







Bahasa
Halaman
Hukum


a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 4
ava i lab le a t www.sc iencedi rec t .com
journa l homepage : www.e lsev ie r . com/ loca te /ac toec
Original article
On the role of Posidonia oceanica beach wrack formacroinvertebrates of a Tyrrhenian sandy shore
Isabella Colombinia, Miguel Angel Mateob, Oscar Serranob, Mario Fallacia,Elena Gagnarlia, Laura Serranob, Lorenzo Chelazzia,*aIstituto per lo Studio degli Ecosistemi, CNR, Via Madonna del Piano 10, 50019 Sesto Fiorentino Firenze, ItalybCentro de Estudios Avanzados de Blanes Consejo Superior de Investigaciones Cientıficas, c/ Acces a la Cala St. Francesc,
14, 17300 - Blanes, Girona, Spain
a r t i c l e i n f o
Article history:
Received 2 May 2008
Accepted 17 July 2008
Published online 18 September 2008
Keywords:
Mediterranean sandy beach
Wrack colonisation
Species succession
Abundance
Multi-isotopic analysis
* Corresponding author. Tel.: þ39 055 522591E-mail address: [email protected] (L. Ch
1146-609X/$ – see front matter ª 2008 Elsevidoi:10.1016/j.actao.2008.07.005
a b s t r a c t
The use of Posidonia oceanica beach wrack by macroinvertebrates of the sandy beach at
Burano (Tuscany, Italy) was assessed by following the colonisation dynamics of the wrack
and analysing the stable isotopes ‘scenario’ of the main local carbon and nitrogen sources
and consumers. One-hundred experimental cylinders, filled with P. oceanica wrack, were
placed on the beach and sampled over a 1-month period. Abundance and species richness
of macroinvertebrates in wracks varied through time. Wrack was colonised by crustaceans
almost immediately after deployment of the experimental cylinders. The amphipod Tali-
trus saltator largely dominated the faunal assembly and, together with the isopod Tylos
europaeus, occupied the wracks closer to the sealine. These were followed by dipterans,
staphylinids, pselaphids and tenebrionids that occurred in drier wracks higher up on the
eulittoral. Moisture content of the wrack and sand decreased through space and time. This
was the primary factor explaining the spatial and temporal changes observed in macro-
invertebrate abundance, with species colonising or abandoning wracks according to
thresholds of environmental parameters. Isotopic analysis clearly established the absence
of any direct dietary link between P. oceanica wrack and macroinvertebrates. Terrestrial
food sources were also discarded. Both our experimental data and a literature search
showed that the organic matter from seston as filtered by the sand is the most plausible
carbon and nitrogen source for beach food webs. Even if P. oceanica wrack is not a trophic
source for macroinvertebrates, it is vitally important as a physical structure that provides
detritivorous and predatory species with refuge from environmentally stressful conditions.
ª 2008 Elsevier Masson SAS. All rights reserved.
1. Introduction the carbon synthesised by primary producers may not be
Understanding the functioning of coastal ecosystems needs
an accurate knowledge of materials and energy flow between
compartments (Mateo et al., 2006). An important fraction of
2; fax: þ39 055 5225920.elazzi).er Masson SAS. All rights
exploited locally but, instead, be transported far away to fuel
the food webs of other ecosystems. This is particularly true in
highly dynamic ecosystems such as marine coastal
environments.
reserved.

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 4 33
The sea–land interface is a complex area where terrestrial
and marine ecosystems interact intensely in several ways.
Sandy shores occupy approximately three-quarters of the
shorelines of the world (Bascom, 1980) and are known to have
very little in situ primary production (Inglis, 1989; McLachlan
and Brown, 2006). Marine allochthonous inputs are thus of
primary importance for beaches that receive substantial
carbon and nitrogen inputs from the sea in the form of macro-
and micro-detritus from various origins. Among stranded
macrophytes, seagrasses have an important role in controlling
coastal sedimentary processes (De Falco et al., 2003), in oper-
ating as important nutrient sinks for nearshore communities
(Ochieng and Erftemeijer, 1999; Mateo et al., 2003; McLachlan
and Brown, 2006), in influencing the geomorphology of sandy
shores by attenuating waves, reducing energy to beaches and
protecting from erosion (Hemminga and Nieuwenhuize, 1990),
in acting as refuges and in supporting prey resources that are
the basic element of food webs for vertebrate predators such as
lizards, shorebirds, foxes, coyotes etc. (Polis and Hurd, 1996;
Dugan et al., 2003; Colombini and Chelazzi, 2003).
There is a substantial literature on seagrass meadows of
Posidonia oceanica (L.) Delile, an endemic and dominant species
of the Mediterranean. This mainly focuses on seagrass growth
patterns, fate of production, dynamics of organic deposits,
leaf litter decomposition, carbon cycling and bacterial carbon
of sediments (Cebrian et al., 1997; Mateo et al., 1997; Marba
et al., 2002; Holmer et al., 2004). Recently, some attention has
been given to the quantities and functions of beach-cast
material (Colombini et al., 2000; Dugan et al., 2003; Jaramillo
et al., 2006; Olabarria et al., 2007) and in particular of P. oceanica
of Mediterranean areas (Mateo et al., 2003). The importance of
beach casts, their colonisation by macroinvertebrate species
and subsequent successional patterns was reviewed by
Colombini and Chelazzi (2003). With the exception of Inglis
(1989) and Olabarria et al. (2007), none of the other authors
have used experimentally deployed wrack patches to analyse
the dynamics of the associated fauna but have studied extant
ones. Moreover, information on spatial and temporal vari-
ability of macroinvertebrate fauna associated with the wrack
as affected by several environmental parameters obtained
during the same experiments is very poor. The stable isotopes
approach to analyse the trophic links between carbon sources
and consumers in ecosystems is nowadays a common prac-
tice. It assumes that the isotopic ratio of a certain carbon or
nitrogen source is transferred to its consumers undergoing
a predictable metabolic fractionation. This predictability
allows us to infer the theoretical diet of consumers by cor-
recting for this fractionation. While numerous isotopic studies
have focussed on the trophic pathways of infralittoral
communities associated with seagrass meadows (Vizzini and
Mazzola, 2003; Holmer et al., 2004; Hyndes and Lavery, 2005;
Carlier et al., 2007), those focussing on supralittoral commu-
nities of sandy beaches are quite rare (Adin and Riera, 2003;
Ince et al., 2007). Assessing the potential transference of
energy and matter from marine sub-tidal to supralittoral
zones is important because it can represent the basis for
sustaining the high biomass and diversity observed in sandy
beach ecosystems (Dugan et al., 2003; Ince et al., 2007).
In this paper we examine the possible role of P. oceanica
wrack as provision of shelter and/or as a trophic niche for the
macroinvertebrate fauna of a sandy beach. For this study the
beach adjacent to the Burano Lagoon (Tuscany, Italy) was
selected because of the conspicuous number of behavioural
and ecological studies that have been conducted there on
beach macrofauna (Scapini et al., 1992; Fallaci et al., 1996,
2002, 2003; Aloia et al., 1999; Colombini et al., 2005). The
colonisation dynamics of experimental wrack by the local
fauna was followed in time (33-day experiment) and space
(grid, sand and wrack compartments) as affected by key
environmental factors (moisture, pH, and conductivity).
Possible trophic links between P. oceanica wrack and macro-
invertebrates were assessed using an isotopic approach. Other
possible sources of carbon and nitrogen from both terrestrial
and marine environments were considered as well.
2. Materials and methods
2.1. Sampling site
The study site was located in a 30-km stretch of coast between
the Ansedonia promontory (southern Tuscany, Grosseto
province) and the Fiora River mouth (northern Latium, Viterbo
province). In this area the coast is characterised by
a morphologically homogeneous beach (oriented from north-
west to south-east) and is exposed (exposure rating ¼ 11.5
according to McLachlan, 1980). In front of this stretch of coast
there is a P. oceanica meadow of ca. 30 km2 at a mean depth of
18 m (Bianchi et al., 1993) (Fig. 1). Close to the promontory of
Ansedonia lies a protected area (WWF Italy, Burano Oasis)
where beach access to the public is controlled. For this reason
the study was conducted here and experiments were carried
out in an area far away (42�2305000N, 11�2204000E) from the beach
entrance. At this point the eulittoral zone included the part of
the beach devoid of vegetation, where beach wrack material
was stranded during heavy sea storms. The mean width of
this zone was 15 m, beach slope was 24.5% and its profile
varied according to wave action, sea currents and wind. Mean
grain size was 1.295 F according to the method of Folk and
Ward (1957). The supralittoral zone extended for 10 m across
the beach and formed the foredune with pioneer plants. The
extralittoral zone, characterised by a dune ca. 8 m in height,
was covered by a typical Mediterranean maquis. The area was
characterised by microtidal excursions with a tidal range of
less than 30 cm (Colombini et al., 2003).
During the period of the study the sand was covered with
drift material, mainly of fluvial origins, but also with scattered
P. oceanica detritus (agaegropiles, leaf litter, and rhizomes).
Nevertheless, a well-developed P. oceanica ‘banquette’ did not
occur, although they could occasionally achieve significant
sizes during sea storms (50-cm piles 3 m in width along a 300-
m stretch of coastline).
2.2. Experimental design
The beaches at Feniglia and Tagliata (north-west and south-
east with respect to the Ansedonia promontory) are
commonly loaded with large banquettes of P. oceanica depos-
ited in areas close to the promontory by the dominant south-
eastern currents. On the Feniglia beach about 150 kg of dried P.

Tagliata
Feniglia
Ansedonia
Burano Lagoon
N
Italy
30 m
50 m
Site locationPosidonia oceanica meadow
5 km
42°23’50”N
11°22’40”E
Tyrrhenian Sea
Fig. 1 – Study site near the Burano lagoon, Tuscany, Italy. The dashed lines indicate bathymetric levels.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 434
oceanica leaf litter was collected from a large wrack banquette
along a homogeneous beach fascia. In the laboratory the
collected material was placed over nets and dried at 25 �C with
circulating air for 2 months in order to exclude all extant
macrofauna.
Field experiments were carried out from 11 May to 13 June
2005 during a period of theoretically stable climatic condi-
tions. Prior to deployment, P. oceanica leaf litter was soaked in
seawater for 6 h to ensure a homogeneous initial degree of
humidity. One-hundred cylinders (50 cm high, 20 cm in
diameter) were made from green plastic net (9-mm mesh size,
big enough for macroinvertebrates to have free access to the
wrack and to prevent P. oceanica leaf litter from falling out).
Half of the cylinders height was buried into the sand letting
the other half stand above the sand surface. These halves
were separated by a net made of the same material as the
cylinders. The half above the sand was filled to the brim with
1273.5 � 31.2 g (mean and SE) fresh weight of P. oceanica leaf
litter. The cylinders were covered with a plastic lid pressed
with a 1-kg sand bag to obtain similar compacted experi-
mental wracks and to prevent the material from being blown
out by the wind. The leaf compartment of the experimental
cylinder will be hereafter referred to as ‘wrack’. The sand
compartment below the wrack will be referred to as ‘below
wrack sand’(BWS).
A grid, consisting of 100 transects separated from each
other by 3 m and laid perpendicularly to the shoreline was
designed. For each transect, beginning at 1 m from the
shoreline, five positions, at 3-m intervals, were determined
over the eulittoral. The grid therefore resulted in 500 positions
and covered a rectangular area of the beach 297 � 12 m. At the
beginning of the experiment, of the 500 possible positions 100
cylinders were placed over the grid, 20 for each of the five
distances from the shoreline. The position of the latter was
determined according to a computer random generated
distribution pattern.
2.3. Sampling procedures
In order to assess species succession after the deposition of
the material, two experimental cylinders plus two control
sand samples for each distance from the sea were collected
between 08:00 and 10:00 am at 1, 2, 3, 4, 5, 8, 13, 19, 26, and 33-
day intervals from the onset of the experiment. A higher
sampling frequency was conducted during the first days of the
experiment because greater microclimatic changes (especially
for moisture contents) were expected in the wrack in this
period (Messana et al., 1977). After the fifth sampling session
a 5–7-day sampling interval was carried out except for day 8
when a sudden heavy rainfall occurred and the experimental
wracks were checked. Controls consisted of sand cores (25 cm
in height, 20 cm in diameter) sampled at a 1-m distance from
each corresponding experimental cylinder parallel to the
shore. Control cores will be hereafter referred to as ‘control
sand’ (CS). All samples (wrack, BWS, and CS) were collected
separately in thermally-sealed plastic bags. All macro-
invertebrates found were immediately extracted using sieves.
For sand samples 1-mm mesh size was used, whereas for
wrack samples leaves were separated with sieves of 4 mm and
specimens remaining in or escaping the mesh were hand
collected. Individuals were sorted and, when possible, taxo-
nomically classified to species level. A sex ratio was deter-
mined for the two Phaleria species. A size–frequency analysis
was applied to check whether the larvae of Phaleria spp. could
be grouped in different classes according to their size (total
length). This was executed by using the probability paper
method (Harding, 1949). Reliability was tested by employing
both chi-squared and G tests ( p < 0.05) (Zar, 1984). Computa-
tion was executed using ANAMOD 1.6 software (Nogueira,
1992).
Wrack samples were immediately weighed after collection.
Sub-samples (ca. 100 g each) of wrack and sand (for BWS and
CS) were collected and stored in the refrigerator prior to

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 4 35
determination of moisture content (%), conductivity (mS/cm)
and pH according to standard methods (Black, 1973). Moisture
was calculated as the difference between wet and dry weight
(before and after drying at 105 �C until constant weight).
Conductivity and pH were measured after stirring a suspen-
sion of 5 g of sample for 15 min in 100 or 25 ml of demineral-
ised water for P. oceanica and sand samples, respectively.
2.4. Isotopic analysis
At the experimental site, five pitfall traps were deployed at 3-
m intervals from the shore to the base of the dune and kept
operative for 24 h to collect individuals that were used for
stable isotope determination for food web analysis. For this
analysis some macroinvertebrates were selected according to
their trophic level (detritivores, predators). Individuals were
kept alive for 24 h to allow evacuation of gut contents and
then were frozen at �20 �C. For amphipods and isopods two
age class were selected. For amphipods the two classes were
chosen according to the number of articles of the second
antennal flagellum: <10 articles (juveniles) and >20 articles
(adults). For isopods the two classes were selected according
to the maximum width of the 4th pereionit:<3 mm (juveniles)
and >5 mm (adults). The intermediate classes were not
selected because the aim was to assess clear differences
between the two age classes. Prior to isotopic analysis the
specimens were dried at 70 �C until constant weight and
stored in a dry, dark environment.
For isotopic analysis freshly deposited P. oceanica was also
collected along the shoreline in front of the experimental area.
Samples of the most abundant vascular plants occurring in
the foredune (Anthemis maritima L., Cakile maritima Scop.,
Crucianella maritima L., Elymus farctus (Viv.) Runem., Otanthus
maritimus (L.) Hoffmanns et Links, Salsola kali L., Sporobolus
pungens (Schreber) Kunt, Medicago marina L.) and in the dune
system (Juniperus oxycedrus L., J. phoenicea L.) adjacent to the
experimental area were collected in order to assess their
possible use as food for the beach community of macro-
invertebrates or if P. oceanica wrack deposits contributed in
some way as humus for dune vegetation.
Each replicate of macroinvertebrates consisted of 4–10
individuals. An aliquot of ca. 0.7 mg of that composite sample
was used for isotopic determination. The elimination of
carbonates by acidification prior to isotopic analysis was
considered only for crustacean species (Mateo et al., 2008;
Serrano et al., 2008). Acidification consisted of the addition of
1-M HCl drop-by-drop and left in acid for 3 h to eliminate
possible traces of carbonates (Nieuwenhuize et al., 1994). Non-
acidified aliquots were used for d15N determination. P. oceanica
leaf was gently scraped to eliminate epiphytes. A ca. 1.5-mg
dry weight (DW) aliquot of P. oceanica leaf litter from a 5-g DW
powdered bulk sample was used. Only green leaf material
from four individuals of each species of dune vascular plants
was used. For each individual 1 g DW leaf material was dried,
milled and from this material 1.2 mg was encapsulated for
isotopic analysis. The intact portion (about 70 g) of all control
sand samples (n ¼ 100) was sieved using a 125-mm sieve to
extract sand organic matter. Prior to sieving at each distance
from the sea shore (1, 4, 7, 10, and 13 m) samples were grouped
into five cumulated samples for each distance so as to obtain
an adequate organic content for the isotope analysis. The
material passing the sieve was acidified as described above.
Around 6 mg of the resulting material were used to obtain the
isotopic ratios of the sand organic matter (SOMb). A mean
beach SOMb was calculated for 1–4 m (SOMb1) and for 7–13 m
(SOMb2) from the sea shore. The first two distances were
separated from the remaining ones because they were under
the major influence of the sea.
Carbon and nitrogen isotopic signatures were measured
from the gasses evolved from sample combustion in a Fin-
nigan Delta S isotope ratio mass spectrometer (Conflo II
interface) at the Scientific–Technical Services of the Univer-
sity of Barcelona. Isotopic values are reported in the d notation
relative to the standards Vienna Pee Dee Belemnite for carbon
and air for nitrogen (dsample ¼ [(Rsample/Rstandard) � 1] � 1000,
R ¼ 13C/12C, or R ¼ 15N/14N). Analytical precision based on the
standard deviation of internal standards (atropine, IAEA CH3,
CH6, CH7, and USGS40 – analytical grade L-glutamic acid, for
carbon, and atropine, IAEA N1, NO3, N2, and USGS40, for
nitrogen) ranged from 0.06 to 0.11& (mean ¼ 0.09&) for
carbon, and from 0.06 to 0.28& (mean ¼ 0.16&) for nitrogen.
2.5. Numerical procedures and statistical analysis
All data were checked for normality (Kolmogorov–Smirnov
test) and for variance homogeneity (Levene’s test) (Zar, 1984).
For experimental cylinders and controls a two-factor
ANOVA was performed to assess differences of species rich-
ness and ln(n þ 1) transformed abundance of macro-
invertebrates in space (five distances) and in time (ten days).
To compare species richness of experimental cylinders with
that of controls a paired-sample t-test was used. Spearman’s
rank correlation was used to test the association between
macroinvertebrate zonation in the experimental wracks and
time. This was obtained considering the day of capture and
distance from the shoreline for each individual.
To identify the spatial and temporal distributions of the 10
most abundant taxa Hotelling’s confidence ellipses (Batsche-
let, 1981) were calculated at a 95% level of probability. To
assess the differences between the distributions of the
selected taxa Hotelling’s two-sample test was used (Batsche-
let, 1981). A one-factor ANOVA was performed to ascertain if
differences were dependent on both spatial and temporal
factors or only on one factor.
The environmental parameters (moisture content,
conductivity and pH) of the different compartments were
correlated with space and time using Spearman’s rank
correlation.
In order to test the relationships between macrofauna and
environmental parameters, a presence–absence approach
was used associated to univariate parametric t-tests. In this
analysis, pairs of data series of the parameters were
compared, one series with values of all samples where at least
one individual was recorded, and another series with values
where no individuals occurred. For macroinvertebrate abun-
dances the statistical requirements of normality and variance
homogeneity were met when the data were ln(n þ 1) trans-
formed and zero counts were eliminated from the data series.
Furthermore, multiple linear regressions were carried out
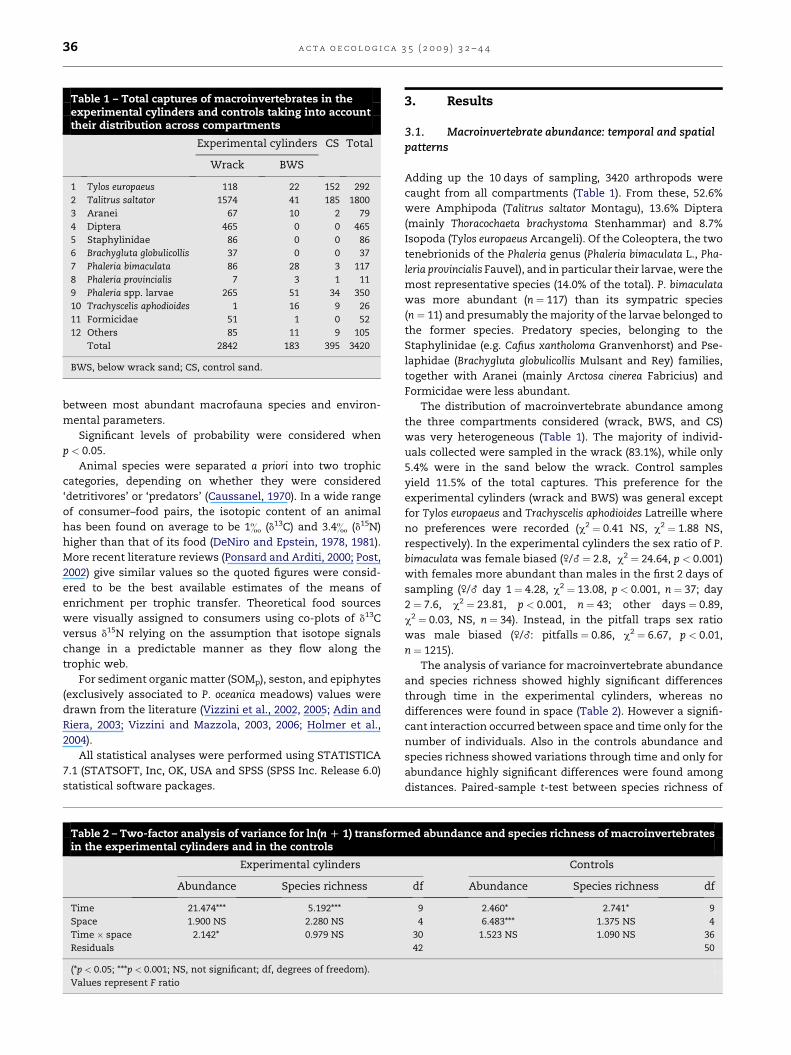
Table 1 – Total captures of macroinvertebrates in theexperimental cylinders and controls taking into accounttheir distribution across compartments
Experimental cylinders CS Total
Wrack BWS
1 Tylos europaeus 118 22 152 292
2 Talitrus saltator 1574 41 185 1800
3 Aranei 67 10 2 79
4 Diptera 465 0 0 465
5 Staphylinidae 86 0 0 86
6 Brachygluta globulicollis 37 0 0 37
7 Phaleria bimaculata 86 28 3 117
8 Phaleria provincialis 7 3 1 11
9 Phaleria spp. larvae 265 51 34 350
10 Trachyscelis aphodioides 1 16 9 26
11 Formicidae 51 1 0 52
12 Others 85 11 9 105
Total 2842 183 395 3420
BWS, below wrack sand; CS, control sand.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 436
between most abundant macrofauna species and environ-
mental parameters.
Significant levels of probability were considered when
p < 0.05.
Animal species were separated a priori into two trophic
categories, depending on whether they were considered
‘detritivores’ or ‘predators’ (Caussanel, 1970). In a wide range
of consumer–food pairs, the isotopic content of an animal
has been found on average to be 1& (d13C) and 3.4& (d15N)
higher than that of its food (DeNiro and Epstein, 1978, 1981).
More recent literature reviews (Ponsard and Arditi, 2000; Post,
2002) give similar values so the quoted figures were consid-
ered to be the best available estimates of the means of
enrichment per trophic transfer. Theoretical food sources
were visually assigned to consumers using co-plots of d13C
versus d15N relying on the assumption that isotope signals
change in a predictable manner as they flow along the
trophic web.
For sediment organic matter (SOMp), seston, and epiphytes
(exclusively associated to P. oceanica meadows) values were
drawn from the literature (Vizzini et al., 2002, 2005; Adin and
Riera, 2003; Vizzini and Mazzola, 2003, 2006; Holmer et al.,
2004).
All statistical analyses were performed using STATISTICA
7.1 (STATSOFT, Inc, OK, USA and SPSS (SPSS Inc. Release 6.0)
statistical software packages.
Table 2 – Two-factor analysis of variance for ln(n D 1) transformin the experimental cylinders and in the controls
Experimental cylinders
Abundance Species richness
Time 21.474*** 5.192***
Space 1.900 NS 2.280 NS
Time � space 2.142* 0.979 NS
Residuals
(*p < 0.05; ***p < 0.001; NS, not significant; df, degrees of freedom).
Values represent F ratio
3. Results
3.1. Macroinvertebrate abundance: temporal and spatialpatterns
Adding up the 10 days of sampling, 3420 arthropods were
caught from all compartments (Table 1). From these, 52.6%
were Amphipoda (Talitrus saltator Montagu), 13.6% Diptera
(mainly Thoracochaeta brachystoma Stenhammar) and 8.7%
Isopoda (Tylos europaeus Arcangeli). Of the Coleoptera, the two
tenebrionids of the Phaleria genus (Phaleria bimaculata L., Pha-
leria provincialis Fauvel), and in particular their larvae, were the
most representative species (14.0% of the total). P. bimaculata
was more abundant (n ¼ 117) than its sympatric species
(n ¼ 11) and presumably the majority of the larvae belonged to
the former species. Predatory species, belonging to the
Staphylinidae (e.g. Cafius xantholoma Granvenhorst) and Pse-
laphidae (Brachygluta globulicollis Mulsant and Rey) families,
together with Aranei (mainly Arctosa cinerea Fabricius) and
Formicidae were less abundant.
The distribution of macroinvertebrate abundance among
the three compartments considered (wrack, BWS, and CS)
was very heterogeneous (Table 1). The majority of individ-
uals collected were sampled in the wrack (83.1%), while only
5.4% were in the sand below the wrack. Control samples
yield 11.5% of the total captures. This preference for the
experimental cylinders (wrack and BWS) was general except
for Tylos europaeus and Trachyscelis aphodioides Latreille where
no preferences were recorded (c2 ¼ 0.41 NS, c2 ¼ 1.88 NS,
respectively). In the experimental cylinders the sex ratio of P.
bimaculata was female biased (\/_ ¼ 2.8, c2 ¼ 24.64, p < 0.001)
with females more abundant than males in the first 2 days of
sampling (\/_ day 1 ¼ 4.28, c2 ¼ 13.08, p < 0.001, n ¼ 37; day
2 ¼ 7.6, c2 ¼ 23.81, p < 0.001, n ¼ 43; other days ¼ 0.89,
c2 ¼ 0.03, NS, n ¼ 34). Instead, in the pitfall traps sex ratio
was male biased (\/_: pitfalls ¼ 0.86, c2 ¼ 6.67, p < 0.01,
n ¼ 1215).
The analysis of variance for macroinvertebrate abundance
and species richness showed highly significant differences
through time in the experimental cylinders, whereas no
differences were found in space (Table 2). However a signifi-
cant interaction occurred between space and time only for the
number of individuals. Also in the controls abundance and
species richness showed variations through time and only for
abundance highly significant differences were found among
distances. Paired-sample t-test between species richness of
ed abundance and species richness of macroinvertebrates
Controls
df Abundance Species richness df
9 2.460* 2.741* 9
4 6.483*** 1.375 NS 4
30 1.523 NS 1.090 NS 36
42 50

0
2
4
6
8
10
12
14
16
0 5 10 15 20 25 30 35Day
Distan
ce fro
m sh
orelin
e (m
)
21
34 5
6
7 9
10
11
2
1
Fig. 2 – Hotelling’s confidence ellipse (95%) of mean day of
appearance and across-shore mean zonation of the
different beach macroinvertebrates in the experimental
cylinders (empty ellipses) and controls (shaded ellipses).
The numbers near the ellipses indicate the taxa as
reported in Table 1.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 4 37
experimental cylinders and that of controls showed higher
values for the former (t ¼ 15.99, p < 0.001, n ¼ 86).
In the experimental cylinders the distance of macro-
invertebrates from the sealine increased throughout the
period of the experiment (rs ¼ 0.617, p < 0.001, n ¼ 3025). In
particular, the analysis of the 10 most abundant taxa through
space and time (Fig. 2) showed that the two crustaceans were
the first species to massively invade the experimental cylin-
ders, showing a mean day of presence (mdp) of 1.40 days and
2.13 days after the beginning of the experiment for T. saltator
and T. europaeus, respectively (Fig. 2). Furthermore, crusta-
ceans had the tendency to occupy the experimental cylinders
closer to the sea with a mean zonation of 1.56 and 2.63 m from
the shoreline for the two species, respectively. Hotelling’s
two-sample test showed highly significant difference between
Table 3 – Hotelling’s two-sample tests and one-factor ANOVA o(n1, capture number of the first taxon; n2, capture number of th
Hotelling’s t
Experimental cylinders
T. europaeus vs. T. saltator 26.775***
Staphylinidae vs. B. globulicollis 0.922 NS
Staphylinidae vs. P. bimaculata 5.780**
P. bimaculata vs. Phaleria spp. larvae 9.546***
P. bimaculata female vs. male 12.265***
P. bimaculata female vs. Phaleria spp. larval stage 1 28.556***
P. bimaculata female vs. Phaleria spp. larval stage 2 7.703***
P. bimaculata female vs. Phaleria spp. larval stage 3 2.270 NS
Phaleria spp. larval stage 1 vs. stage 2 8.303***
Phaleria spp. larval stage 1 vs. stage 3 3.185*
Phaleria spp. larval stage 2 vs. stage 3 0.993 NS
Controls
T. europaeus vs. T. saltator 29.119***
Experimental cylinders vs. controls
T. europaeus 71.870***
T. saltator 764.933***
*p < 0.01, **p < 0.01, ***p < 0.001; NS, not significant.
isopods and amphipods (Table 3) in space and time. Control
samples showed differences between T. saltator and T. euro-
paeus (Fig. 2, Table 3) with a mdp of 16.29 and 12.67 days,
respectively. For T. saltator the results obtained for the
experimental cylinders and CS were different but only from
a temporal point of view, whereas for T. europaeus differences
were both in space and time.
Non-crustacean species colonised experimental cylinders
higher up on the eulittoral with dipterans already appearing
after day 2 (mdp ¼ 2.41 days). Predatory species, such as
staphylinids and B. globulicollis contemporaneously appeared
(mdp ¼ 4.29 and 4.32 days, respectively) (Table 3) after the
dipterans, whereas the most important tenebrionid species
occurred in the wracks after day 5 and 23 (mdp ¼ 5.42 and
23.59 days for P. bimaculata, and T. aphodioides, respectively)
but at a higher distance from the shoreline compared to
predatory species (Fig 2, Table 3). P. bimaculata (Table 3)
showed temporal differences between sex (females
mdp ¼ 3.17, males mdp ¼ 11.73) and with its larvae
(mdp ¼ 8.58), with females differing in mean day of appear-
ance from the first and second larval stage (stage 1
mdp ¼ 10.42, stage 2 mdp ¼ 6.75, stage 3 mdp ¼ 4.88). For-
micidae and Aranei occurred on the experimental cylinders
after day 7 and 11 (mdp ¼ 7.32 and 11.62 days, respectively).
3.2. Environmental parameters
Overall, moisture content in all compartments showed
a decrease both in time and along the sea–land axis and
a significant negative rank correlation occurred in all cases
(Fig. 3). Wrack and BWS moisture decreased from 67 to 14%
and from 2.0 to 0.8%, respectively, during the experimental
period, while in CS the decrease in moisture was from 2.5 to
1.4% for the same period. Wrack material near the seashore
(samples at 1-m distance) was 1.5 times more humid than the
f spatial and temporal distributions of macroinvertebratese second taxon)
est One-factor ANOVA n1 n2
Time Space
14.355*** 49.063*** 140 1615
0.002 NS 1.807 NS 86 37
1.115 NS 9.513 NS 86 114
11.131*** 4.899* 114 316
24.010*** 1.325 NS 84 30
48.941*** 2.388 NS 84 166
13.696*** 0.751 NS 84 133
1.323 NS 3.194 NS 84 17
14.179*** 0.561 NS 166 133
6.336* 0.692 NS 166 17
0.864 NS 1.449 NS 133 17
6.218* 58.101*** 152 185
98.162*** 7.244** 140 152
1529.664*** 0.232 NS 1615 185

Wrack
ba
c d
e f
rs = -0.252p < 0.05
01020304050607080
Mo
istu
re (%
)
rs= -0.817p < 0.001
Below wrack sand
00.5
11.5
22.5
33.5
4
Mo
istu
re (%
)
rs = -0.296p < 0.01
rs= -0.665p < 0.001
Control sand
00.5
11.5
22.5
33.5
14.5
5
0 5 10 15 20 25 30 35Day
Mo
istu
re (%
)
rs= -0.375p < 0.001
0 2 4 6 8 10 12 14Distance from shoreline (m)
rs = -0.565p < 0.001
Fig. 3 – Mean moisture contents with 95% confidence limits are shown in the three compartments (wrack n [ 84, BWS
n [ 86, CS n [ 100) during the course of the experiment and at increasing distance from the shoreline. Spearman’s rank
correlation coefficients and their probability are also reported.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 438
rest of the material. BWS and CS showed similar decreasing
patterns, from 3.4% of moisture near the shore to 1.4% next to
the dune.
Conductivity decreased significantly in time only for wrack
(rs ¼ �0.348, p < 0.01, n ¼ 84) and in time and space for CS
(time: rs ¼ �0.265, p < 0.01, n ¼ 100; space: rs ¼ �0.581,
p < 0.001, n ¼ 100). There was no significant pattern in
conductivity in BWS.
The pH was lowest in the P. oceanica wrack (7.8 on average)
compared to other compartments. For wrack there was a very
slight decreased (ca. 0.1 units) in time and space (time:
rs ¼ �0.319, p < 0.01, n ¼ 84; space: rs ¼ �0.295, p < 0.01,
n ¼ 84), whereas for CS (ca. 0.2 units) a slight increase was
recorded during the course of the experiment (rs ¼ 0.300,
p < 0.01, n ¼ 100) and from the shoreline to the dune
(rs ¼ 0.275, p < 0.01, n ¼ 100). Instead, no consistent pattern in
time or space was observed in the pH of BWS.
3.3. Relationship between macroinvertebrates andenvironmental parameters
In general, the univariate t-tests (Table 4) showed significantly
higher values in the moisture contents of samples where
macroinvertebrates were present compared to those where
they were absent. Instead, tenebrionid species were present
where moisture contents were lower. Crustacean species
preferentially colonised wrack and sand samples with mois-
ture contents ranging from 50 to 75% and from 2 to 5%,
respectively. P. bimaculata with its larvae, were mainly found
in BWS when moisture contents were lower than 2.5%. Higher
values of conductivity occurred in wrack where T. europaeus
and Diptera were present and in CS where the two crusta-
ceans occurred. For the groups where univariate t-tests were
significant, a mean pH value of 7.85 was found in samples
where individuals were present. In particular, in wrack
samples pH values were higher where taxa were present
compared to where they were absent, while in CS the opposite
occurred.
Multiple regression analysis between macroinvertebrates
and environmental parameters (Table 5) showed that T. sal-
tator had positive correlations with moisture contents in the
experimental cylinders and in controls and a negative one
with conductivity of the wrack. In controls T. europaeus was
more abundant where higher sand moisture contents
occurred, whereas in the experimental cylinders isopods
were negatively correlated with conductivity in the wrack

Table 4 – Univariate t-tests for independent samplescomparing environmental parameters (moisture content,conductivity, pH) with presence or absence ofmacroinvertebrates for the three compartmentsconsidered
Moisture Conductivity pH
Wrack (n ¼ 84)
T. europaeus 4.56*** 2.00* –
T. saltator 5.32*** – 3.54***
Diptera 7.57*** 2.99** 3.66***
Staphylinidae 7.60*** – –
B. globulicollis 4.41*** – 2.52*
P. bimaculata – – 2.51*
T. aphodioides �4.01*** – –
BWS (n ¼ 86)
T. europaeus 4.30*** – –
T. saltator 4.78*** – –
Staphylinidae 2.38* – –
B. globulicollis 3.00** – –
P. bimaculata �2.60* – –
Phaleria spp. Larvae �3.33** – –
T. aphodioides �3.27** – –
CS (n ¼ 100)
T. europaeus 2.48* 3.98*** �3.60***
T. saltator 2.79** 3.74*** �2.63**
t values are positive when means of parameter where taxa were
present were higher than those where they were absent and
negative when means were lower where individuals were present.
Non-significant taxa are not shown and dashes indicate non-
significant differences. For further explanations see Tables 1 and 2.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 4 39
and positively with pH of the sand. For the tenebrionid
P. bimaculata a positive relationship with moisture contents
of the wrack was found, whereas its larvae were negatively
correlated with sand conductivity of the experimental
cylinders.
3.4. Stable isotopes
The carbon isotopic composition of the macroinvertebrates
captured over the experimental area ranged from �25.6&
measured for the predatory carabid Scarites buparius (Forster)
to �19.0& recorded in the isopod T. europaeus (Table 6).
Table 5 – Multiple linear regressions between ln(n D 1) transfoparameters (moisture, conductivity, pH) of wrack and sand
Wrack
Moisture (%) Conductivity (mS cm�1) Moistu
Experimental cylinders
T. europaeus – �0.424* –
T. saltator 0.078** �0.670* 0.97
P. bimaculata 0.012** – –
Phaleria spp. larvae – – –
Controls
T. europaeus 0.60
T. saltator 1.08
Non-significant taxa and parameters are not shown. Dashes indicate n
Table 2.
Nitrogen isotopic ratios were lowest, 3.4&, in S. buparius and
highest, 11.7&, in the predatory coleopteran Parallelomorphus
laevigatus (Fabricius). The average d13C and d15N values for
macroinvertebrates feeding on detritus (excluding the Tene-
brionidae Pimelia bipunctata Fabricius) were respectively
�20.8 � 0.36& (SE) and 8.3 � 0.4&, while the same average
figures for the four predatory species captured were
�22.7 � 1.1& and 8.2 � 1.8&, respectively.
All sand plants collected in the area presented largely
distinct isotopic signatures from those of macroinvertebrates
(Table 6). With the exception of C. maritima and S. kali, all of
them yielded negative d15N values. Carbon isotopic signatures
in the plants were on average more negative than in the
macroinvertebrates (�25.6 and �21.8&, respectively). The
genus Juniperus presented distinct carbon and nitrogen
isotopic signatures from the rest of the plants. The Cheno-
podiacea S. kali and the Brassicacea C. maritima showed
exceptionally high nitrogen isotopic ratios (around 6&) and
the maximum and the minimum d13C values (�12.2 and
�30.8&, respectively).
P. oceanica leaf was highly enriched in 13C with respect to the
rest of the sample analysed (�12.7& on average). The literature
review on isotopic signatures of SOMp and seston associated
with P. oceanica meadows, and on P. oceanica epiphytes, yielded
d13C and d15N values ranging from �20.6 to �15.9& and from
4.9 to 6.3& (Table 6). While epiphytes and SOMp were enriched
in 13C with respect to the macroinvertebrates (ca. 4–6&), the
average d13C values for seston and macroinvertebrates were
similar. The average d15N for macroinvertebrates was higher
(ca. 3&) than that of seston. The total SOMb was �21.6 and
4.1& for d13C and d15N values, respectively. Furthermore,
proceeding landwards d13C of SOMb significantly decreased
(SOMb ¼ �0.364 � distance � 19.084, R2 ¼ 0.835, p < 0.001,
n ¼ 24).
4. Discussion
This experiment, conducted in the field in semi-natural
conditions, is an attempt to follow species succession and
colonisation of wracks through time keeping fixed patch size,
an important variable influencing macrofaunal abundance. In
rmed abundance without zero counts and environmental
Sand Constant R2 n
re (%) Conductivity (mS cm�1) pH
– 0.803* – 0.325* 28
3* – – – 0.475** 24
– – 0.647** 0.200** 34
�4.016** – 2.546*** 0.102** 69
5*** – – – 0.359*** 36
0*** – – �1.502* 0.590*** 22
on-significant regression coefficients. For further explanations see
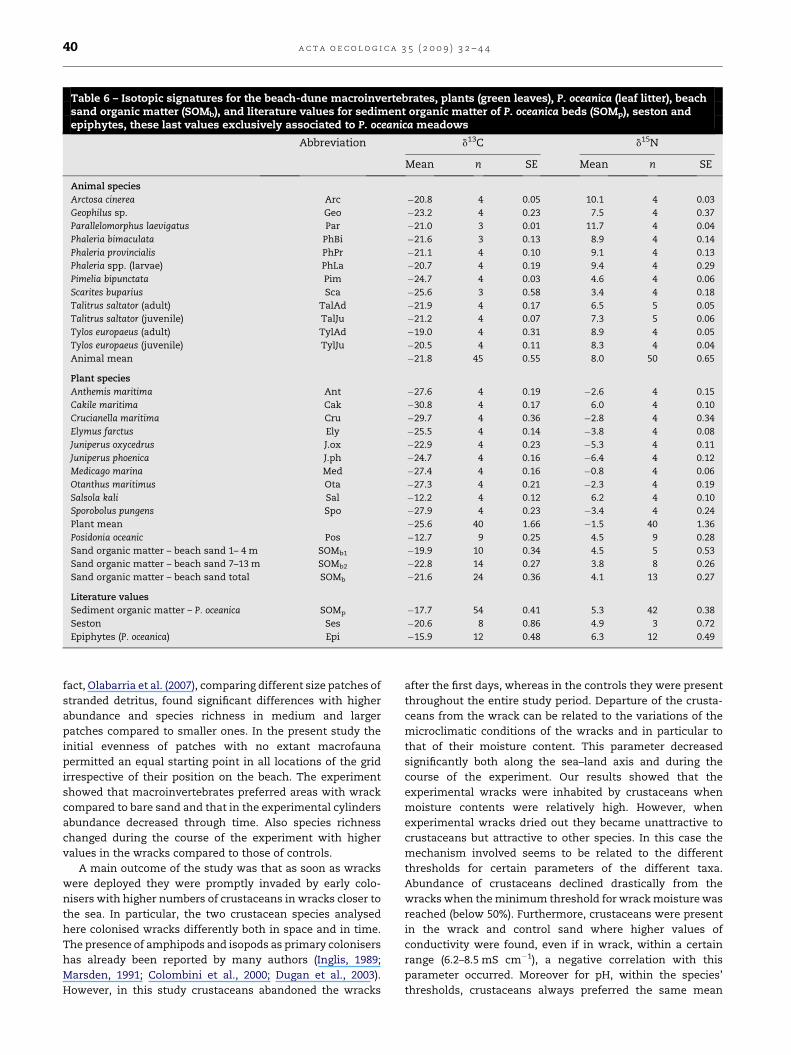
Table 6 – Isotopic signatures for the beach-dune macroinvertebrates, plants (green leaves), P. oceanica (leaf litter), beachsand organic matter (SOMb), and literature values for sediment organic matter of P. oceanica beds (SOMp), seston andepiphytes, these last values exclusively associated to P. oceanica meadows
Abbreviation d13C d15N
Mean n SE Mean n SE
Animal species
Arctosa cinerea Arc �20.8 4 0.05 10.1 4 0.03
Geophilus sp. Geo �23.2 4 0.23 7.5 4 0.37
Parallelomorphus laevigatus Par �21.0 3 0.01 11.7 4 0.04
Phaleria bimaculata PhBi �21.6 3 0.13 8.9 4 0.14
Phaleria provincialis PhPr �21.1 4 0.10 9.1 4 0.13
Phaleria spp. (larvae) PhLa �20.7 4 0.19 9.4 4 0.29
Pimelia bipunctata Pim �24.7 4 0.03 4.6 4 0.06
Scarites buparius Sca �25.6 3 0.58 3.4 4 0.18
Talitrus saltator (adult) TalAd �21.9 4 0.17 6.5 5 0.05
Talitrus saltator (juvenile) TalJu �21.2 4 0.07 7.3 5 0.06
Tylos europaeus (adult) TylAd �19.0 4 0.31 8.9 4 0.05
Tylos europaeus (juvenile) TylJu �20.5 4 0.11 8.3 4 0.04
Animal mean �21.8 45 0.55 8.0 50 0.65
Plant species
Anthemis maritima Ant �27.6 4 0.19 �2.6 4 0.15
Cakile maritima Cak �30.8 4 0.17 6.0 4 0.10
Crucianella maritima Cru �29.7 4 0.36 �2.8 4 0.34
Elymus farctus Ely �25.5 4 0.14 �3.8 4 0.08
Juniperus oxycedrus J.ox �22.9 4 0.23 �5.3 4 0.11
Juniperus phoenica J.ph �24.7 4 0.16 �6.4 4 0.12
Medicago marina Med �27.4 4 0.16 �0.8 4 0.06
Otanthus maritimus Ota �27.3 4 0.21 �2.3 4 0.19
Salsola kali Sal �12.2 4 0.12 6.2 4 0.10
Sporobolus pungens Spo �27.9 4 0.23 �3.4 4 0.24
Plant mean �25.6 40 1.66 �1.5 40 1.36
Posidonia oceanic Pos �12.7 9 0.25 4.5 9 0.28
Sand organic matter – beach sand 1– 4 m SOMb1 �19.9 10 0.34 4.5 5 0.53
Sand organic matter – beach sand 7–13 m SOMb2 �22.8 14 0.27 3.8 8 0.26
Sand organic matter – beach sand total SOMb �21.6 24 0.36 4.1 13 0.27
Literature values
Sediment organic matter – P. oceanica SOMp �17.7 54 0.41 5.3 42 0.38
Seston Ses �20.6 8 0.86 4.9 3 0.72
Epiphytes (P. oceanica) Epi �15.9 12 0.48 6.3 12 0.49
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 440
fact, Olabarria et al. (2007), comparing different size patches of
stranded detritus, found significant differences with higher
abundance and species richness in medium and larger
patches compared to smaller ones. In the present study the
initial evenness of patches with no extant macrofauna
permitted an equal starting point in all locations of the grid
irrespective of their position on the beach. The experiment
showed that macroinvertebrates preferred areas with wrack
compared to bare sand and that in the experimental cylinders
abundance decreased through time. Also species richness
changed during the course of the experiment with higher
values in the wracks compared to those of controls.
A main outcome of the study was that as soon as wracks
were deployed they were promptly invaded by early colo-
nisers with higher numbers of crustaceans in wracks closer to
the sea. In particular, the two crustacean species analysed
here colonised wracks differently both in space and in time.
The presence of amphipods and isopods as primary colonisers
has already been reported by many authors (Inglis, 1989;
Marsden, 1991; Colombini et al., 2000; Dugan et al., 2003).
However, in this study crustaceans abandoned the wracks
after the first days, whereas in the controls they were present
throughout the entire study period. Departure of the crusta-
ceans from the wrack can be related to the variations of the
microclimatic conditions of the wracks and in particular to
that of their moisture content. This parameter decreased
significantly both along the sea–land axis and during the
course of the experiment. Our results showed that the
experimental wracks were inhabited by crustaceans when
moisture contents were relatively high. However, when
experimental wracks dried out they became unattractive to
crustaceans but attractive to other species. In this case the
mechanism involved seems to be related to the different
thresholds for certain parameters of the different taxa.
Abundance of crustaceans declined drastically from the
wracks when the minimum threshold for wrack moisture was
reached (below 50%). Furthermore, crustaceans were present
in the wrack and control sand where higher values of
conductivity were found, even if in wrack, within a certain
range (6.2–8.5 mS cm�1), a negative correlation with this
parameter occurred. Moreover for pH, within the species’
thresholds, crustaceans always preferred the same mean

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 4 41
value that occurred among the higher values in wrack and
among the lower ones in control sand, thus confirming the
findings of Colombini et al. (2005). Generally, talitrids are
known to possess great behavioural plasticity being capable of
reducing unfavourable diurnal conditions, such as extreme
temperatures and desiccation, by changing their position both
vertically and horizontally on the beach (Scapini et al., 1992;
Fallaci et al., 2003). So when talitrids experienced unsuitable
microclimatic conditions in the wracks these were abandoned
immediately.
The presence of the dipteran species (T. brachystoma) on the
more landward wracks already after 2 days confirms the
findings of Hodge and Arthur (1997) for other species of British
seaweed flies (Coelopa frigida Fabricius, Coelopa pilipes Haliday,
Thoracochaeta zosterae Stenhammar;) in which all three
dipteran species had arrived on wracks by day 2. Coelopa larvae
generally fed on bacterial populations (Cullen et al., 1987) and
their developmental rate was generally associated with
temperature conditions (Phillips et al., 1995). The same pattern
probably occurred for T. brachystoma in the P. oceanica leaves.
Consequently, the presence of eggs and larvae of dipterans in
the wrack attracted predators, such as staphylinids (C. xan-
tholoma) and pselaphids (B. globulicollis), as was demonstrated
by feeding experiments on C. xantholoma in which a strict
predator–prey relationship occurred (Backlund, 1945).
Tenebrionids (P. bimaculata and its larvae) were mid-
invaders and preferentially occupied experimental wracks
higher up on the eulittoral in relation to their preference for
lower moisture contents in the sand (Aloia et al., 1999;
Colombini et al., 2005). The differences found in the sex ratios
in the experimental wracks compared to the active population
(see pitfall traps) indicated that females were attracted to the
experimental cylinders before males suggesting that wracks,
or sand beneath the wracks, might be used as sites for ovi-
deposition. Another fact supporting this hypothesis was the
appearance of the first larval stage a week after female colo-
nisation. The other larger larval stages invaded the wracks
slightly after adults meaning that wracks could attract pre-
existing larvae and be used as an important nursery ground by
these tenebrionids. The tenebrionid T. aphodioides was defin-
itively a late invader appearing on wracks only in the last
period of the experiment. A process of interspecific facilitation
has been suggested by some authors (Schoenly and Reid, 1987;
Heard, 1994) with late species benefiting from the presence of
the earlier arrival of primary colonists. The presence of early
arrivals modifies the resource qualitatively and makes it more
suitable for later invaders. Another process probably involved
is reciprocal avoidance due to the temporal separation that
reduced levels of interspecific interference at the food source.
In general, the presence of a specific fauna in the wrack is thus
the result of a combination of factors such as species adap-
tation to specific microclimatic conditions of the wracks, and
the need to find an adequate food source benefiting from the
presence of other species (both bacteria and invertebrates) on
the wrack. Changes in the microclimatic conditions of the
wrack suggest that beach deposits might represent an
important but quite transient resource for food and shelter for
resident macroinvertebrates as was also shown for stranded
wracks of beaches in tropical and temperate areas (Colombini
et al., 2000; Colombini and Chelazzi, 2003; Orr et al., 2005).
With the exception of seston and sand organic matter
(SOMb), the rest of the potential sources of organic carbon
considered in this study showed clearly distinct d13C vs. d15N
values from the macroinvertebrate fauna. Accepting the
broadly mean trophic enrichment in d13C of 1& (DeNiro and
Epstein, 1978) and in d15N of 3.4& (DeNiro and Epstein, 1981;
Minagawa and Wada, 1984), the expected mean isotopic ratios
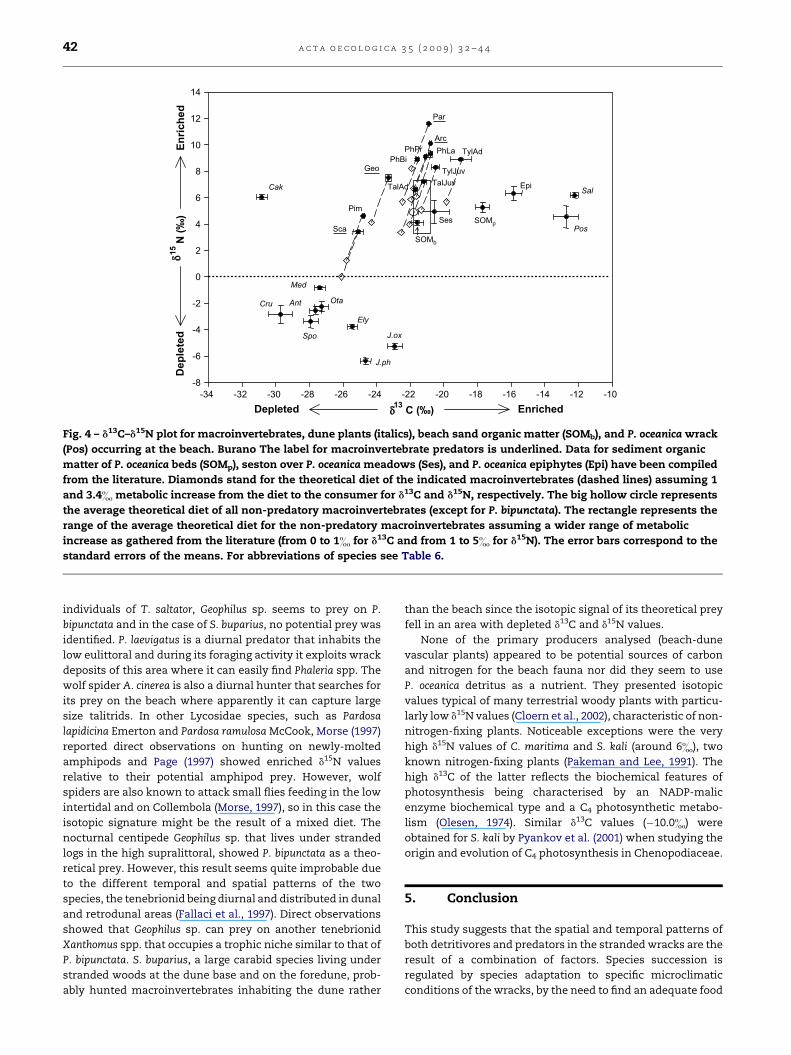
for the preferentially exploited food resource by the non-
predatory macroinvertebrate species would be around �21.8
and 4.9& for d13C and d15N, respectively (Fig. 4). The only
exception was P. bipunctata that presented a theoretical diet of
�23.7 and 1.2&. This species was distributed in a more land-
ward area compared to other scavengers (Fallaci et al., 1997;
Aloia et al., 1999) and thus probably foraged on more terres-
trial resources. Adults and juveniles of T. saltator clearly did
not directly feed on P. oceanica leaves as shown by the isotopic
results, but probably took refuge in wracks in search of shelter
and suitable microclimatic conditions during the day. This
finding is in accordance with the results of Adin and Riera
(2003) that showed that neither dune plants (Agropyrum jun-
ceum, L., Atriplex laciniata L.) nor seagrass (Zostera marina L.)
were used as food sources by T. saltator. These authors sug-
gested that several macroalgal species occurring in the
strandline could contribute to the diet of T. saltator even if this
amphipod preferentially utilised Fucus sp. as main food
source. P. oceanica was the most d13C enriched and this result
was in good agreement with other data reported in the liter-
ature (Vizzini et al., 2002; Carlier et al., 2007). There are several
rationales supporting the theory that seagrasses do not
significantly contribute to the basis of the food web in marine
habitats because they are too C13-enriched compared to
primary consumers (Gacia et al., 2002; Vizzini et al., 2002;
Hyndes and Lavery, 2005) and this seems to occur also for
beach stranded P. oceanica leaves. Tylos europaeus presented
a theoretical diet with values very close to those obtained for
SOMb, especially for juveniles. A similar result was also
obtained for another oniscid, Tylos ponticus Grebnitzky, for
which a marine type of diet (with more algae compared to
seagrass) was suggested by both the isotopic results and the
feeding preference experiments (Dias and Hassall, 2005). Our
findings at Burano strongly suggest that the seston of the
water column is the main organic matter supplier for sandy
beaches with low macroalgae inputs, in particular for the
areas closer to the seashore (Fig. 4). This is supported by the
clear impoverishment of the SOMb in d13C from the seashore
to the dune, probably reflecting the change in the proportion
of organic matter contributed by the seston or by the dune
vegetation. The seston, carried by the water washing the
shore, seems to be retained by the sand that acts as a particle
filter. The isotopic composition of the SOMb1, collected at 1–
4 m from the shoreline, is similar to that of the seston, and the
average isotopic composition of the SOMb, all over the studied
area, matches perfectly with the theoretical diet and range
(see box in Fig. 4) of the detritus feeders considered in this
study assuming a wider range of metabolic increase as gath-
ered from the literature (from 0 to 1& for d13C and from 1 to 5&
for d15N, Minagawa and Wada, 1984; Post, 2002).
The theoretical diets calculated for the predators have
shown that P. laevigatus seems to feed predominantly on
Phaleria spp., the diet of A. cinerea seems based purely on adult
-34 -32 -30 -28 -26 -24 -22 -20 -18 -16 -14 -12 -10-8
-6
-4
-2
0
2
4
6
8
10
12
14
Cak
Par
ArcTylAd
TylJuvTalJuvTalAd
Pos
Sal
Sca
GeoPhBi
PhLa
Pim
PhPr
Cru
Med
OtaAnt
Spo J.ox
J.ph
Ely
SOMp
Epi
Ses
SOMb
C (‰)Depleted Enriched 13
N (‰
)D
ep
leted
E
nrich
ed
15
Fig. 4 – d13C–d15N plot for macroinvertebrates, dune plants (italics), beach sand organic matter (SOMb), and P. oceanica wrack
(Pos) occurring at the beach. Burano The label for macroinvertebrate predators is underlined. Data for sediment organic
matter of P. oceanica beds (SOMp), seston over P. oceanica meadows (Ses), and P. oceanica epiphytes (Epi) have been compiled
from the literature. Diamonds stand for the theoretical diet of the indicated macroinvertebrates (dashed lines) assuming 1
and 3.4& metabolic increase from the diet to the consumer for d13C and d15N, respectively. The big hollow circle represents
the average theoretical diet of all non-predatory macroinvertebrates (except for P. bipunctata). The rectangle represents the
range of the average theoretical diet for the non-predatory macroinvertebrates assuming a wider range of metabolic
increase as gathered from the literature (from 0 to 1& for d13C and from 1 to 5& for d15N). The error bars correspond to the
standard errors of the means. For abbreviations of species see Table 6.
a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 442
individuals of T. saltator, Geophilus sp. seems to prey on P.
bipunctata and in the case of S. buparius, no potential prey was
identified. P. laevigatus is a diurnal predator that inhabits the
low eulittoral and during its foraging activity it exploits wrack
deposits of this area where it can easily find Phaleria spp. The
wolf spider A. cinerea is also a diurnal hunter that searches for
its prey on the beach where apparently it can capture large
size talitrids. In other Lycosidae species, such as Pardosa
lapidicina Emerton and Pardosa ramulosa McCook, Morse (1997)
reported direct observations on hunting on newly-molted
amphipods and Page (1997) showed enriched d15N values
relative to their potential amphipod prey. However, wolf
spiders are also known to attack small flies feeding in the low
intertidal and on Collembola (Morse, 1997), so in this case the
isotopic signature might be the result of a mixed diet. The
nocturnal centipede Geophilus sp. that lives under stranded
logs in the high supralittoral, showed P. bipunctata as a theo-
retical prey. However, this result seems quite improbable due
to the different temporal and spatial patterns of the two
species, the tenebrionid being diurnal and distributed in dunal
and retrodunal areas (Fallaci et al., 1997). Direct observations
showed that Geophilus sp. can prey on another tenebrionid
Xanthomus spp. that occupies a trophic niche similar to that of
P. bipunctata. S. buparius, a large carabid species living under
stranded woods at the dune base and on the foredune, prob-
ably hunted macroinvertebrates inhabiting the dune rather
than the beach since the isotopic signal of its theoretical prey
fell in an area with depleted d13C and d15N values.
None of the primary producers analysed (beach-dune
vascular plants) appeared to be potential sources of carbon
and nitrogen for the beach fauna nor did they seem to use
P. oceanica detritus as a nutrient. They presented isotopic
values typical of many terrestrial woody plants with particu-
larly low d15N values (Cloern et al., 2002), characteristic of non-
nitrogen-fixing plants. Noticeable exceptions were the very
high d15N values of C. maritima and S. kali (around 6&), two
known nitrogen-fixing plants (Pakeman and Lee, 1991). The
high d13C of the latter reflects the biochemical features of
photosynthesis being characterised by an NADP-malic
enzyme biochemical type and a C4 photosynthetic metabo-
lism (Olesen, 1974). Similar d13C values (�10.0&) were
obtained for S. kali by Pyankov et al. (2001) when studying the
origin and evolution of C4 photosynthesis in Chenopodiaceae.
5. Conclusion
This study suggests that the spatial and temporal patterns of
both detritivores and predators in the stranded wracks are the
result of a combination of factors. Species succession is
regulated by species adaptation to specific microclimatic
conditions of the wracks, by the need to find an adequate food

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 4 43
source by benefiting from the presence of other species (both
bacteria and macroinvertebrates) on the wrack and by the
need to find an adequate refuge from the stressful environ-
mental conditions.
Furthermore, the study has clearly shown that P. oceanica
wrack is not a dietary source for the macroinvertebrate fauna
of Burano beach, as already shown in the marine habitat, but
it is a physical structure of vital importance for detritivorous
and predatory species as refuge from environmentally
stressful conditions.
This study should be complemented with further research
taking into consideration the effect of wrack size, seasonality
and bacterial presence on the abundance and species richness
of beach macroinvertebrates.
Acknowledgments
The authors are grateful to Guillermo Mendoza, Dani Martin
(Centre of Advanced Studies of Blanes, CEAB, CSIC), and to
Rafa Marce for their help in some statistical aspects of the data
analysis. This study has been supported partially by funds of
a bilateral programme between CNR (Italy) and CSIC (Spain),
and partially by the EU-INCO project WADI (CT2005-15226).
We are also grateful to Fabio Cianchi of the WWF Burano Oasis
for his assistance. The experiments performed for this study
comply with the current laws of Italy.
r e f e r e n c e s
Adin, R., Riera, P., 2003. Preferential food source utilization amongstranded macroalgae by Talitrus saltator (Amphipoda,Talitridae): a stable isotopes study in the northern coast ofBrittany (France). Estuar. Coast. Shelf Sci. 56, 91–98.
Aloia, A., Colombini, I., Fallaci, M., Chelazzi, L., 1999. Behaviouraladaptations to zonal maintenance of fivespecies of tenebrionidsliving along a Tyrrhenian sandy shore. Mar. Biol. 133, 473–487.
Backlund, H.O., 1945. The wrack fauna of Sweden and Finland:ecology and chorology. Opusc. Entomol. Suppl 5 1–236.
Bascom, W., 1980. Waves and Beaches. Anchor Press/Doubleday,New York.
Batschelet, E., 1981. Circular Statistic in Biology. Academic Press,London.
Bianchi, C.N., Cinelli, F., Morri, C., 1993. La carta bionomica deiMari Toscani. Regione Toscana.
Black, C.A., 1973. Methods of Soil Analysis. Part 2: Chemical andMicrobiological Properties. American Society of Agronomy,Madison, WI.
Carlier, A., Riera, P., Amouroux, J.M., Bodiou, J.Y., Gremare, A.,2007. Benthic trophic network in the Bay of Banyuls-sur-Mer(northwest Mediterranean, France): an assessment based onstable carbon and nitrogen isotopes analysis. Estuar. Coast.Shelf Sci. 72, 1–15.
Caussanel, C., 1970. Contribution a l’etude du peuplement d’uneplage et d’une dune landaise. Vie Milieu. 21, 59–104.
Cebrian, J., Duarte, C.M,, Marba, N., Enriquez, S., 1997. Magnitudeand fate of production of four co-occurring westernMediterranean seagrass species. Mar. Ecol. Prog. Ser. 155, 29–44.
Cloern, J.E., Canuel, E.A., Harris, D., 2002. Stable carbon andnitrogen isotope composition of aquatic and terrestrial plants
of the San Francisco Bay estuarine system. Limnol. Oceanogr.47, 713–729.
Colombini, I., Chelazzi, L., 2003. Influence of marineallochthonous input of sandy beach communities. Oceanogr.Mar. Biol. Annu. Rev. 41, 115–159.
Colombini, I., Aloia, A., Fallaci, M., Pezzoli, G., Chelazzi, L., 2000.Temporal and spatial use of stranded wrack by themacrofauna of a tropical sandy beach. Mar. Biol. 136, 531–541.
Colombini, I., Fallaci, M., Milanesi, F., Scapini, F., Chelazzi, L.,2003. Comparative diversity analysis in sandy littoralecosystems of the western Mediterranean. Estuar. Coast. ShelfSci. 58S, 93–104.
Colombini, I., Fallaci, M., Chelazzi, L., 2005. Micro-scaledistribution of some arthropods inhabiting a Mediterraneansandy beach in relation to environmental parameters. ActaOecol. 28, 249–265.
Cullen, S.J., Young, A.M., Day, T.H., 1987. Dietary requirements ofseaweed flies (Coelopa frigida). Estuar. Coast. Shelf Sci. 24,701–710.
De Falco, G., Molinaroli, N.E., Baroli, M., Bellacicco, S., 2003. Grainsize and compositional trends from Posidonia oceanicameadows to beach shore, Sardinia, western Mediterranean.Estuar. Coast. Shelf Sci. 58, 299–309.
DeNiro, M.J., Epstein, S., 1978. Influence of diet on the distributionof carbon isotopes in animals. Geochim. Cosmochim. Acta 42,495–506.
DeNiro, M.J., Epstein, S., 1981. Influence of diet on the distributionof nitrogen isotopes in animals. Geochim. Cosmochim. Acta45, 341–351.
Dias, N., Hassall, M., 2005. Food, feeding and growth rates ofperacarid macro-decomposers in a Ria Formosa saltmarsh,southern Portugal. J. Exp. Mar. Biol. Ecol. 325, 84–94.
Dugan, J.E., Hubbard, D.M., McCrary, M.D., Pierson, M.O., 2003.The response of macrofauna communities and shorebirds tomacrophyte wrack subsidies on exposed sandy beaches ofsouthern California. Estuar. Coast. Shelf Sci. 58S, 25–40.
Fallaci, M., Colombini, I., Taiti, S., Chelazzi, L., 1996.Environmental factors influencing the surface activity andzonation of Tylos europaeus (Crustacea: Oniscidea) ona Tyrrhenian sandy beach. Mar. Biol. 125, 751–763.
Fallaci, M., Colombini, I., Palesse, L., Chelazzi, L., 1997. Spatial andtemporal strategies in relation to environmental constraintsof four tenebrionids inhabiting a Mediterranean coastal dunesystem. J. Arid. Environ. 37, 45–64.
Fallaci, M., Aloia, A., Colombini, I., Chelazzi, L., 2002. Populationdynamics and life history of two Phaleria species (Coleoptera,Tenebrionidae) living on the Tyrrhenian sandy coast of centralItaly. Acta Oecol. 23, 69–79.
Fallaci, M., Colombini, I., Lagar, M., Scapini, F., Chelazzi, L., 2003.Distribution patterns of different age classes and sexes ina Tyrrhenian population of Talitrus saltator (Montagu). Mar.Biol. 142, 101–110.
Folk, R.L., Ward, W.C., 1957. Brazos river bar: a study in thesignificance of grain size parameters. J. Sediment. Petrol. 27,3–26.
Gacia, E., Duarte, C.M., Middelburg, J.J., 2002. Carbon and nutrientdeposition in a Mediterranean seagrass (Posidonia oceanica)meadow. Limnol. Oceanogr. 47, 23–32.
Harding, J.P., 1949. The use of the probability paper for thegraphical analysis of polymodal frequency distributions.J. Mar. Biol. Assoc. U.K. 28, 141–153.
Heard, S.B., 1994. Processing chain ecology: resource conditionand interspecific interactions. J. Anim. Ecol. 63, 451–464.
Hemminga, M.A., Nieuwenhuize, J., 1990. Seagrass wrack-induced dune formation on a tropical coast (Banq d’Arguin,Mauritania). Estuar. Coast. Shelf Sci. 31, 499–502.
Hodge, S., Arthur, W., 1997. Asymmetric interaction betweenspecies of seaweed fly. J. Anim. Ecol. 66, 743–754.

a c t a o e c o l o g i c a 3 5 ( 2 0 0 9 ) 3 2 – 4 444
Holmer, M., Duarte, C.M., Boschker, H.T.S., Barron, C., 2004.Carbon cycling and bacterial carbon sources in pristine andimpacted Mediterranean seagrass sediments. Aquat. Microb.Ecol. 36, 227–237.
Hyndes, G.A., Lavery, P.S., 2005. Does transported seagrassprovide an important trophic link in unvegetated nearshoreareas? Estuar. Coast. Shelf Sci. 63, 633–643.
Ince, R., Hyndes, G., Lavery, P.S., Vanderklift, M.A., 2007. Marinemacrophytes directly enhance abundance of sandy beachfauna through provision of food and habitat. Estuar. Coast.Shelf Sci. 74, 77–86.
Inglis, G., 1989. The colonisation and degradation of strandedMacrocystis pyrifera (L.) C. Ag. by the macrofauna of a NewZealand sandy beach. J. Exp. Mar. Biol. Ecol. 125, 203–217.
Jaramillo, E., De La Huz, R., Duarte, C., Contreras, H., 2006. Algalwrack deposits and macroinfaunal arthropods on sandybeaches of the Chilean coast. Rev. Chil. Hist. Nat. 79,337–351.
Marba, N., Hemminga, M.A., Mateo, M.A., Duarte, C.M., Mass, Y.E.M., Terrados, J., Garcia, E., 2002. Carbon and nitrogentranslocation between seagrass ramets. Mar. Ecol. Prog. Ser.226, 287–300.
Marsden, I.D., 1991. Kelp–sandhopper interactions on a sandbeach in New Zealand. II. Population dynamics of Talorchestiaquoyana (Milne–Edwards). J. Exp. Mar. Biol. Ecol. 152, 75–90.
Mateo, M.A., Romero, J., Perez, M., Littler, M.M., Littler, D.S., 1997.Dynamics of millenary organic deposits resulting from thegrowth of the Mediterranean seagrass Posidonia oceanica.Estuar. Coast. Shelf Sci. 44, 103–110.
Mateo, M.A., Sanchez-Lizaso, J.-L., Romero, J., 2003. Posidoniaoceanica ‘‘banquettes’’: a preliminary assessment of therelevance for meadow carbon and nutrients budget. Estuar.Coast. Shelf Sci. 56, 85–90.
Mateo, M.A., Cebrian, J., Dunton, K., Mutchler, T., 2006. Carbonflux in seagrass ecosystems. In: Larkum, A.W.D., Orth, R.J.,Duarte, C.M. (Eds.), Seagrasses: Biology, Ecology andConservation. Springer Publishing, Netherlands, pp. 159–192.
Mateo, M.A., Serrano, O., Serrano, L., Michener, R.H., 2008. Effectsof sample preparation on stable isotope ratios of carbon andnitrogen in marine invertebrates: implications for food webstudies using stable isotopes. Oecologia 157, 105–115.
McLachlan, A., 1980. The definition of sandy beaches in relationto exposure: a simple rating system. S. Afr. J. Sci. 76, 137–138.
McLachlan, A., Brown, A., 2006. The Ecology of Sandy Shores,second ed. Academic Press.
Messana, G., Chelazzi, G., Chelazzi, L., Ercolini, A., Ferrara, F.,Messeri, P., Pardi, P., Vannini, M., 1977. Researches on thecoast of Somalia. The shore and the dune of San Uanle. 12.Physical environment: microclimate and soil. Monitore. Zool.Ital. NS (Suppl 9), 147–181.
Minagawa, M., Wada, E., 1984. Stepwise enrichment of 15N alongfood chains: further evidence and the relation between d15Nand animal age. Geochim. Cosmochim. Acta 48, 1135–1140.
Morse, D.H., 1997. Distribution, movement, and activity patternsof an intertidal wolf spider Pardosa lapidicina population(Araneae, Lycosidae). J. Arachnol. 25, 1–10.
Nieuwenhuize, J., Maas, Y.E.M., Middelburg, J.J., 1994. Rapidanalysis of organic carbon and nitrogen in particulatematerials. Mar. Chem. 45, 217–224.
Nogueira, A.J.A., 1992. ANAMOD Extraccao dos componentesmodais de distribuicoes de frequecias de variaveisbiometricas. Departamento de Zoologia Facultade de Cienciase Tecnologia, Universidade de Coimbra.
Ochieng, C.A., Erftemeijer, P.L.A., 1999. Accumulation of seagrassbeach cast along the Kenyan coast: a quantitative assessment.Aquat. Bot. 65, 221–238.
Olabarria, C., Lastra, M., Garrido, J., 2007. Succession ofmacrofauna on microalgal wrack of an exposed sandy beach:effects of patch size and site. Mar. Environ. Res. 63, 19–40.
Olesen, P., 1974. Leaf anatomy and ultrastructure of chloroplastsin Salsola kali L. as related to the C4-pathway ofphotosynthesis. Bot. Notiser. 127, 352–363.
Orr, M., Zimmer, M., Jelinski, D.E., Mews, M., 2005. Wrackdecomposition on different beach types: spatial and temporalvariation in the pattern of subsidy. Ecology 86, 1496–1507.
Page, H.M., 1997. Importance of vascular plant and algalproduction to macroinvertebrate consumers in a SouthernCalifornia salt marsh. Estuar. Coast. Shelf Sci. 45, 823–834.
Pakeman, R.J., Lee, J.A., 1991. The ecology of the strandlineannuals Cakile maritima and Salsola kali. I. Environmentalfactors affecting plant performance. J. Ecol. 79, 143–153.
Phillips, D.S., Leggett, M., Wilcockson, R., Day, T.H., Arthur, W.,1995. Coexistence of competing species of seaweed flies: therole of temperature. Ecol. Entomol. 20, 65–74.
Polis, G.A., Hurd, S.D., 1996. Allochthonous input across habitats,subsidised consumers and apparent trophic cascades:examples from the ocean–land interface. In: Polis, G.A.,Winemiller, K.O. (Eds.), Food Webs: Integration of Patterns andDynamics. Chapman and Hall, London, pp. 275–285.
Ponsard, S., Arditi, R., 2000. What can stable isotopes (d15N andd13C) tell about the food web of soil macro-invertebrates?Ecology 81, 852–864.
Post, D.M., 2002. Using stable isotopes to estimate trophicposition: models, methods and assumptions. Ecology 83,703–718.
Pyankov, V., Ziegler, H., Kuz’min, A., Edwards, G., 2001. Origin andevolution of C4 photosynthesis in the tribe Salsoleae(Chenopodiaceae) based on anatomical and biochemical typesin leave and cotyledons. Plant Syst. Evol. 230, 43–74.
Scapini, F., Chelazzi, L., Colombini, I., Fallaci, M., 1992. Surfaceactivity, zonation and migration of Talitrus saltator ona Mediterranean beach. Mar. Biol. 112, 573–581.
Schoenly, K., Reid, W., 1987. Dynamics of heterotrophicsuccession in carrion arthropod assemblages: discrete seriesor a continuum of change? Oecologia 73, 192–202.
Serrano, O., Serrano, L., Mateo, M.A., Colombini, I., Chelazzi, L.,Gagnarli, E., Fallaci, M., 2008. Acid washing effect on elementaland isotopic composition of whole beach arthropods:implications for food web studies using stable isotopes. ActaOecol 34, 89–96.
Vizzini, S., Mazzola, A., 2003. Seasonal variations in the stablecarbon and nitrogen isotope ratio 13C/12C and 15N/14N) ofprimary producers and consumers in a westernMediterranean coastal lagoon. Mar. Biol. 142, 1009–1018.
Vizzini, S., Mazzola, A., 2006. Sources and transfer of organic matterin food webs of a Mediterranean coastal environment: evidencefor spatial variability. Estuar. Coast. Shelf Sci. 66, 459–467.
Vizzini, S., Sara, G., Michener, R.H., Mazzola, A., 2002. The roleand contribution of Posidonia oceanica (L.) Delile organic matterfor secondary consumers as revealed by carbon and nitrogenstable isotope analysis. Acta Oecol. 23, 277–285.
Vizzini, S., Savona, B., Do Chi, T., Mazzola, A., 2005. Spatialvariability of stable carbon and nitrogen isotope ratios ina Mediterranean coastal lagoon. Hydrobiologia 550, 73–82.
Zar, J.H., 1984. Biostatistical Analysis, second ed. Prentice HallInternational Inc., Englewood Cliffs, NJ.
Top Related
Copyright © 2022 FDOKUMEN

