






Bahasa
Halaman
Hukum


RESEARCH PAPER
Morphological and genetic distinctiveness of metallicolousand non-metallicolous populations of Armeria maritima s.l.(Plumbaginaceae) in PolandA. Abratowska1*, P. Wasowicz1,2*, P. T. Bednarek3, J. Telka1 & M. Wierzbicka1
1 Department of Molecular Plant Physiology, Institute of Botany, Faculty of Biology, University of Warsaw, Warsaw, Poland
2 Department of Biology, Faculty of Biology and Environmental Sciences, Cardinal Stefan Wyszynski University in Warsaw, Warsaw, Poland
3 Institute of Plant Breeding & Acclimatization – National Research Institute, Radzikow, Błonie, Poland
INTRODUCTION
Soils enriched in heavy metals are among the most inhospita-ble environments to both plant and animal life. Such inor-ganic pollution is known to affect organisms on every level ofbiological organisation, leading to toxicity from molecular(Przedpełska-Wasowicz & Wierzbicka 2010) to ecosystem(Banasova et al. 2006) scale. There are, however, plant speciesthat are able to grow on soils containing heavy metal concen-trations that are lethal to many organisms. These plants arewell adapted to extremely polluted soil and are known asmetallophytes, which are of great interest in themselves(Baker & Brooks 1989; Baker & Walker 1990; Ernst 1990;Punz 1995) and also due to their potential use in phytoreme-diation (Raskin et al. 1997; Salt et al. 1998; Meagher 2000;Ernst 2005; Sheoran et al. 2011).
Sites heavily polluted with zinc, lead, cadmium or otherhighly toxic metals are found all over the world. In Europe,their distribution is highly disjunct and associated with for-mer industrial activities, such as mining and smelting. TheOlkusz Ore-bearing Region (southern Poland) is among themost important and most frequently studied metalliferoussites in Central Europe. Historical evidence indicates that sil-
ver has been mined and smelted in the region for more than700 years, while zinc and lead mining started here in 18thcentury (Grodzinska & Szarek-Łukaszewska 2002). Theseindustrial activities have left behind hectares of polluted soilsthat have been colonised by plants and other organisms forcenturies. One such is a metalliferous waste heap in Bolesław,which established 130 years ago (Szarek-Łukaszewska &Niklinska 2002). Moreover, the whole region, and the oldwaste heap in particular, can be considered an ideal place tostudy plant adaptation and microevolution to heavy metals(Wierzbicka & Panufnik 1998; Grodzinska & Szarek-Łukas-zewska 2002; Wierzbicka & Rostanski 2002; Załecka & Wie-rzbicka 2002; Wierzbicka & Pielichowska 2004; Przedpełska &Wierzbicka 2007; Olko et al. 2008), which have been shownto be rapid at both molecular and phenotypic levels in suchpolluted environments (Bone & Farres 2001).
Armeria maritima is widespread across metalliferous sitesin the Olkusz Ore-bearing Region (Dobrzanska 1955;Grodzinska & Szarek-Łukaszewska 2002; Nowak et al. 2011).It is well known that plants in local populations within theregion differ morphologically from those growing on non-metalliferous soils elsewhere in Poland. Szafer (1946) sug-gested that A. maritima accessions from the region should be
Keywords
DNA methylation; epigenetics; heavy metals;
metAFLP; metallophyte; microevolution.
Correspondence
A. Abratowska, Department of Molecular
Plant Physiology, Institute of Botany, Faculty
of Biology, University of Warsaw,
Miecznikowa Str. 1, 02-096 Warsaw, Poland.
E-mail: [email protected]
Editor
M. Hawkesford
*These authors contributed equally to this
work.
Received: 11 March 2011; Accepted: 13
October 2011
doi:10.1111/j.1438-8677.2011.00536.x
ABSTRACT
Patterns of morphological, genetic and epigenetic variation (DNA methylation pat-tern) were investigated in metallicolous (M) and non-metallicolous (NM) popula-tions of Armeria maritima. A morphological study was carried out using plantsfrom six natural populations grown in a greenhouse. Morphological variation wasassessed using seven traits. On the basis of this study, three representative popula-tions were selected for molecular analyses using metAFLP to study sequence- andmethylation-based DNA variation. Only one morphological trait (length of outerinvolucral bracts) was common to both metallicolous populations studied; however,the level of variation was sufficient to differentiate between M and NM populations.Molecular analyses showed the existence of naturally occurring epigenetic variationin A. maritima populations, as well as structuring into distinct between and withinpopulation components. We show that patterns of population genetic structure dif-fered depending on the information used in the study. Analysis of sequence-basedinformation data demonstrates the presence of three well-defined and geneticallydifferentiated populations. Methylation-based data show that two major groups ofindividuals are present, corresponding to the division into M and NM populations.These results were confirmed using different analytical approaches, which suggestthat the DNA methylation pattern is similar in both M populations. We hypothesisethat epigenetic processes may be involved in microevolution leading to developmentof M populations in A. maritima.
Plant Biology ISSN 1435-8603
Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands 1

classified as A. maritima (Mill.) Willd. subsp. halleri (Wallr.)A. Love & D. Love, because of their morphological similarityto plants from metalliferous sites in the Harz (Germany).This classification was widely accepted (Szafer 1946; Do-brzanska 1955; Pawłowski 1963; Pinto da Silva 1972; Lefebvre1974; Lefebvre & Kakes 1981; Szafer et al. 1986; Rutkowski1998; Mirek et al. 2002), although it has recently been chal-lenged (Wierzbicka & Słysz 2005).
Taxonomic status of different A. maritima populationsscattered across metalliferous sites in Europe has been a hot-spot of botanical research. In general, the European taxa ofthe A. maritima complex are poorly distinguishable morpho-logically due to large phenotypic plasticity and intermediatecharacter of many populations (Szafer 1946; Pawłowski 1963;Pinto da Silva 1972; Woodel & Dale 1993), and thus taxon-omy of A. maritima based on morphology alone can be mis-leading. Classification of metallicolous populations ofA. maritima in Europe has been based on morphologicaltraits (Lefebvre 1974; Richards et al. 1989), esterases (Lefeb-vre & Kakes 1981), flavonoid profiles (Lauranson et al. 1995),DNA content and pollen size (Vekemans et al. 1996), allo-zymes (Vekemans & Lefebvre 1997), cytogenetic characters(Coulaud et al. 1999), and molecular markers such as RAPD(Baumbach & Hellwig 2003) and AFLP (Baumbach 2005;Baumbach & Hellwig 2007). These studies showed that thesuggestion of a single subspecies growing on metalliferoussoils (A. maritima subsp. halleri s.l.) should be rejected dueto the lack of consistency of characteristics throughout thegeographical range and close genetic distance to particularmetallicolous (halleri, bottendorfensis, hornburgensis, ‘eifeliaca’,‘calaminaria’) and non-metallicolous (elongata) populations(Baumbach & Hellwig 2007). According to the latest studies,metallicolous populations of A. maritima in Central Europeshould not be classified as distinct subspecies or a complextaxon A. maritima subsp. halleri s.l., but as edaphic varietiesof A. maritima subsp. elongata (Baumbach & Hellwig 2007).
We undertook research on metallicolous and non-metallicolouspopulations of A. maritima in Poland in order to distinguishplants in metalliferous sites within the Olkusz Ore-bearingRegion. Apart from comparison of morphological traits ofplants from two metalliferous sites (Bolesław, Poland, and azinc–lead slag heap in Plombieres, Belgium) and four non-metallicolous sites in Poland, we adopted a genetic approachusing amplified fragment length polymorphisms (AFLP)(Vos et al. 1995). AFLP have been widely used to assessgenetic variability in the plant kingdom (Ronikier 2002;Rouf-Mian et al. 2005; Wassom & Tranel 2005), althoughthe technique has several shortcomings connected with thepredominantly nuclear origin of AFLP, and problems withhomoplasy remain a major issue in analysis and interpreta-tion of AFLP data (Meudt & Clarke 2007). The AFLP modi-fication applied here (metAFLP), employing enzymeisoschizomers differing in their recognition of C-methyla-tion, is capable of detecting variation in both the DNAsequence and its methylation pattern (Bednarek et al. 2007).Using metAFLP, we attempted to clarify whether changes inDNA methylation contribute to the distinction between me-tallicolous and non-metallicolous populations of A. mariti-ma. Recent studies have shown that epigenetic modificationstriggered by heavy metals may result in changes in geneexpression, thus leading to a modification of the phenotype
(Aina et al. 2004; Lukens & Zhan 2007; Vandegehuchte &Janssen 2011). Epigenetic changes have been also hypothes-ised to influence microevolutionary processes in populationson contaminated sites (Aina et al. 2004; Lukens & Zhan2007; Vandegehuchte & Janssen 2011).
We addressed the following questions: (i) are A. maritimaplants from zinc–lead waste heaps in Bolesław morphologicallyand genetically related to plants from zinc–lead slag heap inPlombieres or to non-metallophyte plants from unpollutedsites in Poland; (ii) is the DNA sequence polymorphism andDNA methylation pattern equally informative in genetic struc-ture of metallicolous (M) and non-metallicolous (NM) popu-lations; and (iii) does DNA methylation play a role in theemergence of variation between populations of A. maritima?
MATERIAL AND METHODS
Study species
Armeria maritima (Mill.) Willd. (Plumbaginaceae) is a com-mon herb throughout Europe in a wide variety of habitats,from coastal sandy soils to saline and metalliferous sites(Pinto da Silva 1972; Woodel & Dale 1993). Within the spe-cies, two types of population exist: metallicolous populations(M) on heavy metal-enriched metalliferous soils, and non-metallicolous populations (NM) on non-metalliferous soils.The species is highly variable morphologically and thusnumerous intraspecific classifications have been elaboratedfor the taxon. In most of Europe it is an obligatory outbreed-ing species with a dimorphic incompatibility system, but thiscan break down, leading to the emergence of facultative self-fertile plants (Baker 1966; Lefebvre 1973, 1976; Kornas &Medwecka-Kornas 2002), as found in some European Mpopulations (Lefebvre 1972, 1976; Richards et al. 1989; Veke-mans et al. 1992 after Lauranson et al. 1995) and influencingtheir genetic variability.
Plant sampling
Six A. maritima populations were sampled: two metallicolouspopulations in Poland and Belgium (labelled M1 and M2,respectively) and four non-metallicolous populations fromPoland (labelled NM1 to NM4). Detailed information onpopulations sampled is given in Table 1.
In Bolesław (population M1), where the 130-year-old wasteheap is located, pollution can be regarded as extreme, withheavy metal concentrations in the substrate reaching2000 mgÆkg)1 DW for lead, 150 mgÆkg)1 DW for cadmiumand 30,000–50,000 mgÆkg)1 DW for zinc. Substrate pH was7.6 (Szarek-Łukaszewska & Niklinska 2002). The populationin Plombieres (M2) is in an area containing major zinc andlead ores (La Calamine region, Belgium), where processing ofthese ores has resulted in formation of large slagheaps (Swen-nen et al. 1994). Within this region, we sampled a zinc–leadslagheap having population of A. maritima (Duvigneaud &Saintenoy-Simon 1998). Concentrations of heavy metals andpH are very variable within Plombieres heap (Auquier & deLeval 1974; Simon 1978; Ernst & Nelissen 2000; Lock et al.2003), and the mosaic character of the site (different struc-ture and chemical composition of the substrate) influencesthe vitality of plants. Chemical analyses showed that our
Metallicolous and non-metallicolous populations of A. maritima Abratowska, Wasowicz, Bednarek, Telka & Wierzbicka
2 Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands

plants were on soil with total concentration of heavy metalsreaching 7000–13,000 mgÆkg)1 DW lead, 16–28 mgÆkg)1 DWcadmium and 16,000–34,000 mgÆkg)1 DW zinc; substrate pHwas about 6.0.
Seeds of A. maritima were collected in bulk from severalhundred specimens in each of the six study populations.
Plant cultivation
Seeds were sown in garden soil and germinated in a phyto-tron chamber, with a photoperiod of 16-h light ⁄ 8-h dark at20 ± 1 �C. Seedlings were transferred to single pots of gardensoil and cultivated in a greenhouse with the same photope-riod at 25 ± 5 �C for 12 months.
Morphological analyses
Morphological measurements were performed on plants ofthe same biological age, determined by emergence of the firstinflorescence-bearing stem. The following traits were mea-sured on 50 plants from each population: leaf length andwidth, number of leaves in the rosette, length of stalks, inflo-rescence diameter, bud length and outer involucral bractlength. The general appearance of the plant and the flowercolour were also recorded.
For each quantitative trait, the mean and standard devia-tion, as well as the median value and 5–95 percentile rangewere calculated. The Kruskal–Wallis test was used to test forsignificant differences between populations (a = 0.05). Tovisualise the degree of morphological similarity ⁄ dissimilaritybetween individuals from sampled populations, a multidi-mensional approach was employed. Principal componentsanalysis (PCA) using the covariance matrix was performed inStatistica version 9 (Statsoft Inc., Tulsa, USA) and resultswere visualised using Sigma Plot 9.0 graphing software(Systat Software Inc., Chicago, IL, USA).
Molecular analyses
Leaves were collected from 48 4-month-old plants cultivatedin the greenhouse. Leaves were dried in silica gel and storedfrozen. Genomic DNA was extracted using the Plant DNeasy
Mini Kit (Qiagen Venlo, Netherlands), and quantified spec-trophotometrically. DNA integrity and purity were verifiedusing 1% agarose gel electrophoresis in TBE buffer. Agarosegels were stained with ethidium bromide (0.1 lgÆml)1).
The metAFLP procedure followed that described in Bedn-arek et al. (2007). Samples of equal DNA content (0.5 lg)were digested with Acc65I ⁄ MseI. Simultaneously, 0.5 lg DNAfrom the same sample was treated with KpnI ⁄ MseI. Acc65Iand KpnI are isoschizomers with respect to recognition of C-methylation. Acc65I is insensitive to dam methylation but itsactivity is blocked by both dcm and CpG methylation. KpnIis insensitive to all forms of methylation. After ligation ofadaptors (Table 2), samples underwent pre-selective andselective amplification. For the selective PCR, we used oneprimer labelled with 32P at the 5¢-end. For the selectiveamplification, seven selective primer combinations were used:CpG-GCA ⁄ M-CAG, CpG-GGC ⁄ M-CTC, CpG-GTG ⁄ M-CAT,CpG-GTT ⁄ M-CCC, CpXpG-AGA ⁄ M-CAC, CpXpG-AGC ⁄ M-CTA and CpXpG-TGC ⁄ M-CCA. The selective primersequences used are listed in Table 2. Thermal profiles fol-lowed Vos et al. (1995). The products of selective amplifica-tion were separated on 7% denaturing polyacrylamide gels,and the signal was visualised by autoradiography.
AFLP patterns (Acc65I ⁄ MseI and KpnI ⁄ MseI separately)were compiled into 0 ⁄ 1 matrices, where ‘0’ is absence and‘1’ is presence of a band. Monomorphic bands wereremoved from the analyses. We obtained two data matricesfor methylation-insensitive (KpnI ⁄ MseI) and methylation-sensitive (Acc65I ⁄ MseI) enzyme combinations. Parameters ofgenetic variation were calculated for both matrices: thenumber of polymorphic loci (#Ploc), proportion of poly-morphic loci (%PLP), Nei’s gene diversity (Hj) and theShannon information index (I) were calculated for eachpopulation as a measure of within-population diversity.Nei’s genetic distance was used to estimate genetic distancebetween populations (Lynch & Milligan 1994). Estimates of#Ploc, %PLP, Hj and Nei’s genetic distance between popula-tions were made using AFLP-SURV 1.0 (Vekemans et al.2002). The Shannon information index was calculated asI = )R[pi log2(pi)], where pi is the relative frequency of theith AFLP fragment in a population, using popgene 1.32(Yeh & Boyle 1997).
Table 1. Locations of populations sampled in the present study.
population location habitat
GPS coordinates
NiN E
M1 Bolesław
S Poland
Zinc–lead waste heap;
calamine community
50�17¢31.63¢¢ 19�28¢24.71¢¢ �500
M2 Plombieres
E Belgium
Zinc–lead slag heap; 50�44¢02.40¢¢ 05�58¢10.67¢¢ �2000
NM1 Manasterz
SE Poland
Dry meadow 50�07¢34.53¢¢ 22�40¢12.35¢¢ <500
NM2 Kamienczyk
Central Poland
Dry meadow 52�35¢36.44¢¢ 21�33¢42.78¢¢ >5000
NM3 Osowiec
NE Poland
Semi-dry meadow 53�29¢25.35¢¢ 22�38¢05.49¢¢ �2000
NM4 Bablin
W Poland
Semi-dry meadow 52�40¢30.60¢¢ 16�43¢08.48¢¢ �500
M = metallicolous population; NM = non-metallicolous population; Ni = estimated population size.
Abratowska, Wasowicz, Bednarek, Telka & Wierzbicka Metallicolous and non-metallicolous populations of A. maritima
Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands 3

Patterns of genetic structure were investigated on both datamatrices obtained using methylation-insensitive (KpnI ⁄ MseI)and methylation-sensitive (Acc65I ⁄ MseI) enzyme combina-tion. The neighbour-joining (NJ) trees were constructed forall sampled individuals using Treecon 1.3b (Van de Peer &De Wachter 1994) on the basis of Nei & Li (1979) geneticdistances. Estimation of the robustness of the branches wasdone using 1000 bootstrap replicates.
Genetic structure was investigated using a Bayesianapproach with structure 2.2.3 (Pritchard et al. 2000), theno admixture model and uncorrelated allele frequencies. Sim-
ulations were run using a burn-in length and MCMC(Markov Chain Monte Carlo) of 150,000 and 300,000,respectively. The range of possible Ks was tested from 1 to10. Each simulation was run 10 times in order to quantifythe amount of variation of the likelihood for each K. Theuppermost hierarchical level of the genetic structure was esti-mated using the ad hoc statistic DK (Evanno et al. 2005).Computations were made with the BioPortal project (Kumaret al. 2009).
Analysis of molecular variance was performed with Arle-
quin 3.5 (Excoffier & Lischer 2010). The statistical signifi-cance of the respective fixation indices (FST, FSC and FCT)was tested with 10,000 permutations (Excoffier et al. 1992).
RESULTS
Morphological analysis
We investigated morphological characters in plants cultivatedin a greenhouse. Generally, plants flowered after 120 days(4 months) of cultivation, and reproduced vegetativelythrough the formation of new ‘daughter’ rosettes. Plants fromM and NM populations differed with respect to their generalappearance. Plants from the M2 population formed twice asmany ‘daughter’ rosettes as plants from the other popula-tions. Plants from both M populations were smaller thanthose from NM populations. Flower colour varied frombright to dark pink, although darker flowers were more fre-quent in the M2 population, while pink and bright pink flow-ers were more frequent in the remaining five populations.
Plants in all the six populations showed large variabilityfor almost all measured morphological traits (Table 3). Thehigh degree of morphological variability generally preventeddifferentiation between M and NM populations. The range ofvariation for most of the investigated morphological charac-ters was highly overlapping, as evidenced in the 5–95 percen-tile ranges (Table 3). However, the length of the outerinvolucral bracts was an exception that enabled clear differen-tiation between M and NM populations. In both M populations,
Table 2. Adaptor and primer sequences used in the study.
adaptors ⁄ primers sequence 5¢ fi 3¢
Acc65I adaptor I CTC GTA GCA TGC GTA CA
Acc65I adaptor II GTA CTG TAC GCA TGC TAC
KpnI adaptor I CTC GTA GCA TGC GTA CAG TAC
KpnI adaptor II ACT CGT ACG ACT GT
MseI adaptor I TAC TCA GGA CTC ATC
MseI adaptor II GAG TCC TGA GTA GCA G
Acc65I ⁄ KpnI pre-selective primer GCA TGC GTA CAG TAC C
MseI pre-selective primer GAT GAG TCC TGA GTA AC
CpG-GCA CAT GCG TAC AGT ACC GCA
CpG-GGC CAT GCG TAC AGT ACC GGC
CpG-GTG CAT GCG TAC AGT ACC GTG
CpG-GTT CAT GCG TAC AGT ACC GTT
CpXpG-AGA CAT GCG TAC AGT ACC AGA
CpXpG-AGC CAT GCG TAC AGT ACC AGC
CpXpG-TGC CAT GCG TAC AGT ACC TGC
M-CAC GAT GAG TCC TGA GTA AC A C
M-CAG GAT GAG TCC TGA GTA AC A G
M-CAT GAT GAG TCC TGA GTA AC A T
M-CCA GAT GAG TCC TGA GTA AC C A
M-CCC GAT GAG TCC TGA GTA AC C C
M-CTA GAT GAG TCC TGA GTA AC T A
M-CTC GAT GAG TCC TGA GTA AC T C
Table 3. Variability of seven morphological traits of A. maritima plants in the studied populations. Median (bold) and mean values (normal font) are given;
5–95 percentile range is given in parenthesis (n = 50).
population
leaf length
(mm)
leaf width
(mm)
number of
leaves
length of
stalk (cm)
diameter of
inflorescence (mm)
length of
inflorescence
bud (mm)
length of outer
involucral bracts
(mm)
M1 84.5b
85.1 (70.0–100.0)
4.0a
4.1 (3.4–4.8)
85.0b
87.7 (38.0–140.0)
27.6b
28.2 (17.0–38.8)
23.0b
23.2 (21.0–26.0)
6.0a,b
5.5 (4.0–7.0)
6.5a
6.2 (5.0–7.0)
M2 66.5a
66.0 (59.0–74.0)
3.3b
3.3 (2.6–4.0)
186.0a
192.6 (120.0–270.0)
18.5a
18.7 (11.4–26.4)
20.0a
19.2 (13.0–23.0)
6.0a
6.0 (5.0–7.0)
6.0a
6.7 (4.0–9.0)
NM1 98.5c
96.4 (80.0–114.0)
2.1c
2.2 (1.7–3.0)
100.0b
105.5 (60.0–160.0)
31.0b
31.3 (20.0–40.0)
24.0b
24.2 (21.0–27.0)
5.0b
5.1 (4.0–6.0)
10.0b
11.0 (8.0–16.0)
NM2 100.0c
98.8 (83.0–118.0)
3.3b
3.3 (2.7–4.0)
74.5c
73.7 (50.0–99.0)
38.5c
38.5 (27.0–51.0)
25.0b
24.3 (22.0–27.0)
6.0a
5.6 (4.0–8.0)
10.0b
11.3 (8.0–16.0)
NM3 89.0b
86.3 (61.0–105.0)
2.0c
2.0 (1.6–2.4)
96.5b
97.7 (61.0–144.0)
30.0b
30.0 (25.0–38.0)
26.0c
26.2 (24.0–28.0)
5.0b
5.2 (4.0–6.0)
10.0b
10.9 (8.0–15.0)
NM4 125.5d
123.0 (90.0–160.0)
3.3b
3.3 (2.6–4.1)
75.0c
75.7 (49.0–110.0)
35.5b,c
34.9 (18.0–50.0)
25.5c
25.7 (24.0–27.0)
5.0b
5.4 (4.0–7.0)
10.0b
10.5 (7.0–15.0)
M = metallicolous population; NM = non-metallicolous population.
Letters in superscript denote significant differences as assessed using the Kruskal–Wallis test (a = 0.05).
Metallicolous and non-metallicolous populations of A. maritima Abratowska, Wasowicz, Bednarek, Telka & Wierzbicka
4 Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands

plants had significantly shorter (P < 0.05, Kruskal–Wallistest) outer involucral bracts of the inflorescence bud com-pared with plants from NM populations (Table 3). The outerbracts of the inflorescence bud in plants from M populationswere blunt-ended and shorter than the bud, whereas in NMpopulations the outer bracts were sharp-ended and twice aslong as the bud.
The PCA on the data matrix containing measurementsfrom morphological analyses showed clear morphological dif-ferentiation between the analysed populations. Population M2
was clearly separated from the remaining five populations(Fig. 1). Non-metallicolous populations (NM1–NM4),although slightly differentiated morphologically, formed awell-delineated group. Population M1 exhibited intermediatecharacteristics between the complex of NM populations andpopulation M1 (Fig. 1), as also confirmed with canonical dis-criminant analysis (data not shown). One population fromthe NM group (NM1) as well as both M populations wereselected to further analysis. Thus individuals representingalmost the whole spectrum of edaphic and morphologicalvariation were included in genetic and epigenetic analyses.
Molecular analyses
Robust profiles were obtained for both methylation-insensi-tive (KpnI ⁄ MseI) and methylation-sensitive (Acc65I ⁄ MseI)enzyme combinations. One primer combination produced30–80 fragments in each population. In total, 502 unambigu-ously scored fragments were found in the 136 individualsanalysed for both enzyme combinations. Application of themethylation-insensitive combination (KpnI ⁄ MseI) gave 211segregating fragments (#loc), while the methylation-sensitiveenzymes (Acc65I ⁄ MseI) gave 219 fragments (Table 4).
The number of polymorphic loci (#Ploc) was highest inpopulation M1, regardless of the enzyme combination used(Table 4). Similar variability was observed in population M2,while the lowest number of polymorphic loci was found inpopulation NM1. The proportion of polymorphic loci
(%PLP) followed the same pattern of variation, and rangedbetween 84.8 in population M1 to 37.4 in population NM1
(Table 4) for the methylation-insensitive enzyme combina-tion. Application of the methylation-sensitive combinationdid not affect the pattern of variation. Nei’s gene diversityindex (Hj), calculated on the basis of data from the methyla-tion-insensitive enzyme combination, varied from 0.2922 inpopulation M1 to 0.1810 in population NM1. For the methyl-ation-sensitive enzyme, the index followed the same patternof variation (Table 4). The Shannon information index (I)calculated for the methylation-insensitive enzyme combina-tion varied from 0.3601 in population M2 to 0.2107 in popu-lation NM1; the index calculated for data acquired using themethylation-sensitive combination varied from 0.3453 inpopulation M1 to 0.2282 in NM1 (Table 4).
Nei’s genetic distance between investigated populationswas calculated on the basis of results obtained for bothenzyme combinations. The methylation-insensitive enzymecombination (KpnI ⁄ MseI) yielded almost equal distancesbetween populations, while the methylation-sensitive enzymecombination (Acc65I ⁄ MseI) led to substantial decrease indistances between M populations and distances between Mand NM populations were even larger (Table 5).
The unrooted NJ tree constructed with data from themethylation-insensitive enzyme combination revealed theexistence of three well separated groups of individuals corre-sponding to the three analysed populations (M1, M2 andNM1), with almost equal length of the branches (Fig. 2a).The NJ from the methylation-sensitive enzyme combination
Fig. 1. Results of PCA showing morphological differentiation between
investigated populations of A. maritima. Percentage of the total variance
explained by each component is given in parentheses. M, metallicolous
population; NM, non-metallicolous population.
Table 4. Genetic variation in investigated populations of A. maritima cal-
culated using data matrices from methylation-insensitive (KpnI ⁄ MseI) and
methylation-sensitive (Acc65I ⁄ MseI) enzyme combination.
population n #loc #Ploc %PLP Hj SE I
KpnI ⁄ MseI
Bolesław (M1) 44 211 179 84.8 0.2922 0.0122 0.3601
Plombieres (M2) 47 211 175 82.9 0.2658 0.0122 0.3273
Manasterz (NM1) 45 211 79 37.4 0.1810 0.0129 0.2107
Acc65I ⁄ MseI
Bolesław (M1) 44 219 193 88.1 0.2851 0.0115 0.3453
Plombieres (M2) 47 219 180 82.2 0.2648 0.0124 0.3182
Manasterz (NM1) 45 219 93 42.5 0.1991 0.0127 0.2282
n = number of analysed individuals; #loc = number of loci; #Ploc = num-
ber of polymorphic loci at the 5% level; %PLP = proportion of polymor-
phic loci at the 5% level; Hj = expected heterozygosity (Nei’s gene
diversity); SE = standard error of Hj; I = Shannon information index;
M = metallicolous population; NM = non-metallicolous population.
Table 5. Nei’s genetic distance between populations of A. maritima
based on data matrix from the analysis using methylation-insensitive
(KpnI ⁄ MseI – above diagonal) and methylation-sensitive (Acc65I ⁄ MseI –
below diagonal) enzyme combination.
Bolesław (M1) Plombieres (M2) Manasterz (NM1)
Bolesław (M1) – 0.2205 0.2720
Plombieres (M2) 0.1913 – 0.2798
Manasterz (NM1) 0.3534 0.3280 –
M = metallicolous population; NM = non-metallicolous population.
Abratowska, Wasowicz, Bednarek, Telka & Wierzbicka Metallicolous and non-metallicolous populations of A. maritima
Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands 5
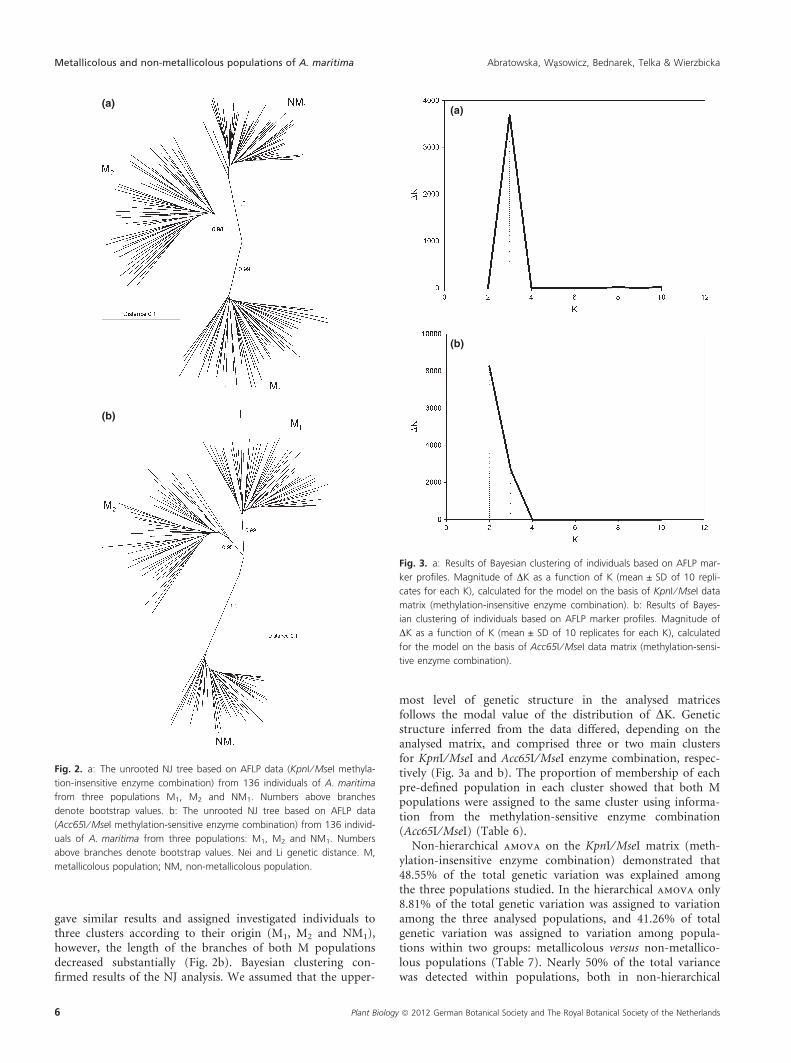
gave similar results and assigned investigated individuals tothree clusters according to their origin (M1, M2 and NM1),however, the length of the branches of both M populationsdecreased substantially (Fig. 2b). Bayesian clustering con-firmed results of the NJ analysis. We assumed that the upper-
most level of genetic structure in the analysed matricesfollows the modal value of the distribution of DK. Geneticstructure inferred from the data differed, depending on theanalysed matrix, and comprised three or two main clustersfor KpnI ⁄ MseI and Acc65I ⁄ MseI enzyme combination, respec-tively (Fig. 3a and b). The proportion of membership of eachpre-defined population in each cluster showed that both Mpopulations were assigned to the same cluster using informa-tion from the methylation-sensitive enzyme combination(Acc65I ⁄ MseI) (Table 6).
Non-hierarchical amova on the KpnI ⁄ MseI matrix (meth-ylation-insensitive enzyme combination) demonstrated that48.55% of the total genetic variation was explained amongthe three populations studied. In the hierarchical amova only8.81% of the total genetic variation was assigned to variationamong the three analysed populations, and 41.26% of totalgenetic variation was assigned to variation among popula-tions within two groups: metallicolous versus non-metallico-lous populations (Table 7). Nearly 50% of the total variancewas detected within populations, both in non-hierarchical
(a)
(b)
Fig. 2. a: The unrooted NJ tree based on AFLP data (KpnI ⁄ MseI methyla-
tion-insensitive enzyme combination) from 136 individuals of A. maritima
from three populations M1, M2 and NM1. Numbers above branches
denote bootstrap values. b: The unrooted NJ tree based on AFLP data
(Acc65I ⁄ MseI methylation-sensitive enzyme combination) from 136 individ-
uals of A. maritima from three populations: M1, M2 and NM1. Numbers
above branches denote bootstrap values. Nei and Li genetic distance. M,
metallicolous population; NM, non-metallicolous population.
(a)
(b)
Fig. 3. a: Results of Bayesian clustering of individuals based on AFLP mar-
ker profiles. Magnitude of DK as a function of K (mean ± SD of 10 repli-
cates for each K), calculated for the model on the basis of KpnI ⁄ MseI data
matrix (methylation-insensitive enzyme combination). b: Results of Bayes-
ian clustering of individuals based on AFLP marker profiles. Magnitude of
DK as a function of K (mean ± SD of 10 replicates for each K), calculated
for the model on the basis of Acc65I ⁄ MseI data matrix (methylation-sensi-
tive enzyme combination).
Metallicolous and non-metallicolous populations of A. maritima Abratowska, Wasowicz, Bednarek, Telka & Wierzbicka
6 Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands

and hierarchical analysis. Non-hierarchical amova for theAcc65I ⁄ MseI matrix (methylation-sensitive enzyme combina-tion) explained 50.32% of the total genetic variation amongthe three populations. Hierarchical amova assigned 22.21%of total genetic variance to that among two groups (metallic-olous versus non-metallicolous populations), and 31.81%among populations within these two groups. Nearly 50% ofthe total variance was due to variance within populations inboth non-hierarchical and hierarchical analysis (Table 7).Application of the methylation-sensitive enzyme combinationled to a 2.5-fold increase in the percentage of total geneticvariation explained by variation among metallicolous andnon-metallicolous populations.
DISCUSSION
Taxonomy of Armeria maritima on metalliferous sites in Eur-ope has been discussed repeatedly (Lefebvre 1972, 1974; Lef-ebvre & Kakes 1981; Richards et al. 1989; Vekemans et al.1992, 1996; Lauranson et al. 1995; Vekemans & Lefebvre1997; Coulaud et al. 1999; Brown 2001; Baumbach & Hellwig2003, 2007; Baumbach 2005). Populations of A. maritima onmetal-enriched soils were previously classified into five differ-ent microendemic subspecies: A. maritima subsp. halleri(Wallr.) A. & D. Love, A. maritima subsp. calaminaria (F.Petri) W. Ernst, A. maritima subsp. eifeliaca (F. Petri) Lefeb-vre, A. maritima subsp. hornburgensis (A. Schulz) Rothm.,and A. maritima subsp. bottendorfensis (A. Schulz) Rothm.According to this taxonomic treatment, plants growing onheavy metal-enriched sites in the Olkusz region (populationM1) would be classified as A. maritima subsp. halleri (Szafer1946; Dobrzanska 1955; Pawłowski 1963; Pinto da Silva 1972;Lefebvre 1974; Lefebvre & Kakes 1981; Szafer et al. 1986; Rut-kowski 1998; Mirek et al. 2002). Lefebvre (1974) suggestedthat taxonomic classification of the above listed microendem-ic subspecies is far from satisfactory due to difficulties result-ing from large phenotypic variation within its geographicrange in Europe. However, it was demonstrated that a set ofmorphological traits, or at least its frequency within a givenpopulation, could be used to differentiate between plantsfrom M and NM sites (Lefebvre 1974). On the basis of ourmorphological studies, we concluded that only one morpho-logical trait (length of outer involucral bracts) was commonto both metallicolous populations studied. The remainingtraits were highly variable within and among populations.However, the level of ‘informative’ morphological polymor-phism was still large enough to differentiate between M andNM populations, as evidenced in multidimensional statisticalanalyses (PCA and CDA). Therefore, morphological similari-ties between M populations, although slight, do exist even inplants grown under uniform greenhouse conditions. We
Table 6. Proportion of membership of each pre-defined population in
each of the clusters identified by Bayesian clustering and obtained after
analysis of the data matrix from methylation-insensitive (KpnI ⁄ MseI) and
methylation-sensitive (Acc65I ⁄ MseI) enzyme combination.
KpnI ⁄ MseI
population
inferred clusters
number of individuals1 2 3
Bolesław (M1) 1.0 0 0 47
Plombieres (M2) 0 1.0 0 44
Manasterz (NM1) 0 0 1.0 45
Acc65I ⁄ MseI
population
inferred clusters
number of individuals1 2
Bolesław (M1) 1.0 0 47
Plombieres (M2) 1.0 0 44
Manasterz (NM1) 0 1.0 45
M = metallicolous population; NM = non-metallicolous population.
Table 7. Analysis of molecular variance (AMOVA) for investigated populations of A. maritima using methylation-insensitive (KpnI ⁄ MseI) and methylation-sen-
sitive (Acc65I ⁄ MseI) enzyme combination. Presented statistics are: degrees of freedom (df), sum of squares (SS), variance components (VC), percentage of
total variation explained by each component (%V). Significance levels are based on 10,000 permutations.
source of variation df SS VC %V P
KpnI ⁄ MseI
all populations Among populations 2 1881.312 20.28320 48.55 **
Within populations 133 2859.357 21.49893 51.45 **
Total 135 4740.669 41.78213
two groups: (M) versus (NM) Among groups 1 1052.479 3.79305 8.81 **
Among populations within groups 1 828.833 17.76291 41.26 **
Within populations 133 2859.357 21.49893 49.93 **
Total 135 4740.669 43.05489
Acc65I ⁄ MseI
all populations Among populations 2 2077.971 22.43859 50.32 **
Within populations 133 2946.515 22.15425 49.68 **
Total 135 5024.485 44.59283
two groups: (M) versus (NM) Among groups 1 1359.207 10.70336 22.21 **
Among populations within groups 1 718.763 15.32675 31.81 **
Within populations 133 2946.515 22.15425 45.98 **
Total 135 5024.485 48.18436
M = metallicolous population; NM = non-metallicolous population.
**P < 0.01 (ARLEQUIN 3.5).
Abratowska, Wasowicz, Bednarek, Telka & Wierzbicka Metallicolous and non-metallicolous populations of A. maritima
Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands 7

expect that similarities could be even stronger in plants col-lected in the field. This is the probable reason why plantsfrom metallicolous population in Poland were previouslyclassified as A. maritima subsp. halleri.
The use of molecular markers cast new light on taxonomyof metallicolous populations of A. maritima in Europe. Veke-mans & Lefebvre (1997), using indirect genetic estimates,showed that metal-tolerant populations of A. maritimaevolved independently, and that the pattern of gene flowbetween populations is consistent with a model of isolationby distance, with considerable gene flow between neighbour-ing populations. These findings, showing no reduction ingene flow between geographically close tolerant and non-tolerant populations, as well as previous research on morpho-logical and reproductive differentiation between populationswithin A. maritima complex, questioned the relevance of thedescription of endemic metallicolous taxa. Genetic polymor-phism using RAPD (Baumbach & Hellwig 2003) and AFLP(Baumbach 2005; Baumbach & Hellwig 2007) approachesconfirmed these findings and suggested that isolation by dis-tance rather than substrate type may be of crucial importancein shaping population genetic structure within A. maritima.However, these previous studies focused only on sequence-based genetic variation in the context of the origin of metal-licolous populations within A. maritima. Our work aimed toinvestigate genetic differentiation between metallicolous andnon-metallicolous populations, as inferred from informationon both DNA sequence and methylation (epigenetic) pat-terns. From the sequence variation, we concluded that theinvestigated populations were equally distant genetically, asexpected from the hypothesis of independent, multipleevolutionary origins of metallicolous populations withinA. maritima. Interestingly, analysis of data acquired using amethylation-sensitive enzyme combination showed thatgenetic distance between metallicolous populations decreasedsubstantially, while the distance between M and NM popula-tions was even larger. These findings were additionally con-firmed using clustering methods (NJ and Bayesian approach)that showed the presence of two, well-defined groups of indi-viduals. Both methods showed that individuals from the twometallicolous populations, even though geographically distantand genetically (as far as DNA sequence is concerned) welldifferentiated, formed one well-supported group. To ourknowledge, this has never been reported previously. The pres-ence of stronger genetic structure between M and NM popu-lations obtained from the methylation-sensitive enzymecombination was additionally confirmed using amovas onboth data matrices (obtained with sequence- and methyla-tion-based information). The percentage of total genetic vari-ation explained by the variation among metallicolous andnon-metallicolous populations increased 2.5-fold after theapplication of the methylation-sensitive enzyme combination.
Although numerous studies have focused on intraspecificepigenetic variation in model and cultivated species (see e.g.Ashikawa 2001; Cervera et al. 2002; Keyte et al. 2006; Vaughnet al. 2007; Salmon et al. 2008), our knowledge on theamount and structuring of methylation-based DNA variationin natural plant populations is still very scarce (Herrera &Bazaga 2010). Recent studies have shown that naturallyoccurring, methylation-based epigenetic variation is struc-tured into distinct between- and within-population compo-
nents, analogous to sequence-based genetic variation (Herrera& Bazaga 2010), and our results confirm these findings. Wefound that a significant amount of epigenetic variation(about 50% according to amovas) can be attributed to varia-tion within populations. This extensive within-populationvariance has been suggested to be important in terms ofmicroevolutionary potential (Kalisz & Purugganan 2004). Wealso found significant epigenetic differentiation between Mand NM populations. The presence of multilocus epigeneticdifferentiation has only been reported for a very limitednumber of plant species (Li et al. 2008; Herrera & Bazaga2010).
It is widely accepted that DNA methylation, through con-trol of gene expression, can induce phenotypic alterations tomorphological (Manning et al. 2006; Marfil et al. 2009) andphysiological (Sha et al. 2005; Gimenez et al. 2006) traits. Itshould be stressed that both metallicolous populations, hav-ing similar patterns of epigenetic variation, also showed somesimilarities in morphological and physiological (metal toler-ance) characteristics. Although no simple connection betweenmethylation pattern and observed similarities can be madeon the basis of our study, we believe that this finding maypave the way for further research on the connection bet-ween epigenetic variation and the origin of metallicolouspopulations.
Changes in DNA methylation patterns are considered partof plant defence against heavy metal stress (Lukens & Zhan2007), and heavy metal-induced epigenetic changes in specificDNA sequences have been observed in Trifolium repens andCannabis sativa (Aina et al. 2004). It has been hypothesisedthat inheritance of heavy metal-induced epigenetic modifica-tions over generations may stimulate microevolutionary pro-cesses under stress conditions (Vandegehuchte & Janssen2011). Therefore, investigating patterns of DNA methylationin natural populations could be considered a first step inassessing the significance of epigenetic change in microevolu-tion (Kalisz & Purugganan 2004; Richards 2006). This taskhas been greatly facilitated by development of relatively sim-ple and inexpensive methods to study natural patterns ofgenome-wide DNA methylation in various organisms (e.g.Reyna-Lopez et al. 1997; Bednarek et al. 2007) We suggestthat it cannot be excluded that epigenetic processes may bepartly responsible for the observed differentiation betweenpopulations from metalliferous and non-metalliferous sites.
CONCLUSIONS
We found that morphology of plants from two heavy metal-polluted regions differed from those growing on unpollutedsoils. These differences, although slight, were sufficient to dis-criminate between the investigated M and NM populationsin Armeria maritima. Sequence-based genetic variationshowed that the M and NM populations were strongly differ-entiated genetically. We showed that the pattern of geneticstructure in the investigated populations changed consider-ably in relation to the pattern of DNA methylation. Weshowed the existence of naturally occurring epigenetic varia-tion in A. maritima populations and that epigenetic variationis structured into distinct between- and within-populationcomponents. We expect that further studies on epigeneticvariation in M and NM populations of A. maritima will cast
Metallicolous and non-metallicolous populations of A. maritima Abratowska, Wasowicz, Bednarek, Telka & Wierzbicka
8 Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands

a new light on our understanding of microevolutionary pro-cesses that lead to the development of metallicolous popula-tions. Our results may indicate a role for epigenetic changesin plant microevolution in response to heavy-metal pollution,although this requires further studies and evaluation. Theresults suggest the need for further taxonomic studies inorder to elucidate the status of A. maritima in the OlkuszOre-bearing Region.
ACKNOWLEDGEMENTS
We thank Dr. Damien Ertz (Curator de Plombieres, NationalBotanic Garden of Belgium, Meise) for facilitating collectionof plant samples from Plombieres, and Dr. Jean-FrancoisHermanns (Theux, Belgium) for valuable assistance. We are
grateful to colleagues from The Society Nouvelle-Montagne,ADL Plombieres and AG Bergbaufolgelandschaften e. V. forsharing their broad experience in the study of metallophytes.We also thank Prof. Krystyna Grodzinska, Dr. Gra_zynaSzarek-Łukaszewska (Institute of Botany, Polish Academy ofSciences, Cracow) and Prof. Adam Rostanski (Department ofPlant Systematics, University of Silesia, Katowice) for inspira-tion and long-term collaboration. We sincerely thank the edi-tor and two anonymous reviewers for insightful commentsthat considerably improved the paper.
The research was financially supported by a grant from theMinistry of Science and Higher Education of the Republic ofPoland (No. 2 P04C 036 28).
REFERENCES
Aina R., Sgorbati S., Santagostino A., Massimo L.,
Ghiani A., Citterio S. (2004) Specific hypomethyla-
tion of DNA is induced by heavy metals in white
clover and industrial hemp. Physiologia Plantarum,
121, 472–480.
Ashikawa I. (2001) Surveying CpG methylation at 5¢-CCGG in the genomes of rice cultivars. Plant
Molecular Biology, 45, 31–39.
Auquier P., de Leval J. (1974) Teneur en zinc, plomb,
cuivre et manganese, in situ et en culture experi-
mentale des Festuca des terrains calaminaires belges.
Bulletin de la Societe Royale de Botanique de Belgi-
que, 107, 375–386.
Baker H.G. (1966) The evolution, functioning and
breakdown of heteromorphic incompatibility sys-
tems. 1. The Plumbaginaceae. Evolution, 20, 349–
368.
Baker A.J.M., Brooks R.R. (1989) Terrestrial higher
plants which hyperaccumulate metallic elements –
a review of their distribution, ecology and phyto-
chemistry. Biorecovery, 1, 81–216.
Baker A.J.M., Walker P.L. (1990) Ecophysiology of
metal uptake by tolerant plants. In: Shaw J. (Ed.),
Heavy metal tolerance in plants: evolutionary aspects.
CRC Press, Boca Raton, FL, pp. 155–177.
Banasova V., Horak O., Ciamporova M., Nadubinska
M., Lichtscheidl I. (2006) The vegetation of metal-
liferous and non-metalliferous grasslands in two
former mine regions in Central Slovakia. Biologia,
61, 433–439.
Baumbach H. (2005) Genetische Differenzierung mit-
teleuropaischer Schwermetallsippen von Silene vul-
garis, Minuartia verna und Armeria maritima unter
Berucksichtigung biogeographischer, montanhistori-
scher und physiologischer Aspekte. PhD Thesis,
Dissertationes Botanicae 398, J. Cramer, Berlin-
Stuttgart.
Baumbach H., Hellwig F.H. (2003) Genetic variation
within and among metal-tolerant and non-tolerant
populations of Armeria maritima (Mill.) Willd. s.l.
(Plumbaginaceae) in Central and Northeast Ger-
many. Plant Biology, 5, 186–193.
Baumbach H., Hellwig F.H. (2007) Genetic differenti-
ation of metallicolous and non-metallicolous Arme-
ria maritima (Mill.) Willd. taxa (Plumbaginaceae)
in Central Europe. Plant Systematics and Evolution,
269, 245–258.
Bednarek P.T., Orłowska R., Koebner R.M.D., Zimny
J. (2007) Quantification of the tissue-culture
induced variation in barley (Hordeum vulgare L.).
BMC Plant Biology, 7, 10.
Bone E., Farres A. (2001) Trends and rates of micro-
evolution in plants. Genetica, 112, 165–182.
Brown G. (2001) The heavy-metal vegetation of
north-western mainland Europe. Botanische Jahrbu-
cher fur Systematik, Pflanzengeschichte und Pflanzen-
geographie, 123, 63–110.
Cervera M.T., Ruiz-Garcia L., Martinez-Zapater J.M.
(2002) Analysis of DNA methylation in Arabidopsis
thaliana based on methylation-sensitive AFLP
markers. Molecular Genetics and Genomics, 268,
543–552.
Coulaud J., Barghi N., Lefebvre C., Siljak-Yakovlev S.
(1999) Cytogenetic variation in populations of Ar-
meria maritima (Mill.) Willd. in relation to geo-
graphical distribution and soil stress tolerances.
Canadian Journal of Botany, 77, 673–685.
Dobrzanska J. (1955) Floristic-ecological studies on
calamine flora of Bolesław and Olkusz vicinities.
Acta Societatis Botanicorum Poloniae, 24, 357–500
(in Polish with English summary).
Duvigneaud J., Saintenoy-Simon J. (1998) Les haldes
calaminaires de Plombieres (province de Liege, Bel-
gique). Les Naturalistes Belges, 79, 24–32.
Ernst W.H.O. (1990) Mine vegetation in Europe. In:
Shaw J. (Ed.), Heavy metal tolerance in plants: evo-
lutionary aspects. CRC Press, Boca Raton, FL, pp.
21–38.
Ernst W.H.O. (2005) Phytoextraction of mine wastes
– options and impossibilities. Chemie der Erde –
Geochemistry, 65 (Suppl. 1), 29–42.
Ernst W.H.O., Nelissen H.J.M. (2000) Life-cycle
phases of a zinc- and cadmium-resistant ecotype
of Silene vulgaris in risk assessment of polymetal-
lic mine soils. Environmental Pollution, 107, 329–
338.
Evanno G., Regnaut S., Goudet J. (2005) Detecting
the number of clusters of individuals using the
software STRUCTURE: a simulation study. Molecu-
lar Ecology, 14, 2611–2620.
Excoffier L., Lischer H.E.L. (2010) Arlequin suite ver.
3.5: a new series of programs to perform popula-
tion genetics analyses under Linux and Windows.
Molecular Ecology Research, 10, 564–567.
Excoffier L., Smouse P.E., Quattro J.M. (1992) Analy-
sis of molecular variance inferred from metric dis-
tances among DNA haplotypes: application to
human mitochondrial DNA restriction data. Genet-
ics, 86, 991–1000.
Gimenez C., Palacios G., Colmenares A. (2006) Musa
methylated DNA sequences associated with toler-
ance to Mycosphaerella fijiensis toxins. Plant Molec-
ular Biology Reporter, 24, 33–43.
Grodzinska K., Szarek-Łukaszewska G. (2002) Zinc–
lead waste heaps in the vicinity of Olkusz – past,
present and future. Kosmos, 51, 127–138 (in Polish
with English summary).
Herrera C.M., Bazaga P. (2010) Epigenetic differentia-
tion and relationship to adaptive genetic divergence
in discrete populations of the violet Viola cazorlen-
sis. New Phytologist, 187, 867–876.
Kalisz S., Purugganan M.D. (2004) Epialleles via DNA
methylation: consequences for plant evolution.
Trends in Ecology and Evolution, 19, 309–314.
Keyte A.L., Percifield R., Liu B., Wendel J.F. (2006)
Intraspecific DNA methylation polymorphism in
cotton (Gossypium hirsutum L.). Journal of Heredity,
97, 444–450.
Kornas J., Medwecka-Kornas A. (2002) Geography
of plants. Polish Scientific Publishing, Warsaw (in
Polish).
Kumar S., Skjaeveland A., Orr R.J., Enger P., Ruden
T., Mevik B.H., Burki F., Botnen A., Shalchian-
Tabrizi K. (2009) AIR: a batch-oriented web pro-
gram package for construction of supermatrices
ready for phylogenomic analyses. BMC Bioinformatics,
10, 7.
Lauranson J., Vekemans X., Lefebvre C., Jay M.
(1995) Flavonoid profile variations in Armeria
maritima (Mill.) Willd. Biochemical Systematics and
Ecology, 23, 319–329.
Lefebvre C. (1972) Armeria sur une mine de cuivre
au Pays de Galles et sur une mine de zinc clans les
Pyrenees. Bulletin de la Societe Royale de Botanique
de Belgique, 105, 207–212.
Lefebvre C. (1973) Outbreeding and inbreeding in a
zinc–lead mine population of Armeria maritima.
Nature, 243, 96–97.
Lefebvre C. (1974) Population variation and taxon-
omy in Armeria maritima with special reference to
heavy-metal tolerant populations. New Phytologist,
73, 209–219.
Lefebvre C. (1976) Breeding system and population
structure of Armeria maritima (Mill.) Willd. on a
zinc–lead mine. New Phytologist, 77, 187–192.
Lefebvre C., Kakes P. (1981) Variation electrophore-
tique des esterases des feuilles d’ Armeria maritima
(Mill.) Willd: quelques aspects taxonomiques et
evolutifs. Bulletin de la Societe Royale de Botanique
de Belgique, 114, 31–40.
Li Y.D., Shan X.H., Liu X.M., Hu L.J., Guo W.L., Liu
B. (2008) Utility of the methylation-sensitive
amplified polymorphism (MSAP) marker for detec-
Abratowska, Wasowicz, Bednarek, Telka & Wierzbicka Metallicolous and non-metallicolous populations of A. maritima
Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands 9

tion of DNA methylation polymorphism and epige-
netic population structure in a wild barley species
(Hordeum brevisubulatum). Ecological Research, 23,
927–930.
Lock K., Janssens F., Janssen C.R. (2003) Effects of
metal contamination on the activity and diversity
of springtails in an ancient Pb–Zn mining area in
Plombieres, Belgium. European Journal of Soil
Biology, 39, 25–29.
Lukens L.N., Zhan S. (2007) The plant genome’s
methylation status and response to stress: implica-
tions for plant improvement. Current Opinion in
Plant Biology, 10, 317–322.
Lynch M., Milligan B. (1994) Analysis of population-
genetic structure using RAPD markers. Molecular
Ecology, 3, 91–99.
Manning K., Tor M., Poole M., Hong Y., Thompson
A.J., King G.J., Giovannoni J.J., Seymour G.B.
(2006) A naturally occurring epigenetic mutation
in a gene encoding an SBP-box transcription factor
inhibits tomato fruit ripening. Nature Genetics, 38,
948–952.
Marfil C.F., Camadro E.L., Masuelli R.W. (2009) Phe-
notypic instability and epigenetic variability in a
diploid potato of hybrid origin, Solanum ruiz-lealii.
BMC Plant Biology, 9, 21.
Meagher R.B. (2000) Phytoremediation of toxic ele-
mental and organic pollutants. Current Opinions in
Plant Biology, 3, 153–162.
Meudt H.M., Clarke A.C. (2007) Almost forgotten or
latest practice? AFLP applications, analyses and
advances Trends in Plant Science, 12, 106–117.
Mirek Z., Piekos-Mirkowa H., Zajac A., Zajac M.
(2002) Flowering plants and pteridophytes of Poland.
A checklist. Institute of Botany, Polish Academy of
Sciences, Cracow.
Nei M., Li W.H. (1979) Mathematical model for
studying genetic variation in terms of restriction
endonucleases. Proceedings of the National Academy
of Sciences of the United States of America, 76,
5269–5273.
Nowak T., Kapusta P., Jedrzejczyk-Korycinska M.,
Szarek-Łukaszewska G., Godzik B. (2011) The vascu-
lar plants of the Olkusz Ore-bearing region. Institute
of Botany, Polish Academy of Sciences, Cracow.
Olko A., Abratowska A., _Zyłkowska A., Wierzbicka
M., Tukiendorf A. (2008) Armeria maritima from a
calamine heap – initial studies on physiologic–met-
abolic adaptations to metal-enriched soil. Ecotoxi-
cology and Environmental Safety, 69, 209–218.
Pawłowski B. (Ed.) (1963) Flora of Poland. Vascular
plants of Poland and neighbouring territories, Vol.
X. Institute of Botany, Polish Academy of Sciences,
Cracow (in Polish).
Pinto da Silva A.R. (1972) Armeria Willd. In: Tutin
T.G., Heywood V.H., Burges N.A., Moore D.M., Val-
entine D.H., Walters S.M., Webb D.A. (Eds), Flora
Europaea. Diapensiaceae to Myoporaceae. Cambridge
University Press, Cambridge, pp. 29–38.
Pritchard J.K., Stephens M., Donnelly P. (2000) Infer-
ence of population structure using multilocus geno-
type data. Genetics, 155, 945–959.
Przedpełska E., Wierzbicka M. (2007) Arabidopsis
arenosa (Brassicaceae) from a lead–zinc waste heap
in southern Poland – a plant with high tolerance to
heavy metals. Plant and Soil, 299, 43–53.
Przedpełska-Wasowicz E., Wierzbicka M. (2010) Gat-
ing of aquaporins by heavy metals in Allium cepa
L. epidermal cells. Protoplasma, 207, 186–194.
Punz W. (1995) Metallophytes in the Eastern Alps
with special emphasis on higher plants growing on
calamine and copper localities. Phyton – Annales
Rei Botanicae, 35, 295–309.
Raskin I., Smith R.D., Salt D.E. (1997) Phytoremedia-
tion of metals: using plants to remove pollutants
from the environment. Current Opinion in Biotech-
nology, 8, 221–226.
Reyna-Lopez G.E., Simpson J., Ruiz-Herrera J. (1997)
Differences in DNA methylation patterns are
detectable during the dimorphic transition of fungi
by amplification of restriction polymorphisms.
Molecular and General Genetics, 253, 703–710.
Richards E.J. (2006) Inherited epigenetic variation –
revisiting soft inheritance. Nature Reviews Genetics,
7, 395–401.
Richards A.J., Lefebvre C., Macklin M.G., Nicholson
A., Vekemans X. (1989) The population genetics of
Armeria maritima (Mill.) Willd on the River South
Tyne, UK. New Phytologist, 112, 281–293.
Ronikier M. (2002) The use of AFLP markers in con-
servation genetics – a case study on Pulsatilla ver-
nalis in the Polish lowlands. Cellular and Molecular
Biology Letters, 7, 677–684.
Rouf-Mian M.A., Zwonitzer J.C., Chen Y., Saha M.C.,
Hopkins A.A. (2005) AFLP diversity within and
among hardinggrass populations. Crop Science, 45,
2591–2597.
Rutkowski L. (1998) Key for identification of vascular
plants in Polish Lowlands. Polish Scientific Pub-
lisher, Warsaw (in Polish).
Salmon A., Clotault J., Jenczewski E., Chable V.,
Manzanares-Dauleux M.J. (2008) Brassica oleracea
displays a high level of DNA methylation polymor-
phism. Plant Science, 174, 61–70.
Salt D.E., Smith R.D., Raskin I. (1998) Phytoremedia-
tion. Annual Review of Plant Physiology and Plant
Molecular Biology, 49, 643–668.
Sha A.H., Lin X.H., Huang J.B., Zhang D.P. (2005)
Analysis of DNA methylation related to rice adult
plant resistance to bacterial blight based on methyl-
ation-sensitive AFLP (MSAP) analysis. Molecular
Genetics and Genomics, 273, 484–490.
Sheoran V., Sheoran A.S., Poonia P. (2011) Role of
hyperaccumulators in phytoextraction of metals
from contaminated sites: a review. Critical Reviews
in Environmental Science and Technology, 41, 168–
214.
Simon E. (1978) Heavy metals in soils, vegetation
development and heavy metal tolerance in plant
populations from metalliferous areas. New Phytolo-
gist, 81, 175–188.
Swennen R., van Keer I., de Vos W. (1994) Heavy
metal contamination in overbank sediments of the
Geul River (East Belgium): its relation to former
Pb–Zn mining activities. Environmental Geology, 24,
12–21.
Szafer W. (1946) The genus Armeria Willd. in Poland
(La genre Armeria en Pologne). Acta Societatis
Botanicorum Poloniae, 17, 7–28.
Szafer W., Kulczynski S., Pawłowski B. (1986) Polish
plants. Polish Scientific Publisher, Warsaw (in
Polish).
Szarek-Łukaszewska G., Niklinska M. (2002) Concen-
tration of alkaline and heavy metals in Biscutella
laevigata L. and Plantago lanceolata L. growing on
calamine spoils (S. Poland). Acta Biologica Craco-
viensia. Series: Botanica, 44, 29–38.
Van de Peer Y., De Wachter Y. (1994) TREECON for
Windows: a software package for the construction
and drawing of evolutionary trees for the Microsoft
Windows environment. Computer Applications in
the Biosciences, 10, 569–570.
Vandegehuchte M.B., Janssen C.R. (2011) Epigenetics
and its implications for ecotoxicology. Ecotoxicology,
20, 607–624.
Vaughn M.W., Tanurdzic M., Lippman Z., Jiang H.,
Carrasquillo R., Rabinowicz P.D., Dedhia N.,
McCombie W.R., Agier N., Bulski A., Colot V.,
Doerge R.W., Martienssen R.A. (2007) Epigenetic
natural variation in Arabidopsis thaliana. PLoS
Biology, 5, 1617–1629.
Vekemans X., Lefebvre C. (1997) On the evolution of
heavy-metal tolerant populations in Armeria mariti-
ma: evidence from allozyme variation and repro-
ductive barriers. Journal of Evolutionary Biology, 10,
175–191.
Vekemans X., Lambert A., Lefebvre C. (1992) Isozyme
variation at the populational level in Armeria mari-
tima. Belgian Journal of Botany, 125, 270–275.
Vekemans X., Lefebvre C., Coulaud J., Blaise S.,
Gruber W., Siljak-Yakovlev S., Brown G. (1996)
Variation in nuclear DNA content at the species
level in Armeria maritima. Hereditas, 124, 237–
242.
Vekemans X., Beauwens T., Lemaire M., Roland-Ruiz
I. (2002) Data from amplified fragment length
polymorphism (AFLP) markers show indication of
size homoplasy and of a relationship between
degree of homoplasy and fragment size. Molecular
Ecology, 11, 139–151.
Vos P., Hogers R., Bleeker M., Reijans M., van de Lee
T., Hornes M., Frijters A., Pot J., Peleman J.,
Kuiper M., Zabeau M. (1995) AFLP: a new tech-
nique for DNA fingerprinting. Nucleic Acids
Research, 23, 4407–4414.
Wassom J.J., Tranel P.J. (2005) Amplified fragment
length polymorphism-based genetic relationships
among weedy Amaranthus species. Journal of
Heredity, 96, 410–416.
Wierzbicka M., Panufnik D. (1998) The adaptation of
Silene vulgaris to growth on a calamine waste heaps
(S. Poland). Environmental Pollution, 101, 415–426.
Wierzbicka M., Pielichowska M. (2004) Adaptation of
Biscutella laevigata L., a metal hyperaccumulator, to
growth on a zinc–lead waste heap in southern
Poland. I. Differences between waste-heap and
mountain populations. Chemosphere, 54, 1663–1674.
Wierzbicka M., Rostanski A. (2002) Microevolution-
ary changes in ecotypes of calamine waste heap
vegetation near Olkusz, Poland: a review. Acta Bio-
logica Cracoviensia. Series: Botanica, 44, 7–19.
Wierzbicka M., Słysz A. (2005) Does Armeria mariti-
ma subsp. halleri (Plumbaginaceae) occur in
Poland? Polish Botanical Studies, 19, 105–117.
Woodel S.R.J., Dale A. (1993) Biological flora of the
British Isles. Armeria maritima (Mill.) Willd. (Stat-
ice armeria L.; S. maritima Mill.). Journal of Ecol-
ogy, 81, 573–588.
Yeh F.C., Boyle T.J.B. (1997) Population genetic anal-
ysis of codominant and dominant markers and
quantitative traits. Belgian Journal of Botany, 129,
157.
Załecka R., Wierzbicka M. (2002) The adaptation of
Dianthus carthusianorum to growth on a calamine
waste heap. Plant and Soil, 246, 249–257.
Metallicolous and non-metallicolous populations of A. maritima Abratowska, Wasowicz, Bednarek, Telka & Wierzbicka
10 Plant Biology ª 2012 German Botanical Society and The Royal Botanical Society of the Netherlands
Top Related

Copyright © 2022 FDOKUMEN

