







Bahasa
Halaman
Hukum


Pest Management Science Pest Manag Sci 59:1333–1343 (online: 2003)DOI: 10.1002/ps.766
Multiple mechanisms account for variation inbase-line sensitivity to azole fungicides in fieldisolates of Mycosphaerella graminicolaIoannis Stergiopoulos,1 Johannes GM van Nistelrooy,1 Gert HJ Kema2 andMaarten A De Waard1∗1Laboratory of Phytopathology, Department of Plant Sciences, Wageningen University, PO Box 8025, Binnenhaven 5, 6709 PDWageningen, The Netherlands2Plant Research International, PO Box 16, 6700 AA Wageningen, The Netherlands
Abstract: Molecular mechanisms that account for variation in base-line sensitivity to azole fungicideswere examined in a collection of twenty field isolates, collected in France and Germany, of the wheatpathogen Mycosphaerella graminicola (Fuckel) Schroeter. The isolates tested represent the wide base-line sensitivity to the azole fungicide tebuconazole described previously. The isolates were cross-sensitiveto other azoles tested, such as cyproconazole and ketoconazole, but not to unrelated chemicals likecycloheximide, kresoxim-methyl or rhodamine 6G. Progenies from a genetic cross between an isolatewith an intermediate and a high sensitivity to azoles displayed a continuous range of phenotypes withrespect to cyproconazole sensitivity, indicating that variation in azole sensitivity in this haploid organismis polygenic. The basal level of expression of the ATP-binding cassette transporter genes MgAtr1-MgAtr5from M graminicola significantly varied amongst the isolates tested, but no clear increase in the transcriptlevel of a particular MgAtr gene was found in the less sensitive isolates. Cyproconazole strongly inducedexpression of MgAtr4, but no correlation between expression levels of this gene and azole sensitivity wasobserved. One isolate with intermediate sensitivity to azoles over-expressed CYP51, encoding cytochromeP450 sterol 14α-demethylase from M graminicola. Isolates with a low or high sensitivity to azoles weretested for accumulation of cyproconazole, but no clear correlation between reduced accumulation of thefungicide in mycelium and sensitivity to azoles was observed. Therefore, differences in accumulationcannot account exclusively for the variation in base-line sensitivity of the isolates to azoles. The resultsindicate that multiple mechanisms account for differences in base-line sensitivity to azoles in field isolatesof M graminicola. 2003 Society of Chemical Industry
Keywords: ATP-binding cassette (ABC) transporters; azoles; base-line sensitivity; CYP51; DMIs; fungicideresistance; Septoria tritici
1 INTRODUCTIONAzoles represent a major class of fungicides that hasbeen extensively used over the past three decadesin the control of fungal pathogens of medicaland agricultural importance. They are systemicfungicides with both protective and curative activityin disease control.1 Their mode of action is basedon inhibition of cytochrome P450 sterol 14α-demethylase (P45014DM), a key enzyme involved in thebiosynthesis of ergosterol in fungi. For this reason azolefungicides are also described as sterol demethylationinhibitors (DMIs).2
Because of their site-specific mode of action,the risk of resistance developing to DMIs is
significant. However, laboratory-generated azole-resistant mutants often display reduced fitness withrespect to spore germination, mycelial growth and vir-ulence, and their levels of resistance are low. For thesereasons, resistance development under field condi-tions was considered unlikely.3–5 Nevertheless, DMIresistance emerged in several pathogen populations,although relatively slowly as compared to other classesof fungicide.6 One of the first cases of field resistancewas reported for Sphaerotheca fuliginea (Schlecht ex Fr)Poll, the causal agent of cucumber powdery mildew.7
Since then, DMI resistance is also reported in otherfungal pathogens, such as Penicillium digitatum (Pers)Sacc,8 Erysiphe graminis DC f sp hordei Marchal,9
∗ Correspondence to: Maarten A De Waard, Laboratory of Phytopathology, Department of Plant Sciences, Wageningen University, PO Box8025, Binnenhaven 5, 6709 PD Wageningen, The NetherlandsE-mail: [email protected]/grant sponsor: The European Commission, Mobility of Researchers (TMR) Programme—Marie Curie Research Training Grants;contract/grant number: ERBFMBICT983558(Received 5 September 2002; revised version received 25 March 2003; accepted 29 May 2003)Published online 5 August 2003
2003 Society of Chemical Industry. Pest Manag Sci 1526–498X/2003/$30.00 1333

I Stergiopoulos et al
Venturia inaequalis (Cooke) Winter10 and Rhynchospo-rium secalis (Oudem) Davis.11
Analysis of azole resistance in fungi revealed that dif-ferent molecular mechanisms may operate in resistantisolates. In Candida albicans (Robin) Berkhout, sev-eral alterations in CYP51 (also described as ERG11),that encodes P45014DM, have been identified. Theseinclude point mutations in the coding region of CYP51that result in reduced affinity of azoles for the targetenzyme12 or over-expression of this gene resulting inhigher intracellular levels of P45014DM.13–17 Similaralterations in CYP51 have been reported in fungalpathogens of agricultural importance. Point mutationsin CYP51 associated with decreased azole sensitivityhave been described in laboratory-generated mutantsof Ustilago maydis (DC) Corda18 and P digitatum,19
and in field isolates of Uncinula necator (Schw) Burr20
and E graminis f sp hordei.21 Resistance to azoles inP digitatum and V inaequalis correlates positively withincreased expression of CYP51 caused by the pres-ence of tandem repeats in the promoter region ofthis gene.22,23
Reduced accumulation of azoles in cells as aresult of active efflux mechanisms constitutes anotherwell-characterised mechanism of resistance. Earlyreports indicated that fenarimol-resistant strains of Anidulans accumulate less fungicide than the wild-typecontrol.24,25 This reduced accumulation is ascribedto an increased energy-dependent efflux of thecompounds from cells, mediated by ATP-bindingcassette (ABC) transporters.26,27 ABC transporterscomprise one of the best-characterised protein familiesassociated with active excretion of drugs. They utilisethe energy derived from hydrolysis of ATP to drivethe transport of a wide variety of cytotoxic agentsover biological membranes.28 In particular, ABCtransporters of filamentous fungi are known to exhibitan important function in protection against synthetictoxic compounds such as antibiotics, fungicidesand other xenobiotics.29 Overproduction of ABCtransporters can result in pleiotropic effects suchas simultaneous resistance to structurally unrelatedcompounds, a phenomenon described as multi-drugresistance (MDR).30
Despite the extensive knowledge of molecular mech-anisms of azole resistance in laboratory-generatedmutants, the relevance and frequency of such mech-anisms in field populations of fungal pathogens isstill unclear. In P digitatum increased expression lev-els of ABC transporters genes such as PMR1 andPMR5, as well as increased expression of CYP51, weredetected in DMI-resistant isolates.22 In this paperwe studied mechanisms that could potentially explainthe wide variation in base-line sensitivity to azolesin field isolates of the fungus Mycosphaerella gramini-cola (Fuckel) J Schroeter in Cohn (anamorph: Septoriatritici Roberge in Desmaz), the causal agent of septo-ria tritici blotch of wheat. The economic importanceof this disease has dramatically increased over thepast few decades and control management of this
pathogen has widely involved the use of azole fungi-cides. In laboratory-generated azole-resistant mutantsof M graminicola, different mechanisms of resistance toazoles may operate, amongst others, over-expressionof specific ABC transporter genes.31 Resistance devel-opment in M graminicola to azole fungicides underfield conditions has not been reported.32 However,field populations of M graminicola consist of iso-lates with a broad range in sensitivity levels to thesefungicides.33,34 Here we report studies on twenty fieldisolates of M graminicola, sampled in France and Ger-many, that possess a significant difference in base-linesensitivity to the azole fungicide tebuconazole. Anal-ysis of the progeny of a genetic cross between twoisolates with a 100-fold difference in sensitivity tocyproconazole indicated that azole sensitivity in thisfungus is polygenic. Expression studies of ABC trans-porter genes and CYP51 showed that isolates displaylarge differences in basal level of expression of thesegenes. Accumulation of cyproconazole did not cor-relate with azole sensitivity. The results indicate thatmultiple mechanisms may operate in field isolates ofM graminicola that determine base-line sensitivity toazole fungicides.
2 MATERIALS AND METHODS2.1 Fungal strainsEleven field isolates of M graminicola originating fromFrance and nine field isolates derived from Germany(see Table 1) were kindly provided by Dr JM Seng(Biotransfer, Montreuil Cedex, France) and Dr ASuty (Bayer AG, Landwirtschaftszentrum Monheim,Leverkusen, Germany), respectively. The isolates weresampled from wheat fields treated with tebuconazoleand selected for differential sensitivity to this fungicide.Single spore isolates were prepared from bulk samplesand used throughout the whole study. A genetic crossbetween isolates M1 and M3 was made accordingto Kema et al35 and 40 progeny isolates (M1M31–M1M3 40) were obtained. Stock cultures of theisolates were kept as yeast-like cells at −80 ◦C.
2.2 Culture conditionsYeast-like cells of the isolates were grown in liq-uid Yeast Sucrose Medium (YSM; yeast extract10 g litre−1, sucrose 10 g litre−1) at 18 ◦C and140 rev min−1. Mycelium was grown in liquid Cza-pek Dox-Mycological Peptone medium (CzD-MP;Czapek Dox 33.4 g litre−1, mycological peptone5 g litre−1) at 25 ◦C and 140 rev min−1. Toxicity assayswere performed in Potato Dextrose Broth (PDB;24 g litre−1) at 25 ◦C.
2.3 CompoundsThe chemicals used in this study were the azolefungicides cyproconazole (Syngenta, Basel, Switzer-land), ketoconazole (Janssen Research Founda-tion, Beerse, Belgium), and tebuconazole (BayerAG, Landwirtschaftszentrum Monheim, Leverkusen,
1334 Pest Manag Sci 59:1333–1343 (online: 2003)

Variation in base-line sensitivity to azole fungicides
Germany), the antibiotic cycloheximide (Sigma–Aldrich Chemie BV, Zwijndrecht, The Netherlands),the Qo-inhibitor (QoI) kresoxim-methyl (BASF AG,Limburgerhof, Germany) and the dye rhodamine6G (Sigma–Aldrich Chemie BV, Zwijndrecht, TheNetherlands). All compounds were of technical gradeand used as 100× concentrated stock solutions of1000, 562, 316, 178, 100, 56.2, 31.6, 17.8, 10, 5.62,3.16, 1.78, 1, 0.562, 0.316, 0.178 and 0.1 mg litre−1
in methanol.
2.4 Fungicide activity testSensitivity of isolates to compounds was assessed ina microtitre plate test, adapted from Pijls et al36 Cellsuspensions (105 ml−1) of the isolates were preparedin PDB from 3-day-old cultures. Wells of sterileflat-bottomed microtitre plates (Greiner BV, Alphenaan de Rijn, The Netherlands) were filled with cellsuspension (50 µl) and mixed’ with PDB (150 µl)amended with fungicides at different concentrations.PDB with methanol (1%) was used as a control. Alltreatments were tested in fourfold per microtitre plateand tests were repeated at least once. Plates were sealedto prevent evaporation from wells and incubated for5 days as still cultures at 25 ◦C in the presence oflight. Growth was assessed with a microtitre platereader (Bio-Tek Instruments Inc, Vermont, U.S.A.)by measuring cell density at 630 nm. Sensitivities of theisolates to the compounds were expressed as minimuminhibitory concentration (MIC) values. These valueswere the lowest concentration of compound in thescale used that did not allow growth. The variationfactor of isolates for sensitivity to compounds wasdefined as the ratio between the MIC value of themost and least sensitive isolate.
2.5 Expression analysis2.5.1 RNA isolation and northern blot analysisYSM or CzD-MP (20 ml) was inoculated withyeast-like cells derived from stock cultures of theisolates tested. After 3 days of culturing at 18 or25 ◦C and 140 rev min−1, the optical density of thecultures was determined at 600 nm (OD600) andnew cultures (20 ml) with a starting density ofOD600 0.1 were initiated. Three days later, theOD600 of these cultures was adjusted to 0.5 in freshmedium (20 ml) and the cultures were incubatedagain overnight under similar conditions as describedabove. Cyproconazole treatments (10 mg litre−1, 0.1%methanol) were performed at 140 rev min−1 for 1 h.Yeast-like cells or mycelium were grown in YSMat 18 ◦C or CzD-MP at 25 ◦C, respectively. Controltreatments were performed with methanol (0.1%).Fungal biomass was harvested by centrifugation at10 000 rev min−1 and 0 ◦C for 10 min, and instantlyfrozen in liquid nitrogen.
RNA was isolated from frozen biomass with theTRIzol reagent (Life Technologies Inc, Maryland,USA) according to the manufacturer’s instructions.RNA (10 µg, 4.5 µl) was denatured in a mixture
of glyoxal (6 M; 4.5 µl), sodium phosphate (0.1 M;3.0 µl), and dimethyl sulfoxide (13.3 µl) at 50 ◦Cfor 1 h, and subjected to electrophoresis on 1.6%agarose gel in sodium phosphate buffer (10 mM;pH 7.0). Blotting was carried out on Hybond-N+
membranes (Amersham Pharmacia Biotech, LittleChalfont Buckinghamshire, UK) by capillary transferwith 10× SSC solution. Homologous hybridisationswere performed overnight at 65 ◦C in ‘Nasmyth’ssolution’ buffer (dextran sulfate 185 g litre−1, sarcosyl18.5 g litre−1, EDTA 0.011 M, Na2HPO4 0.3 M, NaCl1.1 M; pH 6.2). This solution (5.4 ml) was mixedwith distilled water (4.6 ml) just before use to obtainthe hybridisation buffer (10 ml) while sheared herringsperm DNA (100 µg ml−1) was included as blockingagent. Blots were washed twice in 2× SSC, 0.1× SDSand twice in 0.1× SSC, 0.1× SDS at 65 ◦C for 15 min.
2.5.2 ProbesGene-specific probes of the ABC transporter genesMgAtr1 (Acc AJ243112), MgAtr2 (Acc AJ243113),MgAtr3 (Acc AF364105), MgAtr4 (Acc AF329852)and MgAtr5 (Acc AF364104) used in northern blotanalysis represented a 0.84-kb EcoRI fragment ofMgAtr1, a 0.75-kb SalI fragment of MgAtr2, a 0.85-kb SalI fragment of MgAtr3, an 1.14-kb BamHI/PstIfragment of MgAtr4, and a 0.6-kb EcoRI fragmentof MgAtr5. A 0.65-kb DNA fragment of CYP51 (AccAF263470), encoding P45014DM of M graminicola, wasobtained by PCR using genomic DNA from isolateIPO32337 as template. Primers used for amplificationwere CYPinternal4 (5′-CAG CAC TCT TCA TCTGCG AC-3′) and CYP3′anchor (5′-TCC CTC CTCTCC CAC TTT AC-3′). PCR conditions consistedof 30 cycles of 1 min denaturation at 94 ◦C, 1 minof annealing at 58 ◦C, and 1 min of extension at72 ◦C with an additional extension step of 10 minat 72 ◦C at the end of the reaction. Equal loadingof samples on blots was examined with a 0.6-kb EcoRI fragment of the 18S rRNA gene fromAspergillus niger.
Randomly primed DNA isotopic probes wereprepared by enzymatic incorporation of [α32P]dATP.In each labelling reaction, 50 ng of a probe templatewas used. Probes were purified using the QIAquickNucleotide Removal kit (QIAGEN, Westburg BV,Leusden, The Netherlands) before adding to thehybridisation solution.
2.6 Accumulation of [14C]cyproconazoleThree-day-old mycelial cultures were harvested byfiltering through a 0.85-mm pore sieve and by col-lection of the mycelium on a 0.055-mm pore sieve,washed with phosphate buffer (50 mM; pH 6.0)and re-suspended in the same buffer at a net wetweight of 6 mg litre−1. Cultures were incubated at25 ◦C and 140 rev min−1 for 30 min before addi-tion of [14C]cyproconazole, at an initial externalconcentration of 100 µM. Mycelium from cultures
Pest Manag Sci 59:1333–1343 (online: 2003) 1335
I Stergiopoulos et al
(5 ml) was collected by vacuum filtration at inter-vals of 5, 15, 25, 35, 45, 55, 95 and 105 min afteraddition of [14C]cyproconazole, washed (5×) withphosphate buffer (pH 6.0; 5 ml) and stored in vialscontaining LUMASAFE PLUS (3 ml; Groningen,The Netherlands). Energy-dependency of cyprocona-zole accumulation was examined by addition ofcarbonyl-cyanide 3-chlorophenylhydrazone (CCCP,20 µM) 55 min after addition of [14C]cyproconazole.Radioactivity in the samples was measured witha Beckman LS6000TA liquid scintillation counter(Beckman Instruments Inc, Fullerton, USA). Accu-mulation of [14C]cyproconazole was calculatedas nmol of cyproconazole per mg dry weightof biomass.
2.7 Statistical analysisStatistical analysis of the results was performed usingSPSS for Windows release 10.0.5 (SPSS Inc, Chicago,Illinois, USA) and MicrocalOrigin version 5.0(Microcal Software Inc, Northampton, USA) statisti-cal software. Linear association between sensitivityto different compounds or between sensitivity toazoles and accumulation of [14C]cyproconazole wasexamined using the Pearson’s correlation coefficient.Statistically significant differences between mean val-ues of [14C]cyproconazole accumulation were testedusing the Student–Newman–Keuls (S-N-K) analy-sis at a significance level of 0.05 using 11 degreesof freedom.
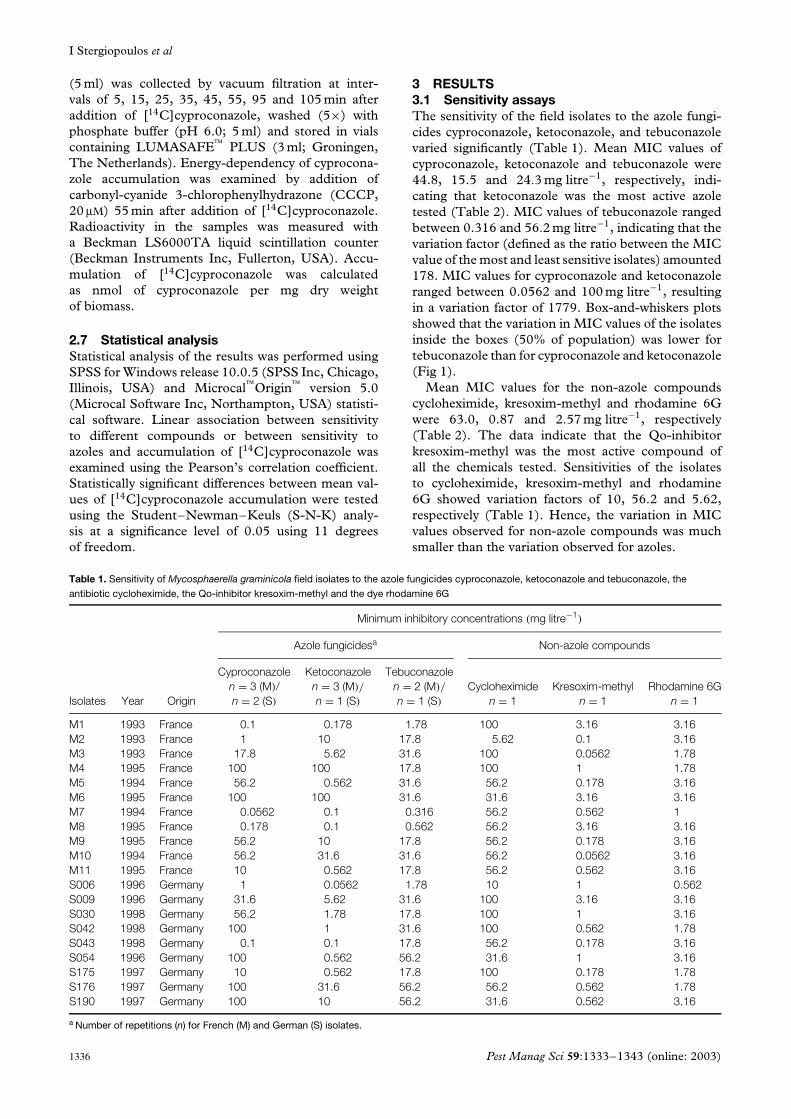
3 RESULTS3.1 Sensitivity assaysThe sensitivity of the field isolates to the azole fungi-cides cyproconazole, ketoconazole, and tebuconazolevaried significantly (Table 1). Mean MIC values ofcyproconazole, ketoconazole and tebuconazole were44.8, 15.5 and 24.3 mg litre−1, respectively, indi-cating that ketoconazole was the most active azoletested (Table 2). MIC values of tebuconazole rangedbetween 0.316 and 56.2 mg litre−1, indicating that thevariation factor (defined as the ratio between the MICvalue of the most and least sensitive isolates) amounted178. MIC values for cyproconazole and ketoconazoleranged between 0.0562 and 100 mg litre−1, resultingin a variation factor of 1779. Box-and-whiskers plotsshowed that the variation in MIC values of the isolatesinside the boxes (50% of population) was lower fortebuconazole than for cyproconazole and ketoconazole(Fig 1).
Mean MIC values for the non-azole compoundscycloheximide, kresoxim-methyl and rhodamine 6Gwere 63.0, 0.87 and 2.57 mg litre−1, respectively(Table 2). The data indicate that the Qo-inhibitorkresoxim-methyl was the most active compound ofall the chemicals tested. Sensitivities of the isolatesto cycloheximide, kresoxim-methyl and rhodamine6G showed variation factors of 10, 56.2 and 5.62,respectively (Table 1). Hence, the variation in MICvalues observed for non-azole compounds was muchsmaller than the variation observed for azoles.
Table 1. Sensitivity of Mycosphaerella graminicola field isolates to the azole fungicides cyproconazole, ketoconazole and tebuconazole, the
antibiotic cycloheximide, the Qo-inhibitor kresoxim-methyl and the dye rhodamine 6G
Minimum inhibitory concentrations (mg litre−1)
Azole fungicidesa Non-azole compounds
Isolates Year Origin
Cyproconazolen = 3 (M)/n = 2 (S)
Ketoconazolen = 3 (M)/
n = 1 (S)
Tebuconazolen = 2 (M)/
n = 1 (S)
Cycloheximiden = 1
Kresoxim-methyln = 1
Rhodamine 6Gn = 1
M1 1993 France 0.1 0.178 1.78 100 3.16 3.16M2 1993 France 1 10 17.8 5.62 0.1 3.16M3 1993 France 17.8 5.62 31.6 100 0.0562 1.78M4 1995 France 100 100 17.8 100 1 1.78M5 1994 France 56.2 0.562 31.6 56.2 0.178 3.16M6 1995 France 100 100 31.6 31.6 3.16 3.16M7 1994 France 0.0562 0.1 0.316 56.2 0.562 1M8 1995 France 0.178 0.1 0.562 56.2 3.16 3.16M9 1995 France 56.2 10 17.8 56.2 0.178 3.16M10 1994 France 56.2 31.6 31.6 56.2 0.0562 3.16M11 1995 France 10 0.562 17.8 56.2 0.562 3.16S006 1996 Germany 1 0.0562 1.78 10 1 0.562S009 1996 Germany 31.6 5.62 31.6 100 3.16 3.16S030 1998 Germany 56.2 1.78 17.8 100 1 3.16S042 1998 Germany 100 1 31.6 100 0.562 1.78S043 1998 Germany 0.1 0.1 17.8 56.2 0.178 3.16S054 1996 Germany 100 0.562 56.2 31.6 1 3.16S175 1997 Germany 10 0.562 17.8 100 0.178 1.78S176 1997 Germany 100 31.6 56.2 56.2 0.562 1.78S190 1997 Germany 100 10 56.2 31.6 0.562 3.16
a Number of repetitions (n) for French (M) and German (S) isolates.
1336 Pest Manag Sci 59:1333–1343 (online: 2003)
Variation in base-line sensitivity to azole fungicides
1000
100
10
1
0.1
0.01
MIC
s (m
g lit
re-1
)
Cyp
roco
nazo
le
Ket
ocon
azol
e
Tebu
cona
zole
Cyc
lohe
xim
ide
Kre
soxi
m-
met
hyl
Rho
dam
ine
6G
Mean50 % of population
Median outliers
Figure 1. Box-and-whiskers plots of MIC values of 20 field isolates ofMycosphaerella graminicola to the azole fungicides cyproconazole,ketoconazole, and tebuconazole, the antibiotic cycloheximide, theQo-inhibitor fungicide kresoxim-methyl, and the dye rhodamine 6G.
Table 2. Sensitivity of field isolates of Mycosphaerella graminicola to
the azole fungicides cyproconazole, ketoconazole and tebuconazole,
the antibiotic cycloheximide, the Qo-inhibitor kresoxim-methyl and
the dye rhodamine 6G
Minimum inhibitory concentrations(mg litre−1)a
Compounds Minimum Maximum Mean Median
Cyproconazole 0.0562 100 44.83 44.25Ketoconazole 0.0562 100 15.50 11.66Tebuconazole 0.316 56.2 24.36 17.80Cycloheximide 5.62 100 63.00 56.20Kresoxim-methyl 0.0562 3.16 0.87 0.79Rhodamine 6G 0.562 3.16 2.57 3.16
a Average of 20 isolates tested.
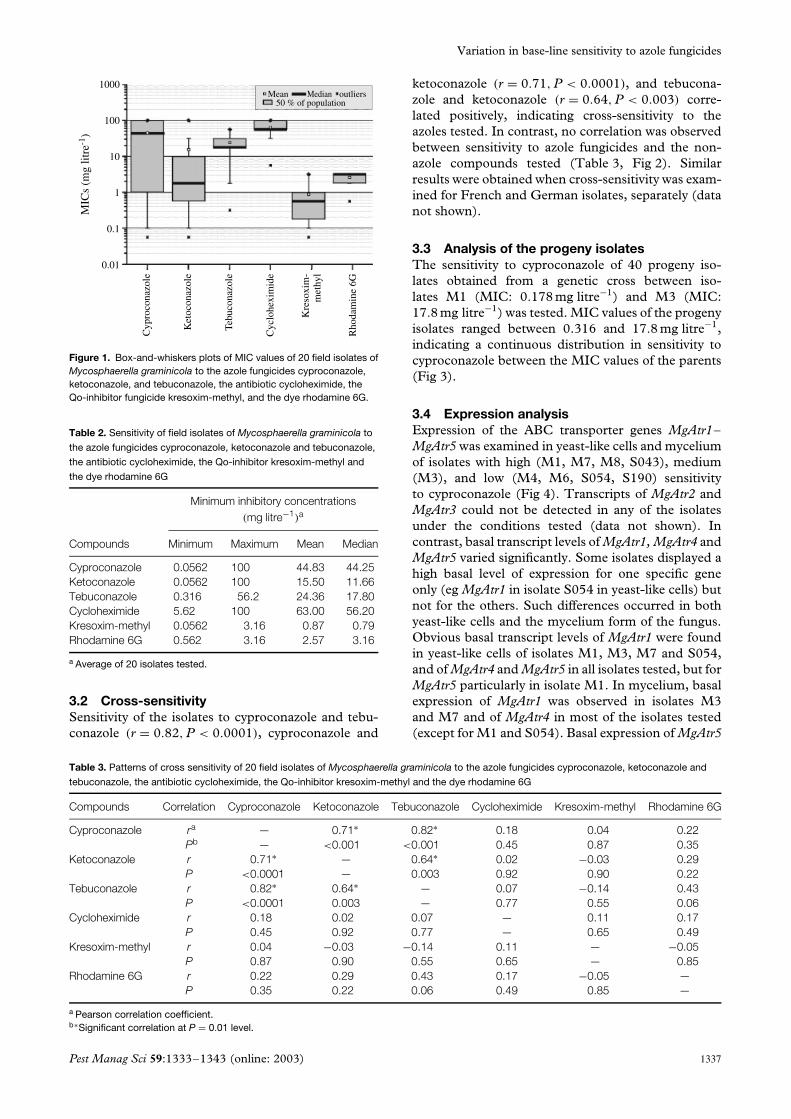
3.2 Cross-sensitivitySensitivity of the isolates to cyproconazole and tebu-conazole (r = 0.82, P < 0.0001), cyproconazole and
ketoconazole (r = 0.71, P < 0.0001), and tebucona-zole and ketoconazole (r = 0.64, P < 0.003) corre-lated positively, indicating cross-sensitivity to theazoles tested. In contrast, no correlation was observedbetween sensitivity to azole fungicides and the non-azole compounds tested (Table 3, Fig 2). Similarresults were obtained when cross-sensitivity was exam-ined for French and German isolates, separately (datanot shown).
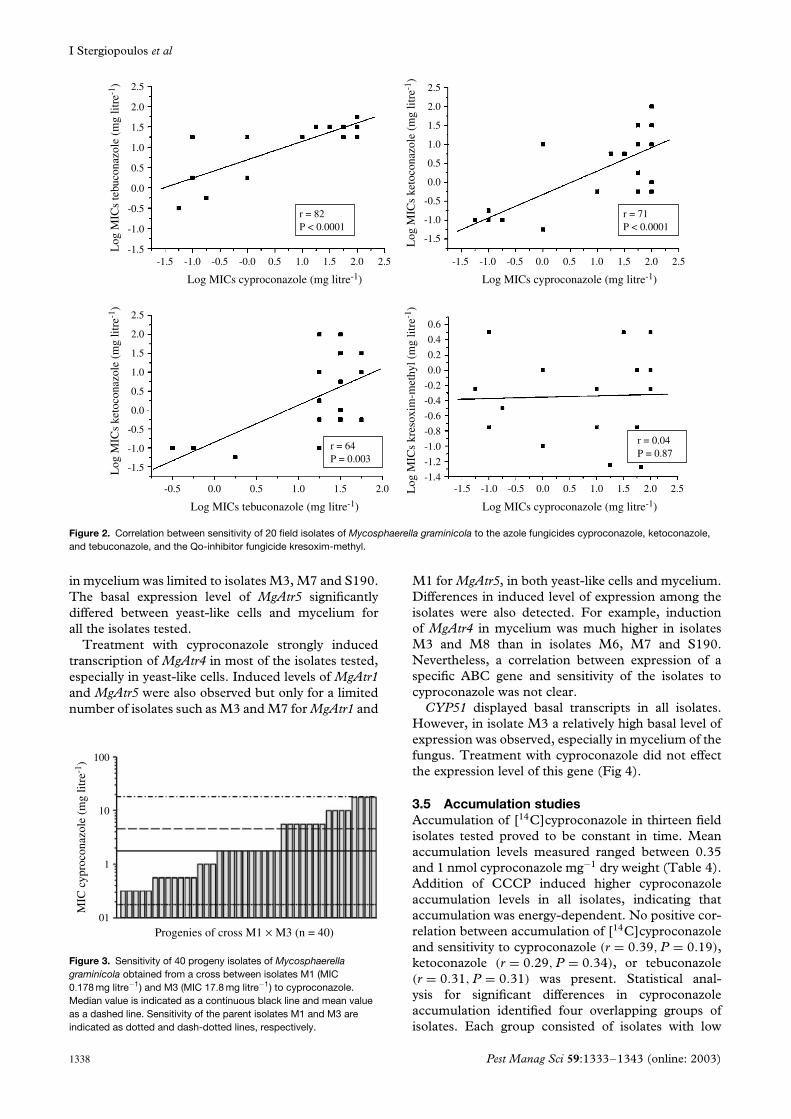
3.3 Analysis of the progeny isolatesThe sensitivity to cyproconazole of 40 progeny iso-lates obtained from a genetic cross between iso-lates M1 (MIC: 0.178 mg litre−1) and M3 (MIC:17.8 mg litre−1) was tested. MIC values of the progenyisolates ranged between 0.316 and 17.8 mg litre−1,indicating a continuous distribution in sensitivity tocyproconazole between the MIC values of the parents(Fig 3).
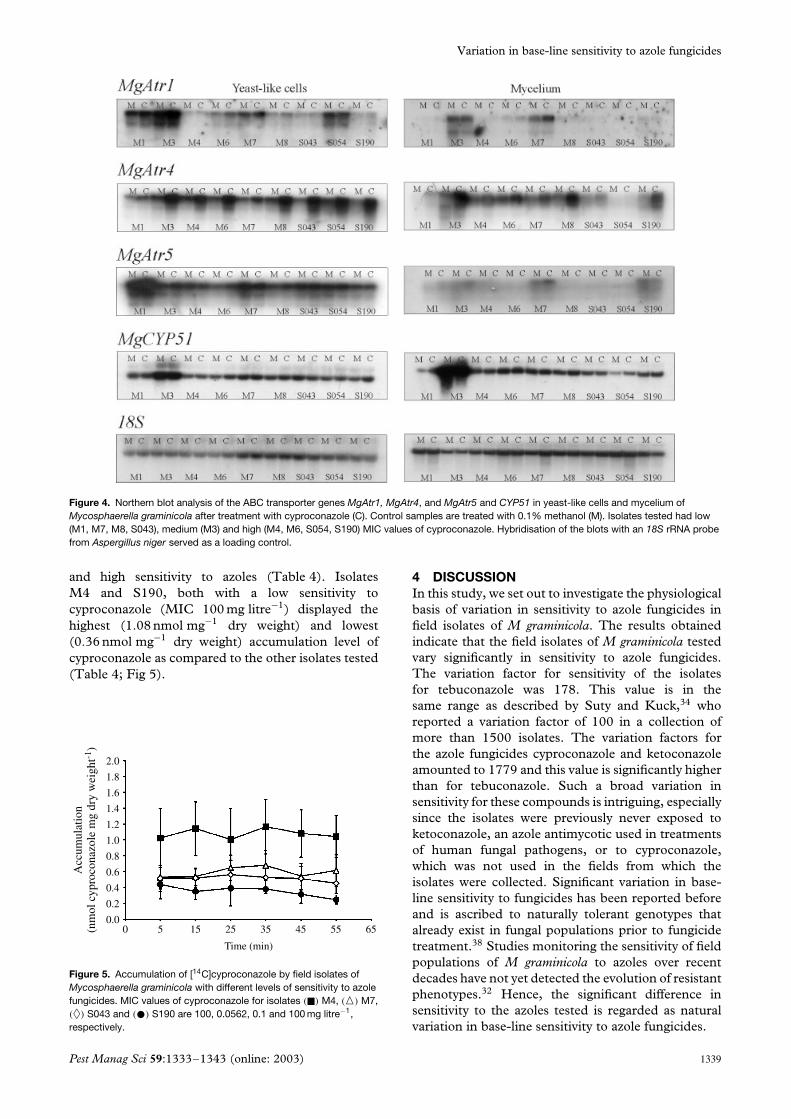
3.4 Expression analysisExpression of the ABC transporter genes MgAtr1–MgAtr5 was examined in yeast-like cells and myceliumof isolates with high (M1, M7, M8, S043), medium(M3), and low (M4, M6, S054, S190) sensitivityto cyproconazole (Fig 4). Transcripts of MgAtr2 andMgAtr3 could not be detected in any of the isolatesunder the conditions tested (data not shown). Incontrast, basal transcript levels of MgAtr1, MgAtr4 andMgAtr5 varied significantly. Some isolates displayed ahigh basal level of expression for one specific geneonly (eg MgAtr1 in isolate S054 in yeast-like cells) butnot for the others. Such differences occurred in bothyeast-like cells and the mycelium form of the fungus.Obvious basal transcript levels of MgAtr1 were foundin yeast-like cells of isolates M1, M3, M7 and S054,and of MgAtr4 and MgAtr5 in all isolates tested, but forMgAtr5 particularly in isolate M1. In mycelium, basalexpression of MgAtr1 was observed in isolates M3and M7 and of MgAtr4 in most of the isolates tested(except for M1 and S054). Basal expression of MgAtr5
Table 3. Patterns of cross sensitivity of 20 field isolates of Mycosphaerella graminicola to the azole fungicides cyproconazole, ketoconazole and
tebuconazole, the antibiotic cycloheximide, the Qo-inhibitor kresoxim-methyl and the dye rhodamine 6G
Compounds Correlation Cyproconazole Ketoconazole Tebuconazole Cycloheximide Kresoxim-methyl Rhodamine 6G
Cyproconazole ra — 0.71∗ 0.82∗ 0.18 0.04 0.22Pb — <0.001 <0.001 0.45 0.87 0.35
Ketoconazole r 0.71∗ — 0.64∗ 0.02 −0.03 0.29P <0.0001 — 0.003 0.92 0.90 0.22
Tebuconazole r 0.82∗ 0.64∗ — 0.07 −0.14 0.43P <0.0001 0.003 — 0.77 0.55 0.06
Cycloheximide r 0.18 0.02 0.07 — 0.11 0.17P 0.45 0.92 0.77 — 0.65 0.49
Kresoxim-methyl r 0.04 −0.03 −0.14 0.11 — −0.05P 0.87 0.90 0.55 0.65 — 0.85
Rhodamine 6G r 0.22 0.29 0.43 0.17 −0.05 —P 0.35 0.22 0.06 0.49 0.85 —
a Pearson correlation coefficient.b∗Significant correlation at P = 0.01 level.
Pest Manag Sci 59:1333–1343 (online: 2003) 1337
I Stergiopoulos et al
2.5
2.0
1.5
1.0
0.5
0.0
-0.5
-1.0
-1.5-1.5 -1.0 -0.5 -0.0 0.5 1.0 1.5 2.0 2.5
Log
MIC
s te
buco
nazo
le (
mg
litre
-1) 2.5
2.0
1.5
1.0
0.5
0.0
-0.5
-1.0
-1.5Log
MIC
s ke
toco
nazo
le (
mg
litre
-1)
0.6
0.0
0.4
0.2
-0.2
-0.4
-0.6
-0.8
-1.0
-1.2
-1.4-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5L
og M
ICs
kres
oxim
-met
hyl (
mg
litre
-1)
Log MICs cyproconazole (mg litre-1)
-1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5
Log MICs cyproconazole (mg litre-1)
Log MICs cyproconazole (mg litre-1)
-0.5 0.0 0.5 1.0 1.5 2.0
Log MICs tebuconazole (mg litre-1)
2.5
2.0
1.5
1.0
0.5
0.0
-0.5
-1.0
-1.5Log
MIC
s ke
toco
nazo
le (
mg
litre
-1)
r = 82P < 0.0001
r = 71P < 0.0001
r = 64P = 0.003
r = 0.04P = 0.87
Figure 2. Correlation between sensitivity of 20 field isolates of Mycosphaerella graminicola to the azole fungicides cyproconazole, ketoconazole,and tebuconazole, and the Qo-inhibitor fungicide kresoxim-methyl.
in mycelium was limited to isolates M3, M7 and S190.The basal expression level of MgAtr5 significantlydiffered between yeast-like cells and mycelium forall the isolates tested.
Treatment with cyproconazole strongly inducedtranscription of MgAtr4 in most of the isolates tested,especially in yeast-like cells. Induced levels of MgAtr1and MgAtr5 were also observed but only for a limitednumber of isolates such as M3 and M7 for MgAtr1 and
100
10
1
01
MIC
cyp
roco
nazo
le (
mg
litre
-1)
Progenies of cross M1 × M3 (n = 40)
Figure 3. Sensitivity of 40 progeny isolates of Mycosphaerellagraminicola obtained from a cross between isolates M1 (MIC0.178 mg litre−1) and M3 (MIC 17.8 mg litre−1) to cyproconazole.Median value is indicated as a continuous black line and mean valueas a dashed line. Sensitivity of the parent isolates M1 and M3 areindicated as dotted and dash-dotted lines, respectively.
M1 for MgAtr5, in both yeast-like cells and mycelium.Differences in induced level of expression among theisolates were also detected. For example, inductionof MgAtr4 in mycelium was much higher in isolatesM3 and M8 than in isolates M6, M7 and S190.Nevertheless, a correlation between expression of aspecific ABC gene and sensitivity of the isolates tocyproconazole was not clear.
CYP51 displayed basal transcripts in all isolates.However, in isolate M3 a relatively high basal level ofexpression was observed, especially in mycelium of thefungus. Treatment with cyproconazole did not effectthe expression level of this gene (Fig 4).
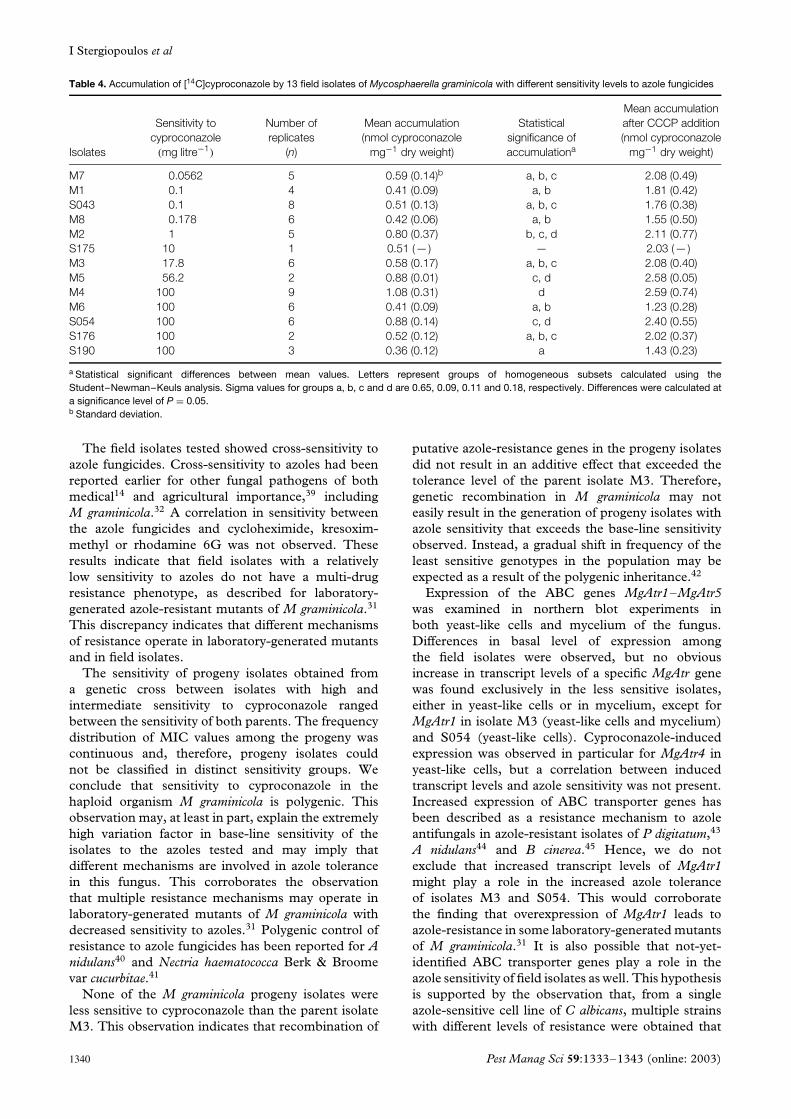
3.5 Accumulation studiesAccumulation of [14C]cyproconazole in thirteen fieldisolates tested proved to be constant in time. Meanaccumulation levels measured ranged between 0.35and 1 nmol cyproconazole mg−1 dry weight (Table 4).Addition of CCCP induced higher cyproconazoleaccumulation levels in all isolates, indicating thataccumulation was energy-dependent. No positive cor-relation between accumulation of [14C]cyproconazoleand sensitivity to cyproconazole (r = 0.39, P = 0.19),ketoconazole (r = 0.29, P = 0.34), or tebuconazole(r = 0.31, P = 0.31) was present. Statistical anal-ysis for significant differences in cyproconazoleaccumulation identified four overlapping groups ofisolates. Each group consisted of isolates with low
1338 Pest Manag Sci 59:1333–1343 (online: 2003)
Variation in base-line sensitivity to azole fungicides
Figure 4. Northern blot analysis of the ABC transporter genes MgAtr1, MgAtr4, and MgAtr5 and CYP51 in yeast-like cells and mycelium ofMycosphaerella graminicola after treatment with cyproconazole (C). Control samples are treated with 0.1% methanol (M). Isolates tested had low(M1, M7, M8, S043), medium (M3) and high (M4, M6, S054, S190) MIC values of cyproconazole. Hybridisation of the blots with an 18S rRNA probefrom Aspergillus niger served as a loading control.
and high sensitivity to azoles (Table 4). IsolatesM4 and S190, both with a low sensitivity tocyproconazole (MIC 100 mg litre−1) displayed thehighest (1.08 nmol mg−1 dry weight) and lowest(0.36 nmol mg−1 dry weight) accumulation level ofcyproconazole as compared to the other isolates tested(Table 4; Fig 5).
2.0
1.8
1.6
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0.00 5 15 25 35 45 55 65
Acc
umul
atio
n(n
mol
cyp
roco
nazo
le m
g dr
y w
eigh
t-1)
Time (min)
Figure 5. Accumulation of [14C]cyproconazole by field isolates ofMycosphaerella graminicola with different levels of sensitivity to azolefungicides. MIC values of cyproconazole for isolates (�) M4, (�) M7,(♦) S043 and (ž) S190 are 100, 0.0562, 0.1 and 100 mg litre−1,respectively.
4 DISCUSSIONIn this study, we set out to investigate the physiologicalbasis of variation in sensitivity to azole fungicides infield isolates of M graminicola. The results obtainedindicate that the field isolates of M graminicola testedvary significantly in sensitivity to azole fungicides.The variation factor for sensitivity of the isolatesfor tebuconazole was 178. This value is in thesame range as described by Suty and Kuck,34 whoreported a variation factor of 100 in a collection ofmore than 1500 isolates. The variation factors forthe azole fungicides cyproconazole and ketoconazoleamounted to 1779 and this value is significantly higherthan for tebuconazole. Such a broad variation insensitivity for these compounds is intriguing, especiallysince the isolates were previously never exposed toketoconazole, an azole antimycotic used in treatmentsof human fungal pathogens, or to cyproconazole,which was not used in the fields from which theisolates were collected. Significant variation in base-line sensitivity to fungicides has been reported beforeand is ascribed to naturally tolerant genotypes thatalready exist in fungal populations prior to fungicidetreatment.38 Studies monitoring the sensitivity of fieldpopulations of M graminicola to azoles over recentdecades have not yet detected the evolution of resistantphenotypes.32 Hence, the significant difference insensitivity to the azoles tested is regarded as naturalvariation in base-line sensitivity to azole fungicides.
Pest Manag Sci 59:1333–1343 (online: 2003) 1339
I Stergiopoulos et al
Table 4. Accumulation of [14C]cyproconazole by 13 field isolates of Mycosphaerella graminicola with different sensitivity levels to azole fungicides
Isolates
Sensitivity tocyproconazole(mg litre−1)
Number ofreplicates
(n)
Mean accumulation(nmol cyproconazole
mg−1 dry weight)
Statisticalsignificance ofaccumulationa
Mean accumulationafter CCCP addition(nmol cyproconazole
mg−1 dry weight)
M7 0.0562 5 0.59 (0.14)b a, b, c 2.08 (0.49)M1 0.1 4 0.41 (0.09) a, b 1.81 (0.42)S043 0.1 8 0.51 (0.13) a, b, c 1.76 (0.38)M8 0.178 6 0.42 (0.06) a, b 1.55 (0.50)M2 1 5 0.80 (0.37) b, c, d 2.11 (0.77)S175 10 1 0.51 (—) — 2.03 (—)M3 17.8 6 0.58 (0.17) a, b, c 2.08 (0.40)M5 56.2 2 0.88 (0.01) c, d 2.58 (0.05)M4 100 9 1.08 (0.31) d 2.59 (0.74)M6 100 6 0.41 (0.09) a, b 1.23 (0.28)S054 100 6 0.88 (0.14) c, d 2.40 (0.55)S176 100 2 0.52 (0.12) a, b, c 2.02 (0.37)S190 100 3 0.36 (0.12) a 1.43 (0.23)
a Statistical significant differences between mean values. Letters represent groups of homogeneous subsets calculated using theStudent–Newman–Keuls analysis. Sigma values for groups a, b, c and d are 0.65, 0.09, 0.11 and 0.18, respectively. Differences were calculated ata significance level of P = 0.05.b Standard deviation.
The field isolates tested showed cross-sensitivity toazole fungicides. Cross-sensitivity to azoles had beenreported earlier for other fungal pathogens of bothmedical14 and agricultural importance,39 includingM graminicola.32 A correlation in sensitivity betweenthe azole fungicides and cycloheximide, kresoxim-methyl or rhodamine 6G was not observed. Theseresults indicate that field isolates with a relativelylow sensitivity to azoles do not have a multi-drugresistance phenotype, as described for laboratory-generated azole-resistant mutants of M graminicola.31
This discrepancy indicates that different mechanismsof resistance operate in laboratory-generated mutantsand in field isolates.
The sensitivity of progeny isolates obtained froma genetic cross between isolates with high andintermediate sensitivity to cyproconazole rangedbetween the sensitivity of both parents. The frequencydistribution of MIC values among the progeny wascontinuous and, therefore, progeny isolates couldnot be classified in distinct sensitivity groups. Weconclude that sensitivity to cyproconazole in thehaploid organism M graminicola is polygenic. Thisobservation may, at least in part, explain the extremelyhigh variation factor in base-line sensitivity of theisolates to the azoles tested and may imply thatdifferent mechanisms are involved in azole tolerancein this fungus. This corroborates the observationthat multiple resistance mechanisms may operate inlaboratory-generated mutants of M graminicola withdecreased sensitivity to azoles.31 Polygenic control ofresistance to azole fungicides has been reported for Anidulans40 and Nectria haematococca Berk & Broomevar cucurbitae.41
None of the M graminicola progeny isolates wereless sensitive to cyproconazole than the parent isolateM3. This observation indicates that recombination of
putative azole-resistance genes in the progeny isolatesdid not result in an additive effect that exceeded thetolerance level of the parent isolate M3. Therefore,genetic recombination in M graminicola may noteasily result in the generation of progeny isolates withazole sensitivity that exceeds the base-line sensitivityobserved. Instead, a gradual shift in frequency of theleast sensitive genotypes in the population may beexpected as a result of the polygenic inheritance.42
Expression of the ABC genes MgAtr1–MgAtr5was examined in northern blot experiments inboth yeast-like cells and mycelium of the fungus.Differences in basal level of expression amongthe field isolates were observed, but no obviousincrease in transcript levels of a specific MgAtr genewas found exclusively in the less sensitive isolates,either in yeast-like cells or in mycelium, except forMgAtr1 in isolate M3 (yeast-like cells and mycelium)and S054 (yeast-like cells). Cyproconazole-inducedexpression was observed in particular for MgAtr4 inyeast-like cells, but a correlation between inducedtranscript levels and azole sensitivity was not present.Increased expression of ABC transporter genes hasbeen described as a resistance mechanism to azoleantifungals in azole-resistant isolates of P digitatum,43
A nidulans44 and B cinerea.45 Hence, we do notexclude that increased transcript levels of MgAtr1might play a role in the increased azole toleranceof isolates M3 and S054. This would corroboratethe finding that overexpression of MgAtr1 leads toazole-resistance in some laboratory-generated mutantsof M graminicola.31 It is also possible that not-yet-identified ABC transporter genes play a role in theazole sensitivity of field isolates as well. This hypothesisis supported by the observation that, from a singleazole-sensitive cell line of C albicans, multiple strainswith different levels of resistance were obtained that
1340 Pest Manag Sci 59:1333–1343 (online: 2003)

Variation in base-line sensitivity to azole fungicides
displayed distinct over-expression patterns of at leastfour genes involved in azole resistance. These includedthe ABC transporter genes CDR1 and CDR2, themajor facilitator gene MDR1, and ERG11, the geneencoding the target enzyme of the azoles in theergosterol biosynthetic pathway.46 The differentialexpression pattern of MgAtr1–MgAtr5 in the twodimorphic states of the fungus, yeast-like cells andmycelium, has been described previously,47,48 andmay reflect differences in cell wall and membranecomposition that can influence ABC transporteractivity.49–51
The expression studies demonstrate a high basaltranscript level of CYP51 in isolate M3. This isolatehas an intermediate sensitivity to azoles as compared toother isolates tested. Increased expression of CYP51is described as a mechanism of resistance to azolesin many fungi.52 In C albicans and S cerevisiae,azole tolerance as a result of enhanced transcriptionor amplification of the CYP51 (ERG11) gene hasalso been observed.13,14,16,17 In P digitatum and Vinaequalis, tandem repeats in the promoter region ofCYP51 enhanced transcription of this gene and thecopy number of these repeats correlated positivelywith CYP51 expression and tolerance to azoles.22,23
It is possible that a similar mechanism is operating inisolate M3.
Point mutations in the open reading frame of theCYP51 gene of filamentous fungi and yeasts can alsoconfer resistance to azoles.12 However, the sequenceof the CYP51 gene from laboratory-generated azole-resistant mutants of M graminicola and their parentalstrains was the same and did not show any ofthe point mutations involved in resistance to azoleantifungal agents in C albicans,12 laboratory mutantsof U maydis18 and P italicum,19 and in field isolatesof U necator20 and E graminis f sp hordei.21,31 Allelicvariations in the CYP51 sequence of field isolates ofM graminicola, make it difficult to assess whether thebase-line sensitivity is affected by point mutations.
Uptake experiments demonstrated that accumula-tion levels of [14C]cyproconazole by field isolates ofM graminicola did not correlate with sensitivity toazoles. Individual isolates with decreased sensitiv-ity to azoles, such as S190, showed a relatively lowaccumulation level of [14C]cyproconazole, but otherisolates with decreased sensitivity to azoles, such asM4, display a relatively high accumulation level ofthe fungicide. Hence, reduced accumulation of azolesfrom mycelium cannot account exclusively for the dif-ferences in base-line sensitivity in the isolates tested.The absence of a correlation between accumulation ofcyproconazole and azole sensitivity may reflect the highgenetic diversity of M graminicola field populations53
that accounts for morphological and physiological dif-ferences. These differences can have a significant effecton accumulation of [14C]cyproconazole and may maskthe variations in accumulation mediated by changesin ABC transporter activity. Studies on laboratory-generated azole-resistant mutants of M graminicola
showed that mutants occur with either a decreased oran increased accumulation of azoles as compared totheir parent isolates.31 Such a relation cannot easily bemade for field isolates, since the genetic relationshipbetween field isolates tested is virtually unknown. Forthese reasons it is still possible that the mechanism ofresistance in the least-sensitive field isolates is medi-ated by changes in ABC transporter activity, resultingin either a decreased or increased accumulation of thefungicide. Active drug disposal in vacuoles or in lipidbodies could possibly explain the relatively high accu-mulation level observed for some of the least sensitivefield isolates and might relate to overexpression ofABC transporters localised in vacuolar membranes.54
This has been described for azole-resistant strains of Umaydis that accumulated triadimenol to higher levelsas compared to sensitive strains.55 Alternative mecha-nisms of resistance, such as alterations in the target-siteof azoles can also not be excluded. For these reasonswe anticipate that a high correlation between azolesensitivity and accumulation in field isolates of Mgraminicola can not easily be established. However,previous literature data suggest that such a correla-tion is present.56,57 The discrepancy with the work ofJoseph-Horne et al56 can be ascribed to the low num-ber of isolates tested. The work of Gisi et al57 is difficultto judge since data that support their conclusion werenot published.
In summary, our studies suggest that variation insensitivity to azoles in field isolates of M graminicola isbased on multiple mechanisms. These may includedecreased accumulation of the fungicide in thecytoplasm, increased accumulation of the fungicide asa result of sequestration of the fungicide in cellularcompartments and overproduction of P45014DM.These multiple mechanisms would corroborate thepolygenic nature of azole sensitivity as shown by theanalysis of a genetic cross of M graminicola isolates.
ACKNOWLEDGEMENTSWe kindly acknowledge Dr JM Seng (Biotransfer,Montreuil Cedex, France) and Dr A Suty (Bayer AG,Landwirtschaftszentrum Monheim, Leverkusen, Ger-many) for providing the field isolates of Mycosphaerellagraminicola and ECP Verstappen for technical assis-tance with the crosses of M graminicola. We are gratefulto Professor Dr PJGM De Wit (WUR) and Dr L-HZwiers for critical reading of the manuscript and theWageningen Mycosphaerella group for key discussionsand suggestions during this work. Ioannis Stergiopou-los was financially supported by the Training andMobility of Researchers (TMR) Programme—MarieCurie Research Training Grants, The European Com-mission (Contract No ERBFMBICT983558).
REFERENCES1 Kuck KH and Scheinpflug H, Biology of sterol-biosynthesis
inhibiting fungicides, in Chemistry of plant protection, ed byHaug G and Hoffman H, Springer-Verlag, Berlin (1986).
Pest Manag Sci 59:1333–1343 (online: 2003) 1341

I Stergiopoulos et al
2 Sisler HD and Ragsdale NN, Biochemical and cellular aspectsof the antifungal action of ergosterol biosynthesis inhibitors,in Proc Symposium British Mycological Society, CambridgeUniversity Press, Cambridge, UK, pp 257–282 (1984).
3 De Waard MA and Fuchs A, Resistance to ergosterol biosyn-thesis inhibitors, II. Genetic and physiological aspects, inFungicide resistance in crop protection, ed by Dekker J andGeorgopoulos SG, Pudoc, Wageningen, The Netherlands,pp 87–100 (1982).
4 Fuchs A and De Waard MA, Resistance to ergosterol biosyn-thesis inhibitors, I. Chemistry and phenomenological aspects,in Fungicide resistance in crop protection, ed by Dekker J andGeorgopoulos SG, Pudoc, Wageningen, The Netherlands,pp 71–86 (1982).
5 Dekker J, Impact of fungicide resistance of disease controltoxicity, in Proc Brit Crop Prot Conf—Pests and Diseases, BCPC,Farnham, Surrey, UK, pp 857–863 (1981).
6 De Waard MA, Resistance to fungicides which inhibit sterol14α-demethylation, an historical perspective, in Fungicideresistance, ed by Heaney S, Slawson D, Hollomon DW,Smith M, Russell PE and Parry DW, BCPC, Farnham,Surrey, UK, pp 3–10 (1994).
7 Schepers HTAM, Fitness of isolates of Sphaerotheca fuliginearesistant or sensitive to fungicides which inhibit ergosterolbiosynthesis. Neth J Pl Path 91:65–76 (1985).
8 Eckert JK, Penicillium digitatum biotypes with reduced sensitivityto imazalil. Phytopathology 77:1728 (1987).
9 Heaney SP, Population dynamics of DMI fungicide sensitivitychanges in barley powdery mildew, in Fungicide resistance inNorth America, ed by Delp CJ, APS Press, St Paul, Minnesota,pp 89–92 (1988).
10 Hildebrand PD, Lockhart CL, Newberry RJ and Ross RG,Resistance of Venturia inaequalis to bitertanol and otherdemethylation-inhibiting fungicides. Canad J Plant Pathol10:311–316 (1988).
11 Kendall SJ, Hollomon DW, Cooke LR and Jones DR, Changesin sensitivity to DMI fungicides in Rhynchosporium secalis.Crop Prot 12:357–362 (1993).
12 Marichal P, Koymans L, Willemsens S, Bellens D, Verhas-selt P, Luyten W, Borgers M, Ramaekers FC, Odds FCand Vanden Bossche H, Contribution of mutations inthe cytochrome P450 14α-demethylase (Erg11p, Cyp51p)to azole resistance in Candida albicans. Microbiology145:2701–2713 (1999).
13 Vanden Bossche H, Marichal P and Odds FC, Molecularmechanisms of drug resistance in fungi. Trends Microbiol2:393–400 (1994).
14 White T, Increased mRNA levels of ERG16, CDR, and MDR1correlate with increases in azole resistance in Candida albicansisolates from a patient infected with human immunodeficiencyvirus. Antimicrob Agents Chemoth 41:1482–1487 (1997).
15 Franz R, Kelly SL, Lamb DC, Kelly DE, Ruhnke M andMorschhauser J, Multiple molecular mechanisms contributeto a stepwise development of fluconazole resistance inclinical Candida albicans strains. Antimicrob Agents Chemother42:3065–3072 (1998).
16 Kontoyiannis DP, Sagar N and Hirschi KD, Overexpressionof Erg11p by the regulatable GAL1 promoter confersfluconazole resistance in Saccharomyces cerevisiae. AntimicrobAgents Chemother 43:2798–2800 (1999).
17 Perea S, Lopez-Ribot JL, Kirkpatrick WR, McAtee RK, Santil-lan RA, Martinez M, Calabrese D, Sanglard D and Patter-son TF, Prevalence of molecular mechanisms of resistanceto azole antifungal agents in Candida albicans strains dis-playing high-level fluconazole resistance isolated from humanimmunodeficiency virus-infected patients. Antimicrob AgentsChemother 45:2676–2684 (2001).
18 Joseph-Horne T, Manning NJ, Hollomon D and Kelly SL,Defective sterol delta 5(6)desaturase as a cause of azoleresistance in Ustilago maydis. FEMS Microbiol Let: 1–2 (1995).
19 Van Nistelrooy JGM, Vanden Brink JM, Van Kan JA, VanGorcom RFM and De Waard MA, Isolation and molec-ular characterization of the gene encoding eburicol 14α-demethylase (CYP51) from Penicillium italicum. Mol Gen Genet250:725–733 (1996).
20 Delye C, Laigret F and Corio-Costet MF, A mutation in the14α-demethylase gene of Uncinula necator that correlates withresistance to a sterol biosynthesis inhibitor. Appl EnvironMicrobiol 63:2966–2970 (1997).
21 Delye C, Bousset L and Corio-Costet MF, PCR cloning anddetection of point mutations in the eburicol 14α-demethylase(CYP51) gene from Erysiphe graminis f sp hordei, a ‘recalcitrant’fungus. Curr Genet 34:399–403 (1998).
22 Hamamoto H, Hasegawa K, Nakaune R, Lee YJ, Makizumi Y,Akutsu K and Hibi T, Tandem repeat of a transcriptionalenhancer upstream of the sterol 14α-demethylase gene(CYP51) in Penicillium digitatum. Appl Environ Microbiol66:3421–3426 (2000).
23 Schnabel G and Jones AL, The 14α-demethylase (CYP51A1)gene is overexpressed in Venturia inaequalis strains resistant tomyclobutanil. Phytopathology 91:102–110 (2001).
24 De Waard MA and Van Nistelrooy JGM, Mechanism ofresistance to fenarimol in Aspergillus nidulans. Pestic BiochemPhysiol 10:219–229 (1979).
25 De Waard MA and Van Nistelrooy JGM, An energy-dependentefflux mechanism for fenarimol a fungicide in a wild-typestrain and fenarimol-resistant mutants of Aspergillus nidulans.Pestic Biochem Physiol 13:255–266 (1980).
26 Del Sorbo G, Andrade AC, Van Nistelrooy JG, Van Kan JA,Balzi E and De Waard MA, Multidrug resistance in Aspergillusnidulans involves novel ATP-binding cassette transporters.Mol Gen Genet 254:417–426 (1997).
27 Andrade AC, Del Sorbo G, Van Nistelrooy JGM and DeWaard MA, The ABC transporter AtrB from Aspergillus nidu-lans mediates resistance to all major classes of fungicides andsome natural toxic compounds. Microbiol UK 146:1987–1997(2000).
28 Higgins CF, ABC transporters: from microorganisms to man.Annu Rev Cell Biol 8:67–113 (1992).
29 Stergiopoulos I, Zwiers L-H and De Waard MA, Secretion ofnatural and synthetic toxic compounds from filamentous fungiby membrane transporters of the ATP-binding cassette andMajor facilitator superfamily. Eur J Plant Pathol 108:719–734(2002).
30 Gottesman MM and Pastan I, Biochemistry of multidrugresistance mediated by the multidrug transporter. Annu RevBiochem 62:385–427 (1993).
31 Zwiers L-H, Stergiopoulos I, Van Nistelrooy JGM and DeWaard MA, ABC transporters and azole susceptibility inlaboratory strains of the wheat pathogen Mycosphaerellagraminicola. Antimicrob Agents Chemother 46:3900–3906(2002).
32 Gisi U, Hermann D, Ohl L and Steden C, Sensitivity profilesof Mycosphaerella graminicola and Phytophthora infestanspopulations to different classes of fungicides. Pestic Sci51:290–298 (1997).
33 Gisi U and Hermann D, Sensitivity behaviour of Septoria triticipopulations on wheat to cyproconazole, in Fungicide resistance,ed by Heaney S, Slawsun D, Hollomon DW, Sunith M,Russell PE and Parry DW, BCPC, Farnham, Surrey, UK,pp 11–18 (1994).
34 Suty A and Kuck KH, Sensitivity of wheat leaf spot Septoriatritici to tebuconazole, in Proc Brit Crop Prot Conf—Pests andDiseases, BCPC, Farnham, Surrey, UK, pp 689–694 (1996).
35 Kema GHJ, Verstappen ECP, Todorova M and Waalwijk C,Successful crosses and molecular tetrad and progeny analysesdemonstrate heterothallism in Mycosphaerella graminicola.Curr Genet 30:251–258 (1996).
36 Pijls CFN, Shaw MW and Parker A, A rapid test to evaluatein vitro sensitivity of Septoria tritici to flutriafol, using amicrotitre plate reader. Plant Pathol 43:726–732 (1994).
37 Kema GHJ and Van Silfhout CH, Genetic variation for viru-lence and resistance in the wheat-Mycosphaerella graminicola
1342 Pest Manag Sci 59:1333–1343 (online: 2003)

Variation in base-line sensitivity to azole fungicides
pathosystem. III. Comparative seedling and adult plant exper-iments. Phytopathology 87:266–272 (1997).
38 Brent KJ, Monitoring fungicide resistance: purposes, proceduresand progress, in Resistance ‘91: Achievements and developmentsin combating pesticide resistance, ed by Denholm I, Devon-shire AL and Hollomon DW, Elsevier Science Publishers,London, pp 1–18 (1992).
39 Georgopoulos SG, Cross-resistance, in Fungicide resistance incrop protection, ed by Dekker J and Georgopoulos SG, Pudoc,Wageningen, The Netherlands, pp 53–59 (1982).
40 Van Tuyl JM, Genetics of fungal resistance to systemic fungicides,Mededelingen Landbouwhogeschool Wageningen, Veen-man H and Zonen BV, Wageningen, The Netherlands,pp 65–99 (1977).
41 Kalamarakis AE, De Waard MA, Ziogas BN and Georgopou-los SG, Resistance to fenarimol in Nectria haematococca varcucurbitae. Pestic Biochem Physiol 40:212–220 (1991).
42 Russell PE, Fungicide resistance: occurrence and management.J Agric Sci 124:317–323 (1995).
43 Nakaune R, Adachi K, Nawata O, Tomiyama M, Akutsu K andHibi T, A novel ATP-binding cassette transporter involvedin multidrug resistance in the phytopathogenic fungusPenicillium digitatum. Appl Environ Microbiol 64:3983–3988(1998).
44 Andrade AC, Van Nistelrooy JGM, Peery RB, Skatrud PL andDe Waard MA, The role of ABC transporters from Aspergillusnidulans in protection against cytotoxic agents and in antibioticproduction. Mol Gen Genet 263:966–977 (2000).
45 Hayashi K, Schoonbeek H, Sugiura H and De Waard MA,Multidrug resistance in Botrytis cinerea associated withdecreased accumulation of the azole fungicide oxpoconazoleand increased transcription of the ABC transporter geneBcatrD. Pestic Biochem Physiol 70:168–179 (2001).
46 Cowen LE, Sanglard D, Calabrese D, Sirjusingh C, Ander-son JB and Kohn LM, Evolution of drug resistance inexperimental populations of Candida albicans. J Bacteriol182:1515–1522 (2000).
47 Zwiers L-H and De Waard MA, Characterization of theABC transporter genes MgAtr1 and MgAtr2 from the
wheat pathogen Mycosphaerella graminicola. Fund Genet Biol30:115–125 (1998).
48 Stergiopoulos I, Gielkens MM, Goodall SD, Venema K and DeWaard MA, Molecular cloning and characterisation of threenew ATP-binding cassette transporter genes from the wheatpathogen Mycosphaerella graminicola. Gene 289:141–149(2002).
49 Hitchcock CA, Barrett-Bee KJ and Russell NJ, The lipidcomposition of azole-sensitive and azole-resistant strains ofCandida albicans. J Gen Microbiol 132:2421–2431 (1986).
50 Sinicrope FA, Dudeja PK, Bissonnette BM, Safa AR and Bra-situs TA, Modulation of P-glycoprotein-mediated drug trans-port by alterations in lipid fluidity of rat liver canalicularmembrane vesicles. J Biol Chem 267:24 995–25 002 (1992).
51 Van den Hazel HB, Pichler H, do Valle Matta MA, Leitner E,Goffeau A and Daum G, PDR16 and PDR17, two homolo-gous genes of Saccharomyces cerevisiae, affect lipid biosynthesisand resistance to multiple drugs. J Biol Chem 274:1934–1941(1999).
52 Joseph-Horne T and Hollomon DW, Molecular mechanism ofazole resistance in fungi. FEMS Microbiol Let 149:141–149(1997).
53 Chen RS and McDonald BA, Sexual reproduction plays a majorrole in the genetic structure of populations of the fungusMycosphaerella graminicola. Genet 142:1119–1127 (1996).
54 Rea PA, MRP subfamily ABC transporters from plants andyeast. J Exper Bot 50(Special Iss):895–913 (1999).
55 Wellmann H and Schauz K, DMI-resistance in Ustilago maydis:II. Effect of triadimefon on regenerating protoplasts andanalysis of fungicide uptake. Pestic Biochem Physiol 46:55–64(1993).
56 Joseph-Horne T, Hollomon D, Manning N and Kelly SL,Investigation of the sterol composition and azole resistancein field isolates of Septoria tritici. Appl Environ Microbiol62:184–190 (1996).
57 Gisi U, Chin KM, Knapova G, Farber RK, Mohr U, Parisi S,Sierotzki H and Steinfeld U, Recent developments in elu-cidating modes of resistance to phenylamide, DMI andstrobilurin fungicides. Crop Prot 19:863–872 (2000).
Pest Manag Sci 59:1333–1343 (online: 2003) 1343
Top Related
Copyright © 2022 FDOKUMEN

