







Bahasa
Halaman
Hukum


GPH
YMa
MHUb
Mc
Bd
D7
AenoucrdcpmurserdptgptgpwwmpdP
Khp
*EAcGpp
Neuroscience 138 (2006) 133–148
0d
ENE EXPRESSION PROFILING AND ANALYSIS OF SIGNALINGATHWAYS INVOLVED IN PRIMING AND DIFFERENTIATION OF
UMAN NEURAL STEM CELLSTsgwfvKfigawnccs(GtpDnipdg2
rndltg2pn2ne
nbpilhT
. CAI,a P. WU,b M. OZEN,d Y. YU,c J. WANG,d
. ITTMANNd AND M. LIUa*
Alkek Institute of Biosciences and Technology, and Department ofedical Biochemistry and Genetics, Texas A&M University Systemealth Science Center, 2121 West Holcombe, Houston, TX 77030,SA
Departments of Neuroscience and Cell Biology, University of Texasedical Branch, Galveston, TX 77555, USA
Department of Radiation Oncology, University of Texas Medicalranch, Galveston, TX 77555, USA
Department of Pathology, Baylor College of Medicine and Michael EeBakey Department of Veterans Affairs Medical Center, Houston, TX7030, USA
bstract—Human neural stem cells have the ability to differ-ntiate into all three major cell types in the CNS includingeurons, astrocytes and oligodendrocytes. The multipotencyf human neural stem cells shed a light on the possibility ofsing stem cells as a therapeutic tool for various neurologi-al disorders including neurodegenerative diseases and neu-otrauma that involve a loss of functional neurons. We haveiscovered previously a priming procedure to direct primarilyultured human neural stem cells to differentiate into almosture neurons when grafted into adult CNS. However, theolecular mechanism underlying this phenomenon is stillnknown. To unravel transcriptional changes of human neu-al stem cells upon priming, cDNA microarray was used totudy temporal changes in human neural stem cell genexpression profile during priming and differentiation. As aesult, transcriptional levels of 520 annotated genes wereetected changed in at least at two time points during theriming process. In addition, transcription levels of morehan 3000 hypothetical protein encoding genes and ESTenes were modulated during the priming and differentiationrocesses of human neural stem cells. We further analyzedhe named genes and grouped them into 14 functional cate-ories. Of particular interest, key cell signal transductionathways, including the G-protein-mediated signaling path-ays (heterotrimeric and small monomeric GTPase path-ays), the Wnt signaling pathway and the TGF-� pathway, areodulated by the neural stem cell priming, suggesting im-ortant roles of these key signaling pathways in priming andifferentiation of human neural stem cells. © 2005 IBRO.ublished by Elsevier Ltd. All rights reserved.
ey words: gene expression array, human neural stem cells,NSCs, stem cell priming and differentiation, cell signalingathways, GTP-binding proteins.
Corresponding author. Tel: �1-713-677-7505; fax: �1-713-677-7512.-mail address: [email protected] (M. Liu).bbreviations: bFGF, basic fibroblast growth factor; Ct, threshold cy-le; Cy3, cyanine 3; Cy5, cyanine 5; EST, expressed sequence tag;�12, G-protein alpha 12; hNSC, human neural stem cell; PBS,
shosphate-buffered saline; RAR, retinoic acid receptor; RGS6, G-rotein signaling 6.
306-4522/06$30.00�0.00 © 2005 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2005.11.041
133
he discovery of neurogenesis and the existence of neuraltem cells in certain regions of CNS convinced us that theeneration of new neurons does happen and is a processhere endogenous neural stem cells differentiate into
unctional neuronal subtypes (Altman and Das, 1965; Al-arez-Buylla and Garcia-Verdugo, 2002; Gage, 2000;empermann and Gage, 2000). Neural stem cells isolated
rom CNS could differentiate into all three types of cells,ncluding neurons, astrocytes and oligodendrocytes whenrafted into developing CNS or neurogenic areas of thedult CNS (Shihabuddin et al., 2000; Svendsen and Cald-ell, 2000; Svendsen and Smith, 1999). However, in non-eurogenic areas of the adult CNS, grafted neural stemells either remain undifferentiated or mainly become glialells (Cao et al., 2002). In vitro cell culture studies havehown that a small percentage of human neural stem cellshNSCs) differentiate into small unipolar or bipolarABAergic or glutamergic neurons while the majority of
hem become astroglial cells under a regular differentiationrocess by withdrawing growth factors (Wu et al., 2002).irectional differentiation of hNSCs into specific type ofeurons, such as cholinergic neurons, is then a critical
ssue to be addressed in our study using hNSCs as aotential therapeutic tool since many neurodegenerativeiseases result from the loss of functional neurons but notlial cells (Lindvall et al., 2004; McKay, 2004; Wu et al.,002).
To make stem cells applicable for cell therapy of neu-odegenerative diseases, an in vitro cell culture systemeeds to be established to induce neural stem cells toifferentiate into neurons in a maximum extent. Along this
ine, our group developed an in vitro priming method byreating fetal hNSCs with a cocktail of basic fibroblastrowth factor (bFGF), heparin and laminin (Wu et al.,002). This simple priming procedure resulted in a nearlyure population of neurons after being transplanted intoon-neurogenic areas of adult rat intact CNS (Wu et al.,002). This technique provides a potential advance oneural stem cell therapy to treat neurodegenerative dis-ases, such as Alzheimer’s and Lou Gehrig’s diseases.
The fact that primed hNSCs differentiate mainly intoeurons while unprimed hNSCs remain undifferentiated orecome astroglial cells when grafted into adult CNS,rompts us to suspect that the priming procedure may
nitiate complex patterns of transcriptional changes (regu-ation of gene expression and key signaling molecules) inNSCs that are different from those in unprimed cells.hus it is of great interest to study the overall gene expres-
ion profile during the process of priming for a betterved.

uhslptmWptAfqswccWmcmwmgfp
C
T8oap(FctbLL
imw0
P
T2nppabbvFuww(
I
Cp3XaaBCrglrcm(Nne
Ih
FlgIa2o0tfruC(er12hF10hHr
A
Ssd(Ghsstswtsof
Y. Cai et al. / Neuroscience 138 (2006) 133–148134
nderstanding of the mechanism underlying its effect onNSCs. The knowledge of the transcriptional program andignaling pathways involved in the priming process willead us to a more explicit view of the distinct expressionrofiles between neuronal and glial differentiation. Al-hough microarray analysis has been performed to studyouse neural progenitors’ differentiation (Luo et al., 2002;en et al., 2002), it is unclear what genes and signaling
athways play important roles in the priming processeshat lead to the generation of neurons from grafted hNSCs.dditionally, why would the priming processes be different
rom a regular differentiation of hNSCs? To answer theseuestions, we have analyzed the temporal gene expres-ion profile and changes in cell signal transduction path-ays of hNSCs upon priming and differentiation using theDNA microarray technique. Our data indicate that keyellular signaling pathways, such as G-protein pathways,nt signaling pathway, FGF and TGF-� pathways, areodulated during the priming and differentiation pro-
esses. Our study reported here is the first one to deter-ine temporal changes in transcription and signaling path-ays of hNSCs during priming and differentiation usingicroarray analysis. Such a systematic characterization oflobal transcriptional and signaling pathway changes of-ers an insight on possible molecular mechanisms of hNSCriming and differentiation into functional neurons.
EXPERIMENTAL PROCEDURES
ulture of hNSCs
he K048 line of hNSCs, originally derived from the forebrain of an-week human fetus, was provided by C. N. Svendsen, Universityf Wisconsin (Madison, WI, USA) (Svendsen et al., 1998; Wu etl., 2002). The culture was maintained according to our previousublication (Wu et al., 2002) with some modifications. Briefly, cells7–8�104/cm2) were cultured in a basic medium including DMEM:12 (3:1, Life Technologies), 15 mM HEPES (Sigma), 1.5% glu-ose (Sigma) and 1� penicillin/streptomycin (Sigma); which washen supplemented with 20 ng/ml EGF (R&D Systems), 10 ng/mlFGF (R&D Systems), 2.5 �g/ml heparin (Invitrogen), 10 ng/mlIF (Chemicon), N2 (Bottenstein and Sato, 1979) and 2 mM-glutamine (Sigma) (Bottenstein and Sato, 1979). Cells werencubated with 8.5% CO2 at 37 °C. About half of the growth
edium was changed every 3–4 days. Expanded neurospheresere dissociated into single cells once every 8–10 days with.025% Trypsin and mechanical trituration.
riming and differentiation of hNSCs
he FHL priming procedure was described previously (Wu et al.,002), which included hNSCs cultured adhesively with a combi-ation of bFGF, heparin and laminin. Two to three days post-assaging, hNSCs (passage 14–28) were seeded at 6�106 cells/er T75 culture flask pre-coated with 0.01% poly-D-lysine (Sigma)nd 0.5 �g/cm2 laminin (Invitrogen). Cells were incubated with aasic medium plus N2, and then supplemented with 20 ng/mlFGF, 5 �g/ml heparin and 1 �g/ml laminin (Invitrogen). A halfolume of medium was changed with fresh medium containingHL every 1–2 days. Cells were primed for the desired intervalsntil harvesting. For those cells to be differentiated further, cellsere first primed for 6 days, and then allowed differentiation byithdrawal of FHL and cultured in a basic medium with B27
Invitrogen) for 10 days. g
mmunofluorescent staining
ells were fixed with 4% paraformaldehyde. After several times ofhosphate-buffered saline (PBS) rinsing, cells were incubated for0 min in 3% normal serum/5% bovine serum albumin/0.1% Triton-100 in PBS. This blockage of nonspecific binding and perme-bilization was followed by an overnight incubation with primaryntibodies, mouse anti-class III �-tubulin (TuJ1) (1:4000, CovanceAbCo, Richmond, CA, USA) or mouse anti-human Nestin (1:200,. A. Messam, NIH) (Messam et al., 2000) at 4 °C. After three
inses with PBS for 20 min each, samples were incubated withoat anti-mouse IgG conjugated with Alexa 594 fluorophore (Mo-
ecular Probes, Eugene, OR, USA) at a 1:300 dilution for 1 h atoom temperature in the dark. After three 10-min rinses with PBS,ell nuclei were counterstained with 1 �g/ml DAPI (Sigma) for 5in at room temperature, and then mounted with Fluoromount G
Fisher) under glass coverslips. Images were captured with aikon Eclipse E1000 epifluorescent microscope, which was con-ected to a Photometrics Coolsnap FX Monochrome digital cam-ra and a MetaMorph processing system.
solation of total RNA and hybridization on bayloruman 18K chips
ollowing the removal of medium in the culture flasks, cells wereysed and the total RNA was extracted by RNeasy Midi kit (Qia-en, Valencia, CA) according to the manufacturer’s instruction.ntegrity and purity of RNA were checked by spectrophotometrynd RNA electrophoresis. The cDNA reverse transcription from0 �g total RNA and fluorescent labeling reactions were carriedut using cyanine 5 (Cy5) labeled nucleotides for the control daycDNA (unprimed control) and cyanine 3 (Cy3) labeled nucleo-
ides for experimental samples. Bacteria reference RNA was usedor internal hybridization controls. The mixture of Cy5 and Cy3eactions was purified and concentrated by PCR purification col-mns (Qiagen). Labeled cDNAs were eluted with 60 �l 2� SSC.hips were pre-hybridized for 45 min in pre-hybridization buffer
5� SSC, 0.1% SDS, 0.1% BSA) at 42 °C, washed with 100%thanol and deionized/distilled H2O once each to remove saltesidue, and then centrifuged in a 50 ml tube for 1 min at700 r.p.m. to remove excess liquid. cDNAs were denatured by a-minute incubation in boiling water before loading onto Bayloruman 18K Chips (Baylor College of Medicine Microarray Coreacility, Houston, TX, USA) and the hybridization was allowed for8 h at 65 °C in hybridization buffer (50% formamide, 10� SSC,.2%SDS) in Genomic Systems Hybridization station. Afterybridization and washing, chips were washed with ethanol and
2O and centrifuged for 1 min to remove the liquid and saltesidues.
nalysis of results
lides were scanned immediately in Axon 4000A dual channelcanner (Axon Instruments, Foster City, CA, USA). The originalata were then extracted by GenePix v. 4.01 Software packageAxon Instruments) and analyzed with Gene Spring v. 7.0 (Siliconenetics, Redwood City, CA, USA). Data with low signal intensity,igh background and high variability were labeled with flags. Arraypecific data normalization was performed using locally weightedcatterplot smoothing (LOWESS) method Berger et al., 2004. Ahree-step filtering was carried out to eliminate false positive re-ults. First, only data with flag absent in at least four of seven chipsere selected. Second, among these flag filtered genes, only
hose with normalized intensities at 1000 units or higher wereelected for further analysis. Finally only genes with two-fold changesr more were selected for data interpretation. t-Test was per-ormed using Gene Spring v.7.0 to validate the data. Functional
rouping was based on literature search (www.ncbi.nlm.nih.gov).
En
V
QteMitfd7cc2ts(iGcoftloc(R2
P
Hwtp(apapu1gi(susomrscbs
F6
Y. Cai et al. / Neuroscience 138 (2006) 133–148 135
ach block of the chips contains bacterial control spikes as inter-al controls.
erification of microarray data with RT-PCR
uantitative PCR was carried out by adding 5 �l of template cDNAo a final 25 �l reaction volume containing 3 mM MgCl2, 0.4 �M ofach forward and reverse primers, and 2.5 �l of LC-FastStart DNAaster Sybr Green 1 (Roche). Real-time PCR was done using the
Cycler instrument (Bio-Rad) with optimized PCR reaction condi-ions. The amplification of selected genes was carried out asollows: a 3-min hot start at 95 °C followed by 40 cycles ofenaturation at 95 °C for 30 s, annealing at 55 °C for 20 s, and a2 °C extension for 30 s. The amplification protocol for �-actin wasarried out as follows: a 3-min hot start at 95 °C followed by 40ycles of denaturation at 95 °C for 30 s, annealing at 56 °C for0 s, and a 72 °C extension for 30 s. Each time point includedhree (day 2, day 5, Pdf10 and Udf10) or four (day 0) individualamples for triplicate. Each assay included a negative controlwater instead of template cDNA), and the experiment was donen duplicate. The fluorescence emitted by the reporter (Sybrreen) dye was detected online in real time, and the thresholdycle (Ct) of each sample was recorded as a quantitative measuref the amount of PCR product in the sample. The Ct value is theractional cycle number at which the fluorescence generated byhe reporter dye exceeds a fixed level above baseline. The se-ected genes’ signals were normalized against the relative quantityf �-actin and expressed as �Ct�(Ctselected gene�Ct�-actin). Thehange in selected genes’ signals relative to the reference signalcontrol sample) was expressed as �� Ct�(�Ctcontrol��Ctsample).elative changes in expression were then calculated as[���Ct].
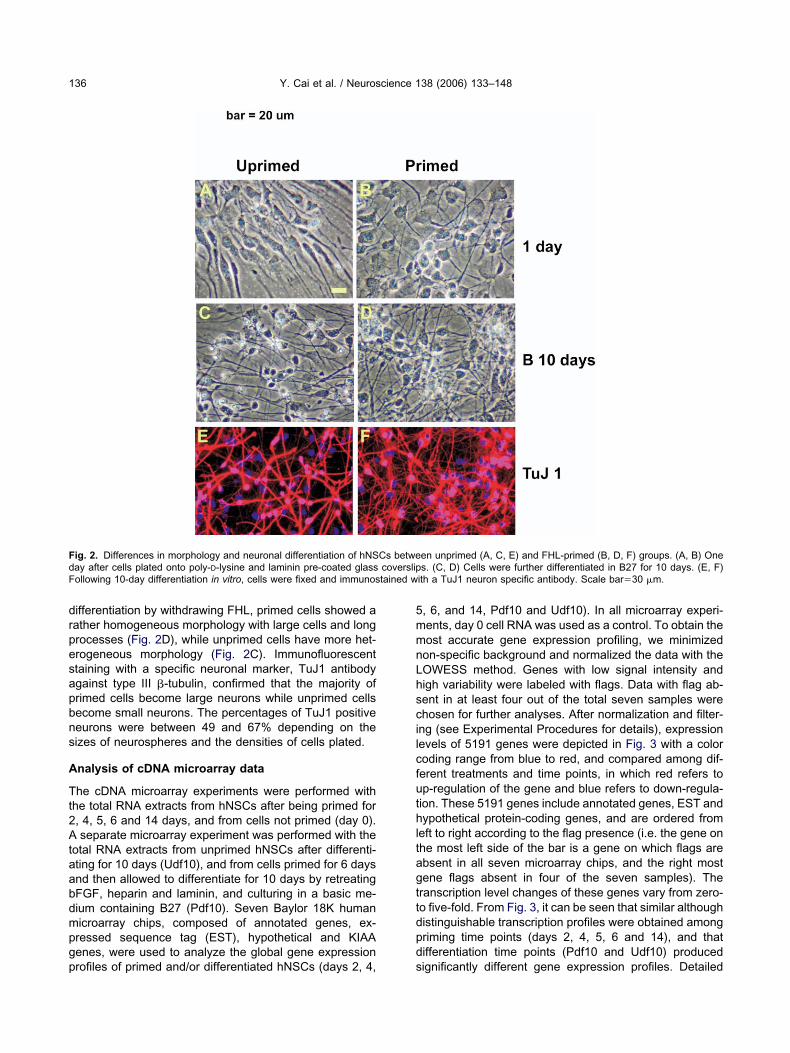
ig. 1. Time course of FHL-priming. Human fetal neural stem cells were primedays (FHL6d), and 14 days (FHL14d). Scale bar�50 �m.
RESULTS
riming and differentiation of hNSCs
uman fetal neural stem cells (hNSCs, passage 14–28)ere treated with the FHL-priming procedure (a combina-
ion of bFGF, heparin and laminin) that we developedreviously (Wu et al., 2002). At different time intervals1–14 days), total RNAs were collected for microarraynalyses. A subgroup of cells was subjected first to FHL-riming for 6 days, and then to a 10-day further differenti-tion by withdrawing FHL and cultivation in medium sup-lemented with B27 (Pdf10). Control groups includednprimed hNSCs only and unprimed cells followed by a0-day differentiation (Udf10). As reported previously by ourroup (Wu et al., 2002), hNSCs treated with FHL showed
mmediate spreading of cells from seeded neurospheresFig. 1). Most of the cells are large and round or square inhape (Figs. 1 and 2B) in contrast to the spindle shape of thenprimed cells (Fig. 2A). When priming prolongs, cellspread further without too much morphological changesver the period of 1–6 days post-priming. However, treat-ent with FHL for a longer period of time (days 11–14, Fig. 1)
esulted in cell round up and the reformation of neuro-pheres. The vast majority of these long-term FHL-treatedells showed immunoreactive with a specific nestin anti-ody, indicating that they are in a stem or progenitor celltage (Wu et al., 2003; data not shown). Following a further
d with bFGF, heparin and laminin for 1 day (FHL1d), 3 days (FHL3d),
drpesapbns
A
Tt2Ataabdmpgp
5mmnLhscilcfuthltagttdpd
Fd coverslipF tained w
Y. Cai et al. / Neuroscience 138 (2006) 133–148136
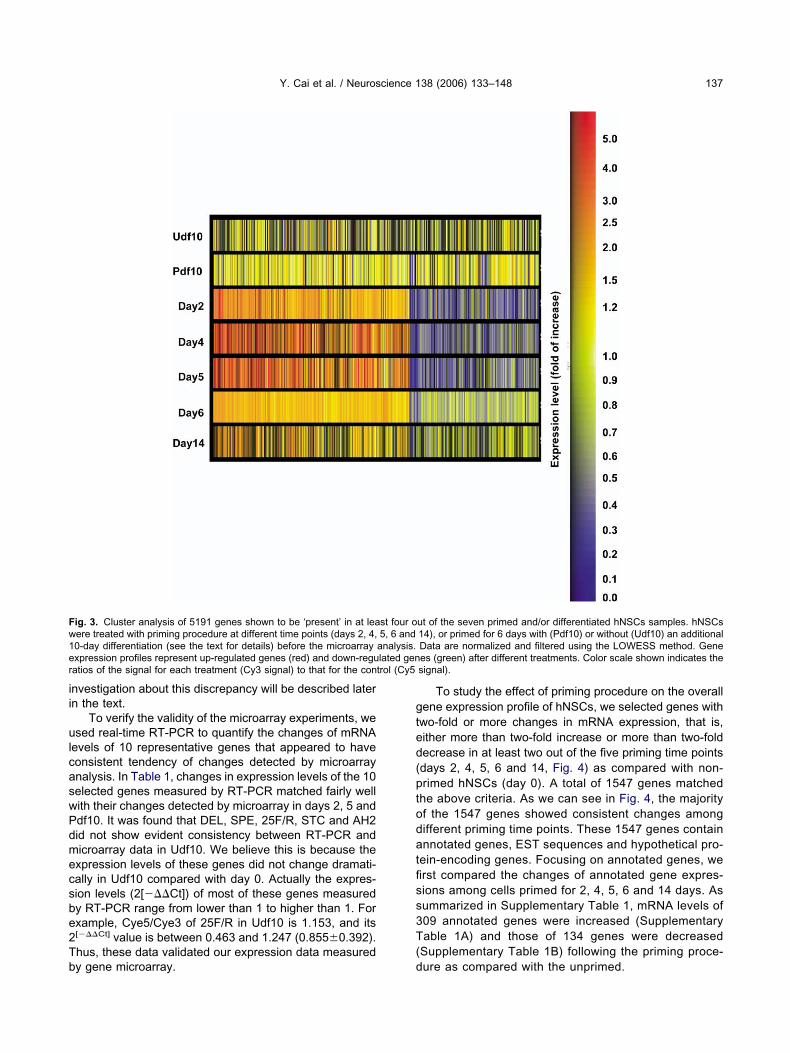
ifferentiation by withdrawing FHL, primed cells showed aather homogeneous morphology with large cells and longrocesses (Fig. 2D), while unprimed cells have more het-rogeneous morphology (Fig. 2C). Immunofluorescenttaining with a specific neuronal marker, TuJ1 antibodygainst type III �-tubulin, confirmed that the majority ofrimed cells become large neurons while unprimed cellsecome small neurons. The percentages of TuJ1 positiveeurons were between 49 and 67% depending on theizes of neurospheres and the densities of cells plated.
nalysis of cDNA microarray data
he cDNA microarray experiments were performed withhe total RNA extracts from hNSCs after being primed for, 4, 5, 6 and 14 days, and from cells not primed (day 0).separate microarray experiment was performed with the
otal RNA extracts from unprimed hNSCs after differenti-ting for 10 days (Udf10), and from cells primed for 6 daysnd then allowed to differentiate for 10 days by retreatingFGF, heparin and laminin, and culturing in a basic me-ium containing B27 (Pdf10). Seven Baylor 18K humanicroarray chips, composed of annotated genes, ex-ressed sequence tag (EST), hypothetical and KIAAenes, were used to analyze the global gene expression
ig. 2. Differences in morphology and neuronal differentiation of hNSay after cells plated onto poly-D-lysine and laminin pre-coated glassollowing 10-day differentiation in vitro, cells were fixed and immunos
rofiles of primed and/or differentiated hNSCs (days 2, 4, s
, 6, and 14, Pdf10 and Udf10). In all microarray experi-ents, day 0 cell RNA was used as a control. To obtain theost accurate gene expression profiling, we minimizedon-specific background and normalized the data with theOWESS method. Genes with low signal intensity andigh variability were labeled with flags. Data with flag ab-ent in at least four out of the total seven samples werehosen for further analyses. After normalization and filter-
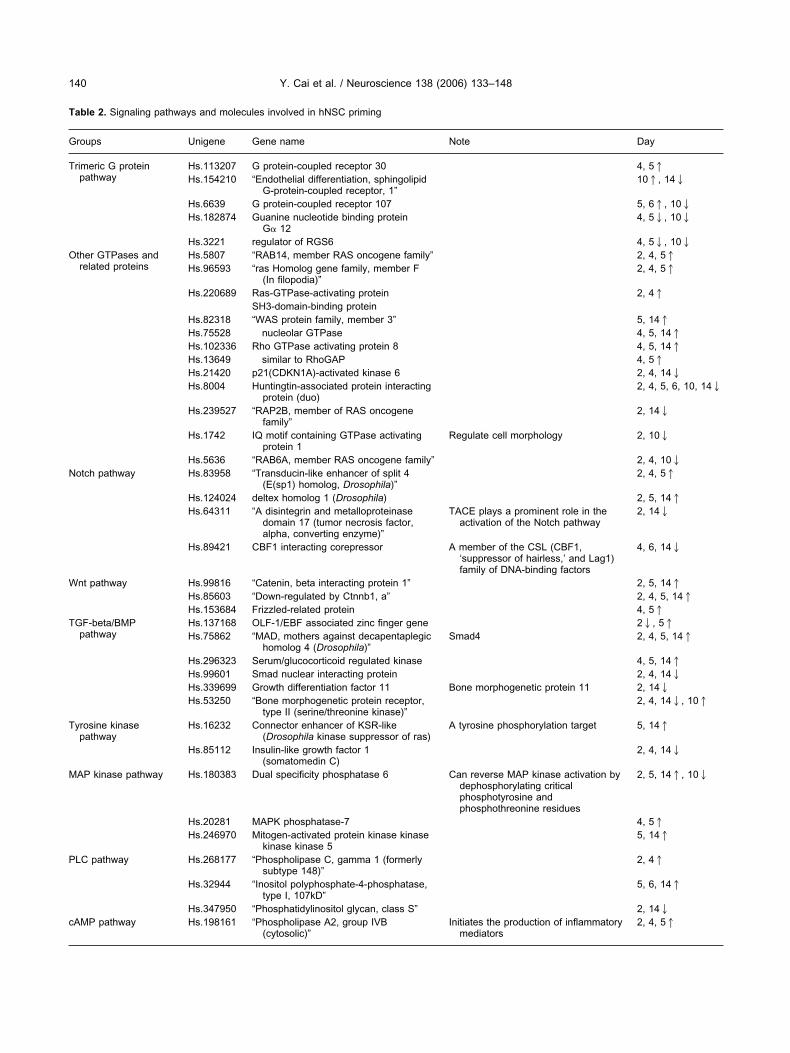
ng (see Experimental Procedures for details), expressionevels of 5191 genes were depicted in Fig. 3 with a coloroding range from blue to red, and compared among dif-erent treatments and time points, in which red refers top-regulation of the gene and blue refers to down-regula-ion. These 5191 genes include annotated genes, EST andypothetical protein-coding genes, and are ordered from
eft to right according to the flag presence (i.e. the gene onhe most left side of the bar is a gene on which flags arebsent in all seven microarray chips, and the right mostene flags absent in four of the seven samples). Theranscription level changes of these genes vary from zero-o five-fold. From Fig. 3, it can be seen that similar althoughistinguishable transcription profiles were obtained amongriming time points (days 2, 4, 5, 6 and 14), and thatifferentiation time points (Pdf10 and Udf10) produced
en unprimed (A, C, E) and FHL-primed (B, D, F) groups. (A, B) Ones. (C, D) Cells were further differentiated in B27 for 10 days. (E, F)
ith a TuJ1 neuron specific antibody. Scale bar�30 �m.
Cs betwe
ignificantly different gene expression profiles. Detailed
ii
ulcaswPdmecsbe2Tb
gted(ptodatfiss3T(
Fw1e lated genr trol (Cy5
Y. Cai et al. / Neuroscience 138 (2006) 133–148 137
nvestigation about this discrepancy will be described latern the text.
To verify the validity of the microarray experiments, wesed real-time RT-PCR to quantify the changes of mRNA
evels of 10 representative genes that appeared to haveonsistent tendency of changes detected by microarraynalysis. In Table 1, changes in expression levels of the 10elected genes measured by RT-PCR matched fairly wellith their changes detected by microarray in days 2, 5 anddf10. It was found that DEL, SPE, 25F/R, STC and AH2id not show evident consistency between RT-PCR andicroarray data in Udf10. We believe this is because thexpression levels of these genes did not change dramati-ally in Udf10 compared with day 0. Actually the expres-ion levels (2[���Ct]) of most of these genes measuredy RT-PCR range from lower than 1 to higher than 1. Forxample, Cye5/Cye3 of 25F/R in Udf10 is 1.153, and its[���Ct] value is between 0.463 and 1.247 (0.8550.392).hus, these data validated our expression data measured
ig. 3. Cluster analysis of 5191 genes shown to be ‘present’ in at leaere treated with priming procedure at different time points (days 2, 4,0-day differentiation (see the text for details) before the microarrayxpression profiles represent up-regulated genes (red) and down-reguatios of the signal for each treatment (Cy3 signal) to that for the con
y gene microarray. d
To study the effect of priming procedure on the overallene expression profile of hNSCs, we selected genes withwo-fold or more changes in mRNA expression, that is,ither more than two-fold increase or more than two-foldecrease in at least two out of the five priming time pointsdays 2, 4, 5, 6 and 14, Fig. 4) as compared with non-rimed hNSCs (day 0). A total of 1547 genes matched
he above criteria. As we can see in Fig. 4, the majorityf the 1547 genes showed consistent changes amongifferent priming time points. These 1547 genes containnnotated genes, EST sequences and hypothetical pro-
ein-encoding genes. Focusing on annotated genes, werst compared the changes of annotated gene expres-ions among cells primed for 2, 4, 5, 6 and 14 days. Asummarized in Supplementary Table 1, mRNA levels of09 annotated genes were increased (Supplementaryable 1A) and those of 134 genes were decreased
Supplementary Table 1B) following the priming proce-
ut of the seven primed and/or differentiated hNSCs samples. hNSCs14), or primed for 6 days with (Pdf10) or without (Udf10) an additionalData are normalized and filtered using the LOWESS method. Genees (green) after different treatments. Color scale shown indicates thesignal).
st four o5, 6 andanalysis.
ure as compared with the unprimed.

So
OopitlgtatepacdNop
EG
Ir(eta
rctardbradPdo
Oi
Bhiniepbaihamatr(1pdihpdddicPritp
ci
ceespo
T
G
DCSD2SITAD
G
DCSD2SITAD
Y. Cai et al. / Neuroscience 138 (2006) 133–148138
ignal transduction pathways involved in primingf hNSCs
ne focus of our study is to characterize a complex seriesf signaling programs potentially involved in the primingrocess of hNSCs. A better understanding of these signal-
ng pathways at a cellular level will be of great assistanceo the elucidation of the mechanisms underlying stem cellineage specification and to the discovery of potential tar-ets for stem cell therapy, gene therapy and pharmaceu-icals. In our study, mRNA levels of 43 transcription factorsnd DNA binding proteins were changed at two or moreime points during priming and differentiation. One hundredleven signal transduction pathway components or relatedroteins were changed in their mRNA expression levels int least two time points. These signaling moleculeshanged during hNSC priming were classified into 11 well-efined pathways, including G-protein signaling, Wnt,otch, and TGF-� pathways (Table 2). Brief discussionsn each of these signaling pathways involved in hNSCriming processes are given below.
ffects of hNSC priming on heterotrimeric-protein-coupled signaling pathways
t has been reported that heterotrimeric G-protein-coupledeceptors regulated asymmetric cell division, cell polarityKnoblich, 2001; Cai et al., 2005) and stem cell differ-ntiation (Strubing et al., 1997), which are fundamental
o neural stem cell differentiation. According to our micro-
able 1. Verification of cDNA microarray data by real-time qPCR
ene D2 D5
Cye5/Cye3 2[���Ct] Cye5/Cye3 2[���Ct]
EL 3.268 3.1931.63 2.909 3.5841.04TN 3.323 5.3082.05 2.564 1.5240.433PE 1.741 1.920.518 2.525 2.9790.748RCT 2.973 3.3051.733 2.65 2.0340.8425F/R 2.597 1.3580.429 2.571 1.6720.183TC 0.251 0.3370.174 0.317 0.1330.074
GF 0.399 0.50.197 0.83 0.6440.119A3 0.24 0.3490.163 0.219 0.8020.102H2 2.963 4.3941.394 2.394 6.3841.759uo 0.148 0.3250.115 0.139 0.3850.172
ene Pdf10 Udf10
Cye5/Cye3 2[���Ct] Cye5/Cye3 2[���Ct]
EL 1.083 1.3580.729 1.405 0.780.134TN 1.143 2.310.928 2.1 2.1060.671PE 1.363 1.5240.388 0.871 1.1550.287RCT 1.692 1.1680.193 1.078 1.2240.4985F/R 1.271 1.0530.682 1.153 0.8550.392TC 0.88 0.0710.049 1.299 0.690.38
GF 0.564 0.3870.131 2 1.6240.823A3 0.834 0.450.19 1.246 1.0590.245H2 1.318 1.9350.538 0.468 1.3560.686uo 0.22 0.4320.122 1.937 2.4551.31
rray data, expression levels of two G-protein-coupled a
eceptors (GPCR 30 and GPCR 107) (Table 2) werehanged at several time points, indicating a potential func-ion of these two receptors in the priming process. Inddition, heterotrimeric G-protein alpha 12 (G�12) and theegulator of G-protein signaling 6 (RGS6) were decreaseduring the priming and differentiation processes. It haseen reported that G�12 and RGS6 were involved in neu-onal differentiation of PC12 and Neuro 2A cell lines (Liu etl., 2002; Sarma et al., 2003). In contrast, we detectedecreased mRNA levels of G�12 and RGS6 at day 4 anddf10 of hNSCs. This discrepancy may result from theifferences in cell types and culturing conditions imposedn cells.
ther GTPases and their signaling pathways arenvolved in hNSC priming
esides heterotrimeric G protein, other types of GTPasesave also been shown to regulate cell fate determination
n certain tissues and systems. As shown in Table 2, aumber of small GTPases and their regulators are either
ncreased or decreased during hNSC priming and differ-ntiation. For example, Rab14, Ras-GTPase-activatingrotein SH3 domain-binding protein, WASP family mem-er 3, nucleolar GTPase, Rho GAP, and Rho GTPase-ctivating protein 8 (RhoGAP8) were all increased dur-
ng priming and differentiation processes. On the otherand, p21-activated protein kinase 6 (PAK6), huntingtin-ssociated protein interacting protein (Duo), Rap2b, IQotif containing GTPase activating protein 1 (IQGAP1),nd Rab6A were all decreased during priming or differen-iation. It has been demonstrated that GTPases play a keyole in neural differentiation and cell fate determinationArakawa et al., 2003; Bryan et al., 2004; Liu and Jessell,998). For example, activation of Rho family GTPasesromotes neural differentiation, neurite outgrowth, andendritic spine formation in primary cultured neurons and
n neuroblastoma cells (Bryan et al., 2004). On the otherand, in response to IGF-1 stimulation, Rho GTPases and130 Rho GAP were shown to be essential in cell fateetermination during adipogenesis and myogenesis (Sor-ella et al., 2003). The expression of Rab3 was increaseduring differentiation of bone marrow macrophages (BMMs)
nto osteoclasts (Abu-Am et al., 1999). Our data indi-ated that the expression levels of several small GT-ases (Rho, Rab, and Ras family members) and their
egulators (GAPs and GEFs) were changed during prim-ng of hNSCs (Table 2), suggesting important roles ofhese GTPases and their signaling pathways during theriming processes of hNSCs.
AMP and retinoic acid signaling pathwaysn hNSC priming
AMP/CREB pathway is a key regulator of neuronal differ-ntiation and plasticity (Beyer and Karolczak, 2000; Gintyt al., 1994; Heasley et al., 1991; Piper et al., 2000). Ashown in Table 2, the expression levels of several cAMPathway-related genes were modulated during the primingf hNSCs, both phospholipase A2 and A kinase (PRKA)
nchor protein 2 (Paralemmin 2) are upregulated while
PsaeA(ioctd
rcfc(nrsm
Nh
Nu(ctwTgtldprg
vd
FS g time pou ted gene
Y. Cai et al. / Neuroscience 138 (2006) 133–148 139
KA regulatory subunit II was down-regulated. It has beenhown that phospholipase A2 induced neurite outgrowthnd neuronal differentiation in Rat PC12 cells (Nakashimat al., 2003), and the regulation of secreted phospholipase2 is associated with differentiation of murine epidermis
Gurrieri et al., 2003). cAMP and cAMP-dependent signal-ng pathways have been shown to play a key role in neuriteutgrowth and differentiation (Sanchez et al., 2004; Sta-howiak et al., 2003; Tojima et al., 2003). Our data indicatehat the priming process of hNSCs regulates cAMP-depen-ent signaling pathway.
Hormonal and nuclear receptors such as retinoic acideceptors (RAR) and peroxisome proliferator-activated re-eptor- (PPAR-) have dramatic effects on neuronal dif-erentiation and the regeneration of different tissues, in-luding neurogenesis, adipogenesis, and hematopoiesisCollins, 2002). As shown in Table 2, the mRNA levels of aumber of genes involved in retinoic acid and other nucleareceptor pathways are mostly up-regulated during priming,uggesting that priming procedure does affect the RAR-
ig. 4. Cluster analyses of 1547 genes with normalized expression levelected genes were detected in at least two out of the five priminp-regulation (red) or down-regulation (green) by priming. The annota
ediated pathway in hNSCs. s
otch and Wnt signaling pathways are involved inNSC priming
otch signaling pathway has been reported widely to reg-late cell fate determination, cell polarity and differentiationOhishi et al., 2002). As shown in Table 2, Notch pathwayomponents were regulated differently. For example, theranscription factor enhancer of split and the ligand deltexere upregulated while CBF1 interacting corepressor andACE were down-regulated during priming of hNSCs, sug-esting that priming could regulate different components ofhe Notch signaling pathway at different times. It is be-ieved that Notch pathway inhibits a specific pathway ofifferentiation and induces differentiation along alternativeathways and that Notch pathway is involved in asymmet-ic cell division and cell polarity (Le Borgne and Schweis-uth, 2003; Ohishi et al., 2002, 2003).
Wnt signaling pathway has been reported to be in-olved in extensive biological processes such as animalevelopment, oncogenesis, cell polarity establishment and
ased (red) or decreased (blue) by two-fold or more in priming hNSCs.ints (days 2, 4, 5, 6 and 14). Gene expression patterns representlist of these genes is shown in Supplementary Table 1.
els incre
o on (Peifer and Polakis, 2000; Shulman et al., 1998; van
T
G
T
O
N
W
T
T
M
P
c
Y. Cai et al. / Neuroscience 138 (2006) 133–148140
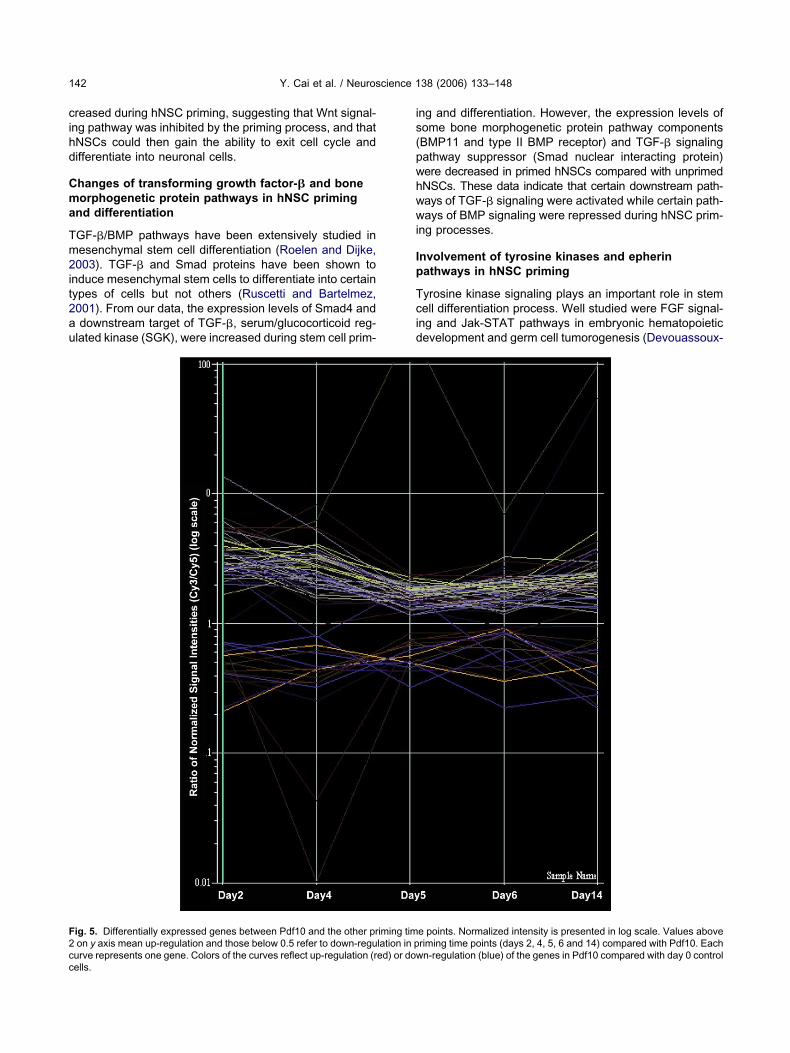
able 2. Signaling pathways and molecules involved in hNSC priming
roups Unigene Gene name Note Day
rimeric G proteinpathway
Hs.113207 G protein-coupled receptor 30 4, 51Hs.154210 “Endothelial differentiation, sphingolipid
G-protein-coupled receptor, 1”101, 142
Hs.6639 G protein-coupled receptor 107 5, 61, 102Hs.182874 Guanine nucleotide binding protein
G� 124, 52, 102
Hs.3221 regulator of RGS6 4, 52, 102ther GTPases andrelated proteins
Hs.5807 “RAB14, member RAS oncogene family” 2, 4, 51Hs.96593 “ras Homolog gene family, member F
(In filopodia)”2, 4, 51
Hs.220689 Ras-GTPase-activating protein 2, 41SH3-domain-binding protein
Hs.82318 “WAS protein family, member 3” 5, 141Hs.75528 nucleolar GTPase 4, 5, 141Hs.102336 Rho GTPase activating protein 8 4, 5, 141Hs.13649 similar to RhoGAP 4, 51Hs.21420 p21(CDKN1A)-activated kinase 6 2, 4, 142Hs.8004 Huntingtin-associated protein interacting
protein (duo)2, 4, 5, 6, 10, 142
Hs.239527 “RAP2B, member of RAS oncogenefamily”
2, 142
Hs.1742 IQ motif containing GTPase activatingprotein 1
Regulate cell morphology 2, 102
Hs.5636 “RAB6A, member RAS oncogene family” 2, 4, 102otch pathway Hs.83958 “Transducin-like enhancer of split 4
(E(sp1) homolog, Drosophila)”2, 4, 51
Hs.124024 deltex homolog 1 (Drosophila) 2, 5, 141Hs.64311 “A disintegrin and metalloproteinase
domain 17 (tumor necrosis factor,alpha, converting enzyme)”
TACE plays a prominent role in theactivation of the Notch pathway
2, 142
Hs.89421 CBF1 interacting corepressor A member of the CSL (CBF1,‘suppressor of hairless,’ and Lag1)family of DNA-binding factors
4, 6, 142
nt pathway Hs.99816 “Catenin, beta interacting protein 1” 2, 5, 141Hs.85603 “Down-regulated by Ctnnb1, a” 2, 4, 5, 141Hs.153684 Frizzled-related protein 4, 51
GF-beta/BMPpathway
Hs.137168 OLF-1/EBF associated zinc finger gene 22, 51Hs.75862 “MAD, mothers against decapentaplegic
homolog 4 (Drosophila)”Smad4 2, 4, 5, 141
Hs.296323 Serum/glucocorticoid regulated kinase 4, 5, 141Hs.99601 Smad nuclear interacting protein 2, 4, 142Hs.339699 Growth differentiation factor 11 Bone morphogenetic protein 11 2, 142Hs.53250 “Bone morphogenetic protein receptor,
type II (serine/threonine kinase)”2, 4, 142, 101
yrosine kinasepathway
Hs.16232 Connector enhancer of KSR-like(Drosophila kinase suppressor of ras)
A tyrosine phosphorylation target 5, 141
Hs.85112 Insulin-like growth factor 1(somatomedin C)
2, 4, 142
AP kinase pathway Hs.180383 Dual specificity phosphatase 6 Can reverse MAP kinase activation bydephosphorylating criticalphosphotyrosine andphosphothreonine residues
2, 5, 141, 102
Hs.20281 MAPK phosphatase-7 4, 51Hs.246970 Mitogen-activated protein kinase kinase
kinase kinase 55, 141
LC pathway Hs.268177 “Phospholipase C, gamma 1 (formerlysubtype 148)”
2, 41
Hs.32944 “Inositol polyphosphate-4-phosphatase,type I, 107kD”
5, 6, 141
Hs.347950 “Phosphatidylinositol glycan, class S” 2, 142AMP pathway Hs.198161 “Phospholipase A2, group IVB
(cytosolic)”Initiates the production of inflammatory
mediators2, 4, 51

dWflpsWW(
ib2(r2(l
T
G
R
E
O
Y. Cai et al. / Neuroscience 138 (2006) 133–148 141
e Wetering et al., 2002). As shown in Table 2, all threent pathway genes, beta-catenin interacting protein 1,
rizzled-related protein (SFRP), and a gene down-regu-ated by Ctnnb1 (Drctnnb1a), were up-regulated during theriming process. It is widely believed that activation of Wntignaling pathway induced self-renewal of stem cells, andnt proteins acted as cell growth factors (Reya, 2003;illert et al., 2003). Beta-catenin interacting protein one
able 2. Continued
roups Unigene Gene name
Hs.42322 A kinase (PRKA) anchor pro
Hs.365523 “Protein kinase, cAMP-deperegulatory, type II, alpha”
etinoic acid and othernuclear receptorpathways
Hs.325081 GRASP protein
Hs.74002 Nuclear receptor coactivator
Hs.183858 Transcriptional intermediaryHs.110849 Estrogen-related receptor alHs.118138 “Nuclear receptor subfamily
member 2”Hs.250505 “Retinolc acid receptor, alphHs.15589 PPAR binding protein
phrin pathway Hs.89839 EphA1
Hs.125124 EphB2Hs.30942 Ephrin-B2
ther differentiation-related molecules
Hs.36137 “Hepatocyte nuclear factor 3Hs.54452 “Zinc finger protein, subfami
1 (lkaros)”
Hs.32425 Early B-cell factor
Hs.55967 Short stature homeobox 2
Hs.78061 Transcription factor 21
Hs.29397 “Zinc finger protein, subfami5 (Pegasus)”
Hs.99348 Distal-less hormobox 5Hs.23823 Halry/enhancer-of-split relate
YRPW motif-likeHs.22030 Paired box gene 5 (B-cell lin
specific activator protein)
Hs.241558 Ariadne homolog 2 (Drosoph
Hs.198 Paired box gene 3 (Waardensyndrome 1)
Hs.3844 LIM domain only 4Hs.137064 Cytoplasmic polyadenylation
binding protein
CTNNBIP1) negatively regulates the Wnt pathway by inhib- d
ting the interaction of beta-catenin with Tcf4 and repressingeta-catenin-Tcf4-mediated transactivation (Tago et al.,000). The expression level of down-regulated by Ctnnb1 aDrctnnb1a) was dramatically increased by a reduction ofepressing beta-catenin expression (Kawasoe et al.,000). And furthermore, secreted frizzled-related proteinsSFRP) inhibit paracrine WNT/frizzled signaling in over-oaded myocardium (Schumann et al., 2000). From our
Note Day
Essential for dissemination of cAMPsignals in highly polarizedepithelium in association withF-actin
2, 4, 5, 141
2, 142
A retinoic acid-induced gene.GRP1-associated scaffold protein
2, 4, 5, 141
Enhances the transcriptional activity ofmultiple hormone receptors in thepresence of ligands
2, 4, 5, 61
51, 1022, 4, 51
1, 4, 5, 141
4, 516, 142
Mediates developmental events,particularly in the nervous system
2, 5, 141
4, 51Ephrin ligand 4, 51
” 2, 4, 5, 141May play an important role in CD4
versus CD8 lineage commitmentdecisions
2, 4, 51
Participates in the regulation of thepre-B and B lymphocyte-specificMB1 gene
2, 4, 5, 141
Involved in pattern formation in bothinvertebrate and vertebrate species
2, 4, 51
Important in cell fate determinationand tissue differentiation in theembryo
2, 4, 51
Regulator of lymphocyte differentiation 4, 51
4, 512, 5, 141
A transcription factor that is expressedat early, but not late, stages ofB-cell differentiation duringembryogenesis
10, 141
Up-regulated during retinoicacid-induced granulocyticdifferentiation of acuteleukemia cells
2, 4, 5, 6, 141
2, 41
4, 142Controls germ cell differentiation by
regulating the formation ofsynaptonemal complexes
2, 51
tein 2
ndent,
1
factor 1pha1, group
a”
, gammaly 1A,
ly 1A,
d with
eage
ila)
burg
element
ata, the expression of all these three genes was in-
cihd
Cma
Tm2it2au
is(pwhwwi
Ip
Tcid
F2cc
Y. Cai et al. / Neuroscience 138 (2006) 133–148142
reased during hNSC priming, suggesting that Wnt signal-ng pathway was inhibited by the priming process, and thatNSCs could then gain the ability to exit cell cycle andifferentiate into neuronal cells.
hanges of transforming growth factor-� and boneorphogenetic protein pathways in hNSC priming
nd differentiation
GF-�/BMP pathways have been extensively studied inesenchymal stem cell differentiation (Roelen and Dijke,003). TGF-� and Smad proteins have been shown to
nduce mesenchymal stem cells to differentiate into certainypes of cells but not others (Ruscetti and Bartelmez,001). From our data, the expression levels of Smad4 anddownstream target of TGF-�, serum/glucocorticoid reg-
lated kinase (SGK), were increased during stem cell prim-
ig. 5. Differentially expressed genes between Pdf10 and the other pron y axis mean up-regulation and those below 0.5 refer to down-regu
urve represents one gene. Colors of the curves reflect up-regulation (red) or dowells.
ng and differentiation. However, the expression levels ofome bone morphogenetic protein pathway componentsBMP11 and type II BMP receptor) and TGF-� signalingathway suppressor (Smad nuclear interacting protein)ere decreased in primed hNSCs compared with unprimedNSCs. These data indicate that certain downstream path-ays of TGF-� signaling were activated while certain path-ays of BMP signaling were repressed during hNSC prim-
ng processes.
nvolvement of tyrosine kinases and epherinathways in hNSC priming
yrosine kinase signaling plays an important role in stemell differentiation process. Well studied were FGF signal-
ng and Jak-STAT pathways in embryonic hematopoieticevelopment and germ cell tumorogenesis (Devouassoux-
e points. Normalized intensity is presented in log scale. Values aboveriming time points (days 2, 4, 5, 6 and 14) compared with Pdf10. Each
iming timlation in p
n-regulation (blue) of the genes in Pdf10 compared with day 0 control

SKteirtthiirGttahePttam
oag
Ek
MdtZptficphwbpaap
T
G
T
CE
S
LM
T
R
R
TC
Y. Cai et al. / Neuroscience 138 (2006) 133–148 143
hisheboran et al., 2003; Moroni et al., 2002; Nosaka anditamura, 2000). During hNSC priming, several protein
yrosine kinase related proteins displayed changes in theirxpression (Table 2, tyrosine kinase pathway). Particularly
nteresting are that the expression levels of two ephrineceptors, Eph A1 and Eph B2, are increased at differentime points, suggesting that ephrin pathway might be ac-ivated and play an important role in the priming process ofNSCs. Both microarray data and microscopic images
ndicated that neurite outgrowth and cell migration, the twomportant steps in hNSC differentiation into functional neu-ons, happened under the influence of priming process.rowth factors, cytokines, and their receptors are impor-
ant regulators of stem cell differentiation and proliferationhrough autocrine and paracrine functions (Tarasenko etl., 2004). Abnormal signaling of these pathways leads toematopoietic disorders, tumorigenesis, and neuronal dis-ases (Biscardi et al., 2000; Dawbarn and Allen, 2003;unt, 1992). Moreover, Ephrin receptor kinases constitute
he largest family of receptors tyrosine kinases. Eph recep-ors and the downstream signaling pathways may not play
role in cell proliferation, but have important functions inultiple biological processes such as cell migration, ax-
able 3. Genes changed in opposite directions during priming and di
roups Unigene Gene name
ranscription factor and DNAbinding protein
Hs.21599 Kruppel-like fa
ytoskeleton related Hs.167 Microtubule-asCM and cell adhesion Hs.3314 “Selenoprotein
Hs.287820 Fibronectin 1Hs.188401 Annexin A10
ignaling Hs.125790 Leuclne-rich reHs.23643 Mst3 and SOKHs.274382 “Protein kinaseHs.247302 Twisted gastruHs.180383 Dual specificityHs.136309 SH3-domain GHs.183858 Transcriptional
igands Hs.170310 “Cat eye syndretabolism Hs.1869 Phosphoglucom
Hs.77910 3-Hydroxy-3-mHs.79345 “Coagulation faHs.251871 CTP synthase
ranslation and proteinprocessing
Hs.177486 “Amyloid betadisease)”
Hs.173125 Peptidylprolyl IHs.211539 “Eukaryotic tra
NA binding, transcription andprocessing
Hs.169900 “Poly(A) bindinHs.54780 “TranscriptionHs.118194 Debranching e
eceptor and membrane protein Hs.154210 “Endothelial difHs.6639 G protein-coupHs.53250 “Bone morphog
kinase)”Hs.50984 Sarcoma amplHs.22180 5-HydroxytryptHs.5422 glycoprotein MHs.199067 v-erb-b2 Erythr
ransporter Hs.169902 “Solute carrier
nal guidance and embryonic development (Bruckner etl., 1997; Drescher et al., 2000; Flanagan and Vanderhae-hen, 1998; Pasquale et al., 1997).
ffect of hNSC priming on mitogen-activated proteininase (MAPK) and Phospholipase C (PLC) pathways
APK pathway is one of the earliest studied signal trans-uction pathways, regulating cell proliferation and differen-iation in a complex pattern (Seger and Krebs, 1995;hang and Liu, 2002). During neural stem cell primingrocesses, the expression levels of mitogen-activated pro-ein kinase kinase kinase kinase 5 (MEKK5), dual speci-city phosphatase 6, and MAPK phosphatase-7 were in-reased at different time points of priming, suggesting thatriming process induces hNSCs for differentiation by in-ibiting the self-renewal of hNSCs through MAPK path-ay. The MAPK pathway can be coupled to and activatedy many other signaling pathways, such as tyrosine kinaseathway and G-protein-coupled receptor pathway (Segernd Krebs, 1995; Zhang and Liu, 2002). Recently, Uddin etl. (2004) reported that specific isoforms of p38 MAPKromoted the late-stage differentiation of primary erythroid
on
Day
iquitous) 101, 142
protein 2 42, 101a, 1” 22, 101
52, 1012, 142, 101
taining 2 2, 142, 101kinase 2, 142, 101
on-inducible double-stranded RNA dependent” 22, 101101, 142
tase 6 2, 5, 141, 102endophilin B1 101, 142iary factor 1 51, 102mosome region, candidate 1” 2, 142, 101
2, 142, 101ryl-coenzyme A synthase 1 (soluble) 4, 141, 102procoagulant component (hemophilia A)” 101, 142
51, 102ursor protein (protease nexin-II, Alzheimer 41, 102
F (cyclophilin F) 41, 102nitiation factor 2, subunit 3 (gamma, 52kD)” 22, 101, cytoplasmic 4 (inducible form)” 2, 41, 102n factor, RNA polymerase I” 2, 142, 101molog 1 (S. cerevisiae) 101, 142
on, sphingolipid G-protein-coupled receptor, 1” 101, 142tor 107 5, 61, 102
rotein receptor, type II (serine/threonine 2, 4, 142, 101
ence 2, 142, 101rotonin) receptor 6 22, 101
22, 101leukemia viral oncogene homolog 3 (avian) 22, 101(facilitated glucose transporter), member 1” 2, 4, 51, 102
fferentiati
ctor 7 (ub
sociatedP, plasm
peal-con1-related, interferlationphospha
RB2-likeintermed
ome chroutase 1
ethylglutactor VIII,
(A4) prec
somerasenslation Ig proteinterminationzyme hoferentiatiled recepenetic p
ified sequamine (se6Boblasticfamily 2
ell cycle Hs.175043 Cyclin M4 2, 142, 101
pt2
ieca
Tacphbew
FitrU
Y. Cai et al. / Neuroscience 138 (2006) 133–148144
rogenitors. Activation of Erk signaling pathway is inhibi-ory to self-renewal of embryonic stem cells (Burdon et al.,002).
The PLC pathways are important for many physiolog-cal and pathological processes. The two products of PLCnzymes, IP3 and DAG, are key regulators of intracellularalcium concentration, which is essential for cell growthnd differentiation (Whitfield et al., 1995). As shown in
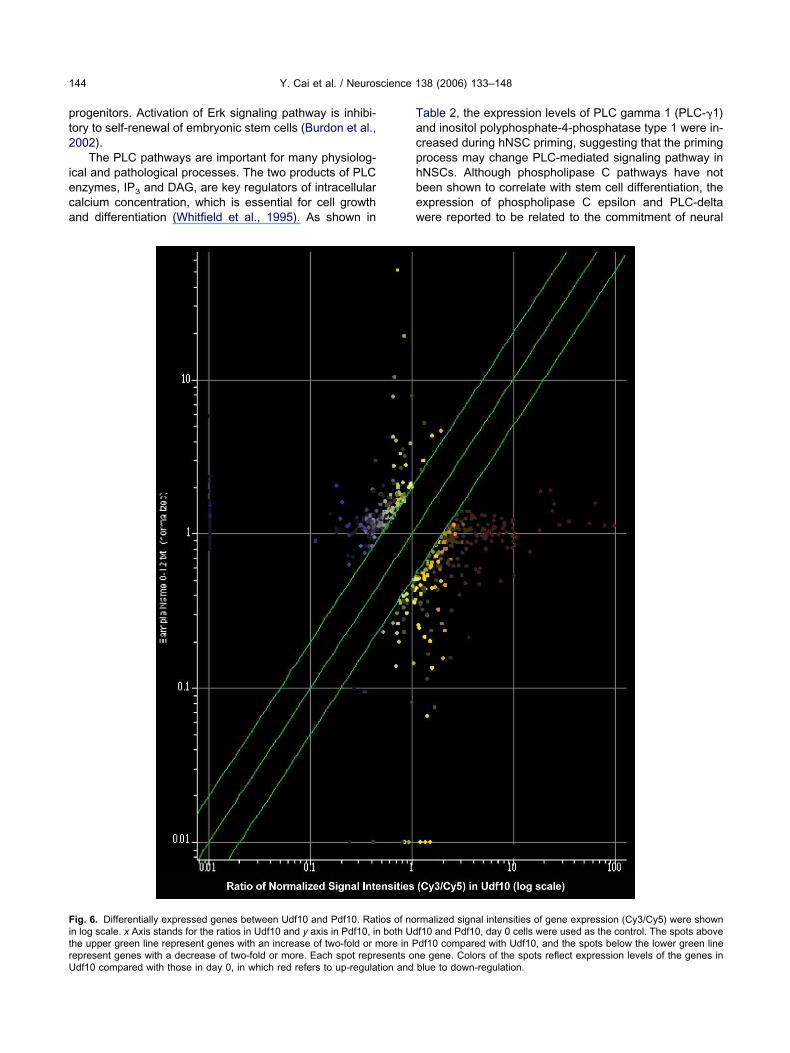
ig. 6. Differentially expressed genes between Udf10 and Pdf10. Ratn log scale. x Axis stands for the ratios in Udf10 and y axis in Pdf10, inhe upper green line represent genes with an increase of two-fold or m
epresent genes with a decrease of two-fold or more. Each spot represents ondf10 compared with those in day 0, in which red refers to up-regulation andable 2, the expression levels of PLC gamma 1 (PLC-1)nd inositol polyphosphate-4-phosphatase type 1 were in-reased during hNSC priming, suggesting that the primingrocess may change PLC-mediated signaling pathway inNSCs. Although phospholipase C pathways have noteen shown to correlate with stem cell differentiation, thexpression of phospholipase C epsilon and PLC-deltaere reported to be related to the commitment of neural
rmalized signal intensities of gene expression (Cy3/Cy5) were shownf10 and Pdf10, day 0 cells were used as the control. The spots abovedf10 compared with Udf10, and the spots below the lower green line
ios of noboth Udore in P
e gene. Colors of the spots reflect expression levels of the genes inblue to down-regulation.

pm
Pg
Arnuitditdwmfa
Dh
Tott
Afpal0te(dgsf
Dh
Pccnhudprp
T
G
T
O
JNW
T
R
O
Y. Cai et al. / Neuroscience 138 (2006) 133–148 145
recursor cells and skin stem cells, respectively (Naka-ura et al., 2003; Wu et al., 2003).
riming of hNSCs up-regulates differentiation relatedenes/molecules
s shown in Table 2, most of the genes that have beeneported to play a role in differentiation process in botheuronal cells and non-neuronal cells have been up-reg-lated during the priming processes of hNSCs. Interest-
ngly, the ortholog of mouse integral membrane glycopro-ein LIG-1, which has been shown to play a role in glialifferentiation and development, was down-regulated dur-
ng priming process of hNSCs, suggesting that the primingreatment of hNSCs down-regulated genes involved in glialifferentiation and development. These results correlateell with our previous data that primed hNSCs differentiateainly into neurons while unprimed hNSCs remain undif-
erentiated or become astroglial cells when grafted intodult CNS.
ifferent gene expression patterns between primedNSCs and Pdf10 cells
o study the effect of the additional 10-day differentiationn primed hNSCs, we compared the microarray data ob-ained from Pdf10 with those from primed cells at differentime points (days 2, 4, 5, 6 and 14) without differentiation.
able 4. Signaling pathways differentially regulated between Udf10 a
roups Genbank
rimeric G protein pathway AA775047
AA779406N34362N93476
ther GTPases and related proteins AA043653AA454558N74995R34013
AK STAT pathway R23241otch Pathway AA733033nt pathway AA486838
AA478738AA486067R83038
yrosine kinase pathway AA446994AA134862
etinoic acid and other nuclear receptor pathways AA074201R38548
ther differentiation-related molecules AA151708N50828N48089H82431AA703652AA465719H42504H84264AA460466
s shown in Fig. 5, expression levels of 67 genes wereound to be changed more than two-fold in Pdf10 com-ared with primed cells in at least two time points. Gener-lly, curves above two on y axis refer to genes up-regu-
ated in corresponding priming time points and those below.5 refer to genes down-regulated. Furthermore, we foundhat the mRNA levels of 32 annotated genes were differ-ntially regulated between Pdf10 and the primed cellsTable 3). Such observation is predictable since a furtherifferentiation after a priming procedure most likely affectsene expression patterns, which corresponds well with theignificant morphological differences between primed andurther differentiated hNSCs (Fig. 2).
ifferential gene expression profiles in differentiatedNSC with or without priming
reviously, we have shown that primed hNSCs could be-ome large multipolar cholinergic neurons while unprimedells become only small unipolar or bipolar GABAergiceurons in vitro (Wu et al., 2002). Furthermore, primedNSCs become pure neurons when grafted in CNS whilenprimed hNSCs differentiate mainly into glial cells. Theiscrepancies in differentiation of hNSCs with and withoutriming suggest that the priming process may differentiallyegulate gene expression through distinct cellular signalingathways. Along this line, we compared the microarray
change Gene name
042 Guanine nucleotide binding protein (G protein), alphainhibiting activity polypeptide 2
996 G protein beta subunit-likeRegulator of G-protein signaling 5
203 Endothelial differentiation, sphingolipidG-protein-coupled receptor, 1
006 RAS protein activator-like 2379 ADP-ribosylation factor 1 GTPase activating protein306 RAB35, member RAS oncogene family491 RAB40C, member RAS oncogene family074 STAT-induced STAT inhibitor-20856 Notch homolog 1, translocation-associated779 Secreted frizzled-related protein 417 Catenin, beta interacting protein 138 Transducin (beta)-like 1X-linked49 Low density lipoprotein receptor-related protein 5025 Fibroblast growth factor receptor 4294 Insulin receptor substrate 2207 Nuclear receptor subfamily 2, group F, member 6474 Retinoic acid induced 16035 Homeobox (expressed in ES cells) 1236 Zinc-fingers and homeoboxes 1763 Ring finger protein (C3H2C3 type) 6124 Prospero-related homeobox 1223 Slit homolog 3 (Drosophila)247 Intersectin 2386 Musashi homolog 1 (Droaophila)403 Sine oculis homeobox homolog 3 (Drosophila)424 Cyclin-dependent kinase 5, regulatory subunit 1 (p35)
nd Pdf10
Fold
2.
2.151
0.
2.3.0.0.2.0.2.0.0.0.2.0.0.0.2.2.2.4.0.0.0.0.0.
AI364369 0.483 Pre-B-cell leukemia transcription factor 1

daubc
dmadagprgUrcstcc
lartaUdtipiPpttu
ItafiiupficaGSCddsp
petasmt
AfRtI
A
A
A
A
B
B
B
B
B
B
B
C
C
C
D
D
D
F
Y. Cai et al. / Neuroscience 138 (2006) 133–148146
ata between two types of hNSCs: 1) primed for 6 daysnd then differentiated for 10 days (Pdf10) and 2)nprimed but differentiated for 10 days (Udf10). Again, inoth sets of data, day 0 stem cells were used as theontrol, i.e. hNSCs neither being primed nor differentiated.
To elucidate the differential gene expression profiles inifferentiated hNSCs with or without priming, normalizedicroarray data of Pdf10 and Udf10 were compared andnalyzed. As shown in Fig. 6, 523 genes were found to beifferentially regulated by two-fold or more between Pdf10nd Udf10. Dots above the upper green line stand forenes up-regulated by more than two-fold in Pdf10 com-ared with Udf10, and those below the lower green lineeflect genes decreased in Pdf10. A full list of annotatedenes that are differentially regulated between Pdf10 anddf10 is presented in Supplementary Table 2. To elucidate
egulation of the signaling pathways that confers to the dis-repancy between Pdf10 and Udf10, we analyzed theignaling molecules that were expressed differently be-ween these two groups. As shown in Table 4, unprimedells (Udf10) have a distinct signaling regulation patternompared with the primed cells (Pdf10).
Multiple signaling pathways were differentially regu-ated between Pdf10 and Udf10. GTPases, Wnt, Notchnd JAK-STAT pathways, known to play very importantoles in stem cell differentiation and/or cell fate determina-ion (Hoang, 2004; Nosaka and Kitamura, 2000; Radtke etl., 2004), were found differentially changed betweendf10 and Pdf10. For example, Notch 1 was found to beown-regulated in Udf10 when compared with Pdf10. Fur-
hermore, expression levels of several Wnt pathway inhib-ting genes were found to be decreased in Udf10 com-ared with Pdf10. These data indicate that the physiolog-
cal and morphological difference between Udf10 anddf10 may stem from regulation of these distinct signalingathways. In other words, different phenotypic differentia-
ion between primed and unprimed hNSCs may result fromhe recruitment of different molecule clusters and the reg-lation of different signaling pathways.
DISCUSSION
n this study, we have analyzed the gene expression pat-erns and cell signaling pathways during hNSC primingnd differentiation using cDNA microarray technique. Werst examined the changes in gene expression levels dur-ng priming process of hNSCs and then compared cellsndergoing differentiation with and without the primingrocesses. Real-time quantitative PCR was used for veri-cation of microarray data. Signaling molecules/proteinshanged during the priming process of hNSCs were char-cterized into 11 subgroups and pathways, including the-protein pathway, the Wnt/Notch pthway, the TGF-�/mad pathway, and the growth factor receptor pathway.orrelating to the different morphological and phenotypicifferentiations, hNSCs with or without priming showedistinct patterns in overall gene expression and specificignaling pathways. The knowledge of the transcriptional
rogram and signaling patterns involved in the primingrocess will lead us to a more explicit view of the distinctxpression profiles between neuronal and glial differentia-ion. This is the first temporal study on gene expressionlterations of hNSCs priming and differentiation andhould shed light on our future characterization of theolecular mechanism of hNSC differentiation into func-
ional neurons.
cknowledgments—This work is partially supported by a grantrom the Mission Connect of the Institute for Rehabilitation andesearch (TIRR) Foundation and a grant (5R01HL064792) from
he National Institutes of Health/National Heart, Lung, and Bloodnstitute to M. Liu.
REFERENCES
bu-Am Y, Teitelbaum SL, Chappel JC, Schlesinger P, Ross FP(1999) Expression and regulation of RAB3 proteins in osteoclastsand their precursors. J Bone Miner Res 14:1855–1860.
ltman J, Das GD (1965) Autoradiographic and histological evidenceof postnatal hippocampal neurogenesis in rats. J Comp Neurol124:319–335.
lvarez-Buylla A, Garcia-Verdugo JM (2002) Neurogenesis in adultsubventricular zone. J Neurosci 22:629–634.
rakawa Y, Bito H, Furuyashiki T, Tsuji T, Takemoto-Kimura S,Kimura K, Nozaki K, Hashimoto N, Narumiya S (2003) Control ofaxon elongation via an SDF-1alpha/Rho/mDia pathway in culturedcerebellar granule neurons. J Cell Biol 161:381–391.
erger JA, Hautaniemi S, Jarvinen AK, Edgren H, Mitra SK, Astola J(2004) Optimized LOWESS normalization parameter selection forDNA microarray data. BMC Bioinformatics 5:194
eyer C, Karolczak M (2000) Estrogenic stimulation of neurite growthin midbrain dopaminergic neurons depends on cAMP/protein ki-nase A signalling. J Neurosci Res 59:107–116.
iscardi JS, Ishizawar RC, Silva CM, Parsons SJ (2000) Tyrosinekinase signalling in breast cancer: epidermal growth factor receptorand c-Src interactions in breast cancer. Breast Cancer Res2:203–210.
ottenstein JE, Sato GH (1979) Growth of a rat neuroblastoma cell linein serum-free supplemented medium. Proc Natl Acad Sci U S A76:514–517.
ruckner K, Pasquale EB, Klein R (1997) Tyrosine phosphorylation oftransmembrane ligands for Eph receptors. Science 275:1640–1643.
ryan B, Kumar V, Stafford LJ, Cai Y, Wu G, Liu M (2004) GEFT, aRho-family guanine nucleotide exchange factor, regulates neuriteoutgrowth and dendritic spine formation. J Biol Chem 279:45824–45832.
urdon T, Smith A, Savatier P (2002) Signalling, cell cycle and pluri-potency in embryonic stem cells. Trends Cell Biol 12:432–438.
ai Y, Stafford LJ, Bryan BA, Mitchell D, Liu M (2005) G-protein-activated phospholipase C-beta, new partners for cell polarity pro-teins Par3 and Par6. Oncogene 24:4293–4300.
ao Q, Benton RL, Whittemore SR (2002) Stem cell repair of centralnervous system injury. J Neurosci Res 68:501–510.
ollins SJ (2002) The role of retinoids and retinoic acid receptors innormal hematopoiesis. Leukemia 16:1896–1905.
awbarn D, Allen SJ (2003) Neurotrophins and neurodegeneration.Neuropathol Appl Neurobiol 29:211–230.
evouassoux-Shisheboran M, Mauduit C, Tabone E, Droz JP,Benahmed M (2003) Growth regulatory factors and signalling pro-teins in testicular germ cell tumours. Apmis 111:212–224; discus-sion 224.
rescher U (2000) Excitation at the synapse: Eph receptors team upwith NMDA receptors. Cell 103:1005–1008.
lanagan JG, Vanderhaeghen P (1998) The ephrins and Eph recep-
tors in neural development. Annu Rev Neurosci 21:309–345.
G
G
G
H
H
K
K
K
L
L
L
L
L
M
M
M
N
N
N
O
O
P
P
P
P
R
R
R
R
S
S
S
S
S
S
S
S
S
S
S
S
T
T
T
U
Y. Cai et al. / Neuroscience 138 (2006) 133–148 147
age FH (2000) Mammalian neural stem cells. Science 287:1433–1438.
inty DD, Bonni A, Greenberg ME (1994) Nerve growth factor acti-vates a Ras-dependent protein kinase that stimulates c-fos tran-scription via phosphorylation of CREB. Cell 77:713–725.
urrieri S, Furstenberger G, Schadow A, Haas U, Singer AG, Gho-mashchi F, Pfeilschifter J, Lambeau G, Gelb MH, Kaszkin M (2003)Differentiation-dependent regulation of secreted phospholipasesA2 in murine epidermis. J Invest Dermatol 121:156–164.
easley LE, Benedict S, Gleavy J, Johnson GL (1991) Requirement ofthe adenovirus E1A transformation domain 1 for inhibition of PC12cell neuronal differentiation. Cell Regul 2:479–489.
oang T (2004) The origin of hematopoietic cell type diversity. Onco-gene 23:7188–7198.
awasoe T, Furukawa Y, Daigo Y, Nishiwaki T, Ishiguro H, Fujita M,Satoh S, Miwa N, Nagasawa Y, Miyoshi Y, Ogawa M, Nakamura Y(2000) Isolation and characterization of a novel human gene,DRCTNNB1A, the expression of which is down-regulated by beta-catenin. Cancer Res 60:3354–3358.
empermann G, Gage FH (2000) Neurogenesis in the adult hip-pocampus. Novartis Found Symp 231:220–235; discussion 235–241, 302–306.
noblich JA (2001) Asymmetric cell division during animal develop-ment. Nat Rev Mol Cell Biol 2:11–20.
e Borgne R, Schweisguth F (2003) Unequal segregation of Neural-ized biases Notch activation during asymmetric cell division. DevCell 5:139–148.
indvall O, Kokaia Z, Martinez-Serrano A (2004) Stem cell therapy forhuman neurodegenerative disorders: how to make it work. NatMed 10 (Suppl):S42–S50.
iu JP, Jessell TM (1998) A role for rhoB in the delamination of neuralcrest cells from the dorsal neural tube. Development 125:5055–5067.
iu Z, Chatterjee TK, Fisher RA (2002) RGS6 interacts with SCG10and promotes neuronal differentiation. Role of the G gamma sub-unit-like (GGL) domain of RGS6. J Biol Chem 277:37832–37839.
uo Y, Cai J, Liu Y, Xue H, Chrest FJ, Wersto RP, Rao M (2002)Microarray analysis of selected genes in neural stem and progen-itor cells. J Neurochem 83:1481–1497.
cKay RD (2004) Stem cell biology and neurodegenerative disease.Philos Trans R Soc Lond B Biol Sci 359:851–856.
essam CA, Hou J, Major EO (2000) Coexpression of nestin in neuraland glial cells in the developing human CNS defined by a human-specific anti-nestin antibody. Exp Neurol 161:585–596.
oroni E, Dell’Era P, Rusnati M, Presta M (2002) Fibroblast growthfactors and their receptors in hematopoiesis and hematologicaltumors. J Hematother Stem Cell Res 11:19–32.
akamura Y, Fukami K, Yu H, Takenaka K, Kataoka Y, Shirakata Y,Nishikawa S, Hashimoto K, Yoshida N, Takenawa T (2003) Phos-pholipase Cdelta1 is required for skin stem cell lineage commit-ment. EMBO J 22:2981–2991.
akashima S, Ikeno Y, Yokoyama T, Kuwana M, Bolchi A, Ottonello S,Kitamoto K, Arioka M (2003) Secretory phospholipases A2 induceneurite outgrowth in PC12 cells. Biochem J 376:655–666.
osaka T, Kitamura T (2000) Janus kinases (JAKs) and signal trans-ducers and activators of transcription (STATs) in hematopoieticcells. Int J Hematol 71:309–319.
hishi K, Katayama N, Shiku H, Varnum-Finney B, Bernstein ID(2003) Notch signalling in hematopoiesis. Semin Cell Dev Biol14:143–150.
hishi K, Varnum-Finney B, Bernstein ID (2002) The notch pathway:modulation of cell fate decisions in hematopoiesis. Int J Hematol75:449–459.
asquale EB (1997) The Eph family of receptors. Curr Opin Cell Biol9:608–615.
eifer M, Polakis P (2000) Wnt signaling in oncogenesis andembryogenesis: a look outside the nucleus. Science 287:1606–
1609.iper DR, Mujtaba T, Rao MS, Lucero MT (2000) Immunocytochem-ical and physiological characterization of a population of culturedhuman neural precursors. J Neurophysiol 84:534–548.
unt CJ (1992) Regulation of hematopoietic cell function by proteintyrosine kinase-encoding oncogenes: a review. Leuk Res 16:551–559.
adtke F, Wilson A, Mancini SJ, MacDonald HR (2004) Notch regu-lation of lymphocyte development and function. Nat Immunol5:247–253.
eya T (2003) Regulation of hematopoietic stem cell self-renewal.Recent Prog Horm Res 58:283–295.
oelen BA, Dijke P (2003) Controlling mesenchymal stem cell differ-entiation by TGFBeta family members. J Orthop Sci 8:740–748.
uscetti FW, Bartelmez SH (2001) Transforming growth factor beta,pleiotropic regulator of hematopoietic stem cells: potential physio-logical and clinical relevance. Int J Hematol 74:18–25.
anchez S, Jimenez C, Carrera AC, Diaz-Nido J, Avila J, Wandosell F(2004) A cAMP-activated pathway, including PKA and PI3K, reg-ulates neuronal differentiation. Neurochem Int 44:231–242.
arma T, Voyno-Yasenetskaya T, Hope TJ, Rasenick MM (2003)Heterotrimeric G-proteins associate with microtubules during dif-ferentiation in PC12 pheochromocytoma cells. FASEB J 17:848–859.
chumann H, Holtz J, Zerkowski HR, Hatzfeld M (2000) Expression ofsecreted frizzled related proteins 3 and 4 in human ventricularmyocardium correlates with apoptosis related gene expression.Cardiovasc Res 45:720–728.
eger R, Krebs EG (1995) The MAPK signaling cascade. FASEB J9:726–735.
hihabuddin LS, Horner PJ, Ray J, Gage FH (2000) Adult spinal cordstem cells generate neurons after transplantation in the adult den-tate gyrus. J Neurosci 20:8727–8735.
hulman JM, Perrimon N, Axelrod JD (1998) Frizzled signaling andthe developmental control of cell polarity. Trends Genet 14:452–458.
ordella R, Jiang W, Chen GC, Curto M, Settleman J (2003) Modula-tion of Rho GTPase signaling regulates a switch between adipo-genesis and myogenesis. Cell 113:147–158.
tachowiak EK, Fang X, Myers J, Dunham S, Stachowiak MK (2003)cAMP-induced differentiation of human neuronal progenitor cells ismediated by nuclear fibroblast growth factor receptor-1 (FGFR1).J Neurochem 84:1296–1312.
trubing C, Rohwedel J, Ahnert-Hilger G, Wiedenmann B, HeschelerJ, Wobus AM (1997) Development of G protein-mediated Ca2�channel regulation in mouse embryonic stem cell-derived neurons.Eur J Neurosci 9:824–832.
vendsen CN, Caldwell MA (2000) Neural stem cells in the developingcentral nervous system: implications for cell therapy through trans-plantation. Prog Brain Res 127:13–34.
vendsen CN, Smith AG (1999) New prospects for human stem-celltherapy in the nervous system. Trends Neurosci 22:357–364.
vendsen CN, ter Borg MG, Armstrong RJ, Rosser AE, Chandran S,Ostenfeld T, Caldwell MA (1998) A new method for the rapid andlong term growth of human neural precursor cells. J NeurosciMethods 85:141–152.
ago K, Nakamura T, Nishita M, Hyodo J, Nagai S, Murata Y, AdachiS, Ohwada S, Morishita Y, Shibuya H, Akiyama T (2000) Inhibitionof Wnt signaling by ICAT, a novel beta-catenin-interacting protein.Genes Dev 14:1741–1749.
arasenko YI, Yu Y, Jordan PM, Bottenstein J, Wu P (2004) Effect ofgrowth factors on proliferation and phenotypic differentiation ofhuman fetal neural stem cells. J Neurosci Res 78:625–636.
ojima T, Kobayashi S, Ito E (2003) Dual role of cyclic AMP-depen-dent protein kinase in neuritogenesis and synaptogenesis duringneuronal differentiation. J Neurosci Res 74:829–837.
ddin S, Ah-Kang J, Ulaszek J, Mahmud D, Wickrema A (2004)
Differentiation stage-specific activation of p38 mitogen-activated
v
W
W
W
W
W
Z
S
S
Y. Cai et al. / Neuroscience 138 (2006) 133–148148
protein kinase isoforms in primary human erythroid cells. Proc NatlAcad Sci U S A 101:147–152.
an de Wetering M, de Lau W, Clevers H (2002) WNT signaling andlymphocyte development. Cell 109 (Suppl):S13–S19.
en T, Gu P, Minning TA, Wu Q, Liu M, Chen F, Liu H, Huang H(2002) Microarray analysis of neural stem cell differentiation in thestriatum of the fetal rat. Cell Mol Neurobiol 22:407–416.
hitfield JF, Bird RP, Chakravarthy BR, Isaacs RJ, Morley P (1995)Calcium-cell cycle regulator, differentiator, killer, chemopreventor,and maybe, tumor promoter. J Cell Biochem Suppl 22:74–91.
illert K, Brown JD, Danenberg E, Duncan AW, Weissman IL, Reya T,Yates JR 3rd, Nusse R (2003) Wnt proteins are lipid-modified andcan act as stem cell growth factors. Nature 423:448–452.
u D, Tadano M, Edamatsu H, Masago-Toda M, Yamawaki-Kataoka Y,
Terashima T, Mizoguchi A, Minami Y, Satoh T, Kataoka T (2003) tNeuronal lineage-specific induction of phospholipase Cepsilonexpression in the developing mouse brain. Eur J Neurosci17:1571–1580.
u P, Tarasenko YI, Gu Y, Huang LY, Coggeshall RE, Yu Y (2002)Region-specific generation of cholinergic neurons from fetal hu-man neural stem cells grafted in adult rat. Nat Neurosci 5:1271–1278.
hang W, Liu HT (2002) MAPK signal pathways in the regulation of cellproliferation in mammalian cells. Cell Res 12:9–18.
APPENDIX
upplementary data
upplementary data associated with this article can be found, in
he online version, at doi: 10.1016/j.neuroscience.2005.11.041.(Accepted 8 November 2005)(Available online 18 January 2006)
Top Related
Copyright © 2022 FDOKUMEN

