






Bahasa
Halaman
Hukum


www.elsevier.com/locate/gene
Gene 349 (2005
Re-analysis of a human hepatitis B virus (HBV) isolate from an
East African wild born Pan troglodytes schweinfurthii: Evidence for
interspecies recombination between HBV infecting
chimpanzee and human
Emmanuil N. Magiorkinis, Gkikas N. Magiorkinis,
Dimitrios N. Paraskevis, Angelos E. Hatzakis*
National Retrovirus Reference Center, Department of Hygiene and Epidemiology, University of Athens, School of Medicine,
Mikras Asias 75, GR-11527, Athens, Greece
Received 9 September 2004; received in revised form 22 November 2004; accepted 15 December 2004
Available online 10 March 2005
Received by G. Pesole
Abstract
According to current estimates, hepatitis B virus (HBV) has infected 2 billion people worldwide and among them, 360 million suffer from
chronic HBV infection. Except humans, HBV or HBV-like viruses have also been isolated from different species of apes and mammals.
Although recombination has been described to occur extensively between different genotypes within the human HBV lineage, no
recombination event has ever been reported between human and non-human primate HBV sequences. It was our objective to perform an
exhaustive search for recombination between human and non-human primate HBV strains among all available full-length human and non-
human primate HBV sequences, using bootscanning and phylogenetic analyses. Intriguingly, we found that an HBV sequence isolated from a
wild born Pan troglodytes schweinfurthii in East Africa–FG–is a recombinant consisting of HBV infecting chimpanzee (ChHBV) and human
genotype C. More specifically, in a fragment of approximately 500 nt (positions 551–1050 spanning half of the RT domain of pol, which
overlaps with half of the coding region of the small surface protein), FG grouped with HBV genotype C, while in the rest of the genome it
grouped with ChHBV sequences. Phylogenetic analyses showed that in the latter region FG was more closely related to the Pan troglodytes
troglodytes subspecies, forming an outlier to this group. Moreover, we show evidence that the recombination event occurred after the initial
dispersion of HBV genotype C in humans. Finally, our findings point out that although rare recombination between HBV viruses infecting
different species occurs.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Hepadnaviruses; Mosaic; Primates; Phylogenetic analysis
0378-1119/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.gene.2004.12.021
Abbreviations: ChHBV, chimpanzee hepatitis B virus; G, gamma;
GiHBV, gibbons hepatitis B virus; GTR, general time reversible model;
HBV, hepatitis B virus; HBV-DNA, hepatitis B virus DNA; HBsAg,
hepatitis B virus surface antigen; MC, Monte Carlo; ML, maximum
likelihood; OuHV, orang-utan hepatitis virus; pol, polymerase gene; RT,
reverse transcriptase; SHBs, small hepatitis B virus surface protein;
WMHBV, woolly monkey hepatitis B virus.
* Corresponding author. Tel.: +30 210 7462090; fax: +30 210 7462190.
E-mail address: [email protected] (A.E. Hatzakis).
1. Introduction
Hepatitis B virus (HBV) is the prototype member of a
family of viruses designated as Hepadnaviridae. This
family consists of enveloped DNA viruses with a partially
double-stranded genome of approximately 3182–3221 nt in
length. HBV sequences isolated from humans have been
classified into eight genotypes A–H (Norder et al., 1994;
Stuyver et al., 2000; Okamoto et al., 1988; Arauz-Ruiz et
al., 2002). The distribution of human HBV genotypes
) 165–171

E.N. Magiorkinis et al. / Gene 349 (2005) 165–171166
differs greatly with the geographic origin: genotypes A, D
and G circulate globally (Norder et al., 1993a; Stuyver et al.,
2000), B and C prevail in East and South East Asia
(Okamoto et al., 1988), genotype E predominates in West
Africa (Norder et al., 1994), while genotypes F and H
circulate among native Amerindians in Central and South
America (Norder et al., 1993b; Arauz-Ruiz et al., 2002).
HBV viruses have been also isolated from different
primate species, such as chimpanzees (Vaudin et al., 1988;
Hu et al., 2000; MacDonald et al., 2000; Takahashi et al.,
2000), lowland gorillas (Grethe et al., 2000), gibbons (Norder
et al., 1996; Grethe et al., 2000), orang-utans (Warren et al.,
1999; Verschoor et al., 2001), as well as from woolly
monkeys (Lanford et al., 1998). Chimpanzee subspecies
inhabit in West, West-Central and East Africa; gibbons and
orang-utans in East and South Eastern Asia, while woolly
monkeys inhabit in South America. The last group (woolly
monkey hepatitis B virus orWMHBV) is themost genetically
divergent among primate hepadnaviruses providing, thus, an
outlier to the other lineages. According to previous studies,
the prevalence of active HBV-infection (detection of both
HBV-DNA and HBsAg) is 10.5% in chimpanzees, 39% in
lowland gorillas, 26% in gibbons and 20% in orang-utans,
while the respective combined rates for woolly monkeys is
30% (Starkman et al., 2003). These findings are in
accordance with another study reporting a high prevalence
of active or past HBV infection in chimpanzees and gorillas
(32% and 30%, respectively) and the absence of HBV
infection in old world monkeys (Makuwa et al., 2003).
Until now, recombination has been documented in
several cases among HBV genotypes isolated from humans,
as for example between genotypes A and D, A and C, B and
C or between D and C (Bollyky et al., 1996; Morozov et al.,
2000; Owiredu et al., 2001; Hannoun et al., 2000; Cui et al.,
2002). Interestingly, HBV genotypes E and G are also
shown to be recombinant consisting partially of D and A,
respectively (Bowyer and Sim, 2000; Fares and Holmes,
2002). On the other hand, although several cross-species
transmission events have been reported which occurred
between different primate species, such as between gibbons
and chimpanzees, chimpanzees and gorillas (Norder et al.,
1996; Grethe et al., 2000) and between chimpanzees and
humans (Takahashi et al., 2000; Hu et al., 2000), no
recombination events have ever been documented between
human and non-human primate HBV sequences.
It was our objective to investigate all available full-length
HBV sequences for evidence of recombination between
human and non-human primate HBV sequences.
2. Materials and methods
2.1. Materials
Full-length HBV sequences (550 sequences) were down-
loaded from the GenBank database (http://www.ncbi.nih.
gov). The following sequences were used as reference panel
using the currently available HBV genotype nomenclature:
(a) human sequences: genotypes A (U87742, M57663), B
(AB010289, D00329), C (AY123041, AF068756), D
(AF151735, X02496), E (X75657, AB032431), F
(AY090455, X69798), G (AB064313, AF160501) and H
(AY090454, AB059659); (b) non-human primate sequen-
ces: HBV infecting chimpanzees (ChHBV) (D00220,
AF242586), gibbons (GiHBV) (AJ131568, AY077736),
orang-utans (OuHV) (AF193863, AF193864) and woolly
monkeys (WMHBV) (AY226578, AF046996). Additional
HBV sequences for human genotype C and ChHBV used
for phylogenetic were: genotype C (Y18858, X04615,
AF223960, AY306136), ChHBV from Pan troglodytes
troglodytes (AF242585, AF222322, AY330911) and Pan
troglodytes vellerosus (AY330912, AF305327). Both strains
D00220 and AF242586 have been isolated from Pan
troglodytes verus.
2.2. Methods
Full genomic sequences were aligned using the Clustal
W program version 1.81 (Higgins and Sharp, 1988;
Thompson et al., 1994). An exploratory bootscanning
analysis was performed using a sliding window of 400 nt
moving in steps of 50 nt and maximum likelihood
estimated distances (F84 model with transitions/trans-
versions=2, default value) as implemented in Simplot
(3.2 beta version) (http://sray.med.som.jhmi.edu/Raysoft/
Simplot). The F84 model was used since it is the most
complex model available in the current version of
Simplot. HBV genotypes E and G that have been referred
to be recombinant were included separately in the
bootscanning analysis of each of the rest 526 HBV
sequences (Bowyer and Sim, 2000; Fares and Holmes,
2002).
For a single HBV-sequence (isolate name FG, accession
number AF498266) for which according to bootscanning
plots there was evidence for recombination between human
and non-human primates, separate phylogenetic analyses
were performed for the putative non-recombinant fragments,
based on maximum likelihood using Tamura-Nei substitu-
tion model (Tamura and Nei, 1993) including a G-
distributed rates heterogeneity among sites, as implemented
in Treepuzzle (Schmidt et al., 2002). Phylogenetic trees
were also reconstructed using Bayesian inference with
the GTR model and a G-distributed rates heterogeneity
among sites as implemented in MrBayes (v 3.0)
(Huelsenbeck and Ronquist, 2001). Four Metropolis
Coupled Markov chains Monte Carlo (MC3) were run
for 106 generations with a burnin of 105 generations. An
intergenotype recombination breakpoint was defined as a
region of approximately 100–200 nt in length in which
the HBV genotype classification was different in the
adjacent pieces (supported by N0.9 posterior probability
by Bayesian inference of phylogeny and N75% ML

E.N. Magiorkinis et al. / Gene 349 (2005) 165–171 167
puzzling steps). To assess for any statistically significant
differences of alternative topologies in different frag-
ments, we used the Shimodaira test as implemented in
Treepuzzle.
3. Results
3.1. Results of the exploratory analysis: FG sequence is a
recombinant consisting of ChHBV and human HBV
genotype C
Exploratory bootscanning analysis, including 550 full-
length available HBV sequences isolated from humans and
primates, showed evidence that an HBV sequence isolated
from Pan troglodytes schweinfurthii (Vartanian et al.,
2002) (isolate name FG, accession number AF498266)
was potentially recombinant consisting of HBV sequences
from chimpanzees (ChHBV) and humans (HBV genotype
C) (Fig. 1). More specifically, sequence FG clustered
significantly within the ChHBV lineage in two different
regions spanning nucleotides 1–550 and 1051–3182. On the
other hand, the region between nucleotides 551–1050,
spanning half of the RT domain of pol (aa 139–306), which
overlaps with half of the coding region of the small surface
protein (SHBs) (aa 183–226), clustered significantly with
HBV genotype C (Fig. 1A). Phylogenetic analysis, using
ML and Bayesian methods (Fig. 1C), further confirmed the
previous findings about discordant phylogenetic clustering
in fragments 1–550, 1051–3182 and 551–1050. Moreover,
according to the Shimodaira test, there was evidence for
significant difference ( pb0.05) between candidate topolo-
gies in different pieces of the alignment (regions 1–550,
1051–3182 and 551–1050).
3.2. Analysis of the region clustering with ChHBV in
comparison with other chimpanzee subspecies
To examine in detail the phylogenetic relationship
between the FG sequence and the ChHBV subspecies,
we concatenated the genomic regions clustered with
ChHBV (1–550 and 1051–3182) and performed boot-
scanning and phylogenetic analyses using additional
ChHBV reference sequences isolated from different sub-
species (Pan troglodytes troglodytes, Pan troglodytes
vellerosus and Pan troglodyte verus). Bootscanning plot
showed that FG sequence was more closely related to the
ChHBV Pan troglodytes troglodytes group (genomic
regions 1051–16401 and 1951–3182) for the longest part
of the alignment, whereas in two short fragments (111–550
and 1641–1950) no significant clustering (bootstrap support
b75%) was found between FG and any of the other ChHBV
groups (data not shown). Phylogenetic analyses in the latter
1 Nucleotide positions correspond to the original FG sequence.
regions indicated that ChHBV sequences from the different
chimpanzee subspecies did not form monophyletic clusters,
probably due to limited phylogenetic signal in the align-
ment, thus suggesting that in these regions further classi-
fication of the HBV sequences from chimpanzees is not
possible (data not shown).
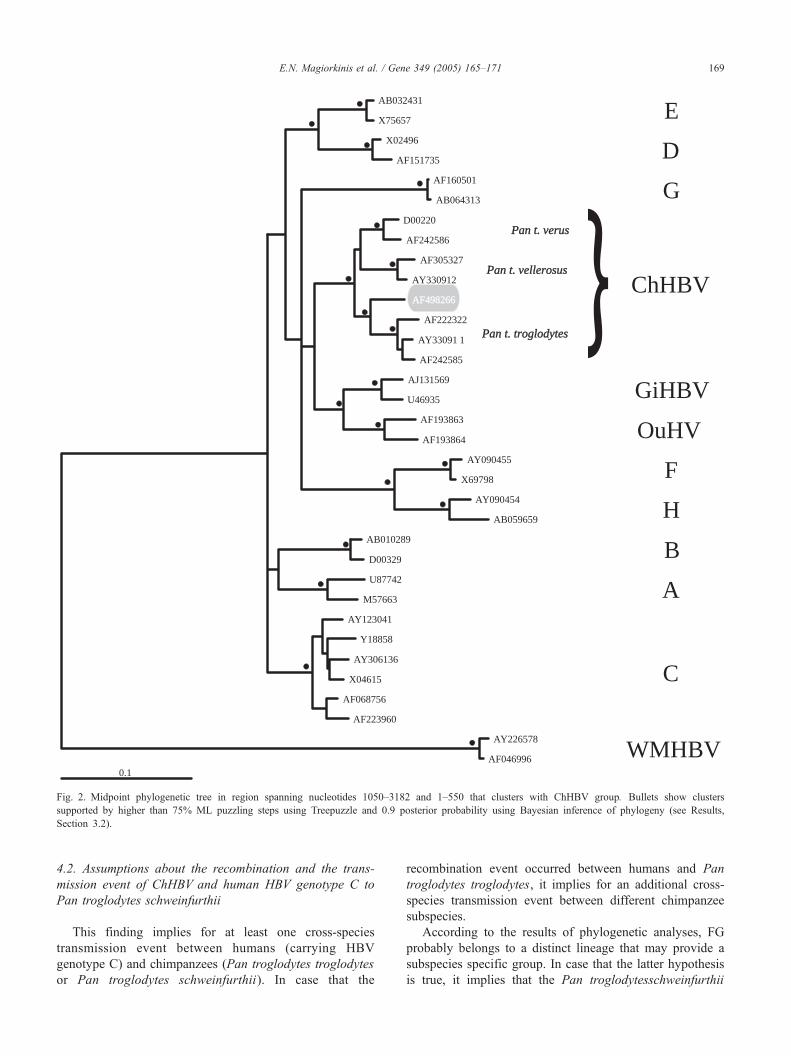
Phylogenetic analysis in the concatenated region (1–550
and 1051–3182) showed that FG clustered with the ChHBV
sequences from Pan troglodytes troglodytes subspecies,
albeit forming an outlier to the latter group (Fig. 2). The
evolutionary distances between the FG and the Pan
troglodytes troglodytes group were comparable to the
distances between different chimpanzee subspecies (e.g.
between Pan troglodytes verus and Pan troglodytes
vellerosus) (Fig. 2). These findings suggest that the partial
FG sequence belongs to a potentially new ChHBV
subgroup, which may provide a subspecies specific group
for Pan troglodytes schweinfurthii. However, given the
limited sequence information available for this subspecies, it
is not possible to identify the precise origin of the partial FG
sequence.
3.3. Analysis of the region clustering with human HBV
genotype C
In contrary to the phylogenetic clustering of FG sequence
within the ChHBV group, where it appeared to be an
outgroup to the Pan troglodytes troglodytes cluster, in
region (551–1050 nt) FG fell within the HBV genotype C
group (Fig. 1C), thus suggesting that the recombination
event occurred after the initial spread of human HBV
genotype C.
4. Discussion
4.1. Summary of analysis
Throughout an exhaustive search of all full-length
available HBV sequences, we found evidence for
recombination between human and non-human primate
HBV strains, in an HBV sequence isolated from a
chimpanzee which belongs to the Pan troglodytes
schweinfurthii subspecies. Further analysis showed that
the HBV isolate consisted of partial human HBV
genotype C and ChHBV, which was more closely related
to the Pan troglodytes troglodytes group. We should note
that the recombinant HBV sequence, FG, was isolated
from a chimpanzee living in the wild without co-culturing
the virus, in vitro (Vartanian et al., 2002), thus suggesting
that recombination could neither be due to laboratory
contamination nor to any human intervention. Although
the latter hypothesis cannot be entirely excluded, it
appears as a less plausible explanation given that the
HBV strain was isolated from a chimpanzee living in the
wild.

WMHBV
F
H
C
OuHV
GiHBV
B
D
ChHBV
G
A
E
WMHBV
F
H
C
OuHV
GiHBV
B
D
ChHBV
G
A
E1-550 551-1050 1051-3182
0
20
40
60
80
100
0 500 1000 1500 2000 2500 3000
WMHBV
F
H
C
OuHBV
GiHBV
B
D
ChHBV
G
A
E
C
A B
WMHBV
F
H
C
OuHV
GiHBV
B
D
ChHBV
G
A
E
Nucleotides
Boo
stra
p va
lues
0.1
X75657
AB032431
AF160501
AB064313
M57663
U87742
AAF498266F498266
AF242586
D00220
X02496
AF151735
D00329
AB010289
AY123041
AF068756
AY077736
HB131568
AF193864
AF193863
X69798
AY090455
AB059659
AY090454
AF046996
AY226578 0.1
X75657
AB032431
AF160501
AB064313
D00329
AB010289
M57663
U87742
AF242586
D00220
AY077736
HB131568
AF193864
AF193863
AF068756
AF498266AF498266
AY123041
X02496
AF151735
X69798
AY090455
AB059659
AY090454
AF046996
AY226578 0.1
X75657
AB032431
X02496
AF151735
D00329
AB010289
M57663
U87742
AY123041
AF068756
AF160501
AB064313
X69798
AY090455
AB059659
AY090454
AF498266AF498266
AF242586
D00220
AY077736
HB131568
AF193864
AF193863
AF046996
AY226578
Pres1 PreS2
S
PreC
P
C
X
Fig. 1. (A) Schematic representation of the HBV-genome showing regions with discordant phylogenetic clustering of the FG recombinant sequence. (B) Bootscanning plot of the FG sequence against representative
HBV sequences from humans and primates. (C) Midpoint phylogenetic trees. Bullets indicate clusters supported by higher than 75% ML puzzling steps using Treepuzzle and 0.9 posterior probability using
Bayesian inference of phylogeny (see Results, Section 3.1).
E.N.Magiorkin
iset
al./Gene349(2005)165–171
168
E
D
C
ChHBV
Pan t. verusPan t. verus
Pan t. vellerosusPan t. vellerosus
Pan t. troglodytesPan t. troglodytes
OuHV
GiHBV
F
H
B
G
WMHBV
A
0.1
AB032431
X75657
X02496
AF151735
AF160501
AB064313
D00220
AF242586
AF305327
AY330912
AF498266AF498266
AF222322
AY33091 1
AF242585
AJ131569
U46935
AF193863
AF193864
AY090455
X69798
AY090454
AB059659
AB010289
D00329
U87742
M57663
AY123041
Y18858
AY306136
X04615
AF068756
AF223960
AY226578
AF046996
Fig. 2. Midpoint phylogenetic tree in region spanning nucleotides 1050–3182 and 1–550 that clusters with ChHBV group. Bullets show clusters
supported by higher than 75% ML puzzling steps using Treepuzzle and 0.9 posterior probability using Bayesian inference of phylogeny (see Results,
Section 3.2).
E.N. Magiorkinis et al. / Gene 349 (2005) 165–171 169
4.2. Assumptions about the recombination and the trans-
mission event of ChHBV and human HBV genotype C to
Pan troglodytes schweinfurthii
This finding implies for at least one cross-species
transmission event between humans (carrying HBV
genotype C) and chimpanzees (Pan troglodytes troglodytes
or Pan troglodytes schweinfurthii). In case that the
recombination event occurred between humans and Pan
troglodytes troglodytes, it implies for an additional cross-
species transmission event between different chimpanzee
subspecies.
According to the results of phylogenetic analyses, FG
probably belongs to a distinct lineage that may provide a
subspecies specific group. In case that the latter hypothesis
is true, it implies that the Pan troglodytesschweinfurthii

E.N. Magiorkinis et al. / Gene 349 (2005) 165–171170
subspecies is also infected with ChHBV virus in the wild
probably introduced at a similar time frame as in the other
chimpanzee subspecies, whereas we should note that, given
the limited ChHBV sequence information available until
now, we cannot estimate the precise origin of the partial FG
sequence.
Interestingly, the recombination breakpoints in FG were
mapped within the overlapping region of the RT domain of
pol and the small surface protein and in the non-overlapping
region of pol. Recombination breakpoints have also been
reported similarly for other intergenotypic HBV recombi-
nants in humans (Owiredu et al., 2001; Fares and Holmes,
2002; Cui et al., 2002).
4.3. The HBV genotype distribution in Africa indicates that
recombination between ChHBV and human HBV genotype
C is an extremely rare event—lack of data concerning
genotypic diversity accounts for the limited knowledge of
recombination events
Recombination between human HBV genotype C and
ChHBV seems to be a rare event, since chimpanzees inhabit
in Africa, where only HBV genotypes A, D and E are
reported to circulate (Norder et al., 1993a, 1994). Interest-
ingly, Hu et al. (2000) recently reported the infection of a
wild-caught chimpanzee with HBV genotype C, while it
was not possible to clarify whether the infection occurred in
the wild, or if it was due to the injection with contaminated
human serum. Moreover, Kramvis et al. (2002) reported
similarly an HBV sequence belonging to genotype C
isolated from a patient in South Africa, who according to
the designation of the HBV sequence in the GenBank
database probably originated from Eastern Asia. We should
note here that only a small fraction of the full-length HBV
sequences available worldwide originated from Africa and,
moreover, that only a small number of ChHBV sequences is
available so far from the troglodytes troglodytes and Pan
troglodytes schweinfurthii chimpanzees subspecies. Thus,
we should expect more recombinants and especially
between ChHBV and HBV genotypes A, D and E
circulating in Africa, as soon as more HBV and ChHBV
sequences are available in the future.
Our findings, therefore, show evidence about the first
interspecies recombinant, for which in contrast to the HBV
recombinants reported frequently among humans (Owiredu
et al., 2001; Fares and Holmes, 2002; Cui et al., 2002),
parental viruses are not circulating in the same geographic
area, thus suggesting that it might have been generated by
occasional exposure of humans in ChHBV from either Pan
troglodytes troglodytes or Pan troglodytes schweinfurthii, or
most possibly vice versa (chimpanzees got transmitted from
humans with HBV genotype C). Interestingly, the partial FG
sequence fell within genotype C suggesting that the
recombination event between human HBV and ChHBV
occurred after the initial disperse of the genotype C in
humans.
4.4. Conclusions—future perspectives
In conclusion, the only currently available ChHBV
virus from Pan troglodytes schweinfurthii was found to be
recombinant consisting of HBV genotype C and ChHBV
sequences. Our results are in favour for a subspecies
specific ChHBV infection in Pan troglodytes schweinfur-
thii. However, this issue will be clarified when the
prevalence of ChHBV infection is estimated and when
additional virus sequences become available from this
chimpanzee subspecies.
Acknowledgements
This work was supported by the Hellenic Scientific
Society for the Study of AIDS and Sexually Transmitted
Diseases. DP was supported by the Hellenic Center for
Infectious Diseases Control (HCIDC) of the Ministry of
Health and Welfare. MM and GM were supported by the
Hellenic Scientific Society for the Study of AIDS and
Sexually Transmitted Diseases.
References
Arauz-Ruiz, P., Norder, H., Robertson, B.H., Magnius, L.O., 2002.
Genotype H: a new Amerindian genotype of hepatitis B virus revealed
in Central America. J. Gen. Virol. 83, 2059–2073.
Bollyky, P.L., Rambaut, A., Harvet, P.H., Holmes, E.C., 1996. Recombi-
nation between sequences of hepatitis B virus from different genotypes.
J. Mol. Evol. 42, 97–102.
Bowyer, S.M., Sim, J.G.M., 2000. Relationships within and between
genotypes of hepatitis B virus at points across the genome: footprints of
recombination in certain isolates. J. Gen. Virol. 81, 379–392.
Cui, C., et al., 2002. The dominant hepatitis B virus genotype identified in
Tibet is a C/D hybrid. J. Gen. Virol. 83, 2773–2777.
Fares, M.A., Holmes, E.C., 2002. A revised evolutionary history of
hepatitis B virus (HBV). J. Mol. Evol. 54, 807–814.
Grethe, S., Heckel, J.O., Rietschel, W., Hufert, F.T., 2000. Molecular
epidemiology of hepatitis B virus variants in nonhuman primates.
J. Virol. 74, 5377–5381.
Hannoun, C., Norder, H., Lindh, M., 2000. An aberrant genotype revealed
in recombinant hepatitis B virus strains from Vietnam. J. Gen. Virol. 81,
2267–2272.
Higgins, D.G., Sharp, P.M., 1988. CLUSTAL: a package for performing
multiple sequence alignment on a microcomputer. Gene 15, 237–244.
Hu, X., Margolis, H.S., Purcell, R.H., Ebert, J., Robertson, B.H., 2000.
Identification of hepatitis B virus indigenous to chimpanzees. Proc.
Natl. Acad. U. S. A. 97, 1661–1664.
Huelsenbeck, J.P., Ronquist, F., 2001. MR BAYES: Bayesian inference of
phylogenetic trees. Bioinformatics 17, 754–755.
Kramvis, A., Weitzmann, L., Owiredu, W.K.B.A., Kew, M.C., 2002.
Analysis of the complete genome of subgroup AV hepatitis B virus
isolates from South Africa. J. Gen. Virol. 83, 835–839.
Lanford, R.E., Chavez, D., Brasky, K.M., Burns III, R.B., Rico-Hesse, R.,
1998. Isolation of a hepadnavirus from the woolly monkey, a New
World primate. Proc. Natl. Acad. U. S. A. 95, 5757–5761.
MacDonald, D.M., Holmes, E.C., Lewis, J.C., Simmonds, P., 2000.
Detection of hepatitis B virus infection in wild-born chimpanzees
(Pan troglodytes verus): phylogenetic relationships with human and
other primate genotypes. J. Virol. 74, 4253–4257.

E.N. Magiorkinis et al. / Gene 349 (2005) 165–171 171
Makuwa, M., et al., 2003. Occurrence of hepatitis viruses in wild-born non-
human primates: a 3 year (1998–2001) epidemiological survey in
Gabon. J. Med. Primatol. 32, 307–314.
Morozov, V., Pisareva, M., Groudinin, M., 2000. Homologous recombi-
nation between different genotypes of hepatitis B virus. Gene 260,
55–65.
Norder, H., et al., 1993a. Genetic relatedness of hepatitis B viral strains of
diverse geographical origin and natural variations in the primary
structure of the surface antigen. J. Gen. Virol. 74, 1341–1348.
Norder, H., Courouce, A.M., Magnius, L.O., 1993b. Complete nucleotide
sequences of six hepatitis B viral genomes encoding the surface antigen
subtypes ayw4, adw4q-, and adrq- and their phylogenetic classifica-
tion. Arch. Virol., Suppl. 8, 189–199.
Norder, H., Courouce, A.-M., Magnius, L.O., 1994. Complete genomes,
phylogenetic relatedness, and structural proteins of six strains of the
hepatitis B virus, four of which represent two new genotypes. Virology
198, 489–503.
Norder, H., Ebert, J.W., Fields, H.A., Mushahwar, I.K., Magnius, L.O.,
1996. Complete sequencing of a gibbon hepatitis B virus genome
reveals a unique genotype distantly related to the chimpanzee hepatitis
B virus. Virology 218, 214–223.
Okamoto, H., et al., 1988. Typing hepatitis B virus by homology in
nucleotide sequence: comparison of surface antigen subtypes. J. Gen.
Virol. 69, 2575–2583.
Owiredu, W.K., Kramvis, A., Kew, M.C., 2001. Hepatitis B virus DNA in
serum of healthy black African adults positive for hepatitis B surface
antibody alone: possible association with recombination between
genotypes A and D. J. Med. Virol. 64, 441–454.
Schmidt, H.A., Strimmer, K., Vingron, M., von Haeseler, A., 2002. TREE-
PUZZLE: maximum likelihood phylogenetic analysis using quartets
and parallel computing. Bioinformatics 18, 502–504.
Starkman, S.E., MacDonald, D.M., Lewis, J.C.M., Holmes, E.C., Sim-
monds, P., 2003. Geographic and species association of hepatitis B virus
genotypes. Virology 314, 381–393.
Stuyver, L., et al., 2000. A new genotype of hepatitis B virus: complete
genome and phylogenetic relatedness. J. Gen. Virol. 81, 67–74.
Takahashi, K., Brotman, B., Usuda, S., Mishiro, S., Prince, A.M., 2000.
Full-genome sequence analyses of hepatitis B virus (HBV) strains
recovered from chimpanzees infected in the wild: implications for an
origin of HBV. Virology 267, 58–64.
Tamura, K., Nei, M., 1993. Estimation of the number of nucleotide
substitutions in the control region of mitochondrial DNA in humans and
chimpanzees. Mol. Biol. Evol. 10, 512–526.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. Clustal W: improving
the sensitivity of progressive multiple sequence alignment through
sequence weighting position-specific gap penalties and weight matrix
choice. Nucleic Acids Res. 22, 4673–4680.
Vartanian, J.P.M., et al., 2002. Identification of a hepatitis B virus genome
in wild chimpanzees (Pan troglodytes schweinfurthi) from East Africa
indicates a wide geographical dispersion among equatorial African
primates. J. Virol. 76, 11155–11158.
Vaudin, M., Wolstenholme, A.J., Tsiquaye, K.N., Zuckerman, A.J.,
Harrison, T.J., 1988. The complete nucleotide sequence of the genome
of a hepatitis B virus isolated from a naturally infected chimpanzee.
J. Gen. Virol. 69, 1383–1389.
Verschoor, E.J., Warren, K.S., Langenhuijzen, S., Swan, R.A., Heriyanto,
Heeney, J.L., 2001. Analysis of two genomic variants of orang-utan
hepadnavirus and their relationship to other primate hepatitis B-like
viruses. J. Gen. Virol. 82, 893–897.
Warren, K.S., Heeney, J.L., Swan, R.A., Heriyanto, Verschoor, E.J., 1999.
A new group of hepadnavirus naturally infecting orangutans (Pongo
pygmaeus). J. Virol. 73, 7860–7865.
Top Related

Copyright © 2022 FDOKUMEN

