







Bahasa
Halaman
Hukum


1 23
Bioprocess and BiosystemsEngineering ISSN 1615-7591 Bioprocess Biosyst EngDOI 10.1007/s00449-014-1303-5
High-productivity lipid productionusing mixed trophic state cultivationof Auxenochlorella (Chlorella)protothecoides
Hamid Rismani-Yazdi, KristinH. Hampel, Christopher D. Lane, BenA. Kessler, Nicholas M. White, KennethM. Moats & F. C. Thomas Allnutt

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL PAPER
High-productivity lipid production using mixed trophic statecultivation of Auxenochlorella (Chlorella) protothecoides
Hamid Rismani-Yazdi • Kristin H. Hampel • Christopher D. Lane •
Ben A. Kessler • Nicholas M. White • Kenneth M. Moats •
F. C. Thomas Allnutt
Received: 28 May 2014 / Accepted: 7 October 2014
� Springer-Verlag Berlin Heidelberg 2014
Abstract A mixed trophic state production process for
algal lipids for use as feedstock for renewable biofuel
production was developed and deployed at subpilot scale
using a green microalga, Auxenochlorella (Chlorella)
protothecoides. The process is composed of two separate
stages: (1) the photoautotrophic stage, focused on biomass
production in open ponds, and (2) the heterotrophic stage
focused on lipid production and accumulation in aerobic
bioreactors using fixed carbon substrates (e.g., sugar). The
process achieved biomass and lipid productivities of 0.5
and 0.27 g/L/h that were, respectively, over 250 and 670
times higher than those obtained from the photoautotrophic
cultivation stage. The biomass oil content (over 60 %
w/DCW) following the two-stage process was predomi-
nantly monounsaturated fatty acids (*82 %) and largely
free of contaminating pigments that is more suitable for
biodiesel production than photosynthetically generated
lipid. Similar process performances were obtained using
cassava hydrolysate as an alternative feedstock to glucose.
Keywords Auxenochlorella (Chlorella) protothecoides �Algae biofuel � Heterotrophic � Photoautotrophic �Fermentation
Introduction
There has been a surge of activity in recent years focused on
new methods and their validation for the production of
economically and environmentally viable biofuels using
microalgae. A contributing factor to the increased activity
around algal biofuels was the increasing price of petroleum,
which peaked at US$145 per barrel in July of 2008. R&D
activity continues and improvements to the unit processes
surrounding algal biofuel production have occurred, such as
improvement in light penetration for algae production in
ponds [1], novel harvesting methods [2], extraction of lipids
[3, 4], hydrothermal liquefaction [5], innovative bioreactors
[4], and catalytic gasification [6]. Nonetheless, recent
analyses indicate that technical and economic hurdles
remain (as well as political obstacles) to profitable com-
mercialization [7]. Continued innovation and research are
required to overcome the remaining technological and
commercial barriers. It is clear that there is a need to replace
fossil fuels with renewable and sustainable fuels and that
H. Rismani-Yazdi � K. H. Hampel � C. D. Lane �B. A. Kessler � N. M. White � K. M. Moats �F. C. Thomas Allnutt
Phycal Inc., 51 Alpha Park, Highland Heights, OH 44143, USA
K. H. Hampel
e-mail: [email protected]
C. D. Lane
e-mail: [email protected]
B. A. Kessler
e-mail: [email protected]
N. M. White
e-mail: [email protected]
K. M. Moats
e-mail: [email protected]
F. C. Thomas Allnutt
e-mail: [email protected]
Present Address:
H. Rismani-Yazdi (&)
Novozymes North America Inc., P.O. Box 576, Franklinton,
NC 27525, USA
e-mail: [email protected]
Present Address:
F. C. Thomas Allnutt
BrioBiotech LLC, P.O. Box 26, Glenelg, MD 21737, USA
123
Bioprocess Biosyst Eng
DOI 10.1007/s00449-014-1303-5
Author's personal copy

algal biofuels still have tremendous potential to contribute
to this positive outcome if these hurdles can be overcome.
Mixed trophic state background
Traditionally, algal mass culturing is photoautotrophic,
with a complete reliance on solar energy and inorganic
carbon (carbon dioxide or bicarbonate) for conversion into
sugars and complex carbon compounds. Photoautotrophic
growth is, by its nature, self-limiting due to shading that
occurs as cell density increases (limiting light penetration)
and has other associated problems, such as pond water loss
through evaporation and insufficient gas exchange [8].
While the majority of algae are photoautotrophic, a sur-
prising number are capable of heterotrophic or mixotrophic
growth [9–13]. This led to efforts to look at the hetero-
trophic or mixotrophic production of biofuels from algae
[10]. Production of biomass through heterotrophy, mixo-
trophy, or using mixed trophic states provides an oppor-
tunity for improved productivity but requires an
inexpensive source of fixed carbon usable by algae (e.g.,
hydrolysates or partially purified sugars) to be commer-
cially relevant. With the increasing focus on the production
of ethanol from cellulosic feedstock and starch, it is
interesting to note that algae represent a way to produce
triglycerides from these same sugar sources that, once
transesterified, can be burned directly as transportation
fuels or act as an ideal feedstock for the production of drop-
in replacements for existing transportation fuels (i.e.,
having equivalent energy content and properties).
Employing microalgae cultivation systems that use
renewable inputs to generate biofuels is one potential
approach to improving the ability to reach economic targets
for production of algal biofuels. The production of algal
biomass based on mixotrophic growth, simultaneously
using light and fixed carbon for growth, is well known [14–
17]. Mixotrophic production systems for algae, based on
deep ponds and supplied with fixed carbon (e.g., glucose),
have been applied commercially for the production of
Chlorella for the health food markets [18]. Alternatively, a
mixed trophic cultivation process has been recently pro-
posed for the production of biofuels [19–21]. The mixed
trophic state systems use multiple trophic processes that are
separated either by time or location (for example, photo-
autotrophic growth then heterotrophic growth). A mixed
trophic state system is the focus of this work and will be
referred to as the Heteroboost process (HTB). This HTB
process has a photoautotrophic stage of algae cultivation in
open ponds that is followed by a concentration step and
then a heterotrophic growth stage in an aerobic bioreactor.
The process takes advantage of the ability of the sun to
provide cheap energy to photosynthetically fix inorganic
carbon from either the atmosphere or industrial emissions,
yet is able to rapidly accumulate oil to high density during
the heterotrophic growth using renewable inputs from
agricultural production of a fixed carbon feedstock (e.g.,
sugar, acetate or glycerol). The advantages anticipated in
the HTB process are the spatial and temporal separation of
inexpensive mass culture of algae on a continuous basis
(the photoautotrophic stage) and rapid lipid accumulation
in a more concentrated and constrained system (the het-
erotrophic bioreactor). The HTB process contrasts with
traditional photoautotrophic mass culturing systems where
the cells are grown to the highest density possible under
photoautotrophic conditions then subjected to stress (e.g.,
nutrient limitation such as nitrogen) to induce lipid syn-
thesis. The traditional photoautotrophic approach is limit-
ing in that it cannot be run as a continuous process and
requires additional time to induce lipid synthesis through
induction of the nutrient stress. The HTB process uses the
photoautotrophic step for biomass generation in open
ponds, which can be run in a continuous manner at the
optimized maximal production rate; this is an important
difference from the straight photoautotrophy and nutrient
stress approach traditionally used.
The ability of the microalga Auxenochlorella (Chlo-
rella) protothecoides to grow both photoautotrophically
and heterotrophically, when combined with its high pho-
tosynthetic efficiency of 8 % [22] and ability to accumulate
large quantities of neutral lipids (up to 70 %) [23] at rel-
atively high cell densities using a range of fixed carbon
substrates [24], make it a suitable candidate for the HTB
process. A recent review of the genus Chlorella discovered
that species traditionally ascribed to Chlorella were spread
over two classes of chlorophytes, the Trebouxiophyceae
and the Chlorophyceae [25]. The main species used in the
current study was placed in the Trebouxiophyceae and
renamed as Auxenochlorella protothecoides. An extensive
academic and patent background has been developed
around the heterotrophic growth and lipid production in A.
protothecoides [10, 21, 26]. The objective of this study was
to develop and evaluate a mixed trophic state cultivation
strategy for high productivity mass culturing and lipid
production by A. protothecoides at subpilot scale.
Materials and methods
Organism and medium composition
Auxenochlorella (Chlorella) protothecoides KRT1009, a
single-celled green alga belonging to the phylum Chloro-
phyta, was selected from UTEX 25 obtained from the
University of Texas Culture Collection. UTEX 25 as
received contained mixed cell types, a clonal line of A.
protothecoides was isolated from this mother culture and
Bioprocess Biosyst Eng
123
Author's personal copy

verified using the cleaved amplified polymorphic sequence
techniques [27]. For photoautotrophic growth, the medium
was composed of 0.5 g NH4Cl, 1.44 g K2HPO4, 0.72 g
KH2PO4, 50 mg tetrasodium EDTA, 20 mg MgSO4�7H2O,
10 mg CaCl2�2H2O, 10 mg FeCl3�6H2O, 5 mg H3BO3,
1.25 mg ZnSO4�7H2O, 0.38 MnSO4�H2O, 0.25 mg CoCl2�6H2O, 0.25 mg Na2MoO4�2H2O, 0.08 mg CuSO4�5H2O, and
100 lg thiamine hydrochloride per liter of water.
Subculture and seed cultivation
Seed cultures were grown in an inoculum train as shown in
Fig. 1. Seed cultures were maintained on agar plates and
transferred regularly to new plates such that the age of the
cells on the plate at the time of starting the first flask was
consistent. The stages of the inoculum train are presented
in Table 1: (1) 25 mL volume in a 250-mL flask, (2)
250 mL in a 500-mL flask, (3) 2 L volume in a 2-L flat-
bottom flask, and (4) 20 L volume in a carboy. Based on
historical data, cultures were transferred to maintain an
exponentially growing population up to the carboy stage.
Stages 1 and 2 were maintained at 28 ± 2 �C, a constant
140 ± 10 lmol/m2/s light intensity from an Eye Hortilux
metal halide bulb (Iwasaki MT1000B-D/HOR/HTL-BL1)
in a SunTubeTM six-inch reflector (Sunlight Supply, Van-
couver, WA), 5,000 ppm ambient CO2 concentration, and
180 rpm shaking speed. Stage 3 was maintained at the
same light and temperature levels, but was bubbled with a
sterile-filtered air/CO2 mix (approximately 1 % (w/w)
CO2) at 1 L/min. The gas flow provided both the mixing
and carbon source. Aseptic technique was used to maintain
an aseptic culture until the carboy stage. The Stage 4 car-
boy was maintained at the same temperature, light, and
CO2 conditions as Stage 3, but with a higher total gas flow
rate that was set manually to provide optimal mixing.
Flasks and carboys were grown in triplicate vessels.
Photoautotrophic growth in raceway ponds
The carboy volumes were combined and mixed before
being redistributed into two 2,400 L raceway ponds that
were approximately 4.5 m long, 1.2 m wide and 0.45 m
deep, and were operated at 15 cm volume depth with 800 L
working volume. Raceway ponds were grown in duplicate.
Mixing was provided through a paddlewheel with a liquid
linear velocity of no less than 0.2 cm/s throughout the
pond. Eye Hortilux metal halide bulbs (Iwasaki) in Sun-
TubeTM six-inch reflectors (Sunlight Supply) were used to
provide the culture with an average of 700 lmol/m2/s
incident light intensity. Carbon dioxide was provided
through a sintered metal sparger into each pond based on a
feedback loop controlling a solenoid valve. The valve was
Fig. 1 A schematic diagram of
the HTB mixed trophic state
process and the seed train for
production of inoculum for
ponds in the subpilot plant
Bioprocess Biosyst Eng
123
Author's personal copy

opened when the pH of the culture was above the pH set-
point value of 6.7. Temperature was controlled at
28 ± 2 �C using an immersion chiller. The two ponds were
operated for 5.4 days and achieved a volumetric produc-
tivity of 0.04 ± 0.01 g/L/day and an aerial productivity of
4.9 ± 2.0 g/m2/day, before being harvested.
Biomass dewatering
Photoautotrophically grown cells in the ponds (1,600 L)
were harvested and transferred to a centrifuge feed tank. A
stacked disc centrifuge (Alfa Laval MAPX207) was used to
dewater and concentrate the cells. The centrifuge had 7.6 L
of solids capacity and operated at 8009g at 24 ± 2 �C.
Centrifuge discharge frequency was adjusted to obtain the
desired biomass concentration for the heterotrophic growth
stage. The concentrated biomass was collected and trans-
ferred rapidly to the bioreactors.
Heterotrophic growth (Fed-batch operation)
Heterotrophic growth of A. protothecoides was conducted
in 10 L Biostat B fermentors (Sartorius, Bohemia, NY).
The bioreactor was filled with 5 L of the concentrated cell
slurry, and the pH was adjusted and set to 6.5, using 3 M
NaOH. The cultivation temperature was controlled at
28 �C, and dissolved oxygen (DO) was set at C20 %. The
aeration flow rate was kept at 4 L/min. Agitation was
controlled automatically in response to the DO level. A
proprietary organic antifoam DF 204 (BASF, Florham
Park, NJ) was used for foam control as needed. A con-
centrated glucose or cassava starch hydrolysate solution
(500–600 g glucose-equivalent/L) was fed manually into
the bioreactor using a peristaltic pump to maintain the
residual glucose concentration in the bioreactor between 10
and 30 g/L throughout the fermentation. Stock solutions
were continuously stirred with a magnetic stirrer. Samples
from the bioreactor were collected periodically for cell
growth determination, glucose analysis, and lipid
extraction.
Preparation of cassava starch hydrolysate
Pure cassava starch was used for preparing the cassava
starch hydrolysate as an alternative substrate to lab-grade
glucose for A. protothecoides fermentation. The lique-
faction and saccharification processes were done in a 50 L
Letsch stainless steel reactor (Letsch Corp., Springfield,
MO). For the liquefaction, the substrate slurry (20 % w/v
on dry starch basis, DSB), was titrated to pH 5.7 with 6 N
NaOH, and alpha amylase at 0.02 % w/w DSB (Spezyme
Alpha, Genencor, Palo Alto, CA) was added to the slurry.
The mixture was heated to 85 �C and held for 90 min
under continuous agitation. The liquefied starch was then
saccharified (pH 4.3 at 55 �C for 24 h) using glucoamy-
lase at 0.05 % w/w DSB (Optidex L-400, Genencor, Palo
Alto, CA). Samples of the mixture were analyzed by
HPLC, as described below, to determine the glucose-
equivalent sugar content of the mixture. The syrup was
diluted to the desired glucose-equivalent concentration
with water and sterilized by autoclaving before being used
in the fermentation.
Biomass measurement
Cell growth was estimated by measuring absorbance at
750 nm (A750) or the dry cell weight (DCW, g/L). The
DCW was determined by removing a sample that would
provide at least 10 mg of DCW (estimated empirically).
This sample was filtered through a pre-weighed 0.4 lm
glass fiber filter (Whatman), rinsed three times with 5 mL
ammonium bicarbonate solution (125 mM), and weighed
wet. The sample was then put into a Shimadzu MOC 120H
electronic moisture balance (Shimadzu Corp, Kyoto,
Japan) and run until the dry weight was constant. The
DCW was determined by correction for the weight of the
filter and the volume of sample filtered.
Table 1 Conditions for
photoautotrophic production of
A. protothecoides biomass
Inoculum
stages
Vessel Vessel
capacity
(L)
Culture
volume
(L)
Light
intensity
(lmol/m2/s)
Starting
density
(A750)
Final
density
(A750)
Duration
(days)
Stage 1 Erlenmeyer
flask
0.25 0.025 140 0.08 ± 0.0 1.37 ± 0.08 1.8
Stage 2 Erlenmeyer
flask
0.50 0.25 140 0.19 ± 0.0 0.75 ± 0.03 1.0
Stage 3 Flat-bottom
flask
2 2 140 0.08 ± 0.01 3.06 ± 0.75 4.2
Stage 4 Carboy 20 20 140 0.31 ± 0.02 2.67 ± 0.67 7.1
Pond Raceway
pond
2,400 800 700 0.34 ± 0.08 1.26 ± 0.11 5.4
Bioprocess Biosyst Eng
123
Author's personal copy

Glucose analysis
For glucose analysis, the culture broth supernatant or cas-
sava hydrolysate was diluted to the appropriate concen-
tration using HPLC grade water and filtered through
0.45 lm cellulose-acetate filters (Whatman). Samples were
analyzed on a Thermo Scientific HPLC system equipped
with a Surveyor LC pump plus quaternary gradient pump,
Surveyor Autosampler, Surveyor IR Plus detector, and
ChromQuest 5.0 software (Thermo/Finnigan, Waltham,
Massachusetts, USA). Separation was achieved on a
300 mm 9 7.8 mm, 9 lm particle size Aminex HPX-87H
column (Bio Rad, Hercules, CA, USA). The column was
held at 55 �C using a heating blanket. The mobile phase
was 5 mM H2SO4 with a flow rate of 0.6 mL/min. Injec-
tion was achieved using a 20-lL injection loop. Glucose
quantification was achieved through comparison to a glu-
cose standard curve created from ultra-pure glucose
(Sigma-Aldrich, St. Louis, MO) at concentrations ranging
from 0.5 to 10 g/L glucose. The glucose calibration curve
was generated by plotting the glucose chromatogram peak
areas against the glucose concentration, and the linearity of
the line was evaluated using least squares regression.
Lipid extraction
Lipid extraction was conducted using a modified Bligh and
Dyer method [28]. Briefly, lyophilized cell pellet
(50–300 mg) was mixed with a 1:2 chloroform/methanol
solution in a 50-mL glass centrifuge tube and five 6.35 mm
glass beads were added. The samples were vortexed for
1 h. After mixing, an additional aliquot of water was added
to each sample to bring the ratio to 1:2:0.8 chloroform/
methanol/water and samples centrifuged at 560 9g for
10 min at room temperature using a Beckman G6B cen-
trifuge. The chloroform layer was collected, and the sol-
vent was removed by evaporation under N2. Extracted
material was weighed using a high precision analytical
balance (Sartorius ED124S) with 0.1 mg sensitivity to
provide a gravimetric measurement of total lipid.
Determination of fatty acid methyl esters (FAMES)
Transesterification of the lipid extract was achieved by
placing a lipid sample into a screw-capped glass test tube,
adding 5.3 mL of methanol and 0.58 mL of 12 M sulfuric
acid. Additionally, 1 mL of a solution containing 2.2 mg/
mL (final concentration of 0.74 mg/mL) nonadecanoic acid
was added to each test tube as an internal standard. Sam-
ples were capped and placed in a dry heat bath at 75 �C for
90 min. Every 20 min the samples were removed and
mixed vigorously for 20 s. Samples were cooled to room
temperature and neutralized using KOH. Hexane was
added to extract the FAMEs from the methanol phase. The
hexane phase was filtered through anhydrous sodium sul-
fate. FAME analysis was performed using a Trace GC
Ultra Gas Chromatography system equipped with flame
ionization detector, AS 3000 auto sampler, and Chrom-
Quest 5.0 software (Thermo Fisher). The oven was
equipped with a 30 m 9 0.32 mm 9 0.5 lm BPX-90
column (SGE Analytical Science, USA). The identity of
each FAME peak was determined by comparing the
retention times of the compounds to the retention time of
FAME standards (Sigma-Aldrich, St. Louis, MO). The
internal standard (nonadecanoic methyl ester) method was
used to quantify each FAME peak by relative area. All
analytical measurements for biomass, glucose, lipid and
FAME were performed in duplicate and results are pre-
sented as Mean ± SD.
Nile red staining and microscopy
Cells were stained with Nile Red dye and imaged using a
fluorescent microscope. Nile Red stain is a neutral oil stain
that permeates cell membrane and targets triglycerides, or
neutral oils [29]. Microscopy was performed with a Nikon
Eclipse E200 light microscope equipped with an Amscope
MD900 digital camera. The dye was excited with a Mer-
cury lamp and measured with an absorbance and emission
filter sets of 450–490 and 500–515 nm, respectively.
Results and discussion
A mixed trophic state cultivation strategy for mass cul-
turing and lipid production by A. protothecoides was
designed and tested at subpilot scale. The process, referred
to here as the HTB process, consists of a photoautotrophic
open pond cultivation of algae that is followed by con-
centration and a heterotrophic stage (Fig. 1). The advan-
tages anticipated in such an approach are the spatial and
temporal separation of inexpensive mass culture of algae
on a continuous basis (the photoautotrophic stage) and high
productivity lipid accumulation in a more concentrated and
constrained system (the heterotrophic bioreactor). The
HTB process uses the photoautotrophic step for mass cul-
ture in open ponds, which can be run in a continuous
manner, and then provides a concentrated algal suspension
that is fed with fixed carbon (e.g., glucose) to induce rapid
lipid accumulation. The inoculum is produced in aseptic
culture then aseptically and sequentially transferred to
progressively larger photoautotrophic culture systems until
it reaches the carboy stage where asepsis is no longer
maintained (Fig. 1). The carboy is transferred to open
ponds where the bulk of the photoautotrophically produced
biomass is generated.
Bioprocess Biosyst Eng
123
Author's personal copy

Photoautotrophic growth
A representative photoautotrophic growth profile of A.
protothecoides in open raceway ponds is shown in Fig. 2a,
and key performance indicators of the process are listed in
Table 2. Photoautotrophic cultivation of A. protothecoides
in open ponds resulted in a final biomass concentration of
0.3 ± 0.1 g/L and an aerial biomass productivity of
4.9 ± 2.0 g/m2/day (Table 2). During the photoautotrophic
growth in open ponds, A. protothecoides biomass
concentration reached 0.3 ± 0.1 g/L, with the lipid content
of 17.7 ± 2.3 %, on the basis of dry cell weight, and the
average volumetric lipid productivity of 0.4 ± 0.1 mg/L/h.
At the end of the photoautotrophic stage, algal biomass
from the ponds was harvested by centrifugation to obtain a
highly concentrated seed for the heterotrophic phase of the
process.
Heterotrophic growth
During the heterotrophic stage of the HTB process, con-
centrated biomass of photoautotrophically grown A. prot-
othecoides was added as inoculum to 10 L bioreactors and
grown under fed-batch conditions using glucose as the sole
fixed carbon substrate. A representative growth profile for
photoautotrophically cultivated A. protothecoides grown
on glucose in the bioreactor is shown in Fig. 2b, and key
performance indicators of the process are listed in Table 3.
Under heterotrophic growth conditions, cells changed color
from green to yellow through degradation of chlorophyll
and produced on average 116.7 ± 15.5 DCW/L biomass
containing over 60 % lipid (Table 2). The accumulated
lipids were deposited as intracellular lipid bodies, which
could easily be detected by a fluorescent probe, Nile red
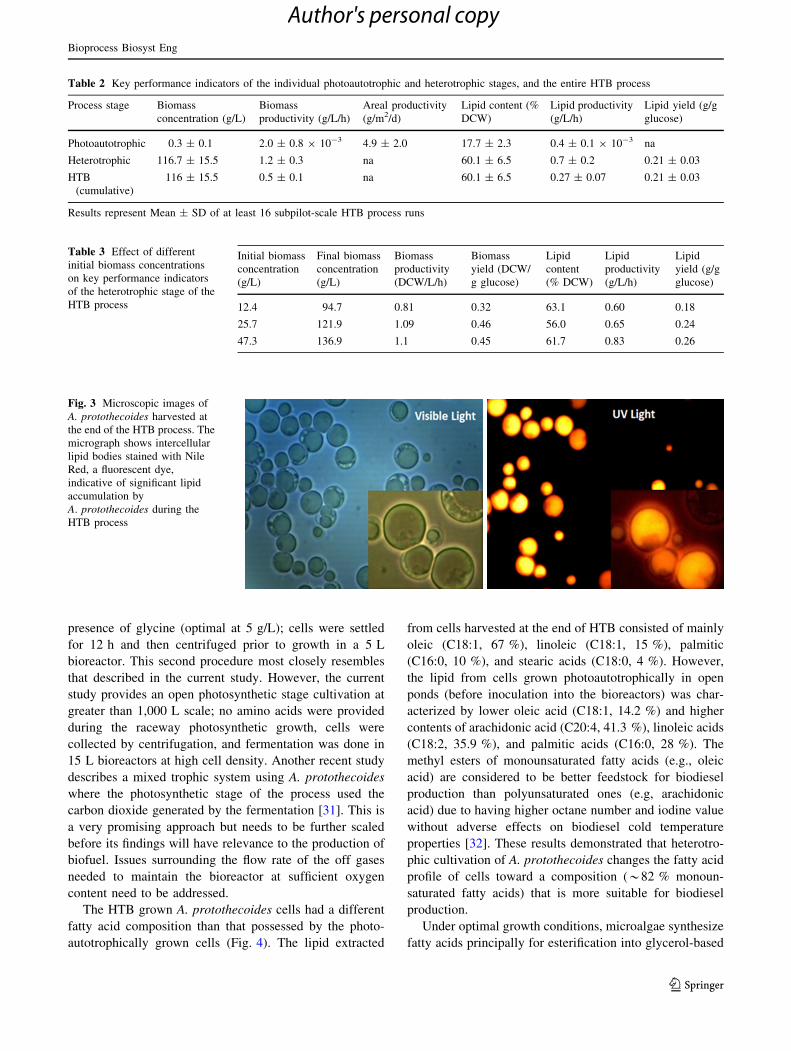
(Fig. 3). The volumetric rate of lipid production during the
heterotrophic stage was 0.7 ± 0.2 g/L/h, which was more
than three orders of magnitude higher than that achieved
during photoautotrophic cultivation stage. Similarly, the
biomass productivity in the heterotrophic stage of the
process was 600-fold higher than that achieved under
photoautotrophic conditions (Table 2). These significantly
higher biomass and lipid productivities during the hetero-
trophic stage of the process lend the HTB process signifi-
cant performance advantages over growing oleaginous
algae solely photoautotrophically. The cumulative biomass
and lipid productivities of the HTB process are presented in
Table 2 and were over 250 and 600 times higher, respec-
tively, than those obtained from the photoautotrophic cul-
tivation stage.
The concept of using a mixed trophic state system for
production of biofuels was described in a recent patent
[19]. The methods described in the Oyler patent were
poorly defined and no data were provided with any algal
strain. Subsequent patent applications by Sayre [30] and
Wu and Xiong [21] provided more detail on a mixed tro-
phic state production process. The process described by
Sayre provided data using a mixed trophic growth system
at shake flask scale where the cells were grown aseptically
photosynthetically, concentrated, then grown heterotro-
phically on glycerol [30]; no scaled up data were provided.
The process of Wu and Xiong [21] described shake flask
and small fermentor scale mixed trophic growth of A.
protothecoides. The phototrophic growth was done in the
Fig. 2 Photoautotrophic and heterotrophic growth of A. prototheco-
ides KRT1009. a Representative growth of A. protothecoides
KRT1009 in an open 800 L raceway pond with 15 cm depth and
2.6 m2 culture area. Pond bacterial contamination was 120 CFU/mL,
and areal productivity was 7.3 g/m2/day. b Representative growth and
lipid production by A. protothecoides KRT1009 using glucose as the
source of substrate in a 10 L bioreactor. The bioreactor was seeded
with photoautotrophically grown cells (40 g/L) concentrated by
centrifugation from open ponds. Bioreactor initial volume was 5 L,
and temperature, pH and dissolved oxygen were controlled at 28 �C,
6.5 and 22 %, respectively. The bioreactors were run as a fed-batch
for 75 h
Bioprocess Biosyst Eng
123
Author's personal copy
presence of glycine (optimal at 5 g/L); cells were settled
for 12 h and then centrifuged prior to growth in a 5 L
bioreactor. This second procedure most closely resembles
that described in the current study. However, the current
study provides an open photosynthetic stage cultivation at
greater than 1,000 L scale; no amino acids were provided
during the raceway photosynthetic growth, cells were
collected by centrifugation, and fermentation was done in
15 L bioreactors at high cell density. Another recent study
describes a mixed trophic system using A. protothecoides
where the photosynthetic stage of the process used the
carbon dioxide generated by the fermentation [31]. This is
a very promising approach but needs to be further scaled
before its findings will have relevance to the production of
biofuel. Issues surrounding the flow rate of the off gases
needed to maintain the bioreactor at sufficient oxygen
content need to be addressed.
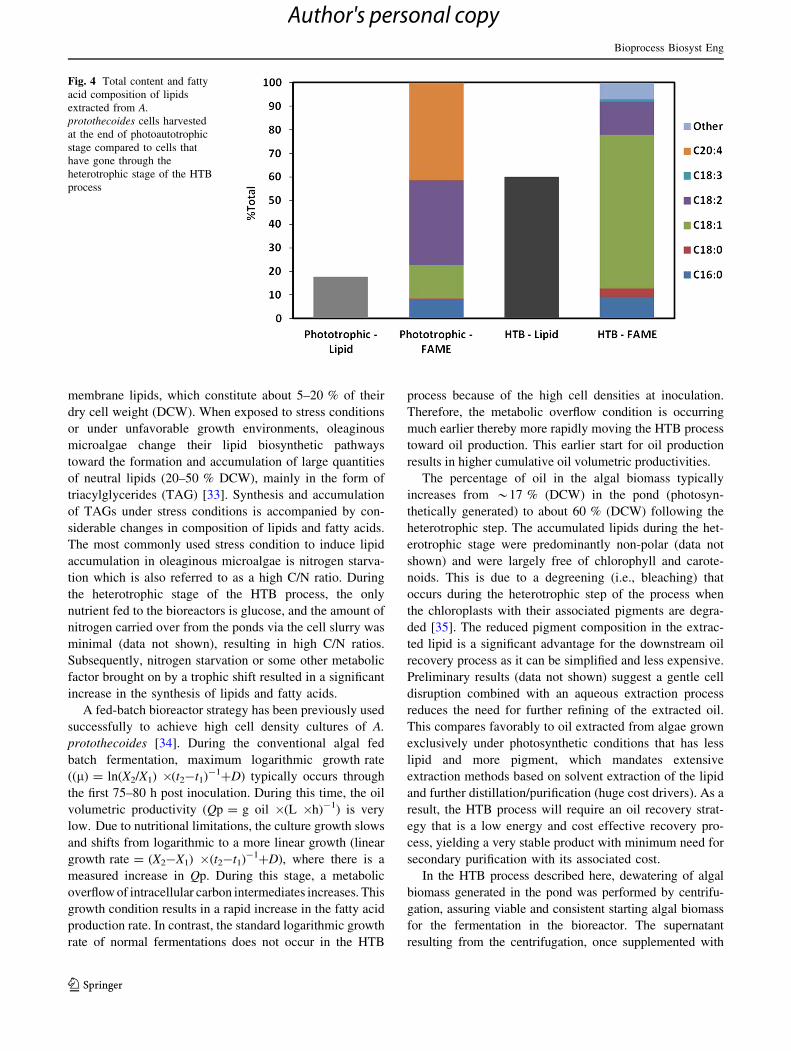
The HTB grown A. protothecoides cells had a different
fatty acid composition than that possessed by the photo-
autotrophically grown cells (Fig. 4). The lipid extracted
from cells harvested at the end of HTB consisted of mainly
oleic (C18:1, 67 %), linoleic (C18:1, 15 %), palmitic
(C16:0, 10 %), and stearic acids (C18:0, 4 %). However,
the lipid from cells grown photoautotrophically in open
ponds (before inoculation into the bioreactors) was char-
acterized by lower oleic acid (C18:1, 14.2 %) and higher
contents of arachidonic acid (C20:4, 41.3 %), linoleic acids
(C18:2, 35.9 %), and palmitic acids (C16:0, 28 %). The
methyl esters of monounsaturated fatty acids (e.g., oleic
acid) are considered to be better feedstock for biodiesel
production than polyunsaturated ones (e.g, arachidonic
acid) due to having higher octane number and iodine value
without adverse effects on biodiesel cold temperature
properties [32]. These results demonstrated that heterotro-
phic cultivation of A. protothecoides changes the fatty acid
profile of cells toward a composition (*82 % monoun-
saturated fatty acids) that is more suitable for biodiesel
production.
Under optimal growth conditions, microalgae synthesize
fatty acids principally for esterification into glycerol-based
Fig. 3 Microscopic images of
A. protothecoides harvested at
the end of the HTB process. The
micrograph shows intercellular
lipid bodies stained with Nile
Red, a fluorescent dye,
indicative of significant lipid
accumulation by
A. protothecoides during the
HTB process
Table 2 Key performance indicators of the individual photoautotrophic and heterotrophic stages, and the entire HTB process
Process stage Biomass
concentration (g/L)
Biomass
productivity (g/L/h)
Areal productivity
(g/m2/d)
Lipid content (%
DCW)
Lipid productivity
(g/L/h)
Lipid yield (g/g
glucose)
Photoautotrophic 0.3 ± 0.1 2.0 ± 0.8 9 10-3 4.9 ± 2.0 17.7 ± 2.3 0.4 ± 0.1 9 10-3 na
Heterotrophic 116.7 ± 15.5 1.2 ± 0.3 na 60.1 ± 6.5 0.7 ± 0.2 0.21 ± 0.03
HTB
(cumulative)
116 ± 15.5 0.5 ± 0.1 na 60.1 ± 6.5 0.27 ± 0.07 0.21 ± 0.03
Results represent Mean ± SD of at least 16 subpilot-scale HTB process runs
Table 3 Effect of different
initial biomass concentrations
on key performance indicators
of the heterotrophic stage of the
HTB process
Initial biomass
concentration
(g/L)
Final biomass
concentration
(g/L)
Biomass
productivity
(DCW/L/h)
Biomass
yield (DCW/
g glucose)
Lipid
content
(% DCW)
Lipid
productivity
(g/L/h)
Lipid
yield (g/g
glucose)
12.4 94.7 0.81 0.32 63.1 0.60 0.18
25.7 121.9 1.09 0.46 56.0 0.65 0.24
47.3 136.9 1.1 0.45 61.7 0.83 0.26
Bioprocess Biosyst Eng
123
Author's personal copy
membrane lipids, which constitute about 5–20 % of their
dry cell weight (DCW). When exposed to stress conditions
or under unfavorable growth environments, oleaginous
microalgae change their lipid biosynthetic pathways
toward the formation and accumulation of large quantities
of neutral lipids (20–50 % DCW), mainly in the form of
triacylglycerides (TAG) [33]. Synthesis and accumulation
of TAGs under stress conditions is accompanied by con-
siderable changes in composition of lipids and fatty acids.
The most commonly used stress condition to induce lipid
accumulation in oleaginous microalgae is nitrogen starva-
tion which is also referred to as a high C/N ratio. During
the heterotrophic stage of the HTB process, the only
nutrient fed to the bioreactors is glucose, and the amount of
nitrogen carried over from the ponds via the cell slurry was
minimal (data not shown), resulting in high C/N ratios.
Subsequently, nitrogen starvation or some other metabolic
factor brought on by a trophic shift resulted in a significant
increase in the synthesis of lipids and fatty acids.
A fed-batch bioreactor strategy has been previously used
successfully to achieve high cell density cultures of A.
protothecoides [34]. During the conventional algal fed
batch fermentation, maximum logarithmic growth rate
((l) = ln(X2/X1) � (t2-t1)-1?D) typically occurs through
the first 75–80 h post inoculation. During this time, the oil
volumetric productivity (Qp = g oil � (L � h)-1) is very
low. Due to nutritional limitations, the culture growth slows
and shifts from logarithmic to a more linear growth (linear
growth rate = (X2-X1) � (t2-t1)-1?D), where there is a
measured increase in Qp. During this stage, a metabolic
overflow of intracellular carbon intermediates increases. This
growth condition results in a rapid increase in the fatty acid
production rate. In contrast, the standard logarithmic growth
rate of normal fermentations does not occur in the HTB
process because of the high cell densities at inoculation.
Therefore, the metabolic overflow condition is occurring
much earlier thereby more rapidly moving the HTB process
toward oil production. This earlier start for oil production
results in higher cumulative oil volumetric productivities.
The percentage of oil in the algal biomass typically
increases from *17 % (DCW) in the pond (photosyn-
thetically generated) to about 60 % (DCW) following the
heterotrophic step. The accumulated lipids during the het-
erotrophic stage were predominantly non-polar (data not
shown) and were largely free of chlorophyll and carote-
noids. This is due to a degreening (i.e., bleaching) that
occurs during the heterotrophic step of the process when
the chloroplasts with their associated pigments are degra-
ded [35]. The reduced pigment composition in the extrac-
ted lipid is a significant advantage for the downstream oil
recovery process as it can be simplified and less expensive.
Preliminary results (data not shown) suggest a gentle cell
disruption combined with an aqueous extraction process
reduces the need for further refining of the extracted oil.
This compares favorably to oil extracted from algae grown
exclusively under photosynthetic conditions that has less
lipid and more pigment, which mandates extensive
extraction methods based on solvent extraction of the lipid
and further distillation/purification (huge cost drivers). As a
result, the HTB process will require an oil recovery strat-
egy that is a low energy and cost effective recovery pro-
cess, yielding a very stable product with minimum need for
secondary purification with its associated cost.
In the HTB process described here, dewatering of algal
biomass generated in the pond was performed by centrifu-
gation, assuring viable and consistent starting algal biomass
for the fermentation in the bioreactor. The supernatant
resulting from the centrifugation, once supplemented with
Fig. 4 Total content and fatty
acid composition of lipids
extracted from A.
protothecoides cells harvested
at the end of photoautotrophic
stage compared to cells that
have gone through the
heterotrophic stage of the HTB
process
Bioprocess Biosyst Eng
123
Author's personal copy

nutrients and cleared from potential growth inhibitors, can
be recycled back to the ponds for many cycles without
additional water treatment, resulting in significant reduction
in water treatment cost. While this has not been done yet at
large scale, small-scale recycling of spent medium using
6.5 L aquaria was used to demonstrate this ability. Starting
with fresh medium, cells were grown and then collected by
centrifugation and the spent medium reused in the same
system for growth of A. protothecoides. These experiments
were performed in triplicate and compared to aquaria pro-
vided only with fresh medium. Using three cycles of growth
and reuse, there was no significant difference in the recycled
medium growth rate (8.0 g/m2/d) and that using the fresh
medium (7.9 g/m2/d).
Microbial contamination of algal biomass is inevitable
when the culture is grown in non-aseptic conditions, such
as raceways and open ponds. The expectation is that bac-
terial contamination will be transferred from the open
photosynthetic culture to the bioreactor by way of the
concentrated inoculum. It was a pleasant surprise that in
the bioreactors the A. protothecoides dominated and pro-
ductivity was not negatively impacted by bacterial load.
The bacteria are still present and had an average daily
doubling time of 0.62 (STD 0.01; % CV 1.3 %), while the
A. protothecoides had a doubling time of 1.35 (STD 0.11;
% CV 8.15 %). In the scaled up process outlined in this
study, on harvest from the bioreactor, the contaminating
organisms present in the broth have been very low, to the
extent that no significant differences in key performance
indicators were observed when results were compared with
those obtained from bioreactors seeded with axenic cul-
tures (data not shown). These results are consistent with
reports of a recent study demonstrating that A. prototh-
ecoides is able to grow well in non-sterilized wastewater
with no effects of bacterial contamination on its growth
rate [36]. An interesting observation is that when this
process was run with Chlorella vulgaris the bioreactor was
overrun with bacteria that crashed the culture.
Effect of initial biomass concentration (i.e, inoculum
size) on fermentation
Lipid production by A. protothecoides during the hetero-
trophic stage of the HTB process was carried out with
different inoculum sizes to determine the effect of initial
biomass concentration on key performance indicators of
the fermentation process. The initial inocula sizes evalu-
ated were 12, 25, and 47 g/L at the starting volume of 5 L
in 10 L bioreactors. Figure 5 follows the changes in bio-
mass concentration and lipid content during the course of
120 h fermentation at various inoculum sizes, and the key
performance indicators of the fermentation are summarized
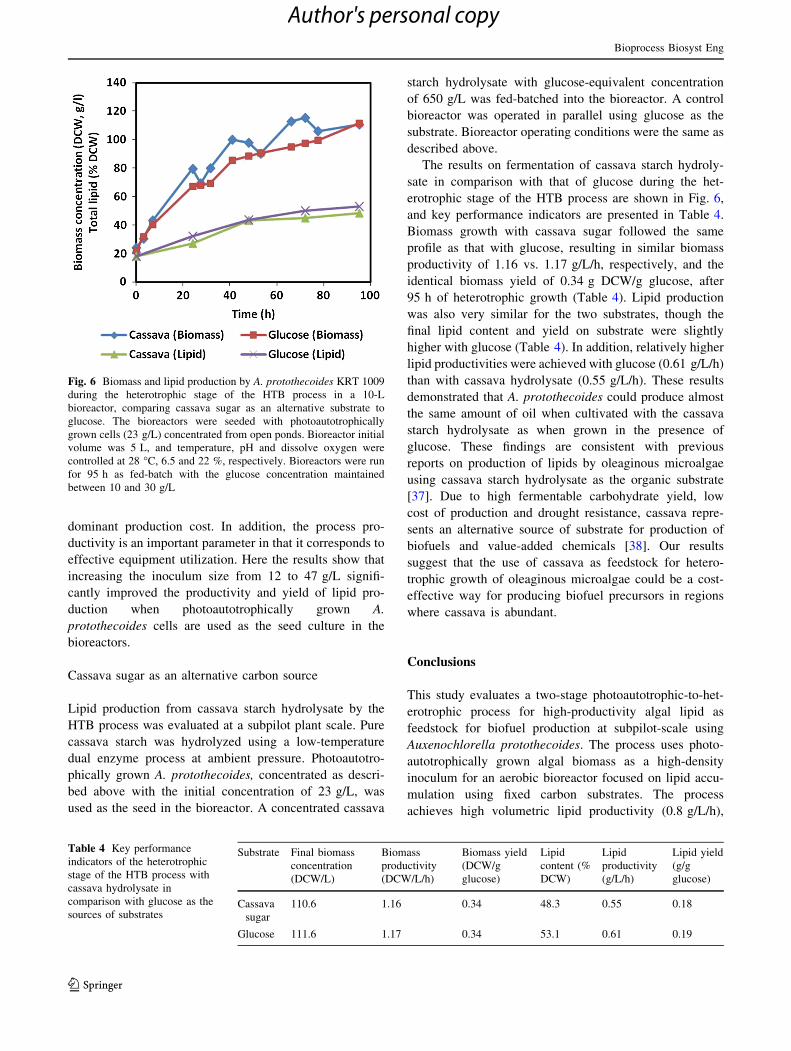
in Table 3. The final biomass concentration increased with
increase in the initial inoculum size. The increase in bio-
mass concentration corresponded to higher biomass pro-
ductivities and higher yield of biomass on glucose at larger
inoculum sizes. The 25 and 47 g/L inoculum sizes resulted
in similar biomass productivities (1.09 vs. 1.1 g/L/h) and
biomass yields (0.46 vs. 0.45 g/g), which were higher than
the values achieved with the 12 g/L inoculum size, 0.81 g/
L/h and 0.32 g/g, respectively (Table 3). The late (*80 h)
decline in biomass concentration from the lowest inoculum
size tested (i.e. 12 g/L) could be due to competition for
nutrients with contaminating bacteria. This, however, was
not verified experimentally. The inoculum size affected the
lipid production, especially in terms of lipid productivity
and lipid yield. Lipid productivity and lipid yield increased
considerably with an increase in inoculum size, where the
highest values achieved for these parameters were 0.83 (g/
L/h) and 0.26 (g/g), respectively, by the 47 g/L inoculum
size (Table 3). The highest lipid content (63 % DCW) was
obtained by 12 g/L inoculum size, whereas the 25 g/L
inoculum resulted in the least lipid accumulation (56 %
DCW) by the cells.
Lipid production by oleaginous organisms (e.g., A.
protothecoides) via HTB growth must have the primary
objective of maximizing the lipid yield on the substrate
(i.e., glucose), because the cost of the carbon source is the
Fig. 5 Effect of different initial biomass concentrations on biomass
and lipid production by A. protothecoides KRT 1009 during the
heterotrophic stage of the HTB process in a 10-L bioreactor. The
bioreactors were seeded with photoautotrophically grown cells (12.4,
25.7, and 47.3 g/L) concentrated by centrifugation from open ponds.
Bioreactor was run as fed-batch with the glucose concentration
maintained between 10 and 30 g/L. Bioreactor initial volume was
5 L, and temperature, pH and dissolved oxygen were controlled at
28 �C, 6.5 and 22 %, respectively. The bioreactors were run as a fed-
batch for 120 h
Bioprocess Biosyst Eng
123
Author's personal copy
dominant production cost. In addition, the process pro-
ductivity is an important parameter in that it corresponds to
effective equipment utilization. Here the results show that
increasing the inoculum size from 12 to 47 g/L signifi-
cantly improved the productivity and yield of lipid pro-
duction when photoautotrophically grown A.
protothecoides cells are used as the seed culture in the
bioreactors.
Cassava sugar as an alternative carbon source
Lipid production from cassava starch hydrolysate by the
HTB process was evaluated at a subpilot plant scale. Pure
cassava starch was hydrolyzed using a low-temperature
dual enzyme process at ambient pressure. Photoautotro-
phically grown A. protothecoides, concentrated as descri-
bed above with the initial concentration of 23 g/L, was
used as the seed in the bioreactor. A concentrated cassava
starch hydrolysate with glucose-equivalent concentration
of 650 g/L was fed-batched into the bioreactor. A control
bioreactor was operated in parallel using glucose as the
substrate. Bioreactor operating conditions were the same as
described above.
The results on fermentation of cassava starch hydroly-
sate in comparison with that of glucose during the het-
erotrophic stage of the HTB process are shown in Fig. 6,
and key performance indicators are presented in Table 4.
Biomass growth with cassava sugar followed the same
profile as that with glucose, resulting in similar biomass
productivity of 1.16 vs. 1.17 g/L/h, respectively, and the
identical biomass yield of 0.34 g DCW/g glucose, after
95 h of heterotrophic growth (Table 4). Lipid production
was also very similar for the two substrates, though the
final lipid content and yield on substrate were slightly
higher with glucose (Table 4). In addition, relatively higher
lipid productivities were achieved with glucose (0.61 g/L/h)
than with cassava hydrolysate (0.55 g/L/h). These results
demonstrated that A. protothecoides could produce almost
the same amount of oil when cultivated with the cassava
starch hydrolysate as when grown in the presence of
glucose. These findings are consistent with previous
reports on production of lipids by oleaginous microalgae
using cassava starch hydrolysate as the organic substrate
[37]. Due to high fermentable carbohydrate yield, low
cost of production and drought resistance, cassava repre-
sents an alternative source of substrate for production of
biofuels and value-added chemicals [38]. Our results
suggest that the use of cassava as feedstock for hetero-
trophic growth of oleaginous microalgae could be a cost-
effective way for producing biofuel precursors in regions
where cassava is abundant.
Conclusions
This study evaluates a two-stage photoautotrophic-to-het-
erotrophic process for high-productivity algal lipid as
feedstock for biofuel production at subpilot-scale using
Auxenochlorella protothecoides. The process uses photo-
autotrophically grown algal biomass as a high-density
inoculum for an aerobic bioreactor focused on lipid accu-
mulation using fixed carbon substrates. The process
achieves high volumetric lipid productivity (0.8 g/L/h),
Fig. 6 Biomass and lipid production by A. protothecoides KRT 1009
during the heterotrophic stage of the HTB process in a 10-L
bioreactor, comparing cassava sugar as an alternative substrate to
glucose. The bioreactors were seeded with photoautotrophically
grown cells (23 g/L) concentrated from open ponds. Bioreactor initial
volume was 5 L, and temperature, pH and dissolve oxygen were
controlled at 28 �C, 6.5 and 22 %, respectively. Bioreactors were run
for 95 h as fed-batch with the glucose concentration maintained
between 10 and 30 g/L
Table 4 Key performance
indicators of the heterotrophic
stage of the HTB process with
cassava hydrolysate in
comparison with glucose as the
sources of substrates
Substrate Final biomass
concentration
(DCW/L)
Biomass
productivity
(DCW/L/h)
Biomass yield
(DCW/g
glucose)
Lipid
content (%
DCW)
Lipid
productivity
(g/L/h)
Lipid yield
(g/g
glucose)
Cassava
sugar
110.6 1.16 0.34 48.3 0.55 0.18
Glucose 111.6 1.17 0.34 53.1 0.61 0.19
Bioprocess Biosyst Eng
123
Author's personal copy

lipid contents (63 % w/DCW), and yield (yp/s 0.26), which
compares favorably to conventional heterotrophic algae
growth (yp/s 0.10–0.15). The process reduces overall cost
and improves oil quality compared to the strictly photo-
autotrophic process. Future efforts must focus on opti-
mizing lipid yields, improving productivity, and reducing
feedstock cost.
Acknowledgments This research project was supported under the
Department of Energy grant DE-FE-0000888 awarded by National
Energy Technology Laboratory. Additional support was supplied by a
large group of excellent support staff at Phycal in the R&D group who
worked tirelessly to help enable this process.
Conflict of interest All authors were former employees of Phycal
Inc. and paid in part from a grant supplied by the US Department of
Energy.
References
1. Heinrich JM, Niizawa I, Botta FA, Trombert AR, Irazoqui HA
(2012) Analysis and design of photobioreactors for microalgae
production II: experimental validation of a radiation field simu-
lator based on a Monte Carlo algorithm. Photochem Photobiol
88(4):952–960. doi:10.1111/j.1751-1097.2012.01149.x
2. Xu L, Guo C, Wang F, Zheng S, Liu C-Z (2011) A simple and
rapid harvesting method for microalgae by in situ magnetic
separation. Bioresour Technol 102(21):10047–10051. doi:10.
1016/j.biortech.2011.08.021
3. Halim R, Danquah MK, Webley PA (2012) Extraction of oil from
microalgae for biodiesel production: a review. Biotechnol
Advances 30(3):709–732. doi:10.1016/j.biotechadv.2012.01.001
4. Mercer P, Armenta R (2011) Developments in oil extraction from
microalgae. Eur J Lipid Sci Technol. doi:10.1002/ejlt.201000
455
5. Faeth J, Valdez P, Savage P (2013) Fast hydrothermal liquefac-
tion of Nannochloropis sp. to produce biocrude. Energy Fuels
27:1391–1398
6. Onwudili JA, Lea-Langton AR, Ross AB, Williams PT (2013)
Catalytic hydrothermal gasification of algae for hydrogen pro-
duction: composition of reaction products and potential for
nutrient recycling. Bioresour Technol 127:72–80. doi:10.1016/j.
biortech.2012.10.020
7. Rampton R, Zabarenko D (2012) Algae biofuel not sustainable
now, review says. msnbccom
8. Leite GB, Abdelaziz AEM, Hallenbeck PC (2013) Algal biofuels:
challenges and opportunities. Bioresour Technol 145:134–141.
doi:10.1016/j.biortech.2013.02.007
9. Samejima H, Myers J (1958) On the heterotrophic growth of
Chlorella pyrenoidosa. J Gen Microbiol 18(1):107–117
10. Miao X, Wu Q (2006) Biodiesel production from heterotrophic
microalgal oil. Bioresour Technol 97(6):841–846. doi:10.1016/j.
biortech.2005.04.008
11. Azam F, Hemmingsen BB, Volcani BE (1974) Role of silicon in
diatom metabolism. V. Silicic acid transport and metabolism in
the heterotrophic diatom Nitzschia alba. Arch Microbiol
97(2):103–114
12. Lewin J, Hellebust JA (1975) Heterotrophic nutrition of the
marine pennate diatom Navicula pavillardi Hustedt. Can J
Microbiol 21(9):1335–1342
13. Chojnacka K, Noworyta A (2004) Evaluation of Spirulina sp.
growth in photoautotrophic, heterotrophic and mixotrophic
cultures. Enzyme Microb Technol 34(5):461–465. doi:10.1016/j.
enzmictec.2003.12.002
14. Fabregas J, Garcia D, Lamela T, Morales ED, Otero A (1999)
Mixotrophic production of phycoerythrin and exopolysaccharide
by the microalga Porphyridium cruentum. Cryptogamie Algolo-
gie 20(2):89–94
15. Kamjunke N, Tittel J (2009) Mixotrophic algae constrain the loss
of organic carbon by exudation. J Phycol 45(4):807–811. doi:10.
1111/j.1529-8817.2009.00707.x
16. Sforza E, Cipriani R, Morosinotto T, Bertucco A, Giacometti GM
(2012) Excess CO2 supply inhibits mixotrophic growth of
Chlorella protothecoides and Nannochloropsis salina. Bioresour
Technol 104:523–529. doi:10.1016/j.biortech.2011.10.025
17. Hu B, Min M, Zhou W, Du Z, Mohr M, Chen P, Zhu J, Cheng Y,
Liu Y, Ruan R (2012) Enhanced mixotrophic growth of mi-
croalga Chlorella sp. on pretreated swine manure for simulta-
neous biofuel feedstock production and nutrient removal.
Bioresour Technol 126C:71–79. doi:10.1016/j.biortech.2012.09.
031
18. Ogbonna JC, Masui H, Tanaka H (1997) Sequential hetero-
trophic/autotrophic cultivation—an efficient method of producing
Chlorella biomass for health food and animal feed. J Appl Phycol
9:359–366
19. Oyler JR (2008) Two-stage process for producing oil from mic-
roalgae. USA Patent, 8,475,543
20. Sayre R, Pereira S (2010) Molecuar approaches for the optimi-
zation of biofuel productions. USA Patent, 20100317073
21. Wu Q, Xiong W (2009) Method for producing biodiesel from an
alga. USA Patent, 20090298159
22. Krohn BJ, McNeff CV, Yan B, Nowlan D (2011) Production of
algae-based biodiesel using the continuous catalytic Mcgyan
process. Bioresour Technol 102(1):94–100. doi:10.1016/j.bior
tech.2010.05.035
23. Heredia-Arroyo T, Wei W, Hu B (2010) Oil accumulation via
heterotrophic/mixotrophic Chlorella protothecoides. Appl Bio-
chem Biotechnol 162(7):1978–1995. doi:10.1007/s12010-010-
8974-4
24. Xu H, Miao X, Wu Q (2006) High quality biodiesel production
from a microalga Chlorella protothecoides by heterotrophic
growth in fermenters. J Biotechnol 126(4):499–507. doi:10.1016/
j.jbiotec.2006.05.002
25. Huss V, Frank C, Hartmann E, Hirmer M, Kloboucek A, Seidel
B, Wenzeler P, Kessler E (1999) Biochemical taxonomy and
molecular phylogeny of the genus Chlorella sensu lato (Chloro-
phyta). J Phycol 35:587–598
26. Miao X, Wu Q (2004) High yield bio-oil production from fast
pyrolysis by metabolic controlling of Chlorella protothecoides.
J Biotechnol 110(1):85–93. doi:10.1016/j.jbiotec.2004.01.013
27. Konieczny A, Ausubel FM (1993) A procedure for mapping
Arabidopsis mutations using co-dominant ecotype-specific PCR-
based markers. Plant J 4(2):403–410
28. Bligh EG, Dyer WJ (1959) A rapid method of total lipid
extraction and purification. Can J Biochem Physiol
37(8):911–917
29. Greenspan P, Mayer EP, Fowler SD (1985) Nile red: a selective
fluorescent stain for intracellular lipid droplets. J Cell Biol
100(3):965–973
30. Sayre RT (2009) Optimization of biofuel production. USA Patent
US20090181438
31. Santos CA, Nobre B, Lopes da Silva T, Pinheiro HM, Reis A
(2014) Dual-mode cultivation of Chlorella protothecoides
applying inter-reactors gas transfer improves microalgae biodie-
sel production. J Biotechnol 184:74–83. doi:10.1016/j.jbiotec.
2014.05.012
32. Ramos MJ, Fernandez CM, Casas A, Rodriguez L, Perez A
(2009) Influence of fatty acid composition of raw materials on
Bioprocess Biosyst Eng
123
Author's personal copy

biodiesel properties. Bioresour Technol 100(1):261–268. doi:10.
1016/j.biortech.2008.06.039
33. Hu Q, Sommerfeld M, Jarvis E, Ghirardi M, Posewitz M, Seibert
M, Darzins A (2008) Microalgal triacylglycerols as feedstocks for
biofuel production: perspectives and advances. Plant J
54(4):621–639. doi:10.1111/j.1365-313X.2008.03492.x
34. Xiong W, Li X, Xiang J, Wu Q (2008) High-density fermentation
of microalga Chlorella protothecoides in bioreactor for microbio-
diesel production. Appl Microbiol Biotechnol 78(1):29–36.
doi:10.1007/s00253-007-1285-1
35. Hortensteiner S, Chinner J, Matile P, Thomas H, Donnison IS
(2000) Chlorophyll breakdown in Chlorella protothecoides:
characterization of degreening and cloning of degreening-related
genes. Plant Mol Biol 42(3):439–450
36. Ramos Tercero EA, Sforza E, Morandini M, Bertucco A (2014)
Cultivation of Chlorella protothecoides with urban wastewater in
continuous photobioreactor: biomass productivity and nutrient
removal. Appl Biochem Biotechnol 172(3):1470–1485. doi:10.
1007/s12010-013-0629-9
37. Lu Y, Zhai Y, Liu M, Wu Q (2010) Biodiesel production from
algal oil using cassava (Manihot esculenta Crantz) as feedstock.
J Appl Phycol 22(5):573–578. doi:10.1007/s10811-009-9496-8
38. Shetty J, Chotani G, Gand D, Bates D (2007) Cassava as an
alternative feedstock in the production of renewable transporta-
tion fuels. Intl Sugar J 109(1307):663–677
Bioprocess Biosyst Eng
123
Author's personal copy
Top Related
Copyright © 2022 FDOKUMEN

