG, Garcia AB (2003) Accumula-
tion of salt protein in rice plants as a response to environmental
stresses. Plant Sci 164:623–628
Debez A, Chaıbi W, Bouzid S (2001) Effet de NaCl et de regulateurs
de croissance sur la germination d’Atriplex halimus L. Cahiers
Agric 10:135–138
Debez A, Ben Hamed K, Grignon C, Abdelly C (2004) Salinity
effects on germination, growth, and seed production of the
halophyte Cakile maritima. Plant Soil 262:179–189
Essa TA (2002) Effect of salinity stress on growth and nutrient.
Composition of Three Soybean (Glycine max L. Merrill)
Cultivars. J Agr Crop Sci 188:86–93
Flowers TJ, Hajibagheri MA, Clipson NJW (1986) Halophytes. Quart
Rev Biol 61:313–337
Flowers TJ, Yeo AR (1995) Breeding for salinity resistance in crop
plants: where next? Aust. J Plant Physiol 22:875–884
Fougere F, Rudulier D, Streeter JG (1991) Effects of salt stress on
amino acid, organic acid, and carbohydrate composition of roots,
bacteroids, and cytosol of alfalfa (Medicago sativa L.). Plant
Physiol 96:1228–1236
Gadallah MAA (1999) Effects of proline and glycinebetaine on Viciafaba response to salt stress. Biol Plant 42:249–257
Glenn EP, Brown JJ, Blumwald E (1999) Salt tolerance and crop
potential of halophytes. Crit Rev Plant Sci 18:227–255
Gunasekera CP, Martin LD, Siddique KHM, Walton GH (2006)
Genotype by environment interactions of Indian mustard (Bras-sica juncea L.) and canola (B. napus L.) in Mediterranean-type
environments. Crop growth and seed yield Europ J Agr 25:1–12
Hewitt EJ (1966) Sand and water culture methods used in the study of
plant nutrition. Common wealth Bureau of Horticulture. Tech
Com 22:431–446
Katembe WJ, Ungar IA, Mitchell JP (1998) Effect of salinity on
germination and seedling growth of two Atriplex species(Chenopodiaceae). Ann Bot 82:167–175
Khan MA, Gul B, Weber D (2002) Seed germination in the great
basin halophyte Salsola iberica. Can J Bot 80:650–655
Iyengar ERR, Reddy MP (1996) Photosynthesis in high salt-tolerant
plants. In: Pesserkali M (ed) Hand book of photosynthesis.
Marshal Dekar, Baten Rose, pp 56–65
Lieth H, Moschenko M, Lohmann M, Koyro HW, Hamdy A (1999)
Halophyte uses in different climates. I. Ecological and ecophys-
iological studies. Backhuys Publishers, Leiden
Maggio A, Reddy MP, Joly RJ (2000) Gas exchange and solute
accumulation in the halophyte Salvadora persica grown at
moderate salinity. Env Exp Bot 44:31–38
Marschner H (1995) Mineral nutrition of higher plants. Academic,
London
Mc Cready RM, Guggolz J, Silviera V, Owes HS (1950) Determi-
nation of starch and amylase in vegetables, application to peas.
Annal Chem 22:1156–1158
Meloni DA, Gulotta MR, Martinez CA, Oliva MA (2004) The effect
of salt stress on growth, nitrate reduction and proline and glycine
betaine accumulation in Prosopis alba. Braz J Plant Physiol
16(1):39–46
Moghaieb REA, Saneoka H, Fujita K (2004) Effect of salinity on
osmotic adjustment, glycine betaine accumulation and the
betaine aldehyde dehydrogenase gene expression in two halo-
phytic plants, Salicornia europaea and Suaeda maritima. Plant
Sci 166:1345–1349
Murakeozy EP, Nagy Z, Duhaz C, Buchereau A, Tuba Z (2003)
Seasonal changes in the levels of compatible osmolytes in three
Acta Physiol Plant (2007) 29:375–384 383
123

halophytic species of inland saline vegetation in Hungary. J Plant
Physiol 160:395–401
Noe GB, Zedler JB (2000) Differential effects of four abiotic factors
on the germination of salt marsh annuals. Am J Bot 87:1679–
1682
O’Leary JW (1984) The role of halophytes in irrigated agriculture. In:
Staples RC, Toenniessen GH (eds) Salinity tolerance in plants:
strategies for crop improvement. Wiley, New York, pp 285–300
Petrusa LM, Winicov I (1997) Proline status in salt-tolerant and salt-
sensitive alfalfa cell lines and plants in response to NaCl. Plant
Physiol Biochem 35:303–310
Popp M, Smirnoff N (1995) Polyol accumulation and metabolism
during water deficit. In: Smirnoff N (ed) Environment and plant
metabolism: flexibility and acclimation. Bios Scientific, Oxford,
pp 199–215
Slama I, Ghnaya T, Messedi D, Hessini K, Labidi N, Savoure A,
Abdelly C (2007) Effect of sodium chloride on the response of
the halophyte species Sesuvium portulacastrum grown in man-
nitol-induced water stress. J Plant Res (in press)
Staub AM (1963) Extraction, identification et dosages des glucides
dans les extraits d’organes et les corps bacteriens, In: Masson et
Compagnie (eds) Techniques de laboratoire (Tome 1 et 2), Paris,
pp 1307–1366
Tipirdamaz R, Gagneul D, Duhaze C, Aınouche A, Monnier C,
ozkum D, Larher F (2005) Clustering of halophytes from an
inland salt marsh in Turkey according to their ability to
accumulate sodium and nitrogenous osmolytes. Env Exp Bot
57(1–2):139–153
Torrecillas A, Leon A, Del Amor F, Martinez-Monpean MC (1984)
Determinacion rapida de clorofila en discos foliares de limonero.
Fruits 39:617–622
Ungar IA (1996) Effect of salinity on seed germination, growth, and
ion accumulation of Atriplex patula (Chenopodiaceae). Am J Bot
83:604–607
384 Acta Physiol Plant (2007) 29:375–384
123
Top Related
Copyright © 2022 FDOKUMEN

