







Bahasa
Halaman
Hukum


Lower Mississippian (Osagean) brachiopods from the Santiago Formation,
Oaxaca, Mexico: stratigraphic and tectonic implications
D. Navarro-Santillana,*, F. Sour-Tovara, E. Centeno-Garcıab
aMuseo de Paleontologıa de la Facultad de Ciencias, UNAM, Ciudad Universitaria, Mexico City, CP 04510, MexicobInstituto de Geologıa, UNAM, Ciudad Universitaria, Mexico DF, CP 04510, Mexico
Received 31 October 2001; accepted 31 January 2002
Abstract
Four species of spiriferid brachiopods from the Santiago Formation of Oaxaca are described. The species described belong to the orders
Athyridida (Actinoconchus lamellosus ), Spiriferida (Torynifer pseudolineatus ), and Spiriferinida (cf. Syringothyris sp. and Punctospirifer
gigas new species).
The association between A. lamellosus and T. pseudolineatus indicates an Early Mississippian (Osagean) age for the Santiago Formation.
The Carboniferous succession in the area was originally divided into the Santiago Formation (formed by lower and upper members) and the
Ixtaltepec Formation. The studied fauna belongs to the lower member of the Santiago Formation. Because Pennsylvanian brachiopods were
found in strata of the upper member, it is now considered to have more affinity with the Ixtaltepec Formation. Therefore, we propose a change
in the stratigraphy of the area to include the upper member of the Santiago Formation in the Ixtaltepec Formation. This change in the
stratigraphy is supported because a major low-angle normal fault places the former upper member of the Santiago Formation on the lower
member and indicates a tectonic contact between both members of the original Santiago Formation.
Brachiopods from the Santiago Formation are similar to those in the Midcontinent province located in the eastern and central regions of the
United States. The affinity of these Mississippian faunas has important tectonic implications, because Ordovician and Silurian faunas of
Oaxaquia have strong biogeographic affinities with Gondwanan–European provinces. Therefore, there may have been a geographic
connection between southeastern Mexico and North America during Lower Carboniferous times. This could change previous interpretations,
which suggest that the collision of Oaxaquia with North America occurred during Permian time. q 2002 Elsevier Science Ltd. All rights
reserved.
Keywords: Lower Mississippian; Osagean; Brachiopods; Oaxaco; Mexico; Santiago Formation
1. Introduction
The most complete Paleozoic section of south central
Mexico crops out north of Nochixtlan, Oaxaca (Fig. 1),
where it rests unconformably on the Grenvillean-age
Oaxaca complex (Robison and Pantoja-Alor, 1968). This
complex, together with the Huiznopala and Novillo
complexes, has been interpreted as a microcontinent called
Oaxaquia. This cratonic block seems to be allochthonous
with respect to the North America craton throughout its
Precambrian–Early Paleozoic evolution (Stewart et al.,
1993, 1999; Ortega-Gutierrez et al., 1995; Boucot et al.,
1997). The Paleozoic succession of Oaxaca that rests on
southern Oaxaquia is composed of basal Tinu Formation of
Tremadocian age (Robison and Pantoja-Alor, 1968; Orte-
ga-Gutierrez et al., 1995). This is overlain by the Santiago
Formation of Early Mississippian age, followed by the
Ixtaltepec Formation, which contains abundant Lower-
Middle Pennsylvanian invertebrates (Pantoja-Alor, 1970;
Sour-Tovar and Quiroz-Barroso, 1991; Quiroz-Barroso and
Perrilliat, 1997, 1998). The sequence continues into red
beds of the Yododene Formation of unknown age and is
capped by Cretaceous limestone.
The Santiago Formation was formally divided into two
members by Pantoja-Alor (1970), who considered it to be
Early Mississippian due to the presence of the brachiopods
Kitakamithyris and Rotaia. In recent paleontologic studies,
ichnofossils of the genus Scalarituba (Sour-Tovar and
Quiroz-Barroso, 1990, 1991) and the pelecypod ?Streblo-
chondria of Mississippian age have been recognized
(Quiroz-Barroso, 1995). The fossil assemblages of the
0895-9811/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved.
PII: S0 89 5 -9 81 1 (0 2) 00 0 47 -0
Journal of South American Earth Sciences 15 (2002) 327–336
www.elsevier.com/locate/jsames
* Corresponding author.
E-mail address: [email protected] (D. Navarro-
Santillan).

Santiago Formation, their degree of preservation, and the
sedimentological features of the rocks indicate reef and
perireef communities (paleoenviroments) (Quiroz-Barroso,
1995) similar to those studied in other Carboniferous
outcrops of the world, such as the ‘Brachiopod Calcarenite
Community’ in England (McKerrow, 1978). Brachiopods
are the most abundant and diverse group among the Upper
Paleozoic fauna from the Oaxaca area. They are associated
with tabulate corals, sponges, bryozoans, pterioid bivalves,
gastropods, and crinoids. The purpose of this study is to
describe the spiriferid and athyridid brachiopods from the
Santiago Formation and discuss their stratigraphic and
paleobiogeographic significance. New paleontological find-
ings that lead us to redefine the stratigraphy of this unit are
also discussed.
2. Location
The Santiago Formation crops out in several canyons and
washes located between 178290 –178340N lat. and 978050 –
978080W long. (Figs. 1 and 2), all of them in the Nochixtlan
municipality, northwest of Oaxaca City. These sites can be
reached by Federal Highway 190. The type section is
Fig. 1. (a) Geographic distribution of Oaxaquia and its position with respect the North American craton and the Ouachita belt. (b) Location of Santiago
Ixtaltepec and distribution of terranes in southern Mexico (modified from Campa-Uranga and Coney, 1983; Ortega-Gutierrez et al., 1995).
Fig. 2. Geologic map of Paleozoic rocks of the Santiago Ixtaltepec region (modified from Pantoja-Alor, 1970; Centeno-Garcıa and Keppie, 1999) and location
of the fossiliferous localities.
D. Navarro-Santillan et al. / Journal of South American Earth Sciences 15 (2002) 327–336328

located in the Las Pulgas wash bed, 600 m north of the town
of Santiago.
3. Stratigraphy
Paleozoic sedimentary rocks from the Santa Maria Tinu
and Santiago Ixtaltepec localities were described by
Pantoja-Alor (1970), who stated that the Lower Paleozoic
is represented by the Tinu Formation of Cambrian–
Ordovician age (Figs. 2 and 3). This formation discordantly
overlies metamorphic rocks of the Precambrian Oaxaca
complex (Fig. 2) and is made up of interbedded limestone
and black shale containing abundant trilobites of Tremado-
cian age (Robison and Pantoja-Alor, 1968). Upper Paleo-
zoic rocks were grouped in three formations by
Pantoja-Alor (1970): the Santiago Formation of Early
Mississippian age, the Ixtaltepec Formation of Middle
Pennsylvanian age, and the Yododene Formation of Late
Pennsylvanian or Permian age (Fig. 3).
The Santiago Formation (Figs. 2 and 3) originally was
described as a series of marine rocks composed of
limestone, sandstone, siltstone, and shale that overlie, in
angular inconformity, clastic rocks of the Tinu Formation.
However, bedding-parallel shearing indicates that
this contact is a normal low-angle detachment fault
(Centeno-Garcıa and Keppie, 1999). Pantoja-Alor (1970)
suggested that the Santiago Formation transitionally
changes into overlying clastics of the Ixtaltepec Formation.
On the basis of lithologic differences, the Santiago
Formation was divided into two easily distinguishable
members (Pantoja-Alor, 1970): a lower calcareous member,
with interbedded thin strata of shale and marl in its upper
part, and an upper lutitic member, with interbedded strata of
siltstone, sandstone, and limestone.
As a part of this work, we measured the type section of
the Santiago Formation in the Las Pulgas canyon, 500 m
north of Santiago Ixtaltepec, and we reexamined the
stratigraphy and fossil content of this unit. We obtained a
thickness of 157 m instead of the 192 m previously
proposed (Pantoja-Alor, 1970). The lower 67 m belong to
the lower calcareous member, described by Pantoja-Alor
(1970). No Mississippian fossils were found in the
remaining 90 m of the upper lutitic member of the Santiago
Formation, but some specimens of the brachiopods
Rugosochonetes granulifer, Composita ovata, and Neospir-
ifer sp. were, which indicates a Pennsylvanian age for this
unit.
On the basis of the fossil content, we propose to reduce
the thickness of the Santiago Formation (Fig. 3) and extend
the lower limit of the Ixtaltepec Formation to include the
upper member of the Santiago Formation. We also suggest
Fig. 3. (a) Stratigraphic column proposed by Pantoja-Alor (1970) and (b) stratigraphic column proposed herein (showing changes made to the stratigraphy of
the Santiago and Ixtaltepec formations).
D. Navarro-Santillan et al. / Journal of South American Earth Sciences 15 (2002) 327–336 329

avoiding the use of the member divisions of the Santiago
Formation. The revised Santiago Formation, as defined
herein, is characterized in its lower part by limestone
containing tabulate coral banks associated with brachiopods
and demosponges belonging to the order Lithistida. These
communities are similar to those found in several ‘Osagean’
outcrops in France (D. Vachard, pers. comm.). The reef
banks are interbedded with calcareous sandstone nearly
12 m thick that contains the same fossil association. The
upper part of the unit is approximately 55 m thick and
contains most of the brachiopods described herein.
The contact between the Santiago and Ixtaltepec
Formations, as defined here, is a shear zone that strongly
deforms the shale and calcareous sandstone of the upper part
of the Santiago Formation and tectonically places the
limestone or siltstone of the Ixtaltepec Formation over the
sheared Santiago Formation. This slide is similar to that
observed at the Tinu–Santiago contact. Both structures
represent top-to-the-north sliding that places younger units
over older as a result of extensional tectonics (Centeno-
Garcıa and Keppie, 1999). The Ixtaltepec Formation
consists of interbedded shale and sandstone with some
limestone beds and contains abundant Lower-Middle
Pennsylvanian invertebrates, which indicate a shallow
marine environment (Sour-Tovar and Quiroz-Barroso,
1991; Quiroz-Barroso and Perrilliat, 1997, 1998).
4. Age and paleobiogeographic remarks
Brachiopods described herein were mainly collected
from the middle and upper parts of the lower member of the
Santiago Formation. Of the four species described here,
Actinoconchus lamellosus and Torynifer pseudolineatus
suggest an Osagean (Early Mississippian) age. These rocks
can be correlated with the outcrops of the Vicente Guerrero
Formation near Ciudad Victoria, Tamaulipas (Fig. 1)
(Carrillo-Bravo, 1959; Stewart et al., 1999) and with the
lower part of the Patlanoaya Formation, southeast of Izucar
de Matamoros, Puebla (Villasenor-Martınez et al., 1987).
Several other outcrops in North America yield similar
species, including the Escabrosa limestone of Bisbee
Quadrangle in Arizona (Girty, 1904), the Keokuk limestone
of the Mississippi River Valley in Illinois (Weller, 1914),
the Boone limestone of St. Joe, Arkansas (Girty, 1915), and
the Cuyahoga (Root et al., 1961) and Logan (Hyde, 1953;
Root et al., 1961) Formations in Ohio, as well as the
Chappel Formation in Texas (Plummer, 1950). The
association of A. lamellosus and T. pseudolineatus has
been found in all these units, except for the Patlanoaya, and
all have been dated as Osagean. The localities from the
United States are part of the Midcontinent province, except
for the Escabrosa limestone, which belongs to the
Appalachian province.
The finding of the A. lamellosus–T. pseudolineatus
association changes the paleogeographic and tectonic
interpretations of the geographic configuration of Mexico
during the Upper Paleozoic, as well as the possible origin of
its tectonostratigraphic terranes. During Early Mississippian
time, the lithospheric pieces where the Paleozoic outcrops
of Oaxaca are located had a latitudinal position very similar
to that of the epicontinental sea represented by the
Midcontinent province. Pennsylvanian fossils studied from
Oaxaca are also similar to those from the east central
regions of the United States (Sour-Tovar, 1994; Quiroz-
Barroso, 1995; Quiroz-Barroso and Perilliat, 1997, 1998).
Thus, we can propose that, during the Carboniferous, the
Midcontinent province may have extended to the south,
including the localities of Oaxaquia. Because marine
communities in Oaxaca have a faunistic association very
similar to those of the North American Midcontinent,
Oaxaquia is considered allochthonous with respect to North
America, at least until the Silurian (Stewart et al., 1993).
Some authors have proposed that its position in Permian
times was still allochthonous with respect to North America
because of the overlap of South America with southern
Mexico and Central America when Pangea is reconstructed
(e.g. Bullard et al., 1965). The similarity of Early
Mississippian faunas in Oaxaquia to those of south central
North America suggests a close approach of the Oaxaquia
block to Laurentia by this time. Reconstruction of Mexico
along the western margin of Pangea, as proposed by Pindell
(1985), places a collisional belt along Oaxaquia during the
Late Paleozoic. However, our data suggest that the
Oaxaquia block was either accreted before the Early
Mississippian or added to North America by other
mechanisms (transform faults?). Moreover, because the
faunal province of Oaxaca and the Midcontinent is different
from that of the Ouachitas, our data suggest that the
Oaxaquia block may not have collided with the Ouachita
belt.
5. Systematic paleontology
Fossil material studied here is deposited in the Museo de
Paleontologıa, Facultad de Ciencias, Universidad Nacional
Autonoma de Mexico, as part of the collection FCMP/E1.
5.1. A. lamellosus (Leveille, 1835)
Phylum Brachiopoda (Dumeril, 1806)
Class Articulata (Huxley, 1869)
Order Athyridida Boucot (Johnson and Staton, 1964)
Suborder Athyrididina Boucot (Johnson and Staton, 1964)
Superfamily Athyridoidea (Davidson, 1881)
Family Athyrididae (Davidson, 1881)
Subfamily Athyridinae (Davidson, 1881)
Genus Actinoconchus (M’Coy, 1844)
Type species Spirifera planosulcata (Phillips, 1836)
Actinoconshus Lamellosus (Leveille, 1835) Figs. 4.1–4.5.
D. Navarro-Santillan et al. / Journal of South American Earth Sciences 15 (2002) 327–336330

5.1.1. Synonymy
Spirifer lamellosus (Leveille, 1835, p. 39, Figs. 21–23).
Athyris lamellosus (Leveille) (Davidson, 1857, pp.
79–80, plate 16, Fig. 1 and plate 17, Fig. 6).
Athyris lamellosa (Leveille) (Davidson, 1859, plate 16,
Fig. 1a,b).
Athyris (Actinoconchus ) cf. lamellosa (Leveille) (Sibly,
1906, p. 374, plate XXXII, Fig. 1a,b).
Actinoconchus cf. lamellosa (Leveille) (Minato, 1951,
pp. 380–381, plate 1, Fig. 6a,b).
Actinoconchus lamellosa (Leveille) (Minato, 1952, p.
173, plate 11, Fig. 6).
A. lamellosus (Leveille) (Brunton, 1980, pp. 225–226,
Figs. 16 and 17).
5.1.2. Description
Shell is up to 41.1 mm wide and 28.5 mm long;
transversely subelliptical in outline, moderately and sub-
equally biconvex, short hinge line; open delthyrium;
rounded auricles; and uniplicate anterior commisure.
Pedicle valve is slightly flattened along midline in the
posterior half of the shell. Flattening changes gradually to a
shallow, poorly defined mesial sulcus in anterior part;
anterior margin in this part of the shell sometimes folds over
opposite valve to produce a mesial linguiform extension of
moderate size. Posterior tip of the apex is small, curved, and
in close contact with the umbo of the brachial valve. Hinge
teeth are internally prominent, curved at the top, and
supported by very short dental plates; between plates, there
is a deep, transversely striated pedicle cavity.
Brachial valve is slightly flattened in the anterior part
along mesial line; this flattening gradually transforms the
anterior region into a shallow mesial fold that is very
prominent near the anterior margin.
Surfaces of both valves are marked by concentric,
parallel, lamelliform extensions, often mostly destroyed,
leaving only their bases. Internally, dental sockets are wide
and deep, and crura are long and convergent with primary
lamellae curving abruptly at their origin.
5.1.3. Discussion
A. lamellosus is easily recognized by its size and shell
form; the length of the hinge line is shorter than the widest
part of shell. It has rounded cardinal extremities and is
mainly characterized by strong and concentric subparallel
lamellar extensions separated by a distance of 3–5 mm.
The studied material included sixteen specimens: five
juvenile forms in different developmental stages, five
internal molds of pedicle valves (three fragmented and
two nearly complete), one internal mold of a brachial valve,
one external impression of a pedicle valve, and four external
impressions of brachial valves.
The specimen numbers are as follows: FCMP/E1-767,
E1783, E1-821a–b, E1-1462, E1-1882, E1-1883a–b, E1-
1884, E1-894, E1-1895, E1-1896, E1-1897, E1-1898, E1-
1899, E1-1900, E1-1901 and E1-1938.
5.2. T. pseudolineatus (Hall, 1858)
Order Spiriferida (Waagen, 1883)
Suborder Delthyridina (Ivanova, 1972)
Superfamily Reticularioidea (Waagen, 1883)
Family Elythidae (Frederiks, 1924)
Subfamily Toryniferinae (Carter, 1994)
Genus Torynifer (Hall and Clarke, 1894)
Type species Spirifer pseudolineatus (Hall, 1858) from the
Lower Mississippian of the United States.
Torynifer Pseudolineatus (Hall, 1858). Fig. 4.6–4.9.
5.2.1. Synonymy
Spirifer pseudolineatus (Hall, 1858, p. 645, plate 20, Fig.
4).
Spirifera pseudolineata (Hall, 1883, plate 36, Figs.
28–30).
Reticularia pseudolineatus (Hall) (Beede, 1906, p.
1317–1318, plate 21, Fig. 5 and plate 20, Fig. 6a).
Reticularia pseudolineata (Hall) (Grabau and Shimer,
1909, p. 339, Fig. 433).
Torynifer pseudolineata (Hall) (Cooper, 1944, p. 327,
plate 126, Figs. 4–8, 16, and 17).
T. pseudolineatus (Hall) (Ivanova, 1959, p. 54, text Fig.
6Z).
5.2.2. Description
The shell is large, with both valves strongly convex,
particularly in posterior region; transversely subelliptical in
outline; lateral margins symmetrically rounded, as are
cardinal extremities. Length of hinge line is about two-
thirds of the shell width.
Pedicle valve surface curves abruptly from umbonal
region to cardinal margin. Mesial sulcus is ill-defined,
shallow at its origination in umbonal region, and of
moderate width and obsolete at peak. Umbo is very small,
sharp, curved, and projecting beyond the brachial valve;
lateral margins are not well defined; surface is vertically
striated; cardinal area is very small and arched; delthyrium
is triangular and open; hinge teeth are supported by a pair of
strong dental plates that extend about one-fourth of valve
length, diverging at an angle of 308–408; between the dental
lamellae, a long medial septum extends half, or slightly
more, of the valve length, and reaches two or three times the
length of the dental plates; interior covered by fine radial
lines, which number 2–5 per mm.
Brachial valve is less convex than pedicle valve; surface
is abruptly curved from umbonal region and smooth in
anterior and lateral parts, where the shell is compressed
through cardinal extremities; mesial fold usually impercep-
tible or obsolete, except near anterior margin, where it is
D. Navarro-Santillan et al. / Journal of South American Earth Sciences 15 (2002) 327–336 331
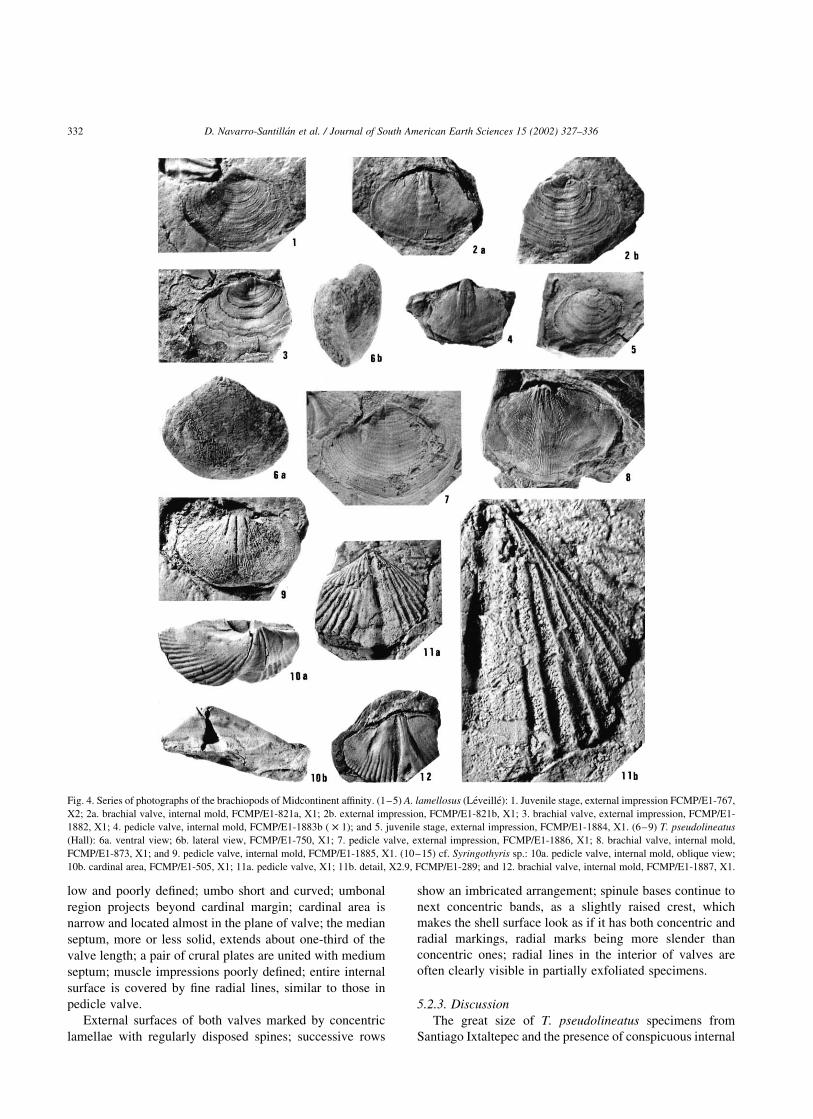
low and poorly defined; umbo short and curved; umbonal
region projects beyond cardinal margin; cardinal area is
narrow and located almost in the plane of valve; the median
septum, more or less solid, extends about one-third of the
valve length; a pair of crural plates are united with medium
septum; muscle impressions poorly defined; entire internal
surface is covered by fine radial lines, similar to those in
pedicle valve.
External surfaces of both valves marked by concentric
lamellae with regularly disposed spines; successive rows
show an imbricated arrangement; spinule bases continue to
next concentric bands, as a slightly raised crest, which
makes the shell surface look as if it has both concentric and
radial markings, radial marks being more slender than
concentric ones; radial lines in the interior of valves are
often clearly visible in partially exfoliated specimens.
5.2.3. Discussion
The great size of T. pseudolineatus specimens from
Santiago Ixtaltepec and the presence of conspicuous internal
Fig. 4. Series of photographs of the brachiopods of Midcontinent affinity. (1–5) A. lamellosus (Leveille): 1. Juvenile stage, external impression FCMP/E1-767,
X2; 2a. brachial valve, internal mold, FCMP/E1-821a, X1; 2b. external impression, FCMP/E1-821b, X1; 3. brachial valve, external impression, FCMP/E1-
1882, X1; 4. pedicle valve, internal mold, FCMP/E1-1883b ( £ 1); and 5. juvenile stage, external impression, FCMP/E1-1884, X1. (6–9) T. pseudolineatus
(Hall): 6a. ventral view; 6b. lateral view, FCMP/E1-750, X1; 7. pedicle valve, external impression, FCMP/E1-1886, X1; 8. brachial valve, internal mold,
FCMP/E1-873, X1; and 9. pedicle valve, internal mold, FCMP/E1-1885, X1. (10–15) cf. Syringothyris sp.: 10a. pedicle valve, internal mold, oblique view;
10b. cardinal area, FCMP/E1-505, X1; 11a. pedicle valve, X1; 11b. detail, X2.9, FCMP/E1-289; and 12. brachial valve, internal mold, FCMP/E1-1887, X1.
D. Navarro-Santillan et al. / Journal of South American Earth Sciences 15 (2002) 327–336332
radial lines distinguish them clearly from T. setigera, T.
salemensis (Weller, 1914), T. montanus (Shaw, 1962), T.
eufastigium (Carter, 1987), and T. spinosus (Stainbrook,
1947), all Mississippian species described from North
America. Moreover, in T. setigera and T. eufastigium, the
mesial sinus and fold are deeper, and apparently, T.
salemensis has no internal radial ribs. There are no other
comparable species of Torynifer.
The studied material included 27 specimens: two whole
specimens with articulated valves; two external impressions
and three internal molds of pedicle valves; eight internal
molds of brachial valves; 11 fragments of internal molds of
pedicle valves; and one fragment of an external impression.
The specimen numbers are as follows: FCMP/E1-263,
E1-282, E1-731, E1-743, E1-750, E1-769, E1-780, E1-794,
E1-796, E1-838, E1-841, E1-849, E1-864, E1-868, E1-873,
E1-1885, E1-1886, E1-1902–1910, and E1-1911.
5.3. Cf. Syringothyris sp
Order Spiriferinida (Ivanova, 1972)
Suborder Spiriferinidina (Ivanova, 1972)
Superfamily Syringothyridoidea (Frederiks, 1926)
Family Syringothyrididae (Frederiks, 1926)
Subfamily Syringothyridinae (Frederiks, 1926)
Genus cf. Syringothyris (Winchell, 1863)
Type species Syringothyris typa (Winchell, 1863) from the
Lower Mississippian of Iowa, USA. cf. Syringothyris sp.
Figs. 4.10–4.12 and 5.1–5.3.
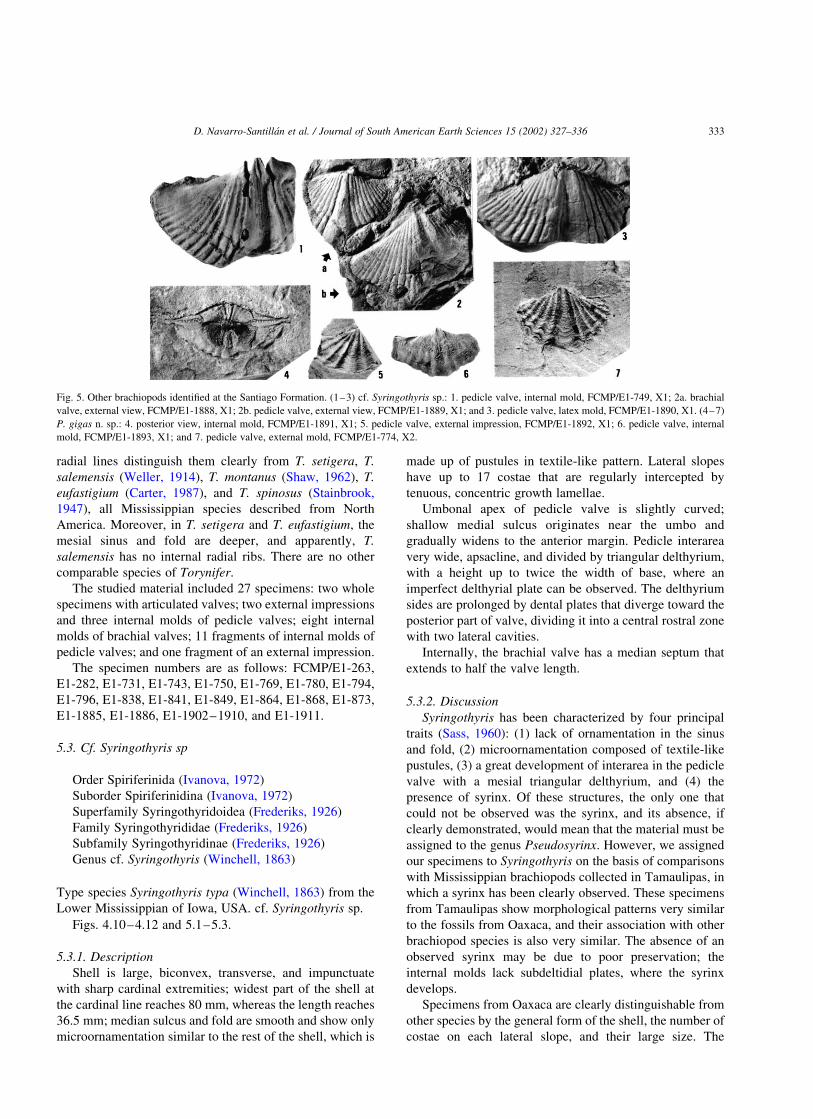
5.3.1. Description
Shell is large, biconvex, transverse, and impunctuate
with sharp cardinal extremities; widest part of the shell at
the cardinal line reaches 80 mm, whereas the length reaches
36.5 mm; median sulcus and fold are smooth and show only
microornamentation similar to the rest of the shell, which is
made up of pustules in textile-like pattern. Lateral slopes
have up to 17 costae that are regularly intercepted by
tenuous, concentric growth lamellae.
Umbonal apex of pedicle valve is slightly curved;
shallow medial sulcus originates near the umbo and
gradually widens to the anterior margin. Pedicle interarea
very wide, apsacline, and divided by triangular delthyrium,
with a height up to twice the width of base, where an
imperfect delthyrial plate can be observed. The delthyrium
sides are prolonged by dental plates that diverge toward the
posterior part of valve, dividing it into a central rostral zone
with two lateral cavities.
Internally, the brachial valve has a median septum that
extends to half the valve length.
5.3.2. Discussion
Syringothyris has been characterized by four principal
traits (Sass, 1960): (1) lack of ornamentation in the sinus
and fold, (2) microornamentation composed of textile-like
pustules, (3) a great development of interarea in the pedicle
valve with a mesial triangular delthyrium, and (4) the
presence of syrinx. Of these structures, the only one that
could not be observed was the syrinx, and its absence, if
clearly demonstrated, would mean that the material must be
assigned to the genus Pseudosyrinx. However, we assigned
our specimens to Syringothyris on the basis of comparisons
with Mississippian brachiopods collected in Tamaulipas, in
which a syrinx has been clearly observed. These specimens
from Tamaulipas show morphological patterns very similar
to the fossils from Oaxaca, and their association with other
brachiopod species is also very similar. The absence of an
observed syrinx may be due to poor preservation; the
internal molds lack subdeltidial plates, where the syrinx
develops.
Specimens from Oaxaca are clearly distinguishable from
other species by the general form of the shell, the number of
costae on each lateral slope, and their large size. The
Fig. 5. Other brachiopods identified at the Santiago Formation. (1–3) cf. Syringothyris sp.: 1. pedicle valve, internal mold, FCMP/E1-749, X1; 2a. brachial
valve, external view, FCMP/E1-1888, X1; 2b. pedicle valve, external view, FCMP/E1-1889, X1; and 3. pedicle valve, latex mold, FCMP/E1-1890, X1. (4–7)
P. gigas n. sp.: 4. posterior view, internal mold, FCMP/E1-1891, X1; 5. pedicle valve, external impression, FCMP/E1-1892, X1; 6. pedicle valve, internal
mold, FCMP/E1-1893, X1; and 7. pedicle valve, external mold, FCMP/E1-774, X2.
D. Navarro-Santillan et al. / Journal of South American Earth Sciences 15 (2002) 327–336 333

exception is S. transversa (Minato, 1951), from which they
differ only in the width–length ratio of the shell, which is
smaller in S. transversa. These comparisons suggest that the
material of cf. Syringothyris sp. may represent a new
species. However, because of the generic imprecision, it
was decided not to name it (Figs. 4 and 5).
The studied material included pedicle valves (seven
external molds, nine internal molds, and one external
impression) and brachial valves (two external molds and
one external impression). All specimens show different
development stages, and most are incomplete.
The specimen numbers are as follows: FCMP/E1-289,
E1-505, E1-746, E1-749, E1-786, E1-788, E1-798, E1-800,
E1-803, E1-804, E1-808, E1-844, E1-857, E1-865, E1-881,
E1-1881, E1-1887–1890, E1-1912–1922, and E1-1923.
5.4. Punctospirifer gigas
Superfamily Pennospiriferinoidea (Dagys, 1972)
Family Punctospiriferidae (Waterhouse, 1975)
Genus Punctospirifer (North, 1920)
Type species Punctospirifer scabricosta (North, 1920) from
the Lower Carboniferous (Visean) of England.
Punctospirifer Gigas (Sour-Tovar and Navarro-Santillan
new species.)
Fig. 5.4–5.7.
5.4.1. Diagnosis
Shell is large for this genus, with dimensions up to
40.4 mm wide and 19.4 mm long for brachial valve and
37.3 mm wide and 18.9 mm long in pedicle valve; five ribs
on each side of sinus and fold, an average of six lamellar
growth lines for each five mm; shells lack plications in sinus
and a furrow on fold.
5.4.2. Description
Spiriferoid shell, clearly biconvex, is small in size but
large for genus; semicircular in outline; nearly twice as wide
as long; maximum width at hinge line; cardinal extremities
slightly rounded and mucronate auricles. Commissure
sulcate; sulcus of pedicle valve and fold of brachial valve
are well defined, moderately developed, and rounded;
lateral slopes of both valves with up to five costae and
five furrows on each side of fold and sulcus. There are up to
six radial growth lines for each 5 mm, perpendicular to the
costation, and crossing the fold and sulcus. This gives the
shell an imbricated appearance; microornamentation also
includes spines and granulations; shell punctate.
Pedicle valve is convex; narrow and well-defined
interarea; apex small and slightly curved; medial sulcus,
which originates at the apex, is shallow, rounded, narrow in
posterior region, and gradually widens to anterior region.
Internally, a pair of slightly divergent dental plates; middle
septum well developed and thick at base.
Brachial valve slightly less convex than pedicle valve;
small apex at umbo; mesial fold originates at umbo, striated,
rounded, and does not surpass the rest of shell; middle
septum extends half the valve length.
At the hinge line, there is a series of teeth forming the
hinge; three for each mm.
5.4.3. Discussion
P. gigas was compared with P. scabricosta (North, 1920;
Campbell, 1959), P. subtextus (Girty, 1915), P. kentuck-
yensis (Dunbar and Condra, 1932; Elias, 1957), P.
transversus (Elias, 1957; Lane, 1962; Easton, 1962;
Sour-Tovar, 1994), P. sulcifer, P. globosa (Sanders,
1958), P. amblys (Cvancara, 1958), P. solidirostris (Weller,
1914; Carter, 1972, 1987), P. campestris (Lane, 1962), P.
monroensis (Carter, 1990), P. (Reticulariina ) spinosa
(Easton, 1962), P. depressus (Root et al., 1961), P. cf.
similis (Hyde, 1953), and P. acutus (Carter, 1968) and
differs from all of them mainly by its larger size, its five
costae on each lateral slope (though some specimens of P.
globosa, P. depressus, and P. (Reticulariina ) spinosa have
five plications laterally), and the smaller number of growth
lamellae per millimeter. It is also distinguished from P.
globosa, P. sulcifer, and P. solidirostris, which have a small
plication in the sulcus and a small furrow in the fold.
The studied material included nine external impressions
and seven internal molds of pedicle valves, six external
impressions and three internal molds of brachial valves, and
one specimen that shows the posterior part of an internal
mold of one specimen.
5.4.4. Etymology
Gigas is a Latin word of Greek origin that refers to the
large size of individuals in this taxonomic unit.
The syntypes are as follows: FCMP/ E1-774, E1-1891,
E1-1892a, and E1-1893
The specimen numbers are as follows: FCMP/E1-72, E1-
301, E1-748, E1-784, E1-797, E1-861, E1-862, E1-885, E1-
897, E1-1892b, E1-1924–1936, and E1-1937.
Acknowledgments
The authors thank to H. Hernandez-Campos for photo-
graphic work and L. Chavez-Garcıa for his technical
assistance. They also express their appreciation to J.T.
Dutro Jr., S.A. Quiroz-Barroso, R.B. Blodgett, and F.J.
Vega-Vera for their critical reviews of the manuscript.
References
Beede, J.W., 1906. Fauna of the Salem limestone of Indiana. In: Cummings,
E.R., Beede, J.W. (Eds.), Indiana Department of Geology and Natural
Resources, 30th Annual report (for 1905), pp. 1189–1486.
Boucot, A.J., Blodgett, R.B., Stewart, J.H., 1997. European province late
Silurian brachiopods from the Ciudad Victoria area, Tamaulipas,
D. Navarro-Santillan et al. / Journal of South American Earth Sciences 15 (2002) 327–336334

northeastern Mexico. In: Klapper, G., Murphy, M.A., Talent, J.A.
(Eds.), Paleozoic Sequence Stratigraphy, Biostratigraphy: Studies in
Honor of J. Granville (‘Jess’) Johnson, Geological Society of America,
Special Paper, 321., pp. 273–293.
Boucot, A.J., Johnson, J.G., Staton, R.D., 1964. On some Atrypoid,
Retzioid, and Athyridoid brachiopoda. Journal of Paleontology 38 (5),
805–822.
Brunton, C.H.C., 1980. Type specimens of some upper Palaeozoic
Athyridide brachiopods. Bulletin of the British Museum (Natural
History) Geology 34 (4), 219–234.
Bullard, E.C., Everett, J.E., Smith, A.G., 1965. The fit of the continents
around the Atlantic. Philosophical Transactions of the Royal Society of
London, Series A 258, 41–51.
Campa-Uranga, M.F., Coney, P.J., 1983. Tectonostratigraphic terranes and
mineral resource distribution of Mexico. Canadian Journal of Earth
Sciences 20, 1040–1051.
Campbell, K.S.W., 1959. The type species of three upper Palaeozoic
punctate spiriferoids. Palaeontology 1 (4), 351–363.
Carrillo-Bravo, J., 1959. Notas sobre el Paleozoico de la region de Ciudad
Victoria, Tamaulipas. Boletın de la Asociacion Mexicana de Geologos
Petroleros 11, 1–98.
Carter, J.L., 1968. New genera and species of Early Mississippian
brachiopods from the Burlington limestone. Journal of Paleontology
42 (5), 1140–1152.
Carter, J.L., 1972. Early Mississippian brachiopods from the Gilmore city
limestone of Iowa. Journal of Paleontology 46 (4), 473–491.
Carter, J.L., 1987. Lower Carboniferous brachiopods from the Banff
Formation of Western Alberta. Geological Survey of Canada, Bulletin
378, 183.
Carter, J.L., 1990. New brachiopods (Brachiopoda: Articulata) from the
Late Osagean of the Upper Mississippi Valley. Annals of Carnegie
Museum 59 (3), 219–247.
Carter, J.L., 1994. Subfamily Toryniferinae. In: Carter, J.L., Johnson, J.G.,
Gourvennec, R., Hong-Fei, H. (Eds.), A Revised Classification of the
Spiriferid Brachiopods, Annals of Carnegie Museum, 63(4)., pp.
327–374.
Centeno-Garcıa, E., Keppie, J.D., 1999. Latest Paleozoic-Early Mesozoic
structures in the central Oaxaca terrane of Southern Mexico:
deformation near a triple junction. Tectonophysics 301, 231–242.
Cooper, G.A., 1944. Phylum Brachiopoda. In: Shimer, H.W., Shrock, R.R.
(Eds.), Index Fossils of North America, pp. 277–365.
Cvancara, A.M., 1958. Invertebrates fossils from the lower Carboniferous
of New South Wales. Journal of Paleontology 32 (5), 846–888.
Dagys, A.S., 1972. Yavleniya metakhoreza sredi triasovykh spiriferinid,
Akademia Nauk SSSR, Sibirskoye Otdelenie, Trudy Instituta Geologii i
Geofiziki, 111., 36 pp..
Davidson, T., 1857. Notes sur les genres Athyris ( ¼ Spirigera ),
Camarophoria, Orthisina et Strophalosia des terrains permiens d’
Anglaterre. Bulletin of Societe Linneenne de Normandie 2, 11–24.
Davidson, T., 1859. A Monograph of the British fossil Brachiopoda, 2, Part
5, no. 2: The Fossil Brachiopoda–Carboniferous, Monograph, 11.
Palaeontographical Society, London, pp. 49–80.
Davidson, T., 1881. On genera and species of spiral-bearing brachiopoda,
from specimens developed by the Rev. Norman Glass: with notes on the
results obtained by Mr George Maw from extensive washings of the
Wenlock and Ludlow Shales of Shropsire. Geological Magazine 8 (1),
1–13.
Dumeril, A.M.C., 1806. Zoologie analytique ou methode naturelle de
classification des animaux, XXIV Allais, Paris, 344 pp..
Dunbar, C.O., Condra, G.E., 1932. Brachiopoda of the Pennsylvanian
System in Nebraska, Nebraska Geological Survey, Bulletin 5, Serie 2,
377 pp..
Easton, W.H., 1962. Carboniferous Formations and Faunas of Central
Montana, United States Geological Survey, Professional Paper 348, 126
pp..
Elias, M.K., 1957. Late Mississippian Fauna from the Redoak Hollow
Formation of Southern Oklahoma, Part 2. Brachiopoda. Journal of
Paleontology 31 (3), 487–527.
Frederiks, G., 1926. Tablitsa dlya opredeleniya rodov semeystva Spirifer-
idae King. Akademia Nauk SSSR, Izvestiya, Ser. 6, 20, pp. 393–423.
Girty, G.H., 1904. Note on the Carboniferous fossils. In: Ransome, F.L.,
(Ed.), The Geology and Ore deposits of the Bisbee Quadrangle,
Arizona, United States Geological Survey, Professional Paper, 21., pp.
46–54.
Girty, G.H., 1915. Faunas of the Boone Limestone at St. Joe, Arkansas,
United States Geological Survey, Bulletin 598, 50 pp..
Grabau, A.W., Shimer, W.H., 1909. North American Index Fossils.
Invertebrates 1, A.G. Seiler, New York, 853 pp..
Hall, J., 1858. Palaeontology of Iowa. In: Hall, J., Whitney, J.D. (Eds.),
Report on the Geological Survey of the State of Iowa; embracing the
results of investigations made during portion of the year 1855–1857,
Vol. 1(2)., pp. 473–724.
Hall, J., 1883. Brachiopoda from the Niagara and Upper Helderberg Groups
of Indiana. Indiana Department of Geology and Natural History, 12th
Annual Report for 1882, pp. 319–375.
Hall, J., Clarke, J.M., 1894. An Introduction to the Study of the
Brachiopoda: 13th Annual Report New York State Geologist for the
year 1893. Palaeontology, 751–943.Part 2.
Huxley, T.H., 1869. An introduction to the classification of animals, John
Churchill and Sons, London, 147 pp..
Hyde, J.E., 1953. In: Fisher, M.M., (Ed.), The Mississippian formations of
central and southern Ohio, Geological Survey of Ohio Bulletin, 51., p.
355.
Ivanova, E.A., 1959. K sistematike i evoliutsiy spiriferid (The systematics
and evolution of Spririferidae (Brachiopoda).). Palaeontologicheskii
Zhurnal 4, 47–64.
Ivanova, E.A., 1972. Osnovnye zakonomernosti evoliutsii spiriferid
(Brachiopoda). (Main features of Spiriferid evolution (Brachiopoda).).
Paleontologicheskii Zhurnal 3, 28–42.
Lane, B.O., 1962. The fauna of the Ely Group in the Illipah Area of Nevada.
Journal of Paleontology 36 (5), 888–911.
Leveille, C., 1835. Apercu geologique de quelques localites tres riches en
coquilles sur les frontieres de France et de Bellgique, Societe
Geologique de France, Memories, 1st ser., 2., pp. 29–49.
McKerrow, W.S., 1978. The Ecology of Fossils, MIT Press, Cambridge,
MA, 383 pp..
M’Coy, F., 1844. A synopsis of the characters of the Carboniferous
limestone fossils of Ireland, University Press, Dublin, 207.
Minato, M., 1951. On the lower Carboniferous fossils of the Kitakami
Massif, northeast Honsyu, Japan. Hokkaido University, Faculty of
Science (Geology and Mineralogy), Journal, Serie IV VII (4), 355–382.
Minato, M., 1952. A further note on the Lower Carboniferous fossils of the
Kitakami Mountainland, Northeast Japan. Hokkaido University,
Faculty of Science (Geology and Mineralogy), Journal, Serie IV VIII
(2), 136–174.
North, F.J., 1920. On Syringothyris Winchell and certain Carboniferous
Brachiopoda referred to Spiriferina d’Orbigny. Geological Society of
London, Quarterly Journal 76 (2), 162–227.
Ortega-Gutierrez, F., Ruiz, F.J., Centeno-Garcia, E., 1995. Oaxaquia, a
Proterozoic microcontinent accreted to North America during the Late
Paleozoic. Geology 23 (12), 1127–1130.
Pantoja-Alor, J., 1970. Rocas sedimentarias paleozoicas de la region
centro-septentrional de Oaxaca. Sociedad Geologica Mexicana. Libro
Guıa de la Excursion Mexico-Oaxaca, 67–84.
Phillips, J., 1836. The Mountain Limestone district. Illustrations of the
Geology of Yorkshire: Part 2, John Murray, London, 253 pp..
Pindell, J.L., 1985. Alleghanian reconstruction and subsequent evolution of
the Gulf of Mexico, Bahamas and Proto-Caribbean. Tectonics 4 (1),
1–39.
Plummer, F.B., 1950. The Carboniferous rocks of the Llano region of
Central Texas. University of Texas, Publication no. 4329, 170 p..
Quiroz-Barroso, S.A., 1995. Bivalvos del Carbonıfero de Nochixtlan,
D. Navarro-Santillan et al. / Journal of South American Earth Sciences 15 (2002) 327–336 335

Oaxaca. Universidad Nacional Autonoma de Mexico, Facultad de
Ciencias, Tesis Doctoral, 86 pp.
Quiroz-Barroso, S.A., Perrilliat, M.C., 1997. Pennsylvanian Nuculoids
(Bivalvia) from the Ixtaltepec Formation, Oaxaca, Mexico. Journal of
Paleontology 71 (3), 400–407.
Quiroz-Barroso, S.A., Perrilliat, M.C., 1998. Pennsylvanian Bivalves from
the Ixtaltepec Formation, Mexico. Journal of Paleontology 72 (6),
1011–1024.
Robison, R.A., Pantoja-Alor, J., 1968. Tremadocian trilobites from the
Nochixtlan Region, Oaxaca, Mexico. Journal of Paleontology 42 (3),
767–800.
Root, S.I., Rodriguez, J., Forsyth, J.L., 1961. Geology of Knox County.
Geological Survey of Ohio, Bulletin 59, 232 p..
Sanders, J.E., 1958. Brachiopoda and Pelecypoda. In: Easton, W.H.,
Sanders, J.E., Knight, J.B., Miller, A.K. (Eds.), Mississippian fauna in
northwestern Sonora, Mexico, Smithsonian Miscellaneous Collections,
119(3)., p. 87.
Sass, D.B., 1960. Some aspects of the paleontology, stratigraphy and
sedimentation of the Corry Sandstone of the northwestern Pennsylva-
nia. Bulletin of American Paleontology 41 (192), 251–381.
Shaw, A.B., 1962. Rhynchonellid brachiopods and a Torynifer from the
Madison Group (Mississippian). Journal of Paleontology 36 (4),
630–637.
Sibly, T.F., 1906. On the Carboniferous Limestone (Avonian) of the
Mendip Area (Somerset), with special reference to the palaeontological
sequence. Geological Society of London, Quaterly Journal 62,
324–380.
Sour-Tovar, F., 1994. Braquiopodos Pensilvanicos del Area de Santiago
Ixtaltepec, Municipio de Nochixtlan, Oaxaca. Universidad Nacional
Autonoma de Mexico, Facultad de Ciencias. Master in Sciences
Dissertation, 55 pp.
Sour-Tovar, F., Quiroz-Barroso, S.A., 1990. Siguiendo la Huella.
Icnofosiles de Mexico. Informacion Cientıfica y Tecnologica 12
(171), 27–32.
Sour-Tovar, F., Quiroz-Barroso, S.A., 1991. Icnofosiles paleozoicos de
Nochixtlan, Oaxaca. Memorias del III Congreso Nacional de
Paleontologıa, Resumenes, Sociedad Mexicana de Paleontologıa, 131
pp.
Stainbrook, M.A., 1947. Brachiopoda of the Percha Shale of New Mexico
and Arizona. Journal of Paleontology 21 (4), 297–328.
Stewart, J.H., Blodgett, R.B., Boucot, A.J., Carter, J.L., 1993. Middle
Paleozoic exotic terrane near Ciudad Victoria, northeastern Mexico,
and the southern margin of Paleozoic North America. 1st, Circum-
Pacific and Circum-Atlantic Terrane Conference Proceedings, Uni-
versidad Nacional Autonoma de Mexico, Instituto de Geologıa,
147–149.
Stewart, J.H., Blodgett, R.B., Boucot, A.J., Carter, J.L., Lopez, R., 1999.
Exotic Paleozoic strata of Gondwana provenance near Ciudad Victoria,
Tamaulipas, Mexico. In: Ramos, V.A., Keppie, J.D. (Eds.), Laurentia–
Gondwana Connections before Pangea, Geological Society of America,
Special Paper, 336., pp. 227–252.
Villasenor-Martinez, A.B., Martinez, C.A., Contreras, M.B., 1987.
Bioestratigrafıa del Paleozoico Superior de San Salvador Patlanoaya,
Puebla, Mexico. Revista de la Sociedad Mexicana de Paleontologıa 1,
390–417.
Waagen, W.H., 1883. Salt Range fossils I. Productus–Limestone fossils.
Geological Survey of India, Memoirs, Palaeontologıa Indica, Serie 13,
vol. 1, Part 4. Brachiopoda: fasc. 2, pp. 391–546.
Waterhouse, J.B., 1975. New Permian and Triassic Brachiopod Taxa.
University of Queensland, Department of Geology, Papers 7 (1), 1–23.
Weller, S., 1914. The Mississippian Brachiopoda of the Mississippi Valley
Basin. Geological Survey of Illinois, Monograph, 1, 508 pp..
Winchell, A., 1863. Descriptions of fossils of the yellow sandstone lying
beneath the Burlington Limestone at Burlington, Iowa. Academy of
Natural Sciences of Philadelphia Proceedings 15 (7), Ser. 2, pp. 2–25.
D. Navarro-Santillan et al. / Journal of South American Earth Sciences 15 (2002) 327–336336
Top Related
Copyright © 2022 FDOKUMEN

