







Bahasa
Halaman
Hukum


Forest Management and Carbon Sequestration in Size-Structured Forests:
The Case of Pinus Sylvestris in Spain
Renan Ulrich Goetz a
Natali Hritonenko b
Ruben Mur a
Àngels Xabadia a
Yuri Yatsenko c
a University of Girona, Campus Montilivi, s/n, 17071 Girona, Spain; [email protected]
(corresponding author), Phone: +34 972-418719, Fax: +34 972-418032
b Prairie View A&M University, Box 519, Prairie View, Texas, USA; [email protected]
c Houston Baptist University, 7502 Fondren Road, Houston, Texas, USA; [email protected]
Abstract 1
The Kyoto Protocol allows Annex I countries to deduct carbon sequestered by land use, land-use 2 change and forestry from their national carbon emissions. Thornley and Cannell (2000) demonstrated 3 that the objectives of maximizing timber and carbon sequestration are not complementary. Based on 4 this finding, this paper develops a model that takes into account the underlying biophysical and 5 economic processes, and determines empirically the optimal selective management regime. The results 6 show that sequestration costs are significantly lower than previous estimates. 7
8
Key words: Kyoto Protocol, selective logging, carbon management, dynamic optimization. 9 10 JEL classification: C61, Q23, Q54. 11
12 Acknowledgements: 13 The authors gratefully acknowledge the support of the Ministerio de Ciencia y Tecnología grant 14 AGL2007-65548, INIA grant SUM 2006-00019-C02-01, and Generalitat de Catalunya grants 15 (XREPP, and 2005 SGR213).16

1
1. Introduction 17
Pioneering work by Adams and Ek (1974), Haight et al. (1985), Haight (1987), and Getz and 18
Haight (1989) numerically determined the optimal selective-logging regime of an uneven-19
aged forest. More recently, several authors extended this line of work by considering not only 20
the net benefits of timber management but also of carbon sequestration. Many of these studies 21
were conducted to determine the costs of carbon sequestration at a national or international 22
level (Adams et al. 1999; Sohngen and Mendelsohn 2003). The decision variables used in the 23
studies included the afforested land, the length of tree rotation and the management intensity. 24
The variable rotation length has been commonly used to assess interactions between timber 25
production and carbon sequestration (van Kooten et al. 1995; Stainback and Alavalapati 26
2002). However, the variable management intensity can only be chosen from a fixed set of 27
alternatives when trees are planted, and cannot be changed while there are standing trees. 28
Hence, these studies did not provide an answer to the question of whether it is optimal to 29
harvest part of the forest during the rotation or to harvest all trees at the end of the rotation 30
length. Likewise, the optimal partial harvest and its timing require a detailed stand analysis. 31
Pohjola and Valsta (2007) analyzed these questions empirically but did not provide a 32
theoretical model and the necessary conditions for its solution. The paper by Pohjola and 33
Valsta (2007) presents an important step forward for the detailed stand analysis method but it 34
does not consider the fact that the individual trees compete among each other for scarce 35
resources such as light, space and nutrients, i.e., it does not take into account the effect of 36
density. In addition, this article contributes to the present literature on stand management by 37
considering sequestration of carbon not only in timber but also in wood products and forest 38
soil. Finally, the paper integrates further the economic and biophysical model so that 39
decisions (logging) taken during the dynamic optimization path affect the future evolution of 40
the forest and vice versa, i.e., the evolution of the forest affects future logging patterns. 41

2
The results of the study show that carbon sequestration costs for forests seem to be 42
significantly lower when the forest management regime is changed than when land use is 43
changed, i.e., a change from agriculture to forestry. Therefore, the study suggests reanalyzing 44
the role of forest carbon sequestration as a part of climate change policies within the context 45
of changing the management regime rather than changing land use. 46
47
1.1 Features of this study 48
Since the tree density (trees per hectare) and the distribution of the size within the stand 49
(Keyfitz 1968) affect the biological growth of the individual trees, the number of planted trees 50
should be determined endogenously and the effect of intraspecific competition should be 51
considered in the model. If the forest reproduces naturally, the density of the forest can be 52
regulated by thinning, i.e., by the selective logging of younger trees. Xabadia and Goetz 53
(2007) and Goetz et al. (2007) determined the optimal selective-logging regime for timber 54
production of a size-structured forest in which planting and intraspecific competition were 55
taken into account; they did not, however, consider the costs and benefits of carbon 56
sequestration. 57
The modeling of a size-structured forest can include the fact that the price of timber increases 58
with the size of the trees because the timber of larger trees can be used for higher-value 59
products, such as furniture. Consequently, the price of timber can be considered as a function 60
of the size of the tree. This aspect is not only important for correctly determining the net 61
benefits of timber production but also for accurately measuring the permanence of the 62
sequestered carbon in the final product. This study explicitly takes into account the different 63
permanence times and analyzes their economic and biological effect on the optimal logging 64
regime. 65

3
Another important part of the carbon cycle is the carbon sequestered in the soil. The potential 66
of forest soils to sequester carbon is well known (Rasse et al. 2001). Matthews (1993), for 67
instance, reports that the change from agricultural land to forest land led, after 200 years, to 68
an increase in soil carbon from 30 t C/ha initially to 70 t C/ha. 69
Hence, the density of the stand, intraspecific competition, the size of the tree, and soil carbon 70
are essential for an accurate description of the biological growth process of the tree (partial 71
integrodifferential equation), of the life cycle of carbon in the different compartments of the 72
forest, and for the correct modeling of economic factors. 73
These factors have been considered previously on their own but not together in a single study. 74
With respect to the numerical determination of the optimal logging regime, the seminal 75
contributions by Adams and Ek (1974), Haight et al. (1985), Haight (1987) and Getz and 76
Haight (1989) describe the evolution of an age (size) distributed forest by a set of difference 77
equations in time for a number of age (size) cohorts. Since the evolution of the forest over 78
time is continuous, the set of difference equations presents an approximation (discretization 79
over time and age) of the underlying biological processes. The discretization procedure 80
applied has shown to be realistic and has been employed widely; however, the set of 81
difference equations has been set up ad hoc and is not derived from the underlying partial 82
differential equation. Therefore, little can be said about the size of the error in the 83
approximation. This paper attempts to overcome this shortcoming by deriving a set of 84
difference equations from a partial integrodifferential equation, as proposed in the 85
mathematical biology literature. In a numerical analysis, de Roos (1988) shows that the 86
derived set of difference equations can be considered a good approximation of the underlying 87
partial integrodifferential equation. Moreover, the use of a single partial integrodifferential 88
equation allows the economic and biophysical model to be integrated to a high degree. The 89
previously published models incorporated data obtained from stand projection tables or stand 90

4
projection programs (biophysical model). However, partial harvesting affects the future 91
growth of the forest, so that the data must be updated whenever trees are logged. Hence, the 92
modified evolution of the forest affects the choice of the optimal partial harvesting regime. 93
The mutual interdependency of the economic and biophysical model requires the future 94
evolution of the forest within the economic decision model to be continuously updated. The 95
interlocking of the economic and biophysical model is only possible if the biophysical 96
component of the economic model not only consists of data but also of processes that govern 97
the evolution of the forest. The set of difference equations employed in the previous literature 98
can only be integrated in the economic model if the parameters of all difference equations are 99
estimated simultaneously for a sufficiently large variety of initial size distributions. However, 100
applicability may be limited since this process would require estimating a set of usually more 101
than 10 difference equations subject to constraints that restrict the movement of the trees 102
between the difference equations (cohorts). Hence, apart from the possible approximation 103
error, it already seems more reasonable from a practical point of view to estimate a single 104
partial integrodifferential equation than a large set of difference equations. 105
The paper is organized as follows: the next section describes the dynamics of a forest, and 106
presents the economic decision problem; an empirical analysis is conducted in Section 3 to 107
determine the optimal selective-logging regime of a privately owned forest with respect to 108
timber and carbon sequestration; and conclusions are presented in Section 4. 109

5
2. The bioeconomic model 110
The dynamics of the bioeconomic model reflects the change in the density of the size-111
structured trees x(t,l) over calendar time t and size l, and also the changes in soil carbon s(t) 112
over time. Before we can specify the precise mathematical presentation of these two 113
equations, we need to introduce some notation. 114
The size of the tree is measured by the diameter at breast height and is denoted by l. The set 115
of the admissible values of the variable l consists of the interval [l0, lm], where l0 corresponds 116
to the minimum vital diameter of the tree and lm is the maximum diameter that the trees can 117
reach. In the case of a completely managed forest, no natural reproduction takes place, and all 118
young trees have to be planted. Therefore, the parameter l0 corresponds to the diameter of the 119
trees when they are planted. Although the theoretical and empirical analysis is framed within 120
the context of a completely managed forest, it is quite possible to extend the framework to the 121
case in which trees reproduce naturally. In this case l0 can be interpreted as the diameter at 122
which trees are selected for upgrowth. The flux of logged trees is denoted by u(t,l), and the 123
flux of planted young trees at time t with diameter l0 by p(t). We assume that a diameter-124
distributed forest can be fully characterized by the number of trees and by the distribution of 125
the diameter of the trees. In other words, space and the local environmental conditions of the 126
trees are not taken into account.[1] 127
As the density function x(t,l) indicates the population density with respect to the structuring 128
variable l at time t, the number of trees in the forest at time t is given by 129
0
( ) ( , ) .ml
l
X t x t l dl= ∫ (1) 130
The term E(t) denotes environmental characteristics that affect the growth rate of the diameter 131
of individual trees. In the absence of pests, these environmental characteristics are given by 132

6
the local conditions where the tree is growing, and by the neighboring trees. Since our model 133
does not consider space, the term E(t) exclusively represents the competition between 134
individuals, i.e., the competition between individuals for space, light and nutrients. 135
Environmental pressure can be expressed, for example, by the total number of trees or the 136
basal area of all trees of the stand. Hence, a large (small) basal area of the stand signifies high 137
(low) intraspecific competition of the trees (Trasobares et al. 2004).[2] Thus, the change in 138
diameter of the tree over time is described by the function g(E(t),l), i.e., 139
( ( ), ),g E t ldldt
= (2) 140
where the basal area E(t) is determined based on the functional relationship between diameter 141
and basal area:
0
2( ) ( , )4
ml
lE t l x t l dlπ
= ∫ . 142
The instantaneous mortality rate, δ(E(t),l), describes the rate at which the probability of 143
survival of an l-sized tree in the presence of intraspecific competition E(t) decreases with 144
time. Based on the well-known McKendrick equation for age-structured populations 145
(McKendrick 1926), the dynamics of a diameter-distributed forest can be described by the 146
following partial integrodifferential equation as discussed by de Roos (1997) or by Metz and 147
Diekmann (1986): 148
( , ) [ ( ( ), ) ( , )] ( ( ), ) ( , ) ( , ),x t l g E t l x t l E t l x t l u t lt l
δ∂ ∂+ = − −
∂ ∂ t∈[0, T), l∈[l0, lm], (3) 149
subject to the boundary conditions x(0,l) = x0(l), l∈[l0, lm], and x(t,l0) = p(t), t∈[0, T). The first 150
two terms of Equation (3) represent the change in the tree density over time and diameter. The 151
term ( ( ) ( ))g x l∂ ⋅ ⋅ ∂ = ( ) ( ) g x l g l x∂ ∂ + ∂ ∂ takes into account the interdependence between 152
diameter and time, i.e., it represents the temporal change in diameter multiplied by the change 153
in tree density with respect to diameter plus the temporal change in diameter with respect to 154

7
diameter multiplied by the tree density.[3] Thus, the forest dynamics are described by the flux 155
of tree density with respect to diameter and time, which is equal to the terms of the right hand 156
side of Equation (3), given by tree mortality and logging. 157
To consider, in addition, the net benefits of carbon sequestration in the economic model, it is 158
necessary to analyze how the carbon content of the forest ecosystem changes over time. The 159
change in the carbon content is given by ,dz dt db dt ds dt= + and depends on the change in 160
carbon sequestered in the biomass captured by db dt , and the change in the carbon content in 161
the soil ds dt . With respect to the dynamics of soil carbon, we denote the above-ground 162
volume of the biomass of the forest by 0
0( ) ( , )ml
l
V t l x t l dlβγ= ∫ , where the strictly positive 163
parameters β and γ0 have to be chosen according to the species of the tree and the available 164
empirical data. Once function V is defined, we are in a position to specify the soil carbon 165
dynamics, which are given by 166
( )( ) ( ), ( ) ,ds t h V t s tdt
= t∈[0, T), s(0) = s0. (4) 167
Function h reflects to what extent the growth or harvest of trees and the current amount of soil 168
carbon affect the change in the soil carbon over time. 169
The amount of sequestered carbon in the biomass is given by 0
1( ) ( , )ml
l
b t l x t l dlβγ= ∫ . b(t) is 170
calculated based on the above-ground volume of the forest and the constant γ1, and the ratio γ1 171
/ γ0 reflects the carbon content per cubic meter of the biomass and is more or less constant 172
throughout the growth process of the trees. Before stating the complete bioeconomic model 173
we need to define the function ( ( , ), ( , ))B x t l u t l . This represents the net benefits of timber 174
production as a function of the standing and the harvested trees, with Bu > 0 and Bx < 0, 175

8
where a subscript of a variable with respect to a function indicates the partial derivative of the 176
function with respect to this variable. Finally, we denote the price of carbon by ρ1 and the cost 177
of planting young trees by ρ2. [4] 178
The decision problem (DP) of the manager or forest owner is to find the optimal trajectories 179
of the control variables u(t,l) and p(t), which in turn allows the optimal trajectories of the state 180
variables x(t,l) and s(t), and variables b(t), E(t) and V(t) to be obtained. We assume that the 181
owner or manager of the forest maximizes the joint net benefits of timber production and 182
carbon sequestration denoted by J. The literature currently distinguishes between two main 183
accounting methods for carbon. In the first one (the flow method) the valuation of carbon 184
sequestration is based on the flux of carbon in the ecosystem, and the price of carbon within a 185
period of time. That is, if the net storage of carbon ( ) ( )db t dt ds t dt+ is either positive or 186
above a predetermined reference value at time t [5], carbon sequestration provides a value, 187
and it is given by the net storage of carbon times the price of carbon. This approach 188
corresponds to the concept of “additionality”, which requires that only carbon sequestered 189
above the status quo should be considered. However, if the net storage of carbon is negative 190
due to extensive logging activities, or if it is below a predetermined reference value, the value 191
of the carbon balance is negative. The second accounting method (the stock method) is based 192
on the concept of a rental rate of carbon which corresponds to the value of storing one ton of 193
carbon for a period of time, and is equal to the price of carbon times the discount rate 194
(Sohngen and Mendelsohn 2003). If there is a predetermined reference value of the carbon 195
stock, then only the tons of carbon that are additionally sequestered are taken into account to 196
determine the value of carbon sequestration. In this study we opted for the flow method since 197
the sequestered carbon in the biomass and the wood product can be easily accounted for in the 198
modeling framework presented below. 199

9
The economic benefits of carbon sequestration in the presence of a strictly positive discount 200
rate reside in the fact that carbon is sequestered at time t and released at a later point in time. 201
The sequestration occurs initially in the tree and once the tree is cut the carbon remains 202
sequestered in the wood product. Therefore, we have two types of carbon parking: first in the 203
tree and second in the timber product. Unfortunately, the carbon flow db/dt only takes into 204
account the carbon sequestered in the tree, not in the wood product. According to the 205
definition of db/dt, the sequestered carbon is released immediately after the tree has been cut. 206
Thus, the term db/dt does not take into account the fact that the sequestered carbon continues 207
to be sequestered during the lifetime PT of the wood product (permanence time). When trees 208
are cut the amount of sequestered carbon is reduced. In the case of an instantaneous release of 209
carbon at time t, the loss of the discounted monetary value of the stored carbon would be 210
given by 1
0
1 1 ( , )ml
rtb
l
u e l u t l dlβρ γ−= ∫ . However, since the sequestered carbon is not released 211
immediately, but gradually over PT years and, according to the release function ( )PTω , the 212
loss of the discounted monetary value of the stored carbon of the logged trees in year t is 213
given by 2
0
( )1 1
0
( ) ( , )ml PT
r tb
l
u e PT d l u t l dlτ βω τ ρ γ− +⎛ ⎞= ⎜ ⎟
⎝ ⎠∫ ∫ .[6] The term in the inner brackets takes 214
into account the fact that the postponed release of the sequestered carbon leads to a lower loss 215
in value in comparison with the loss resulting from an immediate release of the sequestered 216
carbon after the tree is logged. Let 0
( ) [0,1]PT
re PT dτυ ω τ−= ∈∫ denote the share of the 217
sequestered carbon in the biomass that is released from the wood product if all carbon 218
releases are expressed in terms of year t. This allows the amount of the released carbon in 219
terms of year t to be calculated. Therefore, the longer the carbon is sequestered in the wood 220
product, the lower the discounted value of the released carbon. 221

10
A comparison of the payment for carbon releases shows that 222
( )1 2
0
1 1 11 ( , )ml
rt rtb b c
l
u u e l u t l dl e uβρ γ υ ρ− −− = − =∫ . The term cu is needed to account for the 223
carbon that is not released immediately after the trees are logged. Thus the term 224
( )0
1 1 ( , )ml
cl
u l u t l dlβγ υ= −∫ can be incorporated into the carbon flow equation, leading to 225
( ) ( )cdb t dt u ds t dt+ + . Collecting the previously introduced elements, the decision problem 226
takes the form of: 227
( , ), ( )max
u t l p t0
1 20
( ) ( )( ( , ), ( , )) ( ) ( )mlT
rtc
l
db t ds tJ e B x t l u t l dl u t p t dtdt dt
ρ ρ−⎧ ⎫⎪ ⎪⎡ ⎤= + + + −⎨ ⎬⎢ ⎥⎣ ⎦⎪ ⎪⎩ ⎭
∫ ∫ (DP) 228
under the restrictions 229
( , ) [ ( ( ), ) ( , )] ( ( ), ) ( , ) ( , ),x t l g E t l x t l E t l x t l u t lt l
δ∂ ∂+ = − −
∂ ∂ t∈[0, T), l∈[l0, lm], 230
0
2( ) ( , )4
ml
l
E t l x t l dlπ= ∫ , 231
( )( ) ( ), ( ) ,ds t h V t s tdt
= t∈[0, T), s(0) = s0 , 232
0
0( ) ( , )ml
l
V t l x t l dlβγ= ∫ , 233
0
1( ) ( , )ml
l
b t l x t l dlβγ= ∫ , 234
0
1( ) (1 ( )) ( , )ml
cl
u t l l u t l dlβγ υ= −∫ , 235
x(0,l) = x0(l), l∈[l0, lm], x(t,l0) = p(t), t∈[0, T), 236
0≤ u(t,l) ≤ umax(t,l), 0 ≤ p(t) ≤ pmax(t), 237

11
As the decision problem (DP) is based on one partial integrodifferential equation, Equation 238
(3), and one ordinary integrodifferential equation, Equation (4), the necessary conditions 239
presented previously for maximizing function J cannot be applied. Hritonenko et al. (2008) 240
establish the necessary extremum conditions in the form of a maximum principle for 241
optimization problems where a partial integrodifferential equation is present. 242
The necessary conditions for a maximum yield 243
∂J/∂u(t,l)≤0 at u*(t,l)=0, ∂J/∂u(t,l)≥0 at u*(t,l)=umax(t,l), 244
∂J/∂u(t,l)=0 at 0<u*(t,l)<umax(t,l) for almost all (a.a.) t∈[0, T), l∈[l0, lm]; (5) 245
∂J/∂p(t)≤0 at p*(t)=0, ∂J/∂p(t)≥0 at p*(t)=pmax(t), ∂J/∂p(t)=0 at 0<p*(t)<pmax(t), a.a. 246
t∈[0, T), (6) 247
( , ) [ ( ( ), ) ( , )] ( ( ), ) ( , ) ( , ),x t l g E t l x t l E t l x t l u t lt l
δ∂ ∂+ = − −
∂ ∂ (7) 248
x(0,l) = x0(l), l∈[l0, lm], x(t,l0) = p(t), t∈[0, T) , (8) 249
12
1
( , ) ( , )( ( ), ) [ ( ( ), )] ( , ) ( ( , ), ( , )) ( ) ( , )
4x
t l t lg E t l r E t l t l B x t l u t l l t F t l
t lr l β
λ λ πδ λ γ γ ρ
∂ ∂+ = + − + −
∂ ∂+ , (9) 250
λ(T,l) = 0, l∈[l0, lm], λ(t,lm) = 0, t∈[0, T), (10) 251
( )( )1( ) ( ), ( ) ( )
Tr t
t
t e h V s ds
τζ ρ τ τ ζ τ τ− − ∂= +
∂∫ , (11) 252
where λ(t,l) and ( )tζ are unknown dual variables, and 253
( )0( , ) ( ) ( ), ( )F t l l t h V t s tV
βγ ζ ∂=
∂, (12) 254
0
[ ( ( ), ) ( , )]( ) { ( ( ), ) ( , )} ( , )ml
EE
l
g E t l x t lt E t l x t l t l dll
γ δ λ∂= − −
∂∫ . (13) 255
The variables λ(t,l) and ζ(t) present the in situ value of trees and sequestered carbon in the 256
soil, respectively. 257

12
Equations (5) and (6) provide, for an interior solution [7], the following necessary conditions: 258
( , )( ( , ), ( , ))rtu t le B x t l u t l λ− = , (a.a.) t∈[0, T), l∈[l0, lm]; and (14) 259
2 0( ) ( , )rte t t lρ λ−− = , t∈[0,T). (15) 260
According to Equation (14) the discounted net benefits of logging have to be equal to the in 261
situ value of the trees for almost all t and l along the optimal path. Likewise, Equation (15) 262
states that the discounted cost of planting a young tree has to be identical to the in situ value 263
of a young tree along the optimal trajectory. Equation (7) describes the dynamics of the forest 264
as discussed above, and, due to the brevity of this exposition, the discussion is not repeated 265
here. The subsequent line presents the boundary conditions of the state variable x(t,l), i.e., the 266
initial diameter distribution of the trees and the inflow of newly planted trees. Equation (9) 267
indicates that the changes in the value of a standing tree (in situ value) over time and diameter 268
have to correspond to the changes in the value if the tree were cut down. The change in the in 269
situ value over diameter in Equation (9) is a composite expression given by 270
( ( ), ) ( , )g E t l t l lλ∂ ∂ and reflects the physical growth in diameter over time evaluated by the 271
change in the in situ value over diameter. The right-hand side of Equation (9) indicates that if 272
we had cut down trees, the interest earned from the money invested from the sale of these 273
trees would have been rλ and the value of the trees that had not died naturally should have 274
been δλ . Likewise, if we had cut down trees, the maintenance cost of the remaining trees, -Bx 275
> 0, would have decreased, which would contribute positively to the net benefits. In addition, 276
the expression 2 ( )4 l tπ γ reflects the monetary value of the improved conditions for the 277
growth of the remaining trees due to the decrease in intraspecific competition resulting from 278
trees being cut down. Moreover, if we had cut down trees, we would have foregone the 279
interest paid on the money received for sequestering carbon that corresponds to the cut trees, 280

13
leading to a decrease in net benefits, which is given by the term 1 1r l βγ ρ− . Finally, if we had 281
cut down trees the volume of the forest would have decreased, leading to a non-positive 282
change in the carbon content of the soil. The monetary value of this change is given by the 283
term ( , )F t l . The transversality conditions of the decision problem are stated in Equation (10). 284
Equation (11) provides the in situ value of soil carbon, which is given by the price of 285
sequestered carbon plus the discounted value of future changes in soil carbon as its level 286
increases or decreases. 287
As the decision problem (DP) is based on one partial integrodifferential equation, Equation 288
(3), and one ordinary integrodifferential equation, Equation (4), the first order conditions 289
include a system of partial integrodifferential equations. Given the complexity of the resulting 290
equations, an analytical solution of the first order conditions cannot be obtained, and it is 291
necessary to resort to numerical techniques to obtain a solution to the forest manager’s 292
decision problem. 293
Available techniques such as the gradient projection method (Veliov 2003) or the method of 294
finite elements may be appropriate to solve a distributed optimal control problem numerically 295
(Calvo and Goetz 2001). However, all of these methods require programming complex 296
algorithms that are not widely known. Therefore, we propose adapting a different method 297
called the Escalator Boxcar Train, EBT, used by de Roos (1988) to describe the evolution of 298
physiologically-structured populations. He has shown that this technique is an efficient 299
integration technique for structured population models. In contrast to the other available 300
methods, the EBT can be implemented with standard computer software such as GAMS 301
(General Algebraic Modeling System), which is used to solve mathematical programming 302
problems. 303

14
Applying the EBT allows the partial integrodifferential equations of the problem (DP) to be 304
transformed into a set of ordinary differential equations. Besides a brief presentation of the 305
EBT method, Goetz et al. (2007) show how this approach can be extended to account for 306
optimization problems by incorporating decision variables. To transform the decision problem 307
(DP), we first divide the range of the diameter into n equal parts, and define Xi(t) as the 308
number of trees in the cohort i, with i = 0, 1, 2, 3,…, n, that is, the trees whose diameter falls 309
within the limits li and li+1 are grouped in the cohort i. Likewise, we define Li(t) as the average 310
diameter, Ui(t) as the number of cut trees within cohort i, and P(t) as the number of planted 311
trees in cohort 0. Within these provisions the decision problem (DP) can now be formulated 312
as the maximization of the objective functions subject to a set of ordinary differential 313
equations instead of one partial integrodifferential and one ordinary differential equation 314
given by 315
2( ), ( )
00 0 0 0
0
20
max ( ( ), ( )) ( ( ))
subject to
( ) ( )( ( ), ) ( ) ( ), ( ( ), )
( ( ), ) ( ) ( ( ), ) ( ) ( )
( )
)
( )( )
( ( ,
Trt
U ct P t
i ii i i i
B X t U t P t e dt
dX t dL tE L X t U t g E Ldt dt
dX dE t l X t E t l L t P
db t ds tu
tdt dl
dL g E tdt
tdt dt
t t
ρ
δ
δ δ
ρ −⎧ ⎫−⎨ ⎬
⎩ ⎭
= − − =
= −
⎡ ⎤+ + +⎢ ⎥⎣ ⎦
− +
=
∫
0 0 0 0 0 0
00 0
) ( ) ( ( ), ) ( ) ( ( ), ) ( )
(0) , ( ( ), ) ( , ) ( ), ( ), ( ) 0 ( ) ( )i i i i
dl X t g E t l L t E t l L tdl
X X g E t l x t l P t U t P t U t X t
δ+ −
= = ≥ ≤ 316
( ) ( )20
0
( )( ) , ( ), ( ) , (0)4
n
i ii
ds tE t L X h V t s t s sdt
π=
= =∑ , 317
( ) ( )0 10 0
( ) , ( )n n
i i i ii i
V t L X b t L Xβ βγ γ= =
= =∑ ∑ , 318

15
where X and U denote the vectors 1, , nX X X= … and 1, , nU U U= … , respectively, and 319
0X the initial number of tress in each cohort. For numerical reasons the cohort with the 320
smallest trees has to be treated differently than the other cohorts. Hence, we obtain a separate 321
ordinary differential equation for 0X and 0L . Once the system of differential equations is 322
written as a system of difference equations, standard mathematical programming software 323
such as GAMS (General Algebraic Modeling System, (Brooke et al. 1992)) can be employed 324
to solve the problem. 325
However, before the decision problem can be solved, all functions have to be specified. While 326
the specification of the benefit and cost functions is straightforward, the biological functions 327
have to be estimated based on data generated by a biophysical forest growth simulator. For 328
this purpose we employed the bio-physical simulation model GOTILWA (Growth Of Trees Is 329
Limited by Water).[8] This not only simulates the development of the trees, but also the 330
amount of sequestered carbon in the biomass and in the soil. The next section describes the 331
simulation of forest growth by GOTILWA in detail, and presents the solution to the forest 332
manager’s decision problem, i.e., the optimal short- and long-term trajectories of the logging 333
and planting decisions and the evolution of the standing trees in the forest. 334
335
3. Empirical analysis 336
The purpose of the empirical analysis is to demonstrate the applicability of the presented 337
theoretical model for management and policy issues but not a detailed analysis of the land use 338
and land-use change options within existing international agreements with respect to climate 339
change. Initially we determine the selective-logging regime that maximizes the discounted 340
private net benefits from timber production and carbon sequestration of a stand of Pinus 341

16
sylvestris over a time horizon of 200 years. Thereafter, the analysis concentrates on the main 342
aspect of the empirical study by establishing to what extent a change in the price of carbon 343
affects the optimal logging regime. We specify the parameters and the functions in Section 344
3.1, and analyze the optimal management regime in Section 3.2. 345
3.1. Data and specification of functions 346
The net benefit function of the economic model, ( ( , ), ( , ))B x t l u t l , consists of the net revenue 347
from the sale of timber at time t, minus the costs of maintenance, which comprise clearing, 348
pruning, and grinding the residues. The net revenue is given by the sum of the revenue of the 349
timber sale minus logging costs defined as: ( )( ) ( )( )( ) ( )( ) ( ) ( )( )0
n
i i i ii
L t lc L t tv L t U t mc X tρ=
⎡ ⎤⎡ ⎤− −⎢ ⎥ ⎣ ⎦
⎣ ⎦∑ 350
where ( ) ( )0
n
ii
X t X t=
= ∑ . The terms in the first square brackets denote the sum of the revenue of 351
the timber sale minus the cutting costs of each cohort i, and the term in the second square 352
bracket, ( )( )mc X t , accounts for the maintenance costs. The function ( )iLρ denotes how the 353
timber price per cubic meter of wood varies with diameter; ( )itv L is the total volume of a tree 354
as a function of its diameter; ( )ilc L is logging cost. Timber price per cubic meter was taken 355
from a study by Palahí and Pukkala (2003), who analyzed the optimal management of a Pinus 356
sylvestris forest in a clear-cutting regime. They estimated a polynomial function given by 357
( ) 23.24 13.63 , 86.65L Min Lρ ⎡ ⎤= − +⎣ ⎦ , which is an increasing and strictly convex function, for a 358
diameter lower than 65cm. At L = 65 the price reaches its maximum value, and for 65L > it is 359
considered constant. The logging cost comprises logging, pruning, cleaning the underbrush, 360
and collecting and removing residues. Based on the work by Palahí and Pukkala (2003), the 361
logging cost is given by the function ( )( ) 6 exp 4.292 0.506lnlc L L= + −⎡ ⎤⎣ ⎦ , measured in euros 362
per cubic meter of logged timber. Data about maintenance costs were provided by the 363

17
consulting firm Tecnosylva, which elaborates forest management plans throughout Spain. 364
According to the data supplied by Tecnosylva, the maintenance cost function is approximated 365
by ( ( ))mc X t , and is given by 2( ( )) 10 0.0159 0.0000186mc X t X X= + + . The planting cost 366
is linear in the amount of planted trees and is given by C(P) = 0.73P. The thinning and 367
planting period, Δt, is set at 10 years, which is a common practice for a Pinus sylvestris forest 368
(Cañellas et al. 2000). 369
To proceed with the empirical study, various initial diameter distributions of a forest were 370
chosen. These distributions were specified as a transformed beta density function ( )lθ since it 371
is defined over a closed interval and allows a large variety of different shapes of the initial 372
tree diameter distributions to be defined (Mendenhall et al. 1990). The initial forest consists of 373
a population of trees with diameters within the interval 0 cm ≤ l ≤ 50 cm. The density function 374
of the diameter of trees, ( ); ;lθ γ ϕ , is defined over a closed interval, and thus the integral 375
( )1 1
1
1
; ,l
l dlθ γ ϕ+
∫ (16) 376
gives the proportion of trees lying within the range [ , 1)i il l + . We defined l0 = 0 and lm = 80 as 377
the minimum and maximum tree diameters, respectively. For the simulation of the forest 378
dynamics, we concentrate on the diameter interval [0, 50], because thereafter the tree growth 379
rate is very small. This interval was divided into 10 subintervals of identical length. 380
Therefore, the diameters of the trees of each cohort differ at most by 5 cm, and the size of the 381
trees of each cohort can be considered homogeneous. 382
To determine the forest and soil dynamics, the growth of a diameter-distributed stand of Pinus 383
sylvestris without thinning was simulated with the bio-physical simulation model GOTILWA. 384
This model simulates growth and mortality and explores how the life cycle of an individual 385
tree is influenced by the climate, the characteristics of the tree itself, and environmental 386

18
conditions. The model is defined by 11 input files specifying more than 90 parameters related 387
to the site, soil composition, tree species, photosynthesis, stomatal conductance, forest 388
composition, canopy hydrology, and climate. We simulated the growth of the forest based on 389
the previously specified initial diameter distributions. 390
The data generated from the series of simulations allow function ( ), ig E L , which describes the 391
change in diameter over time, to be estimated. This type of function was specified as a von 392
Bertalanffy growth curve (von Bertalanffy 1957), generalized by Millar and Myers (1990), 393
which allows the growth rate of the diameter to vary with environmental conditions specified 394
as the total basal area of all trees. The precise specification of the function is given by 395
( ) ( )( )0 1, i m ig E L l L Eβ β= − − . The exogenous variables of this function are diameter at breast 396
height (Li) and basal area (E) provided by GOTILWA. The parameters β0 and β1 are 397
proportionality constants and were estimated by OLS based on the pooled data. The 398
estimation yielded the growth function ( , ) (80 )(0.0070177 -0.000043079 )i ig E L L E= − . Other 399
functional forms of ( ), ig E L were evaluated as well, but they explained the observed variables 400
to a lesser degree. As GOTILWA only simulates the survival or death of an entire cohort, not 401
the survival or death of an individual tree, it was not possible to obtain an adequate estimation 402
of function ( ), iE Lδ , which describes the mortality of the forest. Nevertheless, information 403
provided by the company Tecnosylva suggests that the mortality rate in a managed forest can 404
be considered almost constant over time and independent of the diameter. Thus, according to 405
the data supplied by Tecnosylva, ( ), iE Lδ was made constant over time and equal to 0.01 for 406
each cohort. 407
The value of the tree volume parameters, ( )itv L , has also been estimated using the data 408
generated with GOTILWA. The tree volume is based on the allometric relation 409

19
( ) 1.7450870.00157387tv L L= . In addition, the data obtained from the GOTILWA simulations 410
allowed us to estimate the change in soil carbon over time as a function of the forest tree 411
volume and the amount of soil carbon. The estimation yielded the 412
function (212.12 ( ))( 0.0322 0.0003385 ( ))h s t V t= − − + . Row and Phelps (1990) suggest that 413
the release of sequestered carbon in the wood product can be described by an extended 414
logistic decay function, which Eggers (2002) specified by ( )
1.2( ) 1.21 5 c PTPT
eω = −
+. In 415
accordance with van Kooten and Bulte (1999) and Eggers (2002) we assumed that a long 416
permanence time corresponds to a half-life period of 50 years. Hence we employed the report 417
values for the function ( )c PT by Eggers (2002) for a long (50), medium (16) and short (1) 418
half-life period. Hence, we obtain for a long, medium and short half-life period that 1 υ− is 419
given by 0.58, 0.28, and 0.08, respectively. This expresses in terms of year t the share of 420
carbon that is retained in the wood product from time t to the end of the lifetime of the 421
product. 422
For a given initial diameter distribution of the trees, and given specifications of the economic 423
and biophysical functions of the model, a numerical solution of the decision problem (DP) 424
can be obtained. To proceed with the empirical study, three different initial diameter 425
distributions of a forest were chosen. They were obtained by varying the parameters γ and ϕ 426
of the beta density function, and are depicted in Figure 1. They stand for a young forest, a 427
mature forest, and a forest in which trees are distributed uniformly over the diameter range. 428
The initial basal area of all three distributions is identical and equal to 25m2/ha so that the 429
resulting optimal management regimes can be compared. 430
Figure 1 here 431
3.2. Analysis of the optimal selective-logging regime 432

20
In this part of the empirical study we determine the optimal management regime of a size-433
distributed forest and examine how the optimal management regime is affected by variations 434
in the carbon price. The optimizations were carried out with the CONOPT3 solver available 435
in the GAMS optimization package. For a given initial distribution, the numerical solution of 436
the problem determines the optimal trajectories of the decision variables: logging, iU , and 437
planting, P; and of the state variables: number of trees, iX , and diameter, iL . Consequently, it 438
also determines the value of the economic variables, such as the net revenue from timber 439
sales, logging costs, planting costs, maintenance costs and revenues from carbon storage. All 440
optimizations were carried out on a per-hectare basis. 441
The empirical analysis begins by calculating the optimal selective-logging regime for the 442
young forest distribution, given a long half-life period and a discount rate of 2%.[9] Figure 2 443
depicts the evolution of the number of standing trees and carbon in the forest over time for 444
different levels of the carbon price. It shows that the price of sequestered carbon, expressed in 445
terms of CO2, has a significant influence on the optimal selective-logging regime. When the 446
price increases from zero (base case) to 20 €/CO2 ton, it is optimal to increase the investment 447
in the forest. Therefore, the number of planted trees increases from 0 to 880 in the first period, 448
and the number of logged trees decreases from 115 to 107.[10] As a result, the number of 449
standing trees in the forest increases, especially in the initial periods of the time horizon, and 450
the amount of carbon sequestered increases in the long run (see Figure 2a). For a price of 0 451
€/CO2 ton, the amount of carbon in the forest ecosystem at the end of the planning horizon, 452
including soil carbon and the carbon stored in the above-ground and below ground biomass, is 453
108.3 tons/ha. However, it increases to 210.6 tons/ha when the carbon price is 20 €/CO2 ton, 454
that is, the carbon stored in the ecosystem in the long run is almost doubled (see Figure 2b). 455
Figure 2 here 456

21
Tables 1 and 2 present a more detailed analysis of the results for the base case, in which net 457
benefits only originate from timber (price of 0 €/CO2 ton) and for the case in which the price 458
is 10 €/CO2 ton. Table 1 contains the evolution of the physical variables over the first 100 459
years, while Table 2 presents the monetary values resulting from optimal forest management. 460
In particular, it is important to analyze the discounted sum of the net benefits obtained from 461
forest management, which we will refer to as simply the net benefits in the remaining part of 462
the paper. It is possible to see in Table 2 that when carbon sequestration does not form part of 463
the management objective, the net benefits over 200 years are about 6763 €/ha. When the 464
benefits and costs of carbon sequestration with a carbon price of 10 €/CO2 ton are 465
incorporated in the formulation of the decision problem, the net benefits increase to 7703 466
€/ha. Although the discounted sum of the net timber benefits decreases from 6762 €/ha to 467
6108 €/ha, the net benefits of carbon sequestration compensates the lower net benefits 468
obtained from the timber sale and the higher maintenance costs due to an increase in the tree 469
density. For an exact calculation of the net benefit of the carbon sequestration it is important 470
to consider only the carbon that is sequestered in addition to the carbon sequestered by the 471
forest that maximizes the net benefits of the timber product. In this way the empirical study 472
only takes into account the “additional” sequestered carbon. 473
Table 1 and 2 here 474
To analyze to what extent this result can be generalized, we conducted a sensitivity analysis 475
for the three initial forest distributions considered, and for different carbon prices, ranging 476
from 0 to 25 €/CO2 ton. Figure 3 depicts the NPV of the different optimization scenarios. The 477
figure corroborates the previously obtained result by showing that the NPV of the benefits of 478
forest management increases with an increase in the price from 0 to 25 €/CO2 ton. However, 479
the increase in the net benefits depends on the shape of the initial forest distribution. For low 480
carbon prices (0 - 10 €/CO2 ton), both a mature and a uniformly distributed forest produces 481

22
higher net benefits compared to a young forest. In contrast, for high carbon prices (over 15 482
€/CO2 ton), the result is reversed. This is due to the fact that for low carbon prices, the timber 483
price is more decisive, so that mature or uniform forests generate higher net benefits than 484
young forests. However, if the carbon price is relatively high its price becomes decisive, and 485
the carbon net benefits of a young and highly productive forest outweigh the reduction in net 486
benefits of timber production. 487
Figure 3 here 488
One might suppose that the results obtained depend strongly on the permanence time, PT, of 489
the sequestered carbon in the wood products since this determines the amount that the forest 490
owner has to pay for the release of carbon once the trees have been logged. To analyze this 491
point in more detail, we conducted a sensitivity analysis with respect to PT for the young 492
distribution, and we calculated the optimal logging regime, given a short lifespan (half-life 493
period equal to 1 year), a medium lifespan (half-life period equal to 16 years) and a long 494
lifespan (half-life period of 50 years). The results show that the PT does not affect the net 495
benefits obtained from forest management. In order to save space we chose not to present 496
these results in graphical form. 497
In addition, we conducted a sensitivity analysis of the discount rate. Figure 4 depicts the 498
change in the net benefits over a time horizon of 200 years for different carbon prices, given 499
discount rates of 2% and 4%. Figure 4 shows that an increase in the carbon price may lead to 500
a substantial increase in the net benefits if the chosen discount rate is low. For instance, for a 501
discount rate of 2%, a price of 5 €/ton of CO2 leads to an increase in the net benefits of 5% 502
compared to the base case, and a price of 25 €/ton of CO2 leads to an increase in the net 503
benefits of 20%. However, the higher the discount rate, the smaller the gains from carbon 504
sequestration. This tendency is amplified even more if the carbon price is relatively small. For 505

23
instance, for a discount rate of 4%, the carbon price needs to be 15 €/ton to obtain a 6% 506
increase in the net benefits compared to the base case. Hence, if the discount rate of the 507
private owner of a forest is above the discount rate used by a social planner, the amount of 508
sequestered carbon is below the socially optimal amount. 509
Figure 4 here 510
Our calculations show that an increase in the carbon price induces management regimes that 511
augment the sequestered carbon in the biomass and soil. However, the increase in the 512
sequestered carbon goes along with a decrease in the net benefits from timber production. The 513
relation between the decrease in the net benefits of timber production and the increase in 514
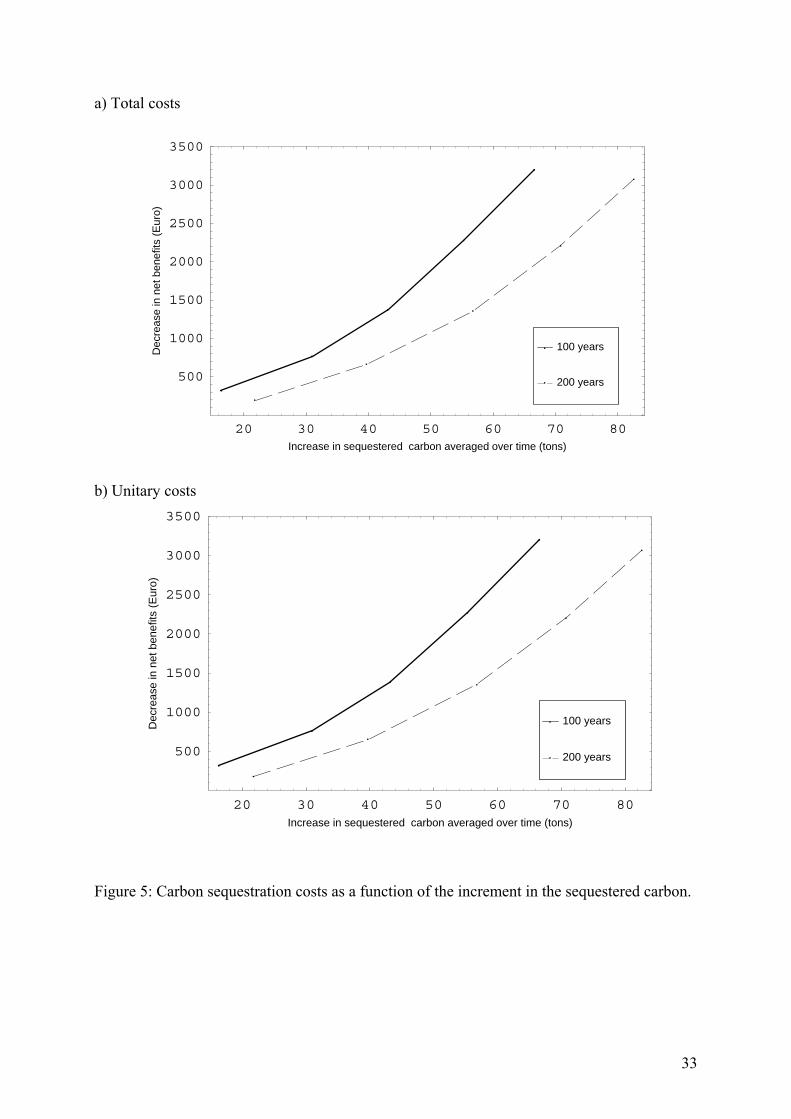
sequestered carbon (price of carbon) over time horizons of 100 and 200 years is presented in 515
Figure 5a. This figure also shows that the decrease in the net benefits of timber is more 516
pronounced over 100 years than over 200 years. This result can be explained by the fact that a 517
large share of the timber net benefits can only be obtained after 100 years because only then 518
the trees have achieved the optimal logging diameter. The decrease in net benefits of timber 519
production results from an increase in the carbon price, which in turn leads to a change in the 520
management regime that favors biomass production at the cost of timber production. 521
Moreover, Figure 5a allows the costs of sequestered carbon to be calculated per ton, and those 522
costs presented in Figure 5b, which shows that the costs of sequestered carbon per ton are 523
between 8.5 and 48.1 € (2.3 – 13.1 €/ton of CO2). Based on an extensive literature review and 524
a meta-analysis, van Kooten and Sohngen (2007) state that the carbon sequestration costs in 525
Europe range between 120 and 710 €/tC.[11] In this respect the sequestration costs presented 526
in this study are clearly at the lower end of previous studies. It may seem that the results of 527
our study cannot be compared directly with the results of the previous literature since 528
previous studies predominantly determined the carbon sequestration costs for afforested land 529
and, consequently, the opportunity costs of the land have to form part of the carbon 530

24
sequestration costs. However, since we are considering a change in the management regime of 531
a standing forest, the opportunity costs of the land affect the aggregate discounted net benefit 532
of timber but not a change in these benefits. In other words, the opportunity costs of the land 533
are independent of the forest management regime and therefore do not need to be taken into 534
account when the costs for additionally sequestered carbon are calculated. 535
Figure 5 here 536
4. Conclusions 537
In this paper we present a theoretical model that allows us to determine the optimal 538
management regime of a diameter-distributed forest. The theoretical model can be formulated 539
as a distributed optimal control problem in which the control variables and the state variable 540
depend on two factors: time and the diameter of the tree. As the resulting necessary conditions 541
of this problem include a system of partial integrodifferential equations, the problem cannot 542
be solved analytically. For this reason, a numerical method is employed. The Escalator 543
Boxcar Train technique can be used to solve a distributed optimal control problem by 544
transforming the independent factor, diameter, into a state variable of the system that evolves 545
over time. In this way, the partial integrodifferential equation is decoupled into a system of 546
ordinary differential equations, converting the distributed optimal control problem into a 547
classic optimization problem that can be solved by utilizing standard mathematical 548
programming techniques. 549
An empirical analysis is conducted to determine the optimal selective-logging regime, that is, 550
the selective-logging scheme that maximizes the present value of net benefits from timber 551
production and carbon sequestration of a privately owned Pinus sylvestris forest, and to 552
evaluate how the optimal management of the forest is affected by variation in the market 553
prices of carbon sequestration. The study is characterized by the fact that the complex growth 554

25
process of the trees and the storage of carbon in the soil and in the biomass are taken into 555
account. 556
The results of our calculations show that an increase in the carbon price leads to an increase in 557
the number of trees in the forest and to an increase in the carbon sequestered in the forest 558
ecosystem. Moreover, it underlines that the accurate assessment of the carbon in the biomass 559
and the soil, and the considered time horizon are crucial for the correct determination of the 560
optimal forest and carbon management regime. In contrast, the permanence time of the 561
sequestered carbon in wood products does not affect the optimal management regime. If the 562
carbon price is relatively low, mature and uniform forests lead to a higher increase in net 563
benefits than young forests. This result is reversed for the case of a young forest. The overall 564
costs of carbon sequestration as a result of a change in the forest management regime are 565
relatively small in comparison with the results presented in the existing literature. Part of this 566
difference can be explained by the fact that the additionally sequestered carbon does not 567
require a change in the land use. Hence, in contrast with afforestation, a change in the 568
management regime does not require the opportunity cost of the land to be imputed. 569
Consequently, carbon sequestration costs are expected to be lower when the forest 570
management regime is changed than when land use is changed. This study presents the 571
magnitude of the decrease in the costs. 572
573
Endnotes574 [1] More complex models which take into account space, for instance, in the form of local
conditions can be solved by heuristic approaches, but not by efficient optimization
techniques (Weintraub et al. 2007).

26
[2] We do not consider space and assume that trees with different sizes are distributed evenly
in the stand. However, if this is not the case, distance dependent competition indices would
be more appropriate (García-Abril et al. 2007).
[3] If we had chosen age as the structuring variable the function g would be constant and
equal to 1 since (age) 1d dt g= = and therefore the aging of the tree by one year would
correspond to one year of calendar time.
[4] In the case of natural reproduction (Karlsson and Örlander 2000) it can be assumed that
the number of seeds that turn into seedlings is sufficiently large, which allows the forest
manager to choose the number of trees with a diameter of l0 by removing any additional
trees. In this case, p(t) denotes the number of upgrowing trees and ρ2 the “nursing” costs.
[5] For example, the reference value may be given by the net storage of carbon resulting from
maximizing the net benefits of timber production alone for an already existing or a newly
planted stand. The management regime that only maximizes the net benefits of timber
production follows from setting the price of carbon equal to zero.
[6] An example of the specification of the function ( )PTω is presented in Section 3.1: Data
and Specification of Functions.
[7] The numerical solution of the problem showed that interior and boundary solutions occur
along the optimal trajectory. In the latter case the Lagrange multipliers take on positive
values.
[8] This program has been developed by C. Gracia and S. Sabaté, University of Barcelona,
Department of Ecology and CREAF (the Centre for Ecological Research and Forestry
Applications), Autonomous University of Barcelona, respectively.
[9] Palahí and Pukkala (2003) and Díaz-Balteiro and Romero (2003) choose a discount rate
of 2% while Creedy and Wurzbacher (2001) choose a discount rate of 4%. Thus, we start
the empirical analysis with a discount rate of 2% and subsequently conduct a sensitivity

27
analysis to evaluate the effect of different discount rates on the optimal management
regime.
[10] These results demonstrate how important it is to include planting as a separate decision
variable.
[11] Van Kooten and Sohngen present their results in US$ which we converted into € based
on an exchange rate of 1.45$/€.

28
Literature Cited
ADAMS, D., R. ALIG, B. MCCARL, J. M. CALLAWAY, and S. WINNET. 1999. Minimum Cost Strategies for Sequestering Carbon in Forests. Land Economics 75(3):360-374.
ADAMS, D. M., and A. R. EK. 1974. Optimizing the management of uneven-aged forest stands. Canadian Journal of Forest Research 4:274-286.
BROOKE, A., D. KENDRICK, and A. MEERAUS. 1992. GAMS: A User's Guide. The Scientific Press, San Francisco.
CALVO, E., and R. GOETZ. 2001. Using distributed optimal control in economics: A numerical approach based on the finite element method. Optimal Control Applications and Methods 22(5/6):231-249.
CAÑELLAS, I., F. MARTÍNEZ-GARCÍA, and G. MONTERO. 2000. Silviculture and dynamics of Pinus sylvestris L. stands in Spain. Investigación Agraria: Sistemas y Recursos Forestales Fuera de serie nº 1:233-253.
CREEDY, J., and A. D. WURZBACHER. 2001. The economic value of a forested catchment with timber, water and carbon sequestration benefits. Ecological Economics 38:71–83.
DE ROOS, A. 1988. Numerical methods for structured population models: The Escalator Boxcar Train. Numerical Methods for Partial Differential Equations 4:173–195.
DE ROOS, A. 1997. A gentle introduction to physiologically structured population models. Chapter 5, pp. 119-204 in Structured Population Models in Marine, Terrestrial and Freshwater Systems, S. Tuljapurkar and H. Caswell (eds.). Chapman and Hall, New York.
DÍAZ BALTEIRO, L., and C. ROMERO. 2003. Forest management optimisation models when carbon captured is considered: a goal programming approach. Forest Ecology and Management 174:447-457.
EGGERS, T. 2002. The Impacts of Manufacturing and Utilisation of Wood Products on the European Carbon Budget. R. Päivinen. Joensuu, European Forest Institute.
GARCÍA-ABRIL, A., S. MARTÍN-FERNÁNDEZ, M. A. GRANDE, and J. A. MANZANERA. 2007. Stand structure, competition and growth of Scots pine (Pinus sylvestris L.) in a Mediterranean mountainous environment. Annals of Forest Science 64:825-30.
GETZ, W. M., and R. G. HAIGHT. 1989. Population Harvesting: Demographic Models of Fish, Forest and Animal Resources. Princeton University Press, Princeton.
GOETZ, R. U., A. XABADIA, and E. CALVO. 2007. A numerical approach to determine the optimal forest management regime in the presence of intra-specific competition. Manuscript, University of Girona, Spain.
HAIGHT, R. E. 1987. Evaluating the efficiency of even-aged and uneven-aged stand management. Forest Science 33:116-34.
HAIGHT, R. E., J. D. BRODIE, and R. M. ADAMS. 1985. Optimizing the sequence of diameter distributions and selection harvests for uneven-aged stand management. Forest Science 31(2):451-462.
HRITONENKO, N., Y. YATSENKO, R. U. GOETZ, and A. XABADIA. 2008. Maximum principle for a size-structured model of forest and carbon sequestration management. Applied Mathematics Letters 21:1090-94.
KARLSSON, C., and G. ÖRLANDER. 2000. Soil scarification shortly before a rich seed-fall improves seedling establishment in seed tree stands of Pinus sylvestris. Scandinavian Journal of Forest Research 15:256-266.
KEYFITZ, N. 1968. Introduction to the Mathematics of Population. Addison-Wesley, New York.

29
MATTHEWS, R. 1993. Towards a methodology for the evaluation of the carbon budget of forests. Mensuration Branch, Forestry Commission, Alice Holt Lodge, Farnham.
MCKENDRICK, A. 1926. Applications of mathematics to medical problems. Proceeding Edinburgh Mathematical Society 44:98-130.
MENDENHALL, W., D. WACKERLY, and R. SCHEAFFER. 1990. Mathematical Statistics with Applications. PWSKent, Boston.
METZ, J., and O. DIEKMANN. 1986. The Dynamics of Physiologically Structured Populations. Springer-Verlag, Heidelberg.
MILLAR, R., and R. MYERS. 1990. Modelling environmentally induced change in growth for Atlantic Canada cod stocks. ICES- International Council for the Exploration of the Sea C.M./G24.
PALAHÍ, M., and T. PUKKALA. 2003. Optimising the management of scots pine (Pinus sylvestris L.) stands in Spain based on individual-tree models. Annals of Forest Science 60:105-114.
POHJOLA, J., and L. VALSTA. 2007. Carbon credits and management of Scots pine and Norway spruce stands in Finland. Forest Policy and Economics 9(7):789-798.
RASSE, D., B. LONGDOZ, and R. CEULEMANS. 2001. TRAP: a modelling approach to below-ground carbon allocation in temperate forests. Plant and Soil 229:281-293.
ROW, C., and R. PHELPS. 1990. Tracing the flow of carbon through the U.S. forest product sector. 19th World Congress of the International Union of Forest Research Organizations. Montreal, Canada.
SOHNGEN, B., and R. MENDELSOHN. 2003. An Optimal Control Model of Forest Carbon Sequestration. American Journal of Agricultural Economics 85 (2):448-457.
STAINBACK, A. G., and J. R. ALAVALAPATI. 2002. Economic analysis of slash pine forest carbon sequestration in the southern U. S. Journal of Forest Economics 8(2):105-117.
THORNLEY, J., and M. CANNELL. 2000. Managing forests for wood yield and carbon storage: a theoretical study. Tree Physiology 20:477-484.
TRASOBARES, A., M. TOMÉ, and T. PUKKALA. 2004. Growth and yield model for Pinus halepensis Mill. in Catalonia, north-east Spain. Annals of Forest Science 61:747-58.
VAN KOOTEN, G. C., C. S. BINKLEY, and G. DELCOURT. 1995. Effect of Carbon Taxes and Subsidies on Optimal Forest Rotation Age and Supply of Carbon Services. American Journal of Agricultural Economics. 77(2):365-74.
VAN KOOTEN, G. C., and B. SOHNGEN. 2007. Economics of forest ecosystem carbon sinks: A review. International Review of Environmental and Resource Economics 1(3):237-269.
VELIOV, V. M. 2003. Newton’s method for problems of optimal control of heterogeneous systems. Optimization Methods and Software 18:387-411.
VON BERTALANFFY, L. 1957. Quantitative laws in metabolism and growth. Quarterly Review of Biology 32:217–231.
WEINTRAUB, A., C. ROMERO, T. BJØRNDAL, and R. EPSTEIN. 2007. Handbook of Operations Research in Natural Resources. Springer, New York, pp.315–544.
XABADIA, A., and R. U. GOETZ. 2007. The optimal selective logging regime and the Faustmann formula. GSE Research Network, Working Paper 353, Barcelona, Spain.

30
0 10 20 30 40diameter at breast height (cm)
50
100
150
200
250
300
num
ber
oftr
ees mature
forest
youngforest
uniform
distribution
Figure 1. Initial distributions used in the study

31
2a)
0 50 100 150 200time
500
1000
1500
2000N
umbe
rof
stan
ding
tree
s
CO2 price of 20 Euro
CO2 price of 10 Euro
CO2 price of 0 Euro
2b)
0 50 100 150 200time
50
100
150
200
250
300
Car
bon
inth
efo
rest
ecos
yste
m(t
ons/
ha)
CO2 price of 20 Euro
CO2 price of 10 Euro
CO2 price of 0 Euro
Figure 2: Variation in the evolution of the main variables over time for different levels of the
carbon price

32
0 5 10 15 20 25Price of CO2 (euros/Tm)
4000
6000
8000
10000
12000
14000
Netbenefits(euros)
matureforest
uniform
distribution
youngforest
Figure 3: Net benefits over 200 years as a function of the CO2 price for different initial
distributions of the forest
0 5 10 15 20 25Price of CO2 (euros/Tm)
5
10
15
20
25
Incrementofthenetbenefits(%)
r=0.04
r=0.02
Figure 4: Change in the net benefits over 200 years as a function of the CO2 price for different
levels of the discount rate
33
a) Total costs
20 30 40 50 60 70 80Increase in sequestered carbon averaged over time (tons)
500
1000
1500
2000
2500
3000
3500D
ecre
ase
inne
tben
efits
(Eur
o)
200 years
100 years
b) Unitary costs
20 30 40 50 60 70 80Increase in sequestered carbon averaged over time (tons)
500
1000
1500
2000
2500
3000
3500
Dec
reas
ein
netb
enef
its(E
uro)
200 years
100 years
Figure 5: Carbon sequestration costs as a function of the increment in the sequestered carbon.

34
Table 1: Optimal selective-logging regime of a young stand (physical variables)
Considering only timber (price =0 €/CO2 ton) Logged
Year Number of trees
Planted trees
Logged trees
volume (m3/ha)
0 820 0 115 98.78 10 797 0 15 9.14 20 763 26 52 38.56 30 773 102 84 58.18 40 780 119 104 67.64 50 796 151 127 79.77 60 815 170 143 87.33 70 806 93 94 58.56 80 768 127 157 112.63 90 801 55 14 8.25 100 789 101 105 64.57
Considering timber and carbon sequestration (price =10 €/CO2 ton) 0 1250 402 87 79.45 10 1341 149 46 33.34 20 1267 0 61 43.15 30 1276 115 93 60.29 40 1303 123 84 53.73 50 1319 156 127 76.86 60 1322 175 158 90.72 70 1286 222 245 134.88 80 1250 243 266 172.15 90 1267 139 110 81.07 100 1282 151 124 91.66

35
Table 2: Optimal selective-logging regime of a young stand (monetary values)
Considering only timber (price =0 €/CO2 ton)
Year
Net revenue from timber
sale (€/ha)(b)
Maintenance cost
(€/ha)
Planting cost
(€/ha)
Carbon in the forest (tons/ha)
Carbon revenue (€/ha)
Net benefit (€/ha)
Discounted net benefit
(€/ha) 0 3019.96 355.74 0 134.35 - 2664.21 2664.21 10 218.92 345.26 0 146.93 - -126.33 -103.64 20 1042.79 329.71 18.7 153.19 - 694.37 467.29 30 1496.9 334.1 74.4 154.15 - 1088.4 600.88 40 1670.21 337.53 86.94 152.13 - 1245.75 564.19 50 1909.38 344.85 110.54 146.26 - 1453.99 540.2 60 2050.62 353.6 124.1 137.79 - 1572.92 479.4 70 1401.55 349.46 67.9 137.13 - 984.19 246.07 80 2979.38 332 92.52 121.42 - 2554.86 524.03 90 191.78 347.14 40.07 134.69 - -195.42 -32.88 100 1525.91 341.63 73.73 133.775 - 1110.55 153.29
Discounted sum over 200 years: 8900.34 1873.29 264.29 - 6762.75 Considering timber and carbon sequestration (price =10 €/CO2 ton)
0 2525.16 590.18 293.49 140.81 0 1641.5 1641.5 10 901.06 648.64 108.87 152.54 262.64 406.19 333.22 20 1131.89 600.47 0 166.5 541.06 1072.47 721.74 30 1493.96 606.6 83.8 177.3 699.73 1503.3 829.93 40 1307.8 623.64 90.09 190.29 479.12 1073.2 486.04 50 1787.5 633.74 113.92 196.67 434.68 1474.51 547.82 60 2027.26 636.05 127.68 198.47 393.82 1657.35 505.13 70 2917.77 612.71 161.87 186.79 -15.09 2128.09 532.08 80 4227.54 590.25 177.58 162.89 2.95 3462.66 710.22 90 2194.64 600.88 101.6 164.22 -66.75 1425.41 239.84 100 2484.47 610.36 110.45 164.08 166.39 1930.05 266.41
Discounted sum over 200 years: 10206.29 3373.43 724.22 1484.82 7703.56
Top Related
Copyright © 2022 FDOKUMEN

