







Bahasa
Halaman
Hukum


Research article
Female and male contribution to egg size in salmonids
SUSANNA PAKKASMAA1*, NINA PEUHKURI, ANSSI LAURILA1,
HEIKKI HIRVONEN and ESA RANTAIntegrative Ecology Unit, Department of Ecology and Systematics, Division of Population Biology,
P.O. Box 17 (Arkadiankatu 7), FIN-00014 University of Helsinki, Finland1Present address: Department of Population Biology, Evolutionary Biology Centre, Uppsala
University, Norbyvagen 18D, SE-75236 Uppsala, Sweden
(*author for correspondence, tel.: +46-18-4716496; fax: +46-18-4716484;
e-mail: [email protected])
Received 23 November 2000; accepted 8 October 2001
Co-ordinating editor: H. Kokko
Abstract. Egg size contributes to other life history traits of an individual. It is traditionally con-
sidered as a maternally determined characteristic to which the male does not have any direct
contribution. However, a recent finding in insects suggests that males can affect egg size also
directly. In fish, the male effect could take place only during egg swelling, as the final egg size is
reached after that. We studied egg size in four freshwater salmonid species (the land-locked Atlantic
salmon, the brown trout, the Arctic charr and the lake trout) right after fertilisation (initial egg size)
and after the swelling phase (final egg size). The results showed that the final egg size is affected not
only by the initial egg size but also by both the female and the male through the process of egg
swelling. This study suggests that paternal contribution may form a previously largely ignored
source of variation in early life history traits in salmonid fish.
Key words: early life history, egg size, egg swelling, female effects, male effects, Salmo, Salvelinus
Introduction
Egg size is a central early life history character influencing the fitness of both
the mother and her offspring (Bernardo, 1996a). Egg size may correlate not
only with the status (e.g., age, size and condition) of the female producing the
eggs, but also with the size and other attributes hatching offspring (Chambers,
1997). In fish, for example, large eggs generally result in large larvae and fry
(Chambers, 1997) having, among other things, wider mouth gape, longer visual
reactive distance and greater swimming speed than is typical for their smaller
conspecifics (Blaxter, 1986; Miller et al., 1988). These traits, in addition to large
size as such, are likely to have survival value in nature, e.g., in terms of in-
creased efficiency to forage and/or to avoid predators.
Traditionally, egg size has been considered as a maternal character deter-
mined by the female’s genotype and the environment she has been exposed to
Evolutionary Ecology 15: 143–153, 2001.� 2002 Kluwer Academic Publishers. Printed in the Netherlands.

(Bernardo, 1996a, b; Chambers and Leggett, 1996; Mousseau and Fox, 1998).
The male’s influence on egg size has been acknowledged in some animals,
where the male contributes to female nutrition through food or nuptial gifts
provided before egg laying and affects thus egg size indirectly (e.g., Gwynne,
1984; Sakaluk, 1986; Daan et al., 1990). Recent observations in the cricket
Gryllus firmus, however, showed that the male can affect egg size through a
direct contribution to the water uptake and/or metabolism of the embryo
before hatching (Weigensberg et al., 1998). In fish, maternal effects on egg size
have been widely recognised (e.g., reviewed by Heath and Blouw, 1998), but
the potential for the male affecting egg size has practically been neglected. In
the present study, we examine the effects of both the female and the male on
egg size in salmonid fishes.
In salmonids, like in most teleost fishes, the final egg size is attained after
swelling following egg activation by water contact or fertilisation. Swelling is
caused by an osmotic influx of water from the surrounding medium, resulting
in the formation of a fluid-filled perivitelline space underneath the expanding,
outer egg membrane (Alderdice, 1988). The extent of swelling differs markedly
among fish species so that the perivitelline space can make up most of the egg
volume (Lønning and Davenport, 1980) or be hardly noticeable (Bolin, 1930).
It is well established that the amount of resources provided by the female
influences egg size (Bernardo, 1996b; Chambers and Legget, 1996). There is
some indication from Atlantic cod (Gadus morhua) that the female may con-
tribute to the final egg size also by influencing the extent of swelling (Kjesbu
et al., 1996). The male contribution to egg size can only be manifested through
the swelling process as fertilisation in salmonids is external and the male does
not provide any resources to the developing embryo, neither directly nor via
the mother. We studied the parental contributions on egg size in four salmo-
nids by including the pre-fertilisation size (female effect) and the post-fertili-
sation size, i.e., the amount of swelling (female and male effects).
Materials and methods
We studied egg size in four freshwater salmonids: land-locked Atlantic salmon
(Salmo salar m. sebago), brown trout (S. trutta m. lacustris), Arctic charr
(Salvelinus alpinus) and lake trout (Sa. namaycush). Salmon, brown trout and
Arctic charr are native to Finland, but lake trout is an imported species
originating from North America. Salmon and brown trout were captured as
breeding adults from the Vuoksi water system, eastern Finland, where both
species occur naturally. In addition to wild fish, we used parental salmon from
a hatchery stock originating from the same area. For Arctic charr and lake
trout, only hatchery-reared parents were available. For each species, we used a
144

complete factorial mating design and created fertilisation matrices by crossing
five to eight females each with five to seven males (Table 1). The matrices
consisted of 25–56 families, each with approximately 300 eggs. The parental
fish from the hatchery stock were randomly chosen, with the condition that the
males and females of each matrix were always from different year classes in
order to avoid matings with close relatives. Individuals from the wild were
randomly assigned to each matrix.
All crossings were made in the hatchery in October–November 1996. Eggs
and milt of the wild fish were stripped at the place of catch and stored on ice for
a variable period but always less than 2 weeks. The eggs were stored together
with ovarian fluids, and milt was stored in plastic bags filled with pure oxygen.
The motility of spermazoa was checked under microscope before being used
for fertilisations, and milt showing at least 70% of progressively swimming
spermatozoa was used. The fish from the hatchery strains were stripped in the
hatchery, and fertilisations were made on the day of stripping or the following
day. The eggs of each female were divided in equally large batches (about 300
eggs) in 0.2 l plastic vials and fertilised. They were photographed immediately
after fertilisation, allowed to swell, and photographed again 4 h later, by which
time the swelling is completed (Kamler, 1992).
Maximum diameter of a sample of eggs (usually >100 eggs per family) was
measured from the photographs with an image analysis programme. Family-
specific average egg sizes before and after swelling were calculated from these
data, and they were used to analyse the parental effects on egg swelling and
thus on the final egg size. Linear regression was used for each matrix separately
in order to analyse how much of the variation in the final egg size can be
explained by variation in the initial egg size. We used matrix-specific residuals
of the linear regression of the final egg size against initial egg size as the
response variable in the analyses. Thus, we controlled for the potential effect of
initial egg size and the associated female effects when analysing egg swelling.
Each matrix was analysed with a two-way analysis of variance (ANOVA)
without replication. The female � male interaction was calculated according to
Table 1. The number of matrices of wild and hatchery fish in each study species and the lengths
and weights of the parental fish. The origin of the fish (hatchery or nature) is indicated.
Species Matrices Origin Females Males
Length (cm) Weight (g) Length (cm) Weight (g)
Salmon 3 Nature 67–78 3050–5770 62–96 1900–7500
7 Hatchery 48–69 1220–4830 12–51 20–1410
Brown trout 3 Nature 48–77 1180–5410 36–86 590–7060
Arctic charr 7 Hatchery 47–65 880–2800 53–67 1590–3230
Lake trout 2 Hatchery 56–66 2630–4370 59–78 1870–5980
145
Tukey (1949). The probabilities obtained from the ANOVAs were combined
using Fisher’s meta-analytic method described in Sokal and Rohlf (1995). Here
the method created an overall test for the significance of female and male
effects on egg swelling. Probabilities were calculated separately for wild and
hatchery-reared salmon.
Results and discussion
As expected, most of the variation in final egg size was explained by the initial
egg size (Table 2). Nonetheless, we found statistically significant male effects,
female effects, and female � male interactions on the final egg size corrected by
the initial egg size (Table 2, Fig. 1A). The female had an effect on the final egg
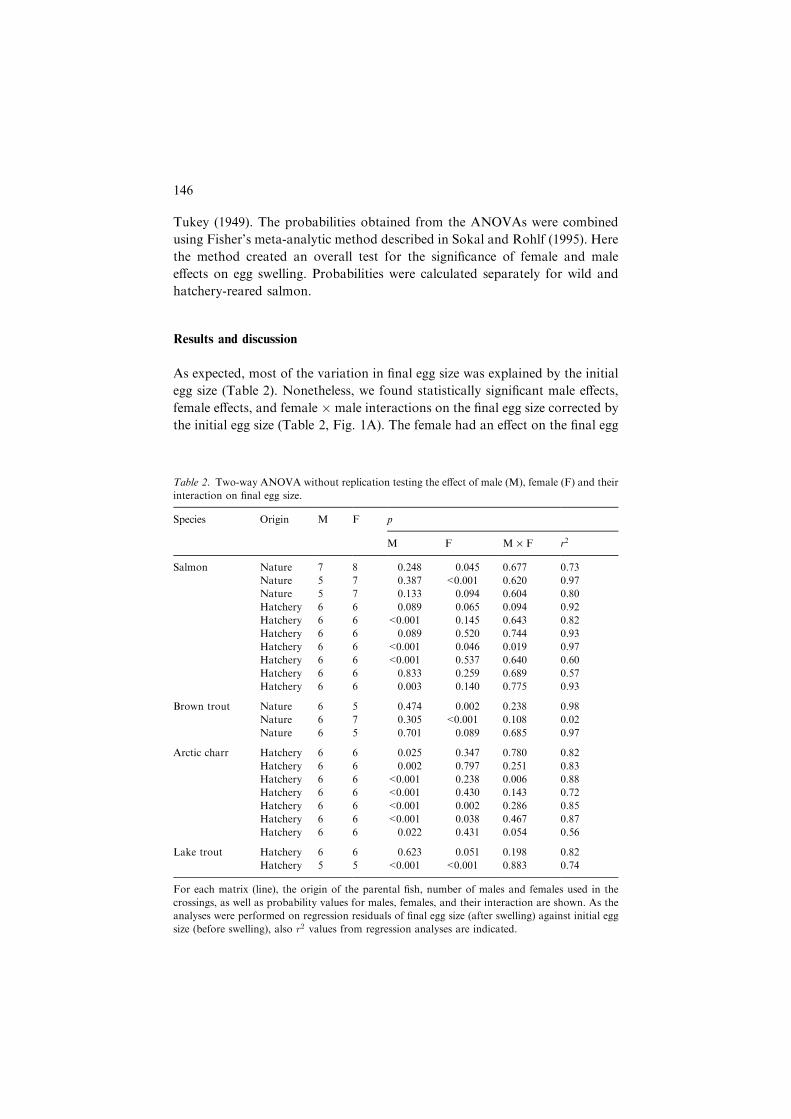
Table 2. Two-way ANOVA without replication testing the effect of male (M), female (F) and their
interaction on final egg size.
Species Origin M F p
M F M · F r2
Salmon Nature 7 8 0.248 0.045 0.677 0.73
Nature 5 7 0.387 <0.001 0.620 0.97
Nature 5 7 0.133 0.094 0.604 0.80
Hatchery 6 6 0.089 0.065 0.094 0.92
Hatchery 6 6 <0.001 0.145 0.643 0.82
Hatchery 6 6 0.089 0.520 0.744 0.93
Hatchery 6 6 <0.001 0.046 0.019 0.97
Hatchery 6 6 <0.001 0.537 0.640 0.60
Hatchery 6 6 0.833 0.259 0.689 0.57
Hatchery 6 6 0.003 0.140 0.775 0.93
Brown trout Nature 6 5 0.474 0.002 0.238 0.98
Nature 6 7 0.305 <0.001 0.108 0.02
Nature 6 5 0.701 0.089 0.685 0.97
Arctic charr Hatchery 6 6 0.025 0.347 0.780 0.82
Hatchery 6 6 0.002 0.797 0.251 0.83
Hatchery 6 6 <0.001 0.238 0.006 0.88
Hatchery 6 6 <0.001 0.430 0.143 0.72
Hatchery 6 6 <0.001 0.002 0.286 0.85
Hatchery 6 6 <0.001 0.038 0.467 0.87
Hatchery 6 6 0.022 0.431 0.054 0.56
Lake trout Hatchery 6 6 0.623 0.051 0.198 0.82
Hatchery 5 5 <0.001 <0.001 0.883 0.74
For each matrix (line), the origin of the parental fish, number of males and females used in the
crossings, as well as probability values for males, females, and their interaction are shown. As the
analyses were performed on regression residuals of final egg size (after swelling) against initial egg
size (before swelling), also r2 values from regression analyses are indicated.
146

size in two wild salmon matrices and one hatchery-reared matrix, two brown
trout matrices, two Arctic charr matrices, and one lake trout matrix. Significant
Figure 1. (A) An example of a salmon matrix in which we found a significant female effect
(F5;24 ¼ 2:69; p ¼ 0:046), male effect (F5;24 ¼ 13:19; p < 0:001) and female �male interaction effect
(F1;24 ¼ 6:32; p ¼ 0:019) on final egg size corrected by the initial egg size. Vertical lines are the
residuals from the regression of final egg size on initial egg size grouped for each female. The
symbols represent different males. (B) Egg swelling for the same matrix as in A. Each circle
indicates the swelling in percentage in the family.
147

male effects were found in four salmon matrices, all of hatchery origin, in all
Arctic charr matrices and one lake trout matrix. The interaction between the
female and the male was significant in one salmon matrix (hatchery-reared fish)
and two Arctic charr matrices. Combined results (Table 3) showed that the
female effect was significant in each species separately, and the male effect was
significant in three out of four species. The female�male interaction was found
to be significant only in Arctic charr.
Salmon had the largest eggs, with the average egg diameter being 6.42 mm
(SD � 0.20) and swelling increased egg diameter on average by 5.2% (SD �1.46) (Fig. 1B). In brown trout, average egg size was 6.30 mm (SD � 0.34) and
swelling 4.2% (SD � 4.22). The Salvelinus sp. had smaller eggs. The average
size of Arctic charr eggs was 5.55 mm (SD � 0.22) and swelling corresponded
to an increase of 4.6% (SD � 1.61), whereas lake trout eggs were on average
5.08 mm (SD � 0.21) and swelling 8.0% (SD � 1.96).
Egg size and its effects on offspring size and size-related fitness character-
istics has been the subject of much research in a number of oviparous organ-
isms (e.g., insects: Fox and Mousseau, 1998; amphibians: Kaplan, 1980, 1998;
Table 3. Combined probability values for female, male, and interaction effect on final egg size for
each species separately. Results for hatchery and wild fish are presented separately as well as
grouped together.
Hatchery Nature Hatchery + nature
�2R ln p 2k p �2R ln p 2k p �2R ln p 2k p
Salmon
Male 57.32 14 0.001 8.72 6 0.238 66.62 20 0.001
Female 20.08 14 0.130 10.92 6 0.125 31.00 20 0.055
M�F 18.85 14 0.207 2.74 6 0.920 19.02 20 0.521
Brown trout
Male 4.58 6 0.677
Female 17.28 6 0.009
M · F 8.07 6 0.238
Arctic charr
Male 50.81 14 <0.001
Female 27.49 14 0.019
M�F 27.11 14 0.019
Lake trout
Male 21.20 4 <0.001
Female 21.60 4 <0.001
M�F 3.49 4 0.558
All species
Male 129.55 32 <0.001 13.30 12 0.348 142.84 44 <0.001
Female 69.18 32 <0.001 28.20 12 0.005 97.38 44 <0.001
M�F 46.88 32 0.043 10.81 12 0.545 57.69 44 0.081
148

fish: Hutchings, 1991; Reznick, 1991; Heath and Blouw, 1998; Einum and
Fleming, 1999). Maternal effects on egg size are widely documented and the
studies have traditionally considered egg size as reflecting the amount of the
resources provided by the mother for the developing embryo (Bernardo,
1996b), thus constraining the size of the hatching individual. In the present
study, we found both the female and the male to affect the final size of the eggs
in three of the four salmonid species. This was despite the fact that we had
controlled for variation in initial egg size by using regression residuals as re-
sponse variables in the analyses. Hence, our results suggest that, at least in
salmonids, the view of egg size as a character entirely determined by the ma-
ternal genotype and phenotype should be revised. Taking into account the
possible male effects on egg size and other life history characters may open new
avenues for research for studies of early life history variation.
Our results indicate that both parents contributed to the final egg size
through the swelling process. Li et al. (1989), on the contrary, suggested that in
salmon swelling is an inherent property of the eggs and is of equal extent even if
the unfertilised eggs differ in size. However, their study design did not allow for
examining the effect of females, nor those of males. In the study of Kjesbu
et al. (1996) with cod, the eggs of individual females tended to show a similar
swelling pattern (measured as egg dry weight/diameter ratio) from year to year.
The authors interpreted such a female effect on the extent of egg swelling to
reflect a strong genetic component. Although the study design we used does not
allow the interpretation of our results in terms of genetic differences among the
females, they are in accordance with those of Kjesbu et al. (1996). The sig-
nificant female � male interaction observed in some matrices in the present
study shows that the final size obtained by the eggs of a certain female may also
depend on the male with whom the eggs have been fertilised. However, the
exact mechanisms by which the female and male affect egg swelling are at
present unknown. The osmotic properties of the egg change when it comes into
contact with water (Alderdice, 1988) and, for example, the sperm of certain
males may have caused the egg to absorb water more efficiently.
The results of our study agree with those of Weigensberg et al. (1998) who
found that, in addition to the indirect effect, the male also had a direct effect on
egg size in the cricket G. firmus. In their study, this effect was a result of
differences in the water uptake and/or metabolism of the embryos fathered by
different males. In fish, egg swelling increases the volume of the perivitelline
fluid and thus depends on the amount of water absorbed by the egg (Alderdice,
1988).
Salmonids spawn on the stream (or lake) bottom, and the eggs develop
buried in the bottom gravel. Therefore, they are likely to be exposed to me-
chanical disturbances and pressure changes by gravel and water movements
(Groot and Alderdice, 1985). The fluid-filled perivitelline space protects the
149

growing embryo from such external disturbances, and thus likely influences its
normal development and survival (Laale, 1980). If the parents influence the
extent of swelling, as our results suggest, this might be reflected in differential
survival of offspring from different families even if the initial egg size might
have been the same. All else being equal, this should select for increased
swelling of the eggs, especially in stream environments where the external
mechanical disturbances are likely to be stronger than in lakes. So far, this
remains speculative as the families in our study could not be incubated sepa-
rately after the photographing and no data on egg survival is therefore avail-
able. A recent study by Lahnsteiner et al. (1999) with brown trout showed a
positive relationship between the average increase in egg size during swelling
and egg viability in batches of eggs from different females. However, it is not
clear whether the process of egg swelling itself produced this outcome or
whether the less swelling eggs were initially less viable. On the other hand, there
is indication that in environments with limited oxygen large egg size may be a
disadvantage (van den Berghe and Gross, 1989). This could be of evolutionary
significance, as incubation of salmonid eggs may take place in low oxygen sites
(Chapman, 1988).
It is unlikely that the volume of the perivitelline fluid (i.e., the extent of
swelling) directly influences larval size. However, it is known that the fluid-
filled perivitelline space (together with the outer egg membrane) acts as a re-
sistance layer for the gas exchange of the developing embryo (Rombough,
1988). Diffusion distances of oxygen will be different in differentially swelled
eggs and may thus influence the oxygen uptake of the embryo and thereby its
growth and well-being. The embryo excretes its metabolic wastes in the peri-
vitelline fluid (Alderdice, 1988), and there may be a minimum volume and thus
minimum egg size needed for normal metabolism. The perivitelline fluid may
serve also other functions (Laale, 1980), but these may not be so clearly con-
nected with the magnitude of swelling (e.g., polyspermy prevention), or be
relevant for salmonids (flotation).
We studied eggs of both hatchery-reared and wild fish. All Arctic charr and
lake trout parents were of hatchery origin, because wild parents were not
available. All brown trout were wild fish, but in salmon, we used both wild and
hatchery-reared parents. We found female effects among both types of fish, but
male effects only among hatchery-reared fish. A possible explanation for this
observation is that the storing may have affected the properties of the gametes
from the wild fish. However, the eggs and milt were stored on ice, and fur-
thermore, sperm motility was checked before fertilisations and only high-
quality sperm was used. On the other hand, the procedures we used for
hatchery fish were closer to the natural as the eggs were fertilised shortly after
stripping. An alternative explanation for finding male effects on egg size only in
the hatchery-reared fish might be related to the fact that the hatchery envi-
150

ronment differs in several respects (e.g., feeding, temperature regimes, inter-
actions between individuals, to mention a few) from the natural environment of
these fishes. For example, Crill et al. (1996) studied the effect of parental en-
vironment on morphological and physiological traits in Drosophila melano-
gaster. They found that the parental environment affected certain traits, and in
particular, egg size was affected by the rearing temperature of the male, so that
the eggs sired by males raised in high temperatures were smaller. Although
Crill et al. (1996) observed cross-generational environmental effects on off-
spring traits, the actual mechanisms how they were mediated are unknown.
Clearly, further studies are needed to settle out this question.
To conclude, we found that, in addition to the female, also the male in
several salmonids influences the egg size through his effect on egg swelling after
fertilisation. Our study thus suggests that paternal contribution may form a
previously largely ignored source of variation in early life history traits.
Keeping in mind the broad scientific interest on maternal effects (Mousseau
and Fox, 1998), to find out the causes and consequences of male effects calls for
further studies. The obvious next steps are to study the occurrence of this
phenomenon in other fish species, as well as the significance of egg swelling and
male-mediated egg size variation on offspring performance and survival in
salmonids. Our study was performed at the family level, and to find out the
magnitude and fitness consequences of egg swelling at the individual level is
another challenge for future research.
Acknowledgements
We thank the Saimaa Fisheries Research and Aquaculture unit of the Finnish
Game and Fisheries Research Institute for providing the facilities for this
study. We also thank Teija Seppa for help in photographing, Jorma Piironen
for discussions and other help, and Erik Petersson for comments on an earlier
version of the manuscript. Our research was funded by the Academy of Fin-
land, Finnish Ministry of Education and the European Commission (FAIR-
CT96-1957).
References
Alderdice, D.F. (1988) Osmotic and ionic regulation in teleost eggs and larvae. In W.S. Hoar and
D.J. Randall (eds) Fish Physiology, Vol. XIA. Academic Press, Inc., London, pp. 163–251.
Bernardo, J. (1996a) The particular maternal effect of propagule size, especially egg size: patterns,
models, quality of evidence and interpretation. Am. Zool. 36, 216–236.
Bernardo, J. (1996b) Maternal effects in animal ecology. Am. Zool. 36, 83–105.
Blaxter, J.H.S. (1986) Development of sense organs and behavior of teleost larvae with special
reference to feeding and predator avoidance. Trans. Am. Fish. Soc. 115, 98–114.
151

Bolin, R.L. (1930) Embryonic development of the labrid fish, Oxyjulis californicus Gunther. Copeia
1930, 122–128.
Chambers, R.C. (1997) Environmental influences on egg and propagule sizes in marine fishes. In
R.C. Chambers and E.A. Trippel, (eds) Early Life History and Recruitment in Fish Populations.
Chapman and Hall, London, pp. 63–102.
Chambers, R.C. and Leggett, W.C. (1996) Maternal influences on variation in egg sizes in tem-
perate marine fishes. Am. Zool. 36, 180–196.
Chapman, D.W. (1988) Critical review of variables used to define effects of fines in redds of large
salmonids. Trans. Am. Fish. Soc. 117, 1–21.
Crill, W.D., Huey, R.B. and Gilchrist, G.W. (1996) Within- and between-generation effects of
temperature on the morphology and physiology of Drosophila melanogaster. Evolution 50, 1205–
1218.
Daan, S., Dijkstra, C. and Tinbergen, J. (1990) Family planning in the kestrel (Falco tinnunculus) –
the ultimate control of covariation of laying date and clutch size. Behaviour 114, 83–116.
Einum, S. and Fleming, I.A. (1999) Maternal effects of egg size in brown trout (Salmo trutta):
norms of reaction to environmental quality. Proc. R. Soc. Lond. B 266, 2095–2100.
Fox, C.W. and Mousseau, T.A. (1998) Maternal effects as adaptations for trans-generational
phenotypic plasticity in insects. In T.A. Mousseau and C.W. Fox (eds) Maternal Effects as
Adaptations. Oxford University Press, New York, pp. 159–177.
Groot, E.P. and Alderdice, D.F. (1985) Fine structure of the external egg membrane of five species
of Pacific salmon and steelhead trout. Can. J. Zool. 63, 552–566.
Gwynne, D.T. (1984) Courtship feeding increases female reproductive success in bushcrickets.
Nature 307, 361–363.
Heath, D.D. and Blouw, D.M. (1998) Are maternal effects in fish adaptive or merely physiological
side effects? In T.A. Mousseau and C.W. Fox (eds) Maternal Effects as Adaptations. Oxford
University Press, New York, pp. 178–201.
Hutchings, J.A. (1991) Fitness consequences of variation in egg size and food abundance in brook
trout Salvelinus fontinalis. Evolution 45, 1162–1168.
Kamler, E. (1992) Early Life History of Fish: an Energetics Approach. Chapman and Hall, London.
Kaplan, R.H. (1980) The implications of ovum size variability for offspring fitness and clutch size
within several populations of salamanders (Ambystoma). Evolution 34, 51–64.
Kaplan, R.H. (1998) Maternal effects, developmental plasticity, and life history evolution: an
amphibian model. In T.A. Mousseau and C.W. Fox (eds) Maternal Effects as Adaptations,
Oxford University Press, New York, pp. 244–260.
Kjesbu, O.S., Solemdal, P., Bratland, P. and Fonn, M. (1996) Variation in annual egg production
in individual captive Atlantic cod (Gadus morhua). Can. J. Fish. Aquat. Sci. 53, 610–620.
Laale, H.W. (1980) The perivitelline space and egg envelopes of bony fishes: a review. Copeia 1980,
210–226.
Lahnsteiner, F., Weismann, T. and Patzner, R.A. (1999) Physiological and biochemical parameters
for egg quality determination in lake trout, Salmo trutta lacustris. Fish Phys. Biochem. 20, 375–
388.
Li, X., Jenssen, E. and Fyhn, H.J. (1989) Effects of salinity on egg swelling in Atlantic salmon
(Salmo salar). Aquaculture 76, 317–334.
Lønning, S. and Davenport, J. (1980) The swelling egg of the rough dab, Hippoglossoides plates-
soides limandoides (Bloch). J. Fish Biol. 17, 359–378.
Miller, T.J., Crowder, L.B., Rice, J.A. and Marschall, E.A. (1988) Larval size and recruitment
mechanisms in fishes: toward a conceptual framework. Can. J. Fish. Aquat. Sci. 45, 1657–1670.
Mousseau, T.A. and Fox, C.W. (1998) The adaptive significance of maternal effects. Trends Ecol.
Evol. 13, 403–407.
Reznick, D. (1991) Maternal effects in fish life histories. In E. Dudley (ed.) Evolutionary Biology.
Discorides Press, Portland, Oregon, pp. 780–793.
Rombough, P.J. (1988) Respiratory gas exchange, aerobic metabolism, and effects of hypoxia
during early life. In W.S. Hoar and D.J. Randall (eds) Fish Physiology, Vol. XIA. Academic
Press, Inc., London, pp. 59–161.
152

Sakaluk, S.K. (1986) Sperm competition and the evolution of nuptial feeding behavior in the
cricket, Gryllodes supplicans (Walker). Evolution 40, 584–593.
Sokal, R.R. and Rohlf, F.J. (1995) Biometry: the Principles and Practice of Statistics in Biological
Research. 3rd edn. W.H. Freeman and Company, New York.
Tukey, J.W. (1949) One degree of freedom for non-additivity. Biometrics 5, 232–242.
van den Berghe, E.P. and Gross, M.R. (1989) Natural selection resulting from female breeding
competition in a Pacific salmon (coho: Oncorhynchus kisutch). Evolution 43, 125–140.
Weigensberg, I., Carriere, Y. and Roff, D.A. (1998) Effects of male genetic contribution and
paternal investment to egg and hatchling size in the cricket, Gryllus firmus. J. Evol. Biol. 11, 135–
146.
153
Top Related
Copyright © 2022 FDOKUMEN

