







Bahasa
Halaman
Hukum


Nucleic Acids Research, 2007, 1–10doi:10.1093/nar/gkm440
Nuclear accumulation of plasmid DNA can beenhanced by non-selective gating of the nuclear poreRoosmarijn E. Vandenbroucke, Bart Lucas, Joseph Demeester,
Stefaan C. De Smedt* and Niek N. Sanders
Laboratory of General Biochemistry and Physical Pharmacy, Ghent University, Harelbekestraat 72, B-9000 Ghent,Belgium
Received January 25, 2007; Revised April 19, 2007; Accepted May 17, 2007
ABSTRACT
One of the major obstacles in non-viral genetransfer is the nuclear membrane. Attempts toimprove the transport of DNA to the nucleus throughthe use of nuclear localization signals or importin-bhave achieved limited success. It has been pro-posed that the nuclear pore complexes (NPCs)through which nucleocytoplasmic transport occursare filled with a hydrophobic phase through whichhydrophobic importins can dissolve. Therefore,considering the hydrophobic nature of the NPCchannel, we evaluated whether a non-selectivegating of nuclear pores by trans-cyclohexane-1,2-diol (TCHD), an amphipathic alcohol that rever-sibly collapses the permeability barrier of the NPCs,could be obtained and used as an alternativemethod to facilitate nuclear entry of plasmid DNA.Our data demonstrate for the first time that TCHDmakes the nucleus permeable for both high mole-cular weight dextrans and plasmid DNA (pDNA) atnon-toxic concentrations. Furthermore, in line withthese observations, TCHD enhanced the transfec-tion efficacy of both naked DNA and lipoplexes. Inconclusion, based on the proposed structure ofNPCs we succeeded to temporarily open the NPCsfor macromolecules as large as pDNAs and demon-strated that this can significantly enhance non-viralgene delivery.
INTRODUCTION
Viral vectors are efficient DNA delivery systems as theypossess natural mechanisms to enter cells, to escape fromendosomes and to transport their DNA into the nucleus.However, they also display important disadvantages, suchas immunogenic response and safety risks when adminis-tered to patients. Non-viral carriers lack these disadvan-tages, but poor transfection efficiencies currently limit the
usefulness of these vectors for gene therapy applications.The low gene transfer capacity of non-viral vectors ismainly due to their inability to translocate the therapeuticDNA into the cell nucleus. Indeed, it has been shown thatmicroinjection of plasmid DNA (pDNA) in the cytoplasmof non-dividing cells resulted in 51% gene expression,while a massive gene expression occurred when the pDNAwas injected in the nucleus (1–3). In dividing cells thenuclear envelope disassembles on a regular base, whichoffers an opportunity for DNA to enter the nucleus (4–6).However, the DNA that is waiting in the cytoplasm forthe next cell division is sensitive to degradation bynucleases. Therefore, methods that can enhance thenuclear uptake of DNA into nuclei of both non-dividingand dividing cells are urgently needed in non-viral-basedgene therapy.Nucleocytoplasmic transport proceeds through the
nuclear pore complexes (NPCs) which form channels inthe nuclear envelope with a diameter of �40 nm (7,8).Vertebrate NPCs have a mass of �125MDa and contain30–50 different proteins, which are called nucleoporins.Small molecules with a molecular weight up to 30 kDa canpassively diffuse through the NPC. In contrast, thetranslocation of larger macromolecules into the nucleusoccurs via an active mechanism involving nuclear trans-port receptors. The majority of the nuclear transportpathways are mediated by receptors of the importinfamily. Proteins or other cargo molecules that carry aclassical nuclear localization sequence (NLS) are recog-nized by importin-a, which subsequently forms a complexthrough its importin-b-binding domain with importin-b(9). NLSs can be highly diverse in nature and rangefrom the short bipartite classical NLS to extendedprotein domains, as is the case for histones or ribosomalproteins (10). To promote the nuclear import of DNA,NLS peptides, NLS-containing proteins and evenimportin-b (11) have been attached to DNA via severalstrategies: electrostatic (12–17) or covalent (18–22) bind-ing, via protein–DNA interaction (23,24), via PNA clamps(15,25–27) and coupled to polymers (28–30), lipids (31–36)or recombinant lambda phage (37). Nevertheless, all these
*To whom correspondence should be addressed. Tel: +32-9-2648047; Fax: +32-9-2648189; Email: [email protected]
� 2007 The Author(s)
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/
by-nc/2.0/uk/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Nucleic Acids Research Advance Access published June 21, 2007 by guest on M
arch 3, 2014http://nar.oxfordjournals.org/
Dow
nloaded from

attempts to improve the transport of DNA to the nucleusthrough the use of NLSs or importin-b have achieved onlylimited success.It has recently been shown that the nuclear uptake of
macromolecules can be enhanced significantly by additionof the amphipathic molecule trans-cyclohexane-1,2-diol(TCHD) (38). The mechanism by which TCHD causesnuclear localization of macromolecules can be explainedbased on the inner channel properties of the nuclear pores.It is believed that these nuclear pores are filled with ahydrophobic phase through which importins, but not inerthydrophilic substrates, can dissolve. The addition ofTCHD causes a temporary, non-selective gating of thepore and allows passage of molecules which wouldotherwise be rejected from passage. In other words, anon-selective gating of the nuclear pore channel by TCHDrenders the actual translocation through the pore channelindependent of nuclear transport receptors. This can beexplained by the fact that TCHD causes disruption of thehydrophobic interactions between the hydrophobic phe-nylalanine–glycine repeats of the nucleoporines, whichconsequently causes collapsing of the permeability barrierof the NPCs. Importantly, the effect of TCHD is reversibleand does not cause damage of the nuclear pores (38).In this paper we studied whether a non-selective gating
of nuclear pores by amphipathic molecules like TCHDcould also be used as an alternative method to facilitatenuclear entry of plasmid DNA. Therefore, we examinedthe effect of TCHD (a) on the nuclear import ofmacromolecules and pDNA and (b) on the transfectionefficiency of naked pDNA and non-viral nanoparticles,such as poly- and lipoplexes. In summary, we found thatTCHD was able to make the nuclear membrane perme-able for both high molecular weight dextrans and pDNAat non-toxic concentrations. Furthermore, TCHDenhanced the transfection efficiency of both nakedpDNA and DOTAP:DOPE-based lipoplexes, but had noeffect on the linear PEI-based polyplexes.
MATERIALS AND METHODS
Materials
Dulbecco’s modified Eagle’s medium (DMEM),L-glutamine (L-Gln), heat-inactivated fetal bovine serum(FBS), phosphate-buffered saline (PBS) and penicillin/streptomycin (P/S) were supplied by GibcoBRL(Merelbeke, Belgium). The secreted alkaline phosphatase(SEAP) expression plasmid was a kind gift fromProf. Dr Tavernier (Ghent University, Belgium) and22 kDa linear polyethyleneimine (PEI) from Prof.Dr Wagner (University of Munich, Germany). ThepGL3-control plasmid and luciferase assay kit wereobtained from Promega (Leiden, The Netherlands).One hundred fifty-eight kilodaltons of tetramethylrhoda-mine isothiocyanate-labeled dextran (TRITC-dextran)and TCHD were purchased from Sigma Aldrich.1,2-dioleoyl-3-trimethylammonium-propane (DOTAP)chloride salt, 1,2-dilinoleoyl-sn-glycero-3-phosphoethanolamine (DOPE) and 1,2-distearoyl-sn-glycero-3-phosphoethanolamine-N-[Amino(Polyethylene
Glycol)2000] (DSPE-PEG2000) were obtained from AvantiPolar Lipids (Alabaster, AL, USA).
Cell culture
A549 (lung carcinoma cells; ATCC number CCL-185) andVero (African green monkey cells; ATCC number CCL-81)cells were cultured inDMEMcontaining 2mML-Gln, 10%heat-inactivated FBS, 100U/ml P/S and grown at 378C ina humidified atmosphere containing 5% CO2.
Fluorescent labeling of pDNA
Plasmid DNA was covalently labeled with Cy5 using theLabelIT kit of Mirus Corporation (Madison, WI, USA)according to the manufacturer’s recommendations.Briefly, LabelIT reagent, containing Cy5, and 100 mgDNA were mixed in 1ml Hepes buffer (20mM Hepes,pH 7.4) at Cy5:DNA ratio (w:w) of 0.5:1 and incubated at378C for 1 h. Subsequently, the labeled pDNA wasseparated from unattached label by precipitation in thepresence of ethanol and 0.5M NaCl and reconstituted in20mM Hepes buffer (pH 7.4).
Microinjection studies
Microinjection studies were conducted using a Femtojet�
microinjector and an Injectman� NI 2 micromanipulator(Eppendorf, Hamburg, Germany). Vero cells were chosenfor these microinjection experiments as they have a well-defined nucleus and large cytoplasm. Vero cells (2.5� 104
cells/cm2) were plated onto sterile glass bottom culturedishes (MatTek Corporation, MA, USA) and allowed toadhere for 1 day. The cells were then washed with PBS andtransferred into 2ml serum-free medium supplementedwith 20mM Hepes (pH 7.4) to improve the bufferingcapacity of the medium during microinjection. Fivemicroliters 158 kDa TRITC-dextran (1mg/ml) orCy5-labeled pDNA (1mg/ml) was back-loaded intoFemptotip II microinjection needles and cells wereinjected using an injection pressure of 100 psi, a back-pressure of 30–50 psi and injection duration of 0.5 s.Where mentioned, the medium was replaced after micro-injection by TCHD-containing serum-free medium, sup-plemented with 20mM Hepes (pH 7.4).
To determine the cellular distribution, the fluorescencein the cells was visualized on different time points aftermicroinjection using a Nikon C1si confocal laser scanningmodule attached to a motorized Nikon TE2000-E invertedmicroscope (Nikon Benelux, Brussels, Belgium). With a60� water immersion objective and the 561 and 638 nmlaser lines for the excitation of the TRITC and Cy5 label,respectively. A non-confocal diascopic DIC (differentialinterference contrast) image was collected simultaneouslywith the confocal images.
For the z-scan analysis of the fluorescence aftercytoplasmic microinjection of the Cy5-labeled pDNA(Cy5-pDNA), the confocal volume (�1 fl) of a BioRadMRC 1024 CLSM (Hemel Hempstadt, UK) equippedwith the ConfoCor 2 fluorescence correlation spectro-scopy (FCS) setup (LSM510 ConfoCor 2, Zeiss,Gottingen, Germany) was positioned in a randomlyselected site in the nucleus. The fluorescence, along the
2 Nucleic Acids Research, 2007
by guest on March 3, 2014
http://nar.oxfordjournals.org/D
ownloaded from

z-axis at this selected XY site and perpendicular to the cellsurface, was recorded with the avalanche photodiodes ofthe ConfoCor 2 system before and every 10min afteraddition of 1% (w/v) TCHD dissolved in serum-freemedium supplemented with 20mM Hepes (pH 7.4).
Preparation of polyplexes and lipoplexes
Polyplexes consisting of 22 kDa linear PEI were preparedas described by Fayazpour et al. (39). Briefly, polyplexeswere prepared in 20mM Hepes pH 7.4 by adding the PEIall at once to the pDNA at a N/P ratio of 10.Subsequently, the mixture was vortexed for 10 s and thepolyplexes were allowed to equilibrate for 30min at roomtemperature prior to use. The final pDNA concentrationin the polyplexes was 0.126 mg/ml.
Liposomes composed of DOTAP:DOPE:DSPE-PEG2000 (molar ratio 5:5:0.2) were prepared as describedpreviously (40). Briefly, appropriate amounts of lipidswere dissolved in chloroform and mixed. The chloroformwas subsequently removed by rotary evaporation at 378Cfollowed by flushing the obtained lipid film with nitrogenduring 30min at room temperature. The dried lipids werethen hydrated by adding Hepes buffer till a final lipidconcentration of 10.2mM. After mixing in the presence ofglass beads, liposome formation was allowed overnightat 48C. Thereafter, the formed liposomes were extruded11 times through two stacked 100 nm polycarbonatemembrane filters (Whatman, Brentfort, UK) at roomtemperature using an Avanti Mini-Extruder (Avanti PolarLipids). The extruded liposomes were subsequently mixedwith pDNA in a �charge ratio of 4 and incubated at roomtemperature for 30min prior to use. The final pDNAconcentration in the lipoplex dispersion was 0.126 mg/ml.
Size and zeta potential measurements
The average particle size and zeta potential of theliposomes, lipoplexes and polyplexes were measured byphoton correlation spectroscopy (PCS) (Autosizer 4700,Malvern, Worcestershire, UK) and particle electrophor-esis (Zetasizer 2000, Malvern), respectively. The liposome,lipoplex and polyplex dispersions were diluted 40-fold inHepes buffer before the particle size and zeta potentialwere measured. The average (�standard error) of theliposomes and lipoplexes was 118� 1 and 242� 6 nm,respectively and their average zeta potential equaled26� 4 and 14� 1mV, respectively. The diameter andzeta potential of the linear PEI polyplexes were 165� 4 nmen 33� 2mV.
Cell viability assay
The influence of TCHD on the cell viability wasdetermined using the CellTiter-Glo� Assay (Promega)according to the manufacturer’s instructions. Briefly,2.5� 104 cells/cm2 were seeded in a 96-well plate andallowed to adhere. After 24 h, cells were washed with PBSand incubated with serum-free medium containingincreasing amounts of TCHD. After 1 h, the TCHD wasremoved and replaced by culture medium. After 48 h, theplate was incubated at room temperature for 30min and100 ml CellTiter-Glo� reagent was added to each well.
After shaking the plate for 10 and 2min incubation atroom temperature, the luminescence was measured on aGloMaxTM 96 luminometer with 1 s integration time.
Transfection experiments
Cells were seeded into 24-well plates at 2.5� 104 cells/cm2
and allowed to attach overnight. Subsequently, the culturemedium was removed, and after two washing steps withserum-free medium, 0.4 mg pDNA, polyplexes or lipo-plexes (both containing 0.4 mg pDNA) were added to eachwell. After 2 h the pDNA or non-viral nanoparticles wereremoved from the cells and the cells were post-incubatedfor 1 h with serum-free medium containing increasingamounts of TCHD. Subsequently, this medium wasreplaced by culture medium and the cells were furtherincubated at 378C. After 48 h both the SEAP (orluciferase) activity, as well as the total cellular proteinconcentration were measured.To determine the SEAP activity, 100ml of the culture
medium above the cells was taken and incubated at 658Cfor 30min. Subsequently, 100 ml dilution buffer (0.1Mglycine, 1mM MgCl2, 0.1mM ZnCl2, pH 10.4) and 15 ml4-methylumbelliferyl phosphate (4-MUP, 5.1 mg/ml indistilled water) was added. The obtained mixtures werethen incubated at 378C and the fluorescence was measuredon a Wallac Victor2 fluorescence plate reader (PerkinElmer-Cetus Life Sciences, Boston, MA) using an excita-tion and emission wavelength of 360 and 449 nm,respectively.To determine the luciferase activity of the cells, cells
were lysed with 80 ml 1�CCLR buffer (Promega) andluciferase activity was determined with the Promegaluciferase assay kit according to the manufacturer’sinstructions. Briefly, 100ml substrate was added to 20 mlsample and after a 2 s delay, the luminescence wasmeasured during 10 s with the GloMaxTM 96luminometer.To correct for the amount of cells per well, the protein
concentration was determined with the BCA kit (Pierce,Rockford, IL, USA). Two hundred microliters mastermix,containing 50 parts reagent A to 1 part B, was mixed with20 ml cell lysate and incubated at 378C for 30min and theabsorbance was measured on a Wallac Victor2 absorbanceplate reader (Perkin Elmer-Cetus Life Sciences) at 590 nm.
Statistics
The experimental data in this report are expressed as mean� standard deviation (SD). One way ANOVA was used todetermine whether data groups differed significantly fromeach other. A P-value 50.05 was considered statisticallysignificant.
RESULTS AND DISCUSSION
TCHD facilitates the nuclear accumulation ofdextrans and plasmid DNA
It has been demonstrated that TCHD enhances the rate ofnuclear entry of the maltose binding protein (38).However, this protein has a rather low molecular weight
Nucleic Acids Research, 2007 3
by guest on March 3, 2014
http://nar.oxfordjournals.org/D
ownloaded from
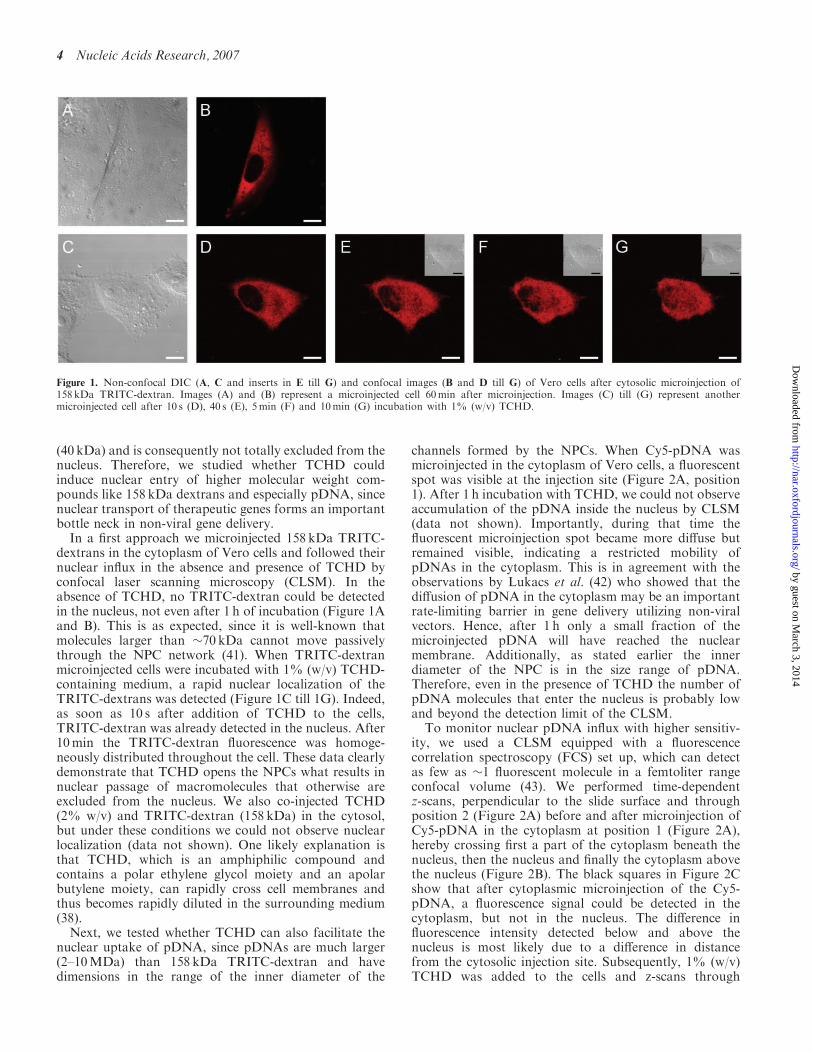
(40 kDa) and is consequently not totally excluded from thenucleus. Therefore, we studied whether TCHD couldinduce nuclear entry of higher molecular weight com-pounds like 158 kDa dextrans and especially pDNA, sincenuclear transport of therapeutic genes forms an importantbottle neck in non-viral gene delivery.In a first approach we microinjected 158 kDa TRITC-
dextrans in the cytoplasm of Vero cells and followed theirnuclear influx in the absence and presence of TCHD byconfocal laser scanning microscopy (CLSM). In theabsence of TCHD, no TRITC-dextran could be detectedin the nucleus, not even after 1 h of incubation (Figure 1Aand B). This is as expected, since it is well-known thatmolecules larger than �70 kDa cannot move passivelythrough the NPC network (41). When TRITC-dextranmicroinjected cells were incubated with 1% (w/v) TCHD-containing medium, a rapid nuclear localization of theTRITC-dextrans was detected (Figure 1C till 1G). Indeed,as soon as 10 s after addition of TCHD to the cells,TRITC-dextran was already detected in the nucleus. After10min the TRITC-dextran fluorescence was homoge-neously distributed throughout the cell. These data clearlydemonstrate that TCHD opens the NPCs what results innuclear passage of macromolecules that otherwise areexcluded from the nucleus. We also co-injected TCHD(2% w/v) and TRITC-dextran (158 kDa) in the cytosol,but under these conditions we could not observe nuclearlocalization (data not shown). One likely explanation isthat TCHD, which is an amphiphilic compound andcontains a polar ethylene glycol moiety and an apolarbutylene moiety, can rapidly cross cell membranes andthus becomes rapidly diluted in the surrounding medium(38).Next, we tested whether TCHD can also facilitate the
nuclear uptake of pDNA, since pDNAs are much larger(2–10MDa) than 158 kDa TRITC-dextran and havedimensions in the range of the inner diameter of the
channels formed by the NPCs. When Cy5-pDNA wasmicroinjected in the cytoplasm of Vero cells, a fluorescentspot was visible at the injection site (Figure 2A, position1). After 1 h incubation with TCHD, we could not observeaccumulation of the pDNA inside the nucleus by CLSM(data not shown). Importantly, during that time thefluorescent microinjection spot became more diffuse butremained visible, indicating a restricted mobility ofpDNAs in the cytoplasm. This is in agreement with theobservations by Lukacs et al. (42) who showed that thediffusion of pDNA in the cytoplasm may be an importantrate-limiting barrier in gene delivery utilizing non-viralvectors. Hence, after 1 h only a small fraction of themicroinjected pDNA will have reached the nuclearmembrane. Additionally, as stated earlier the innerdiameter of the NPC is in the size range of pDNA.Therefore, even in the presence of TCHD the number ofpDNA molecules that enter the nucleus is probably lowand beyond the detection limit of the CLSM.
To monitor nuclear pDNA influx with higher sensitiv-ity, we used a CLSM equipped with a fluorescencecorrelation spectroscopy (FCS) set up, which can detectas few as �1 fluorescent molecule in a femtoliter rangeconfocal volume (43). We performed time-dependentz-scans, perpendicular to the slide surface and throughposition 2 (Figure 2A) before and after microinjection ofCy5-pDNA in the cytoplasm at position 1 (Figure 2A),hereby crossing first a part of the cytoplasm beneath thenucleus, then the nucleus and finally the cytoplasm abovethe nucleus (Figure 2B). The black squares in Figure 2Cshow that after cytoplasmic microinjection of the Cy5-pDNA, a fluorescence signal could be detected in thecytoplasm, but not in the nucleus. The difference influorescence intensity detected below and above thenucleus is most likely due to a difference in distancefrom the cytosolic injection site. Subsequently, 1% (w/v)TCHD was added to the cells and z-scans through
Figure 1. Non-confocal DIC (A, C and inserts in E till G) and confocal images (B and D till G) of Vero cells after cytosolic microinjection of158 kDa TRITC-dextran. Images (A) and (B) represent a microinjected cell 60min after microinjection. Images (C) till (G) represent anothermicroinjected cell after 10 s (D), 40 s (E), 5min (F) and 10min (G) incubation with 1% (w/v) TCHD.
4 Nucleic Acids Research, 2007
by guest on March 3, 2014
http://nar.oxfordjournals.org/D
ownloaded from
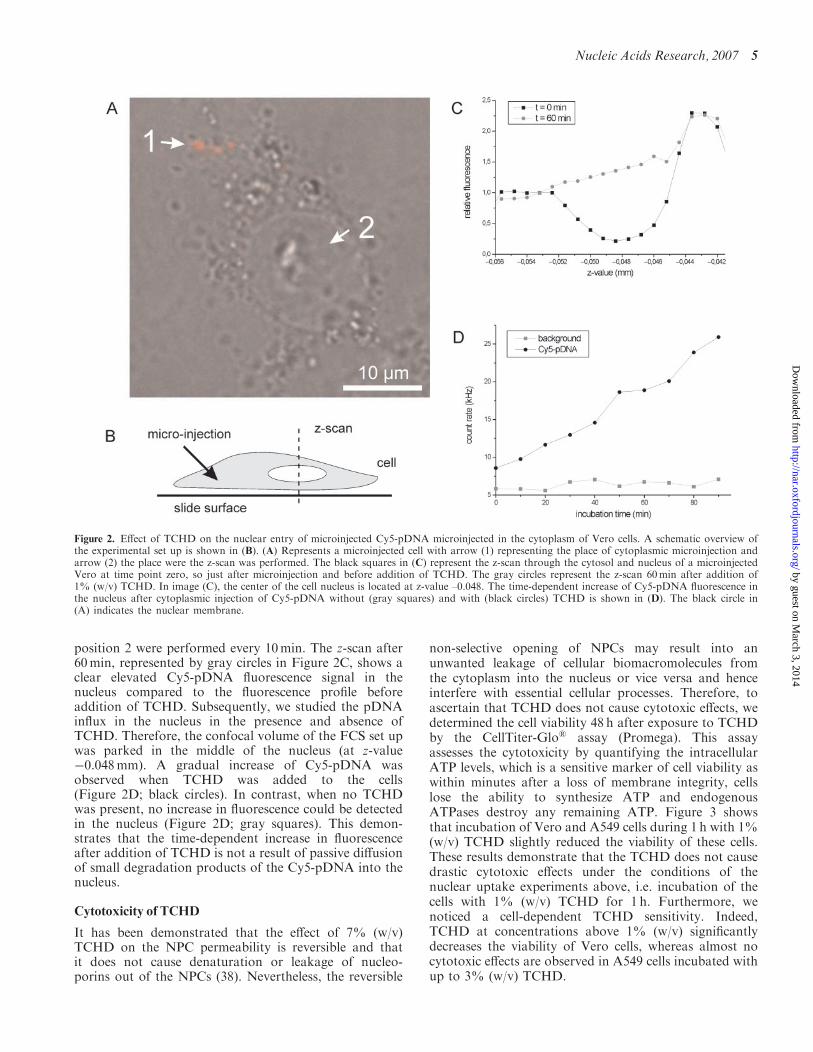
position 2 were performed every 10min. The z-scan after60min, represented by gray circles in Figure 2C, shows aclear elevated Cy5-pDNA fluorescence signal in thenucleus compared to the fluorescence profile beforeaddition of TCHD. Subsequently, we studied the pDNAinflux in the nucleus in the presence and absence ofTCHD. Therefore, the confocal volume of the FCS set upwas parked in the middle of the nucleus (at z-value�0.048mm). A gradual increase of Cy5-pDNA wasobserved when TCHD was added to the cells(Figure 2D; black circles). In contrast, when no TCHDwas present, no increase in fluorescence could be detectedin the nucleus (Figure 2D; gray squares). This demon-strates that the time-dependent increase in fluorescenceafter addition of TCHD is not a result of passive diffusionof small degradation products of the Cy5-pDNA into thenucleus.
Cytotoxicity of TCHD
It has been demonstrated that the effect of 7% (w/v)TCHD on the NPC permeability is reversible and thatit does not cause denaturation or leakage of nucleo-porins out of the NPCs (38). Nevertheless, the reversible
non-selective opening of NPCs may result into anunwanted leakage of cellular biomacromolecules fromthe cytoplasm into the nucleus or vice versa and henceinterfere with essential cellular processes. Therefore, toascertain that TCHD does not cause cytotoxic effects, wedetermined the cell viability 48 h after exposure to TCHDby the CellTiter-Glo� assay (Promega). This assayassesses the cytotoxicity by quantifying the intracellularATP levels, which is a sensitive marker of cell viability aswithin minutes after a loss of membrane integrity, cellslose the ability to synthesize ATP and endogenousATPases destroy any remaining ATP. Figure 3 showsthat incubation of Vero and A549 cells during 1 h with 1%(w/v) TCHD slightly reduced the viability of these cells.These results demonstrate that the TCHD does not causedrastic cytotoxic effects under the conditions of thenuclear uptake experiments above, i.e. incubation of thecells with 1% (w/v) TCHD for 1 h. Furthermore, wenoticed a cell-dependent TCHD sensitivity. Indeed,TCHD at concentrations above 1% (w/v) significantlydecreases the viability of Vero cells, whereas almost nocytotoxic effects are observed in A549 cells incubated withup to 3% (w/v) TCHD.
Figure 2. Effect of TCHD on the nuclear entry of microinjected Cy5-pDNA microinjected in the cytoplasm of Vero cells. A schematic overview ofthe experimental set up is shown in (B). (A) Represents a microinjected cell with arrow (1) representing the place of cytoplasmic microinjection andarrow (2) the place were the z-scan was performed. The black squares in (C) represent the z-scan through the cytosol and nucleus of a microinjectedVero at time point zero, so just after microinjection and before addition of TCHD. The gray circles represent the z-scan 60min after addition of1% (w/v) TCHD. In image (C), the center of the cell nucleus is located at z-value –0.048. The time-dependent increase of Cy5-pDNA fluorescence inthe nucleus after cytoplasmic injection of Cy5-pDNA without (gray squares) and with (black circles) TCHD is shown in (D). The black circle in(A) indicates the nuclear membrane.
Nucleic Acids Research, 2007 5
by guest on March 3, 2014
http://nar.oxfordjournals.org/D
ownloaded from

Influence of TCHD on the transfection efficiency ofnon-viral vectors
Since nuclear uptake of pDNA is considered as one of themajor barriers in non-viral gene delivery and since weshowed that TCHD could cause nuclear uptake of Cy5-pDNA, we wondered whether TCHD could enhance thetransfection efficiency of naked pDNA, cationic poly-plexes and lipoplexes. All transfection experiments wereperformed in two cell types, namely A549 cells (lungcarcinoma cell line) and Vero cells (kidney epithelial cellline), using different TCHD concentrations.The effect of TCHD on the transfection efficiency of
naked pDNA was evaluated by incubating Vero and A549cells with naked pDNA for 2 h. Subsequently, pDNA thatwas not incorporated into the cells was removed bywashing, and the cells were exposed to increasingpercentages (w/v) of TCHD for 1 h (Figure 4). Theincubation with TCHD clearly increased the gene expres-sion. This increase reached a maximal value at a TCHDpercentage of 0.5 and 1.5% in Vero (Figure 4A) and A549cells (Figure 4B), respectively. At these optimal concen-trations, a 3- and 66-fold increase in gene expression wasobserved in Vero and A549 cells, respectively. At higherpercentages no further increase and even a drop in geneexpression was observed. Most likely this indicates thatTCHD at these concentrations affects cellular processesthat are not detected by the MTT assay. Indeed, it hasbeen shown that the sensitivity of the MTT assay dependson the mechanism causing cytotoxicity (44). Between 0and 1.5% (w/v) TCHD, a gradual increase in geneexpression was observed in A549 cells (Figure 4B),which may indicate that the extent of NPC openingby TCHD is concentration dependent. Whether at 1.5%(w/v) TCHD a maximal opening of the NPCs is reached isnot certain, since above this concentration also cytotoxiceffects can play a role. This also explains the lower effectsof TCHD in Vero cells. Indeed, in these cells the optimal
concentration of TCHD to increase gene transfer is 0.5%.Based on the results in A549 cells, we can deduce that atsuch low TCHD concentration the opening of the NPChas not reached its maximum.
When TCHD was incubated together with the nakedpDNA, we also observed an increase in transfectionefficiency, but the effect was less pronounced compared tothe post-incubation experiments described earlier (datanot shown). This could be expected as it takes some timefor pDNA to become internalized by the cells and releasedfrom endocytotic vesicles into the cytoplasm.Additionally, as cytosolic nucleases degrade pDNA, theoptimal moment to add TCHD to the cells is immediatelyafter the release of the pDNA in the cytoplasm.
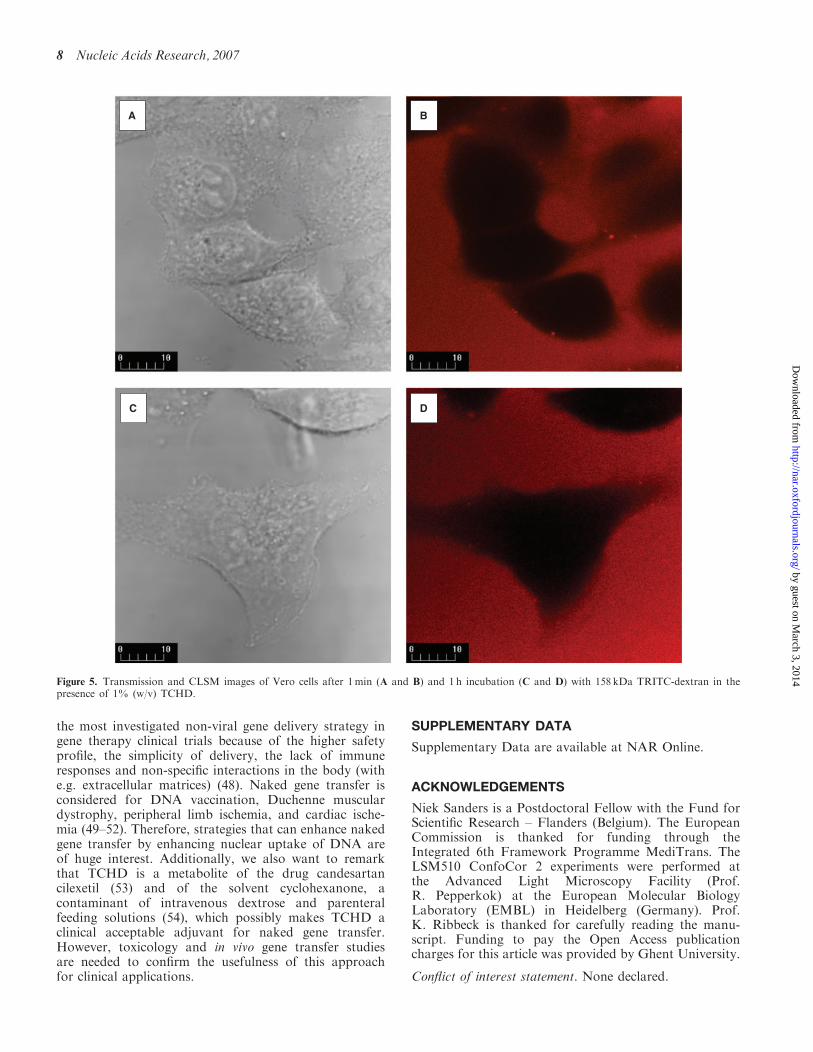
To exclude that the higher gene expression was due to aperforation of the cell membrane by TCHD, which wouldallow an increased internalization of pDNA through theplasma membrane, we tested whether TCHD causescytoplasmic entry of 158 kDa TRITC-dextrans. Figure 5shows that TRITC-dextrans did not enter the cells in theabsence nor in the presence of TCHD (1% w/v).Consequently, we can state that after 1 h incubationTCHD does not cause membrane perforation.
In a next step, we analyzed the effect of TCHD on thetransfection efficiency of linear PEI-based polyplexes, aquite efficient non-viral pDNA carrier that inducesendosomal release via the proton sponge mechanism(45). Surprisingly, none of the incubation protocols withTCHD resulted in a significant increase in gene expressionmediated by linear PEI-based polyplexes even notwhen the Vero and A549 cells were post-incubated with2% (w/v) TCHD (Supplementary Figure 1A and B).These data may indicate that, in agreement with Grosseet al. (46), linear PEI-based polyplexes are mainly releasedfrom the endosomes as intact complexes. Because of thelatter and taking into account the diameter of the linearPEI-based polyplexes (�165 nm), TCHD is not expected
Figure 3. Cell viability of Vero (black) and A549 cells (gray) after 1 h of incubation with TCHD. The asterix (�) represents data that significantlydiffer (P50.05) from the data point 0% (w/v) TCHD.
6 Nucleic Acids Research, 2007
by guest on March 3, 2014
http://nar.oxfordjournals.org/D
ownloaded from

to be able to enhance the nuclear transport and hencetransfection efficacy of such linear PEI-based polyplexes.
The endosomal escape mechanism of DOTAP:DOPE-based lipoplexes is based on a different mechanism andresults in the release of uncomplexed pDNA in the cytosol(47). This implies that, when the free pDNA reaches thenuclear membrane, TCHD should be able to induce itsnuclear translocation, similar to the naked pDNAtransfections. Therefore, we analyzed the effect ofTCHD on the transfection efficiency of DOTAP:DOPE-based lipoplexes. The same incubation protocols and cellswere used as in the experiments above. Incubating the cellswith TCHD for 1 h immediately after incubation with thelipoplexes, indeed caused an increase in transfectionefficiency in both Vero and A549 cells (Figure 6A andB). A maximal increase was observed when the Vero andA549 cells were incubated with 0.5 and 1.5% (w/v)TCHD, respectively. This increase is lower than observedwith naked pDNA transfection, which can be attributed tocell division. Compared to naked pDNA transfection,transfection with lipoplexes introduces a higher amount of
pDNA in the cytoplasm, which can translocate to thenucleus with higher probability either by entry through thepores or during cell division. Hence, the few extra copiesthat enter the nucleus after treating the cells with TCHDwill not tremendously increase gene expression. Incontrast, the amount of pDNA that reaches the cytoplasmand subsequently the nucleus is extremely low in case ofnaked pDNA transfection. Therefore, if TCHD can causenuclear translocation of only one pDNA molecule, thiseffect is much more spectacular.
CONCLUSION
In conclusion, our study demonstrates for the first timethat the amphipathic alcohol TCHD can be employed toenhance the nuclear uptake of pDNA by reversiblycollapsing the permeability barrier of the NPCs at non-toxic concentrations. Furthermore, our transfection datashow that TCHD has huge potential to enhance especiallynaked gene transfer. Although less efficient than carrier-mediated gene delivery, naked gene transfer is currently
Figure 4. Transfection efficiency of naked pDNA in Vero (A) and A549 (B) cells post-incubated for 1 h with different TCHD concentrations. Theasterisk (�) represents data that significantly differ (P50.05) from the data point 0% (w/v) TCHD.
Nucleic Acids Research, 2007 7
by guest on March 3, 2014
http://nar.oxfordjournals.org/D
ownloaded from
the most investigated non-viral gene delivery strategy ingene therapy clinical trials because of the higher safetyprofile, the simplicity of delivery, the lack of immuneresponses and non-specific interactions in the body (withe.g. extracellular matrices) (48). Naked gene transfer isconsidered for DNA vaccination, Duchenne musculardystrophy, peripheral limb ischemia, and cardiac ische-mia (49–52). Therefore, strategies that can enhance nakedgene transfer by enhancing nuclear uptake of DNA areof huge interest. Additionally, we also want to remarkthat TCHD is a metabolite of the drug candesartancilexetil (53) and of the solvent cyclohexanone, acontaminant of intravenous dextrose and parenteralfeeding solutions (54), which possibly makes TCHD aclinical acceptable adjuvant for naked gene transfer.However, toxicology and in vivo gene transfer studiesare needed to confirm the usefulness of this approachfor clinical applications.
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online.
ACKNOWLEDGEMENTS
Niek Sanders is a Postdoctoral Fellow with the Fund forScientific Research – Flanders (Belgium). The EuropeanCommission is thanked for funding through theIntegrated 6th Framework Programme MediTrans. TheLSM510 ConfoCor 2 experiments were performed atthe Advanced Light Microscopy Facility (Prof.R. Pepperkok) at the European Molecular BiologyLaboratory (EMBL) in Heidelberg (Germany). Prof.K. Ribbeck is thanked for carefully reading the manu-script. Funding to pay the Open Access publicationcharges for this article was provided by Ghent University.
Conflict of interest statement. None declared.
A B
DC
Figure 5. Transmission and CLSM images of Vero cells after 1min (A and B) and 1 h incubation (C and D) with 158 kDa TRITC-dextran in thepresence of 1% (w/v) TCHD.
8 Nucleic Acids Research, 2007
by guest on March 3, 2014
http://nar.oxfordjournals.org/D
ownloaded from

REFERENCES
1. Capecchi,M.R. (1980) High efficiency transformation by directmicroinjection of DNA into cultured mammalian cells. Cell,22, 479–488.
2. Dowty,M.E., Williams,P., Zhang,G., Hagstrom,J.E. and Wolff,J.A.(1995) Plasmid DNA entry into postmitotic nuclei of primary ratmyotubes. Proc. Natl Acad. Sci. USA, 92, 4572–4576.
3. Pollard,H., Remy,J.S., Loussouarn,G., Demolombe,S., Behr,J.P.and Escande,D. (1998) Polyethylenimine but not cationic lipidspromotes transgene delivery to the nucleus in mammalian cells.J. Biol. Chem., 273, 7507–7511.
4. Brunner,S., Sauer,T., Carotta,S., Cotten,M., Saltik,M. andWagner,E. (2000) Cell cycle dependence of gene transfer bylipoplex, polyplex and recombinant adenovirus. Gene Ther.,7, 401–407.
5. Mortimer,I., Tam,P., MacLachlan,I., Graham,R.W.,Saravolac,E.G. and Joshi,P.B. (1999) Cationic lipid-mediatedtransfection of cells in culture requires mitotic activity. Gene Ther.,6, 403–411.
6. Wilke,M., Fortunati,E., van den,B.M., Hoogeveen,A.T. andScholte,B.J. (1996) Efficacy of a peptide-based gene delivery systemdepends on mitotic activity. Gene Ther., 3, 1133–1142.
7. Laskey,R.A. (1998) CIBA Medal Lecture. Regulatory roles of thenuclear membrane. Biochem. Soc. Trans., 26, 561–567.
8. Vasu,S.K. and Forbes,D.J. (2001) Nuclear pores and nuclearassembly. Curr. Opin. Cell Biol., 13, 363–375.
9. Gorlich,D., Henklein,P., Laskey,R.A. and Hartmann,E. (1996) A 41amino acid motif in importin-alpha confers binding to importin-beta and hence transit into the nucleus. EMBO J., 15, 1810–1817.
10. Jakel,S., Albig,W., Kutay,U., Bischoff,F.R., Schwamborn,K.,Doenecke,D. and Gorlich,D. (1999) The importin beta/importin7 heterodimer is a functional nuclear import receptor for histoneH1. EMBO J., 18, 2411–2423.
11. Nagasaki,T., Kawazu,T., Tachibana,T., Tamagaki,S. and Shinkai,S.(2005) Enhanced nuclear import and transfection efficiency ofplasmid DNA using streptavidin-fused importin-beta.J. Control Release, 103, 199–207.
12. Collas,P. and Alestrom,P. (1998) Nuclear localization signalsenhance germline transmission of a transgene in zebrafish.Transgenic Res., 7, 303–309.
13. Subramanian,A., Ranganathan,P. and Diamond,S.L. (1999)Nuclear targeting peptide scaffolds for lipofection of nondividingmammalian cells. Nat. Biotechnol., 17, 873–877.
14. Liang,M.R., Alestrom,P. and Collas,P. (2000) Glowing zebrafish:integration, transmission, and expression of a single luciferasetransgene promoted by noncovalent DNA-nuclear transport peptidecomplexes. Mol. Reprod. Dev., 55, 8–13.
15. Bremner,K.H., Seymour,L.W., Logan,A. and Read,M.L. (2004)Factors influencing the ability of nuclear localization sequence
Figure 6. Transfection efficiency of lipoplexes in Vero (A) and A549 (B) cells post-incubated for 1 h with different TCHD concentrations. The asterisk(�) represents data that significantly differ (P50.05) from the data point 0% (w/v) TCHD.
Nucleic Acids Research, 2007 9
by guest on March 3, 2014
http://nar.oxfordjournals.org/D
ownloaded from

peptides to enhance nonviral gene delivery. Bioconjug. Chem.,15, 152–161.
16. Mesika,A., Kiss,V., Brumfeld,V., Ghosh,G. and Reich,Z. (2005)Enhanced intracellular mobility and nuclear accumulation of DNAplasmids associated with a karyophilic protein. Hum. Gene Ther.,16, 200–208.
17. Arenal,A., Pimentel,R., Garcia,C., Pimentel,E. and Alestrom,P.(2004) The SV40T antigen nuclear localization sequence enhancesnuclear import of vector DNA in embryos of a crustacean(Litopenaeus schmitti). Gene, 337, 71–77.
18. Sebestyen,M.G., Ludtke,J.J., Bassik,M.C., Zhang,G., Budker,V.,Lukhtanov,E.A., Hagstrom,J.E. and Wolff,J.A. (1998) DNA vectorchemistry: the covalent attachment of signal peptides to plasmidDNA. Nat. Biotechnol., 16, 80–85.
19. Zanta,M.A., Belguise-Valladier,P. and Behr,J.P. (1999) Genedelivery: a single nuclear localization signal peptide is sufficient tocarry DNA to the cell nucleus. Proc. Natl Acad. Sci. USA,96, 91–96.
20. Neves,C., Byk,G., Scherman,D. and Wils,P. (1999) Coupling of atargeting peptide to plasmid DNA by covalent triple helixformation. FEBS Lett., 453, 41–45.
21. Tanimoto,M., Kamiya,H., Minakawa,N., Matsuda,A. andHarashima,H. (2003) No enhancement of nuclear entry by directconjugation of a nuclear localization signal peptide to linearizedDNA. Bioconjug. Chem., 14, 1197–1202.
22. van der Aa,M., Koning,G., van der Gugten,J., d’Oliveira,C.,Oosting,R., Hennink,W.E. and Crommelin,D.J. (2005) Covalentattachment of an NLS-peptide to linear dna does not enhancetransfection efficiency of cationic polymer based gene deliverysystems. J. Control Release, 101, 395–397.
23. Chan,C.K., Hubner,S., Hu,W. and Jans,D.A. (1998) Mutualexclusivity of DNA binding and nuclear localization signalrecognition by the yeast transcription factor GAL4: implications fornonviral DNA delivery. Gene Ther., 5, 1204–1212.
24. Vaysse,L., Harbottle,R., Bigger,B., Bergau,A., Tolmachov,O. andCoutelle,C. (2004) Development of a self-assembling nucleartargeting vector system based on the tetracycline repressor protein.J. Biol. Chem., 279, 5555–5564.
25. Branden,L.J., Mohamed,A.J. and Smith,C.I. (1999) A peptidenucleic acid-nuclear localization signal fusion that mediates nucleartransport of DNA. Nat. Biotechnol., 17, 784–787.
26. Branden,L.J., Christensson,B. and Smith,C.I. (2001) In vivo nucleardelivery of oligonucleotides via hybridizing bifunctional peptides.Gene Ther., 8, 84–87.
27. Ludtke,J.J., Zhang,G., Sebestyen,M.G. and Wolff,J.A. (1999)A nuclear localization signal can enhance both the nuclear transportand expression of 1 kb DNA. J. Cell Sci., 112 (Pt 12), 2033–2041.
28. Chan,C.K. and Jans,D.A. (1999) Enhancement of polylysine-mediated transferrinfection by nuclear localization sequences:polylysine does not function as a nuclear localization sequence.Hum. Gene Ther., 10, 1695–1702.
29. Chan,C.K., Senden,T. and Jans,D.A. (2000) Supramolecular struc-ture and nuclear targeting efficiency determine the enhancement oftransfection by modified polylysines. Gene Ther., 7, 1690–1697.
30. Carlisle,R.C., Bettinger,T., Ogris,M., Hale,S., Mautner,V. andSeymour,L.W. (2001) Adenovirus hexon protein enhances nucleardelivery and increases transgene expression of polyethylenimine/plasmid DNA vectors. Mol. Ther., 4, 473–483.
31. Hagstrom,J.E., Sebestyen,M.G., Budker,V., Ludtke,J.J., Fritz,J.D.and Wolff,J.A. (1996) Complexes of non-cationic liposomes andhistone H1 mediate efficient transfection of DNA without encap-sulation. Biochim. Biophys. Acta, 1284, 47–55.
32. Aronsohn,A.I. and Hughes,J.A. (1998) Nuclear localization signalpeptides enhance cationic liposome-mediated gene therapy.J. Drug Target, 5, 163–169.
33. Murray,K.D., Etheridge,C.J., Shah,S.I., Matthews,D.A.,Russell,W., Gurling,H.M. and Miller,A.D. (2001) Enhanced catio-nic liposome-mediated transfection using the DNA-binding peptidemu (mu) from the adenovirus core. Gene Ther., 8, 453–460.
34. Keller,M., Harbottle,R.P., Perouzel,E., Colin,M., Shah,I.,Rahim,A., Vaysse,L., Bergau,A., Moritz,S. et al. (2003)Nuclear localisation sequence templated nonviral gene delivery
vectors: investigation of intracellular trafficking events of LMD andLD vector systems. Chembiochem, 4, 286–298.
35. Preuss,M., Tecle,M., Shah,I., Matthews,D.A. and Miller,A.D.(2003) Comparison between the interactions of adenovirus-derivedpeptides with plasmid DNA and their role in gene delivery mediatedby liposome-peptide-DNA virus-like nanoparticles.Org. Biomol. Chem., 1, 2430–2438.
36. Wiseman,J.W., Scott,E.S., Shaw,P.A. and Colledge,W.H. (2005)Enhancement of gene delivery to human airway epithelial cellsin vitro using a peptide from the polyoma virus protein VP1.J. Gene Med., 7, 759–770.
37. Akuta,T., Eguchi,A., Okuyama,H., Senda,T., Inokuchi,H.,Suzuki,Y., Nagoshi,E., Mizuguchi,H., Hayakawa,T. et al. (2002)Enhancement of phage-mediated gene transfer by nuclear localiza-tion signal. Biochem. Biophys. Res. Commun., 297, 779–786.
38. Ribbeck,K. and Gorlich,D. (2002) The permeability barrier ofnuclear pore complexes appears to operate via hydrophobicexclusion. EMBO J., 21, 2664–2671.
39. Fayazpour,F., Lucas,B., varez-Lorenzo,C., Sanders,N.N.,Demeester,J. and De Smedt,S.C. (2006) Physicochemical andtransfection properties of cationic hydroxyethylcellulose/DNAnanoparticles. Biomacromolecules, 7, 2856–2862.
40. Sanders,N.N., Van,R.E., De Smedt,S.C. and Demeester,J. (2001)Structural alterations of gene complexes by cystic fibrosis sputum.Am. J. Respir. Crit. Care Med., 164, 486–493.
41. Galy,V., Mattaj,I.W. and Askjaer,P. (2003) Caenorhabditis elegansnucleoporins Nup93 and Nup205 determine the limit of nuclearpore complex size exclusion in vivo. Mol. Biol. Cell, 14, 5104–5115.
42. Lukacs,G.L., Haggie,P., Seksek,O., Lechardeur,D., Freedman,N.and Verkman,A.S. (2000) Size-dependent DNA mobility in cyto-plasm and nucleus. J. Biol. Chem., 275, 1625–1629.
43. Schwille,P. (2001) Fluorescence correlation spectroscopy and itspotential for intracellular applications. Cell Biochem. Biophys.,34, 383–408.
44. Weyermann,J., Lochmann,D. and Zimmer,A. (2005) A practicalnote on the use of cytotoxicity assays. Int. J. Pharm., 288, 369–376.
45. Boussif,O., Lezoualc’h,F., Zanta,M.A., Mergny,M.D.,Scherman,D., Demeneix,B. and Behr,J.P. (1995) A versatile vectorfor gene and oligonucleotide transfer into cells in culture andin vivo: polyethylenimine. Proc. Natl Acad. Sci. USA,92, 7297–7301.
46. Grosse,S., Thevenot,G., Monsigny,M. and Fajac,I. (2006) Whichmechanism for nuclear import of plasmid DNA complexed withpolyethylenimine derivatives? J. Gene Med., 8, 845–851.
47. Koltover,I., Salditt,T., Radler,J.O. and Safinya,C.R. (1998) Aninverted hexagonal phase of cationic liposome-DNA complexesrelated to DNA release and delivery. Science, 281, 78–81.
48. Wolff,J.A. and Budker,V. (2005) The mechanism of naked DNAuptake and expression. Adv. Genet., 54, 3–20.
49. Davis,H.L., Schleef,M., Moritz,P., Mancini,M., Schorr,J. andWhalen,R.G. (1996) Comparison of plasmid DNA preparationmethods for direct gene transfer and genetic immunization.Biotechniques, 21, 92–99.
50. Romero,N.B., Benveniste,O., Payan,C., Braun,S., Squiban,P.,Herson,S. and Fardeau,M. (2002) Current protocol of a researchphase I clinical trial of full-length dystrophin plasmid DNA inDuchenne/Becker muscular dystrophies. Part II: clinical protocol.Neuromuscul. Disord., 12(Suppl. 1), S45–S48.
51. Tsurumi,Y., Takeshita,S., Chen,D., Kearney,M., Rossow,S.T.,Passeri,J., Horowitz,J.R., Symes,J.F. and Isner,J.M. (1996) Directintramuscular gene transfer of naked DNA encoding vascularendothelial growth factor augments collateral development andtissue perfusion. Circulation, 94, 3281–3290.
52. Symes,J.F. (2001) Gene therapy for ischemic heart disease:therapeutic potential. Am. J. Cardiovasc. Drugs, 1, 159–166.
53. Yamamoto,K., Kitayoshi,T., Nishimura,S., Chatani,F. andWatanabe,T. (2003) Absence of interactive effects of trans-1,2-cyclohexanediol, a major metabolite of the side-chain ofcandesartan cilexetil, on digoxin-induced arrhythmias in dogs.J. Pharmacol. Sci., 92, 387–399.
54. Mills,G.A. and Walker,V. (1990) Urinary excretion of cyclohex-anediol, a metabolite of the solvent cyclohexanone, by infants in aspecial care unit. Clin. Chem., 36, 870–874.
10 Nucleic Acids Research, 2007
by guest on March 3, 2014
http://nar.oxfordjournals.org/D
ownloaded from
Top Related
Copyright © 2022 FDOKUMEN

