







Bahasa
Halaman
Hukum


This article was downloaded by: [Valentina Losi]On: 25 May 2012, At: 14:08Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Advances in Oceanography andLimnologyPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/taol20
Nematodes as indicators ofenvironmental quality in seagrass(Posidonia oceanica) meadows of theNW Mediterranean SeaValentina Losi a , Monica Montefalcone a , Mariapaola Moreno a ,Elisa Giovannetti a , Luigi Gaozza a , Matia Grondona a & GiancarloAlbertelli aa Dipartimento di Scienze della Terra, dell'Ambiente e della Vita(DISTAV), Università di Genova, Corso Europa 26, 16132 Genova,Italy
Available online: 25 May 2012
To cite this article: Valentina Losi, Monica Montefalcone, Mariapaola Moreno, Elisa Giovannetti,Luigi Gaozza, Matia Grondona & Giancarlo Albertelli (2012): Nematodes as indicators ofenvironmental quality in seagrass (Posidonia oceanica) meadows of the NW Mediterranean Sea,Advances in Oceanography and Limnology, 3:1, 69-91
To link to this article: http://dx.doi.org/10.1080/19475721.2012.670666
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representationthat the contents will be complete or accurate or up to date. The accuracy of anyinstructions, formulae, and drug doses should be independently verified with primarysources. The publisher shall not be liable for any loss, actions, claims, proceedings,

demand, or costs or damages whatsoever or howsoever caused arising directly orindirectly in connection with or arising out of the use of this material.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

Advances in Oceanography and LimnologyVol. 3, No. 1, June 2012, 69–91
Nematodes as indicators of environmental quality in seagrass
(Posidonia oceanica) meadows of the NW Mediterranean Sea
Valentina Losi*, Monica Montefalcone, Mariapaola Moreno, Elisa Giovannetti,Luigi Gaozza, Matia Grondona and Giancarlo Albertelli
Dipartimento di Scienze della Terra, dell’Ambiente e della Vita (DISTAV),Universita di Genova, Corso Europa 26, 16132 Genova, Italy
(Received 30 November 2011; final version received 24 February 2012)
Meiofauna in sediments colonized by Posidonia oceanica in the Ligurian Sea (NWMediterranean) was investigated to evaluate its use as an early warning indicatorof environmental disturbance. Descriptors commonly used in seagrass healthassessment are mainly related to the plant (e.g., phenological parameters) and/orto the meadow (e.g., structural parameters) and are subjected to long-termresponse times. Conversely, meiofauna is considered a good bioindicator witha rapid response to environmental variations, due to its main features (e.g., shortlife cycle, scarce mobility, presence/absence of tolerant/sensitive species).Meiofaunal community, sediment characteristics and organic matter descriptorsin three meadows located in urbanized coastal areas were contrasted with thoseobserved in three meadows located within Marine Protected Areas. Samplingswere carried out in two distinct periods, at the beginning and at the end of thesummer season, in order to individuate early changes in the meiofaunalassemblages. The meiofauna and, particularly, the nematode assemblagecomposition and diversity, highlighted differences among meadows at the endof the summer not detectable by organic matter descriptors and structuralparameters of the meadow (e.g. shoot density). Nematodes are, thus, proposed asappropriate biological quality elements (BQEs) able to show early responsesto environmental disturbance.
Keywords: Meiofauna; Nematodes; Posidonia oceanica; early warning response;biological quality element; Mediterranean Sea
Introduction
The endemic Posidonia oceanica (L.) Delile, the most important and abundant seagrassin the Mediterranean Sea, has shown an alarming decline in many areas during the lastcentury [1–3], especially in the North-Western Mediterranean, where many meadows havebeen definitively lost in recent decades [4, 5]. Regression of meadows results from bothdirect human impacts, through physical damages to the meadows [6] and indirect humaneffects, trough the impact on water quality and sediments dynamic [7].
In Liguria, an administrative region of NW Italy, the intense urban and industrialdevelopment of the coastline since the 1960s, as well as the high tourist pressure, led to astrong regression of the P. oceanica meadows [8–10], which originally developed as awide and continuous strip along the coast [11]. Indeed, Montefalcone et al. [3] estimated
*Corresponding author. Email: [email protected]
ISSN 1947–5721 print/ISSN 1947–573X online
� 2012 Taylor & Francis
http://dx.doi.org/10.1080/19475721.2012.670666
http://www.tandfonline.com
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

that the 40% of the original extent of the Ligurian meadows has been lost, with the 10%in the last 16 years. This constant rate of decline is dramatic considering the ecologicalimportance of P. oceanica meadows and the legal protection they are subjected: meadowshave been defined as priority natural habitat under the European Community Directive92/43/EEC [12], whose conservation requires the designation of special areas identifiedas Sites of Community Interest (SCIs). P. oceanica meadows are among the mostproductive marine ecosystems, they protect coastal areas and stabilise beaches andseafloors weakening the water movement, support a very rich associated community,playing an important role in structuring pelagic and benthic assemblages, and createcomplex trophic interactions among organisms [13 and refs. therein].
Due to the key role of seagrasses in coastal areas, maintenance of existing meadowsis essential [1], especially considering that those ecosystems are fragile, subjected to manyhuman disturbances and characterized by long time recovery. European Community asksfor those habitats included in SCIs to be maintained in a satisfactory condition and to beproperly managed; thus, to ensure the effective conservation of seagrasses, appropriatemonitoring programs are required. In this perspective early warning indicators, able torapidly detect environmental alterations, would allow prompt management actions thatmight avoid alterations hardly recoverable at the plant and/or the meadow level.
Descriptors commonly used to evaluate seagrass meadows health status are mainlyrelated to the plant (e.g., phenological parameters) and/or to the meadow (e.g., structuralparameters) and are, therefore, subjected to long-term response times. Giovannetti et al.[14] proposed the use of the community structure of leaves’ epiphytes as an early warningindicator, able to respond in a short time to environmental disturbance.
In this study we evaluated the use of meiofaunal community, with a particularattention to Nematodes, as early warning indicators of environmental disturbances.Meiofauna, representing the main link between higher and lower trophic levels of foodwebs [15], plays an important role in the dynamic of seagrass meadows, where it isinvolved in the decomposition of organic matter and regeneration of nutrients andrepresents a food source for higher trophic levels [16]. The meiofaunal community hasproven to be extremely useful in assessing the effects of anthropogenic disturbancein marine sediments due to its main features (e.g., short life cycle, rapid development,scarce mobility, absence of pelagic life stages, presence of tolerant/sensitive species)[e.g., 17–20]. Within meiofaunal taxa, Nematodes offer a variety of possibilities forassessing changes in community structure, due to their high structural and functionaldiversity. As species or genus composition is comparatively more sensitive to anydisturbance [21], Nematodes have already been employed in biomonitoring studies andresulted suitable indicators for pollution-induced disturbances of benthic ecosystems[e.g., 22–26]. To date, very few studies investigated meiofaunal organisms of P. oceanicameadows [20, 27–32].
The main aims of this paper were: (i) evaluate the diversity of meiofaunal and nematodecommunity in P. oceanica meadows subjected to a different degree of human disturbance;(ii) compare the response of meiofaunal organisms to disturbance with the response of somecommonly adopted descriptors at the plant (e.g., leaf biometry) and at the meadow (e.g.,shoot density) level. The meiofauna and nematode communities of sediments associated tothree Ligurian P. oceanicameadows located inMarine Protected Areas were compared withthose associated to three meadows located closely to urbanised coastal areas. Since theexponential increase in the tourist frequentation during the summer season (from June toSeptember) in Liguria region has been shown to enhance urban sewage outfalls and to
70 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

negatively affect water quality [14], samplings were carried out in two distinct periods, at thebeginning and at the end of the summer season, in order to evaluate the short time responseof the meiofaunal community to this kind of disturbance.
Materials and methods
Study area
The study was carried out at a region-wide spatial scale in six Posidonia oceanica meadowsalong the Ligurian coast (NW Mediterranean Sea). Following the same sampling designadopted by Giovannetti et al. [14], three P. oceanica meadows were selected among thoselocated in virtually ‘‘natural’’ areas, where natural and human sources of disturbance werelow [33, 34] and where the degree of naturalness of marine coastal environment, i.e. healthstatus of coastal ecosystems [35], was high to very high, so that they were included inMarine Protected Areas (afterward called ‘‘MPA’’). Similarly, three meadows wereselected in urbanised areas having a comparatively lower degree of naturalness, a higherdegree of human presence and where major sources of disturbance occur (afterward called‘‘Urban’’). The meadows of Mortola (MO), Camogli (CA) and Monterosso al Mare (MM)belong to the MPA condition, whereas the meadows of Arma di Taggia (AT), Bogliasco(BO) and Pieve Ligure (PL) to the Urban condition (Figure 1). In particular, the meadowsof Bogliasco (BO) and Pieve Ligure (PL) were located in correspondence of urban sewageswhere the drainage pipes cross the meadow extent and discharge at about 20m depthwhere the meadow still occurs. The Arma di Taggia (AT) meadow was located incorrespondence of the mouth of the Armea stream, a small river that conveys theindustrial and urban waters into the sea.
Field activities
Samplings were carried out in two distinct periods: at the beginning (May 2007) and at theend (September 2007) of the summer season. In each meadow two sites (1 and 2), spaced
Figure 1. Geographic location of the six meadows investigated indicated by their code:MO¼Mortola; AT¼Arma di Taggia; BO¼Bogliasco; PL¼Pieve Ligure; CA¼Camogli;MM¼Monterosso al Mare.
Advances in Oceanography and Limnology 71
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

about 500m from each other, were randomly selected at the intermediate meadow depth(from about 12 to 15m depth), which was considered as the optimal depth for the plantgrowth [36, 37]. For each site and in each sampling period four replicate cores of sedimentsamples were collected by SCUBA divers to assess sediment grain size, sedimentaryorganic matter concentration and biochemical composition as well as abundance andcomposition of meiofaunal assemblages, with particular attention to Nematodes, withinthe same sites that formed the subject of the previous study by Giovannetti et al. [14]. Allanalyses were carried out on the surface sediments layer (0–2 cm).
The health status of the six meadows was assessed by Giovannetti et al. [14], whommeasured, contextually with our sediment samplings, shoot density, P. oceanica and deadmatte cover to be computed in the Conservation Index [38] and sampled ten shoots for leafphenology and epiphyte community analyses.
Grain size
A sediment aliquot (about 200 g) for each sampling was pre-treated with an H2O2
solution (10%) for a period of about 24–48 h, in order to eliminate the soluble salts and theorganic matter in the sediments, and was dried at 60�C for 48 h. Particle size profileswere determined by dry-sieving analysis, using a set of standard sieves (Endecotts Ltd)following the Udden and Wentworth scale [39]. The disaggregated sample was passedthrough the sieves, shaken by an automatic shaker, for a standard time period (usually15min). The sample remaining in each sieve was subsequently weighed.
Sedimentary organic matter concentration and biochemical composition
Sediments for the organic matter determinations were stored at �20�C. Total organicmatter (TOM) was determined by ignition loss according to Parker [40]. Proteinconcentration (Prt) was assessed according to the Hartree [41] method, modified byFabiano et al. [42]. Bovine albumin solutions were used as standards and absorbance wasdetermined at 650 nm. Carbohydrate concentration (Cho) was analyzed according toDubois et al. [43], modified for the sediment application by Gerchakov and Hatcher [44].D(þ) glucose solutions were used as standards and absorbance was measured at 490 nm.For each analysis, sediment blanks were performed following the same method, withsediment pre-treated at 550�C for 4 h. Concentrations were expressed as mg g�1 ofsediment dry weight (DW). The value of the protein to carbohydrate ratio (Prt/Cho) waschosen as descriptor of the quality of sediment organic matter.
Meiofaunal community
Samples were fixed with 4% buffered formaldehyde in 0.4 mm pre-filtered seawatersolution. In the laboratory, all meiofaunal samples were rinsed with a gentle jet of freshwater over a 0.5mm sieve to exclude macrofauna, decanted over a 38 mm sieve ten times,centrifuged three times with Ludox HS40 (specific density 1.18 g cm�3, [45]), and stainedwith Rose Bengal (0.5 gL�1). Meiofaunal organisms were counted and identified at themajor taxon level, using a stereomicroscope. The rare taxa were defined as the taxathat represented 51% of the total meiofaunal abundance of all investigated samples.
72 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

Diversity was evaluated as the richness of taxa and as the Shannon-Wiener Index (H’,log 2 based) at each site.
Nematode assemblages
One-hundred Nematodes of each replicate were randomly picked out using fine pin undera stereomicroscope (magnification� 40), transferred from formalin to glycerol througha series of ethanol-glycerol solutions, and mounted on slides in anhydrous glycerine,following the procedure described by Heip et al. [45]. Nematodes were identified at genuslevel by using the pictorial keys of Platt and Warwick [46, 47] and Warwick et al. [48], aswell as the NeMys online identification key [49].
Nematode biomass was estimated using the biovolumes method according to theequation:
V ¼ 530� L�W2
[50], where V is the volume (nl), L is the total length (mm, excluding filiform tails, whenpresent) and W the width (mm) of a given Nematode. Nematodes wet weights (mm WW)were obtained using a specific gravity of 1.13mg nl�1 [51] and converted into dry weighs(mg DW) assuming a dry/weight ratio of 0.25 [52].
Diversity indices (richness of genera and H’) were calculated as for meiofauna.Nematode genera were classified according to Wieser [53] into four feeding groups to
investigate the trophic structure of the community: selective (1A) and non-selective (1B)deposit-feeders, epistrate-feeders (2A), and predators/omnivores (2B). The Index ofTrophic Diversity (ITD) was calculated according to Heip et al. [45], ranging from 0.25(the highest trophic diversity) to 1.0 (the lowest trophic diversity).
On the basis of the threshold values proposed by Moreno et al. [54], the EcologicalQuality Status (EQS) of the six meadows in both periods was assessed using nematode H’,ITD and genera composition as indicators.
Statistical analyses
Differences between the MPA and the Urban meadows in the two distinct periods in grainsize, sedimentary organic matter concentration and biochemical composition, meiofaunaland nematode abundance and diversity, biomass and trophic groups for Nematodes, wereanalysed using 4-way analyses of variance (ANOVAs). Factor 1 was the Period (fixed, twolevels: May and September), factor 2 was the Condition (orthogonal and fixed, two levels:MPA and Urban), factor 3 was the Meadow (random and nested in Condition, threelevels), factor 4 was the Site (random and nested in Meadow, two levels: 1 and 2) with4 replicates per site. Prior to analysis, the normality and homoscedasticity assumptionswere checked using the Shapiro–Wilk and the Bartlett’s test and, when required, the datawere appropriately transformed. When a treatment factor was significant, the differencesbetween levels were determined using the Student–Newman–Keuls test (SNK test).
Asymmetrical multivariate analysis of variance based on permutations(PERMANOVA) [55] was used to examine differences between the MPA and theUrban meadows in the two distinct periods in: (i) meiofaunal taxonomic composition(separately for the entire community and rare taxa), (ii) nematode genera composition and
Advances in Oceanography and Limnology 73
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

(iii) trophic composition of nematode assemblages. Factors and levels considered were thesame described for the univariate analyses.
A SIMPER (Similarity Percentages-species contributions) analysis based on theBray–Curtis similarity matrix was performed to reveal which meiofaunal taxa or nematodegenera contributed mostly to the observed dissimilarity. The turnover diversity(i.e., ß-diversity estimated as % Bray–Curtis dissimilarity; [56]) was also estimated asthe dissimilarity of meiofaunal (separately for the entire community and rare taxa) andnematode assemblage composition between Urban and MPA meadows before and afterthe summer season (in May and September) and data were presence/absence transformedprior to the analysis.
In order to explore the multivariate structure of nematode assemblages, non-metricMulti-Dimensional Scaling (nMDS) ordination was performed based on the Bray–Curtissimilarity matrix.
To ascertain if the grain size, quantity and quality of sedimentary organic matterexplain changes in meiofaunal and nematode assemblages, non-parametric multivariatemultiple regression analyses, based on Euclidean distances, were carried out using theroutine DISTLM forward [57]. The forward selection of the predictor variables wascarried out with tests by permutation. P values were obtained using 4999 permutationsof raw data for the marginal tests (tests of individual variables), while for all of theconditional tests, the routine uses 4999 permutations of residuals under a reduced model.
All statistical tests were performed using PRIMER 6 with PERMANOVAþ add-onsoftware package [58, 59] and GMAV 5.0 software (University of Sydney, Australia).
Results
Posidonia oceanica conservation status
Results on the P. oceanica conservation status are reported in Giovannetti et al. [14]. Theleaf biometry, the shoot density, the Conservation Index and the epiphyte coverage did notdetect any differences between the MPA and the Urban meadows in the two distinctperiods under investigation. In contrast, the Urban meadows showed a considerabledevelopment of the brown algae, such as Myrionema orbiculare and Giraudiasphacelarioides.
Grain size
Sediments varied from medium to very coarse sand, with mean grain size values rangingbetween 0.36� 0.03 and 1.89� 0.18mm, found respectively at Arma di Taggia (AT) andCamogli (CA) in September (Table 1). The grain size showed significant differences amongmeadows (ANOVA, F4,95¼ 6.20, p5 0.01). No significant differences were found betweenUrban and MPA meadows in any period.
Sedimentary organic matter concentration and biochemical composition
Proteins (Prt) showed concentrations ranging between 0.10� 0.01mg g�1 of sediment DWat Pieve Ligure (PL) in May and 0.34� 0.04mg g�1 of sediment DW at Camogli (CA) inMay. Carbohydrate (Cho) concentrations ranged between 0.17� 0.04mg g�1 of sedimentDW at Monterosso al Mare (MM) in May and 0.74� 0.03mg g�1 of sediment DW at
74 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2
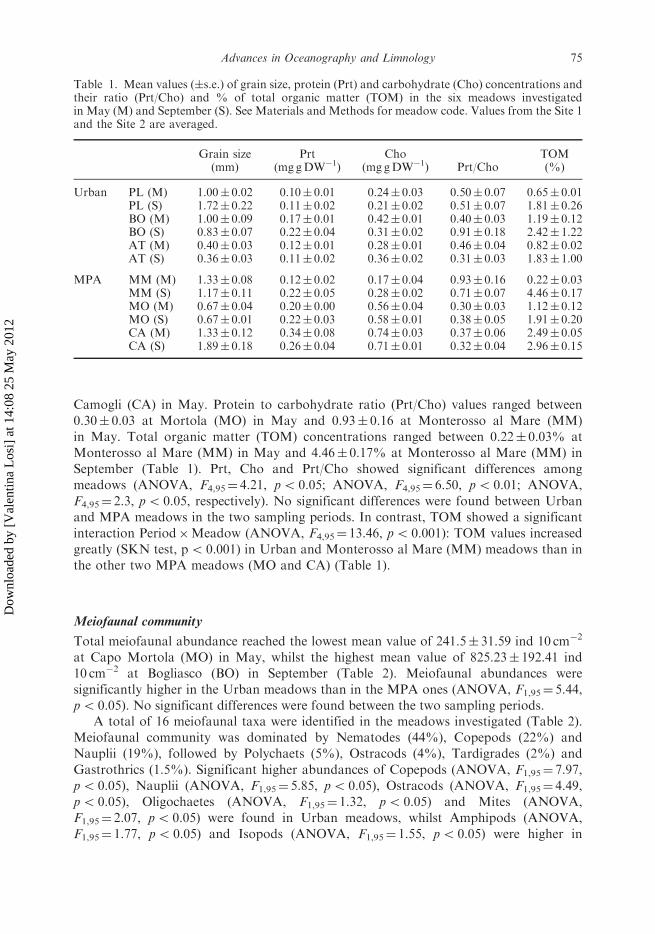
Camogli (CA) in May. Protein to carbohydrate ratio (Prt/Cho) values ranged between0.30� 0.03 at Mortola (MO) in May and 0.93� 0.16 at Monterosso al Mare (MM)in May. Total organic matter (TOM) concentrations ranged between 0.22� 0.03% atMonterosso al Mare (MM) in May and 4.46� 0.17% at Monterosso al Mare (MM) inSeptember (Table 1). Prt, Cho and Prt/Cho showed significant differences amongmeadows (ANOVA, F4,95¼ 4.21, p5 0.05; ANOVA, F4,95¼ 6.50, p5 0.01; ANOVA,F4,95¼ 2.3, p5 0.05, respectively). No significant differences were found between Urbanand MPA meadows in the two sampling periods. In contrast, TOM showed a significantinteraction Period�Meadow (ANOVA, F4,95¼ 13.46, p5 0.001): TOM values increasedgreatly (SKN test, p5 0.001) in Urban and Monterosso al Mare (MM) meadows than inthe other two MPA meadows (MO and CA) (Table 1).
Meiofaunal community
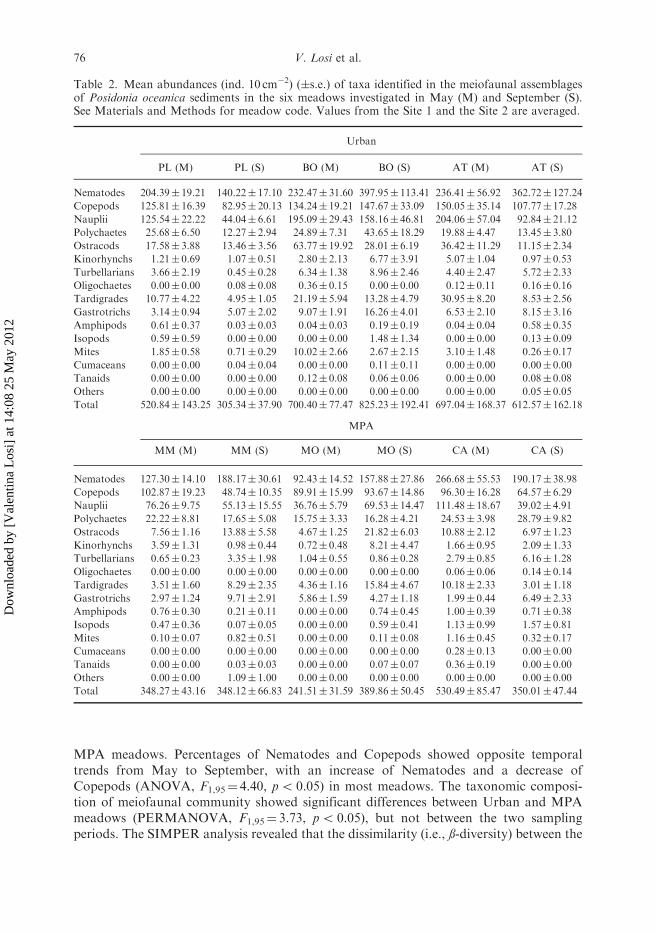
Total meiofaunal abundance reached the lowest mean value of 241.5� 31.59 ind 10 cm�2
at Capo Mortola (MO) in May, whilst the highest mean value of 825.23� 192.41 ind10 cm�2 at Bogliasco (BO) in September (Table 2). Meiofaunal abundances weresignificantly higher in the Urban meadows than in the MPA ones (ANOVA, F1,95¼ 5.44,p5 0.05). No significant differences were found between the two sampling periods.
A total of 16 meiofaunal taxa were identified in the meadows investigated (Table 2).Meiofaunal community was dominated by Nematodes (44%), Copepods (22%) andNauplii (19%), followed by Polychaets (5%), Ostracods (4%), Tardigrades (2%) andGastrothrics (1.5%). Significant higher abundances of Copepods (ANOVA, F1,95¼ 7.97,p5 0.05), Nauplii (ANOVA, F1,95¼ 5.85, p5 0.05), Ostracods (ANOVA, F1,95¼ 4.49,p5 0.05), Oligochaetes (ANOVA, F1,95¼ 1.32, p5 0.05) and Mites (ANOVA,F1,95¼ 2.07, p5 0.05) were found in Urban meadows, whilst Amphipods (ANOVA,F1,95¼ 1.77, p5 0.05) and Isopods (ANOVA, F1,95¼ 1.55, p5 0.05) were higher in
Table 1. Mean values (�s.e.) of grain size, protein (Prt) and carbohydrate (Cho) concentrations andtheir ratio (Prt/Cho) and % of total organic matter (TOM) in the six meadows investigatedin May (M) and September (S). See Materials and Methods for meadow code. Values from the Site 1and the Site 2 are averaged.
Grain size(mm)
Prt(mg gDW�1)
Cho(mg gDW�1) Prt/Cho
TOM(%)
Urban PL (M) 1.00� 0.02 0.10� 0.01 0.24� 0.03 0.50� 0.07 0.65� 0.01PL (S) 1.72� 0.22 0.11� 0.02 0.21� 0.02 0.51� 0.07 1.81� 0.26BO (M) 1.00� 0.09 0.17� 0.01 0.42� 0.01 0.40� 0.03 1.19� 0.12BO (S) 0.83� 0.07 0.22� 0.04 0.31� 0.02 0.91� 0.18 2.42� 1.22AT (M) 0.40� 0.03 0.12� 0.01 0.28� 0.01 0.46� 0.04 0.82� 0.02AT (S) 0.36� 0.03 0.11� 0.02 0.36� 0.02 0.31� 0.03 1.83� 1.00
MPA MM (M) 1.33� 0.08 0.12� 0.02 0.17� 0.04 0.93� 0.16 0.22� 0.03MM (S) 1.17� 0.11 0.22� 0.05 0.28� 0.02 0.71� 0.07 4.46� 0.17MO (M) 0.67� 0.04 0.20� 0.00 0.56� 0.04 0.30� 0.03 1.12� 0.12MO (S) 0.67� 0.01 0.22� 0.03 0.58� 0.01 0.38� 0.05 1.91� 0.20CA (M) 1.33� 0.12 0.34� 0.08 0.74� 0.03 0.37� 0.06 2.49� 0.05CA (S) 1.89� 0.18 0.26� 0.04 0.71� 0.01 0.32� 0.04 2.96� 0.15
Advances in Oceanography and Limnology 75
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2
MPA meadows. Percentages of Nematodes and Copepods showed opposite temporaltrends from May to September, with an increase of Nematodes and a decrease ofCopepods (ANOVA, F1,95¼ 4.40, p5 0.05) in most meadows. The taxonomic composi-tion of meiofaunal community showed significant differences between Urban and MPAmeadows (PERMANOVA, F1,95¼ 3.73, p5 0.05), but not between the two samplingperiods. The SIMPER analysis revealed that the dissimilarity (i.e., ß-diversity) between the
Table 2. Mean abundances (ind. 10 cm�2) (�s.e.) of taxa identified in the meiofaunal assemblagesof Posidonia oceanica sediments in the six meadows investigated in May (M) and September (S).See Materials and Methods for meadow code. Values from the Site 1 and the Site 2 are averaged.
Urban
PL (M) PL (S) BO (M) BO (S) AT (M) AT (S)
Nematodes 204.39� 19.21 140.22� 17.10 232.47� 31.60 397.95� 113.41 236.41� 56.92 362.72� 127.24
Copepods 125.81� 16.39 82.95� 20.13 134.24� 19.21 147.67� 33.09 150.05� 35.14 107.77� 17.28
Nauplii 125.54� 22.22 44.04� 6.61 195.09� 29.43 158.16� 46.81 204.06� 57.04 92.84� 21.12
Polychaetes 25.68� 6.50 12.27� 2.94 24.89� 7.31 43.65� 18.29 19.88� 4.47 13.45� 3.80
Ostracods 17.58� 3.88 13.46� 3.56 63.77� 19.92 28.01� 6.19 36.42� 11.29 11.15� 2.34
Kinorhynchs 1.21� 0.69 1.07� 0.51 2.80� 2.13 6.77� 3.91 5.07� 1.04 0.97� 0.53
Turbellarians 3.66� 2.19 0.45� 0.28 6.34� 1.38 8.96� 2.46 4.40� 2.47 5.72� 2.33
Oligochaetes 0.00� 0.00 0.08� 0.08 0.36� 0.15 0.00� 0.00 0.12� 0.11 0.16� 0.16
Tardigrades 10.77� 4.22 4.95� 1.05 21.19� 5.94 13.28� 4.79 30.95� 8.20 8.53� 2.56
Gastrotrichs 3.14� 0.94 5.07� 2.02 9.07� 1.91 16.26� 4.01 6.53� 2.10 8.15� 3.16
Amphipods 0.61� 0.37 0.03� 0.03 0.04� 0.03 0.19� 0.19 0.04� 0.04 0.58� 0.35
Isopods 0.59� 0.59 0.00� 0.00 0.00� 0.00 1.48� 1.34 0.00� 0.00 0.13� 0.09
Mites 1.85� 0.58 0.71� 0.29 10.02� 2.66 2.67� 2.15 3.10� 1.48 0.26� 0.17
Cumaceans 0.00� 0.00 0.04� 0.04 0.00� 0.00 0.11� 0.11 0.00� 0.00 0.00� 0.00
Tanaids 0.00� 0.00 0.00� 0.00 0.12� 0.08 0.06� 0.06 0.00� 0.00 0.08� 0.08
Others 0.00� 0.00 0.00� 0.00 0.00� 0.00 0.00� 0.00 0.00� 0.00 0.05� 0.05
Total 520.84� 143.25 305.34� 37.90 700.40� 77.47 825.23� 192.41 697.04� 168.37 612.57� 162.18
MPA
MM (M) MM (S) MO (M) MO (S) CA (M) CA (S)
Nematodes 127.30� 14.10 188.17� 30.61 92.43� 14.52 157.88� 27.86 266.68� 55.53 190.17� 38.98
Copepods 102.87� 19.23 48.74� 10.35 89.91� 15.99 93.67� 14.86 96.30� 16.28 64.57� 6.29
Nauplii 76.26� 9.75 55.13� 15.55 36.76� 5.79 69.53� 14.47 111.48� 18.67 39.02� 4.91
Polychaetes 22.22� 8.81 17.65� 5.08 15.75� 3.33 16.28� 4.21 24.53� 3.98 28.79� 9.82
Ostracods 7.56� 1.16 13.88� 5.58 4.67� 1.25 21.82� 6.03 10.88� 2.12 6.97� 1.23
Kinorhynchs 3.59� 1.31 0.98� 0.44 0.72� 0.48 8.21� 4.47 1.66� 0.95 2.09� 1.33
Turbellarians 0.65� 0.23 3.35� 1.98 1.04� 0.55 0.86� 0.28 2.79� 0.85 6.16� 1.28
Oligochaetes 0.00� 0.00 0.00� 0.00 0.00� 0.00 0.00� 0.00 0.06� 0.06 0.14� 0.14
Tardigrades 3.51� 1.60 8.29� 2.35 4.36� 1.16 15.84� 4.67 10.18� 2.33 3.01� 1.18
Gastrotrichs 2.97� 1.24 9.71� 2.91 5.86� 1.59 4.27� 1.18 1.99� 0.44 6.49� 2.33
Amphipods 0.76� 0.30 0.21� 0.11 0.00� 0.00 0.74� 0.45 1.00� 0.39 0.71� 0.38
Isopods 0.47� 0.36 0.07� 0.05 0.00� 0.00 0.59� 0.41 1.13� 0.99 1.57� 0.81
Mites 0.10� 0.07 0.82� 0.51 0.00� 0.00 0.11� 0.08 1.16� 0.45 0.32� 0.17
Cumaceans 0.00� 0.00 0.00� 0.00 0.00� 0.00 0.00� 0.00 0.28� 0.13 0.00� 0.00
Tanaids 0.00� 0.00 0.03� 0.03 0.00� 0.00 0.07� 0.07 0.36� 0.19 0.00� 0.00
Others 0.00� 0.00 1.09� 1.00 0.00� 0.00 0.00� 0.00 0.00� 0.00 0.00� 0.00
Total 348.27� 43.16 348.12� 66.83 241.51� 31.59 389.86� 50.45 530.49� 85.47 350.01� 47.44
76 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2
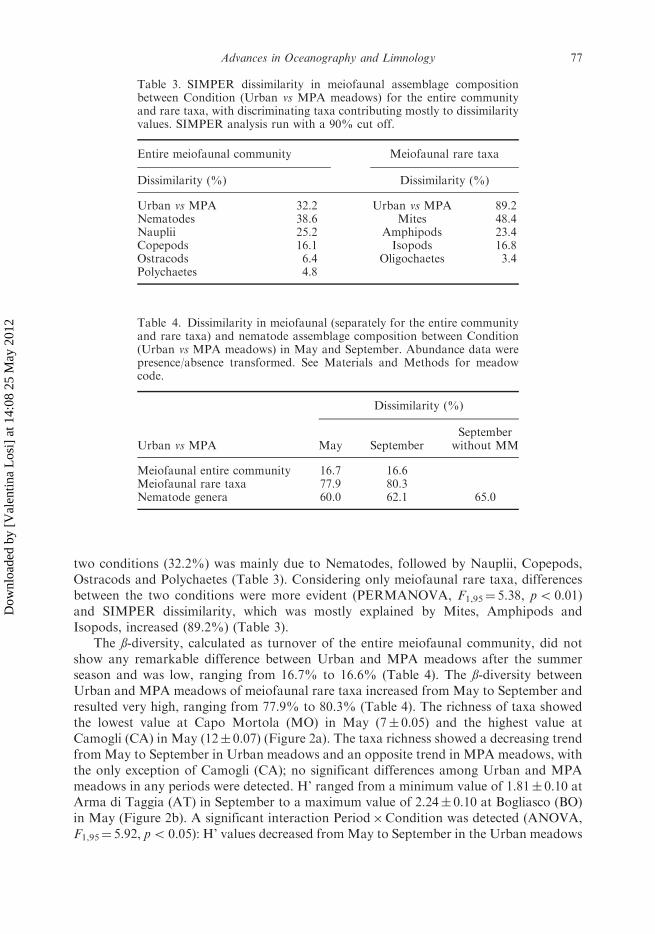
two conditions (32.2%) was mainly due to Nematodes, followed by Nauplii, Copepods,Ostracods and Polychaetes (Table 3). Considering only meiofaunal rare taxa, differencesbetween the two conditions were more evident (PERMANOVA, F1,95¼ 5.38, p5 0.01)and SIMPER dissimilarity, which was mostly explained by Mites, Amphipods andIsopods, increased (89.2%) (Table 3).
The ß-diversity, calculated as turnover of the entire meiofaunal community, did notshow any remarkable difference between Urban and MPA meadows after the summerseason and was low, ranging from 16.7% to 16.6% (Table 4). The ß-diversity betweenUrban and MPA meadows of meiofaunal rare taxa increased from May to September andresulted very high, ranging from 77.9% to 80.3% (Table 4). The richness of taxa showedthe lowest value at Capo Mortola (MO) in May (7� 0.05) and the highest value atCamogli (CA) in May (12� 0.07) (Figure 2a). The taxa richness showed a decreasing trendfrom May to September in Urban meadows and an opposite trend in MPA meadows, withthe only exception of Camogli (CA); no significant differences among Urban and MPAmeadows in any periods were detected. H’ ranged from a minimum value of 1.81� 0.10 atArma di Taggia (AT) in September to a maximum value of 2.24� 0.10 at Bogliasco (BO)in May (Figure 2b). A significant interaction Period�Condition was detected (ANOVA,F1,95¼ 5.92, p5 0.05): H’ values decreased fromMay to September in the Urban meadows
Table 3. SIMPER dissimilarity in meiofaunal assemblage compositionbetween Condition (Urban vs MPA meadows) for the entire communityand rare taxa, with discriminating taxa contributing mostly to dissimilarityvalues. SIMPER analysis run with a 90% cut off.
Entire meiofaunal community Meiofaunal rare taxa
Dissimilarity (%) Dissimilarity (%)
Urban vs MPA 32.2 Urban vs MPA 89.2Nematodes 38.6 Mites 48.4Nauplii 25.2 Amphipods 23.4Copepods 16.1 Isopods 16.8Ostracods 6.4 Oligochaetes 3.4Polychaetes 4.8
Table 4. Dissimilarity in meiofaunal (separately for the entire communityand rare taxa) and nematode assemblage composition between Condition(Urban vs MPA meadows) in May and September. Abundance data werepresence/absence transformed. See Materials and Methods for meadowcode.
Dissimilarity (%)
Urban vs MPA May SeptemberSeptember
without MM
Meiofaunal entire community 16.7 16.6Meiofaunal rare taxa 77.9 80.3Nematode genera 60.0 62.1 65.0
Advances in Oceanography and Limnology 77
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2
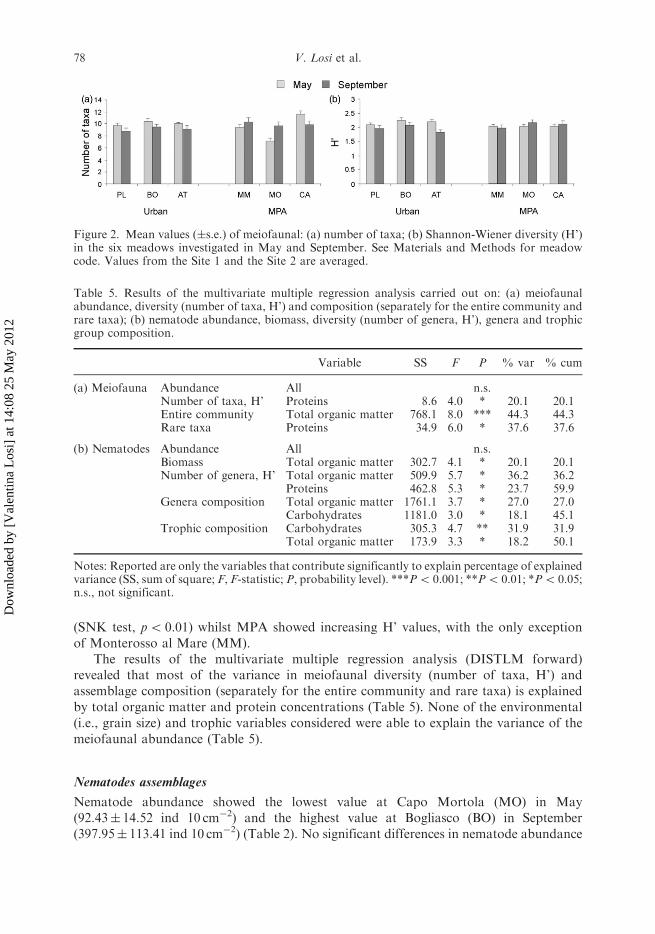
(SNK test, p5 0.01) whilst MPA showed increasing H’ values, with the only exceptionof Monterosso al Mare (MM).
The results of the multivariate multiple regression analysis (DISTLM forward)revealed that most of the variance in meiofaunal diversity (number of taxa, H’) andassemblage composition (separately for the entire community and rare taxa) is explainedby total organic matter and protein concentrations (Table 5). None of the environmental(i.e., grain size) and trophic variables considered were able to explain the variance of themeiofaunal abundance (Table 5).
Nematodes assemblages
Nematode abundance showed the lowest value at Capo Mortola (MO) in May(92.43� 14.52 ind 10 cm�2) and the highest value at Bogliasco (BO) in September(397.95� 113.41 ind 10 cm�2) (Table 2). No significant differences in nematode abundance
Table 5. Results of the multivariate multiple regression analysis carried out on: (a) meiofaunalabundance, diversity (number of taxa, H’) and composition (separately for the entire community andrare taxa); (b) nematode abundance, biomass, diversity (number of genera, H’), genera and trophicgroup composition.
Variable SS F P % var % cum
(a) Meiofauna Abundance All n.s.Number of taxa, H’ Proteins 8.6 4.0 * 20.1 20.1Entire community Total organic matter 768.1 8.0 *** 44.3 44.3Rare taxa Proteins 34.9 6.0 * 37.6 37.6
(b) Nematodes Abundance All n.s.Biomass Total organic matter 302.7 4.1 * 20.1 20.1Number of genera, H’ Total organic matter 509.9 5.7 * 36.2 36.2
Proteins 462.8 5.3 * 23.7 59.9Genera composition Total organic matter 1761.1 3.7 * 27.0 27.0
Carbohydrates 1181.0 3.0 * 18.1 45.1Trophic composition Carbohydrates 305.3 4.7 ** 31.9 31.9
Total organic matter 173.9 3.3 * 18.2 50.1
Notes: Reported are only the variables that contribute significantly to explain percentage of explainedvariance (SS, sum of square; F, F-statistic; P, probability level). ***P5 0.001; **P5 0.01; *P5 0.05;n.s., not significant.
Figure 2. Mean values (�s.e.) of meiofaunal: (a) number of taxa; (b) Shannon-Wiener diversity (H’)in the six meadows investigated in May and September. See Materials and Methods for meadowcode. Values from the Site 1 and the Site 2 are averaged.
78 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2
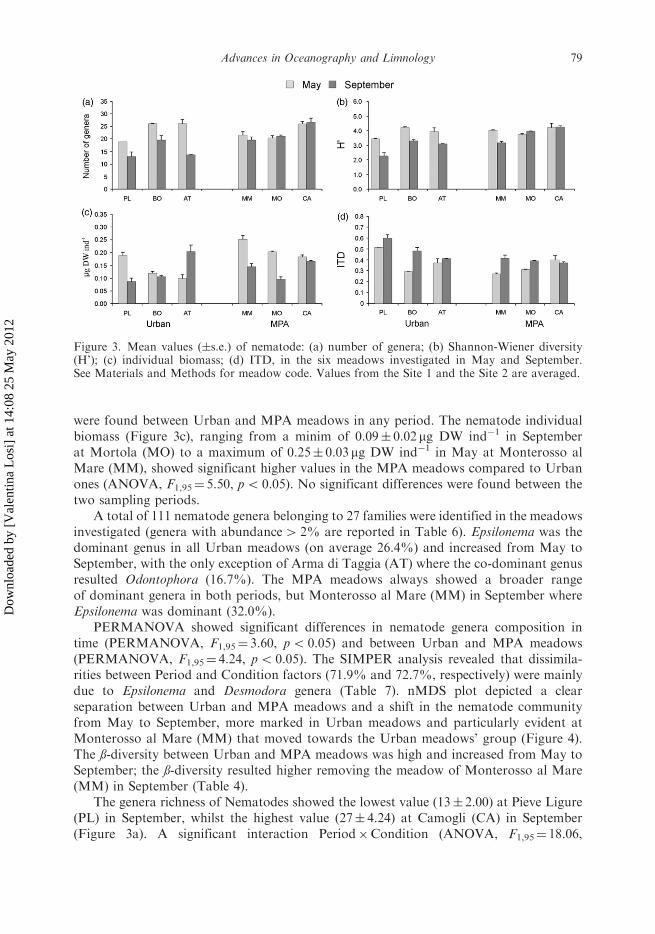
were found between Urban and MPA meadows in any period. The nematode individualbiomass (Figure 3c), ranging from a minim of 0.09� 0.02mg DW ind�1 in Septemberat Mortola (MO) to a maximum of 0.25� 0.03mg DW ind�1 in May at Monterosso alMare (MM), showed significant higher values in the MPA meadows compared to Urbanones (ANOVA, F1,95¼ 5.50, p5 0.05). No significant differences were found between thetwo sampling periods.
A total of 111 nematode genera belonging to 27 families were identified in the meadowsinvestigated (genera with abundance4 2% are reported in Table 6). Epsilonema was thedominant genus in all Urban meadows (on average 26.4%) and increased from May toSeptember, with the only exception of Arma di Taggia (AT) where the co-dominant genusresulted Odontophora (16.7%). The MPA meadows always showed a broader rangeof dominant genera in both periods, but Monterosso al Mare (MM) in September whereEpsilonema was dominant (32.0%).
PERMANOVA showed significant differences in nematode genera composition intime (PERMANOVA, F1,95¼ 3.60, p5 0.05) and between Urban and MPA meadows(PERMANOVA, F1,95¼ 4.24, p5 0.05). The SIMPER analysis revealed that dissimila-rities between Period and Condition factors (71.9% and 72.7%, respectively) were mainlydue to Epsilonema and Desmodora genera (Table 7). nMDS plot depicted a clearseparation between Urban and MPA meadows and a shift in the nematode communityfrom May to September, more marked in Urban meadows and particularly evident atMonterosso al Mare (MM) that moved towards the Urban meadows’ group (Figure 4).The ß-diversity between Urban and MPA meadows was high and increased from May toSeptember; the ß-diversity resulted higher removing the meadow of Monterosso al Mare(MM) in September (Table 4).
The genera richness of Nematodes showed the lowest value (13� 2.00) at Pieve Ligure(PL) in September, whilst the highest value (27� 4.24) at Camogli (CA) in September(Figure 3a). A significant interaction Period�Condition (ANOVA, F1,95¼ 18.06,
Figure 3. Mean values (�s.e.) of nematode: (a) number of genera; (b) Shannon-Wiener diversity(H’); (c) individual biomass; (d) ITD, in the six meadows investigated in May and September.See Materials and Methods for meadow code. Values from the Site 1 and the Site 2 are averaged.
Advances in Oceanography and Limnology 79
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2
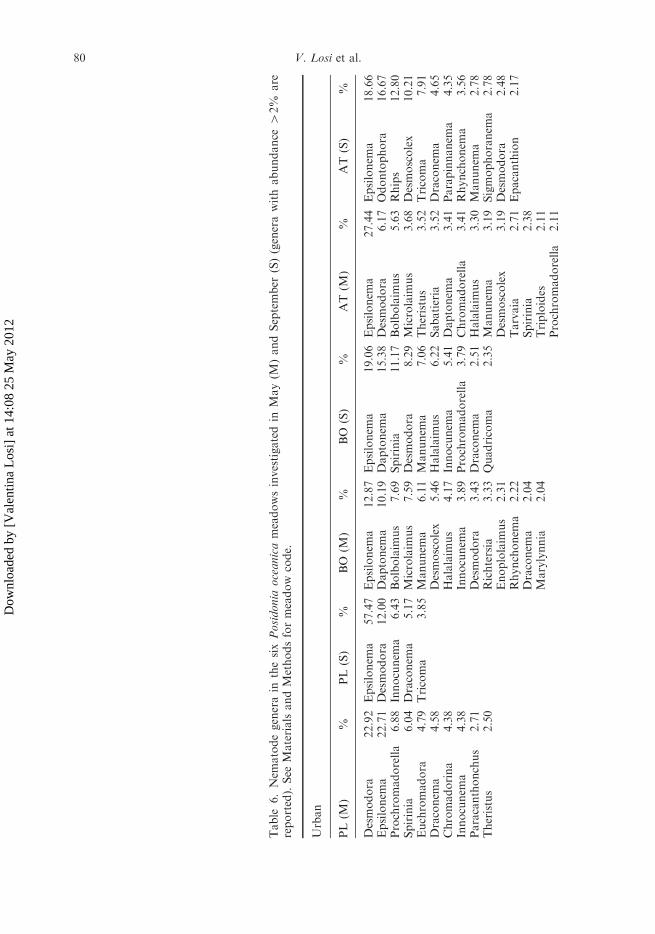
Table
6.Nem
atodegenerain
thesixPosidonia
oceanicameadowsinvestigatedin
May(M
)andSeptember
(S)(generawithabundance
42%
are
reported).See
MaterialsandMethodsformeadow
code.
Urban
PL(M
)%
PL
(S)
%BO
(M)
%BO
(S)
%AT
(M)
%AT
(S)
%
Desmodora
22.92
Epsilonem
a57.47
Epsilonem
a12.87
Epsilonem
a19.06
Epsilonem
a27.44
Epsilonem
a18.66
Epsilonem
a22.71
Desmodora
12.00
Daptonem
a10.19
Daptonem
a15.38
Desmodora
6.17
Odontophora
16.67
Prochromadorella
6.88
Innocunem
a6.43
Bolbolaim
us
7.69
Spirinia
11.17
Bolbolaim
us
5.63
Rhips
12.80
Spirinia
6.04
Draconem
a5.17
Microlaim
us
7.59
Desmodora
8.29
Microlaim
us
3.68
Desmoscolex
10.21
Euchromadora
4.79
Tricoma
3.85
Manunem
a6.11
Manunem
a7.06
Theristus
3.52
Tricoma
7.91
Draconem
a4.58
Desmoscolex
5.46
Halalaim
us
6.22
Sabatieria
3.52
Draconem
a4.65
Chromadorina
4.38
Halalaim
us
4.17
Innocunem
a5.41
Daptonem
a3.41
Parapinnanem
a4.35
Innocunem
a4.38
Innocunem
a3.89
Prochromadorella
3.79
Chromadorella
3.41
Rhynchonem
a3.56
Paracanthonchus
2.71
Desmodora
3.43
Draconem
a2.51
Halalaim
us
3.30
Manunem
a2.78
Theristus
2.50
Richtersia
3.33
Quadricoma
2.35
Manunem
a3.19
Sigmophoranem
a2.78
Enoplolaim
us
2.31
Desmoscolex
3.19
Desmodora
2.48
Rhynchonem
a2.22
Tarvaia
2.71
Epacanthion
2.17
Draconem
a2.04
Spirinia
2.38
Marylynnia
2.04
Triploides
2.11
Prochromadorella
2.11
80 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2
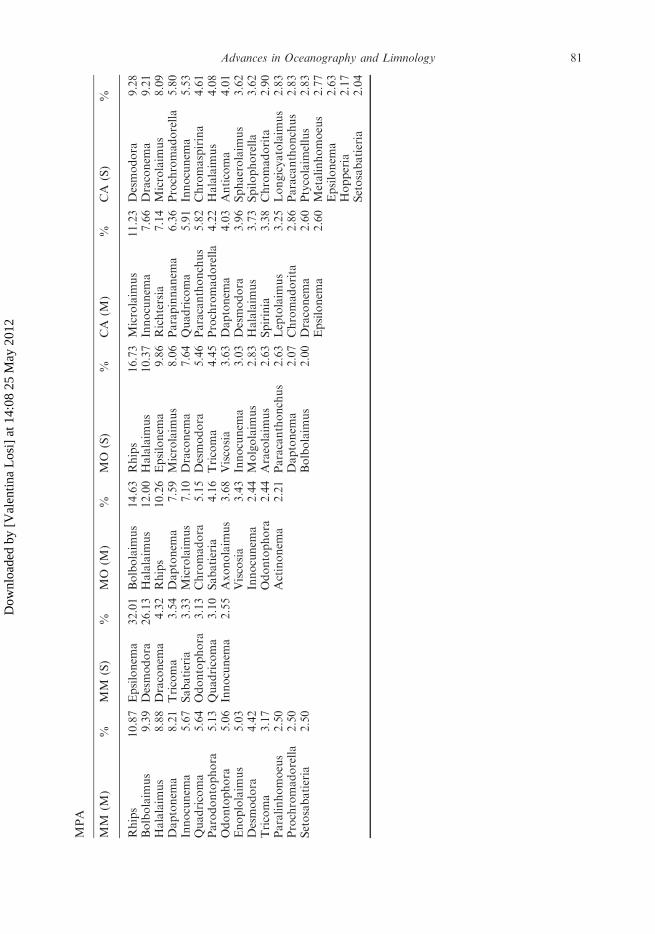
MPA
MM
(M)
%MM
(S)
%MO
(M)
%MO
(S)
%CA
(M)
%CA
(S)
%
Rhips
10.87
Epsilonem
a32.01
Bolbolaim
us
14.63
Rhips
16.73
Microlaim
us
11.23
Desmodora
9.28
Bolbolaim
us
9.39
Desmodora
26.13
Halalaim
us
12.00
Halalaim
us
10.37
Innocunem
a7.66
Draconem
a9.21
Halalaim
us
8.88
Draconem
a4.32
Rhips
10.26
Epsilonem
a9.86
Richtersia
7.14
Microlaim
us
8.09
Daptonem
a8.21
Tricoma
3.54
Daptonem
a7.59
Microlaim
us
8.06
Parapinnanem
a6.36
Prochromadorella
5.80
Innocunem
a5.67
Sabatieria
3.33
Microlaim
us
7.10
Draconem
a7.64
Quadricoma
5.91
Innocunem
a5.53
Quadricoma
5.64
Odontophora
3.13
Chromadora
5.15
Desmodora
5.46
Paracanthonchus
5.82
Chromaspirina
4.61
Parodontophora
5.13
Quadricoma
3.10
Sabatieria
4.16
Tricoma
4.45
Prochromadorella
4.22
Halalaim
us
4.08
Odontophora
5.06
Innocunem
a2.55
Axonolaim
us
3.68
Viscosia
3.63
Daptonem
a4.03
Anticoma
4.01
Enoplolaim
us
5.03
Viscosia
3.43
Innocunem
a3.03
Desmodora
3.96
Sphaerolaim
us
3.62
Desmodora
4.42
Innocunem
a2.44
Molgolaim
us
2.83
Halalaim
us
3.73
Spilophorella
3.62
Tricoma
3.17
Odontophora
2.44
Araeolaim
us
2.63
Spirinia
3.38
Chromadorita
2.90
Paralinhomoeus
2.50
Actinonem
a2.21
Paracanthonchus
2.63
Leptolaim
us
3.25
Longicyatolaim
us
2.83
Prochromadorella
2.50
Daptonem
a2.07
Chromadorita
2.86
Paracanthonchus
2.83
Setosabatieria
2.50
Bolbolaim
us
2.00
Draconem
a2.60
Ptycolaim
ellus
2.83
Epsilonem
a2.60
Metalinhomoeus
2.77
Epsilonem
a2.63
Hopperia
2.17
Setosabatieria
2.04
Advances in Oceanography and Limnology 81
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

p5 0.01) was detected, with the Urban and MPA meadows showing opposite trendin time: the number of genera decreased in Urban meadows (SNK test, p5 0.001) andincreased in MPA ones, with the exception of Monterosso al Mare (MM). SNK testshowed significant higher values of genera richness (p5 0.05) in MPA meadows thanin Urban meadows in September.
H’ ranged from a minimum value of 2.26� 0.50 at Pieve Ligure (PL) in September toa maximum value of 4.22� 0.16 at Camogli (CA) in September (Figure 3b). A significant
Table 7. SIMPER dissimilarity in nematode assemblages composition, withdissimilarity between Period (May vs September) and Condition (Urban vs MPAmeadows), and discriminating genera contributing mostly to dissimilarity values(only contribution 42% are reported). SIMPER analysis run with a 90% cut off.
Dissimilarity (%) Dissimilarity (%)
May vs September 71.9 Urban vs MPA 72.7
Epsilonema 14.3 Epsilonema 15.1Desmodora 6.7 Desmodora 6.4Rhips 5.5 Rhips 5.8Daptonema 4.6 Daptonema 4.0Bolbolaimus 4.3 Microlaimus 3.9Microlaimus 3.8 Bolbolaimus 3.7Draconema 3.4 Halalaimus 3.6Halalaimus 3.4 Odontophora 3.1Odontophora 3.1 Draconema 3.1Innocuonema 2.5 Innocuonema 2.5Tricoma 2.4 Spirinia 2.5Prochromadorella 2.4 Desmoscolex 2.4Spirinia 2.3 Manunema 2.3Manunema 2.1 Prochromadorella 2.3Desmoscolex 2.1 Tricoma 2.2Quadricoma 2.0 Quadricoma 2.0
Figure 4. nMDS of nematode assemblages of Posidonia oceanica sediments in the six meadowsinvestigated in May (M) and September (S). See Materials and Methods for meadow code.
82 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

interaction Period�Meadow (ANOVA, F4,95¼ 14.15, p5 0.001) was recorded: H’ values
decreased (SKN test, p5 0.01) in Urban meadows and Monterosso al Mare (MM)
meadow, whilst increased in the other two MPA meadows (MO and CA) (Figure 3c).
The highest trophic diversity (ITD¼ 0.27� 0.02) was recorded at Monterosso al Mare
(MM) in May, whilst the lowest (ITD¼ 0.60� 0.07) at Pieve Ligure (PL) in September
(Figure 3d). The dominant trophic group was represented by epistrate feeders (2A) (39.0%
of whole community and 38.7% of nematode genera), followed by selective deposit
feeders (1A) (37.5% of whole community and 22.5% of nematode genera) (Figure 5). The
trophic groups showed significant differences between Urban and MPA meadows
(PERMANOVA, F1,95¼ 4.72, p5 0.05). Selective deposit feeders (1A) resulted higher
(ANOVA, F1,23¼ 5.95, p5 0.05) in Urban meadows than in MPA ones (45.7% and
29.6%, respectively) and increased from May to September (ANOVA, F1,23¼ 3.89,
p5 0.05). Non-selective deposit feeders (1B) decreased at most meadows in September
(ANOVA, F1,23¼ 5.21, p5 0.05). Predators (2B) were higher (ANOVA, F1,23¼ 3.56,
p5 0.05) in the MPA meadows than in the Urban meadows (12.5% and 5.7%,
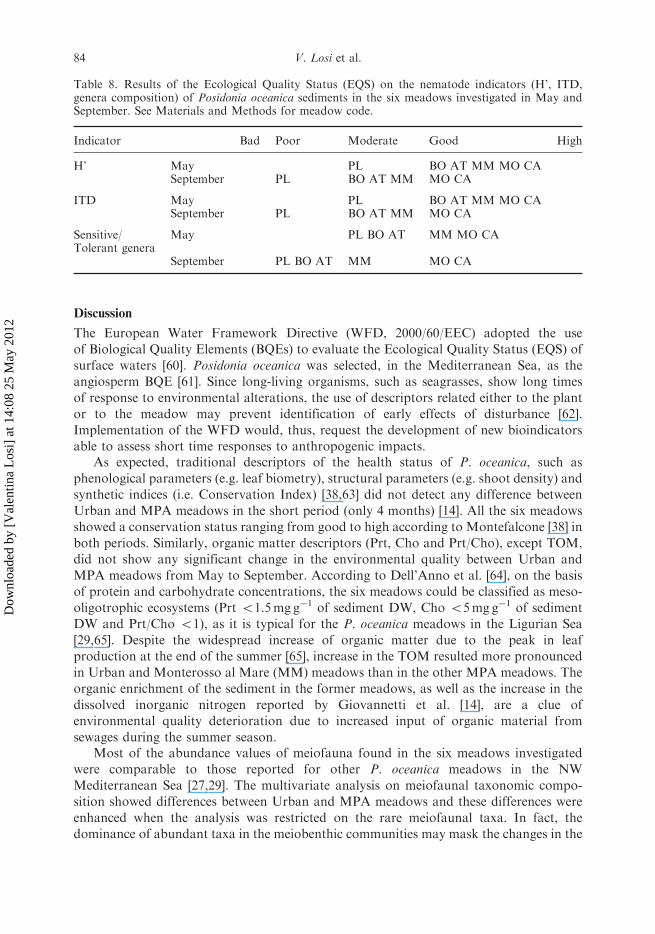
respectively).Using H’ and ITD as indicators, the Ecological Quality Status (EQS) of all the
meadows, but Pieve Ligure (PL), was Good in May; in September, only Urban and
Monterosso al Mare (MM) meadows decreased of one quality class. The genera
composition indicator classified the MPA meadows in Good status and the Urban
meadows in Moderate status in May; in September, only Urban and Monterosso al Mare
(MM) meadows decreased of one quality class (Table 8).The multivariate multiple regression analysis (DISTLM forward) revealed that most of
the variance in nematode biomass, diversity (richness of genera, H’), genera and trophic
group composition is explained by total organic matter, carbohydrate and protein
concentrations (Table 5). None of the environmental and trophic variables considered was
able to explain the variance of the nematode abundance (Table 5).
Figure 5. Trophic structure of nematode assemblages in the six meadows investigated in May (M)and September (S). 1A (selective deposit feeders); 1B (non selective deposit feeders); 2A (epistratefeeders); 2B (predators/omnivores). See Materials and Methods for meadow code.
Advances in Oceanography and Limnology 83
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2
Discussion
The European Water Framework Directive (WFD, 2000/60/EEC) adopted the useof Biological Quality Elements (BQEs) to evaluate the Ecological Quality Status (EQS) of
surface waters [60]. Posidonia oceanica was selected, in the Mediterranean Sea, as theangiosperm BQE [61]. Since long-living organisms, such as seagrasses, show long times
of response to environmental alterations, the use of descriptors related either to the plantor to the meadow may prevent identification of early effects of disturbance [62].
Implementation of the WFD would, thus, request the development of new bioindicatorsable to assess short time responses to anthropogenic impacts.
As expected, traditional descriptors of the health status of P. oceanica, such as
phenological parameters (e.g. leaf biometry), structural parameters (e.g. shoot density) andsynthetic indices (i.e. Conservation Index) [38,63] did not detect any difference between
Urban and MPA meadows in the short period (only 4 months) [14]. All the six meadowsshowed a conservation status ranging from good to high according to Montefalcone [38] in
both periods. Similarly, organic matter descriptors (Prt, Cho and Prt/Cho), except TOM,did not show any significant change in the environmental quality between Urban and
MPA meadows from May to September. According to Dell’Anno et al. [64], on the basisof protein and carbohydrate concentrations, the six meadows could be classified as meso-
oligotrophic ecosystems (Prt 51.5mg g�1 of sediment DW, Cho 55mg g�1 of sedimentDW and Prt/Cho 51), as it is typical for the P. oceanica meadows in the Ligurian Sea
[29,65]. Despite the widespread increase of organic matter due to the peak in leafproduction at the end of the summer [65], increase in the TOM resulted more pronounced
in Urban and Monterosso al Mare (MM) meadows than in the other MPA meadows. Theorganic enrichment of the sediment in the former meadows, as well as the increase in the
dissolved inorganic nitrogen reported by Giovannetti et al. [14], are a clue ofenvironmental quality deterioration due to increased input of organic material from
sewages during the summer season.Most of the abundance values of meiofauna found in the six meadows investigated
were comparable to those reported for other P. oceanica meadows in the NW
Mediterranean Sea [27,29]. The multivariate analysis on meiofaunal taxonomic compo-sition showed differences between Urban and MPA meadows and these differences were
enhanced when the analysis was restricted on the rare meiofaunal taxa. In fact, thedominance of abundant taxa in the meiobenthic communities may mask the changes in the
Table 8. Results of the Ecological Quality Status (EQS) on the nematode indicators (H’, ITD,genera composition) of Posidonia oceanica sediments in the six meadows investigated in May andSeptember. See Materials and Methods for meadow code.
Indicator Bad Poor Moderate Good High
H’ May PL BO AT MM MO CASeptember PL BO AT MM MO CA
ITD May PL BO AT MM MO CASeptember PL BO AT MM MO CA
Sensitive/Tolerant genera
May PL BO AT MM MO CA
September PL BO AT MM MO CA
84 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

relative importance of the other taxa [66]. Dominant meiofaunal taxa (Nematodes,Copepods, Nauplii, Polychaetes, Ostracods) were the same for Urban and MPA meadowsbut, in Urban meadows, they were much more abundant in both periods. Differences weremore manifest considering rare taxa: Mites and Oligochaetes occurred mostly in Urbanmeadows whilst Amphipods and Isopods in MPA ones. Those differences were consistentwith time, being found in both May and September. The higher presence of Isopodsand Amphipods, known as sensitive taxa [67 and refs. therein], could indicate a betterenvironmental quality of MPA meadows compared with Urban meadows. Increase in theturnover diversity on rare taxa composition between Urban and MPA meadows withtime stressed the importance of rare meiofaunal taxa in detecting differences in theenvironmental quality under different trophic state conditions [68].
H’ values showed an opposite temporal trend, decreasing in Urban meadows andincreasing in MPA meadows, with the exception of Monterosso al Mare (MM). Reductionin the meiofaunal diversity in response to human disturbance has already been reported byseveral Authors [e.g., 24, 69–73]. Meiofaunal diversity, however, resulted always very highand confirmed P. oceanica meadows as highly diverse ecosystems.
Working at a higher taxonomic resolution, nematode genera composition highlighteddifferences between Urban and MPA meadows and between the two sampling periods,differently from nematode abundance and biomass. Nematodes of Urban meadowsexperienced a decrease in the diversity, expressed as number of genera and H’. Despite thecomplex links between diversity and disturbance [74, 75], a reduction in the nematodediversity as a response to organic enrichment has already been reported by several Authors[e.g., 76–78]. Nematode community in Urban meadow was different from that in the MPAmeadows already in May, thus suggesting the existence of chronic disturbances affectingthe former meadows. Intensification of disturbance during the summer season amplifiedthose differences, resulting in a shift of nematode community structure in Urban meadows.In only 4 months the increased dominance of genus Epsilonema and the increase orappearance, among the dominant genera, of typical tolerant genera known to resist todifferent kinds of pollution, such as Daptonema [e.g., 79, 80] and Odontophora [e.g., 54, 81,82] were recorded. The increase in Epsilonema may be explained as an opportunisticresponse due to its ability to exploit the food released from sewage discharge. Epsilonemais a small and short Nematode, with a strong tendency towards clumped distributions [83]and is a good competitor adhering to sediments around food patches [84]. It constitutes anaberrant morphotype, called ‘‘stout’’, characterised by a length/width ratio 515 [85] andis heavily cuticularized, all features that make this genus more stable and less vulnerableto disturbance [86]. Several studies pointed out that small, short and stout Nematodes,which are characterized by higher growth rates and a reduced age at first breeding, act asopportunists in shallow and ocean-margin areas [85–87]. Nematodes of MPA meadowsshowed a more heterogeneous community with a comparatively broader range of the mostabundant genera and resulted characterized by the presence of sensitive genera, such asRichtersia, Halalaimus and Bolbolaimus [e.g., 32, 54, 88, 89] in both periods, indicatinggood environmental quality. The nematode assemblages were likely to be related withorganic enrichment associated to sewage discharge, which resulted as the main factorexplaining their variability.
Signals of different environmental quality between Urban and MPA meadows werehighlighted also by the trophic structure of nematode community, which is known to becoupled with the potential food sources available in the sediments [30]. The dominanceof selective deposit feeders, i.e. bacterivorous Nematodes that mainly appear in spots with
Advances in Oceanography and Limnology 85
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

high microbial activity [90], in Urban meadows both in May and September can be viewedas a consequence of the sewage input that favoured development of bacteria in sediments[91, 92]. On the other hand, the dominance of epistrate feeders (2B) in MPA in Mayand September appears as a specific feature of P. oceanica meadows, consistently withthe relevance of vegetal debris, microphytobenthos, epiphytes and phytoplankton inputsof this system [30]. The higher abundance of predators in MPA meadows in both periodsindicates a more heterogeneous and structured community characterized by the highesttrophic levels that might imply a higher habitat complexity [93].
Monterosso al Mare (MM) showed, however, a different behavior from May toSeptember with respect to the other two MPA meadows (MO and CA). In only 4 months itregistered an increase in the total organic matter, a decrease in both meiofaunal andnematode diversity, a decrease of those sensitive rare taxa typical of MPA meadows(i.e., Amphipods and Isopods) and an increase in the rare taxa typical of Urban ones(i.e., Mites). Its nematode community changed from the one characteristic of MPAmeadows, i.e. heterogeneous with sensitive genera and dominated by the epistrate feederstrophic group, to the one described for Urban meadows in September, i.e. dominated byEpsilonema and Desmodora and by selective deposit feeders. This meadow is located infront of the village of Monterosso al Mare (MM) that is one of the most famous localitiesof the ‘‘Cinque Terre’’, a coastal area of the eastern Ligurian Riviera. The legalprotection of this Marine Protected Area resulted ineffective to preserve the P. oceanicameadow of Monterosso al Mare (MM) from intensive tourist frequentation duringthe summer season [94], a pulse disturbance that affects periodically this area. As aconsequence, the choice of Monterosso al Mare (MM) as a Ligurian control area withinthe national monitoring program aimed at reaching objectives of the WFD has beenrecently criticized [95].
The three nematodes indicators used to assess the Ecological Quality Status (EQS),namely H’, ITD and genera composition, were consistent in detecting environmentalquality deterioration from May to September in all the Urban meadows and Monterossoal Mare (MM) meadow. In particular, the indicator based on genera compositionwas able to differentiate environmental quality between MPA and Urban meadowsalready in May.
In conclusion, the meiofauna community and, in particular, the rare taxa, discrim-inating Urban meadows from MPA meadows, can be viewed as an appropriate biologicalquality element (BQE) to be adopted in water quality assessments; however, a lowertaxonomic resolution was likely to hide the short time response we expected in only4 months. On the contrary, the Nematodes changed their community structure during thesummer period and, thus, they should be used as an early warning indicator of changesin environmental quality before that the effects were evident on the plant and/or on thewhole meadow.
Acknowledgments
We thank the staff of the Laboratory of Ecology (University of Genoa) for precious helpwith laboratory analyses and Carlo Bertora, Marco Donato, Roberta Lasagna, Ilaria Lavarello,Ambra Milani, Romina Rivella for their participation in field activities. We are very gratefulto Mariachiara Chiantore and Cristina Misic for valuable comments and suggestions. Weacknowledge the two anonymous referees for their constructive and critical reviews of themanuscript.
86 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

References
[1] N. Marba, C.M. Duarte, M. Holmer, R. Martınez, G. Basterretxea, A. Orfila, A. Jordi, and
J. Tintore, Effectiveness of protection of seagrass (Posidonia oceanica) populations in Cabrera
National Park (Spain), Environ. Conserv. 29 (2002), pp. 509–518.[2] C.F. Boudouresque, G. Bernard, P. Bonhomme, E. Charbonnel, G., Diviacco, A. Meinesz,
G. Pergent, C. Pergent-Martini, S. Ruitton, and L. Tunesi, Preservation et conservation des
herbiers a Posidonia oceanica, RAMOGE Pub., France, 2006.
[3] M. Montefalcone, A. Rovere, V. Parravicini, G. Albertelli, C. Morri, and C.N. Bianchi,
Evaluating change in seagrass meadows: a time-framed comparison of Side Scan Sonar maps,
Aquat. Bot. (2011), in press.[4] N. Marba, C.M. Duarte, J. Cebrian, M.E. Gallegos, B. Olesen, and K. Sand-Jensen, Growth and
population dynamics of Posidonia oceanica on the Spanish Mediterranean coast: elucidating
seagrass decline, Mar. Ecol. Prog. Ser. 137 (1996), pp. 203–213.[5] M. Montefalcone, V. Parravicini, M. Vacchi, G. Albertelli, M. Ferrari, C. Morri, and
C.N. Bianchi, Human influence on seagrass habitat fragmentation in NW Mediterranean Sea,
Estuar. Coast. Shelf S. 86 (2010), pp. 292–298.[6] A. Meinesz, J.R. Lefevre, and J.M. Astier, Impact of coastal development on the infralittoral zone
along the southeastern Mediterranean shore of continental France, Mar. Pollut. Bull. 23 (1991),
pp. 343–347.[7] C.M. Duarte, The future of seagrass meadows, Environ. Conserv. 29 (2002), pp. 192–206.[8] C.N. Bianchi and C. Morri, Marine biodiversity of the Mediterranean Sea: situation, problems
and prospects for future research, Mar. Pollut. Bull. 40 (2000), pp. 367–376.
[9] A. Peirano, V. Damasso, M. Montefalcone, C. Morri, and C.N. Bianchi, Effect of
climate, invasive species and anthropgenic impacts on the growth of the seagrass Posidonia
oceanica (L.) Delile in Liguria (NW Mediterranean Sea), Mar. Pollut. Bull. 50 (2005),
pp. 817–822.
[10] M. Montefalcone, G. Albertelli, C. Morri, and C.N. Bianchi, Patterns of wide-scale
substitution within meadows of the seagrass Posidonia oceanica in NW Mediterranean Sea:
invaders are stronger than natives, Aquat. Conserv.: Mar. Freshwat. Ecosyst. 20 (2010),
pp. 507–515.[11] C.N. Bianchi and A. Peirano, Atlante delle fanerogame marine della Liguria: Posidonia oceanica
e Cymodocea nodosa, ENEA, Centro Ricerche Ambiente Marino, La Spezia, 1995.[12] European Community, Council Directive on the conservation of natural habitats and of wild fauna
and flora, 92/43/EEC, Off. J. Eur. Comm. L 206, 22 July 1992.[13] C. F. Boudouresque, Marine biodiversity in the Mediterranean: status of species, populations and
communities, Sci. Rep. Port-Cros Natl. Park 20 (2004), pp. 97–146.
[14] E. Giovannetti, M. Montefalcone, C. Morri, C.N. Bianchi, and G. Albertelli, Early warning
response of Posidonia oceanica epiphyte community to environmental alterations (Ligurian Sea,
NW Mediterranean), Mar. Pollut. Bull. 60 (2010), pp. 1031–1039.[15] R. Danovaro, C. Gambi, and S. Mirto, Meiofaunal production and energy transfer efficiency in a
seagrass (Posidonia oceanica) bed of the Western Mediterranean, Mar. Ecol. Prog. Ser. 234
(2002), pp. 95–104.[16] M. Armenteros, J.P. Williams, B. Creagh, and N. Capetillo, Spatial and temporal variations of
meiofaunal communities from the western sector of the Gulf of Batabano, Cuba: II. Seagrass
systems, Rev. Biol. Trop. 56 (2008), pp. 55–63.[17] R.M. Warwick, M.R. Carr, K.R. Clarke, J.M. Gee, and R.H. Green, A mesocosm experiment on
the effects of hydrocarbon and copper on a sublittoral soft-bottom sediment meiobenthic
community, Mar. Ecol. Prog. Ser. 46 (1988), pp. 181–191.
[18] P.J. Somerfield, J.M. Gee, and R.M. Warwick, Soft sediment meiofaunal community structure
in relation to a long-term heavy metal gradient in the Fal estuary system, Mar. Ecol. Prog. Ser. 105
(1994), pp. 79–88.
Advances in Oceanography and Limnology 87
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

[19] M. Moreno, L. Vezzulli, V. Marin, P. Laconi, G. Albertelli, and M. Fabiano, The use of
meiofauna diversity as an indicator of pollution in harbours, ICES J. Mar. Sci. 65 (2008),
pp. 1428–1435.
[20] S. Mirto, S. Bianchelli, C. Gambi, M. Krzelj, A. Pusceddu, M. Scopa, M. Holmer, and
R. Danovaro, Fish-farm impact on metazoan meiofauna in the Mediterranean Sea: analysis of
regional vs. habitat effects, Mar. Env. Res. 69 (2010), pp. 38–47.[21] R.M. Warwick and K.R. Clarke, A comparison of some methods for analysing changes in benthic
community structure, J. Mar. Biol. Assoc. U.K. 71 (1991), pp. 225–244.
[22] V.R. Ferris and J.M. Ferris, Thread worms (Nematoda), in Pollution ecology of estuarine
environments, C.W. Hart and S.L.H. Fuller, eds., Academic Press, London, 1979, pp. 1–33.[23] B.C. Coull and M.A. Palmer, Field experimentation in meiofaunal ecology, Hydrobiologia
118 (1984), pp. 1–19.[24] B.C. Coull and G.T. Chandler, Pollution and meiofauna: field, laboratory and mesocosm studies,
Oceanogr. Mar. Biol. Annu. Rev. 30 (1992), pp. 191–271.[25] T. Bongers and H. Ferris, Nematode community structure as a bioindicator in environmental
monitoring, Trends Ecol. Evol. 14 (1999), pp. 224–228.
[26] S. Hoss, E. Claus, P.C. Von der Ohe, M. Brinke, H. Gude, P. Heininger, and W. Traunspurger,
Nematode species at risk - A metric to assess pollution in soft sediments of freshwaters, Environ.
Int. 37 (2011), pp. 940–949.
[27] R. Novak, Spatial and seasonal distribution of the meiofauna in the seagrass Posidonia oceanica,
Neth. J. Sea Res. 16 (1982), pp. 380–388.[28] R. Novak, Ecology of Nematodes in the Mediterranean Seagrass Posidonia oceanica (L.) Delile
1. General Part and Faunistics of the Nematode Community, Mar. Ecol. 10 (1989), pp. 335–363.[29] R. Danovaro, Detritus-Bacteria-Meiofauna interactions in a seagrass bed (Posidonia oceanica) of
the NW Mediterranean, Mar. Biol. 127 (1996), pp. 1–13.[30] R. Danovaro and C. Gambi, Biodiversity and trophic structure of nematode assemblages in
seagrass system: evidence for a coupling with changes in food availability, Mar. Biol. 141 (2002),
pp. 667–677.[31] R. Danovaro, C. Gambi, and S. Mirto, Meiofaunal production and energy transfer efficiency in a
seagrass (Posidonia oceanica) bed of the Western Mediterranean, Mar. Ecol. Prog. Ser. 234
(2002), pp. 95–104.[32] C. Gambi, S. Bianchelli, M. Perez, O. Invers, J. Ruiz, and R. Danovaro, Biodiversity response
to experimental induced hypoxic-anoxic conditions in seagrass sediments, Biodiversity Conserv.
18 (2009), pp. 33–54.[33] A. Peirano and C.N. Bianchi, Decline of the seagrass Posidonia oceanica in response to
environmental disturbance: a simulation-like approach off Liguria (NW Mediterranean Sea),
in The Response of Marine Organisms to their Environments, L.E. Hawkins and S. Hutchinson,
eds., University of Southampton, UK, 1997, pp. 87–95.[34] G. Diviacco and S. Coppo, Atlante degli habitat marini della Liguria: descrizione e cartografia
delle praterie di Posidonia oceanica e dei principali popolamenti marini costieri, Regione Liguria,
Genova, 2007.[35] G. Diviacco, Indagine sulla qualita dell’ambiente naturale costiero marino in Liguria. Stato di
conservazione delle biocenosi bentiche costiere, Regione Liguria, Ufficio Parchi e Aree Protette,
Genova, 1998.[36] H. Augier and G. Cristiani, Indice foliaire et biomasse de l’herbier de posidonies de la zone de
Carry-Sausset (Cote Bleue, Bouches du Rhone-France), in First International Workshop on
Posidonia oceanica Beds, C.F. Boudouresque, A. Jeudy de Grissac, and J. Olivier, eds.,
GIS Posidonie Publ., France, 1984, pp. 245–255.[37] D. Pessani, A. Caltagirone, F. Poncini, M. Vetere, Confronto tra due praterie di Posidonia
oceanica della Riviera Ligure di Levante e di Ponente. 2. La fauna epifita, in Second International
Workshop on Posidonia oceanica Beds, C.F. Boudouresque, A. Meinesz, E. Fresi, eds.,
GIS Posidonie Publ., France, 1989, pp. 161–173.
88 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

[38] M. Montefalcone, Ecosystem health assessment using the Mediterranean seagrass Posidonia
oceanica: A review, Ecol. Indic. 9 (2009), pp. 595–604.
[39] C.K. Wentworth, A scale of grade and class terms for clastic sediments, J. Geol. 30 (1922),
pp. 377–392.
[40] J.G. Parker, A comparison of methods used for the measurement of organic matter in sediments,
Chem. Ecol. 1 (1983), pp. 201–210.[41] E.F. Hartree, Determination of proteins: a modification of the Lowry method that give a linear
photometric response, Anal. Biochem. 48 (1972), pp. 422–427.[42] M. Fabiano, R. Danovaro, and S. Fraschetti, A three-year time series of elemental and
biochemical composition of organic matter in subtidal sandy sediment of the Ligurian Sea
(NW Mediterranean), Cont. Shelf Res. 15 (1995), pp. 1453–1469.[43] M. Dubois, K. Gilles, J.K. Mamilton, P.A. Rebers, and F. Smith, Colorimetric method
for determination of sugar and related substances, Anal. Chem. 28 (1956), pp. 350–356.[44] S.M. Gerchakov and P.G. Hatcher, Improved technique for analysis of carbohydrates in
sediments, Limnol. Oceanogr. 17 (1972), pp. 938–443.[45] C. Heip, M. Vincx, and G. Vranken, The ecology of marine nematodes, Oceanogr. Mar. Biol.
Annu. Rev. 23 (1985), pp. 399–489.[46] H.M. Platt and R.M. Warwick, Free-living Marine Nematodes. Part. I. British Enoplids.
Synopses of the British Fauna, Vol. 28, Cambridge University Press, Cambridge, 1983.[47] H.M. Platt and R.M. Warwick, Free-living Marine Nematodes. Part II. British Chromadorids.
Synopses of the British Fauna, Vol. 38., E.J. Brill/Dr Backhuys, Leiden, 1988.
[48] R.M. Warwick, H.M. Platt, and P.J. Somerfield, Freeliving Marine Nematodes. Part III.
Monhysterids, Synopses of the British Fauna, Vol. 53, Field Studies Council, Shrewsbury, 1998.
[49] M. Steyaert, T. Deprez, M. Raes, T. Bezerra, I. Demesel, S. Derycke, G. Desmet, G. Fonseca,
M.A. Franco, T. Gheskiere, E. Hoste, J. Ingels, T. Moens, J. Vanaverbeke, S. Van Gaever,
S. Vanhove, A. Vanreusel, D. Verschelde, and M. Vincx, 2005. Electronic Key to the free-living
marine Nematodes. Available at http://nemys.ugent.be/[50] R.M. Warwick and R. Price, Ecological and metabolic studies on free-living nematodes from an
estuarine mud-flat, Estuar. Coast. Mar. Sci. 9 (1979), pp. 257–271.[51] W. Wieser, Benthic studies in Buzzards Bay. II. The meiofauna, Limnol. Oceanogr. 5 (1960),
pp. 121–153.
[52] J.V. Juario, Nematode species composition and seasonal fluctuation of a sublitoral meiofauna
community in the German Bight, Veroff. Inst. Meeresforsch. Brem. 15 (1975), pp. 283–337.
[53] W. Wieser, Die Beziehung zwischen Mundhohlengestalt, Ernahrungsweise und Vorkommen bei
freilebenden marinen nematoden. Ark. Zool. 4 (1953), pp. 439–484.[54] M. Moreno, F. Semprucci, L. Vezzulli, M. Balsamo, M. Fabiano, and G. Albertelli, The use of
nematodes in assessing ecological quality status in the Mediterranean coastal ecosystems,
Ecol. Indic. 11 (2011), pp. 328–336.
[55] M.J. Anderson, PERMANOVA: a FORTRAN computer program for permutational multivariate
analysis of variance, Department of Statistics, University of Auckland, New Zealand, 2005.[56] J.S. Gray, The measurement of marine species diversity, with an application to the benthic fauna
of the Norwegian continental shelf, J. Exp. Mar. Biol. Ecol. 250 (2000), pp. 23–49.[57] B.H. McArdle and M.J. Anderson, Fitting multivariate models to community data: a comment on
distance-based redundancy analysis, Ecology 82 (2001), pp. 290–297.[58] K. Clarke and R.N. Gorley, Primer v6: user manual/tutorial, PRIMER-E, Plymouth, United
Kingdom, 2006.[59] M.J. Anderson, R.N. Gorley, and K.R. Clarke, PERMANOVAþ for PRIMER: Guide to
Software and Statistical Methods, PRIMER-E, Plymouth, United Kingdom, 2008.[60] European Community, Council Directive for a legislative frame and actions for the water policy,
2000/60/EC, Off. J. Eur. Comm., L 327, 23 October 2000.[61] Med-GIG, WFD Intercalibration technical report for coastal and transitional waters in the
Mediterranean ecoregion, in WFD Intercalibration Technical Report–Part 3: Coastal and
Advances in Oceanography and Limnology 89
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

Transitional Waters, 2007. Available at http://circa.europa.eu/Public/irc/jrc/jrc_eewai/library?
l¼/intercalibration_2&vm¼detailed&sb¼Title
[62] D. Balata, I. Bertocci, L. Piazzi, and U. Nesti, Comparison between epiphyte assemblages
of leaves and rhizomes of the seagrass Posidonia oceanica subjected to different levels of
anthropogenic eutrophication, Estuar. Coast. Shelf Sci. 79 (2008), pp. 533–540.[63] C. Pergent-Martini, V. Leoni, V. Pasqualini, G.D. Ardizzone, E. Balestri, R. Bedini,
A. Belluscio, T. Belsher, J. Borg, C.F. Boudouresque, S. Boumaza, J.M. Bouquegneau,
M.C. Buia, S. Calvo, J. Cebrian, E. Charbonnel, F. Cinelli, A. Cossu, G. Di Maida, B. Dural,
P. Francour, S. Gobert, G. Lepoint, A. Meinesz, H. Molenaar, M. Mansour, P. Panayotidis,
A. Peirano, G. Pergent, L. Piazzi, M. Pirrotta, G. Relini, J. Romero, J.L. Sanchez-Lizaso,
R. Semroud, P. Shembri, A. Shili, A. Tomasello, and B. Velimirov, Descriptors of Posidonia
oceanica meadows: use and application, Ecol. Indic. 5 (2005), pp. 213–230.
[64] A. Dell’Anno, M.L. Mei, A. Pusceddu, and R. Danovaro, Assessing the trophic state and
eutrophication of coastal marine systems: a new approach based on the biochemical composition of
sediment organic matter, Mar. Pollut. Bull. 44 (2002), pp. 611–622.[65] R. Danovaro, M. Fabiano, and M. Boyer, Seasonal changes of benthic bacteria in a seagrass
(Posidonia oceanica) bed in relation to the origin, composition and fate of the sediment organic
matter, Mar. Biol. 119 (1994), pp. 489–500.[66] S. Bianchelli, C. Gambi, D. Zeppilli, and R. Danovaro,Metazoan meiofauna in deep-sea canyons
and adjacent open slopes: a large-scale comparison with focus on the rare taxa, Deep-Sea Res.
Part I 57 (2010), pp. 420–433.[67] G. Fonseca, P. Hutchings, and F. Gallucci, Meiobenthic communities of seagrass beds (Zostera
capricorni) and unvegetated sediments along the coast of New South Wales, Australia, Estuar.
Coast. Shelf Sci. 91 (2011), pp. 69–77.
[68] A. Pusceddu, S. Bianchelli, C. Gambi, and R. Danovaro, Assessment of benthic trophic
status of marine coastal ecosystems: Significance of meiofaunal rare taxa, Estuar. Coast. Shelf
Sci. 93 (2011), pp. 420–430.[69] D.E. Duplisea and B.T. Hargrave, Response of meiobenthic size-structure, biomass and
respiration to sediment organic enrichment, Hydrobiologia 339 (1996), pp. 161–170.[70] A.D. Kennedy and C.A. Jacoby, Biological indicators of marine environmental health:
meiofauna – a neglected benthic component?, Environ. Monit. Assess. 54 (1999), pp. 47–68.[71] S. Mirto, T. La Rosa, R. Danovaro, and A. Mazzola, Microbial and meiofaunal response to
intensive Mussel-Farm biodeposition in coastal sediment of the western Mediterranean,
Mar. Pollut. Bull. 40 (2000), pp. 244–252.[72] L. Vezzulli, D. Marrale, M. Moreno, and M. Fabiano, Sediment organic matter and meiofauna
community response to long-term fish-farm impact in the Ligurian Sea (Western Mediterranean),
Chem. Ecol. 19 (2003), pp. 431–440.
[73] F. Semprucci, P. Boi, A. Manti, A. Covazzi Harriague, M. Rocchi, P. Colantoni, S. Papa, and
M. Balsamo, Benthic communities along a littoral of the Central Adriatic Sea (Italy), Helgol.
Mar. Res. 64 (2010), pp. 101–115.[74] J.H. Connell and R.D. Slatyer, Mechanisms of succession in natural communities and their role
in community stability and organisation, Am. Nat. 111 (1977), pp. 1119–1144.[75] F. Salas, J. Patrıcio, C. Marcos, M.A. Pardal, A. Perez-Ruzafa, and J.C. Marques, Are
taxonomic distinctness measures compliant to other ecological indicators in assessing ecological
status?, Mar. Pollut. Bull. 52 (2006), pp. 162–174.[76] M. Schratzberger and R.M. Warwick, Effects of the intensity and frequency of organic
enrichment on two estuarine nematode communities, Mar. Ecol. Prog. Ser. 164 (1998), pp. 83–94.[77] S. Mirto, T. La Rosa, C. Gambi, R. Danovaro, and A. Mazzola, Nematode community response
to fish-farm impact in the Western Mediterranean, Environ. Pollut. 116 (2002), pp. 203–214.[78] S.A. Netto and I. Valgas, The response of nematode assemblages to intensive mussel farming
in coastal sediments (Southern Brazil), Environ. Monit. Assess. 162 (2010), pp. 81–93.
90 V. Losi et al.
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2

[79] M. Steyaert, N. Gamer, D. Van Gansbeke, and M. Vincx, Nematode communities from theNorth Sea: environmental controls on species diversity and vertical distribution within the sediment,J. Mar. Biol. Assoc. U.K. 79 (1999), pp. 253–264.
[80] M. Moreno, G. Albertelli, and M. Fabiano, Nematode response to metal, PAHs and organic
enrichment in tourist marinas of the Mediterranean sea, Mar. Pollut. Bull. 58 (2009),pp. 1192–1201.
[81] C. Van Colen, F. Montserrat, K. Verbist, M. Vincx, M. Steyaert, J. Vanaverbeke,
P.M.J. Herman, S. Degraer, and T. Ysebaert, Tidal flat nematode responses to hypoxiaand subsequent macrofauna-mediated alterations of sediment properties, Mar. Ecol. Prog. Ser.381 (2009), pp. 189–197.
[82] A. Hedfi, E. Mahmoudi, F. Boufahja, H. Beyrem, and P. Aıssa, Effects of increasing levels ofnickel contamination on structure of offshore nematode communities in experimental microcosms,Bull. Environ. Contam. Toxicol. 79 (2007), pp. 345–349.
[83] P.J. Somerfield, S.L. Dashfield, and R.M. Warwick, Three-dimensional spatial structure:nematodes in a sandy tidal flat, Mar. Ecol. Prog. Ser. 336 (2007), pp. 177–186.
[84] R. Gingold, S.E. Ibarra-Obando, and A. Rocha-Olivares, Spatial aggregation patterns offree-living marine nematodes in contrasting sandy beach micro-habitats, J. Mar. Biol. Assoc. U.K.
91 (2011), pp. 615–622.[85] K. Soetaert, A. Muthumbi, and C. Heip, Size and shape of ocean margin nematodes:
morphological diversity and depth-related patterns, Mar. Ecol. Prog. Ser. 242 (2002), pp. 179–193.
[86] M. Raes and A. Vanreusel, Microhabitat type determines the composition of nematodecommunities associated with sediment-clogged cold-water coral framework in the PorcupineSeabight (NE Atlantic), Deep-Sea Research I 53 (2006), pp. 1880–1894.
[87] J. Vanaverbeke, K. Soetaert, and M. Vincx, Changes in morphometric characteristics ofnematode communities during a spring phytoplankton bloom deposition, Mar. Ecol. Prog. Ser.273 (2004), pp. 139–146.
[88] M. Schratzberger and S. Jennings, Impacts of chronic trawling disturbance on meiofaunal
communities, Mar. Biol. 141 (2002), pp. 991–1000.[89] M. Steyaert, L. Moodley, J. Vanaverbeke, S. Vandewiele, and M. Vincx, Laboratory
experiments on the infaunal activity of intertidal nematodes, Hydrobiologia 540 (2005),
pp. 217–223.[90] I. De Mesel, S. Derycke, J. Swings, M. Vincx, and T. Moens, Influence of bacterivorous
nematodes on the decomposition of cordgrass, J. Exp. Mar. Biol. Ecol. 296 (2003), pp. 227–242.
[91] R.W. Harvey, R.L. Smith, and L. George, Effect of organic contamination upon microbialdistributions and heterotrophic uptake in a Cape Cod, Mass., Aquifer, Appl. Environ. Microbiol.48 (1984), pp. 1197–1202.
[92] S.G. Horrigan, A. Hagstrom, I. Koike, and F. Azam, Inorganic nitrogen utilization byassemblages of marine bacteria in seawaterculture, Mar. Ecol. Prog. Ser. 50 (1988), pp. 147–150.
[93] R. Sandulli, C. De Leonardis, and J. Vanaverbeke, 2010. Meiobenthic communities in the shallowsubtidal of three Italian Marine Protected Areas, Ital. J. Zool. 77 (2010), pp. 186–196.
[94] M. Montefalcone, G. Albertelli, C. Morri, V. Parravicini, and C.N. Bianchi, Legal protection isnot enough: Posidonia oceanica meadows in marine protected areas are not healthier than thosein unprotected areas of the northwest Mediterranean Sea, Mar. Pollut. Bull. 58 (2009),
pp. 515–519.[95] L. Mangialajo, N. Ruggieri, V. Asnaghi, M. Chiantore, P. Povero, and R. Cattaneo-Vietti,
Ecological status in the Ligurian Sea: The effect of coastline urbanisation and the importance of
proper reference sites, Mar. Pollut. Bull. 55 (2007), pp. 30–41.
Advances in Oceanography and Limnology 91
Dow
nloa
ded
by [
Val
entin
a L
osi]
at 1
4:08
25
May
201
2
Top Related
Copyright © 2022 FDOKUMEN

