





![bronchial-hygiene-therapy.ppt [Read-Only] - Semantic Scholar](https://static.fdokumen.com/doc/165x107/6317b9679076d1dcf80beb6a/bronchial-hygiene-therapyppt-read-only-semantic-scholar.jpg)

Bahasa
Halaman
Hukum


Respiratory Syncytial Virus Inhibits Ciliagenesis inDifferentiated Normal Human Bronchial Epithelial Cells:Effectiveness of N-AcetylcysteineManuel Mata1,2*, Irene Sarrion1, Miguel Armengot3,4, Carmen Carda3, Isidoro Martinez2,5,
Jose A. Melero2,6, Julio Cortijo1,2,3
1 Research Foundation of the University General Hospital of Valencia, Valencia, Spain, 2 Centro de Investigacion Biomedica en Red (CIBER) de Enfermedades Respiratorias,
Valencia, Spain, 3 University of Valencia, Valencia, Spain, 4 University General Hospital of Valencia, Valencia, Spain, 5 Unidad de Interaccion Virus-Celula, Centro Nacional
de Microbiologıa, Instituto de Salud Carlos II, Madrid, Spain, 6 Unidad de Biologıa Viral, Centro Nacional de Microbiologıa, Instituto de Salud Carlos II, Madrid, Spain
Abstract
Persistent respiratory syncytial virus (RSV) infections have been associated with the exacerbation of chronic inflammatorydiseases, including chronic obstructive pulmonary disease (COPD). This virus infects the respiratory epithelium, leading tochronic inflammation, and induces the release of mucins and the loss of cilia activity, two factors that determine mucusclearance and the increase in sputum volume. These alterations involve reactive oxygen species-dependent mechanisms.The antioxidant N-acetylcysteine (NAC) has proven useful in the management of COPD, reducing symptoms, exacerbations,and accelerated lung function decline. NAC inhibits RSV infection and mucin release in human A549 cells. The mainobjective of this study was to analyze the effects of NAC in modulating ciliary activity, ciliagenesis, and metaplasia in primarynormal human bronchial epithelial cell (NHBEC) cultures infected with RSV. Our results indicated that RSV inducedultrastructural abnormalities in axonemal basal bodies and decreased the expression of b-tubulin as well as two genesinvolved in ciliagenesis, FOXJ1 and DNAI2. These alterations led to a decrease in ciliary activity. Furthermore, RSV inducedmetaplastic changes to the epithelium and increased the number of goblet cells and the expression of MUC5AC and GOB5.NAC restored the normal functions of the epithelium, inhibiting ICAM1 expression, subsequent RSV infection throughmechanisms involving nuclear receptor factor 2, and the expression of heme oxygenase 1, which correlated with therestoration of the antioxidant capacity, the intracellular H2O2 levels and glutathione content of NHBECs. The resultspresented in this study support the therapeutic use of NAC for the management of chronic respiratory diseases, includingCOPD.
Citation: Mata M, Sarrion I, Armengot M, Carda C, Martinez I, et al. (2012) Respiratory Syncytial Virus Inhibits Ciliagenesis in Differentiated Normal HumanBronchial Epithelial Cells: Effectiveness of N-Acetylcysteine. PLoS ONE 7(10): e48037. doi:10.1371/journal.pone.0048037
Editor: Hong Wei Chu, National Jewish Health, United States of America
Received February 23, 2012; Accepted September 20, 2012; Published October 31, 2012
Copyright: � 2012 Mata et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permitsunrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: This work was supported by grants SAF2005-00669/SAF2008-03113 (JC), PI10/02294 (MM), and CIBERES (CB06/06/0027) from the Ministry of Scienceand Innovation and the Health Institute ‘Carlos III’ of the Spanish government as well as research grants from regional government (GV2007/287 and AP073/10,Generalitat Valenciana). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing Interests: The authors have declared that no competing interests exist.
* E-mail: [email protected]
Introduction
Human respiratory syncytial virus (RSV; genus Pneumovirus,
family Paramixoviridae) is an important pathogen that causes serious
infection in people of all ages, including children, healthy and sick
adults, and elderly individuals [1,2]. This virus results in persistent
infection, leading to chronic inflammation through mechanisms
involving continuous stimulation of the immune system [3,4,5].
Persistent RSV infection appears to occur in certain individuals
with chronic obstructive pulmonary disease (COPD), where RSV
has been associated with exacerbation of the disease, the main
cause of morbidity in patients with COPD [6,7].
During COPD exacerbations, an increase occurs in sputum
volume, which in the airways is the result of a balance between the
ciliary beat of epithelial cells and mucin production [8]. Both
processes are affected by RSV, which induces the destruction of
ciliated epithelial cells and the expression of MUC5AC, the
predominant mucin gene expressed in human airways
[9,10,11,12].
The involvement of reactive oxygen intermediates (ROIs) as
mediators of the epithelial cell damage seen during exacerbations
seems clear [13]. The source of these oxidants may be leukocytes
via xanthine oxidase, which is increased in influenza-infected
lungs, or lung epithelial cells themselves [13,14,15]. ROIs are
necessary for RSV infection and are involved in the inflammatory
response of host cells [16].
N-acetylcysteine (NAC) is a thiol compound that acts directly as
a free radical scavenger and as a precursor of reduced glutathione
(GSH) [17]. This molecule reduces the number and impact of
COPD exacerbations [18] and also the inflammatory response in
epithelial cells infected with respiratory viruses [19]. Recently, the
effectiveness of NAC for inhibiting MUC5AC induction in A549
cells infected with RSV has been reported [16]. Although the anti-
mucolitic effects of NAC are well established, little is known about
its effects on ciliagenesis in human airway epithelial cells.
The main objective of this study was to analyze the effects of
NAC in an in vitro model of RSV infection developed on air–liquid
PLOS ONE | www.plosone.org 1 October 2012 | Volume 7 | Issue 10 | e48037
interface (ALI)-differentiated normal human bronchial epithelial
cells (NHBECs). We studied the effects of this drug on viral
replication, ciliary activity, ciliagenesis, and mucin production as
well as its antioxidant effects by measuring the total antioxidant
status (TAS), the intracellular H2O2 and glutathione levels and the
expression of nuclear receptor factor 2 (Nrf2), heme oxygenase 1
(HO1), and ICAM1.
Results
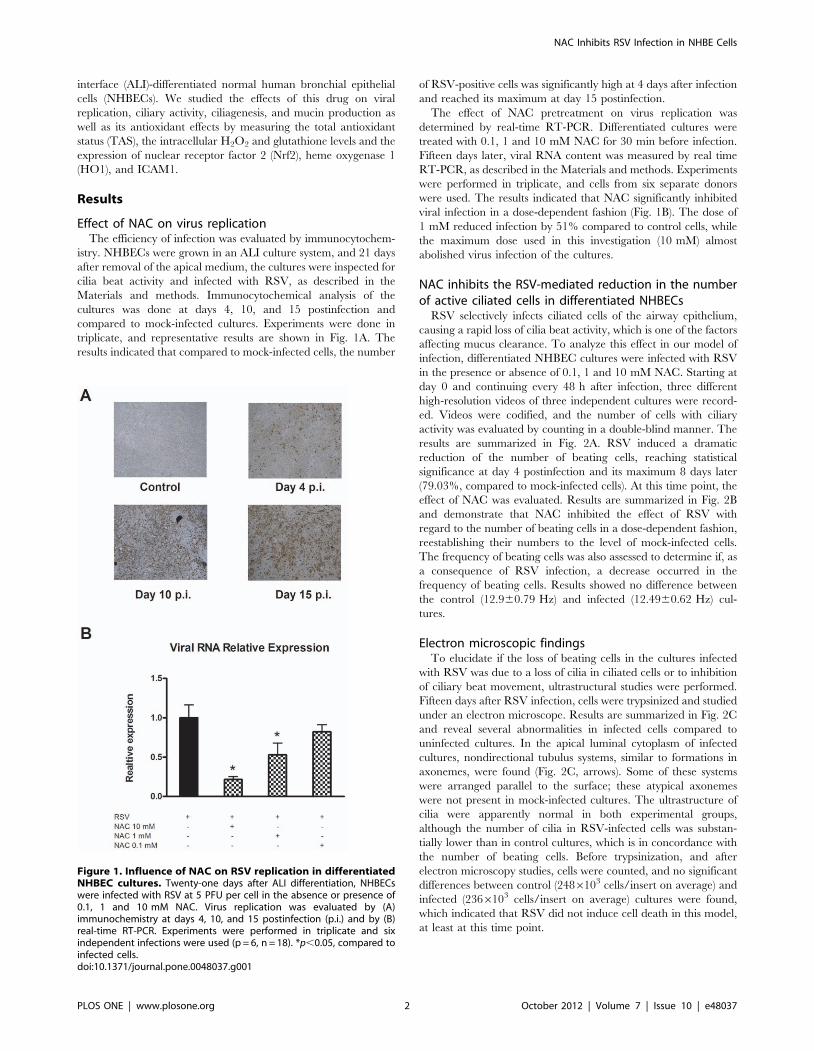
Effect of NAC on virus replicationThe efficiency of infection was evaluated by immunocytochem-
istry. NHBECs were grown in an ALI culture system, and 21 days
after removal of the apical medium, the cultures were inspected for
cilia beat activity and infected with RSV, as described in the
Materials and methods. Immunocytochemical analysis of the
cultures was done at days 4, 10, and 15 postinfection and
compared to mock-infected cultures. Experiments were done in
triplicate, and representative results are shown in Fig. 1A. The
results indicated that compared to mock-infected cells, the number
of RSV-positive cells was significantly high at 4 days after infection
and reached its maximum at day 15 postinfection.
The effect of NAC pretreatment on virus replication was
determined by real-time RT-PCR. Differentiated cultures were
treated with 0.1, 1 and 10 mM NAC for 30 min before infection.
Fifteen days later, viral RNA content was measured by real time
RT-PCR, as described in the Materials and methods. Experiments
were performed in triplicate, and cells from six separate donors
were used. The results indicated that NAC significantly inhibited
viral infection in a dose-dependent fashion (Fig. 1B). The dose of
1 mM reduced infection by 51% compared to control cells, while
the maximum dose used in this investigation (10 mM) almost
abolished virus infection of the cultures.
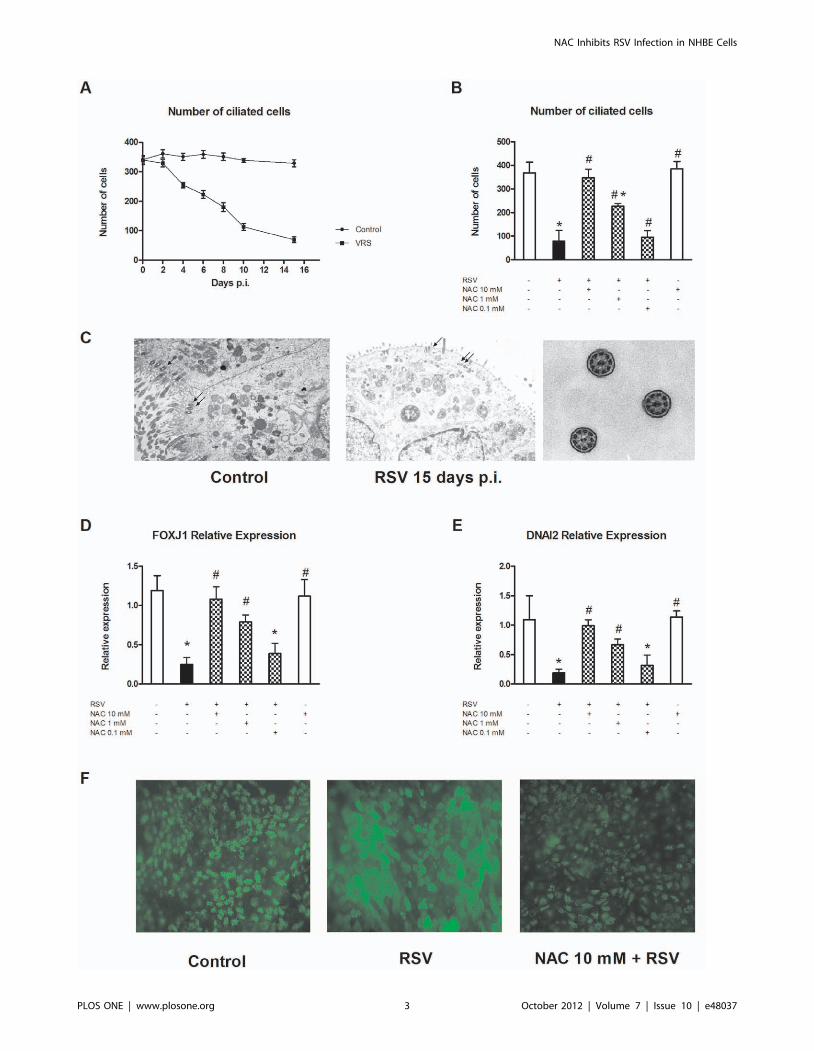
NAC inhibits the RSV-mediated reduction in the numberof active ciliated cells in differentiated NHBECs
RSV selectively infects ciliated cells of the airway epithelium,
causing a rapid loss of cilia beat activity, which is one of the factors
affecting mucus clearance. To analyze this effect in our model of
infection, differentiated NHBEC cultures were infected with RSV
in the presence or absence of 0.1, 1 and 10 mM NAC. Starting at
day 0 and continuing every 48 h after infection, three different
high-resolution videos of three independent cultures were record-
ed. Videos were codified, and the number of cells with ciliary
activity was evaluated by counting in a double-blind manner. The
results are summarized in Fig. 2A. RSV induced a dramatic
reduction of the number of beating cells, reaching statistical
significance at day 4 postinfection and its maximum 8 days later
(79.03%, compared to mock-infected cells). At this time point, the
effect of NAC was evaluated. Results are summarized in Fig. 2B
and demonstrate that NAC inhibited the effect of RSV with
regard to the number of beating cells in a dose-dependent fashion,
reestablishing their numbers to the level of mock-infected cells.
The frequency of beating cells was also assessed to determine if, as
a consequence of RSV infection, a decrease occurred in the
frequency of beating cells. Results showed no difference between
the control (12.960.79 Hz) and infected (12.4960.62 Hz) cul-
tures.
Electron microscopic findingsTo elucidate if the loss of beating cells in the cultures infected
with RSV was due to a loss of cilia in ciliated cells or to inhibition
of ciliary beat movement, ultrastructural studies were performed.
Fifteen days after RSV infection, cells were trypsinized and studied
under an electron microscope. Results are summarized in Fig. 2C
and reveal several abnormalities in infected cells compared to
uninfected cultures. In the apical luminal cytoplasm of infected
cultures, nondirectional tubulus systems, similar to formations in
axonemes, were found (Fig. 2C, arrows). Some of these systems
were arranged parallel to the surface; these atypical axonemes
were not present in mock-infected cultures. The ultrastructure of
cilia were apparently normal in both experimental groups,
although the number of cilia in RSV-infected cells was substan-
tially lower than in control cultures, which is in concordance with
the number of beating cells. Before trypsinization, and after
electron microscopy studies, cells were counted, and no significant
differences between control (2486103 cells/insert on average) and
infected (2366103 cells/insert on average) cultures were found,
which indicated that RSV did not induce cell death in this model,
at least at this time point.
Figure 1. Influence of NAC on RSV replication in differentiatedNHBEC cultures. Twenty-one days after ALI differentiation, NHBECswere infected with RSV at 5 PFU per cell in the absence or presence of0.1, 1 and 10 mM NAC. Virus replication was evaluated by (A)immunochemistry at days 4, 10, and 15 postinfection (p.i.) and by (B)real-time RT-PCR. Experiments were performed in triplicate and sixindependent infections were used (p = 6, n = 18). *p,0.05, compared toinfected cells.doi:10.1371/journal.pone.0048037.g001
NAC Inhibits RSV Infection in NHBE Cells
PLOS ONE | www.plosone.org 2 October 2012 | Volume 7 | Issue 10 | e48037
NAC Inhibits RSV Infection in NHBE Cells
PLOS ONE | www.plosone.org 3 October 2012 | Volume 7 | Issue 10 | e48037

NAC restores the reduction in the expression levels ofDNAI2, FOXJ1, and b-tubulin induced by RSV
Results obtained after analysis of the recorded videos and
electron microscopy studies pointed to interference of virus
infection and ciliagenesis in epithelial cells. To explore this, we
decided to analyze the expression of two important genes involved
in ciliogenesis, the transcriptional factor FOXJ1 and the axonemal
component DNAI2. Our results are presented in Fig. 2D and E,
respectively, and demonstrate that 15 days after RSV infection of
differentiated cultures, a significant inhibition of the expression of
both genes was observed (75% and 81%, respectively). NAC
pretreatment of cultures totally abolished this reduction in a dose-
dependent manner.
To determine if the downregulation of FOXJ1 and DNAI2
correlated with a decrease in ciliated cells, immunofluorescence
studies of b-tubulin were performed. The results obtained are
presented in Fig. 2F. A strong decrease in b-tubulin-positive cells
was observed at day 15 postinfection compared to mock-infected
cells. Pretreatment of cultures with 10 mM NAC strongly
ameliorated this effect.
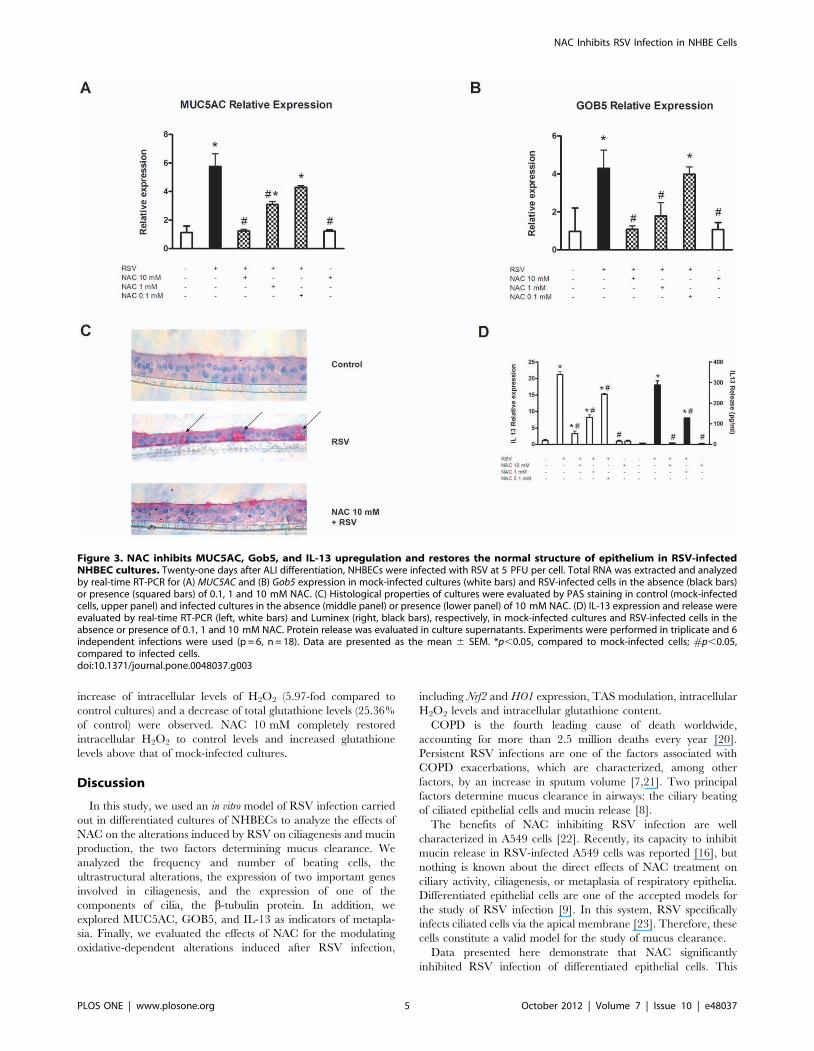
NAC inhibits MUC5AC, GOB5, and IL-13 upregulation inNHBECs infected with RSV: effects on goblet cellmetaplasia
Mucin hypersecretion is another important component that
determines mucus clearance by respiratory epithelium. One of the
effects observed after RSV infection of epithelial cells was an
increase in the expression of mucin MUC5AC. To investigate the
effect of NAC in this model, differentiated NHBECs were infected
with RSV in the absence or presence of 0.1, 1 and 10 mM NAC.
Fifteen days after infection, total RNA was extracted from cells,
and mucin MUC5AC mRNA levels were analyzed by real-time
RT-PCR. Our results indicated that after virus infection, a strong
induction of the expression of this gene occurred (5.76-fold
compared to mock-infected cultures; Fig. 3A). NAC inhibited this
upregulation in a dose-dependent manner.
Gob5 is a Ca2+-dependent chloride channel that regulates
MUC5AC expression. Real-time RT-PCR analysis of the expres-
sion of this gene revealed a significant increase in its expression
(4.29-fold compared to mock-infected cells) that was inhibited by
NAC in a dose-dependent manner (Fig. 3B).
To determine if this increase in MUC5AC expression was due to
an increase in the number of goblet cells, PAS staining was done.
NHBEC cultures were infected with human RSV for 15 days in
the presence or absence of 1 mM NAC. The results are presented
in Fig. 3C. After RSV infection, a decrease occurred in the
thickness of the cultures compared to mock-infected cells, as well
as an increase in the number of PAS-positive cells (arrows
indicated). NAC restored the thickness of the epithelium and the
number of goblet cells to normal levels.
Due to the involvement of IL-13 in goblet cell metaplasia, both
the expression and release levels of this cytokine were measured in
RSV-infected cultures. The results are summarized in Fig. 3D and
indicate a significant induction of expression (21.29-fold compared
to control cells) and release (45.80-fold compared to mock-infected
cells) of IL-13, which was inhibited by NAC pretreatment in a
dose-dependent fashion. The dose of 10 mM completely inhibited
the induction of this mediator at both the gene and protein
expression levels.
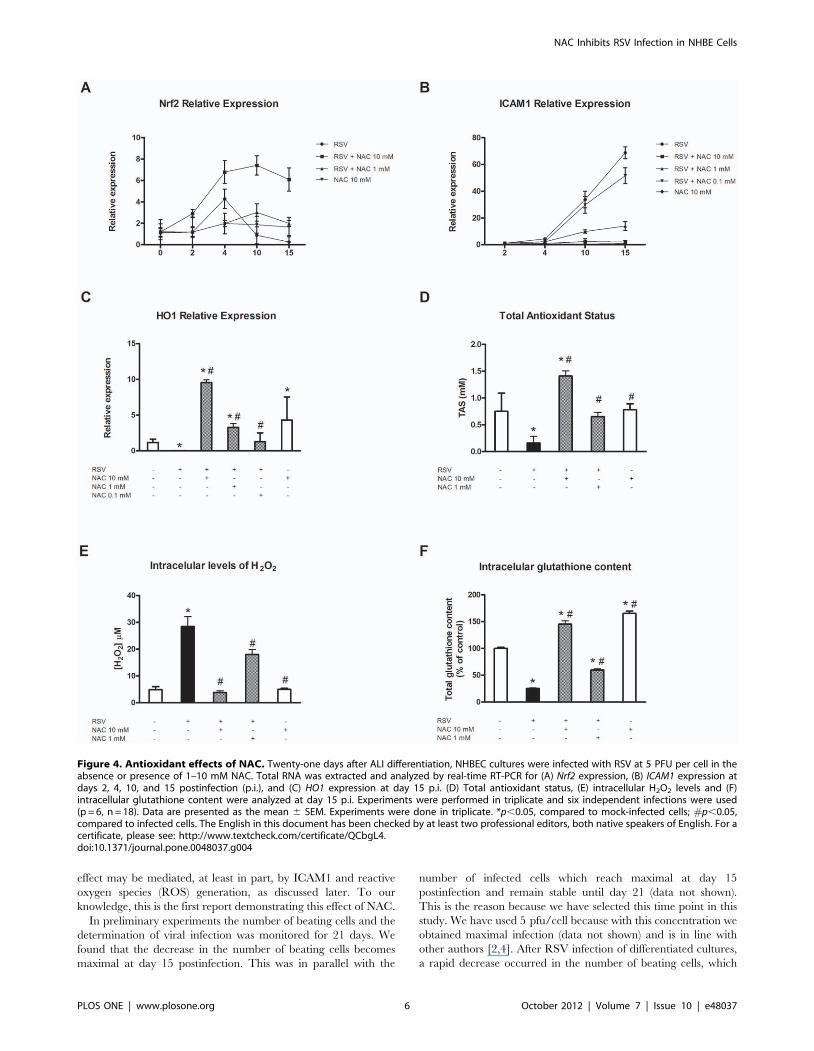
Oxidant events after human RSV infection of NHBECs:effects of NAC
ROI-mediated effects are clearly involved in the pathogenesis of
RSV infection in epithelial cells [16]. In animal models, the early
antioxidant response to viral infection is mediated by Nrf2, which
controls the expression of several antioxidant genes, including
HO1. For this reason, we decided to analyze the expression of this
protein in our model of infection. Differentiated NHBECs were
infected with RSV in the absence or presence of 1 or 10 mM
NAC. Nrf2 expression levels were measured by real-time RT-PCR.
The results obtained indicated that after RSV infection, there was
a rapid induction of this gene by day 4 postinfection followed by a
decrease in mRNA levels to below baseline (0.25-fold compared to
control cells at day 15 postinfection; Fig. 4A). This effect was
attenuated by 1 mM NAC, but in cultures pretreated with 10 mM
NAC, the induction of Nrf2 was maintained until the end of the
experiment. This profile was completely opposite to that found for
ICAM1 expression, as is presented in Fig. 4B. As the levels of Nrf2
decreased, the expression levels of ICAM1 increased.
One of the most important known antioxidant genes is HO1.
Data shown here indicated that the relative expression of this
protein was significantly inhibited in RSV-infected cultures at day
15 postinfection (0.15-fold compared to mock-infected cultures;
Fig. 4C) and that NAC pretreatment not only restored the levels of
HO1, but it also induced the its expression to much higher levels
than those observed in control cells (9.56 and 3.29-fold with 10
and 1 mM NAC, respectively; Fig. 4C).
TAS determines the response capacity of a biological system to
oxidative-mediated events. This status represents the balance
between oxidant and antioxidant molecules and can be measured
using different approaches. We next decided to analyze whether
an alteration of this balance occurred as a consequence of RSV
infection of NHBEC cultures. Differentiated NHBECs were
infected with RSV in the absence or presence of 1 or 10 mM
NAC. Fifteen days later, the antioxidant status of cultures was
determined using a TAS antioxidant assay kit. The results
obtained are summarized in Fig. 4D and indicate that after virus
infection, a decrease occurred in the antioxidant status of the
cultures (21% of that of control), which was completely reversed by
treatment with 1 mM NAC. Pretreatment with 10 mM NAC
induced an increase in TAS of 141% compared to mock-infected
cells.
Finally we decide to explore the effects of NAC on intracellular
H2O2 and glutathione content. Result obtained are shown in
figure 4 (panel E and F respectively). After RSV infection an
Figure 2. Effect of NAC on ciliary activity and ciliagenesis. Twenty-one days after ALI differentiation, NHBECs were infected with RSV at 5 PFUper cell. Every 48 h after infection, three different videos were recorded using a high-speed video camera. (A) Beating cells were counted. Results arepresented as the mean 6 SEM of three independent infections. (B) Fifteen days postinfection (p.i.), the effects of NAC were evaluated in mock-infected cultures (white bars) and RSV-infected cells in the absence (black bars) or presence (squared bars) of 0.1, 1 and 10 mM NAC. (C)Ultrastructural alterations in differentiated NHBEC cultures were evaluated by transmission electron microscopy in control (mock-infected cells, left)and RSV-infected (center) at day 15 p.i. Ultrastructures of cilia (right) were also evaluated. Arrows indicate tubular systems. (D) FOXJ1 and (E) DNAI2expression was analyzed by real-time RT-PCR at day 15 p.i. (F) Cultures were also evaluated for b-tubulin expression by immunofluorescence incontrol (mock-infected cells, left) and RSV-infected cultures in the absence (center) or presence of 10 mM NAC (right). Experiments were performed intriplicate and six independent infections were used (p = 6, n = 18). Data are presented as the mean 6 SEM. *p,0.05, compared to mock-infected cells;#p,0.05, compared to infected cells.doi:10.1371/journal.pone.0048037.g002
NAC Inhibits RSV Infection in NHBE Cells
PLOS ONE | www.plosone.org 4 October 2012 | Volume 7 | Issue 10 | e48037
increase of intracellular levels of H2O2 (5.97-fod compared to
control cultures) and a decrease of total glutathione levels (25.36%
of control) were observed. NAC 10 mM completely restored
intracellular H2O2 to control levels and increased glutathione
levels above that of mock-infected cultures.
Discussion
In this study, we used an in vitro model of RSV infection carried
out in differentiated cultures of NHBECs to analyze the effects of
NAC on the alterations induced by RSV on ciliagenesis and mucin
production, the two factors determining mucus clearance. We
analyzed the frequency and number of beating cells, the
ultrastructural alterations, the expression of two important genes
involved in ciliagenesis, and the expression of one of the
components of cilia, the b-tubulin protein. In addition, we
explored MUC5AC, GOB5, and IL-13 as indicators of metapla-
sia. Finally, we evaluated the effects of NAC for the modulating
oxidative-dependent alterations induced after RSV infection,
including Nrf2 and HO1 expression, TAS modulation, intracellular
H2O2 levels and intracellular glutathione content.
COPD is the fourth leading cause of death worldwide,
accounting for more than 2.5 million deaths every year [20].
Persistent RSV infections are one of the factors associated with
COPD exacerbations, which are characterized, among other
factors, by an increase in sputum volume [7,21]. Two principal
factors determine mucus clearance in airways: the ciliary beating
of ciliated epithelial cells and mucin release [8].
The benefits of NAC inhibiting RSV infection are well
characterized in A549 cells [22]. Recently, its capacity to inhibit
mucin release in RSV-infected A549 cells was reported [16], but
nothing is known about the direct effects of NAC treatment on
ciliary activity, ciliagenesis, or metaplasia of respiratory epithelia.
Differentiated epithelial cells are one of the accepted models for
the study of RSV infection [9]. In this system, RSV specifically
infects ciliated cells via the apical membrane [23]. Therefore, these
cells constitute a valid model for the study of mucus clearance.
Data presented here demonstrate that NAC significantly
inhibited RSV infection of differentiated epithelial cells. This
Figure 3. NAC inhibits MUC5AC, Gob5, and IL-13 upregulation and restores the normal structure of epithelium in RSV-infectedNHBEC cultures. Twenty-one days after ALI differentiation, NHBECs were infected with RSV at 5 PFU per cell. Total RNA was extracted and analyzedby real-time RT-PCR for (A) MUC5AC and (B) Gob5 expression in mock-infected cultures (white bars) and RSV-infected cells in the absence (black bars)or presence (squared bars) of 0.1, 1 and 10 mM NAC. (C) Histological properties of cultures were evaluated by PAS staining in control (mock-infectedcells, upper panel) and infected cultures in the absence (middle panel) or presence (lower panel) of 10 mM NAC. (D) IL-13 expression and release wereevaluated by real-time RT-PCR (left, white bars) and Luminex (right, black bars), respectively, in mock-infected cultures and RSV-infected cells in theabsence or presence of 0.1, 1 and 10 mM NAC. Protein release was evaluated in culture supernatants. Experiments were performed in triplicate and 6independent infections were used (p = 6, n = 18). Data are presented as the mean 6 SEM. *p,0.05, compared to mock-infected cells; #p,0.05,compared to infected cells.doi:10.1371/journal.pone.0048037.g003
NAC Inhibits RSV Infection in NHBE Cells
PLOS ONE | www.plosone.org 5 October 2012 | Volume 7 | Issue 10 | e48037
effect may be mediated, at least in part, by ICAM1 and reactive
oxygen species (ROS) generation, as discussed later. To our
knowledge, this is the first report demonstrating this effect of NAC.
In preliminary experiments the number of beating cells and the
determination of viral infection was monitored for 21 days. We
found that the decrease in the number of beating cells becomes
maximal at day 15 postinfection. This was in parallel with the
number of infected cells which reach maximal at day 15
postinfection and remain stable until day 21 (data not shown).
This is the reason because we have selected this time point in this
study. We have used 5 pfu/cell because with this concentration we
obtained maximal infection (data not shown) and is in line with
other authors [2,4]. After RSV infection of differentiated cultures,
a rapid decrease occurred in the number of beating cells, which
Figure 4. Antioxidant effects of NAC. Twenty-one days after ALI differentiation, NHBEC cultures were infected with RSV at 5 PFU per cell in theabsence or presence of 1–10 mM NAC. Total RNA was extracted and analyzed by real-time RT-PCR for (A) Nrf2 expression, (B) ICAM1 expression atdays 2, 4, 10, and 15 postinfection (p.i.), and (C) HO1 expression at day 15 p.i. (D) Total antioxidant status, (E) intracellular H2O2 levels and (F)intracellular glutathione content were analyzed at day 15 p.i. Experiments were performed in triplicate and six independent infections were used(p = 6, n = 18). Data are presented as the mean 6 SEM. Experiments were done in triplicate. *p,0.05, compared to mock-infected cells; #p,0.05,compared to infected cells. The English in this document has been checked by at least two professional editors, both native speakers of English. For acertificate, please see: http://www.textcheck.com/certificate/QCbgL4.doi:10.1371/journal.pone.0048037.g004
NAC Inhibits RSV Infection in NHBE Cells
PLOS ONE | www.plosone.org 6 October 2012 | Volume 7 | Issue 10 | e48037

was accompanied by ultrastructural alterations of basal axonemal
bodies and a decrease in the number of cilia and b-tubulin-positive
cells. These alterations are consistent with those described by
others [24]. We also analyzed the frequency of beating cells. Other
investigators have reported a rapid decrease in ciliary beat
frequency after RSV infection, such that only 2 h after infection,
the ciliary beat frequency decreased to 0 Hz in infected cells [24].
We found no differences in the frequency of beating cells in RSV-
infected cultures, probably because our system only allowed us to
measure the frequency of cells that were beating, and these cells
likely had not yet been infected by the virus. In this study, we also
analyzed the expression of two important genes related to
ciliagenesis, FOXJ1 and DNAI2. FOXJ1 is a member of the
forkhead/winged-helix family of transcription factors that regu-
lates the expression of several genes implicated in motile cilia
structure, including those encoding axonemal dyneins [25],
calpastatin [26], and components of the central complex like
WDR16 or centrin2 [27]. DNAI2 is implicated to be involved in
the outer dynein arm assembly [28]. The gene expression results of
this study correlated with those observed for the number of ciliated
cells; they showed that the expression levels of both genes may be
indicative of the number of ciliated cells in the airway epithelium
and may be considered as markers of metaplasia in airways. NAC
pretreatment of cultures inhibited, in a dose-dependent fashion, all
of these changes, restoring the normal ciliary activity of cultures.
The viscosity of mucus is determined by different factors,
including mucin release, of which MUC5AC is the most
prominent, in human airways [12]. MUC5AC expression is
controlled by the Ca2+-dependent chloride channel, GOB5 [29].
We observed a significant upregulation of the expression of both
genes in our model, which was inhibited in a dose-dependent
manner by NAC. Histological studies indicated that this increase
in MUC5AC expression was accompanied by changes in the
thickness and density of goblet cells, which were restored by NAC
pretreatment of cultures. These observations were in accordance
with the induction of IL-13 observed after RSV infection, which
has been reported as critical for epithelium metaplasia and with
the cilia loss in human airway epithelium [30,26]. Animal models
studies demonstrate that the induction if IL-13 release is one of the
key regulators of the inflammatory response against RSV infection
[31,32,33] which is in line with data presented here. Because IL-13
expression by airway epithelial cells is controversial, we have used
two different approaches to measure it. On the one hand we have
study the expression of IL-13 gene using Real-Time RT-PCR, and
on the other hand we have measured the protein levels in culture
supernatants using a R&D designed luminex assay. Both systems
have been used to study the expression of this cytokine in epithelial
cells [34,35,36].
All the effects of NAC can be explained by its capacity to inhibit
viral infection, as supported by the results presented here. The cell
surface adhesion molecule ICAM1 is essential for RSV infection of
epithelial cells. This protein binds to F glycoprotein and facilitates
virus entry into the host cell [37]. Increases in ICAM expression
after RSV infection of bronchial epithelial cells has been
previously reported and determines the infectivity of RSV
[38,39]. The expression of ICAM1 is controlled by ROS-mediated
events [40]. Our findings are in line with these observations, as
after RSV infection we detected a strong induction of ICAM1
expression in differentiated NHBEC cultures, which was inhibited
by NAC. The modulation of oxidative stress observed here can
explain the inhibition of ICAM1 overexpression and the
subsequent inhibition of viral infection of differentiated NHBECs.
One known factor involved in the control of antioxidant genes,
like HO-1 or glutathione peroxidase (GPx), is the nuclear factor
Nrf2 [41]. In agreement with observations made in animal models,
we observed a rapid increase in Nrf2 expression followed by a
decrease to levels lower than those observed in non-infected cells
[42]. Probably this pattern indicate the physiological early
response of cultures to virus infection and was dramatically
changed by NAC, which induced HO-1 expression and restored
the antioxidant status of cells. Cho et al. analyzed the upregulation
of HO-1 gene in an animal model of RSV infection and found, in
parallel to Nrf2 pattern, a rapid increase in the expression of this
antioxidant protein, followed by a decrease until levels lower than
those found in the control animals [42]. We have analyzed the
expression of HO-1 at day 15 postinfection and our results
indicate, in concordance to these observed by Cho et al. lower
levels of expression of this gene compared to mock-infected cells.
NAC induces a significantly increase in the levels of HO-1, which
can explain the effects observed at the total antioxidant status and
in the intracellular H2O2 levels of treated cultures compared to
untreated. NAC is a known precursor of glutathione, the most
important intracellular antioxidant molecule [43,44]. Data sup-
ported here indicate that after RSV infection there is a
significantly decrease of glutathione levels which are increased in
NAC treated cultures. This increase explain the restore of
intracellular H2O2 levels and the inhibition of ICAM expression
with subsequent reduction of RSV capacity to infect cultures cells.
In summary, we conclude that NAC inhibited RSV infection in
differentiated NHBEC cultures, restoring normal ciliary activity
and inhibiting mucin release. These results support the beneficial
effects of NAC treatment for the management of respiratory
diseases, including COPD, in which 64% of exacerbations are
associated with respiratory virus infection.
Materials and Methods
Cell model and experimental groupsHuman lung tissue was obtained from patients who had
undergone surgery for lung carcinoma, as previously outlined [45].
Experiments were approved by the local ethics committee and
informed consent was obtained. At the time of operation, lung
function was within normal limits by spirometry. None of the
patients were being chronically treated with theophylline, b-
adrenoceptor agonists, corticosteroids, or anticholinergic drugs.
Bronchia were carefully dissected free from adjoining connective
tissue and lung parenchyma. Human bronchial epithelial cells
were cultured and differentiated in 24-well Transwell inserts
(0.3 cm2; Corning Costar, High Wycombe, UK) under ALI
conditions, as previously described [46]. In brief, a multilayered
bronchial epithelium was obtained by seeding cells (8.256104 cell
per insert) onto polyester inserts (Millipore, Billerica, MA). Cells
were submerged in differentiation medium [50% Dulbecco’s
modified Eagle’s medium (DMEM) in basal epithelial growth
media (BEGM); Clonetics, Wokingham, UK] for the first 7 days.
Cells were then cultured for an additional 21 days with the apical
surface exposed to air, and ciliary activity was inspected every day
until the maximal density of ciliated cells (200–250 ciliated cells
per field) was reached. Cultures were then washed three times with
fresh differentiation culture medium and infected with 100 mL of
the same media containing 26106 plaque forming units (PFU)
RSV per insert. Cultures were incubated for 2 h at 37uC and
washed once with 500 mL differentiation media. In this study, we
used cultures from six different donors. All experiments were done
by triplicates (p = 6, n = 18).
Infected cultures were maintained until day 15 postinfection.
Culture medium was replaced every 48 h. Cilia activity, culture
supernatants, and cells were collected at the indicated time points.
NAC Inhibits RSV Infection in NHBE Cells
PLOS ONE | www.plosone.org 7 October 2012 | Volume 7 | Issue 10 | e48037

In this study, the following experimental groups were included:
control (untreated and mock-infected cells), infected (cells infected
with RSV), infected and treated (cells infected with RSV in the
presence of 0.1, 1 and 10 mM NAC; Sigma-Aldrich, St. Louis,
MO), and only treated (mock-infected cells treated with 0.1, 1 and
10 mM NAC). NAC was dissolved in distilled water as indicated
by the manufacturer, added to the culture medium 1 h before
infection and maintained until the end of the experiment. Cell
toxicity of the three different concentrations of NAC used in this
study was analyzed using the presto blue cell viability reagent
(Invitrogen Ltd., Paisley, UK) following manufacturer’s instruc-
tions. No effect of any of the concentrations tested was observed.
Preparation of the virusRSV (long Strain, ATCC VR-26) obtained from the American
Type Culture Collection (ATCC, Rockville, MD) was propagated
in Hep-2 cells in DMEM (Invitrogen Ltd., Paisley, UK) with 2%
fetal calf serum (DMEM-2; Invitrogen Ltd.), as previously
described [47]. Viruses were purified from clarified culture
supernatants by polyethylene glycol precipitation and centrifuga-
tion in a 30–45–60% discontinuous sucrose gradient in TNE
buffer [48,49]. Virus titters were determined by plaque assay in
Hep-2 cells layered with 0.5% low melting- point agarose (Conda
Laboratories, Madrid, Spain). After 5 days, cells were fixed with
4% formaldehyde (PanreacQuimica, Barcelona, Spain) in phos-
phate-buffered saline (PBS), followed by methanol, and incubated
with a mixture of monoclonal antibodies against the two major
glycoproteins of the virus (2F, 47F, 56F, 021/1G, 021/2G; Sigma-
Aldrich) [47,49]; plaques were visualized using an anti-mouse IgG
horseradish peroxidise-linked whole antibody (Amersham Phar-
macia Biotech Europe GmbH, Freiburg, Germany) and 3-amino-
9-ethylcarbazole (AEC; Sigma-Aldrich). Virus inactivation was
achieved by irradiation with ultraviolet light for 90 min and
confirmed by plaque assay.
Determination of viral infectionIn this study, we used two methods to determine viral infection
of differentiated NHBEC cultures. The first method was
immunocytochemical detection of viral glycoproteins (2F, 47F,
56F, 021/1G, 021/2G), as described above, and the second
method was semiquantitative real-time reverse transcription
polymerase chain reaction (RT-PCR) with primers designed to
amplify nucleoprotein RNA (forward primer, 59-CAT-
GATTCTCCTGATTGTGGGATGA-39; reverse primer, 59-
TCACGGCTGTAAGACCAGATCTAT-39; probe, 59-
CCCCTGCTGCCAATTT-39) [50]. RNA extraction, cDNA
synthesis, and real-time PCR were done as described below.
Ciliary beat and ultrastructure analysisCiliary motility activity and frequency were evaluated as
described previously [51]. Every 48 h after infection, five videos
of each insert were recorded using a high-speed video digital
camera. The number of cells with ciliary activity was measured by
counting under double-blind conditions to minimize experimental
errors due to the observer.
Fifteen days postinfection, cells were trypsinized and processed
for transmission electron microscopy, as outlined previously [51].
Briefly, cells were fixed in 0.03 M phosphate buffer containing
2.5% glutaraldehyde for 1 h. Then, the cells were rinsed in the
same buffer for 30 min and fixed in 1% osmium tetroxide for 1 h.
The most representative areas were selected and examined with a
transmission electron microscope.
Immunofluorescence analysis of b-tubulinFor b-tubulin immunofluorescence analysis, nitrocellulose
membranes (Amersham Pharmacia Biotech Europe GmbH) were
ethanol-fixed and incubated with a mouse monoclonal antibody
against b-tubulin (Sigma-Aldrich) and a secondary rhodamine
anti-mouse fluorescein isothiocyanate (FITC)-conjugated antibody
(Sigma-Aldrich). Images were acquired using a fluorescence
microscope. Five different images of each insert were analyzed.
Goblet cell analysisTo estimate the number of goblet cells in the differentiated
NHBEC cultures, periodic acid-Schiff (PAS) staining was done.
Nitrocellulose membranes were removed from the inserts, fixed,
and paraffin-embedded, as previously outlined [45]. Inserts were
cut in 5-mM sections and stained using the PAS staining system
(Sigma-Aldrich). Finally, sections were mounted with DPX
(Sigma-Aldrich) and analyzed by microscopy.
Determination of MUC5AC, ICAM1, Gob-5, IL-13, Nrf2,and HO1 mRNA expression
Mucin MUC5AC mRNA transcripts were measured by real-
time RT-PCR, as previously reported [45]. Total RNA was
isolated with TRIzol reagent (Invitrogen Ltd.) following the
manufacturer’s instructions. Integrity was measured with a 2100
Bioanalyzer (Agilent Technologies Inc., Santa Clara, CA). Only
extractions with an integrity ratio (28S/18S) near 2.0 were
considered. Two-hundred nanograms of total RNA were retro-
transcribed into cDNA using TaqMan RT reagents (N808-0234;
Applied Biosystems, Foster City, CA), as indicated by the
manufacturer. GAPDH was used as an endogenous control.
Primers and probes for both MUC5AC and GAPDH (Sigma-
Aldrich) were designed using Primer Express software, as
previously reported [45]. For the rest of the genes included in
this study, Assays-on-DemandTM (Applied Biosystems) was used.
The DDCt method was used to obtain semi-comparative data.
Determination of IL-13 protein in supernatantsAt the indicated time points, cultures were washed once with
100 mL sterile PBS, and 65 mL fresh culture medium was added to
the surface. After 1 h of incubation, supernatants were collected
and analyzed for IL-13 release using a multiplex cytometer-based
method (Luminex DX100; Luminex Corp., Austin, TX). Panels
and specific bead-conjugated antibodies were purchased from
Millipore, and calculations were carried out following the
manufacturer’s instructions. Ultrasensitive panels were used for
these experiments.
Determination of Total Antioxidant Status (TAS)TAS was determined using the Total Antioxidant Assay kit
(Cayman Chemical Company, Ann Arbor, MI) following the
manufacturer’s instructions. This assay relies on the ability of
antioxidants in the sample to inhibit the oxidation of ABTS (2,29-
azino-di-[3-ethylbenzthiazoline sulfonate]) to ABTS+ by metmyo-
globin. The capacity of the antioxidants in the sample to prevent
ABTS oxidation is compared with that of Trolox, a water-soluble
tocopherol analog. Results are expressed as molar Trolox
equivalents.
Determination of intracelular levels of H2O2
Intracellular levels of H2O2 were determined using the Amplex
Red Reagent as previously described [16]. In briefly, cultures were
lysed in 100 mM Amplex red solution supplemented with 2 U/ml
HRP and 200 mU/ml superoxide dismutase (OXIS International
NAC Inhibits RSV Infection in NHBE Cells
PLOS ONE | www.plosone.org 8 October 2012 | Volume 7 | Issue 10 | e48037

Inc., Beverly Hills, CA, US) and incubated in the dark for 30 min.
Fluorescence was measured in a plate reader at excitation/
emission wavelengths lEX = 540 nm/lEM = 590 nm, respectively.
Determination of intracellular glutathione contentCultured cells were lysed in chilled 1% SDS lysis buffer. Total
glutathione levels were determined by measuring the conversion of
559-dithiobis(2-nitrobenzoic acid) in the presence of GSH to 2-
nitro-5-thiobenzoic acid, using the total glutathione quantification
kit (Dojindo, Rockville, MD, USA). Sample absorbance was read
at 415 nm and data were normalized to total protein, as
determined by a standard protein concentration assay (Biorad,
Hercules, CA, USA).
Analysis of resultsData are presented as the mean 6 SEM. Statistical analysis of
the results was carried out by analysis of variance (ANOVA)
followed by Tukey’s multiple comparison test or t-test, as
appropriate (GraphPad Software Inc., San Diego, CA). Signifi-
cance was accepted when p,0.05.
Author Contributions
Conceived and designed the experiments: MM JC JAM IM. Performed the
experiments: MM IS MA CC JAM IM. Analyzed the data: MM MA JC.
Contributed reagents/materials/analysis tools: JAM IM. Wrote the paper:
MM.
References
1. Collins PL, Chanock RM, Murphy BR (2001) Respiratory syncytial virus. In:
Knipe D, et al., editors. Virology, 4th ed., New York: Raven Press p. 1443–85.
2. Martınez I, Lombardıa L, Garcıa-Barreno B, Domınguez O, Melero JA (2007)
Distinct gene subsets are induced at different time points after human respiratory
syncytial virus infection of A549 cells. Gen Virol 88(Pt 2):570–81.
3. Krishnan S, Halonen M, Welliver RC (2004) Innate immune responses in
respiratory syncytial virus infections. Viral Immunol 17:220–33.
4. Martınez I, Lombardıa L, Herranza C, Garcıa-Barreno B, Domınguez O, et al.
(2009) Cultures of Hep-2 cells persistently infected by human respiratory
syncytial virus differ in chemokine expression and resistance to apoptosis as
compared to lytic infections of the same cell type. Virology 388:31–41.
5. Di Rosa F, Barnaba V (1998) Persisting viruses and chronic inflammation:
understanding their relation to autoimmunity. Immunol Rev 164:17–27.
6. Wilkinson TM, Donaldson GC, Johnston SL, Openshaw PJ, Wedzicha JA
(2006) Respiratory syncytial virus, airway inflammation, and FEV1 decline in
patients with chronic obstructive pulmonary disease. Am J Respir Crit Care
Med 173:871–6.
7. Wedzycha JA (2004) Role of viruses in exacerbations of chronic obstructive
pulmonary disease. Proc Am Thorac Soc 1:115–20.
8. Malia P, Johnson SL (2006) How viral infections causes exacerbations of airway
diseases. Chest 130(4):1203–10.
9. Tristram DA, Hicks W Jr, Hard R (1998) Respiratory syncytial virus and human
bronchial epithelium. Arch Otolaryngol Head Neck Surg 124(7):777–83.
10. Avadhanula V, Rodriguez CA, Devincenzo JP, Wang Y, Webby RJ, et al. (2006)
Respiratory viruses augment the adhesion of bacterial pathogens to respiratory
epithelium in a viral species- and cell type-dependent manner. J Virol
80(4):1629–36.
11. Fishaut M, Schwartzman JD, McIntosh K, Mostow SR (1978) Behavior of
respiratory syncytial virus in piglet tracheal organ culture. J Infect Dis
138(5):644–9.
12. Takeyama K, Fahy JV, Nadel JA (2001) Relationship of epidermal growth factor
receptors to goblet cell production in human bronchi. Am J Respir Crit Care
Med 163(2):511–6.
13. Akaike TM, Ando T, Oda T, Doi S, Ijiri S, et al. (1990) Dependence on O2-
generation by xanthine oxidase of pathogenesis of influenza virus infection in
mice. J Clin Invest 85:739–45.
14. Jacoby DB, Choi AM (1994) Influenza virus infection induces differential
expression of antioxidant genes in human airway epithelial cells. Free Radic Biol
Med 6:821–4.
15. Kinnula VL, Adler KB, Ackley NJ, Crapo JD (1992) Release of reactive oxygen
species by guinea pig tracheal epithelial cells in vitro. Am J Physiol 262:L708–12.
16. Mata M, Morcillo E, Gimeno J, Cortijo J (2011) N-acetyl-L-cysteine (NAC)
inhibits mucin synthesis and pro-inflammatory mediators in alveolar type II
epithelial cells infected with influenza virus A and B and with respiratory
syncytial virus (RSV). Biochem Pharmacol 82(5):548–55.
17. Cotgreave IA (1997) N-Acetylcysteine: pharmacological considerations and
experimental and clinical applications. Adv Pharmacol 38:205–27.
18. Dekhuijzen PN, van Beurden WJ (2006) The role for N-acetylcysteine in the
management of COPD. Int J Chron Obstruct Pulmon Dis 1(2):99–106.
19. Geiler J, Michaelis M, Naczk P, Leutz A, Langer K, et al. (2010) N-acetyl-L-
cysteine (NAC) inhibits virus replication and expression of pro-inflammatory
molecules in A549 cells infected with highly pathogenic H5N1 influenza A virus.
Biochem Pharmacol 79(3):413–20.
20. Mathers CD, Loncar D (2006) Projections of global mortality and burden of
disease from 2002 to 2030. PLoS Med 3(11):e442.
21. Zeng R, Li C, Li N, Wei L, Cui Y (2011) The role of cytokines and chemokines
in severe respiratory syncytial virus infection and subsequent asthma. Cytokine
53(1):1–7.
22. Carpenter LR, Moy JN, Roebuck KA (2002) Respiratory syncytial virus and
TNF alpha induction of chemokine gene expression involves differential
activation of Rel A and NF-kappa B1. BMC Infect Dis 2:5.
23. Moore ML, Peebles RS, Jr (2006) Respiratory syncytial virus disease
mechanisms implicated by human, animal model, and in vitro data facilitatevaccine strategies and new therapeutics. Pharmacol Ther 112(2):405–24.
24. Philippou S, Otto P, Reinhold P, Elschner M, Streckert HJ (2000) Respiratorysyncytial virus-induced chronic bronchiolitis in experimentally infected calves.
Virchows Arch 436(6):617–21.
25. Thomas J, Morle L, Soulavie F, Laurencon A, Sagnol S, et al. (2010)
Transcriptional control of genes involved in ciliogenesis: a first step in makingcilia. Biol Cell 102(9):499–513.
26. Gomperts BN, Gong-Cooper X, Hackett BP (2004) Foxj1 regulates basal bodyanchoring to the cytoskeleton of ciliated pulmonary epithelial cells. J Cell Sci
117(Pt 8):1329–37.
27. Hirschner W, Pogoda HM, Kramer C, Thiess U, Hamprecht B, et al. (2007)
Biosynthesis of Wdr16, a marker protein for kinocilia-bearing cells, starts at the
time of kinocilia formation in rat, and wdr16 gene knockdown causeshydrocephalus in zebrafish. J Neurochem 101(1):274–88.
28. Loges NT, Olbrich H, Fenske L, Mussaffi H, Horvath J, et al. (2008) DNAI2mutations cause primary ciliary dyskinesia with defects in the outer dynein arm.
Am J Hum Genet 83(5):547–58.
29. Nakanishi A, Morita S, Iwashita H, Sagiya Y, Ashida Y, et al. (2001) Role of
gob-5 in mucus overproduction and airway hyperresponsiveness in asthma. ProcNatl Acad Sci USA 98(9):5175–80.
30. Okada SF, Zhang L, Kreda SM, Abdullah LH, Davis CW, et al. (2011) Couplednucleotide and mucin hypersecretion from goblet cell metaplastic human airway
epithelium. Am J Respir Crit Mol Biol 45(2):253–60.
31. Johnson TR, Parker RA, Johnson JE, Graham BS (2003) IL-13 is sufficient for
respiratory syncytial virus G glycoprotein-induced eosinophilia after respiratory
syncytial virus challenge. J Immunol 170(4):2037–45.
32. Lukacs NW, Moore ML, Rudd BD, Berlin AA, Collins RD, et al. (2006)
Differential immune responses and pulmonary pathophysiology are induced bytwo different strains of respiratory syncytial virus. Am J Pathol 169(3):977–86.
33. Johnson TR, Graham BS (1999) Secreted respiratory syncytial virus Gglycoprotein induces interleukin-5 (IL-5), IL-13, and eosinophilia by an IL-4-
independent mechanism. J Virol Oct;73(10):8485–95.
34. Kinane DF, Shiba H, Stathopoulou PG, Zhao H, Lappin DF, et al. (2006)
Gingival epithelial cells heterozygous for Toll-like receptor 4 polymorphismsAsp299Gly and Thr399ile are hypo-responsive to Porphyromonas gingivalis.
Genes Immun 7(3):190–200.
35. Gangl K, Reininger R, Bernhard D, Campana R, Pree I, et al. (2009) Cigarette
smoke facilitates allergen penetration across respiratory epithelium. Allergy
64(3):398–405.
36. Allahverdian S, Harada N, Singhera GK, Knight DA, Dorscheid DR (2008)
Secretion of IL-13 by airway epithelial cells enhances epithelial repair via HB-EGF. Am J Respir Cell Mol Biol 38(2):153–60.
37. Behera AK, Matsuse H, Kumer M, Kong X, Lockey RF, et al. (2001) Blockingintercellular adhesion molecule-1 on human epithelial cells decreases respiratory
syncytial virus infection. Biochem Biophys Res Commun 280:188–95.
38. Makogoba MW, Sanders ME, Ginther Luce GE, Gugel EA, Dustin ML, et al.
(1998) Functional evidence that intercellular adhesion molecule-1 (ICAM-1) is aligand for LFA-1-dependent adhesion in T cell-mediated cytotoxicity.
Eur J Immunol 18:637–40.
39. Vignola AM, Campbell AM, Chanez P, Bousquet J, Paul-Lacoste P, et al. (1993)
HLA-DR and ICAM-1 expression on bronchial epithelial cells in asthma and
chronic bronchitis. Am Rev Respir Dis 148:689–94.
40. Leverence JT, Medhora M, Konduri GG, Sampath V (2011) Lipopolysaccha-
ride-induced cytokine expression in alveolar epithelial cells: role of PKCf-mediated p47phox phosphorylation. Chem Biol Interact 189(1–2):72–81.
41. Stey C, Steurer J, Bachmann S, Medici TC, Tramer MR (2000) The effect oforal N-acetylcysteine in chronic bronchitis: a quantitative systematic review. Eur
Respir J 16(2):253–62.
42. Cho HY, Imani F, Miller-DeGraff L, Walters D, Melendi GA, et al. (2009)
Antiviral activity of Nrf2 in a murine model of respiratory syncytial virus disease.Am J Respir Crit Care Med 15;179(2):138–50.
NAC Inhibits RSV Infection in NHBE Cells
PLOS ONE | www.plosone.org 9 October 2012 | Volume 7 | Issue 10 | e48037

43. Felton VM, Borok Z and Willis BC (2009) N-acetylcysteine inhibits alveolar
epithelial-mesenchymal transition. Am J Physiol Lung Cell Mol Physiol 297:L805-L812.
44. Gillisen A and Nowak D (1998) Characterization of N-acetylcysteine and
ambroxol in anti-oxidant therapy. Respir Med 92: 609–623.45. Mata M, Sarria B, Buenestado A, Cortijo J, Cerda M, et al. (2005)
Phosphodiesterase 4 inhibition decreases MUC5AC expression induced byepidermal growth factor in human epithelial cells. Thorax 60(2):144–52.
46. Cortijo J, Milara J, Mata M, Donet E, Gavara N, et al (2010) Nickel induces
intracellular calcium mobilization and pathophysiological responses in humancultured airway epithelial cells. Chem Biol Interact 183:25–33.
47. Martınez I, Dopazo J, Melero JA (1997) Antigenic structure of the humanrespiratory syncytial virus G glycoprotein and relevance of hypermutation events
for the generation of antigenic variants. J Gen Virol 78(Pt 10):2419–29.
48. Mbiguino A, Menezes J (1991) Purification of human respiratory syncytial virus:
superiority of sucrose gradient over percoll, renografin, and metrizamide
gradients. J Virol Methods 31(2–3):161–70.
49. Garcıa-Barreno B, Palomo C, Penas C, Delgado T, Perez-Brena P, et al. (1989)
Marked differences in the antigenic structure of human respiratory syncytial
virus F and G glycoproteins. J Virol 63(2):925–32.
50. Herranz C, Melero JA, Martınez I (2011) Reduced innate immune response,
apoptosis, and virus release in cells cured of respiratory syncytial virus persistent
infection. Virology 410(1):56–63.
51. Armengot M, Milara J, Mata M, Carda C, Cortijo J (2010) Cilia motility and
structure in primary and secondary ciliary dyskinesia. Am J Rhinol Allergy
24(3):175–80.
NAC Inhibits RSV Infection in NHBE Cells
PLOS ONE | www.plosone.org 10 October 2012 | Volume 7 | Issue 10 | e48037
Top Related
Copyright © 2022 FDOKUMEN

