







Bahasa
Halaman
Hukum


Phosphoproteomics Profiling Suggests a Role for Nuclear �ΙPKC in
Transcription Processes of Undifferentiated Murine Embryonic
Stem Cells
Helio Miranda Costa-Junior,‡ Nicole Milare Garavello,† Mariana Lemos Duarte,†
Denise Aparecida Berti,† Talita Glaser,† Alexander de Andrade,§ Carlos A. Labate,§
Andre Teixeira da Silva Ferreira,| Jonas Enrique Aguilar Perales,| Jose Xavier-Neto,‡
Jose Eduardo Krieger,‡ and Deborah Schechtman*,†
Instituto de Quımica, Departamento de Bioquımica, Universidade de Sao Paulo, Brazil, Instituto do Coracao,Sao Paulo, Brazil, Departamento de Genetica, ESALQ, Universidade de Sao Paulo, Brazil, and Instituto
Oswaldo Cruz, Rio de Janeiro, Brazil
Received April 20, 2010
Protein kinase C (PKC) plays a key role in embryonic stem cell (ESC) proliferation, self-renewal, anddifferentiation. However, the function of specific PKC isoenzymes have yet to be determined. Of thePKCs expressed in undifferentiated ESCs, �IPKC was the only isoenzyme abundantly expressed in thenuclei. To investigate the role of �ΙPKC in these cells, we employed a phosphoproteomics strategyand used two classical (cPKC) peptide modulators and one �IPKC-specific inhibitor peptide. We identified13 nuclear proteins that are direct or indirect �ΙPKC substrates in undifferentiated ESCs. These proteinsare known to be involved in regulating transcription, splicing, and chromatin remodeling duringproliferation and differentiation. Inhibiting �ΙPKC had no effect on DNA synthesis in undifferentiatedESCs. However, upon differentiation, many cells seized to express �ΙPKC and �ΙPKC was frequentlyfound in the cytoplasm. Taken together, our results suggest that �IPKC takes part in the processes thatmaintain ESCs in their undifferentiated state.
Keywords: Protein kinase C • Peptides • Phosphorylation • Proteomics • 2DE gels • Differentiation •Embryonic stem cells
Introduction
The search for signaling pathways that lead to self-renewal,perpetual proliferation, and differentiation of embryonic stemcells (ESCs) is currently a major research endeavor. Inhibitionof endogenous signaling by GSK3 and MAPK pathways hasbeen shown to maintain ESC in an undifferentiated state,1 whiledynamic phosphorylation events are involved in the early stepsof ESC differentiation into specific cell types.2,3 Therefore, abetter understanding of these key phosphorylation events mayhelp elucidate how signaling pathways control ESC self-renewal/differentiation.
Among different signaling systems, protein kinase C (PKCs)signaling has been shown to participate in ESC proliferation4-6
and differentiation.7,8 Low concentrations of general PKCactivators [1,2, dioctanoyl-sn-glycerol (DOG), and 12-O-tetra-decanoyl forbol 13-acetate (TPA)] induce murine ESC prolifera-tion in a phospholipase C-dependent manner,4 while the
general PKC-inhibitor, bisindolylmaleimide II (Bis II), inhibitsit. However, the exact roles of PKCs in cell signaling are stillunclear, partially because PKCs constitute a family of at least10 serine/threonine kinases for which isozyme-specific modu-lators have only become available in the past 10 years.9-11 ThePKC family of isozymes can be divided according to theiractivation requirements in (1) the classical PKCs (cPKCs) thatincludes PKCR, �I, �II, and γ, which require calcium anddiacylglycerol (DAG) for their activation; (2) the novel PKCsisozymes (nPKCs), PKCδ, ε, η, and θ, which are calcium-independent and DAG-dependent; (3) the atypical PKCs (aP-KCs), � and λ/ι, which are both calcium-independent and DAG-insensitive.12
Although there is little doubt that PKC signaling is relevantto ESC biology, very few PKC substrates have been describedin vivo and there is little information on the specific roles ofdifferent PKC isozymes in ESCs, particularly in undifferentiatedESCs. To shed light on this subject, we devised a systematicapproach to implicate specific PKCs in specific ESC functionsutilizing rationally designed modulators of specific PKC isozymes.These modulators have been shown to interfere with PKCisozyme function, subcellular localization, and protein/proteininteractions.13,14
A particularly useful class of PKC modulators is representedby peptides that interact with isozyme-specific receptors for
* Corresponding Author: Deborah Schechtman, Av. Prof. Lineu Prestes,748, Bloco 10, CEP: 05508-900, Cidade Universitaria, Sao Paulo, Brazil. Phone:(05511) 3091-3810 ext. 224. Fax: (05511) 3091-2186. E-mail: [email protected].
‡ Instituto do Coracao.† Instituto de Quımica, Departamento de Bioquımica, Universidade de
Sao Paulo.§ Departamento de Genetica, ESALQ, Universidade de Sao Paulo.| Instituto Oswaldo Cruz.
10.1021/pr100355k 2010 American Chemical Society Journal of Proteome Research 2010, 9, 6191–6206 6191Published on Web 10/11/2010

activated C kinase (RACK). RACKs are anchoring proteins thatlocalize specific PKC isozymes to distinct intracellular compart-ments. These adaptor proteins promote the interaction be-tween PKCs and their substrates, increasing the specificity ofsignaling cascades.10,15 Upon activation in response to specificstimuli, PKCs translocate to distinct subcellular locations andbind to their isozyme-specific receptor for activated C kinase(RACK).10 PKC isozyme specific peptide inhibitors competewith PKC for RACK interaction and peptide activators functionas agonists, enhancing the interaction between PKC and RACK,and consequently PKC function.9,11
Here, we performed a PKC expression profile that shows thatthe major nuclear PKC in undifferentiated ESCs is �ΙPKC. Togain insight on the function of �ΙPKC in undifferentiated ESCs,we identified direct and indirect �ΙPKC substrates using aphosphoproteomic approach in ESCs treated with �ΙV5-3, ahighly specific �ΙPKC peptide inhibitor. �ΙV5-3 inhibits PKC�Ιtranslocation and function by inhibiting �ΙPKC anchoring toits receptor for activated kinase (RACK),16 without influencingtranslocation and function of the highly similar �ΙΙPKC.16,17
The majority of �ΙPKC substrates we found constitutenuclear proteins involved in transcriptional control, includingchromatin remodeling and splicing processes. In undifferenti-ated ESC, �ΙPKC was localized in the nucleus; however, upondifferentiation, �ΙPKC was also found dispersed throughout thecell and excluded from the nucleus. Our results indicate that�ΙPKC may be involved in the regulation of protein transcrip-tion in undifferentiated ESC.
Experimental Section
Peptides. TAT protein transduction domain peptide, aminoacids 47-57 (TAT47-57) and peptides �C2-4, Ψ�RACK, and�ΙV5-3 coupled to the TAT protein transduction domainpeptide, amino acids 47-57,18 were obtained from KAIPharmaceuticals.
Cell Culture. The feeder independent embryonic stem cellline, E14TG2A (kindly donated by Dr. Joshua Brickman fromthe Institute for Stem Cell Research MRC Centre for Regenera-tive Medicine School of Biological Sciences University ofEdinburgh), was grown on 0.2% gelatin coated plates in GMEM(Sigma) supplemented with 15% fetal bovine serum (FBS),19 1mM sodium pyruvate, 1% MEM nonessential amino acids, 1× 103 U/mL murine leukemia inhibitory factor, 0.1 mM�-mercaptoethanol, 50 U/mL penicillin, and 50 µg/mL strep-tomycin at 37 °C with 5% CO2. Cells were subcultured every2-3 days after they reached 80% confluence. To ensure thatour cell culture conditions maintained the ESCs line in anundifferentiated state, we checked for expression of ESCsspecific markers by FACS, as described.20,21 More than 90% ofthe cells expressed Oct 3/4 and at least 85% expressed SSEA-1(data not shown). The feeder dependent cell line USP 2 (kindlydonated by Dr. Lygia Pereira da Veiga, Instituto de Biosciencias,Universidade de Sao Paulo) was maintained as previouslydescribed.22
Embryonic Stem Cell Differentiation. To differentiate theE14TG2A cells, we used the hanging drop differentiationmethod.23 Briefly, cells were trypsinized for passaging andresuspended in Differentiation medium [DMEM (4.5 g/Lglucose) supplemented with 20% fetal bovine serum and 2 mML-glutamine, 10 µM �-mercaptoethanol, 0.1 mM nonessentialamino acids, 50 U/mL penicillin, 50 µg/mL streptomycin]. Cellswere counted and viability was accessed using Trypan blue.Cells were diluted to 400-1000 cells per 20 µL. Twenty
microliter drops of the ESC suspension were placed on lids of100 mm bacteriological Petri dishes containing 5-10 mL of PBSand cultured as hanging drops for 2 days to form embryoidbodies (EBs). EBs were then rinsed from the lids of Petri dishesand cultured in suspension in 100 mm bacteriological Petridishes with 10 mL of Differentiation medium for 3 days.Individual EBs were then transferred to 24 well tissue culturegrade dishes. At different days of culture, RNA was extractedfrom pools of at least 3 EBs for RT-PCR analysis. For immu-nofluorescence studies, cells were transferred to 24 wellplates containing glass coverslips coated with 3% gelatin andcollected as described bellow.
Real-Time RT-PCR. RNA extraction, reverse transcription,and quantitative real-time PCR were performed as previouslydescribed.24 Briefly, total RNA was isolated with TRIzol Reagentaccording to the manufacturer’s instructions (Invitrogen).cDNA synthesis was performed with random hexamers (HighCapacity cDNA Archive kit-PE Applied Biosystems) and 5 ngof cDNA was used for real-time RT-PCR reaction (SYBR GreenPCR Master Mix-PE Applied Biosystems). All samples wereassayed in triplicate and GAPDH was used as an internalcontrol. The comparative CT (threshold cycle) method was usedfor data analyses. CT indicates the fractional cycle number atwhich the amount of amplified target reaches a fixed threshold,and ∆CT is the difference in threshold cycle for target (Oct 4)and reference (GAPDH). The primer sequences used were: forGAPDH, forward 5′-CAGCAACTCCCACTCTTCC-3′ and reverse5′-CCATGTAGGCCATGAGGTC-3′; for Oct 4, forward 5′-ATGC-CGTGAAGTTGGAGA-3′ and reverse 5′-TGTACCCCAAGGT-GATCCTC-3′.
Preparation of Cell Lysates and Subcellular Fractionation.ESC and total brain lysates were prepared as previouslydescribed.11 For two-dimensional gel eletrophoresis (2DE)studies, 80% confluent cell cultures were treated with 100 nMphorbol ester (PMA), or peptides �C2-4, Ψ�RACK and �ΙV5-3,or the control carrier TAT47-57 (500 nM for 15 min) and celllysates were prepared in 7 M of Urea, 2 M of Thiourea, 4% ofCHAPS (w/v), and 40 mM of DTT. For experiments thatinvolved fractionation of the soluble and particulate fraction,Triton X100 lysates were prepared by ultracentrifugation aspreviously described.11 Lysates of the feeder dependent cellline, USP2, were prepared as above, but first, separation offeeder cells from ESCs was performed by adherence to tissueculture plates.
For nuclear fractionation, cells were washed in cold PBScontaining 5 mM EDTA twice and incubated for 30 min at roomtemperature in solution I [10 mM HEPES, 10 mM KCl, 0.1 mMEDTA, 0.1 mM EGTA, protease inhibitor cocktail (1:300, Sigma# P8340), and phosphatase inhibitors cocktail (1:300, Sigma #P2850 and P5726)]; NP-40 was subsequently added to a finalconcentration of 3%. Cells were centrifuged at 20 000g for 1min and the supernatant containing the soluble fraction(cytosol) was kept at -80 °C until further use. The nuclearfraction (pellet) was solubilized in solution II (20 mM HEPES,420 mM NaCl, 1 mM EGTA, 1 mM EDTA, containing proteaseand phosphatase inhibitors as above) for 30 min on ice andcentrifuged at 20 000g for 10 min. The supernatant with thenuclear proteins was kept at -80 °C. Protein concentration inall cell lysates was determined by the Bradford assay (Bio-RadProtein Assay).
Western Blot. Western blot was performed by transferringSDS-PAGE, or 2D Gels, to nitrocellulose by the method ofTowbin.25 Specific anti-PKC antibodies raised against: the
research articles Costa-Junior et al.
6192 Journal of Proteome Research • Vol. 9, No. 12, 2010

C-terminus of PKC isozymes [R, �Ι, �ΙΙ, γ, δ, ε, �/λ, η, or θ (SantaCruz Biotechnology)], anti-hnRNPK (Chemicon), and anti�-actin and anti-Oct 3/4 (Santa Cruz Biotechnology) were usedat 0.4 µg/mL. Anti-GAPDH (Advanced Immuno Chemical, Inc.)was used at 0.01 µg/µL. Antibodies were diluted in blockingsolution (PBS, 0.1% Tween-20, 10% nonfat milk), for 2 h at roomtemperature. Goat secondary antibodies anti-rabbit IgG andanti-mouse IgG, conjugated to horseradish peroxidase (GE,Healthcare Life Science) were diluted 1:1000 in PBS 0.1%Tween. Immunodetection was performed by chemilumines-cence by exposure of membranes to films.
Confocal Immunofluorescence. For immunofluorescencestudies, cells at 80% confluence, or EBs cultured on 13 mmglass coverslips coated with 3% gelatin were fixed with 4% PFA.Prior to staining, the cells were permeabilized with PBS and0.1% Triton-X100; blocked in PBS, 0.1% Triton-X100, and 1%normal goat serum; and incubated in a humid chamber atroom temperature for 30 min. Cells were subsequently incu-bated in primary antibodies diluted in blocking solutionovernight at 4 °C in a humid chamber. Mouse monoclonalantibody anti-Oct 3/4 (Santa Cruz Biotechnology) and rabbitanti-�ΙPKC were used at 2 µg/mL. Secondary antibodies anti-mouse conjugated with Alexa 555 (4 µg/mL) and anti-rabbitconjugated with Alexa 488 (8 µg/mL) (Molecular Probes) werediluted in blocking solution and incubated in the dark at roomtemperature for 40 min. Immunofluorescence staining wasdetected using a Carl Zeiss 510 LMS confocal system connectedto an Axiovert microscope.
Two-Dimensional Gel Electrophoresis. Protein samples (300µg for analytic gels and 1 mg for preparative gels) were appliedonto 4-7 linear immobilized pH gradient strips (13 cm, GEHealthcare Life Science). Strips were rehydrated for 16 h atroom temperature. Isoelectric focalizations (IEF) were per-formed on an IPGphor III apparatus (GE Healthcare) at 17 kVhIEF. For the second dimension, strips were incubated at roomtemperature, for 20 min in equilibration buffer [6 M urea, 2%(w/v) SDS, 50 mM Tris-HCl, pH 6.8, 30% (v/v) glycerol, and0.001% (w/v) bromophenol blue] with 2% (w/v) DTT, followedby incubation with 4% (w/v) iodoacetamide in equilibriumbuffer, for 20 min. The second dimension was performed invertical SDS-PAGE. Triplicate lysates were prepared for eachtreatment. ESCs of similar passages were treated with eitherPKC modulator peptides or control TAT47-57 peptide. Gels oftriplicate lysates ran together to decrease the variability be-tween gels. Phosphoproteins were detected by staining withPro-Q Diamond (Invitrogen) as per manufacturer’s instructions.Gels were scanned using a Typhoon TRI scanner (GE Health-care Life Science). Gels were then stained with CoomassieBrilliant Blue G250 (CBB)26 and scanned using an UTA-1100scanner and Labscan v 5.0 software (GE Healthcare LifeScience).
Image analysis was performed using the Image MasterSoftware v.5.01 (GE Healthcare Life Science). For each pair ofsamples analyzed, individual spot volumes of triplicate gelswere determined in Pro-Q Diamond stained gels (phosphop-roteins), followed by normalization (individual spot volume/volume of all spots ×100). Spots (of treated samples) thatshowed a change in spot volume of least 1.5-fold as comparedto spots of control (TAT47-57) samples were excised from CBBstained preparative gels and identified by mass spectrometry.Differences between experimental groups were further evalu-
ated by Mann-Whitney t test. Where *p-value < 0.05, **p value< 0.01, and ***p-value < 0.001 were considered statisticallysignificant.
“In-Gel” Protein Digestion. Protein spots were excised fromgels, cut into 1 mm cubes, and washed with water for 15 min.Gel pieces were destained and washed several times with asolution of 50% (v/v) acetonitrile (ACN) and 50 mM ammoniumbicarbonate, until complete removal of the CBB. The 2-DE gelspots were completely dehydrated with 100% (v/v) ACN,rehydrated with 20 mM DTT, and maintained for 40 min at 60°C. This solution was then discarded and replaced with 55 mMiodoacetamide and incubated in the dark for 30 min. Gel sliceswere dehydrated again with 100% ACN and air-dried forcomplete removal of solvent. Protein digestion was carried outwith a solution of 10 ng /µL Sequencing grade Modified Trypsin(Promega), in 25 mM ammonium bicarbonate. Gel pieces wererehydrated with trypsin solution and incubated for 12 h at 37°C. After digestion, gel plugs were extracted twice with 50 µLof 60% (v/v) ACN, 1% (v/v) Formic Acid (FA), and once with50 µL of ACN. All supernatants were combined and vacuum-dried. Peptides were suspended in 12 µL of 1% (v/v) FA formass spectrometry (MS) analysis.27
Protein Identification and Analysis by Mass Spectrometry.Peptide mixtures were identified by online chromatographyusing a Cap-LC coupled to a Q-TOF Ultima API mass spec-trometer (Waters). Five microliters of sample was loaded ontoa NanoEase Trapping column 0.18 × 23.5 mm (Waters) forpreconcentration, followed by peptide separation in a LCnanoease column Symmetry 300 C18 3.5 µm, 75 × 100 mm(Waters). Peptides were eluted in a 60 min linear gradient ofsolvent B [95% (v/v) ACN, 0.1% (v/v) FA in water] at a flowrate of 250 nL/min. Solvent A consisted of 5% (v/v) ACN, 0.1%(v/v) FA in water. All analysis was performed using a positiveion mode at a 3 kV needle voltage. Mass range was set at300-2000 m/z and the MS/MS spectra were acquired for themost intense peaks (g15 counts). Multiple charged precursorions were selected for fragmentation and peptide sequencingwas performed in an automated data-dependent acquisition(DDA) MassLynx software (Waters), switching from the MS toMS/MS mode and then returning to MS mode. The resultingfragmented spectra were processed using the ProteinLynx v4.0software (Waters). MALDI-TOF/TOF MS (Matrix-Assisted LaserDesorption Ionization Time-of-Flight/Time-of-Flight Mass Spec-trometry) was performed as previously described.28 For massspectrometry analysis, the peptides were cocrystallized with 0.3µL of 10 mg/mL R-cyano-4-hydroxycinnamic acid solution in0.1% (w/v) trifluoroacetic acid, 50% (v/v) acetonitrile directlyonto a MALDI target plate. Raw data for protein identificationwere obtained on the 4700 Proteomics Analyzer (AppliedBiosystems, Foster City, CA). Both MS and MS/MS data wereacquired in positive and reflectron modes using a neodymium-doped yttrium aluminum garnet (Nd:YAG) laser with a 200-Hzrepetition rate. Typically, 1600 shots were accumulated forspectra in MS mode while 3000 shots were accumulated forspectra in MS/MS mode. Up to 10 of the most intense ionsignals with signal-to-noise ratio above 20 were selected asprecursors for MS/MS acquisition excluding common trypsinautolysis peaks and matrix ion signals. External calibration inMS mode was performed using a mixture of four peptides: des-Arg1-Bradykinin (m/z ) 904.47), angiotensin I (m/z ) 1296.69),Glu1-fibrinopeptide B (m/z ) 1570.68), and ACTH (18-39) (m/z) 2465.20). MS/MS spectra were externally calibrated using
Role of Nuclear �ΙPKC in Transcription Processes of Murine ESCs research articles
Journal of Proteome Research • Vol. 9, No. 12, 2010 6193
known fragment ion masses observed in the MS/MS spectrumof angiotensin I.
The MASCOT MS/MS Ion Search (www.matrixscience.com)software was used to blast sequences against the Swiss-Prot
and NCBInr databanks. Combined MS-MS/MS searches wereconducted with parent ion mass tolerance at 50 ppm, MS/MSmass tolerance of 0.2 Da, carbamidomethylation of cysteine(fixed modification), and methionine oxidation (variable modi-fication). According to MASCOT probability analysis, only hitswith significant p-values (P < 0.05) were accepted.
Western Blot of Two-Dimensional Gel Electrophoresis.E14TG2A cells were washed in ice-cold PBS buffer scraped witha rubber policeman in 20 mM Tris, pH 7.5, 1 mM EDTA, 1 mMEGTA, 12 mM 2-mercaptoethanol, 10% (v/v) glycerol, 1% (v/v)Triton-X 100, and freshly added Complete Protease InhibitorCocktail (as described above). Samples were precipitated with3 vol of ice-cold acetone at -20 °C for 3 h. Residual acetonewas then removed by air-drying. Proteins were resuspendedin 8 M Urea, 2% (w/v) CHAPS, 0.5% (v/v) IPG Buffer, pH 4-7,and 0.002% (w/v) Bromophenol blue. Protein concentrationwas then determined by Bradford assay (Bio-Rad). Proteinsamples (150 µg) were applied onto 4-7 linear immobilizedpH gradient strips (7 cm, GE Healthcare Life Science). Stripswere rehydrated for 16 h at room temperature. The first-dimension isoelectric focusing was performed as describedabove. The second dimension was performed by SDS-PAGEusing vertical electrophoresis system SE 260 (mini-vertical; GEHeathcare). Gels were transferred to nitrocellulose and Westernblot was performed as described above.
Thymidine Incorporation. Cells at 5 × 103 cells/well wereplated in 24 well plates previously coated with 0.2% gelatin.
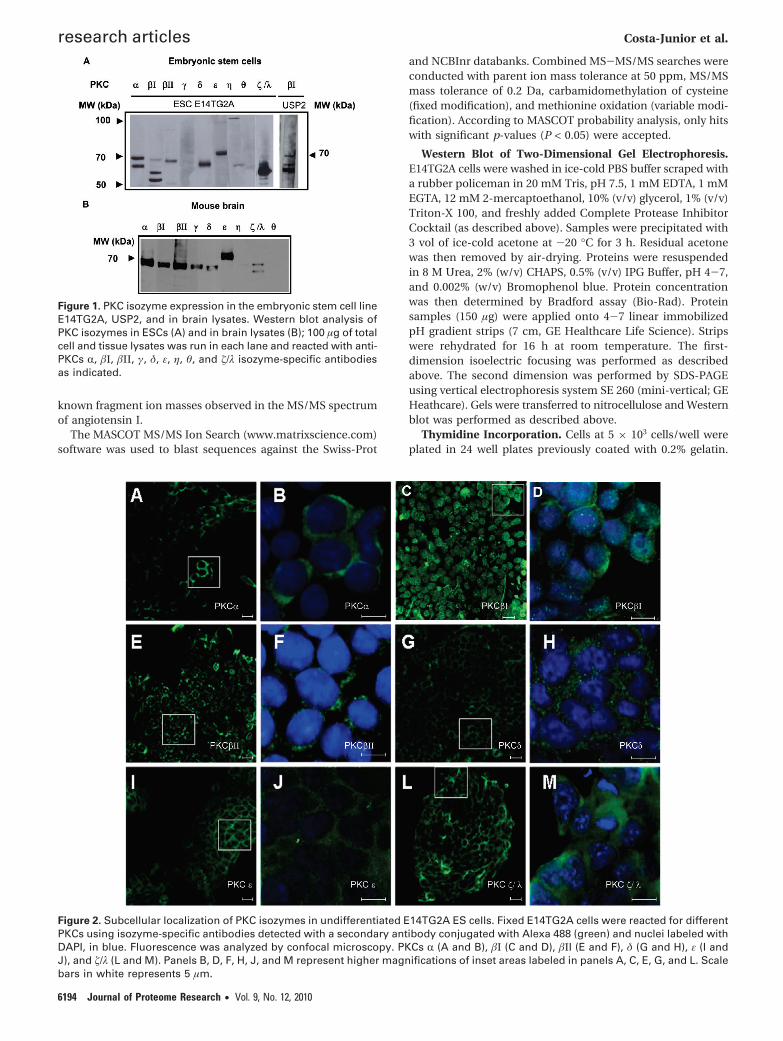
Figure 1. PKC isozyme expression in the embryonic stem cell lineE14TG2A, USP2, and in brain lysates. Western blot analysis ofPKC isozymes in ESCs (A) and in brain lysates (B); 100 µg of totalcell and tissue lysates was run in each lane and reacted with anti-PKCs R, �Ι, �ΙΙ, γ, δ, ε, η, θ, and �/λ isozyme-specific antibodiesas indicated.
Figure 2. Subcellular localization of PKC isozymes in undifferentiated E14TG2A ES cells. Fixed E14TG2A cells were reacted for differentPKCs using isozyme-specific antibodies detected with a secondary antibody conjugated with Alexa 488 (green) and nuclei labeled withDAPI, in blue. Fluorescence was analyzed by confocal microscopy. PKCs R (A and B), �Ι (C and D), �ΙI (E and F), δ (G and H), ε (I andJ), and �/λ (L and M). Panels B, D, F, H, J, and M represent higher magnifications of inset areas labeled in panels A, C, E, G, and L. Scalebars in white represents 5 µm.
research articles Costa-Junior et al.
6194 Journal of Proteome Research • Vol. 9, No. 12, 2010

Four hours after plating, cells were treated with PKC modulatorpeptides in normal culture conditions, as described above.Peptides were further added every 24 h as previously de-scribed.29 At 60 h, 1 µCi/mL of [3H] thymidine/well was addedand cells were further incubated for 12 h. Cells were thenwashed with 1× phosphate buffered saline, incubated with 15%Tricloro acetic acid for 5 min, and washed twice with ethanolfor 5 min; 250 µL of 250 mM NaOH was subsequently addedto each well and cells were incubated for 10 min at roomtemperature. Cells were lysed and scraped with rubber police-man. To 50 µL of cell lysate, 2 mL of scintillation liquid wasadded. Thymidine incorporation was read in a �-counter andresults were calculated relative to control cells cultured in 15%fetal bovine serum as described above.
Results and Discussion
PKC Profiling Indicates That E14TG2A Embryonic StemCells Express PKCs r, �Ι, �ΙΙ, δ, ε, and �/λ. To characterizePKC isozyme expression in the murine ESC line E14TG2A, weused PKC isozyme specific antibodies raised against the PKCC-terminal variable V5 region (approximately last 50 aminoacids). With these antibodies, we determined that the ESC lineE14TG2A expresses PKCs R, �Ι, �ΙΙ, δ, ε, and �/λ (Figure 1).
It has called our attention that in E14TG2A most of the �ΙPKCimmunoreactivity is represented by lower molecular weightforms (approximately 65 and 63 kDa). This contrasts with theother PKCs, in which the full-length form predominates. Sincethe antibodies we utilized recognize the C-terminal V5 domain,low molecular weight �ΙPKC forms must conserve the C-terminal domain, which in turn suggests that they are truncatedat their N-terminal regulatory domains. N-terminally truncated,lower molecular weight forms of �ΙPKC have been previouslydescribed in the nucleus of rapidly dividing, multipotentneoplastic and epithelial cells.30-32 It has been speculated thatthe lack of the regulatory N-terminal region turns thesetruncated forms into constitutively active enzymes,30-32 whichmay play roles in proliferation, or in the maintenance of theundifferentiated phenotype. To establish whether the predomi-nance of truncated �ΙPKC is a feature of ESCs, or a specificfeature of E14TG2A we characterized expression of �ΙPKC inUSP2, another ESC line and in mouse brain lysates. Figure 1shows that lower molecular weight forms of �ΙPKC are indeedpresent in another ESC line, but not in brain lysates (Figure1B).
�ΙPKC Is the Only PKC Isozyme Abundantly Expressed inthe Nucleus of Undifferentiated ESCs. To establish the sub-cellular localization of the different PKC isozymes in theundifferentiated ESC line E14TG2A, we performed immunof-luorescent confocal analysis of PKC expression. Consistent withthe pattern of �ΙPKC described in undifferentiated, multipotentneoplastic and epithelial cells,30-32 in murine morula and inpluripotent cells derived of the inner mass of blastocysts,33 weestablished that �ΙPKC is predominantly expressed inside nucleiof E14TG2A. In some E14TG2A cells, �ΙPKC expression was alsodocumented in filamentous structures. In contrast to �ΙPKC,RPKC is localized throughout the cytoplasm. �ΙΙPKC is con-centrated at a perinuclear region and in a dot like structurewithin the nucleus, reminiscent of pericentrin, as has beenpreviously reported in other cell lines.34 Both PKCs δ and ε arediffusely expressed throughout the cytoplasm, while PKCs RPKCand �/λ are expressed throughout the cells (Figure 2). Insummary, �ΙPKC and �ΙΙPKC stand out from the other PKCsexpressed in undifferentiated E14TG2A cells as the only isozymes
that display restricted subcellular localization. These resultssuggest that, in E14TG2A, �ΙPKC and �ΙΙPKC are poised to playspecific roles linked to the nuclear or perinuclear functions,while the other PKCs appear to be prone to pleotropic roles inthese cells.
�ΙPKC Is Enriched in the Particulate and Nuclear Frac-tions of Undifferentiated ESCs. It is well established that uponactivation PKCs translocate from the soluble to the particulatefraction of the cell.35 It has also been demonstrated thatN-terminally truncated PKCs, which miss their regulatory units,represent constitutively active enzyme forms. To further char-acterize the functional status of PKC signaling in ESCs cells,we fractionated E14TG2A and determined the relative enrich-ment of low and high molecular forms of �ΙPKC in thecytoplasmatic, particulate, and nuclear fractions. We found thatlower molecular weight forms of �ΙPKC are enriched in theparticulate and in the nuclear fractions (Figure 3A), consistentwith the confocal microscopy data depicted in Figure 2. Incontrast, full-length forms of �ΙPKC were predominantly foundin the soluble fraction.
Phorbol Ester Activation Further Enriches �ΙPKC in theNuclear Fraction. To investigate whether the relative distribu-tion of �ΙPKC forms between nucleus and cytoplasm is modi-fied by general PKC activation, we fractionated E14TG2Afollowing treatment with 100 nM PMA (for 10 min). Wedetermined that the N-terminally truncated 63 kDa form of�ΙPKC and the full-length protein increased in the nuclearfraction after PMA and that there was a correspondent decreasein the full-length �ΙPKC in the cytosol (Figure 3B). In summary,
Figure 3. �ΙPKC is expressed in the nucleus of undifferentiatedESCs. Western blot analysis of �ΙPKC in the cytosolic (C) andparticulate (P) fractions and GAPDH levels in the cytosolic fraction(A). E14TG2A cell lysates were fractionated into cytosolic (C) andnuclear (N) fractions in the presence or absence of 100 nM PMAfor 10 min and probed for �ΙPKC. Fractions were also probedfor GAPDH and the transcription factor Oct 3/4 as controls forthe cytosolic and nuclear fractions, respectively (B, upper panel).The amounts of protein in the nuclear fractions were normalizedby the expression of the transcription factor Oct 3/4, and in panelB, bottom panel, we show a quantitative representation of thenuclear translocation of the different molecular weight speciesof �ΙPKC upon activation with 100 nM PMA relative to controlcells.StatisticalsignificancewasdeterminedbytheMann-Whitneyt test where *p < 0.05 as indicated, (averages of 4 independentexperiments were considered).
Role of Nuclear �ΙPKC in Transcription Processes of Murine ESCs research articles
Journal of Proteome Research • Vol. 9, No. 12, 2010 6195

our results indicate that �ΙPKC forms predominate in thenuclear fraction of undifferentiated E14TG2A (Figure 3A) andthat this dominance is further exacerbated after general PKCstimulation by PMA (Figure 3B).
Detection of �ΙPKC Substrates in E14 TG2A Cells UsingPhosphoproteomics. Our PKC profiling and characterizationof subcellular PKC distribution in E14TG2A cells are generallyconsistent with the view that undifferentiated ESCs and othermultipotent cells display an increased concentration of N-terminally truncated, presumably active, �ΙPKC in the nucleus.The nuclear localization of smaller molecular weight speciesof PKC isoenzymes has been previously reported and suggestedto represent the result of nuclear translocation mechanismsoperated by nuclear localization signals exposed at the C-terminus of PKC isoenzymes such as �Ι, R, and δ PKCs.36 Futurestudies are necessary to further characterize these lowermolecular weight forms of �ΙPKC and the processes that leadto �ΙPKC nuclear translocation. While it is possible that �ΙPKCis indeed active in the nucleus of undifferentiated E14TG2A,the evidence is only associative. Therefore, to establish theactivity of �ΙPKC under these conditions, we took a phosphop-roteomic approach. For that we used protein extracts from cellstreated with PKC peptide modulators (Ψ�RACK, a cPKC activa-tor, �C2-4, a cPKC inhibitor, and �ΙV5-3, a �ΙPKC isozyme-specific inhibitor) coupled to the transduction domain of theHIV TAT protein, TAT47-57. In Pro-Q Diamond stained gels, wedetermined that 107 spots had their phosphorylation statusmodulated by cPKC modulator peptides. Of these, 50 spots hadtheir phosphorylation increased with Ψ�RACK and 57 de-
creased with �C2-4. Afterward, Pro-Q Diamond staining gelswere subsequently stained with Coomassie Coloidal Blue.Among the most abundant spots identified by mass spectrom-etry, we established the molecular identity of 11 spots whosephosphorylation was increased by Ψ�RACK and 21 spots whosephosphorylation was decreased by �C2-4. These spots cor-responded to 27 different proteins (Figure 4, SupplementalFigures S1, S3-7, and Table 1). Of these, 27% were nuclearproteins (Table 1). In cells treated with �ΙV5-3, we saw adecrease in the phosphorylation of 11 spots and the disap-pearance of 26 spots, as compared to cells treated with thecontrol TAT47-57. Of these, 14 spots corresponding to 12different proteins were identified and, 67% were nuclearproteins, consistent with the preferential nuclear localizationof �ΙPKC (Figure 5, Supplemental Figures S2, S8-12 and Table2). Some proteins, such as retinoblastoma-binding proteinmRbAp48, heterogeneous nuclear ribonucleoprotein C1/C2,Janus kinase and microtubule-interacting protein 3 (Jakmip3),and R-tubulin chain 1C, were detected with more than onepeptide modulator, supporting their status of cPKC targets. Ofthese, retinoblastoma-binding protein mRbAp48 and R-tubulinwere detected with a �ΙPKC-specific peptide inhibitor (Tables1 and 2).
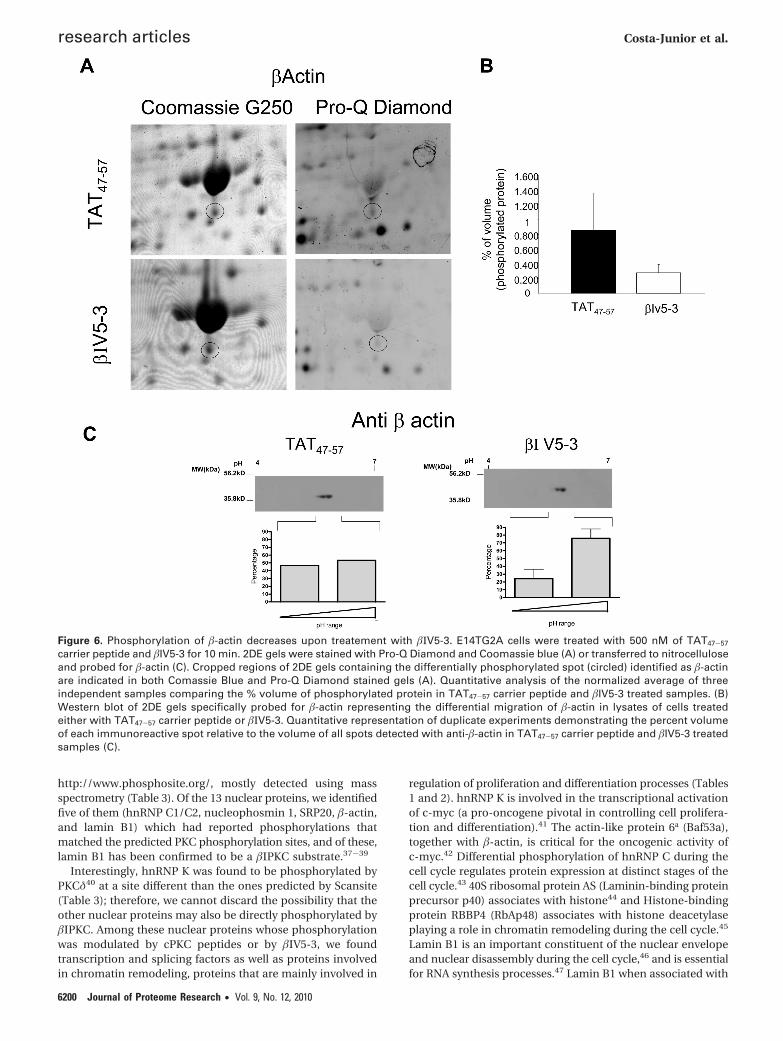
To validate our strategy of detecting �ΙPKC direct andindirect targets, we ran 2D Western blots and specificallyprobed for two �ΙPKC targets, �-actin and hnRNP K (Figures 6and 7). �-Actin ran as two spots equally in lysates TAT47-57
treated cells. After treatment with �ΙV5-3, we verified a markeddecrease in intensity of the more acidic spot, confirming that
Figure 4. Detection of direct and indirect cPKC substrates by phosphoproteomics. Representative 2DE gels of E14TG2A cells treatedwith 500 nM TAT47-57 carrier peptide (A and D), Ψ�RACK (B and E), or �C2-4 (C and F) peptides for 10 min. Gels stained with phospho-specific dye (A-C) and Coomassie blue G250 (D-F). Spots labeled in panels A and D indicate spots used to align Coomassie blue andPro-Q Diamond stained gels. Numbers in panels B, C, E, and F indicate identified spots whose average phosphorylation in two out ofthree independent experiments increased (B) or decreased (C) at least 1.5-fold as compared to TAT47-57 (A). For the protein annotationof the labeled spots in panels B, C, E, and F and statistical significance of the changes in phosphorylation, see Table 1. Enhancedcropped regions of the nuclear proteins identified can be found in Supplementary Figures S1 and S3-7.
research articles Costa-Junior et al.
6196 Journal of Proteome Research • Vol. 9, No. 12, 2010

Tab
le1.
Pro
tein
sId
enti
fied
by
Mas
sS
pec
tro
met
ryW
ho
seP
ho
sph
ory
lati
on
Dec
reas
edo
rIn
crea
sed
wit
hcP
KC
sM
od
ula
tor
Pep
tid
eR
elat
ive
toT
reat
men
tw
ith
TA
T47
-57
a
theo
rica
lex
per
imen
tal
vol
(%)
of
Co
ntr
ol
TA
T47
-57
vol
(%)
of
ψ�R
AC
Ktr
eatm
ent
ψ�R
AC
Kp
rote
inU
nip
rot
met
ho
dse
q(%
)p
ept
sco
reM
Wp
IM
Wp
Iex
p1
exp
2ex
p3
med
iaer
ror
exp
1ex
p2
exp
3m
edia
erro
rp
-val
ue
cellu
lar
loca
tio
n
1R
bA
p48
Q60
972
ESI
Q-T
OF
249
158
524.
9552
4.76
0.19
80.
050
0.40
30.
217
0.17
70.
306
0.05
90.
616
0.32
70.
279
0.11
6n
ucl
eus
2tr
ansp
ort
ing.
lyso
som
alP
5051
6E
SIQ
-TO
F27
1262
695.
4261
5.57
00
00
00.
627
0.91
40.
581
0.70
70.
180
0.01
0m
ito
cho
nd
ria
3m
ann
ose
-6-
ph
osp
hat
ere
cep
tor
bin
din
gp
rote
in1
Q9D
BG
5E
SIQ
-TO
F24
722
447
5.45
515.
60
00
00
0.14
50.
137
0.39
00.
224
0.14
40.
047
cyto
pla
sm
4A
ctin
-lik
ep
rote
in6A
Q9Z
2N8
ESI
Q-T
OF
102
8447
5.39
495.
640
00
00
0.06
30.
058
0.06
80.
063
0.00
50.
001
nu
cleu
s
5A
ctin
.cy
top
lasm
ic1
P60
710
ESI
Q-T
OF
173
243
415.
2940
5.26
00
00
00.
167
0.15
20.
163
0.16
00.
007
0.00
04cy
top
lasm
6R
AC
K1
P68
040
ESI
Q-T
OF
83
5535
7.60
304.
90
00
00
0.05
10.
036
0.06
50.
051
0.01
40.
012
cyto
pla
sm/
mem
bra
ne
7A
DP
-rib
osy
larg
inin
eh
ydro
lase
Q3U
5N4
ESI
Q-T
OF
63
4040
5.43
305.
730
00
00
0.11
50.
173
0.10
10.
130
0.03
70.
013
cyto
pla
sm
8T
CT
P1
sub
un
itγ
P80
318
ESI
Q-T
OF
93
4060
6.28
635.
20
00
00
0.05
50.
051
0.05
20.
052
0.00
20.
0003
cyto
pla
sm9
hn
RP
KC
1/C
2Q
9Z20
4E
SIQ
-TO
F18
1321
934
4.91
384.
940.
078
0.03
70
0.03
80.
039
0.24
00.
132
00.
124
0.12
00.
104
cyto
pla
sm/
nu
cleu
s10
sper
mid
ine
syn
thas
eQ
6467
4E
SIQ
-TO
F11
369
345.
3128
5.4
0.40
60.
085
00.
163
0.21
40.
637
0.65
80
0.43
20.
374
0.12
4cy
top
lasm
11JA
KM
IP3
Q5D
TN
8E
SIQ
-TO
F34
1410
198
5.61
575.
690.
031
0.06
60.
165
0.08
70.
069
0.06
10.
152
0.16
0.12
40.
054
0.14
8go
lgi
theo
rica
lex
per
imen
tal
vol
(%)
of
con
tro
lT
AT
47-
57vo
l(%
)o
f�C
2-4
trea
tmen
t
�C2-
4p
rote
inU
nip
rot
met
ho
dse
q(%
)p
ept
sco
reM
Wp
IM
Wp
Iex
p1
exp
2ex
p3
med
iaer
ror
exp
1ex
p2
exp
3m
edia
erro
rp
-val
ue
cellu
lar
loca
tio
n
1H
SP70
pro
tein
4Q
6131
6E
SIQ
-TO
F28
1288
694
5.15
103
5.35
0.03
10.
066
0.16
50.
087
0.06
90.
008
0.01
30.
115
0.04
50.
060
0.04
7cy
top
lasm
2H
spa9
NP
0346
11E
SIQ
-TO
F32
1890
473
5.91
645.
680.
021
0.03
10.
028
0.02
70.
005
00
00
00.
006
mit
och
on
dri
a/cy
top
lasm
3JA
KM
IP3
Q5D
TN
8E
SIQ
-TO
F31
1315
598
5.61
575.
690.
187
0.38
80.
716
0.43
00.
267
0.09
40.
236
0.16
70.
165
0.07
10.
024
golg
i4
Hsp
60P
6303
8E
SIQ
-TO
F18
1110
261
5.91
585.
320.
305
0.42
10.
351
0.35
90.
058
0.16
80.
057
0.34
50.
190
0.14
40.
124
mit
och
on
dri
a5
Rtu
bu
linP
0521
3E
SIQ
-TO
F26
1013
750
4.94
564.
650.
235
0.20
20.
371
0.26
90.
089
0.12
40.
132
0.12
90.
128
0.00
30.
05cy
tosk
elet
on
6ri
bo
nu
clea
sein
hib
ito
rQ
91V
I7E
SIQ
-TO
F11
780
514.
6951
4.5
0.10
60.
179
0.15
80.
147
0.03
70.
067
0.06
40.
098
0.07
60.
018
0.04
4cy
top
lasm
7R
bA
p48
Q60
972
ESI
Q-T
OF
218
167
524.
9552
4.76
0.19
80.
403
0.05
00.
217
0.17
70.
069
0.07
20.
036
0.05
90.
019
0.10
1n
ucl
eus
8SA
E1
Q9R
1T2
ESI
Q-T
OF
3010
149
385.
2440
5.27
0.43
30.
313
0.33
60.
360
0.06
30.
254
0.14
20.
282
0.22
60.
073
0.03
9n
ucl
eus
9h
nR
NP
C1/
C2
Q9Z
204
ESI
Q-T
OF
135
180
344.
9243
5.44
0.01
30.
027
0.01
50.
018
0.00
70
00
00
0.02
6cy
top
lasm
/n
ucl
eus
10h
nR
NP
C1/
C2
Q9Z
204
ESI
Q-T
OF
135
121
344.
9239
5.53
0.03
20.
025
0.03
80.
031
0.00
60
00
00
0.00
6cy
top
lasm
/n
ucl
eus
11h
nR
NP
C1/
C2
Q9Z
204
ESI
Q-T
OF
268
295
344.
9240
4.89
0.43
30.
313
0.32
50.
357
0.06
60.
254
0.14
20.
215
0.20
40.
056
0.00
9cy
top
lasm
/n
ucl
eus
12n
ucl
eop
ho
smin
1C
AI2
5148
ESI
Q-T
OF
205
4429
4.45
214.
391.
626
1.26
01.
023
1.30
30.
303
0.75
00.
612
1.10
20.
821
0.25
20.
118
nu
cleu
s13
elo
nga
tio
nfa
cto
r1
bet
aO
7025
1E
SIQ
-TO
F20
769
254.
5327
4.62
4.27
92.
532
2.56
23.
125
1.00
01.
831
2.01
31.
689
1.84
50.
162
0.08
1cy
top
lasm
Role of Nuclear �ΙPKC in Transcription Processes of Murine ESCs research articles
Journal of Proteome Research • Vol. 9, No. 12, 2010 6197

�-actin is a target of �ΙPKC (Figure 6). As for hnRNP K, wedetected four spots that displayed reactivity to anti-hnRNP Kantibodies in lysates of cells treated with TAT47-57. Aftertreatment with �ΙV5-3, the more acidic spot almost completelydisappeared, confirming our results with Pro-Q Diamond(phospho-specific stain) (Figure 7), and thus corroborating thathnRNP K is another �ΙPKC target.
To investigate whether the nuclear targets we found are likelyto be direct �ΙPKC substrates, we searched for putative PKCphosphorylation sites in the nuclear proteins identified of bothmurine and human species using Scansite (http://scansite.mit-.edu/). Since PKC isozyme specificity is mostly due to subcel-lular localization of substrates and known PKC consensussequences are highly homologous among the different isozymes,we predicted phosphorylation sites for cPKCs and for nPKCs δand ε using low stringency search mode. As can bee seen inTable 3, potential PKC phosphorylation sites were found in allnuclear proteins identified and most of these were sitesconserved between human and murine proteins except fortranslation initiation factor 3 in which a potential PKC phos-phorylation site was solely found in the human protein. Further,we compared the predicted PKC phosphorylation sites tomatching phosphorylated serines and threonines deposited inT
ab
le1.
Co
nti
nu
ed
theo
rica
lex
per
imen
tal
vol
(%)
of
Co
ntr
ol
TA
T47
-57
vol
(%)
of
ψ�R
AC
Ktr
eatm
ent
ψ�R
AC
Kp
rote
inU
nip
rot
met
ho
dse
q(%
)p
ept
sco
reM
Wp
IM
Wp
Iex
p1
exp
2ex
p3
med
iaer
ror
exp
1ex
p2
exp
3m
edia
erro
rp
-val
ue
cellu
lar
loca
tio
n
14A
PC
-bin
din
gp
rote
inE
B1
Q61
166
ESI
Q-T
OF
287
161
305.
1230
5.32
0.09
10.
098
0.03
40.
074
0.03
50.
040
0.05
60.
044
0.04
70.
008
0.14
6cy
top
lasm
/cy
tosk
elet
on
15p
rote
aso
me
sub
un
itb
eta
typ
e-4
Q55
6Q0
ESI
Q-T
OF
72
6529
5.47
205.
770.
043
0.15
40.
086
0.09
40.
056
0.02
70.
103
0.05
40.
061
0.03
80.
041
cyto
pla
sm/
nu
cleu
s16
AT
Psy
nth
ase
sub
un
itδ,
mit
och
on
dri
al
CA
P19
193
ESI
Q-T
OF
4412
238
195.
5217
5.7
0.07
30.
089
0.25
50.
139
0.10
00.
040
0.06
00.
141
0.08
00.
053
0.08
2m
ito
cho
nd
ria
17p
ero
xire
do
xin
2Q
6117
1E
SIQ
-TO
F13
315
822
5.20
195.
220.
072
0.05
70.
065
0.06
50.
007
0.03
90.
022
0.03
50.
032
0.00
90.
001
cyto
pla
sm18
Ub
iqu
itin
carb
oxy
l-te
rmin
ales
tera
seL3
AA
H48
481
ESI
Q-T
OF
265
151
264.
9621
4.87
0.17
40.
108
0.16
10.
147
0.03
40.
029
0.04
00.
125
0.06
50.
052
0.06
3cy
top
lasm
19T
CT
P1
P63
028
ESI
Q-T
OF
52
4020
4.76
214.
770.
150
0.13
50.
183
0.15
60.
024
0.07
90.
10.
086
0.08
80.
010
0.03
1cy
top
lasm
20H
sp90
b1
NP
0357
61E
SIQ
-TO
F35
1424
893
4.74
884.
830.
187
0.38
70.
716
0.43
00.
266
0.09
40.
235
0.16
60.
165
0.07
00.
102
end
op
lasm
icre
ticu
lum
21ca
lret
icu
linP
1421
1E
SIQ
-TO
F21
817
948
4.34
564.
450.
102
0.11
50.
103
0.10
60.
007
00
00
00.
0008
end
op
lasm
atic
reti
culu
m21
R-t
ub
ulin
M-2
P05
213
ESI
Q-T
OF
75
177
504.
9453
4.45
0.10
20.
1154
890.
103
0.10
60.
007
00
00
00.
0008
cyto
pla
sm/
cyto
skel
eto
n
aSp
ots
nu
mb
ered
inF
igu
re4
are
ind
icat
edto
geth
erw
ith
Un
ipro
tac
cess
ion
nu
mb
er,
the
met
ho
du
sed
for
spo
tid
enti
fica
tio
n,
%o
fse
qu
ence
cove
red
by
pep
tid
es,
nu
mb
ero
fp
epti
des
iden
tifi
ed,
Mas
cot
sco
re,
theo
reti
cal
and
exp
erim
enta
lm
ole
cula
rw
eigh
t(M
W)
and
iso
elet
ric
po
int
(pI)
,%
volu
me
of
con
tro
lT
AT
47-
57an
dtr
eate
dsa
mp
les
(ave
rage
of
thre
eex
per
imen
ts),
stan
dar
der
ror,
and
p-v
alu
esas
det
erm
ined
by
Wh
itn
eyt-
test
wh
ere
*P<
0.05
are
ind
icat
ed.
Figure 5. Detection of direct and indirect �ΙPKC substrates byphosphoproteomics. Representative 2DE gels of E14TG2A cellstreated with 500 nM TAT47-57 carrier peptide (A and C), and �IV5-3peptide (B and D) for 10 min. Gels stained with phospho-specificdye (A and B) and Coomassie blue G250 stained gels (C and D).Spots labeled in panels A and C indicate spots used to alignCoomassie blue and Pro-Q Diamond stained gels. Numbers inpanels B and D indicate spots that were identified, whose averagephosphorylation in three independent experiments decreased atleast 1.5-fold as compared to TAT47-57.. For the protein annotationof the labeled spots in panels B and D and statistical significanceof the changes in phoshphorylation, see Table 2. Enhancedcropped regions of the nuclear proteins identified can be foundin Supplementary Figures S2 and S8-12.
research articles Costa-Junior et al.
6198 Journal of Proteome Research • Vol. 9, No. 12, 2010

Tab
le2.
Pro
tein
sId
enti
fied
by
Mas
sS
pec
tro
met
ryW
ho
seP
ho
sph
ory
lati
on
Dec
reas
edw
ith
the
�IV
5-3
Pep
tid
eR
elat
ive
toT
reat
men
tw
ith
TA
T47
-57
a
MA
LDI
MS-
MS/
ESI
Q-T
OF
theo
rica
lex
per
imen
tal
vol
(%)
of
con
tro
lT
AT
47-
57vo
l(%
)o
f�V
5-3
trea
tmen
t
�V5-
3p
rote
inU
nip
rotv
met
ho
dse
q(%
)P
ept
Sco
reM
WI
MW
pI
exp
1ex
p2
exp
3m
edia
erro
rex
p1
exp
2ex
p3
med
iaer
ror
p-v
alu
ece
lula
rlo
cati
on
1h
nR
NP
KP
6197
9M
ALD
IM
S-M
S/E
SIQ
-TO
F18
/16
6/6
151/
232
515.
3961
5.03
0.08
80.
078
0.07
20.
079
0.00
80
00.
002
0.00
00.
001
0.00
2n
ucl
eus/
cyto
pla
sm2
hn
RN
PK
P61
979
MA
LDI
MS-
MS/
ESI
Q-T
OF
14/9
5/3
90/8
751
5.39
575.
290.
693
0.11
10.
390
0.39
80.
291
0.05
30.
319
0.22
10.
198
0.13
40.
132
nu
cleu
s/cy
top
lasm
2T
CP
-1-�
P80
317
ESI
Q-T
OF
83
3557
6.67
615.
290.
693
0.11
10.
390
0.39
80.
291
0.05
30.
319
0.22
10.
198
0.13
40.
132
cyto
pla
sm3
hn
RN
PF
Q9Z
2X1
MA
LDI
MS-
MS/
ESI
Q-T
OF
8/16
2/8
49/4
546
5.31
485.
310.
751
0.17
20.
327
0.41
70.
299
0.05
50.
376
0.24
30.
225
0.16
10.
178
nu
cleu
s/cy
top
lasm
4SR
P20
P84
104
ESI
Q-T
OF
138
5488
5.15
116
5.38
1.23
50.
274
0.85
90.
789
0.48
40
0.29
20.
258
0.18
30.
159
0.01
8n
ucl
eus
5R
-tu
bu
linis
oty
pe
M-R
-6P
6837
3M
ALD
IM
S-M
S/E
SIQ
-TO
F/2
63/
917
1/13
750
4.94
564.
650.
647
0.65
40.
617
0.63
90.
019
0.56
40.
339
0.30
70.
403
0.13
90.
046
cyto
pla
sm/
cyto
skel
eto
n6
R-t
ub
ulin
iso
typ
eM
-R-2
P05
213
MA
LDI
MS-
MS/
ESI
Q-T
OF
17/4
05/
1321
0/25
450
4.94
554.
360.
647
0.33
70.
617
0.53
40.
171
0.33
90.
307
0.27
20.
306
0.03
30.
094
cyto
pla
sm/
cyto
skel
eto
n6
calr
etic
ulin
P14
211
MA
LDI
MS-
MS/
ESI
Q-T
OF
6/7
2/2
107/
5548
4.34
554.
360.
647
0.33
70.
617
0.53
40.
171
1.34
00.
307
0.60
60.
751
0.53
10.
094
end
op
lasm
atic
reti
culu
m/
pla
sma
mem
bra
ne
7�-
acti
nP
6071
0M
ALD
IM
S-M
S/E
SIQ
-TO
F33
/47
9/19
534/
156
415.
2241
5.52
0.30
90.
926
1.31
90.
852
0.50
90.
354
0.33
40.
146
0.27
80.
114
0.12
2cy
top
lasm
/cy
tosk
elet
on
/n
ucl
eus
8la
min
B1
P14
733
MA
LDI
MS-
MS/
ESI
Q-T
OF
30/3
418
/21
122/
127
675.
1167
5.24
0.43
70.
351
0.42
80.
405
0.04
70.
283
0.27
90.
203
0.25
50.
045
0.04
9cy
tosk
elet
on
/n
ucl
eus
8H
SP�8
Q9J
K92
MA
LDI
MS-
MS/
ESI
Q-T
OF
13/2
86/
1190
/120
715.
2867
5.24
0.43
70.
351
0.42
80.
405
0.04
70.
283
0.27
90.
203
0.25
50.
045
0.04
9cy
top
lasm
9eI
F-3
-εQ
9DC
H4
MA
LDI
MS-
MS/
ESI
Q-T
OF
13/1
93/
444
/143
385.
3349
5.5
0.16
50.
131
0.14
50.
147
0.01
70
00.
005
0.00
10.
003
0.00
2cy
top
lasm
/n
ucl
eus
10E
lon
gati
on
fact
or
1δ
Q68
FG
5M
ALD
IM
S-M
S/E
SIQ
-TO
F22
/95/
313
9/56
736.
0834
5.15
0.14
00.
145
0.10
20.
129
0.02
30.
058
0.06
60.
049
0.05
80.
008
0.18
0cy
top
lasm
1140
Sri
bo
som
alp
rote
inA
SP
1420
6M
ALD
IM
S-M
S/E
SIQ
-TO
F23
/22
5/5
152/
7933
4.80
4.9
420.
221
0.17
90.
208
0.20
30.
021
0.27
30.
051
0.12
30.
149
0.11
30.
211
pla
sma
mem
bra
ne/
cyto
pla
sm/
nu
cleu
s12
AT
Psy
nth
ase.
�P
5648
0M
ALD
IM
S-M
S/E
SIQ
-TO
F41
/31
12/1
561
3/47
556
5.19
475.
110.
199
0.44
60.
048
0.23
10.
200
0.08
80.
289
0.03
50.
137
0.13
30.
077
mit
och
on
dri
a
13H
SP5
P20
029
ESI
Q-T
OF
20/2
78/
1456
9/16
372
5.07
654.
510.
088
0.11
90.
080
0.09
50.
020
00
0.00
60.
002
0.00
30.
007
end
op
lasm
atic
reti
culu
m14
Rb
Ap
48Q
6097
2M
ALD
IM
S-M
S/E
SIQ
-TO
F13
/20
4/5
125/
144
524.
9554
4.57
0.15
80.
228
0.38
10.
255
0.11
40
00.
0380
0.01
270.
022
0.01
nu
cleu
s
aSp
ots
loca
ted
inF
igu
re5
are
ind
icat
edto
geth
erw
ith
Un
ipro
tac
cess
ion
nu
mb
er,
met
ho
du
sed
for
spo
tid
enti
fica
tio
n,
%o
fse
qu
ence
cove
red
by
pep
tid
es,
nu
mb
ero
fp
epti
des
iden
tifi
ed,
Mas
cot
sco
re,
theo
reti
cal
and
exp
erim
enta
lm
ole
cula
rw
eigh
t(M
W)
and
iso
elet
ric
po
int
(pI)
,%
volu
me
of
con
tro
lan
dtr
eate
dsa
mp
les
(ave
rage
of
thre
eex
per
imen
ts),
stan
dar
der
ror,
and
p-v
alu
esas
det
erm
ined
by
Wh
itn
eyt-
test
wh
ere
*P<
0.05
are
ind
icat
ed.
Role of Nuclear �ΙPKC in Transcription Processes of Murine ESCs research articles
Journal of Proteome Research • Vol. 9, No. 12, 2010 6199
http://www.phosphosite.org/, mostly detected using massspectrometry (Table 3). Of the 13 nuclear proteins, we identifiedfive of them (hnRNP C1/C2, nucleophosmin 1, SRP20, �-actin,and lamin B1) which had reported phosphorylations thatmatched the predicted PKC phosphorylation sites, and of these,lamin B1 has been confirmed to be a �ΙPKC substrate.37-39
Interestingly, hnRNP K was found to be phosphorylated byPKCδ40 at a site different than the ones predicted by Scansite(Table 3); therefore, we cannot discard the possibility that theother nuclear proteins may also be directly phosphorylated by�ΙPKC. Among these nuclear proteins whose phosphorylationwas modulated by cPKC peptides or by �ΙV5-3, we foundtranscription and splicing factors as well as proteins involvedin chromatin remodeling, proteins that are mainly involved in
regulation of proliferation and differentiation processes (Tables1 and 2). hnRNP K is involved in the transcriptional activationof c-myc (a pro-oncogene pivotal in controlling cell prolifera-tion and differentiation).41 The actin-like protein 6a (Baf53a),together with �-actin, is critical for the oncogenic activity ofc-myc.42 Differential phosphorylation of hnRNP C during thecell cycle regulates protein expression at distinct stages of thecell cycle.43 40S ribosomal protein AS (Laminin-binding proteinprecursor p40) associates with histone44 and Histone-bindingprotein RBBP4 (RbAp48) associates with histone deacetylaseplaying a role in chromatin remodeling during the cell cycle.45
Lamin B1 is an important constituent of the nuclear envelopeand nuclear disassembly during the cell cycle,46 and is essentialfor RNA synthesis processes.47 Lamin B1 when associated with
Figure 6. Phosphorylation of �-actin decreases upon treatement with �ΙV5-3. E14TG2A cells were treated with 500 nM of TAT47-57
carrier peptide and �IV5-3 for 10 min. 2DE gels were stained with Pro-Q Diamond and Coomassie blue (A) or transferred to nitrocelluloseand probed for �-actin (C). Cropped regions of 2DE gels containing the differentially phosphorylated spot (circled) identified as �-actinare indicated in both Comassie Blue and Pro-Q Diamond stained gels (A). Quantitative analysis of the normalized average of threeindependent samples comparing the % volume of phosphorylated protein in TAT47-57 carrier peptide and �IV5-3 treated samples. (B)Western blot of 2DE gels specifically probed for �-actin representing the differential migration of �-actin in lysates of cells treatedeither with TAT47-57 carrier peptide or �ΙV5-3. Quantitative representation of duplicate experiments demonstrating the percent volumeof each immunoreactive spot relative to the volume of all spots detected with anti-�-actin in TAT47-57 carrier peptide and �IV5-3 treatedsamples (C).
research articles Costa-Junior et al.
6200 Journal of Proteome Research • Vol. 9, No. 12, 2010

chromatin helps preserve nucleoli integrity.48 Nucleophosminhas been shown to be involved in centrosome duplication.49
Localization of SRP20 is also cell-cycle-regulated playing a rolein the control of proliferation.50 In Caenorhabditis elegans SRPprotein phosphorylation is important for embryonic develop-ment.51 Importantly, morula stage murine embryos lackingSRP20 do not develop to blastocysts.52 Sae1 is a key regulatorof PML body [nuclear foci containing the promyelocyticleukemia protein, and plays a role in transcriptional suppres-sion integrity].53 The pluripotency related gene Oct-4 has beenshown to be associated to PML bodies and phosphorylationand sumoylation of specific proteins have been demonstratedto be important for the recruitment of proteins that suppressOct 4 expression to PML bodies.54
In summary, the cPKC and �ΙPKC nuclear targets foundfunctionally are related to cell cycle and nuclear remodelingduring mitosis and proteins that regulate nuclear transcriptionprocesses, related to proliferation and cellular differentiationsuggesting a role for �ΙPKC in these processes.
�ΙV5-3 Does Not Affect ESC Proliferation. To determinewhether �ΙPKC inhibition in undifferentiated ESCs affects theirproliferation, we treated cells with �ΙV5-3 as well as with thecPKC inhibitor peptide �C2-4 and determined DNA synthesis.No effect on DNA synthesis as measured by [3H] thymidineincorporation was observed in cells treated with either �ΙV5-3or �C2-4 as compared to control ΤΑΤ47-57 carrier peptide(Supplemental Figure S13), indicating that inhibition of cPKCs
Figure 7. Phosphorylation of hnRNP K decreases upon treatement with �ΙV5-3. E14TG2A cells were treated with 500 nM of TAT47-57
carrier peptide and �IV5-3 for 10 min. 2DE gels were stained with Pro-Q Diamond and Coomassie blue (A) or transferred to nitrocelluloseand probed for hnRNP K (C). Cropped regions of 2DE gels containing the differentially phosphorylated spots (circled) identified ashnRNP K are indicated in both Comassie Blue and Pro-Q Diamond stained gels (A). Quantitative analysis of the normalized average ofthree independent samples comparing the % volume of phosphorylated protein in TAT47-57 carrier peptide and �IV5-3 treated samplesstatistical significance was determined by the Mann-Whitney t test where * p < 0.05 (B). Western blot of 2DE gels specifically probedfor hnRNP K representing the differential migration of hnRNP K in lysates of cells treated either with TAT47-57 carrier peptide or �ΙV5-3.Quantitative representation of duplicate experiments demonstrating the percent volume of each immunoreactive spot relative to thevolume of all spots detected with anti-hnRNP K in TAT47-57 carrier peptide and �IV5-3 treated samples (C).
Role of Nuclear �ΙPKC in Transcription Processes of Murine ESCs research articles
Journal of Proteome Research • Vol. 9, No. 12, 2010 6201

Table 3. Predicted PKC Phosphorylation Sites of the Nuclear Proteins That Are Differentially Phosphorylated upon Treatment withcPKC or �ΙPKC Modulator Peptides
protein species predicted p-site sequence predicted PKC isozymea validated sitesb
RbAp48 mouse/human T28 YKIWKKNTPFLYDLV vPKCε
mouse/human T221 GKVVDAKTIFTGHTA PKCδ/PKCε
mouse/human T338 TILASSGTDRRLNVW PKCR
hnRNP C1/C2 mouse/human S171 NTSRRGKSGFNSKSG PKCR
mouse S182 SKSGQRGSSSKSGKL PKCR/PKCδ
human S182 SKSGQRGSSKSGKLK PKCR
human S185 GQRGSSKSGKLKGDD PKCR
mouse S186 QRGSSSKSGKLKGDD PKCR
human S222 EKIEKEQSKQAVEMK PKCε
human S241 EEEQSSSSVKKDETN PKCR yesmouse S249 EEEQSSASVKKDETN PKCR/PKCε yesmouse/human T165 RQRVSGNTSRRGKSG PKCR
SAE1 human S86 QFLIRTGSVGRNRAE PKCR
mouse/human S99/S95 GRNRAEASLERAQNL PKCR
human S121 DIEKKPESFFTQFDA PKCε/PKCδ
mouse S125 DVEKKPESFFTKFDA PKC/PKCδ
human S150 DQICHKNSIKFFTGD PKCδ/PKCR/PKC\?\emouse S154 DQICHRNSIKFFTGD PKCδ/PKCR/PKCε
mouse S205 AKRAKLDSSETTMVK PKCδ
human S238 AALKRTTSDYFLLQV PKCδ
human T205 KLDSSETTMVKKKVV PKCR
mouse T209 KLDSSETTMVKKKVL PKCR
human T236 AKAALKRTTSDYFLL PKCδ
human T237 KAALKRTTSDYFLLQ PKCδ
nucleophosmin 1 mouse/human S48 QLSLRTVSLGAGAKD PKCδ
mouse S194 EKVPVKKSVRDTPAK PKCε yeshuman S195 EKAPVKKSIRDTPAK PKCR/PKCε yeshuman S218 GKDSKPSSTPRSKGQ PKCR yesmouse/human S225/S227 PRSKGQESFKKQEKT PKCε/PKCδ/PKCR
mouse/human S252/S254 IKAKMQASIEKGGSL PKCR/PKC\?\e/PKCδ yesmouse/human S258/S260 ASIEKGGSLPKVEAK PKCδ yesmouse/human S291/S293 DLWQWRKSL PKCε
human S293 PIKVTLATLKMSVQP PKCε
mouse/human T78 LKMSVQPTVSLGGFE PKCR yesmouse/human T86 GKDLKPSTPRSKGQE PKCδ
mouse T217 KDSKPSSTPRSKGQE PKCR yeshuman T219 SFKKQEKTPKTPKGP PKCR yesmouse/human T232/T234 SFKKQEKTPKTPKGP PKCR yes
hnRNP K mouse/human S420 IRHESGASIKIDEPL PKCR
mouse/human T9 ETEQPEETFPNTETN PKCε
mouse/human T177 LRENTQTTIKLFQEC PKCR/PKCδ
hnRNP F mouse S237 RMRSGAYSAGYGGYE PKCδ
human S237 RMRPGAYSTGYGGYE PKCδ
human T35 QNFLSDCTIHDGAAG PKCε
SRP20 mouse/human S115 SPPPRRRSPRRRSFS PKCR
mouse/human S120 RRSPRRRSFSRSRSR PKCR/PKCδ
mouse/human S122 SPRRRSFSRSRSRSL PKCR
mouse/human S124 RRRSFSRSRSRSLSR PKCR yesmouse/human S128 FSRSRSRSLSRDRRR PKCR/PKCδ yesmouse/human S130 RSRSRSLSRDRRRER PKCR
mouse/human S138 RDRRRERSLSRERNH PKCR/PKCδ PKCε yesmouse/human S150 RNHKPSRSFSRSRSR PKCR yesmouse/human S154 PSRSFSRSRSRSRSN PKCR yesmouse/human S156 RSFSRSRSRSRSNER PKCR
eIF-3-ε human S212 SLQNGRMSIKAYVST PKCε
40S ribosomal protein AS mouse/human T107 AGRFTPGTFTNQIQA PKCδ
mouse/human T188 EVLRMRGTISREHPW PKCR/PKCε
mouse/human T285 AAPTAQATEWVGATT PKCδ
b actin mouse/human S235 ATAASSSSLEKSYEL PKCδ
mouse/human T202 ERGYSFTTTAEREIV PKCε yesmouse/human T297 RKDLYANTVLSGGTT PKCδ
mouse/human T303 NTVLSGGTTMYPGIA PKCδ
lamin B1 human S210 EDLEFRKSMYEEEIN PKCε
mouse S405/S404 RVTVSRASSSRSVRT PKCR yesmouse S406/S405 VTVSRASSSRSVRTT PKCR yes
research articles Costa-Junior et al.
6202 Journal of Proteome Research • Vol. 9, No. 12, 2010

does not affect proliferation in standard conditions thatmaintain ESCs undifferentiated.
Expression and Subcellular Localization of �ΙPKC duringDifferentiation. To gain further insight on the role of �ΙPKCin undifferentiated ESCs, we looked at �ΙPKC expression inundifferentiated and differentiated ESCs. In most undifferenti-ated, Oct 3/4-positive ESCs, �ΙPKC was found in the nucleus,while in a few of these cells, �ΙPKC was associated to filamen-tous structures (Figure 8A). In contrast, in the Oct 3/4-negativecell counterpart, here assumed to represent differentiated cells,�ΙPKC was excluded from the nucleus and found eitherthroughout the cell, associated mostly to filamentous struc-tures, or in the cytoplasm (Figure 8B).
To determine whether the nuclear pattern of �ΙPKC expres-sion is generally linked to undifferentiated cells, or if it is aparticular feature of E14TG2A growing in two-dimensionallayers, we verified the localization of �ΙPKC in tridimensionallygrowing embryoid bodies (EBs). At 6 days of differentiation,the expression of the embryonic stem cell marker Oct 4, asdetermined by real-time PCR, is 1/10 of the expression ob-served in undifferentiated ESCs (day 0), and at 8 days, expres-sion falls further, reaching a plateau that is maintained untilthe 12th day of differentiation (Figure 9A). In Figure 9, we showthat after 7 days of culture in differentiation medium, �ΙPKCexpression almost completely vanished from the nucleus. Inapproximately half of the cells, �ΙPKC was still expressed in
Table 3. Continued
protein species predicted p-site sequence predicted PKC isozymea validated sitesb
mouse S409/S408 SRASSSRSVRTTRGK PKCR yesmouse T400/T399 PSPSSRVTVSRASSS PKCε/PKCR
mouse T412/T411 SSSRSVRTTRGKRKR PKCR
mouse T413/T412 SSRSVRTTRGKRKRV PKCR
mouse T496 YVLKAGQTVTVWAAN PKCε
human T544 EEVAQRSTVFKTTIP PKCδ
mouse T545 EEVAQRSTVFKTTIP PKCε
humana T575 EEVAQRSTVFKTTIP PKCδ
mouse T550/T549 RSTVFKTTIPEEEEE PKCR/PKCε
Baf53a mouse/human S116 MHVKSEASLHPVLMS PKCδ
mouse/human S375 LSQKTPPSMRLKLIA PKCR
proteasome subunit beta type-4 mouse/human S77 DMLGSYGSLARFRNI PKCR
mouse/human T236 YNRFQIATVTEKGVE PKCε/PKCδ
a Predicted by Scansite (http://scansite.mit.edu). b Validated sites reported in Phosphosite (http://www.phosphosite.org).
Figure 8. Confocal microscopy images of �ΙPKC localization in Oct 3/4 positive and negative cells. E14TG2A cells ∼80% confluent werecultured in conditions that maintained more than 90% of the cells undifferentiated. Cells were fixed and stained for Oct 3/4 (red),�ΙPKC (green), and nuclei labeled with DAPI as described in the Experimental Section. Merged images of Oct 3/4 and �ΙPKC (A lowerpanel left) and merged images of Oct 3/4 and �ΙPKC and labeled nuclei (A lower panel right). Higher magnification images of Oct 3/4positive and negative cells (white arrows) are shown in panel B.
Role of Nuclear �ΙPKC in Transcription Processes of Murine ESCs research articles
Journal of Proteome Research • Vol. 9, No. 12, 2010 6203

the cytoplasm, while in the other half, �ΙPKC was no longervisualized by immunofluorescence. In summary, these resultsdemonstrate that E14TG2A ESCs display an opposite relation-ship between differentiation and nuclear �ΙPKC expression, andthe pattern of strong nuclear expression of �ΙPKC in undif-ferentiated ESCs appears to be related to nuclear transcriptionprocesses predominant under these conditions.
To investigate the role of �ΙPKC in ESCs, we used aproteomics strategy combining cPKC modulator peptides, a�ΙPKC-specific inhibitor peptide, �ΙV5-3, and a phospho-specific dye. Using this strategy, we identified five nuclearproteins whose phosphorylation status was modulated by eithercPKC activator or inhibitor peptides, and eight proteins whosephosphorylation decreased in the presence of the �ΙPKCspecific inhibitor peptide. We cannot completely discard thepossibility that some of the nuclear proteins identified are also�ΙΙPKC substrates, since �ΙΙPKC was also concentrated withina dot in the nucleus. Of the proteins identified, hnRNP K,40
nucleophosmin 1,55 and lamin B139 have been previously
described as PKC substrates, confirming that our strategy wasefficient at capturing �ΙPKC substrates, and hnRNP C and�-actin display putative PKC phosphorylation. Further char-acterization of the identified substrates will determine if theyare direct or indirect �ΙPKC substrates. Nevertheless, theproteins identified herewith have been previously shown to beinvolved in proliferation and/or differentiation processes, whichis consistent with the patterns of subcellular localization thatwe describe in undifferentiated cells. We have also identifiedtranscription regulators, including splicing modulators such asSRP20 and hnRNPs and chromatin remodeling proteins likeRbAp48 and Baf53a. In the nucleus, we found �ΙPKC concen-trated in spots characteristic of splicing complexes,56,57 sup-porting the possible involvement of �ΙPKC in splicing.
Among the changes observed during early events of dif-ferentiation, there is an increase in protein synthesis andtranslation processes.58 Therefore, regulation of translation isan important process for the maintenance of ESC as pluripotentundifferentiated cells and for ESC differentiation. Among the
Figure 9. �ΙPKC localization in embryoid bodies differentiated for 7 days. E14TG2A cells were differentiated by the hanging drop methoddescribed above. Oct4 relative expression was analyzed at different days of differentiation by real-time RT-PCR (A), average of 3independent experiments where *p < 0.05 is statistically different relative to Oct 4 expression at day O (as determined by theMann-Whitney t test). Confocal microscopy of embryoid bodies at 7 days of differentiation (B), higher magnification (C), where �ΙPKCis labeled in green and nuclei in blue.
research articles Costa-Junior et al.
6204 Journal of Proteome Research • Vol. 9, No. 12, 2010

PKC substrates identified, hnRNP K,41 �-actin, Baf 54a,42 andSRP2052 have previously been shown to be important inregulating transcription during differentiation processes. Phos-phorylation has been demonstrated to modulate the activityand subcellular localization of transcription factors, RNAbinding proteins, and splicing regulators such as hnRNPs59,60
and translation initiation factor 3 subunit F.61 Therefore, it ispossible that �ΙPKC phosphorylates and modulates the activityof key regulators of protein translation at the transition ofbetween an undifferentiated ESC and an ESC that is justreceiving its first clues toward a differentiated phenotype (i.e.,a specified cell).62 Supporting this idea, herewith we show, inthe same culture dish, that undifferentiated (Oct 3/4 positive)ESCs express nuclear �ΙPKC, while differentiated cells (Oct 3/4negative) express only cytoplasmic �ΙPKC or not at all. Thisindicates that the nuclear localization of �ΙPKC in undifferenti-ated cells is not due to the serum or growth factors added intothe media but, rather, to the intrinsic gene regulatory programdisplayed by undifferentiated ESCs.
Protein phosphorylation is key in ESC signaling, and inhibi-tion of specific kinases such as GSK3 and MAPK has beenshown to be able to maintain ESC pluripotency.1 Recently,there have been two studies aimed at identifying very earlyphosphorylation events leading to human ESC differentiation.2,3
These studies revealed that most of the proteins phosphoryl-ated in early specification events leading to differentiation areinvolved in the regulation of protein synthesis and translation,nucleic acid binding proteins and transcription factors. Despitethe fact that there are differences between human and murineembryonic stem cells, key signaling pathways and cellularreprogramming of differentiated somatic cells do require thesame set of conserved factors.63,64 It is interesting to point outthat proteins detected in the present study as �ΙPKC targets(direct or indirect) such as heterogeneous nuclear ribonucle-oprotein C1/C2 and K, nucleophosmin 1, elongation factor 1beta (Eef1�), lamin B1 and 40S ribosomal protein AS (Laminin-binding protein precursor p40) have also been shown to bemodulated during differentiation in the two studies mentionedabove.2,3 Since differentiation of EBs is a highly heterogeneousprocess, we did not expect to observe homogeneous patternsof �ΙPKC expression and localization. What we have describedwas a clear trend for cells that have entered the differentiationpath to reverse the undifferentiated mode of �ΙPKC nuclearexpression toward a cytoplasmatic pattern or to shut down�ΙPKC expression altogether. It remains to be shown whetherdifferentiation leads to changes in �ΙPKC expression andsubcellular localization, or whether changes in �ΙPKC expres-sion and localization contribute to ESC differentiation.
In summary, our strategy proved to be effective in detectingin vivo direct and indirect �IPKC isoenzyme specific substratesand our proteomics data and subcellular localization studiessuggest that nuclear �ΙPKC is involved in the maintenance ofthe undifferentiated state in ESCs.
Acknowledgment. We thank Drs. Daria Mochly-Rosen,Alice Vallentin, Frederico Gueiros Filho, Ricardo Giordano,Emer Ferro and Marcelo Damario Gomes for valuablediscussions. H.M.C.-J. was a recipient of a FAPESP postdoctoralfellowship (2006/52062-6). This research was supported byBrazilian agencies grants FAPESP (2005/54188-4) and CNPQ(479673/2007-1) to D.S., FAPERJ and FIOCRUZ to J.E.A.P.
Supporting Information Available: Cropped regionsof 2DE gels containing the differentially phosphorylated spots;
evidence that inhibition of cPKCs with �C2-4 or of �ΙPKC with�ΙV5-3 does not affect ESC proliferation. This material isavailable free of charge via the Internet at http://pubs.acs.org.
References(1) Silva, J.; Barrandon, O.; Nichols, J.; Kawaguchi, J.; Theunissen,
T. W.; Smith, A. Promotion of reprogramming to ground statepluripotency by signal inhibition. PLoS Biol. 2008, 6 (10), e253.
(2) Brill, L. M.; Xiong, W.; Lee, K. B.; Ficarro, S. B.; Crain, A.; Xu, Y.;Terskikh, A.; Snyder, E. Y.; Ding, S. Phosphoproteomic analysis ofhuman embryonic stem cells. Cell Stem Cell 2009, 5 (2), 204–13.
(3) Van Hoof, D.; Munoz, J.; Braam, S. R.; Pinkse, M. W.; Linding, R.;Heck, A. J.; Mummery, C. L.; Krijgsveld, J. Phosphorylationdynamics during early differentiation of human embryonic stemcells. Cell Stem Cell 2009, 5 (2), 214–26.
(4) Quinlan, L. R.; Faherty, S.; Kane, M. T. Phospholipase C and proteinkinase C involvement in mouse embryonic stem-cell proliferationand apoptosis. Reproduction 2003, 126 (1), 121–31.
(5) Han, H. J.; Han, J. Y.; Heo, J. S.; Lee, S. H.; Lee, M. Y.; Kim, Y. H.ANG II-stimulated DNA synthesis is mediated by ANG II receptor-dependent Ca(2+)/PKC as well as EGF receptor-dependent PI3K/Akt/mTOR/p70S6K1 signal pathways in mouse embryonic stemcells. J Cell Physiol 2007, 211 (3), 618–29.
(6) Heo, J. S.; Han, H. J. ATP stimulates mouse embryonic stem cellproliferation via protein kinase C, phosphatidylinositol 3-kinase/Akt, and mitogen-activated protein kinase signaling pathways.Stem Cells 2006, 24( (12), 2637–48.
(7) Ventura, C.; Zinellu, E.; Maninchedda, E.; Fadda, M.; Maioli, M.Protein kinase C signaling transduces endorphin-primed cardio-genesis in GTR1 embryonic stem cells. Circ. Res. 2003, 92 (6), 617–22.
(8) Zhou, X.; Quann, E.; Gallicano, G. I. Differentiation of nonbeatingembryonic stem cells into beating cardiomyocytes is dependenton downregulation of PKC beta and zeta in concert with upregu-lation of PKC epsilon. Dev. Biol. 2003, 255 (2), 407–22.
(9) Mochly-Rosen, D.; Kauvar, L. M. Pharmacological regulation ofnetwork kinetics by protein kinase C localization. Semin. Immunol.2000, 12 (1), 55–61.
(10) Schechtman, D.; Mochly-Rosen, D. Adaptor proteins in proteinkinase C-mediated signal transduction. Oncogene 2001, 20 (44),6339–47.
(11) Schechtman, D.; Mochly-Rosen, D. Isozyme-specific inhibitors andactivators of protein kinase C. Methods Enzymol. 2002, 345, 470–89.
(12) Nishizuka, Y. The heterogeneity and differential expression ofmultiple species of the protein kinase C family. Biofactors 1988, 1(1), 17–20.
(13) Siwko, S.; Mochly-Rosen, D. Use of a novel method to findsubstrates of protein kinase C delta identifies M2 pyruvate kinase.Int. J. Biochem. Cell Biol. 2007, 39 (5), 978–87.
(14) Chen, C. H.; Budas, G. R.; Churchill, E. N.; Disatnik, M. H.; Hurley,T. D.; Mochly-Rosen, D. Activation of aldehyde dehydrogenase-2reduces ischemic damage to the heart. Science 2008, 321 (5895),1493–5.
(15) Rodriguez, M. M.; Ron, D.; Touhara, K.; Chen, C. H.; Mochly-Rosen,D. RACK1, a protein kinase C anchoring protein, coordinates thebinding of activated protein kinase C and select pleckstrin homol-ogy domains in vitro. Biochemistry 1999, 38 (42), 13787–94.
(16) Stebbins, E. G.; Mochly-Rosen, D.; Binding specificity for RACK1resides in the V5 region of beta II protein kinase, C. J. Biol. Chem.2001, 276 (32), 29644–50.
(17) Vallentin, A.; Mochly-Rosen, D. RBCK1, a protein kinase CbetaI(PKCbetaI)-interacting protein, regulates PKCbeta-dependent func-tion. J. Biol. Chem. 2007, 282 (3), 1650–7.
(18) Nagahara, H.; Vocero-Akbani, A. M.; Snyder, E. L.; Ho, A.; Latham,D. G.; Lissy, N. A.; Becker-Hapak, M.; Ezhevsky, S. A.; Dowdy, S. F.Transduction of full-length TAT fusion proteins into mammaliancells: TAT-p27Kip1 induces cell migration. Nat. Med. 1998, 4 (12),1449–52.
(19) Morrison, G. M.; Brickman, J. M. Conserved roles for Oct4homologues in maintaining multipotency during early vertebratedevelopment. Development 2006, 133 (10), 2011–22.
(20) Viswanathan, S.; Benatar, T.; Rose-John, S.; Lauffenburger, D. A.;Zandstra, P. W. Ligand/receptor signaling threshold (LIST) modelaccounts for gp130-mediated embryonic stem cell self-renewalresponses to LIF and HIL-6. Stem Cells 2002, 20 (2), 119–38.
(21) Viswanathan, S.; Benatar, T.; Mileikovsky, M.; Lauffenburger, D. A.;Nagy, A.; Zandstra, P. W. Supplementation-dependent differencesin the rates of embryonic stem cell self-renewal, differentiation,and apoptosis. Biotechnol. Bioeng. 2003, 84 (5), 505–17.
Role of Nuclear �ΙPKC in Transcription Processes of Murine ESCs research articles
Journal of Proteome Research • Vol. 9, No. 12, 2010 6205

(22) Sukoyan, M. A.; Kerkis, A. Y.; Mello, M. R.; Kerkis, I. E.; Visintin,J. A.; Pereira, L. V. Establishment of new murine embryonic stemcell lines for the generation of mouse models of human geneticdiseases. Braz. J. Med. Biol. Res. 2002, 35 (5), 535–42.
(23) Boheler, K. R.; Czyz, J.; Tweedie, D.; Yang, H. T.; Anisimov, S. V.;Wobus, A. M. Differentiation of pluripotent embryonic stem cellsinto cardiomyocytes. Circ. Res. 2002, 91 (3), 189–201.
(24) Pereira, A. C.; Miyakawa, A. A.; Lopes, N. H.; Soares, P. R.; deOliveira, S. A.; Cesar, L. A.; Ramires, J. F.; Hueb, W.; Krieger, J. E.Dynamic regulation of MTHFR mRNA expression and C677Tgenotype modulate mortality in coronary artery disease patientsafter revascularization. Thromb. Res. 2007, 121 (1), 25–32.
(25) Towbin, H.; Staehelin, T.; Gordon, J. Immunoblotting in the clinicallaboratory. J. Clin. Chem. Clin. Biochem. 1989, 27 (8), 495–501.
(26) Candiano, G.; Bruschi, M.; Musante, L.; Santucci, L.; Ghiggeri,G. M.; Carnemolla, B.; Orecchia, P.; Zardi, L.; Righetti, P. G. Bluesilver: a very sensitive colloidal Coomassie G-250 staining forproteome analysis. Electrophoresis 2004, 25 (9), 1327–33.
(27) Fiorani Celedon, P. A.; de Andrade, A.; Meireles, K. G.; Gallo deCarvalho, M. C.; Caldas, D. G.; Moon, D. H.; Carneiro, R. T.;Franceschini, L. M.; Oda, S.; Labate, C. A. Proteomic analysis ofthe cambial region in juvenile Eucalyptus grandis at three ages.Proteomics 2007, 7 (13), 2258–74.
(28) Andrade, H. M.; Murta, S. M.; Chapeaurouge, A.; Perales, J.; Nirde,P.; Romanha, A. J. Proteomic analysis of Trypanosoma cruziresistance to Benznidazole. J. Proteome Res. 2008, 7 (6), 2357–67.
(29) Braun, M. U.; Mochly-Rosen, D. Opposing effects of delta- andzeta-protein kinase C isozymes on cardiac fibroblast proliferation:use of isozyme-selective inhibitors. J. Mol. Cell. Cardiol. 2003, 35(8), 895–903.
(30) Aaltonen, V.; Koivunen, J.; Laato, M.; Peltonen, J. Heterogeneityof cellular proliferation within transitional cell carcinoma: cor-relation of protein kinase C alpha/betaI expression and activity.J. Histochem. Cytochem. 2006, 54 (7), 795–806.
(31) al-Mazidi, H. A.; Kleine, L. P.; Franks, D. J. The presence of anunusual PKC isozyme profile in rat liver cells. Biochem. Cell Biol.1998, 76 (1), 73–82.
(32) Pfaff, I. L.; Wagner, H. J.; Vallon, V. Immunolocalization of proteinkinase C isoenzymes alpha, beta1 and betaII in rat kidney. J. Am.Soc. Nephrol. 1999, 10 (9), 1861–73.
(33) Dehghani, H.; Hahnel, A. C. Expression profile of protein kinaseC isozymes in preimplantation mouse development. Reproduction2005, 130 (4), 441–51.
(34) Chen, D.; Purohit, A.; Halilovic, E.; Doxsey, S. J.; Newton, A. C.Centrosomal anchoring of protein kinase C betaII by pericentrincontrols microtubule organization, spindle function, and cytoki-nesis. J. Biol. Chem. 2004, 279 (6), 4829–39.
(35) Kraft, A. S.; Anderson, W. B. Phorbol esters increase the amountof Ca2+, phospholipid-dependent protein kinase associated withplasma membrane. Nature 1983, 301 (5901), 621–3.
(36) DeVries, T. A.; Neville, M. C.; Reyland, M. E. Nuclear import ofPKCdelta is required for apoptosis: identification of a novel nuclearimport sequence. EMBO J. 2002, 21 (22), 6050–60.
(37) Fiume, R.; Ramazzotti, G.; Teti, G.; Chiarini, F.; Faenza, I.; Mazzotti,G.; Billi, A. M.; Cocco, L. Involvement of nuclear PLCbeta1 in laminB1 phosphorylation and G2/M cell cycle progression. FASEB J.2009, 23 (3), 957–66.
(38) Goss, V. L.; Hocevar, B. A.; Thompson, L. J.; Stratton, C. A.; Burns,D. J.; Fields, A. P. Identification of nuclear beta II protein kinaseC as a mitotic lamin kinase. J. Biol. Chem. 1994, 269 (29), 19074–80.
(39) Hocevar, B. A.; Burns, D. J.; Fields, A. P. Identification of proteinkinase C (PKC) phosphorylation sites on human lamin B. Potentialrole of PKC in nuclear lamina structural dynamics. J. Biol. Chem.1993, 268 (10), 7545–52.
(40) Schullery, D. S.; Ostrowski, J.; Denisenko, O. N.; Stempka, L.;Shnyreva, M.; Suzuki, H.; Gschwendt, M.; Bomsztyk, K. Regulatedinteraction of protein kinase Cdelta with the heterogeneousnuclear ribonucleoprotein K protein. J. Biol. Chem. 1999, 274 (21),15101–9.
(41) Baber, J. L.; Libutti, D.; Levens, D.; Tjandra, N. High precisionsolution structure of the C-terminal KH domain of heterogeneousnuclear ribonucleoprotein K, a c-myc transcription factor. J. Mol.Biol. 1999, 289 (4), 949–62.
(42) Park, J.; Wood, M. A.; Cole, M. D. BAF53 forms distinct nuclearcomplexes and functions as a critical c-Myc-interacting nuclearcofactor for oncogenic transformation. Mol. Cell. Biol. 2002, 22(5), 1307–16.
(43) Pinol-Roma, S.; Dreyfuss, G. Cell cycle-regulated phosphorylationof the pre-mRNA-binding (heterogeneous nuclear ribonucleopro-tein) C proteins. Mol. Cell. Biol. 1993, 13 (9), 5762–70.
(44) Kinoshita, K.; Kaneda, Y.; Sato, M.; Saeki, Y.; Wataya-Kaneda, M.;Hoffmann, A. LBP-p40 binds DNA tightly through associations withhistones H2A, H2B, and H4. Biochem. Biophys. Res. Commun.1998, 253 (2), 277–82.
(45) Nicolas, E.; Morales, V.; Magnaghi-Jaulin, L.; Harel-Bellan, A.;Richard-Foy, H.; Trouche, D. RbAp48 belongs to the histonedeacetylase complex that associates with the retinoblastomaprotein. J. Biol. Chem. 2000, 275 (13), 9797–804.
(46) Vergnes, L.; Peterfy, M.; Bergo, M. O.; Young, S. G.; Reue, K. LaminB1 is required for mouse development and nuclear integrity. Proc.Natl. Acad. Sci. U.S.A. 2004, 101 (28), 10428–33.
(47) Tang, C. W.; Maya-Mendoza, A.; Martin, C.; Zeng, K.; Chen, S.;Feret, D.; Wilson, S. A.; Jackson, D. A. The integrity of a lamin-B1-dependent nucleoskeleton is a fundamental determinant ofRNA synthesis in human cells. J. Cell Sci. 2008, 121 (Pt. 7), 1014–24.
(48) Martin, C.; Chen, S.; Maya-Mendoza, A.; Lovric, J.; Sims, P. F.;Jackson, D. A. Lamin B1 maintains the functional plasticity ofnucleoli. J. Cell Sci. 2009, 122 (Pt. 10), 1551–62.
(49) Wang, W.; Budhu, A.; Forgues, M.; Wang, X. W. Temporal andspatial control of nucleophosmin by the Ran-Crm1 complex incentrosome duplication. Nat. Cell Biol. 2005, 7 (8), 823–30.
(50) Loomis, R. J.; Naoe, Y.; Parker, J. B.; Savic, V.; Bozovsky, M. R.;Macfarlan, T.; Manley, J. L.; Chakravarti, D. Chromatin binding ofSRp20 and ASF/SF2 and dissociation from mitotic chromosomesis modulated by histone H3 serine 10 phosphorylation. Mol. Cell2009, 33 (4), 450–61.
(51) Longman, D.; Johnstone, I. L.; Caceres, J. F. Functional charac-terization of SR and SR-related genes in Caenorhabditis elegans.EMBO J. 2000, 19 (7), 1625–37.
(52) Jumaa, H.; Wei, G.; Nielsen, P. J. Blastocyst formation is blockedin mouse embryos lacking the splicing factor SRp20. Curr. Biol.1999, 9 (16), 899–902.
(53) Daniels, M. J.; Marson, A.; Venkitaraman, A. R. PML bodies controlthe nuclear dynamics and function of the CHFR mitotic checkpointprotein. Nat. Struct. Mol. Biol. 2004, 11 (11), 1114–21.
(54) Park, S. W.; Hu, X.; Gupta, P.; Lin, Y. P.; Ha, S. G.; Wei, L. N.SUMOylation of Tr2 orphan receptor involves Pml and fine-tunesOct4 expression in stem cells. Nat. Struct. Mol. Biol. 2007, 14 (1),68–75.
(55) Beckmann, R.; Buchner, K.; Jungblut, P. R.; Eckerskorn, C.; Weise,C.; Hilbert, R.; Hucho, F. Nuclear substrates of protein kinase C.Eur. J. Biochem. 1992, 210 (1), 45–51.
(56) Gallouzi, I. E.; Brennan, C. M.; Stenberg, M. G.; Swanson, M. S.;Eversole, A.; Maizels, N.; Steitz, J. A. HuR binding to cytoplasmicmRNA is perturbed by heat shock. Proc. Natl. Acad. Sci. U.S.A.2000, 97 (7), 3073–8.
(57) Paul, S.; Dansithong, W.; Kim, D.; Rossi, J.; Webster, N. J.; Comai,L.; Reddy, S. Interaction of muscleblind, CUG-BP1 and hnRNP Hproteins in DM1-associated aberrant IR splicing. EMBO J. 2006,25 (18), 4271–83.
(58) Sampath, P.; Pritchard, D. K.; Pabon, L.; Reinecke, H.; Schwartz,S. M.; Morris, D. R.; Murry, C. E. A hierarchical network controlsprotein translation during murine embryonic stem cell self-renewaland differentiation. Cell Stem Cell 2008, 2 (5), 448–60.
(59) Habelhah, H.; Shah, K.; Huang, L.; Ostareck-Lederer, A.; Burlin-game, A. L.; Shokat, K. M.; Hentze, M. W.; Ronai, Z. ERKphosphorylation drives cytoplasmic accumulation of hnRNP-K andinhibition of mRNA translation. Nat. Cell Biol. 2001, 3 (3), 325–30.
(60) Quaresma, A. J.; Bressan, G. C.; Gava, L. M.; Lanza, D. C.; Ramos,C. H.; Kobarg, J. Human hnRNP Q re-localizes to cytoplasmicgranules upon PMA, thapsigargin, arsenite and heat-shock treat-ments. Exp. Cell Res. 2009, 315 (6), 968–80.
(61) Shi, J.; Hershey, J. W.; Nelson, M. A. Phosphorylation of theeukaryotic initiation factor 3f by cyclin-dependent kinase 11 duringapoptosis. FEBS Lett. 2009, 583 (6), 971–7.
(62) SlackJ. M. W. Essential Developmental Biology. 2nd ed.; BlackwellScience: Malden, MA, 2005; p 448.
(63) Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cellsfrom mouse embryonic and adult fibroblast cultures by definedfactors. Cell 2006, 126 (4), 663–76.
(64) Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.;Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells fromadult human fibroblasts by defined factors. Cell 2007, 131 (5),861–72.
PR100355K
research articles Costa-Junior et al.
6206 Journal of Proteome Research • Vol. 9, No. 12, 2010
Top Related
Copyright © 2022 FDOKUMEN

