






Bahasa
Halaman
Hukum


R E S E A R C H A R T I C L E
Immunogenicityandprotective e⁄cacyofVibrio cholerae outermembranevesicles in rabbitmodelNivedita Roy1, Soumik Barman1, Amit Ghosh2, Amit Pal2, Krishnendu Chakraborty3, Santa Sabuj Das3,Dhira Rani Saha4, Shinji Yamasaki5 & Hemanta Koley1
1Division of Bacteriology, National Institute of Cholera and Enteric Diseases, Kolkata, India; 2Division of Pathophysiology, National Institute of Cholera
and Enteric Diseases, Kolkata, India; 3Division of Clinical Medicine, National Institute of Cholera and Enteric Diseases, Kolkata, India; 4Division of
Electron Microscopy, National Institute of Cholera and Enteric Diseases, Kolkata, India; and 5Graduate School of Life and Environmental Sciences, Osaka
Prefecture University, Osaka, Japan
Correspondence: Hemanta Koley, Division
of Bacteriology, National Institute of Cholera
and Enteric Diseases, P – 33 CIT Road, Scheme
–- XM, Beliaghata, Kolkata 700010, India.
Tel.: 191 33 2353 7470, Ext. 3020; fax: 191
33 2370 5066; e-mail:
Received 25 September 2009; revised 17 April
2010; accepted 26 April 2010.
Final version published online 7 June 2010.
DOI:10.1111/j.1574-695X.2010.00692.x
Editor: Patrik Bavoil
Keywords
outer membrane vesicles (OMVs); Vibrio
cholerae ; oral immunization; colonization;
rabbit model; RITARD model.
Abstract
We show here that oral immunization with purified outer membrane vesicles
(OMVs) of Vibrio cholerae induces a prolonged high rise in the protective antibody
titre. Rabbit immune sera were vibriocidal against the homologous and against
several heterologous V. cholerae strains in vitro. In addition, OMV immunization
conferred significant protective immunity against subsequent bacterial challenges.
Thirty OMV-immunized rabbits were challenged with different V. cholerae strains;
each challenged group contained five immunized and three unimmunized
animals. All the immunized rabbits survived bacterial challenges and were healthy
after 24 h, except the two from each group that received the SG24 and SG06 strains,
respectively, which developed watery diarrhoea. In contrast, all the unimmunized
animals developed cholera-like symptoms, with a death toll of eight within 24 h of
challenge. This is the first report of the induction of protective immunity by
V. cholerae OMVs in a rabbit model (removable intestinal tie-adult rabbit
diarrhoea) that mimics the human disease. Finally, OMVs were found to be
significantly less reactogenic than the live and the heat-killed bacteria. Our studies
show that oral immunization with OMVs of V. cholerae may induce long-term
immunity and may be useful as a ‘nonliving’ vaccine candidate for the future.
Introduction
Enteric bacterial infection is the major cause of diarrhoea,
particularly in the developing world (Mathan, 1998). Chil-
dren and adults living in the less developed world experience
frequent diarrhoeal episodes, which may lead to loss of
working hours, costly hospitalization and sometimes death
(Monroe et al., 1991). Furthermore, travellers to developing
countries are exposed to the high risk of infectious diar-
rhoea, resulting in discomfort and inconvenience. Although
the mortality from diarrhoeal diseases has been significantly
reduced for some time, primarily due to better quality of
drinking water, improved sanitation and the availability of
more effective antimicrobial agents, the disease burden and
as a consequence the morbidity and economic fallout still
remain substantial. Vibrio sp. constitutes one of the leading
causes of acute watery diarrhoea. Cholera, sometimes
known as Asiatic or epidemic cholera, is an infectious
gastroenteritis caused by enterotoxin-producing strains of
the bacteria Vibrio cholerae that leads to epidemic cases of
gastroenteritis and sporadic cases of septicaemia. However,
epidemic V. cholerae isolates are represented by only two
serogroups, namely the O1 and O139, of a total of 209
different serogroups identified so far on the basis of the
somatic O-antigen (Chatterjee & Chaudhuri, 2003).
Non-O1 and non-O139 V. cholerae species have only been
associated with sporadic cases of diarrhoea (Faruque et al.,
1998).
Although cholera can be life threatening, it can be
prevented by proper sanitation and supply of bacteriologi-
cally pure drinking water. However, the economic cost of
infrastructural development to ensure water supply and
sanitation facilities to every citizen is huge and has not yet
been achieved in India, where cholera is endemic. A cheaper
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd.
IMM
UN
OLO
GY
& M
EDIC
AL
MIC
ROBI
OLO
GY

alternative for individual and community-level protection
against cholera might be provided by an economical and
effective vaccine that would mitigate the spread of the
disease. Parenteral cholera vaccines are no longer in use,
but both live and killed whole cell vaccines are currently
available on the market and have been found to be safe,
immunogenic and effective in defined populations. These
include killed whole cell-recombinant cholera toxin B sub-
unit vaccine (WC/rCTB; Dukorals, Crucel/SBL, Sweden)
(Richie et al., 2000), killed whole vaccine (the Vietnamese
vaccine; ORC-Vaxs, VaBiotech) and modified Vietnamese
vaccine (polyvalent killed whole cell; Shancols, Shanta
Biotech, India). The only licensed live oral vaccine,
CVD103-HgR (Orochols, Crucel/Berna Biotech, Switzer-
land), has not been produced since 2004. A lack of efficacy in
certain populations and reactogenicity have remained the
major bottlenecks preventing the wider use of the available
cholera vaccines, particularly in developing countries. Sev-
eral research groups have recently shifted their focus to the
development of subunit vaccines. However, results from
animal experiments suggest that such vaccines may only be
effective when administered with an adjuvant, and serious
concerns have been raised that nonliving protein material
delivered onto mucosal surfaces may induce a state of
tolerance. Even cholera toxin or the B subunit, the two
potent mucosal adjuvants for the induction of antibody
responses, may actually be tolerogenic when it comes to
T-cell-dependent immunity. There exists a need to develop a
new vaccine that can offer long-term protection against
infections with a wide range of V. cholerae serogroups and
that at the same time causes fewer adverse effects.
A wide variety of Gram-negative bacteria, such as Escher-
ichia coli, Neisseria meningitidis, Pseudomonas aeruginosa,
Shigella flexneri, Helicobacter pylori and V. cholerae constitu-
tively secrete outer membrane vesicles (OMVs) during their
growth in vitro (Chatterjee & Das, 1967; Devoe & Gilchrist,
1973; Hoekstra et al., 1976; Kondo et al., 1993; Fiocca et al.,
1999; Kadurugamuwa & Beveridge, 1999). Recent studies
with mice have shown promise for the use of OMVs to
provide protection against infections with N. meningitidis
(Gonzaleza et al., 2006), Salmonella enterica serovar Typhi-
murium (Alaniz et al., 2007), H. pylori (Keenan et al., 2000)
and V. cholerae (Schild et al., 2008). OMVs are spherical,
bilayered lipoproteins with an average diameter of
20–200 nm. They are composed of outer membrane pro-
teins, lipopolysaccharide, outer membrane lipids, periplas-
mic proteins, cytoplasmic proteins, DNA, RNA and other
factors associated with virulence (Horstman & Kuehn, 2000;
Wai et al., 2003; Kuehn & Kesty, 2005; Bauman & Kuehn,
2006; Nevot et al., 2006; Lee et al., 2008).
Studies of OMVs from diverse bacterial strains suggest a
role in the delivery of toxins to host cells, the transfer of
proteins and genetic material between bacterial cells, cell-to-
cell signals and the elimination of competing organisms
(Kuehn & Kesty, 2005; Mashburn-Warren & Whiteley,
2006). Because OMVs are essential to bacterial survival in
the host and pathogenesis, modulation of vesicle formation
and their functions may be a useful objective in relation to
the development of antibiotics as well as vaccines (Henry
et al., 2004; Lee et al., 2008). As outer membrane proteins
and lipopolysaccharide of OMVs may induce host immune
response, detergent-extracted OMVs derived from patho-
genic strains, including N. meningitides, are now undergoing
clinical trials (Fredriksen et al., 1991; de Moraes et al., 1992;
Edwards & Apicella, 2004; Davidsen & Tonjum, 2006;
Gorringe et al., 2009).
A vaccine developed from OMVs of group B meningo-
cocci suspended in saline has been proved to be strongly
immunogenic in mice and humans when administered
intranasally as drops or spray, even without the use of
mucosal adjuvants (Dalseg et al., 1999; Siadat et al., 2007;
Gorringe et al., 2009). Recently, Schild et al. (2009) reported
that V. cholerae OMVs also possess immunogenicity, and
mucosal immunization of mice with these OMVs induces
long-term protective immunity against this gastrointestinal
pathogen in the litters.
In the current study, we thoroughly investigated the
safety, immunogenicity and protective efficacy of OMVs
derived from V. cholerae and their potential use as an orally
administered candidate vaccine.
Materials and methods
Animals
New Zealand White rabbits of either sex weighing 2.0–2.5 kg
were used for oral immunization with OMVs and challenge
experiments in the removable intestinal tie-adult rabbit
diarrhoea (RITARD) model. All rabbits were collected from
the animal resource department of the National Institute of
Cholera and Enteric Diseases, Kolkata. They were individu-
ally caged and maintained at 24 1C with 75% humidity and
fed sterile food and water. Gut sterilization of the animals
was performed by treatment with a course of metronidazole
(125 mg per rabbit day�1) and sulphaquinoxaline sodium
(464 mg per rabbit day�1) for 5 days, and the course was
repeated three times at 2-day intervals to remove giardia and
coccidia from the intestine (Albert et al., 1991; Koley et al.,
1995). Approval was received from the institutional ethical
committee before the animal experiments were performed.
Bacterial strains
OMV antigen was prepared from the reference strain
N16961 (El-tor Inaba). The other V. cholerae strains that
were selected for the heterologous challenge experiments
were O395 (Classical Ogawa), 569B (Classical Inaba), VC20
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd.
19Vibrio cholerae OMVs vaccine candidate

(El-tor Ogawa), SG24 (O139) and SGO6 (O45, non-O1,
non-O139). All strains were stored in 15% glycerol with
brain heart infusion broth (BHIB, Difco) at � 70 1C.
OMV preparation
OMVs were isolated from the reference V. cholerae strain
N16961 following the method of Balsalobre et al. (2006).
Briefly, 1 L of Luria–Bertani broth (LB, Difco) was inocu-
lated with 10 mL of the stationary phase culture of N16961
strain and grown for 8 h to the late exponential phase.
Bacteria were removed by centrifugation (4500 g, 15 min,
4 1C), and the supernatant was filtered by passing it con-
secutively through 0.45-mm and 0.22-mm pore size filters,
respectively. To confirm the absence of viable bacteria, 1 mL
of the filtrate was plated on an LB agar plate, which was
incubated overnight at 37 1C. Protease inhibitors [Complete
EDTA-free protease inhibitor cocktail (Roche), 1 tablet per
1 L of filtrate] were added to the filtrate to prevent protein
degradation. OMVs were subsequently purified from the
filtrate by ultracentrifugation (4 h, 140 000 g, 4 1C) using a
Beckman SW32Ti rotor, washed once with phosphate-
buffered saline (PBS; pH 7.4) and finally resuspended in
625mL of PBS. Protein concentration was determined by the
Modified Lowry Protein Assay Kit (Pierce) and adjusted to
2.5mg mL�1 using PBS. Purified OMVs were stored at
� 80 1C until use.
Electron microscopy
A 5-mL aliquot of OMVs was deposited on the carbon-
coated grid, allowed to settle for approximately 2 min and
washed with Tris buffer solution. Excess fluid was blotted
and covered with a small drop of 2% aqueous uranyl acetate
stain. After a few seconds, this drop was also blotted dry and
the sample was examined under a Tecnai 12 (Bio-Twin
Transmission Electron Microscope; FEI, the Netherlands)
operating at 80 kV (Hayat & Miller, 1990).
Reactogenicity assay
The reactogenicity of the bacteria or the purified vesicles was
assayed by analysing interleukin (IL)-8 induction in HT-29
cells. Total RNA was extracted from the unstimulated cells
and their stimulated counterparts with TrizolTM reagent
(Invitrogen) following the manufacturer’s instructions.
cDNA was prepared from 2 mg of the extracted RNA using
Superscript II reverse transcriptase (Invitrogen), following
the standard protocol. IL-8 was amplified by SYBR-Greens
real-time quantitative PCR using ABI7300 (Applied Biosys-
tems). Relative quantitation was performed using the com-
parative threshold cycle number (CT) method. PCR was
performed with SYBR GreenTM Master MixTM (Applied
Biosystems), where SYBR green is the fluorescent reporter.
The internal control gene GAPDH was amplified simulta-
neously in separate reaction tubes. To eliminate primer
dimers, the fluorescent signal was collected at 82 1C, at
which temperature the primer dimmers melted, but the
PCR products were still in their double-stranded form,
thereby emitting fluorescence. The reaction conditions were
set as follows: initial heating at 95 1C for 5 min, followed by
40 cycles of reactions at 95 1C (1 min), followed by 62 1C
(2.5 min) and finally 82 1C (45 s). Final extension was
carried out at 62 1C for 8 min. The CT of triplicate reactions
was determined using ABI-SDS software and the mean CT of
triplicate reactions was determined. The levels of expression
of the genes of interest were normalized to GAPDH using
the formula 2�DDCT , where �DDCT is DCT (sample)�DCT
(calibrator) and DCT is the CT of the target gene subtracted
from the CT of the housekeeping gene (GAPDH). The
calibrator used in our experiments was the unstimulated
HT-29 cells.
Lipopolysaccharide preparation
Lipopolysaccharide prepared from different V. cholerae
strains was analysed by sodium dodecyl sulphate polyacry-
lamide gel electrophoresis (SDS-PAGE) (Hitchcock &
Brown, 1983). For lipopolysaccharide preparation, over-
night culture of V. cholerae in LB was harvested and washed
with PBS followed by treatment with 50 mL of lysis buffer
containing 2% SDS, 4% 2-mercaptoethanol, 10% glycerol,
1 M Tris (pH 6.8) and bromophenol blue. Lysates were
heated at 100 1C for 10 min. Proteins present in the lysates
were digested by adding 25mg of proteinase K solubilized in
10 mL of lysis buffer and heating the lysates at 60 1C for
60 min. Treated materials were subsequently electrophor-
esed onto SDS-PAGE and the polysaccharide materials were
fixed in the gel by treatment with ethanol–acetic acid
fixative. The gels were developed by silver staining to
visualize the lipopolysaccharide profile of various V. cholerae
strains.
Oral immunization of rabbits
New Zealand White rabbits of either sex (weighing 2–2.5 kg)
were immunized orally with OMVs (N16961) on days 0, 7,
14 and 21 according to the protocol developed by Spira et al.
(1981). The immunized group contained 30 rabbits and the
control group 18 rabbits (Tables 1 and 2). Rabbits were
fasted for 36 h, but water was given ad libitum. Thirty
minutes before the oral immunization, each rabbit was
anaesthetized by intramuscular injection of a mixture of
ketamine (35 mg kg�1 body weight; Sterfil Laboratories Pvt.
Ltd, India) and xylazine (5 mg kg�1 body weight, AstraZe-
neca Pharma India Ltd, India). Intravenous ranitidine
(50 mg kg�1 body weight; Ranbaxy, India) was administered
after 5 min, followed by two boluses of sodium bicarbonate
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd.
20 N. Roy et al.
(15 mL of a 5% solution; SRL, India) at 15-min intervals
introduced directly into the stomach through a feeding tube
(Romsons Sci. and Surg. Ind. Pvt. Ltd, India). The second
bolus was immediately followed by oral administration of
OMVs (50mg in 1 mL PBS) for the experimental rabbits and
the same volume of PBS for the control group. The rabbits
were returned to their cages and given limited amounts of
sterile food and water. Blood samples were collected for the
measurement of immunoglobulin titres before, during and
after immunization.
Serum collection
Blood was collected from the ear veins of rabbits on days 0,
7, 14, 21, 28, 35, 51, 65, 71 and 87. Collected blood was
allowed to clot at room temperature for 30 min and then
kept at 4 1C for 24 h. Serum was separated from the clotted
blood using a sterile Pasteur pipette and centrifuged
(10 min, 1000 g). The supernatant was collected and stored
at � 70 1C after addition of sodium azide to a final
concentration of 0.05% until further use.
Protection study
Postimmunization challenge of the rabbits with bacteria was
done following the protocol developed by Spira et al. (1981).
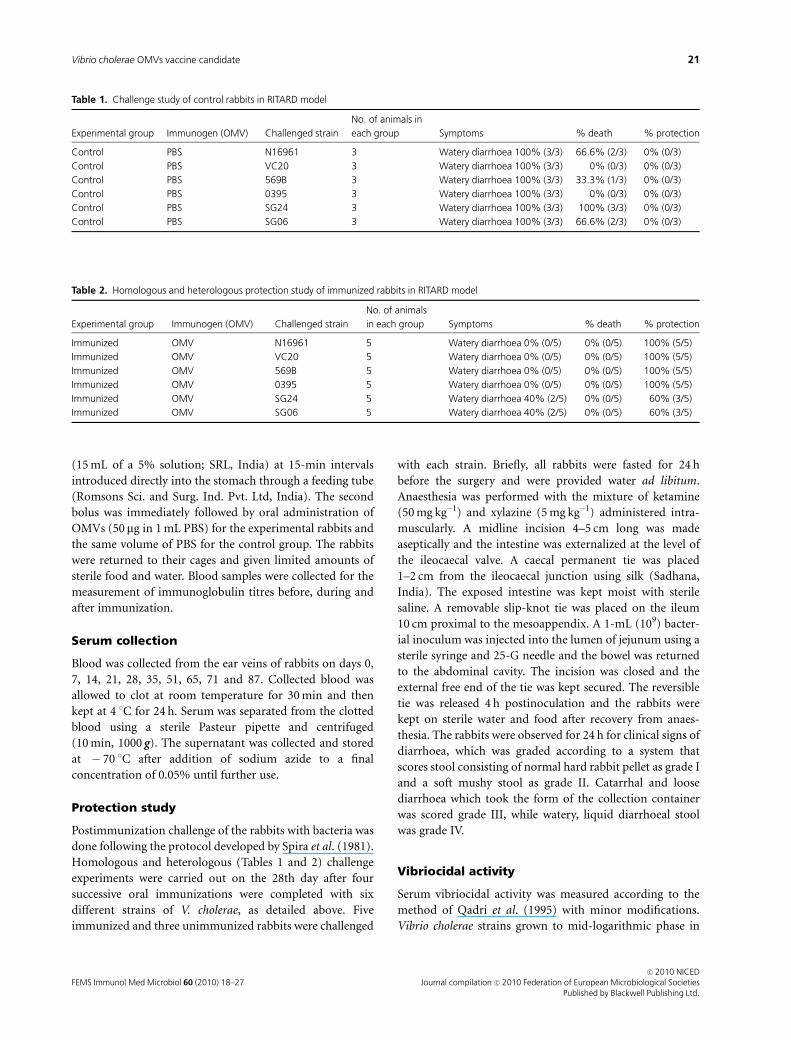
Homologous and heterologous (Tables 1 and 2) challenge
experiments were carried out on the 28th day after four
successive oral immunizations were completed with six
different strains of V. cholerae, as detailed above. Five
immunized and three unimmunized rabbits were challenged
with each strain. Briefly, all rabbits were fasted for 24 h
before the surgery and were provided water ad libitum.
Anaesthesia was performed with the mixture of ketamine
(50 mg kg�1) and xylazine (5 mg kg�1) administered intra-
muscularly. A midline incision 4–5 cm long was made
aseptically and the intestine was externalized at the level of
the ileocaecal valve. A caecal permanent tie was placed
1–2 cm from the ileocaecal junction using silk (Sadhana,
India). The exposed intestine was kept moist with sterile
saline. A removable slip-knot tie was placed on the ileum
10 cm proximal to the mesoappendix. A 1-mL (109) bacter-
ial inoculum was injected into the lumen of jejunum using a
sterile syringe and 25-G needle and the bowel was returned
to the abdominal cavity. The incision was closed and the
external free end of the tie was kept secured. The reversible
tie was released 4 h postinoculation and the rabbits were
kept on sterile water and food after recovery from anaes-
thesia. The rabbits were observed for 24 h for clinical signs of
diarrhoea, which was graded according to a system that
scores stool consisting of normal hard rabbit pellet as grade I
and a soft mushy stool as grade II. Catarrhal and loose
diarrhoea which took the form of the collection container
was scored grade III, while watery, liquid diarrhoeal stool
was grade IV.
Vibriocidal activity
Serum vibriocidal activity was measured according to the
method of Qadri et al. (1995) with minor modifications.
Vibrio cholerae strains grown to mid-logarithmic phase in
Table 2. Homologous and heterologous protection study of immunized rabbits in RITARD model
Experimental group Immunogen (OMV) Challenged strain
No. of animals
in each group Symptoms % death % protection
Immunized OMV N16961 5 Watery diarrhoea 0% (0/5) 0% (0/5) 100% (5/5)
Immunized OMV VC20 5 Watery diarrhoea 0% (0/5) 0% (0/5) 100% (5/5)
Immunized OMV 569B 5 Watery diarrhoea 0% (0/5) 0% (0/5) 100% (5/5)
Immunized OMV 0395 5 Watery diarrhoea 0% (0/5) 0% (0/5) 100% (5/5)
Immunized OMV SG24 5 Watery diarrhoea 40% (2/5) 0% (0/5) 60% (3/5)
Immunized OMV SG06 5 Watery diarrhoea 40% (2/5) 0% (0/5) 60% (3/5)
Table 1. Challenge study of control rabbits in RITARD model
Experimental group Immunogen (OMV) Challenged strain
No. of animals in
each group Symptoms % death % protection
Control PBS N16961 3 Watery diarrhoea 100% (3/3) 66.6% (2/3) 0% (0/3)
Control PBS VC20 3 Watery diarrhoea 100% (3/3) 0% (0/3) 0% (0/3)
Control PBS 569B 3 Watery diarrhoea 100% (3/3) 33.3% (1/3) 0% (0/3)
Control PBS 0395 3 Watery diarrhoea 100% (3/3) 0% (0/3) 0% (0/3)
Control PBS SG24 3 Watery diarrhoea 100% (3/3) 100% (3/3) 0% (0/3)
Control PBS SG06 3 Watery diarrhoea 100% (3/3) 66.6% (2/3) 0% (0/3)
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd.
21Vibrio cholerae OMVs vaccine candidate

TSB (Difco) were centrifuged, washed with PBS and resus-
pended to adjust the OD of the cell suspension to 0.20–0.22
at 600 nm wavelength. Freshly collected serum samples from
the immunized and the unimmunized rabbits were heat
treated for 1 h at 56 1C. Serum samples were serially diluted
in a 96-well plate and 50 mL of each dilution was used for
bactericidal activity. A reaction mixture was prepared using
PBS, bacterial cell and complement (Rockland) at a ratio of
6 : 3 : 1, and 50 mL of the reaction mixture was added to each
well containing the samples. After 1 h of incubation, 100mL
of brain heart infusion broth (BHI broth; Difco) was added
to each well of the 96-well plate and the reaction mixture
was incubated for further 4 h. For negative control reactions,
only PBS and complement was added. The positive control
contained PBS, bacterial cell, complement and El-tor, O1
Inaba, monoclonal antisera (Denka Seiken, Japan). The
reading was taken at 600 nm using an enzyme-linked
immunosorbent assay (ELISA) reader.
Immunoblot against lipopolysaccharide
Lipopolysaccharide of different V. cholerae strains was
resolved by SDS-PAGE and transferred to a nitrocellulose
membrane (Bio-Rad) at 0.25 A and 100 V for 1 h using a
transblot cell (Bio-Rad). The membrane was blocked with
5% skimmed milk (Bio-Rad) at room temperature for 2 h.
After washing with Tris-buffered saline (TBS) containing
0.1% Tween-20, the membrane was incubated with poly-
clonal rabbit antisera raised against OMVs of V. cholerae
N16961 strain at a dilution of 1 : 1000 for 90 min at room
temperature with gentle shaking. The nitrocellulose blot was
washed and incubated with a 1 : 5000 dilution of alkaline
phosphatase-conjugated antirabbit IgG (Sigma Chemical)
for 60 min at 37 1C. The blot was developed with a solution
of nitro blue tetrazolium and 5-bromo-4-chloroindolyl
phosphate.
ELISA
Analysis for the serum IgG antibody was performed using
ELISA, essentially following the method developed by Keren
(1979). Disposable polystyrene (Nunc, Denmark) microtitre
wells were coated with 10 mg of the whole cell lysate of V.
cholerae N16961 strain in 100mL of solution and incubated
for 18 h at 4 1C. Wells were washed three times with PBS-T
(PBS with 0.5% Tween-20). Nonspecific binding sites were
blocked by incubating the wells with 100 mL of 5% BSA
(Sigma Chemical) for 2 h at 37 1C. The wells were washed
three times with PBS-T and incubated with serially diluted
serum samples at 37 1C for 1 h. Following washing, 100mL
each of horseradish peroxidase-conjugated goat antirabbit
immunoglobulin G (Sigma Chemical) and the substrate o-
phenyl-D-amine (OPD) was added to each well and the plate
was incubated at 37 1C. The reaction was stopped after
10 min by adding 25 mL of 5 N sulphuric acid and the
reading was taken at 405-nm wavelength using an ELISA
reader.
Statistical analysis
Frequencies were compared by the chi-squared test, and the
continuous variable was compared using Student’s t-test.
P values � 0.05 and 0.01 were considered statistically
significant and highly significant, respectively.
Results
Vibrio cholerae produces OMVs
A 5-mL amount of OMVs was examined by transmission
electron microscopy, which showed multiple vesicle-like
structures representing OMVs. A few bacteria with vesicles
protruding from their surface were also seen and a repre-
sentative one is shown in Fig. 1.
Immunogenicity of OMVs
OMVs of several Gram-negative bacteria have been sug-
gested as candidates for vaccine development. To study the
immunogenic potential of V. cholerae OMVs, the serum IgG
antibody titre of rabbits against purified OMVs of the
N16961 strain was measured during the preimmunization,
immunization and postimmunization periods. The recipro-
cal titre of OMV-specific serum IgG increased linearly from
day 7 of immunization and reached a plateau around day 21.
This level was maintained during the remaining course of
immunization as well as during the postimmunization
period till the 51st day in most rabbits. In addition, a very
Fig. 1. Electron micrograph of free OMVs as well as those attached to
the bacteria (Vibrio cholerae N16961 strain) (� 20 magnification).
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd.
22 N. Roy et al.

high antibody titre was found until our final measurement
was taken 65 days after immunization was complete (Fig. 2).
Specificity of the antibody response
Bacterial lipopolysaccharide is a major immunogenic molecule
and the cause of the generation of vibriocidal antibodies in the
serum following infection/immunization. The lipopolysac-
charide profiles of the V. cholerae strains used in our studies
except SG24 were identical with respect to the rapid migrating
form. The SG24 strain lacked the long repetitive units of ‘O’
polysaccharide of lipopolysaccharide and was characterized by
the presence of the core oligosaccharide alone. Rabbit antibody
raised against the OMVs of N16961 strain could detect
lipopolysaccharide of several heterologous strains except that
of SG24 and SG06 in a Western blot analysis, suggesting
specificity for the ‘O’ polysaccharide (Fig. 3).
Protective efficacy of OMV sera
Vibriocidal antibody titre is considered to be the best
correlate and a surrogate marker of protection against V.
cholerae infection. Vibriocidal activity of serum is measured
by an in vitro complement fixation test that detects vibrio-
cidal antibody titres raised against the lipopolysaccharide of
V. cholerae. As the membrane of OMVs is derived from the
bacterial cell membrane and contains lipopolysaccharide, we
investigated whether the vesicles would induce vibriocidal
antibodies in rabbits. In our studies, rabbits were immu-
nized on days 0, 7, 14 and 21 and serum was collected on the
28th day to measure vibriocidal activities. While the im-
mune serum exhibited the highest vibriocidal activities
against the homologous strain, significant activities were
also found against most heterologous V. cholerae strains.
Among the latter group, the greatest vibriocidal potential
(�50%) was seen against VC20, 569B and O395 strains. In
contrast, only 22% and 15% killing was observed with SGO6
and SG24 strains, respectively (Fig. 4).
Protective efficacy of OMV immunization
To study the protective efficacy of OMV-specific serum
antibodies in vivo, rabbits were challenged with the homo-
logous and five of the heterologous V. cholerae strains
following oral immunization with OMVs (50mg) on days 0,
7, 14 and 21. On the 28th day, five immunized and three
unimmunized rabbits were challenged with each V. cholerae
strain as described under Materials and methods. Rabbits
were monitored for the next 24 h for the development of the
symptoms of ‘cholera’. All the unimmunized rabbits became
sick and developed moderate to profuse watery diarrhoea
within 16–18 h of challenge with bacteria, and their perineal
region remained constantly wet and soiled with faeces. In
contrast, two each (60%) from the immunized groups
challenged with SG24 and SGO6 strains, respectively, and
the rest that received a challenge with either of the four
V. cholerae strains developed no symptoms of diarrhoea and
were alive and healthy after 24 h (Tables 1 and 2). The above
results suggest 60–100% protective efficacies of the OMV
immunization schedule against the different V. cholerae
strains used in our study.
Reactogenicity of OMVs
Reactogenicity has remained a major bottleneck for many
vaccine strains of V. cholerae and has precluded their accep-
tance for mass vaccination despite considerable protective
efficacy. Reactogenicity is caused by the pro-inflammatory
Fig. 2. Vibrio cholerae OMVs are immunogenic. Five rabbits were
immunized orally on days 0, 7, 14 and 21 with OMVs (50 mg) purified
from V. cholerae N16961 strain. Serum IgG antibody titre was measured
on the days indicated and mean (� SD) values obtained from all the five
rabbits are presented here.
Fig. 3. OMV-induced serum antibody is lipopolysaccharide-specific.
(Left panel) SDS-PAGE of lipopolysaccharide extracted from different
Vibrio cholerae strains stained with Silver stain. (Right panel) Immunoblot
of lipopolysaccharide run in SDS-PAGE and probed with the rabbit
antiserum raised against OMVs of N16961. Lane 1: N16961, lane 2:
VC20, lane 3: 569B, lane 4: O395, lane 5: SG24 and lane 6: SGO6.
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd.
23Vibrio cholerae OMVs vaccine candidate

activities of the bacteria and predominantly results from the
induction of neutrophil chemotactic factor IL-8. To investi-
gate whether OMVs may also be reactogenic, human colon
epithelial cell line HT-29 was treated for 90 min with the
OMVs as well as the live, formalin-treated or heat-killed
bacteria. IL-8 mRNA expression in the cells was analysed by
qRT-PCR. The results showed that the live and the heat-
killed V. cholerae induced the highest levels of IL-8, whereas
the OMVs had the least effect (Fig. 5). These findings
indicate that a vaccine generated from OMVs may be less
reactogenic and better tolerated than the existing vaccines,
which are based on the live or the heat-killed cells.
Discussion
Gram-negative bacteria constitutively secrete OMVs into
the extracellular milieu. However, natural production of
large amounts of OMVs has only been described in a few
bacterial genera. Recent research in this area has revealed
that OMVs may act as intercellular communicasomes in
polyspecies communities by enhancing bacterial survival
and pathogenesis in hosts. Although research in this area
has revealed diverse functions of OMVs, the mechanisms of
vesicle formation and of protein sorting into OMVs, as well
as the pathophysiological role of OMVs, have not been
clearly defined (Lee et al., 2008).
Studies have shown that purified vesicles may induce
protective immune responses against subsequent infections
and may be potential candidates in vaccine development
(Henry et al., 2004). Camilli and colleagues successfully
purified large quantities of OMVs from two different strains
of V. cholerae (Schild et al., 2008). Here, we have purified
OMVs from a third strain, suggesting that many different
V. cholerae strains produce OMVs and that an OMV-based
vaccine may be useful against a wide range of V. cholerae
infections. Schild et al. (2009) used a mouse model to
demonstrate that purified OMVs administered through
oral, intranasal and intraperitoneal routes induce protective
immunity against colonization in the litters. Whereas pre-
natal transfer of anti-OMV immunoglobulins reduces
V. cholerae colonization, immunoglobulins transferred
through breast milk completely protect the neonates from
colonization (Schild et al., 2009). However, protection from
diarrhoea cannot be studied in a mouse model as mice do
not develop symptomatic cholera (Olivier et al., 2007). We
used the RITARD model, which closely resembles human
cholera, and showed that oral immunization with OMVs
completely prevents death and provides significant protec-
tion against the development of diarrhoea in rabbits. In
addition, we have demonstrated for the first time that
OMVs protect against the homologous as well as several
heterologous strains of V. cholerae. This is extremely im-
portant from a vaccination point of view, as different
V. cholerae serogroups and strains may be responsible for
diarrhoeal diseases not only in different geographical loca-
tions, but also at the same site at a particular point of time.
However, we have only used the oral route of immunization
in our studies. Although a previously published study
reported that equivalent protection was conferred by multi-
ple routes of immunization with OMVs (Schild et al., 2009),
it would be important to study the protective efficacy of
other routes besides oral immunization in rabbits. It is
noteworthy that OMVs induced long-term immunity in
both mouse and rabbit models.
Fig. 5. OMVs are the least reactogenic of the vaccines. The reactogeni-
city of different vaccine candidates was analysed by determining their
potency in inducing IL-8 in the cultured HT29 cells. Cells were stimulated
with formalin and heat-killed Vibrio cholera as well live bacteria and
purified vesicles for 90 min and IL-8 mRNA expression was analysed by
qRT-PCR (�Po 0.01, highly significant; 1Po 0.05, significant).
Fig. 4. Vibriocidal activity of OMV-induced antibody. Rabbits were
immunized with OMVs as described under Materials and methods.
Antiserum was collected on day 28 and the homologous and five
heterologous Vibrio cholerae strains were used for the serum vibriocidal
antibody assay as described above. Lane 1: N16961, lane 2: VC20, lane
3: 569B, lane 4: O395, lane 5: SG24 and lane 6: SGO6. The results were
expressed as fold increase compared with the nonimmune serum
(mean� SD of three independent experiments; �Po 0.01).
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd.
24 N. Roy et al.

Earlier studies suggested that the immunogenicity and
protective efficacy of OMVs may result from the proteins
(OMPs) (Cartwright et al., 1999; Borrow et al., 2005).
However, those authors did not investigate whether dena-
turation of the proteins resulted in loss of immunogenicity
and protective efficacy, as happens in the case of killed
whole-cell vaccines (Jertborn, 1992). OMPs may be respon-
sible for OMV-induced immunity in the case of Gram-
negative pathogens. However, the molecule of V. cholerae
that is mainly responsible for the induction of protective
immunity remains to be identified. Local protection in the
intestine is believed to be provided primarily by the secre-
tory IgA, and the serum vibriocidal antibody titre that is
induced by bacterial lipopolysaccharide is considered to be
the surrogate marker of protection. Our studies, for the first
time, strongly indicate that OMV-associated lipopolysac-
charide significantly induces vibriocidal antibody in the
serum. This is underscored by the fact that immunization
with OMVs of the N16961 strain gave rise to significant
vibriocidal antibody titres against heterologous V. cholerae
strains that possess a structurally similar lipopolysaccharide,
but not against the SG24 strain that has a different ‘O’
polysaccharide structure. Lack of response against SG24
strain could also result from the fact that V. cholerae O139
strains, as opposed to other serogroups, are encapsulated,
thus masking lipopolysaccharide from detection by antibo-
dies. Anti-OMV antibody indeed functions as a surrogate
marker of protection against subsequent V. cholerae infec-
tions as indicated by our studies, which showed that the
OMV-immunized rabbits were less protected against cho-
lera-like symptoms caused by the SG24 strains. It is not clear
from our studies whether OMV-specific IgG may directly
protect against gut infection, although previous studies have
suggested that serum IgG may be secreted in the intestine to
confer local protection (Ferrero et al., 1997).
Reactogenicity still remains a major obstacle in the
development of a vaccine that protects against V. cholerae
infection while at the same time causing minimal and
acceptable adverse reactions. Reactogenicity is fairly com-
mon with most vaccine strains of V. cholerae and is believed
to be caused by the proinflammatory reactions induced by
the bacteria. Although the bacterial molecule responsible for
this phenomenon remains to be identified, several virulence
factors may be functioning together, of which the motility-
associated protein flagellin may be the predominant inducer
of reactogenicity. IL-8 secreted by the intestinal mucosal
epithelial cells may be the initial signal for an acute
inflammatory response following bacterial encounter with
the host (Kagnoff & Eckmann, 1997). Recently, it was
suggested that there might be a correlation between the
reactogenicity and the ability of the V. cholerae strains to
induce IL-8 in vitro (Rodrıguez et al., 2001). We observed
significantly less IL-8 induction in the intestinal epithelial
cell line by the OMVs compared with the live bacteria and
the heat-killed whole cells, suggesting that an OMV-based
vaccine may be the least reactogenic vaccine.
Acknowledgements
This study was supported by a contingency grant from
NICED, ICMR. We thank Mr Krishna Biswas for animal
experimentation and Ms Tripti Biswas and Mr Suhasit
Ranjan Ghosh for technical assistance.
References
Alaniz RC, Deatherage BL, Lara JC & Cookson BT (2007)
Membrane vesicles are immunogenic facsimiles of Salmonella
typhimurium that potently activate dendritic cells, prime B and
T cell responses, and stimulate protective immunity in vivo. J
Immunol 179: 7692–7701.
Albert MJ, Alam K, Islam M, Montanaro J, Rahaman AS, Haider
K, Hossain MA, Kibriya AK & Tzipori S (1991) Hafnia alvei, a
probable cause of diarrhea in humans. Infect Immun 59:
1507–1513.
Balsalobre C, Silvan JM, Berglund S, Mizunoe Y, Uhlin BE & Wai
SN (2006) Release of the type I secreted alpha-haemolysin via
outer membrane vesicles from Escherichia coli. Mol Microbiol
59: 99–112.
Bauman SJ & Kuehn MJ (2006) Purification of outer membrane
vesicles from Pseudomonas aeruginosa and their activation of
an IL-8 response. Microbes Infect 8: 2400–2408.
Borrow R, Balmer P & Miller E (2005) Meningococcal surrogates
of protection-serum bactericidal antibody activity. Vaccine 23:
2222–2227.
Cartwright K, Morris R, Rumke H, Fox A, Borrow R, Begg N,
Richmond P & Poolman J (1999) Immunogenicity and
reactogenicity in UK infants of a novel meningococcal vesicle
vaccine containing multiple class 1 (PorA) outer membrane
proteins. Vaccine 17: 2612–2619.
Chatterjee SN & Chaudhuri K (2003) Lipopolysaccharides of
Vibrio cholerae. I. Physical and chemical characterization.
Biochim Biophys Acta 1639: 65–79.
Chatterjee SN & Das J (1967) Electron microscopic observations
on the excretion of cell-wall material by Vibrio cholerae. J Gen
Microbiol 49: 1–11.
Dalseg R, Wedege E, Holst J, Haugen IL, Høiby EA & Haneberg B
(1999) Outer membrane vesicles from group B meningococci
are strongly immunogenic when given intranasally to mice.
Vaccine 14: 2336–2345.
Davidsen T & Tonjum T (2006) Meningococcal genome
dynamics. Nat Rev Microbiol 4: 11–22.
de Moraes JC, Perkins BA, Camargo MC, Hidalgo NT, Barbosa
HA, Sacchi CT, Landgraf IM, Gattas VL, Vasconcelos HG &
Gral IM (1992) Protective efficacy of a serogroup B
meningococcal vaccine in Sao Paulo, Brazil. Lancet 340:
1074–1078.
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd.
25Vibrio cholerae OMVs vaccine candidate

Devoe IW & Gilchrist JE (1973) Release of endotoxin in the form
of cell wall blebs during in vitro growth of Neisseria
meningitidis. J Exp Med 138: 1156–1167.
Edwards JL & Apicella MA (2004) The molecular mechanisms
used by Neisseria gonorrhoeae to initiate infection differ
between men and women. Clin Microbiol Rev 17: 965–981.
Faruque SM, Albert MJ & Mekalanos JJ (1998) Epidemiology,
genetics and ecology of toxigenic Vibrio cholerae. Microbiol
Mol Biol R 62: 1301–1314.
Ferrero RL, Thiberge JM & Labigne A (1997) Local
immunoglobulin G antibodies in the stomach may contribute
to immunity against Helicobacter infection in mice.
Gastroenterology 113: 185–194.
Fiocca R, Necchi V, Sommi P, Ricci V, Telford J, Cover TL & Solcia
E (1999) Release of Helicobacter pylori vacuolating cytotoxin
by both a specific secretion pathway and budding of outer
membrane vesicles. Uptake of released toxin and vesicles by
gastric epithelium. J Pathol 188: 220–226.
Fredriksen JH, Rosenqvist E, Wedege E, Bryn K, Bjune G,
Frøholm LO, Lindbak AK, Møgster B, Namork E & Rye U
(1991) Production, characterization and control of
MenB-vaccine ‘Folkehelsa’: an outer membrane vesicle
vaccine against group B meningococcal disease. NIPH Ann 14:
67–79.
Gonzaleza S, Caballeroa E, Soriab Y, Cobasa K, Granadilloa M &
Pajona R (2006) Immunization with Neisseria meningitidis
outer membrane vesicles prevents bacteremia in neonatal
mice. Vaccine 24: 1633–1643.
Gorringe AR, Taylor S, Brookes C et al. (2009) Phase I safety and
immunogenicity study of a candidate meningococcal disease
vaccine based on Neisseria lactamica outer membrane vesicles.
Clin Vaccine Immunol 16: 1113–1120.
Hayat MA & Miller SE, eds (1990) Negative Staining, pp. 36–48.
McGraw-Hill, New York.
Henry T, Pommier S, Journet L, Bernadac A, Gorvel JP &
Lloubes R (2004) Improved methods for producing outer
membrane vesicles in Gram-negative bacteria. Res Microbiol
155: 437–446.
Hitchcock PJ & Brown TM (1983) Morphological heterogeneity
among polysaccharide lipopolysaccharide chemotypes in
silver-stained polyacrylamide gels. J Bacteriol 154: 269–277.
Hoekstra D, van der Laan JW, de Leij L & Witholt B (1976)
Release of outer membrane fragments from normally growing
Escherichia coli. Biochim Biophys Acta 455: 889–899.
Horstman AL & Kuehn MJ (2000) Enterotoxigenic Escherichia
coli secretes active heat-labile enterotoxin via outer membrane
vesicles. J Biol Chem 275: 12489–12496.
Jertborn M (1992) Cholera: transmission, treatment, prevention.
The 1991 epidemic is the first one in this century in Latin
America. Lakartidningen 89: 3942–3943, 3947.
Kadurugamuwa JL & Beveridge TJ (1999) Membrane vesicles
derived from Pseudomonas aeruginosa and Shigella flexneri can
be integrated into the surfaces of other Gram-negative
bacteria. Microbiology 145: 2051–2060.
Kagnoff MF & Eckmann L (1997) Epithelial cells as sensors for
microbial infection. J Clin Invest 100: 6–10.
Keenan J, Day T, Neal S, Cook B, Perez-Perez G, Allardyce R &
Bagshaw P (2000) A role for the bacterial outer membrane in
the pathogenesis of Helicobacter pylori infection. FEMS
Microbiol Lett 182: 259–264.
Keren DF (1979) Enzyme-linked immunosorbent assay for
Immunoglobulin G and Immunoglobulin A antibodies to
Shigella flexneri antigens. Infect Immun 24: 441–448.
Koley H, Ghosh AN, Paul M, Ghosh AR, Ganguly PK & Nair GB
(1995) Colonization ability & intestinal pathology of rabbits
orally fed with Vibrio cholerae O139 Bengal. Ind J Med Res 101:
57–61.
Kondo K, Takade A & Amako K (1993) Release of the outer
membrane vesicles from Vibrio cholerae and Vibrio
parahaemolyticus. Microbiol Immunol 37: 149–152.
Kuehn MJ & Kesty NC (2005) Bacterial outer outer membrane
vesicles and the host–pathogen interaction. Gene Dev 19:
2645–2655.
Lee EY, Choi DS, Kim KP & Gho YS (2008) Proteomics in Gram-
negative bacterial outer membrane vesicles. Mass Spectrom Rev
27: 535–555.
Mashburn-Warren LM & Whiteley M (2006) Special delivery:
vesicle trafficking in prokaryotes. Mol Microbiol 61: 839–846.
Mathan VI (1998) Diarrhoeal diseases. Brit Med Bull 54: 407–419.
Monroe SS, Stine SE, Gorelkin L, Herrmann JE, Blacklow NR &
Glass RI (1991) Temporal synthesis of proteins and RNAs
during human astrovirus infection of cultured cells. J Virol 65:
641–648.
Nevot M, Deroncele V, Messner P, Guinea J & Mercade E (2006)
Characterization of outer membrane vesicles released by the
psychrotolerant bacterium Pseudoalteromonas antarctica NF3.
Environ Microbiol 8: 1523–1533.
Olivier V, Salzman NH & Satchell KJ (2007) Prolonged
colonization of mice by Vibrio cholerae El Tor O1 depends on
accessory toxins. Infect Immun 75: 5043–5051.
Qadri F, Mohi G, Hossain J, Azim T, Khan AM, Salam MA, Sack
RB, Albert MJ & Svennerholm AM (1995) Comparison of the
vibriocidal antibody response in cholera due to Vibrio cholerae
O139 Bengal with the response in cholera due to Vibrio
cholerae O1. Clin Diagn Lab Immun 2: 685–688.
Richie EE, Punjabi NH, Sidharta YY et al. (2000) Efficacy trial of
single-dose live oral cholera vaccine CVD 103-HgR in North
Jakarta, Indonesia, a cholera-endemic area. Vaccine 18:
2399–2410.
Rodrıguez BL, Rojas A, Campos J, Ledon T, Valle E, Toledo W &
Fando R (2001) Differential interleukin-8 response of
intestinal epithelial cell line to reactogenic and nonreactogenic
candidate vaccine strains of Vibrio cholerae. Infect Immun 69:
613–616.
Schild S, Nelson EJ & Camilli A (2008) Immunization with Vibrio
cholerae outer membrane vesicles induces protective immunity
in mice. Infect Immun 76: 4554–4563.
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd.
26 N. Roy et al.

Schild S, Nelson EJ, Bishop AL & Camilli A (2009)
Characterization of Vibrio cholerae outer membrane vesicles as
a candidate vaccine for cholera. Infect Immun 77: 472–484.
Siadat SD, Kheirandish M, Norouzian D, Behzadiyannejad Q,
Najar Peerayeh S, Zangeneh M & Nejati M (2007) A flow
cytometric opsonophagocytic assay for measurement of
functional antibodies elicited after immunization with outer
membrane vesicle of Neisseria meningitidis serogroup B. Pak J
Biol Sci 10: 3578–3584.
Spira WM, Sack RB & Froehlich JL (1981) Simple adult rabbit
model for Vibrio cholerae and enterotoxigenic Escherichia coli
diarrhea. Infect Immun 32: 739–747.
Wai SN, Lindmark B, Soderblom T, Takade A, Westermark M,
Oscarsson J, Jass J, Richter-Dahlfors A, Mizunoe Y & Uhlin BE
(2003) Vesicle-mediated export and assembly of pore-forming
oligomers of the enterobacterial ClyA cytotoxin. Cell 115:
25–35.
FEMS Immunol Med Microbiol 60 (2010) 18–27
c� 2010 NICEDJournal compilation c� 2010 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd.
27Vibrio cholerae OMVs vaccine candidate
Top Related

Copyright © 2022 FDOKUMEN

