







Bahasa
Halaman
Hukum


Epichloe spp. associated with grasses:new insights on life cycles, dissemination and evolution
Mariusz Tadych1
Marshall S. BergenJames F. White Jr.
Department of Plant Biology and Pathology, RutgersUniversity, New Brunswick, New Jersey 08901-8520
Abstract: Epichloe species with their asexual statesare specialized fungi associated with cool-seasongrasses. They grow endophytically in tissues of aerialparts of host plants to form systemic and mostlyasymptomatic associations. Their life cycles mayinvolve vertical transmission through host seedsand/or horizontal transmission from one plant toother plants of the same species through fungalpropagules. Vertical transmission has been wellstudied, but comparatively little research has beendone on horizontal dissemination. The goal of thisreview is to provide new insights on modes ofdissemination of systemic grass endophytes. Thereview addresses recent progress in research on (i)the process of growth of Epichloe endophytes in thehost plant tissues, (ii) the types and development ofreproductive structures of the endophyte, (iii) therole of the reproductive structures in endophytedissemination and host plant infection processes and(iv) some ecological and evolutionary implications oftheir modes of dissemination. Research in theEpichloe grass endophytes has accelerated in the past25 y and has demonstrated the enormous complexityin endophyte-grass symbioses. There still remain largegaps in our understanding of the role and functionsof these fungi in agricultural systems and understand-ing the functions, ecology and evolution of theseendophytes in natural grass populations.
Key words: Clavicipitaceae, conidiation, endo-phytes, epibiotic, horizontal dissemination, hybridiza-tion, vertical dissemination
INTRODUCTION
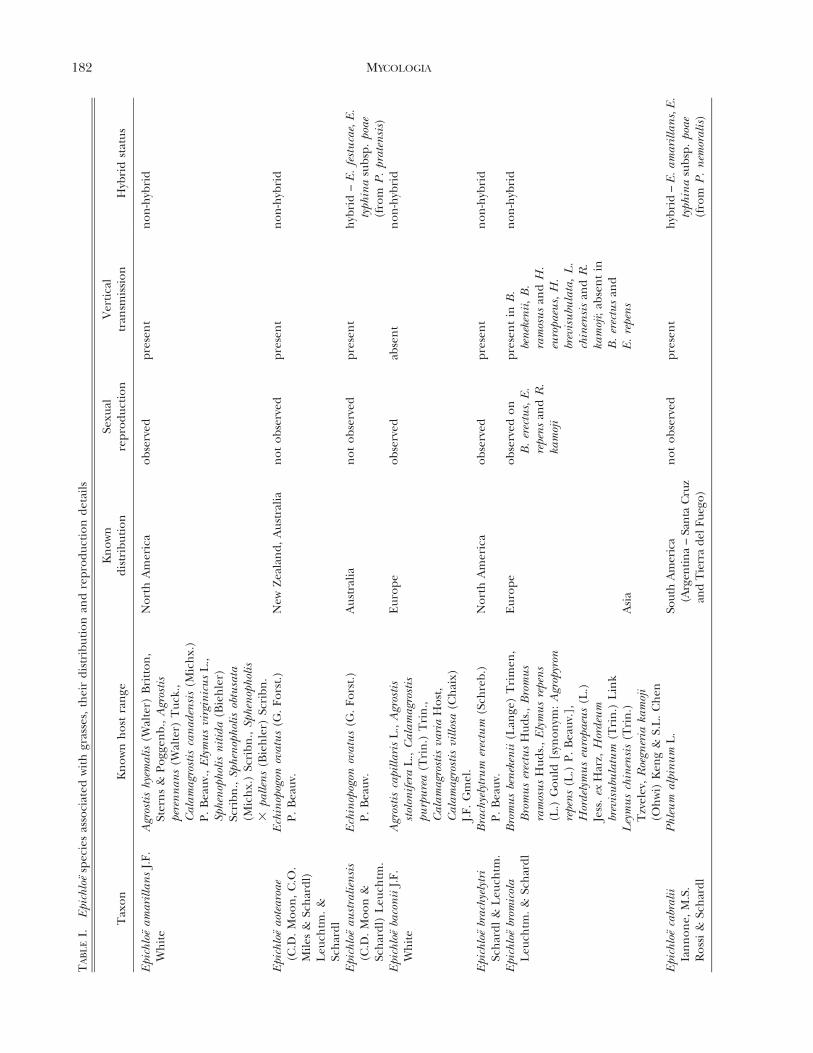
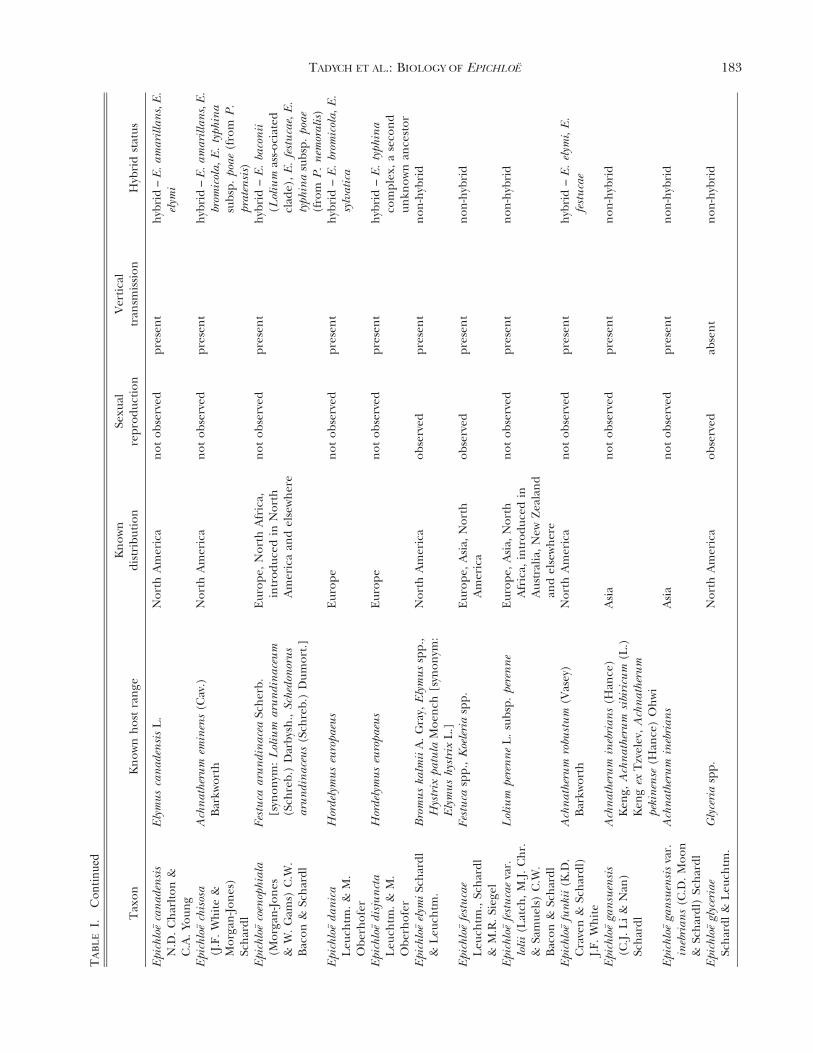
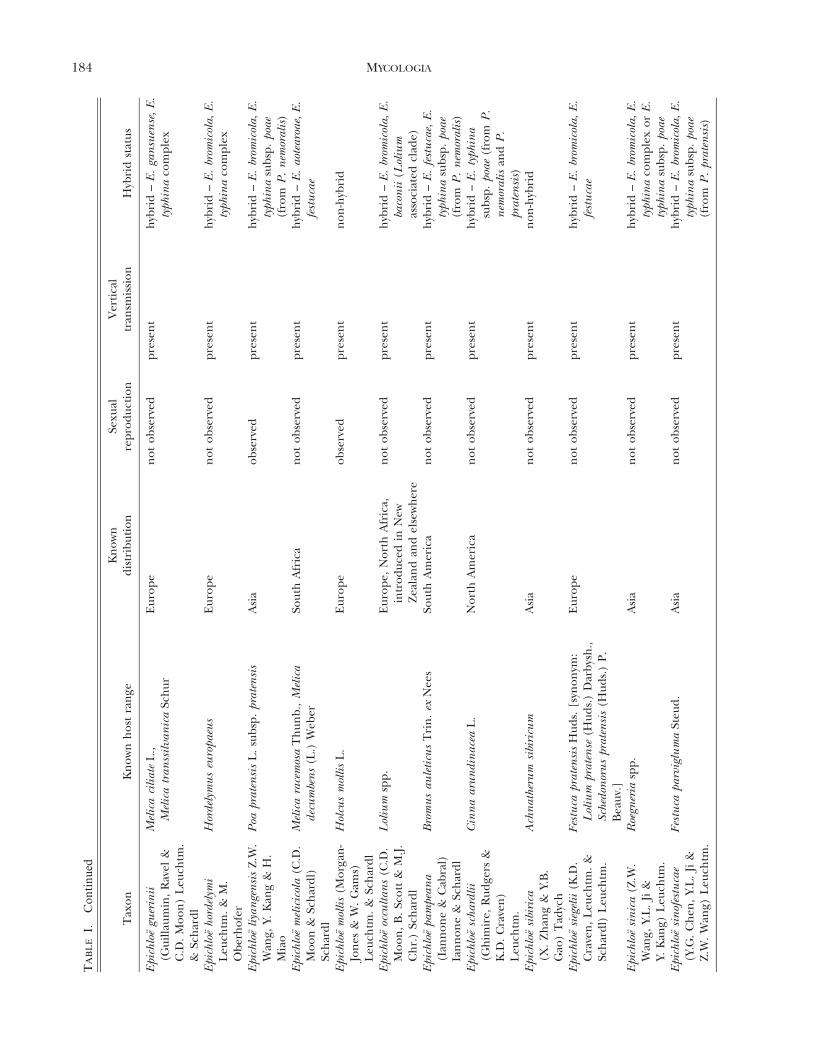
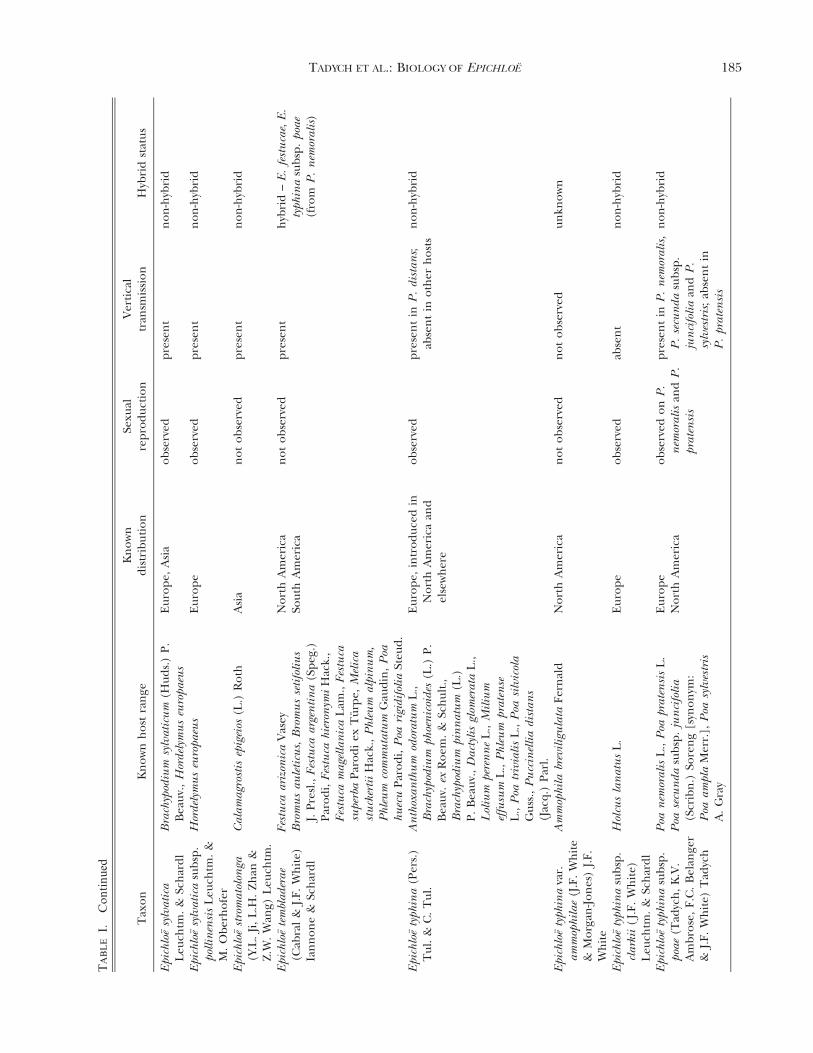
Epichloe (Fr.) Tul. & C. Tul. species (Clavicipitaceae,Hypocreales, Ascomycota), including their asexualstates, are fungal symbionts of cool-season grasses(Pooideae, Poaceae; TABLE I) (Clay 1990, Glenn et al.1996, Schardl et al. 2004, Spatafora et al. 2007,Leuchtmann et al. 2014). It has been estimated that
up to 30% of all cool-season grass species worldwideare associated with Epichloe spp. (Leuchtmann 1992).These fungi are characterized by their endophytichabit to form systemic and asymptomatic associationsthroughout the aerial parts of the host plant, wherethrough intercellular growth of hyphae they colonizethe apoplastic spaces of culms, leaf sheaths and seeds(Clay 1990; White et al. 1991; Christensen et al. 1997,2002, 2008; Christensen and Voisey 2009). Because oftheir systemic growth, species of Epichloe are fre-quently seed-transmitted (TABLE I) to the next gen-eration of the host plant (i.e. when the seedgerminates, the resulting seedling may contain thefungal symbiont) (Sampson 1933, Siegel et al. 1984,White 1988, Clay 1990, White et al. 1991). However,many Epichloe species, in addition to having primarilyendophytic growth, produce fungal structures thatappear externally on the aboveground plant parts(FIG. 1). The external structures consist of: (i)mycelial networks with conidiogenous cells andconidia, growing mostly epibiotically on leaf blades(White et al. 1996, Christensen et al. 1997, Meijer andLeuchtmann 1999, Moy et al. 2000, Craven et al. 2001,Dugan et al. 2002, Moon et al. 2002, Tadych andWhite 2007, Tadych et al. 2007, Iannone et al. 2009,Oberhofer 2012, Alderman 2013), and (ii) light-colored stromata (i.e. dense masses of hyphae, whichusually envelop inflorescences and upper leaf sheathsof flowering culms and prevent flower and seeddevelopment, a syndrome referred to as ‘‘chokedisease’’ (Bradshaw 1959; White 1988; White et al.1991; Schardl et al. 2004, 2009; Schardl 2010;Christensen et al. 2012).
The relationship of the endophytes to hosts ishighly variable and has been suggested to be anantagonism-mutualism continuum depending onparticular host genotype and endophyte-haplotypecombinations and environmental conditions (Saikko-nen et al. 2004, 2006, 2010; Cheplick and Faeth 2009;Brosi et al. 2012). Epichloe endophytes may modify themetabolic activities of colonized hosts (e.g. they maycause a change in levels of antioxidants and othermetabolites in hosts tissues) (Malinowski et al. 1998;Rasmussen et al. 2008, 2009; White and Torres 2010;Brosi et al. 2012; Hamilton and Bauerle 2012;Hamilton et al. 2012; Koulman et al. 2012; Qawasmehet al. 2012a, b; Torres et al. 2012). The endophytesalso produce a range of secondary metabolites (Bushet al. 1997; Clay and Schardl 2002; Schardl et al. 2012,
Submitted 3 Oct 2013; accepted for publication 10 Oct 2013.1 Corresponding author. E-mail: [email protected]
Mycologia, 106(2), 2014, pp. 181–201. DOI: 10.3852/106.2.181# 2014 by The Mycological Society of America, Lawrence, KS 66044-8897Issued 22 April 2014
181
TA
BL
EI.
Epic
hlo
esp
ecie
sas
soci
ated
wit
hgr
asse
s,th
eir
dis
trib
uti
on
and
rep
rod
uct
ion
det
ails
Tax
on
Kn
ow
nh
ost
ran
geK
no
wn
dis
trib
uti
on
Sexu
alre
pro
du
ctio
nV
erti
cal
tran
smis
sio
nH
ybri
dst
atu
s
Epic
hlo
eam
ari
llan
sJ.
F.
Wh
ite
Agr
osti
shye
mali
s(W
alte
r)B
ritt
on
,St
ern
s&
Po
ggen
b.,
Agr
osti
sper
enn
an
s(W
alte
r)T
uck
.,C
ala
magr
osti
sca
naden
sis
(Mic
hx.
)P
.B
eau
v.,
Ely
mu
svir
gin
icu
sL
.,Sphen
ophol
isn
itid
a(B
ieh
ler)
Scri
bn
.,Sphen
ophol
isob
tusa
ta(M
ich
x.)
Scri
bn
.,Sphen
ophol
is3
pall
ens
(Bie
hle
r)Sc
rib
n.
No
rth
Am
eric
ao
bse
rved
pre
sen
tn
on
-hyb
rid
Epic
hlo
eaot
earo
ae
(C.D
.M
oo
n,
C.O
.M
iles
&Sc
har
dl)
Leu
chtm
.&
Sch
ard
l
Ech
inop
ogon
ovatu
s(G
.F
ors
t.)
P.
Bea
uv.
New
Zea
lan
d,
Au
stra
lia
no
to
bse
rved
pre
sen
tn
on
-hyb
rid
Epic
hlo
eau
stra
lien
sis
(C.D
.M
oo
n&
Sch
ard
l)L
euch
tm.
Ech
inop
ogon
ovatu
s(G
.F
ors
t.)
P.
Bea
uv.
Au
stra
lia
no
to
bse
rved
pre
sen
th
ybri
d–
E.
fest
uca
e,E
.ty
phin
asu
bsp
.poa
e(f
rom
P.
pra
ten
sis)
Epic
hlo
eba
con
iiJ.
F.
Wh
ite
Agr
osti
sca
pil
lari
sL
.,A
gros
tis
stol
onif
era
L.,
Cala
magr
osti
spu
rpu
rea
(Tri
n.)
Tri
n.,
Cala
magr
osti
svari
aH
ost
,C
ala
magr
osti
svil
losa
(Ch
aix)
J.F
.G
mel
.
Eu
rop
eo
bse
rved
abse
nt
no
n-h
ybri
d
Epic
hlo
ebr
ach
yely
tri
Sch
ard
l&
Leu
chtm
.B
rach
yely
tru
mer
ectu
m(S
chre
b.)
P.
Bea
uv.
No
rth
Am
eric
ao
bse
rved
pre
sen
tn
on
-hyb
rid
Epic
hlo
ebr
omic
ola
Leu
chtm
.&
Sch
ard
lB
rom
us
ben
eken
ii(L
ange
)T
rim
en,
Bro
mu
ser
ectu
sH
ud
s.,
Bro
mu
sra
mos
us
Hu
ds.
,E
lym
us
repen
s(L
.)G
ou
ld[s
yno
nym
:A
grop
yron
repen
s(L
.)P
.B
eau
v.],
Hor
del
ymu
seu
ropaeu
s(L
.)Je
ss.
exH
arz,
Hor
deu
mbr
evis
ubu
latu
m(T
rin
.)L
ink
Ley
mu
sch
inen
sis
(Tri
n.)
Tzv
elev
,R
oegn
eria
kam
oji
(Oh
wi)
Ken
g&
S.L
.C
hen
Eu
rop
e
Asi
a
ob
serv
edo
nB
.er
ectu
s,E
.re
pen
san
dR
.ka
moj
i
pre
sen
tin
B.
ben
eken
ii,
B.
ram
osu
san
dH
.eu
ropaeu
s ,H
.br
evis
ubu
lata
,L
.ch
inen
sis
and
R.
kam
oji ;
abse
nt
inB
.er
ectu
san
dE
.re
pen
s
no
n-h
ybri
d
Epic
hlo
eca
brali
iIa
nn
on
e,M
.S.
Ro
ssi
&Sc
har
dl
Phle
um
alp
inu
mL
.So
uth
Am
eric
a(A
rgen
tin
a–
San
taC
ruz
and
Tie
rra
del
Fueg
o)
no
to
bse
rved
pre
sen
th
ybri
d–
E.
am
ari
llan
s,E
.ty
phin
asu
bsp
.poa
e(f
rom
P.
nem
orali
s)
182 MYCOLOGIA
TA
BL
EI.
Co
nti
nu
ed
Tax
on
Kn
ow
nh
ost
ran
geK
no
wn
dis
trib
uti
on
Sexu
alre
pro
du
ctio
nV
erti
cal
tran
smis
sio
nH
ybri
dst
atu
s
Epic
hlo
eca
naden
sis
N.D
.C
har
lto
n&
C.A
.Yo
un
g
Ely
mu
sca
naden
sis
L.
No
rth
Am
eric
an
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.am
ari
llan
s,E
.el
ymi
Epic
hlo
ech
isos
a(J
.F.
Wh
ite
&M
org
an-J
on
es)
Sch
ard
l
Ach
nath
eru
mem
inen
s(C
av.)
Bar
kwo
rth
No
rth
Am
eric
an
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.am
ari
llan
s,E
.br
omic
ola
,E
.ty
phin
asu
bsp
.poa
e(f
rom
P.
pra
ten
sis )
Epic
hlo
eco
enop
hia
la(M
org
an-J
on
es&
W.
Gam
s)C
.W.
Bac
on
&Sc
har
dl
Fes
tuca
aru
ndin
ace
aSc
her
b.
[syn
on
ym:
Lol
ium
aru
ndin
ace
um
(Sch
reb
.)D
arb
ysh
.,Sch
edon
oru
saru
ndin
ace
us
(Sch
reb
.)D
um
ort
.]
Eu
rop
e,N
ort
hA
fric
a,in
tro
du
ced
inN
ort
hA
mer
ica
and
else
wh
ere
no
to
bse
rved
pre
sen
th
ybri
d–
E.
baco
nii
(Lol
ium
ass-
oci
ated
clad
e),
E.
fest
uca
e,E
.ty
phin
asu
bsp
.poa
e(f
rom
P.
nem
orali
s)E
pic
hlo
edan
ica
Leu
chtm
.&
M.
Ob
erh
ofe
r
Hor
del
ymu
seu
ropaeu
sE
uro
pe
no
to
bse
rved
pre
sen
th
ybri
d–
E.
brom
icol
a,
E.
sylv
ati
ca
Epic
hlo
edis
jun
cta
Leu
chtm
.&
M.
Ob
erh
ofe
r
Hor
del
ymu
seu
ropaeu
sE
uro
pe
no
to
bse
rved
pre
sen
th
ybri
d–
E.
typhin
aco
mp
lex,
ase
con
du
nkn
ow
nan
cest
or
Epic
hlo
eel
ymi
Sch
ard
l&
Leu
chtm
.B
rom
us
kalm
iiA
.G
ray,
Ely
mu
ssp
p.,
Hys
trix
patu
laM
oen
ch[s
yno
nym
:E
lym
us
hys
trix
L.]
No
rth
Am
eric
ao
bse
rved
pre
sen
tn
on
-hyb
rid
Epic
hlo
efe
stu
cae
Leu
chtm
.,Sc
har
dl
&M
.R.
Sieg
el
Fes
tuca
spp
.,K
oele
ria
spp
.E
uro
pe,
Asi
a,N
ort
hA
mer
ica
ob
serv
edp
rese
nt
no
n-h
ybri
d
Epic
hlo
efe
stu
cae
var.
loli
i(L
atch
,M
.J.
Ch
r.&
Sam
uel
s)C
.W.
Bac
on
&Sc
har
dl
Lol
ium
per
enn
eL
.su
bsp
.per
enn
eE
uro
pe,
Asi
a,N
ort
hA
fric
a,in
tro
du
ced
inA
ust
rali
a,N
ewZ
eala
nd
and
else
wh
ere
no
to
bse
rved
pre
sen
tn
on
-hyb
rid
Epic
hlo
efu
nki
i(K
.D.
Cra
ven
&Sc
har
dl)
J.F
.W
hit
e
Ach
nath
eru
mro
bust
um
(Vas
ey)
Bar
kwo
rth
No
rth
Am
eric
an
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.el
ymi,
E.
fest
uca
e
Epic
hlo
ega
nsu
ensi
s(C
.J.
Li
&N
an)
Sch
ard
l
Ach
nath
eru
min
ebri
an
s(H
ance
)K
eng,
Ach
nath
eru
msi
biri
cum
(L.)
Ken
gex
Tzv
elev
,A
chn
ath
eru
mpek
inen
se(H
ance
)O
hw
i
Asi
an
ot
ob
serv
edp
rese
nt
no
n-h
ybri
d
Epic
hlo
ega
nsu
ensi
sva
r.in
ebri
an
s(C
.D.
Mo
on
&Sc
har
dl)
Sch
ard
l
Ach
nath
eru
min
ebri
an
sA
sia
no
to
bse
rved
pre
sen
tn
on
-hyb
rid
Epic
hlo
egl
ycer
iae
Sch
ard
l&
Leu
chtm
.G
lyce
ria
spp
.N
ort
hA
mer
ica
ob
serv
edab
sen
tn
on
-hyb
rid
TADYCH ET AL.: BIOLOGY OF EPICHLOE 183
TA
BL
EI.
Co
nti
nu
ed
Tax
on
Kn
ow
nh
ost
ran
geK
no
wn
dis
trib
uti
on
Sexu
alre
pro
du
ctio
nV
erti
cal
tran
smis
sio
nH
ybri
dst
atu
s
Epic
hlo
egu
erin
ii(G
uil
lau
min
,R
avel
&C
.D.
Mo
on
)L
euch
tm.
&Sc
har
dl
Mel
ica
cili
ate
L.,
Mel
ica
tran
ssil
van
ica
Sch
ur
Eu
rop
en
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.ga
nsu
ense
,E
.ty
phin
aco
mp
lex
Epic
hlo
ehor
del
ymi
Leu
chtm
.&
M.
Ob
erh
ofe
r
Hor
del
ymu
seu
ropaeu
sE
uro
pe
no
to
bse
rved
pre
sen
th
ybri
d–
E.
brom
icol
a,
E.
typhin
aco
mp
lex
Epic
hlo
eli
yan
gen
sis
Z.W
.W
ang,
Y.K
ang
&H
.M
iao
Poa
pra
ten
sis
L.
sub
sp.
pra
ten
sis
Asi
ao
bse
rved
pre
sen
th
ybri
d–
E.
brom
icol
a,
E.
typhin
asu
bsp
.poa
e(f
rom
P.
nem
orali
s)E
pic
hlo
em
elic
icol
a(C
.D.
Mo
on
&Sc
har
dl)
Sch
ard
l
Mel
ica
race
mos
aT
hu
nb
.,M
elic
adec
um
ben
s(L
.)W
eber
Sou
thA
fric
an
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.aot
earo
ae,
E.
fest
uca
e
Epic
hlo
em
olli
s(M
org
an-
Jon
es&
W.
Gam
s)L
euch
tm.
&Sc
har
dl
Hol
cus
mol
lis
L.
Eu
rop
eo
bse
rved
pre
sen
tn
on
-hyb
rid
Epic
hlo
eoc
cult
an
s(C
.D.
Mo
on
,B
.Sc
ott
&M
.J.
Ch
r.)
Sch
ard
l
Lol
ium
spp
.E
uro
pe,
No
rth
Afr
ica,
intr
od
uce
din
New
Zea
lan
dan
del
sew
her
e
no
to
bse
rved
pre
sen
th
ybri
d–
E.
brom
icol
a,
E.
baco
nii
(Lol
ium
asso
ciat
edcl
ade)
Epic
hlo
epam
pea
na
(Ian
no
ne
&C
abra
l)Ia
nn
on
e&
Sch
ard
l
Bro
mu
sau
leti
cus
Tri
n.
exN
ees
Sou
thA
mer
ica
no
to
bse
rved
pre
sen
th
ybri
d–
E.
fest
uca
e,E
.ty
phin
asu
bsp
.poa
e(f
rom
P.
nem
orali
s)E
pic
hlo
esc
hard
lii
(Gh
imir
e,R
ud
gers
&K
.D.
Cra
ven
)L
euch
tm.
Cin
na
aru
ndin
ace
aL
.N
ort
hA
mer
ica
no
to
bse
rved
pre
sen
th
ybri
d–
E.
typhin
asu
bsp
.poa
e(f
rom
P.
nem
orali
san
dP
.pra
ten
sis )
Epic
hlo
esi
biri
ca(X
.Z
han
g&
Y.B
.G
ao)
Tad
ych
Ach
nath
eru
msi
biri
cum
Asi
an
ot
ob
serv
edp
rese
nt
no
n-h
ybri
d
Epic
hlo
esi
egel
ii(K
.D.
Cra
ven
,L
euch
tm.
&Sc
har
dl)
Leu
chtm
.
Fes
tuca
pra
ten
sis
Hu
ds.
[syn
on
ym:
Lol
ium
pra
ten
se(H
ud
s.)
Dar
bys
h.,
Sch
edon
oru
spra
ten
sis
(Hu
ds.
)P
.B
eau
v.]
Eu
rop
en
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.br
omic
ola
,E
.fe
stu
cae
Epic
hlo
esi
nic
a(Z
.W.
Wan
g,Y.
L.
Ji&
Y.K
ang)
Leu
chtm
.
Roe
gner
iasp
p.
Asi
an
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.br
omic
ola
,E
.ty
phin
aco
mp
lex
or
E.
typhin
asu
bsp
.poa
eE
pic
hlo
esi
nof
estu
cae
(Y.G
.C
hen
,Y.
L.
Ji&
Z.W
.W
ang)
Leu
chtm
.
Fes
tuca
parv
iglu
ma
Steu
d.
Asi
an
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.br
omic
ola
,E
.ty
phin
asu
bsp
.poa
e(f
rom
P.
pra
ten
sis)
184 MYCOLOGIA
TA
BL
EI.
Co
nti
nu
ed
Tax
on
Kn
ow
nh
ost
ran
geK
no
wn
dis
trib
uti
on
Sexu
alre
pro
du
ctio
nV
erti
cal
tran
smis
sio
nH
ybri
dst
atu
s
Epic
hlo
esy
lvati
caL
euch
tm.
&Sc
har
dl
Bra
chyp
odiu
msy
lvati
cum
(Hu
ds.
)P
.B
eau
v.,
Hor
del
ymu
seu
ropaeu
sE
uro
pe,
Asi
ao
bse
rved
pre
sen
tn
on
-hyb
rid
Epic
hlo
esy
lvati
casu
bsp
.pol
lin
ensi
sL
euch
tm.
&M
.O
ber
ho
fer
Hor
del
ymu
seu
ropaeu
sE
uro
pe
ob
serv
edp
rese
nt
no
n-h
ybri
d
Epic
hlo
est
rom
ato
lon
ga(Y
.L.
Ji,
L.H
.Z
han
&Z
.W.
Wan
g)L
euch
tm.
Cala
magr
osti
sep
igei
os(L
.)R
oth
Asi
an
ot
ob
serv
edp
rese
nt
no
n-h
ybri
d
Epic
hlo
ete
mbl
ader
ae
(Cab
ral
&J.
F.
Wh
ite)
Ian
no
ne
&Sc
har
dl
Fes
tuca
ari
zon
ica
Vas
eyB
rom
us
au
leti
cus,
Bro
mu
sse
tifo
liu
sJ.
Pre
sl.,
Fes
tuca
arg
enti
na
(Sp
eg.)
Par
od
i,Fes
tuca
hie
ron
ymi
Hac
k.,
Fes
tuca
mage
llan
ica
Lam
.,Fes
tuca
super
baP
aro
di
exT
urp
e,M
elic
ast
uck
erti
iH
ack.
,P
hle
um
alp
inu
m,
Phle
um
com
mu
tatu
mG
aud
in,
Poa
hu
ecu
Par
od
i,P
oari
gidif
olia
Steu
d.
No
rth
Am
eric
aSo
uth
Am
eric
an
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.fe
stu
cae,
E.
typhin
asu
bsp
.poa
e(f
rom
P.
nem
orali
s)
Epic
hlo
ety
phin
a(P
ers.
)T
ul.
&C
.T
ul.
An
thox
an
thu
mod
oratu
mL
.,B
rach
ypod
ium
phoe
nic
oides
(L.)
P.
Bea
uv.
exR
oem
.&
Sch
ult
.,B
rach
ypod
ium
pin
natu
m(L
.)P
.B
eau
v.,
Dact
ylis
glom
erata
L.,
Lol
ium
per
enn
eL
.,M
iliu
mef
fusu
mL
.,P
hle
um
pra
ten
seL
.,P
oatr
ivia
lis
L.,
Poa
silv
icol
aG
uss
.,P
ucc
inel
lia
dis
tan
s(J
acq
.)P
arl.
Eu
rop
e,in
tro
du
ced
inN
ort
hA
mer
ica
and
else
wh
ere
ob
serv
edp
rese
nt
inP
.dis
tan
s;ab
sen
tin
oth
erh
ost
sn
on
-hyb
rid
Epic
hlo
ety
phin
ava
r.am
mop
hil
ae
(J.F
.W
hit
e&
Mo
rgan
-Jo
nes
)J.
F.
Wh
ite
Am
mop
hil
abr
evil
igu
lata
Fer
nal
dN
ort
hA
mer
ica
no
to
bse
rved
no
to
bse
rved
un
kno
wn
Epic
hlo
ety
phin
asu
bsp
.cl
ark
ii(
J.F
.W
hit
e)L
euch
tm.
&Sc
har
dl
Hol
cus
lan
atu
sL
.E
uro
pe
ob
serv
edab
sen
tn
on
-hyb
rid
Epic
hlo
ety
phin
asu
bsp
.poa
e(T
adyc
h,
K.V
.A
mb
rose
,F
.C.
Bel
ange
r&
J.F
.W
hit
e)T
adyc
h
Poa
nem
orali
sL
.,P
oapra
ten
sis
L.
Poa
secu
nda
sub
sp.
jun
cifo
lia
(Scr
ibn
.)So
ren
g[s
yno
nym
:P
oaam
pla
Mer
r.],
Poa
sylv
estr
isA
.G
ray
Eu
rop
eN
ort
hA
mer
ica
ob
serv
edo
nP
.n
emor
ali
san
dP
.pra
ten
sis
pre
sen
tin
P.
nem
orali
s,P
.se
cun
da
sub
sp.
jun
cifo
lia
and
P.
sylv
estr
is;
abse
nt
inP
.pra
ten
sis
no
n-h
ybri
d
TADYCH ET AL.: BIOLOGY OF EPICHLOE 185

2013; Takach et al. 2012), including ergot alkaloids,indole-deterpenes, aminopyrrolizidines (lolines)and pyrrolopyrazine (peramine) (Gallagher et al.1984, Lyons et al. 1986, Rowan and Gaynor 1986,Bush et al. 1993). Lolines and peramine exhibitinsecticidal properties, whereas ergovaline (ergotalkaloid) and lolitrem B (indole-diterpene) arepoisonous to grazing livestock. For example, ergova-line and lolitrem B produced by E. coenophiala andE. festucae var. lolii have been established as thecausal agent of fescue toxicosis in cattle (Bacon et al.1977, Schmidt et al. 1982) and ryegrass staggers insheep (Fletcher and Harvey 1981, Latch et al. 1984)respectively. In addition, these metabolites arethought to cause the documented complex effectsof Epichloe spp. in enhancing herbivore deterrence(Belesky et al. 1987; Arechevaleta et al. 1989; Clay1990, Bazely et al. 1997, 2007; Malinowski andBelesky 2000; Panaccione et al. 2001; Ahlholmet al. 2002; Clay and Schardl 2002; Faeth 2002,2009; Faeth and Sullivan 2003; Schardl et al. 2004;Lehtonen et al. 2005; Granath et al. 2007; Olejniczakand Lembicz 2007; Rudgers and Clay 2008; Crawfordet al. 2010; Saikkonen et al. 2010; Schardl 2010;Lembicz et al. 2011; Bultman et al. 2012; Faeth andSarri 2012; Gibert et al. 2012; Gundel et al. 2012,2013; Hamilton et al. 2012; Torres et al. 2012;Czarnoleski et al. 2013). Other classes of fungalmetabolites, siderophores (e.g. epichloenin A, fer-riepichloenin A, epichloenin B, also have beenfound in both endophyte-infected plants and endo-phyte mycelia ( Johnson et al. 2007a, b; Koulmanet al. 2007, 2012; Cao et al. 2008). The function ofsiderophores in the Epichloe-grass associations is notfully understood. They have been proposed to play arole in resistance in endophyte-infected grass plants(Rasmussen et al. 2008). It also has been postulatedthat extracellular siderophore, epichloenin A, mayserve as an important molecular/cellular signal forcontrolling fungal growth and hence the symbioticinteraction of E. festuca with L. perenne ( Johnsonet al. 2013).
Epichloe spp. reproduce by three modes: (i) sexualreproduction resulting in stromata development onflowering culms, (ii) seed colonization from systemicinfections or (iii) a mixed mode where both floweringculms choked by stromata and healthy floweringculms with seeds colonized by endophytic myceliumare present. Epichloe-Pooideae symbioses have longbeen considered static associations. However, thebiology and ecology of Epichloe spp. indicate thatthese fungi may have a more complex life cycle andform more dynamic associations with their hosts.
The subject of this review is to discuss the variationsin life cycle among different species of both theT
AB
LE
I.C
on
tin
ued
Tax
on
Kn
ow
nh
ost
ran
geK
no
wn
dis
trib
uti
on
Sexu
alre
pro
du
ctio
nV
erti
cal
tran
smis
sio
nH
ybri
dst
atu
s
Epic
hlo
ety
phin
asu
bsp
.poa
eva
r.aon
iken
kan
aIa
nn
on
e&
Sch
ard
l
Bro
mu
sse
tifo
liu
sSo
uth
Am
eric
a(A
rgen
tin
a–
San
taC
ruz)
no
to
bse
rved
pre
sen
tn
on
-hyb
rid
Epic
hlo
ety
phin
asu
bsp
.poa
eva
r.ca
nari
ensi
s(C
.D.
Mo
on
,B
.Sc
ott
&M
.J.
Ch
r.)
Leu
chtm
.
Lol
ium
edw
ard
iiH
.Sc
ho
lz,
Stie
rst.
&G
aisb
erg
[syn
on
ym:
Lol
ium
can
ari
ense
Steu
d.]
Afr
ica
(Can
ary
Isla
nd
s)n
ot
ob
serv
edp
rese
nt
no
n-h
ybri
d
Epic
hlo
ety
phin
asu
bsp
.poa
eva
r.hu
erfa
na
(J.F
.W
hit
e,G
.T.
Co
le&
Mo
rgan
-Jo
nes
)T
adyc
h&
Leu
chtm
.
Fes
tuca
ari
zon
ica
No
rth
Am
eric
an
ot
ob
serv
edp
rese
nt
no
n-h
ybri
d
Epic
hlo
eu
nci
nata
(W.
Gam
s,P
etri
ni
&D
.Sc
hm
idt)
Leu
chtm
.&
Sch
ard
l
Fes
tuca
pra
ten
sis
[syn
on
ym:
Lol
ium
pra
ten
se,
Sch
edon
oru
spra
ten
sis]
Eu
rop
en
ot
ob
serv
edp
rese
nt
hyb
rid
–E
.br
omic
ola
,E
.ty
phin
asu
bsp
.poa
e(f
rom
P.
nem
orali
s)
186 MYCOLOGIA
sexual and asexual Epichloe spp. associated withgrasses. This review will concentrate on (i) the processof internal growth of Epichloe endophytes in thevegetative and reproductive tissues of the host plant,
(ii) epibiotic growth and development of sexual and/or asexual reproductive structures by these fungi, (iii)the roles of these fungal structures in endophytedissemination and host plant infection processes and
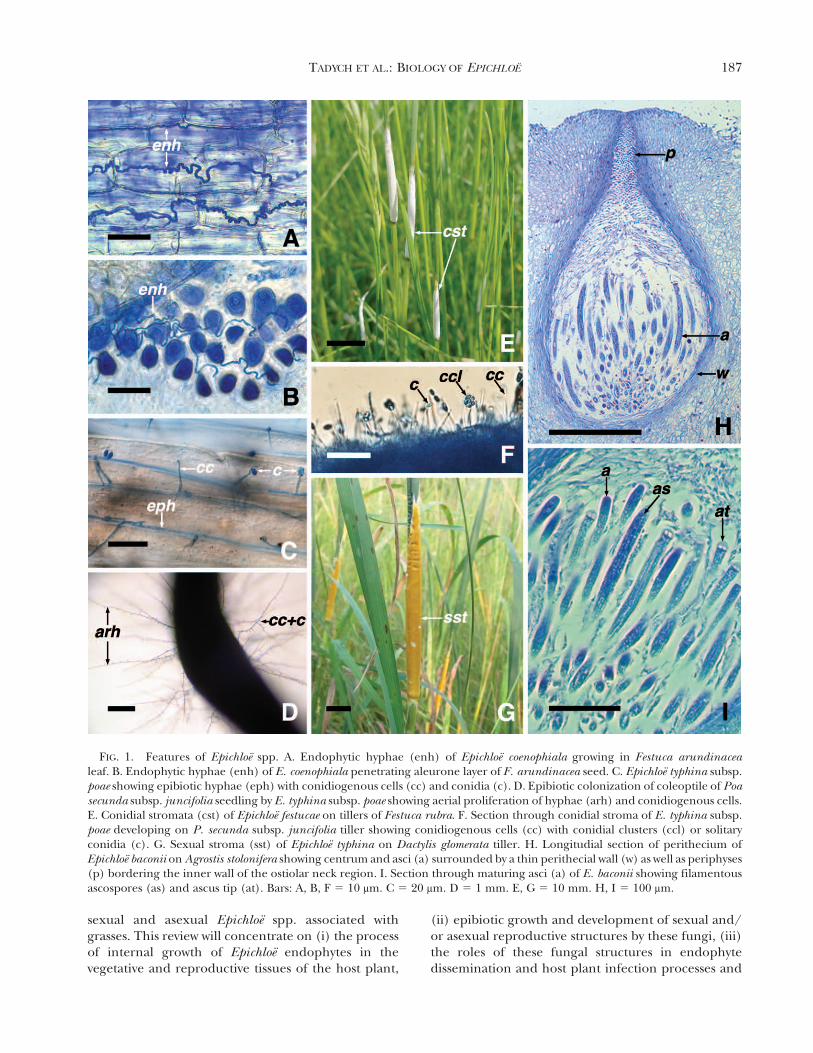
FIG. 1. Features of Epichloe spp. A. Endophytic hyphae (enh) of Epichloe coenophiala growing in Festuca arundinacealeaf. B. Endophytic hyphae (enh) of E. coenophiala penetrating aleurone layer of F. arundinacea seed. C. Epichloe typhina subsp.poae showing epibiotic hyphae (eph) with conidiogenous cells (cc) and conidia (c). D. Epibiotic colonization of coleoptile of Poasecunda subsp. juncifolia seedling by E. typhina subsp. poae showing aerial proliferation of hyphae (arh) and conidiogenous cells.E. Conidial stromata (cst) of Epichloe festucae on tillers of Festuca rubra. F. Section through conidial stroma of E. typhina subsp.poae developing on P. secunda subsp. juncifolia tiller showing conidiogenous cells (cc) with conidial clusters (ccl) or solitaryconidia (c). G. Sexual stroma (sst) of Epichloe typhina on Dactylis glomerata tiller. H. Longitudial section of perithecium ofEpichloe baconii on Agrostis stolonifera showing centrum and asci (a) surrounded by a thin perithecial wall (w) as well as periphyses(p) bordering the inner wall of the ostiolar neck region. I. Section through maturing asci (a) of E. baconii showing filamentousascospores (as) and ascus tip (at). Bars: A, B, F 5 10 mm. C 5 20 mm. D 5 1 mm. E, G 5 10 mm. H, I 5 100 mm.
TADYCH ET AL.: BIOLOGY OF EPICHLOE 187

(iv) some ecological and evolutionary implications ofthe modes of dissemination.
COLONIZATION OF THE HOST PLANT
Endophytic growth.—The internal systemic associationof Epichloe endophytes with grass hosts is formedduring development of the host plant. The endo-phytes proliferate in intercellular spaces (FIG. 1A)and do not form haustoria. Available data suggest thatthey use nutrients from apoplastic fluids as well asfrom adjacent host cells through production of anumber of hydrolytic enzymes (Lindstrom et al. 1993,Tan et al. 2001, Christensen et al. 2002, Moy et al.2002, Li et al. 2005, Bryant et al. 2007). Theendophyte-host association requires close coordina-tion of life cycles of both partners (Herd et al. 1997).
During the vegetative growth of the host grassbeginning with seed germination, the endophytichyphae asymptomatically colonize the developingshoot apex of the seedling (Clay 1990, Christensenet al. 2002). Further hyphal growth through tissues ofthe shoot apical meristem, leaf primordia, sheathsand blades of the leaves results in systemic coloniza-tion of aerial parts of the seedling (FIG. 1A). Inmeristematic tissues hyphal growth is characterized byhyphal tip growth and branching. In contrast, theintercellular hyphae that reside in the apoplast ofgrowing seedlings form extended longitudinal andusually convoluted strands with little branching. Asthe grass cells expand, mycelium within the culm andleaf tissues extend through intercalary extensionrather than tip growth (Christensen et al. 1997,2000, 2008; Herd et al. 1997; Tan et al. 2001). Thismay be the result of shear force on hyphae due to theelongating plant parenchyma cells. This pattern ofgrowth results in synchronization of hyphal growthand plant cell growth. During the development ofthe host inflorescence, endophytic hyphae proliferatefrom the vegetative apex, penetrate into the inflores-cence primordium and floral apex to colonize theovary and ovules. Following fertilization, hyphae enterthe embryo sac and can be found early on the surfaceof the developing embryo. During maturation of seed,hyphae can be found concentrated throughout theembryo and surrounding tissues including plumuleapex, aleurone layer (FIG. 1B) and scutellum of thegrass seed (Freeman 1904, Philipson and Christey1986, White et al. 1991, May et al. 2008); the presenceof the endophyte in grass seeds is not detrimental (i.e.does not affect seed viability) (Siegel et al. 1984).After the seeds are released from the matureinflorescence and germinate, the resulting seedlingscontain the fungal symbiont (Siegel et al. 1984), thusenabling the endophyte to transmit vertically from
maternal plant to the next host generation (Freeman1904, Clay 1990, Afkhami and Rudgers 2008).
Epibiotic growth.—Growth of most Epichloe endo-phytes, regardless of their ability to reproducesexually or asexually, is usually not limited to internaltissues of the host plant. Some Epichloe species arecapable of growing superficially and producingepibiotic reproductive structures (i.e. mycelial net-works with conidiogenous cells and conidia onaboveground parts, mostly on leaf blades) (FIG. 1C–D).These epibiotic structures have been documented ondiverse grass species, including A. hyemalis, Bro.auleticus, Bro. setifolius, D. glomerata, Ech. ovatus,Festuca arvernensis Auquier, Kerguelen & Markgr.-Dann. (synonym: Festuca ovina var. glauca [P. Beauv.]W.D.J. Koch), F. pratensis, Festuca rubra L. (synonym:Festuca rubra var. commutata Gaudin, Festuca ovina var.rubra [L.] Sm.), Hel. europaeus, H. brevisubulatum,Lolium 3 hybridum Hausskn., L. perenne, P. rigidifolia,P. bonariensis (Lam.) Kunth, P. secunda subsp.juncifolia and Poa sp. (White et al. 1996, Christensenet al. 1997, Moy et al. 2000, Craven et al. 2001, Dugan etal. 2002, Moon et al. 2002, Tadych et al. 2007, Iannoneet al. 2009, Oberhofer 2012, Alderman 2013). Theepibiotic hyphae are usually aligned with the longitu-dinal junctions between epidermal cell walls andoccasionally branching, sometimes interconnectedand overlapping (FIG. 1C). The epibiotic conidia arehyaline, aseptate, reniform to ellipsoid, water-transmit-ted and capable of germinating (Tadych et al. 2007,2012).
Stroma formation.—When a host plant colonized byan Epichloe endophyte undergoes reproductive devel-opment (TABLE I), the fungus in many cases has thepotential to form a compact mycelial structure calleda stroma (FIG. 1E–G). Formation of stromata byendophytes results from unrestricted hyphal growthwithin and outside the developing inflorescence,which envelops an emerging inflorescence and theupper leaf sheath of reproductive tillers (White et al.1991, White 1997). The stromata usually preventflower and seed development on affected culms,leading to partial or total sterility of the host plant(Bradshaw 1959). Stromata of Epichloe spp. arestructures where mating takes place but also whereasexual (FIG. 1E–F) and sexual reproductive propa-gules important for fungal dispersal develop(FIG. 1G–I). As the stroma develops, numerouselongated conidiogenous cells arise on a surface(FIG. 1F), which constitute the anamorph (conidialstroma; FIG. 1E). Some Epichloe endophytes, althoughnot known to complete a sexual cycle, may produceconidial stromata on flowering culms (Sampson 1933,White et al. 1992, Ji et al. 2009, Christensen et al.
188 MYCOLOGIA

2012, Oberhofer and Leuchtmann 2012, Tadych et al.2012). The conidial stroma (FIG. 1E) is light, usuallywhite to gray, covered with a dense layer ofunbranched phialidic conidiogenous cells, wherehyaline, one-celled, typically reniform to ellipsoid,conidia are borne (FIG. 1F). These conidia mayfunction as spermatia (Leyronas and Raynal 2008a)(i.e. gametes in a mating process if the fungusundergoes heterotallic sexual reproduction). Epichloestromata are hermaphroditic; both receptive hyphaeand spermatia are formed on the same stroma (White1997). As a consequence of fertilization, the stromathickens, usually become brightly colored, yellow ororange (FIG. 1G) and superficial perithecia (FIG. 1H)develop, containing asci with ascospores (FIG. 1I)
SEXUAL DEVELOPMENT AND REPRODUCTION
Because Epichloe fungi are heterothallic, successfulcompletion of sexual reproduction relies on crossfertilization between two different fungal matingtypes of the species (White and Bultman 1987). Thisresults in the transfer of gametes from a stroma of onemating type to a stroma of the opposite mating type(Raynal 1991b, White 1997, Bultman et al. 1998,Webster and Weber 2007). Spermatia of Epichloespecies are transferred to ascogonia in perithecialprimordia by specialized flies in the genus Botano-phila Lioy (Diptera, Anthomyiidae), which feed anddevelop on Epichloe stromata (Kohlmeyer and Kohl-meyer 1974; Bultman and White 1988; Bultman et al.1995, 1998). Although this pollination-like associationis probably not highly specific, a study of Epichloe-associated Botanophila flies revealed that in Europeand North America at least six distinct fly taxa areinvolved in gamete transfer (Leuchtmann 2007,Lembicz et al. 2013). Specific volatile compounds,that is sesquiterpenoid alcohol (chokol K), methylester (methyl (Z )-3-methyldodec-2-enoate 5 MME)and perhaps other compounds (e.g. diterpenes,trimethylester 5 methyl 2,4,8-trimethylundecanoate5 MTE) emitted by the fungal stroma attract flies tostroma (Schiestl et al. 2006; Steinebrunner et al.2008a, b, 2008c). During their visit, female flies feedon fungal tissues including conidia that eventuallypass through the digestive system (Bultman andLeuchtmann 2003). Feces that are actively depositedon the stromata by the fly during feeding andoviposition contain intact and viable conidia. Theseconidia function as spermatia and are capable ofcross-fertilization of the fungus. Eventually fly larvaehatch and feed on the stroma tissues, includingperithecia. They then drop and pupate in the soil(Bultman et al. 1998). These fungal stromata are thesole source of nourishment for the Botanophila larvae.
In situations of high egg load and an excessive larvalexploitation of stromata, greater larval mortality hasbeen observed, suggesting the existence of somedegree of mutual dependence among both partners,the fly and the fungus (Bultman et al. 2000).
Some field observations indicate, however, thatthere is no correlation between fly abundance andstroma fertilization (Rao and Baumann 2004; Gor-zynska et al. 2010, 2011; Rao et al. 2012). These studiesnoted that in some grass populations no flies, fly eggsor larvae on stromata were detected but fertilizedstromata were reported universally, with peritheciapresent on 85–100% of examined stromata. However,the absence of eggs or larvae of Botanophila onstromata is not necessarily evidence that the flies orother vectors did not visit the stromata. Becausestromata provide food for the Botanophila females,flies could have visited without laying eggs, anddeposited spermatia during such visits which in turnfertilized the stromata (Parker and Bultman 1991). Forinstance, it has been shown that spermatia may adhereto insect body hairs (Bultman et al. 1995).
It has been suggested that in grass populationswhere no flies were observed, another mechanism(i.e. ascospore fertilization) may be responsible forstroma fertilization (Alderman and Rao 2008, Raoet al. 2012). These authors proposed that at thebeginning of growing season the early emergingstromata of E. typhina developing on D. glomerataplants first are fertilized by flies and later thesestromata produced ascospores. They hypothesizedthat ascospores were disseminated by wind and servedas gametes for subsequent fertilization of developingstromata among neighboring fields. Furthermore,Rao et al. (2010, 2012) reported that the gray gardenslug (Deroceras reticulatum Muller; Agriolimacidae,Gastropoda, Mollusca) and reticulate taildropper(Prophysaon andersoni Cooper; Anadenidae, Gastro-poda, Mollusca) forage on D. glomerata plantsinfected by stroma-producing E. typhina and thatthe fungal spermatia are transmitted through the gutof a slug; the authors suggested that this may result incross fertilization of the fungal stromata. This modeof spermatia transport might be limited; the averagebiomass consumed by grove snails (Cepaea nemoralisL.; Helicidae, Gastropoda, Mollusca) feeding on endo-phyte-infected Pu. distans grass seedlings was less thanthat consumed on uninfected ones (Czarnoleski et al.2010).
Abiotic conditions may also play a role in fertiliza-tion. If a stroma of one mating type is sufficientlyclose to a stroma of the opposite mating type, watersplash may be responsible for transfer of spermatiabetween stromata and their fertilization during rainyand windy conditions (Tadych et al. 2007, 2012; Rao
TADYCH ET AL.: BIOLOGY OF EPICHLOE 189

et al. 2012). Therefore, this mechanism could workefficiently, especially in dense populations of stromataand under moist environmental conditions (Rao andBaumann 2004; Gorzynska et al. 2010, 2011; Rao et al.2012). Further studies are required to determine howgametes in Epichloe spp., in both agricultural andnatural grass populations, are transferred.
FACTORS AFFECTING DEVELOPMENT OF FUNGAL
REPRODUCTIVE STRUCTURES
In natural grass populations, sexual reproduction of ahost is rarely completely eliminated by developmentof stromata, even at high colonization frequencies. Inmany species of Epichloe, the abundance of stromatavaries among infected plant species as well as amongindividual infected plants or their tillers (White andChambless 1991; Bucheli and Leuchtmann 1996;Kover and Clay 1998; Groppe et al. 1999; Meijer andLeuchtmann 1999, 2000; Yanagida et al. 2005;Cheplick and Faeth 2009; Lembicz and Olejniczak2009; Oberhofer and Leuchtmann 2012; Tadych et al.2012) and is regulated by specific host genotype-endophyte co-evolution and environmental factors.
Fungal factors.—Population-structure studies of E.sylvatica associated with Brp. sylvaticum revealed thatstromatal development is controlled by the fungalgenotype (Bucheli and Leuchtmann 1996; Meijer andLeuchtmann 1999, 2001). In E. sylvatica, both stroma-forming strains and non-stroma-forming strains donot belong to a single random mating population butare differentiated genetically into separate subpopu-lations despite their occasional co-occurrence onindividual host plants. Epichloe typhina subsp. poaeas revealed by morphology and phylogenetic analysisis associated with four species of grasses (i.e. P.nemoralis, P. pratensis, P. secunda subsp. juncifolia, P.sylvestris). Nevertheless, this endophyte shows allpossible variations in reproductive modes dependingon the host species (Tadych et al. 2012). In Poapratensis the fungus forms stromata but is not seed-transmitted. In P. nemoralis and P. secunda subsp.juncifolia the fungus produces stromata but it alsoenters seeds. In P. sylvestris, fungal stromata have notbeen reported but the fungus transmits throughseeds. Perithecia with viable ascospores have beenobserved only rarely on stromata of P. nemoralis andP. pratensis (A. Leuchtmann pers comm), whileperithecia with ascospores have not been reportedon P. secunda subsp. juncifolia. The most vigorousfungal endophytes may affect plant growth andreproduction as well as the number of stromataproduced (Groppe et al. 1999). Based on the quantityof Epichloe DNA in vegetative plant tissue, Groppe
et al. (1999) found that plants of Bro. erectus infectedby E. bromicola with the highest concentration of thefungus in the vegetative plant tissues had the highestnumber of stromata (3.4 per plant) with no inflores-cences whereas infected plants with the lowestconcentration of the fungus in the vegetative planttissues had the fewest stromata (0.5 per plant) and themost fertile inflorescences (0.3 per plant). However,this value was much lower than values found foruninfected plants (1.1 inflorescences per plant).
Host factors.—Groppe et al. (1999) found that thenumber of stromata produced by E. bromicola in Bro.erectus plant communities was greater when higherplant species diversity was observed. In another studyon E. bromicola infecting three different Bromus spp.,Brem and Leuchtmann (2003) found evidence thatthe 26 E. bromicola isolates from Bromus hostscollected from natural populations in Switzerlandand France were genetically differentiated accordingof their hosts. Although different host strains wereinterfertile, the strains differed in their reproductivemodes (Schardl and Leuchtmann 2005). The E.bromicola strain naturally infecting Bro. erectus wasalways stroma-forming and failed to transmit in hostseeds, whereas its strains from Bro. benekenii and Bro.ramosus did not form stromata and were seedtransmitted. Furthermore, reciprocal inoculations ofhost plant seedlings showed that asexual strains fromBro. benekenii and Bro. ramosus were incapable ofinfecting Bro. erectus whereas the stroma-formingisolates from Bro. erectus were broadly compatiblewith all three host species. Brem and Leuchtmann(2003) proposed that asexual forms arose from withinsexual populations on Bro. erectus after host shifts toBro. benekenii and Bro. ramosus. In another studyLembicz et al. (2010) found that the sexual form of E.bromicola also occurs on El. repens in Poland,apparently the result of a host shift where thereproductive mode remained constant.
A correlation between plant genotype and stromaexpression also has been found in studies on Brp.sylvaticum infected by E. sylvatica (Meijer andLeuchtmann 2000) and on H. patula infected by E.elymi. (Tintjer et al. 2008). Meijer and Leuchtmann(2000) investigated stroma expression in nine geno-types of Brp. sylvaticum and found that in the firstyear 74.1% of the total variance in stroma productionwas explained by host genotype while this variancecomponent declined to 27.5% in the second year ofobservation. Another grass endophyte, E. festucae,forms stromata on F. rubra (Leuchtmann et al.1994). However, separate field surveys of populationsof this endophyte associated with F. rubra and Festucaovina L. in grasslands of the northern Europe,
190 MYCOLOGIA

European subarctic and the European Mediterraneansemi-desert did not yield stromata with fruiting bodyformation but instead revealed predominantly non-recombining asexual or clonal population structures(Arroyo Garcıa et al. 2002, Bazely et al. 2007, Wali et al.2007). These observations suggest that under condu-cive conditions entire populations of Epichloe spp. canthrive as asymptomatic endophytes perhaps dependingentirely on their asexual reproduction via their hostseeds or on production of epibiotic conidia fordispersal.
In populations of Pu. distans, a perennial Euro-Siberian halophyte grass (Hughes and Hallidays1980), the infection by E. typhina seems to appearonly in plants in anthropogenic habitats (Lembicz1998; Lembicz et al. 2009, 2011). The absence of E.typhina colonization in natural populations of thehost, as suggested by Lembicz et al. (2011), was aresult of widespread distribution of host genotypeswhich are incompatible with E. typhina; on the otherhand, if latently infected, the genotype preventeddevelopment of stromata (Meijer and Leuchtmann2000, Lembicz et al. 2009). In the host genotypeinfected by E. typhina, the number of tillers bearingstromata increased significantly with age of the hostplant (Lembicz et al. 2011). To counteract thenegative influence of choke disease the Pu. distansgrass populations in anthropogenic habitats andinfected by E. typhina produced additional tillerswith stromata in late summer (Lembicz et al. 2009,2011). These tillers did not appear in grass popula-tions of natural saline habitats (Lembicz 1998). Thefungus on these late summer tillers initially developedstromata at a lower frequency, but their numberincreased significantly with the age of the plants(Lembicz et al. 2009, 2011). These late summerstromata were always white and did not produceperithecia with asci and ascospores. The researchersspeculated that the lack of perithecia might be due tothe absence of the vector, the adult Botanophilaflies, in late summer (Bultman et al. 1998, but seeGorzynska et al. 2011). In addition, the authorshypothesized that the production of fungal reproduc-tive structures in late summer might not provide anybenefits to the fungus, E. typhina, or to the plant, Pu.distans (Lembicz et al. 2009, 2011). However, latesummer stromata give rise to abundant conidialstromata and may support local dissemination of thefungus and possibly be responsible for maintenanceof new infections in germinating seeds and develop-ing seedlings of the host grass in late summer or earlyautumn (Tadych et al. 2012). As observed by otherresearchers ( Jackowiak 1984, 1996; Jackowiak et al.1991; Moravcova and Frantık 2002), the new seedlingsof Pu. distans can appear in the field throughout the
region under favorable humid and temperatureconditions in September. High humidity, fluctuatingtemperature of 15/6 C and lower constant tempera-ture 15 C (Moravcova and Frantık 2002) optimal forPu. distans seed germination, are aligned withoptimal infection conditions for Epichloe spp. ( Ju etal. 2006). Clearly the relationship of E. typhina withPu. distans requires further study.
Environmental and geographical factors.—Experimen-tal data suggest that environmental factors may notonly influence the distribution and occurrence ofEpichloe endophytes (Novas et al. 2007, Iannone et al.2013) but also the expression of their reproductivestructures. Groppe et al. (2001) showed that stromaformation increased in E. bromicola infecting Bro. erectusfollowing experimental fragmentation of the hostpopulation. During a 2 y study in natural grasspopulations, the proportion of El. hystrix plants bearingE. elymi stromata was significantly different betweenyears, populations and demonstrated a year by popula-tion interaction, suggesting that stroma expression canvary with environmental conditions (Tintjer et al. 2008).The survey on the distribution of E. typhina in naturalpopulations of C. purpurea in Sweden revealed that thefungal stromata in the field were present only innutrient-rich and wet habitats and totally absent fromnutrient-poor and dry habitats (Wennstrom 1996).However, Sun et al. (1990) reported that the additionof nitrogen fertilizer had a negative effect in theproduction of stromata by E. festucae in infected F.rubra plants. In contrast, observations on Brp. sylvaticumcolonized by E. sylvatica revealed that fertilized plantson average showed slightly higher stroma expressionthan unfertilized plants (Meijer and Leuchtmann2000); less consistent increases in stroma expressionin fertilized plants were reported when the infectedplants were exposed to elevated concentration of CO2.On the other hand, Groppe et al. (1999) found elevatedatmospheric CO2 enhanced the production of fungalstromata in E. bromicola-infected Bro. erectus plants butinhibited inflorescence production in uninfectedplants. Lembicz et al. (2011) proposed that higherconcentrations of CO2 that occurred in anthropogenichabitats occupied by Pu. distans grass colonized by E.typhina might be responsible for higher stromaproduction by the fungus.
Kirby (1961) suggested that other factors thatstimulate plant growth might reduce stroma formationbecause rapidly growing flowering culms can escapefungal infection. In contrast, White and Chambless(1991) and White et al. (1991) found that growth offungal isolates on certain sugars in vitro were highlycorrelated with the number of stromata produced onhost plants. A regulatory mechanism involving sugars
TADYCH ET AL.: BIOLOGY OF EPICHLOE 191

predicts that factors that alter the relative sugarconcentrations in the meristems also might alterstroma expression (White et al. 1991). Otherenvironmental factors also may influence stromaexpression (e.g. the effect of shading). Meijer andLeuchtmann (2000) observed a moderate increase ofstroma production in E. sylvatica-infected Brp.sylvaticum in a 2 y study. The authors proposed thatsuccessful horizontal transmission of stroma-formingstrains in shaded environment had contributed toincreased stroma expression in the shade. Undercontrolled conditions, Alderman (2013) has shownthat stroma formation in E. typhina infected D.glomerata plants could be induced by vernalization ofplants at 8 C and 8 h light for 16 wk, following byplacing the plants in a growth chamber at 15 C and16 h light 2 wk before transferring the plants to agreenhouse, where stromata usually appeared withinseveral weeks.
The distribution of stromata of Epichloe spp. oftendoes not follow the distribution of their mostcommon host grasses. In Fennoscandia, Epichloespecies are widely associated with more than 25 grassspecies (Vare and Itamies 1995, Saikkonen et al. 2000,Bazely et al. 2007, Granath et al. 2007, Wali et al.2007). However, pronounced geographical variationand uneven expression of stromata on a specific grasshost species have been observed. For example, thestroma-producing E. typhina colonizing D. glomeratais common in southern Finland, Norway and Swedenbut rare in north Fennoscandia, although the grasshost occurs in pastures and meadows throughout theregion. A similar pattern was found for Ph. pretenseand several other grass genera widely distributed inFennoscandia, including the northernmost part ofthe region, and colonized by Epichloe spp. (Eriksson1967, Koponen and Makela 1976, Eckblad andTorkelsen 1989, Vare and Itamies 1995). At higherlatitudes of Fennoscandia (65uN and higher) stroma-ta were reported solely on species of Calamagrostisonly, including Calamagrostis lapponica (Wahlenb.)Hartm, C. purpurea and Calamagrostis stricta (Timm)Koeler (Eriksson 1967, Koponen and Makela 1976,Eckblad and Torkelsen 1989). Farther south thestromata were found only on Calamagrostis canescens(G.H. Weber) Roth (60uN) and C. epigeios (64uN) insouthern Finland (Koponen and Makela 1976). Insouthern Norway the white conidial stroma predom-inate in May, June and July but in the north also canbe found as late as mid-August (Eckblad andTorkelsen 1989). The orange perithecia stromatamay appear as early as the end of May and as late asthe beginning of October. Peak formation of peri-thecia is in the first two weeks of July. Thus theasexual and the sexual forms of stromata may occur
simultaneously, even on different culms of the sameplant. However, the teleomorph in the north is rare,perhaps because the development of the fungus isslow as a result of the late spring (Eckblad andTorkelsen 1989). Variations in stroma developmenton grasses in different regions may be a complexinteraction between fungal and plant genetics andenvironmental factors such as soil nutrients, water,temperature and day length.
DISSEMINATION OF FUNGAL PROPAGULES
AND INFECTION OF GRASS HOSTS
The mode of dissemination of Epichloe symbiontsvaries depending on host plant, endophyte popula-tion and endophyte species (White 1988, White andChambless 1991, Brem and Leuchtmann 2003,Leuchtmann 2003, Schardl and Moon 2003, Rodri-guez et al. 2009, Oberhofer and Leuchtmann 2012,Tadych et al. 2012). The vertically transmittedsymbiont is disseminated through the maternal germline, while the horizontally transmitted symbiont istaken up from the environment anew by each hostgeneration (Rodriguez et al. 2009, Bright andBulgheresi 2010). Until recently it was believed thatin asexual lineages of Epichloe endophytes the fungigrow systemically throughout most of the above-ground parts of the host plant, are completelyasymptomatic and do not form any external struc-tures for horizontal dissemination (Sampson 1933,Clay 1988, Schardl et al. 2004, Schardl 2010). It wasthought that dissemination between host generationsis achieved by vegetative growth of mycelium into theseed embryo (Sampson 1933, Siegel et al. 1984, Clay1990, Clay and Schardl 2002, Cheplick and Faeth2009, Schardl 2010). However, some of these asexualendophytes also are able to produce external fungalstructures, such as epibiotic mycelial networks withconidiogenous cells and conidia, growing mostly onleaf blades of the host plants (White et al. 1996,Christensen et al. 1997, Moy et al. 2000, Craven et al.2001, Dugan et al. 2002, Moon et al. 2002, Tadychet al. 2007, Iannone et al. 2009, Oberhofer 2012,Alderman 2013) and conidiogenous cells with conidiadeveloping on partial/full asexual stromata (Whiteet al. 1991, Ji et al. 2009, Christensen et al. 2012,Oberhofer and Leuchtmann 2012, Tadych et al. 2012).The presence of epiphytic fungal structures bearingconidia suggests that some of these endophytes alsomight have the ability for plant-to-plant spread viahorizontal transmission of the conidia; thus theirdependence on the host-mediated vertical transmis-sion may not be absolute. More recently it has beendemonstrated that conidia of E. typhina subsp. poae,frequently formed on the surface of leaves of P.
192 MYCOLOGIA

secunda subsp. juncifolia or on the conidial stromataon this host, are spread by water splash and areresponsible for infection of germinating seeds anddeveloping seedlings. As a consequence, such infec-tions cause systemic colonization of the host plant bythis endophyte (Tadych et al. 2007, 2012).
As referenced earlier, sexual Epichloe species onundeveloped stromata produce conidia but afterfertilization perithecia with viable ascospores develop.Mature ascospores are forcibly ejected from perithe-cia and are transmitted by wind (White 1997).Ascospore dispersal predominantly occurs at nightwith high relative humidity and plant water pressure(Ingold 1948, Raynal 1991a, Welch and Bultman1993, White et al. 1993, White and Camp 1995,Webster and Weber 2007). It has been hypothesizedthat ascospores mediate horizontal transmission ofEpichloe endophytes (Chung and Schardl 1997; Bremand Leuchtmann 1999; Leuchtmann 2003; Leyronasand Raynal 2008a, b; Schardl et al. 2009; Schardl2010), but it has not been confirmed whetherinfection occurs directly from ascospores or byinfective primary or secondary conidia formed afteriterative cycles of conidiation of ascospores (Baconand Hinton 1988, 1991; Leuchtmann 2003). There-fore, the exact mechanisms of contagious infection byascospores are still not fully understood (Leucht-mann 2003, Webster and Weber 2007). Chung andSchardl (1997) proposed that the mechanism ofinfection of grasses by ascospores is invasion of theovule via stigma colonization, a mechanism similar tothat demonstrated for Claviceps purpurea (Fr.) Tul.Western and Cavett (1959) proposed another mech-anism, where ascospores or conidia may infectwounds. In experiments to demonstrate infectionChung and Schardl (1997) and Western and Cavett(1959) found infection rates of 1.3% and 5.4%
respectively. However, inconsistencies in reproducingthe results obtained in these experiments leaves somedoubt about the actual mechanism of infection as wellas site of infection (Brem 2001, Leyronas and Raynal2008b).
In contrast, exposure of D. glomerata seeds tofertilized stroma of E. typhina resulted in an infectionrate of 25% (Leyronas and Raynal 2008b). Theresearchers hypothesized that germinating ascosporesor conidia produced by ascospores might be respon-sible for the infection. In another study, although therate of new infections was relatively low, 5.3% ofyoung seedlings of Brp. sylvaticum, growing in Petridishes and exposed to ascospores-ejecting stromata ofE. sylvatica, became infected (Brem 2001). Similarly,under natural conditions Brem and Leuchtmann(1999), using stroma-producing E. sylvatica coloniz-ing Brp. sylvaticum, demonstrated contagious spread
of the endophytic fungus to mature uninfected Brp.sylvaticum plants growing nearby infected plants. Thenumber of new infections observed in that study waspositively correlated with the amount of stromataobserved within each local population, an infectionrate of 34% at one site and 17% at another site wasreported (Brem and Leuchtmann 1999). Theseresults also were reproduced in experimental plots,in which Brp. sylvaticum plants infected by stroma-producing E. sylvatica served as the inoculum sourceand surrounded by mature uninfected plants; thefrequency of new infection of these previouslyuninfected plants was 16–25% (Brem 2001). Theseauthors concluded that infection of vegetative tillersof the grass host was frequent and hypothesized thatascospores invade the meristematic zone of tillerbuds.
Tadych et al. (2012) revealed that germinatingseeds and seedlings of P. secunda subsp. juncifoliabecome infected when exposed to E. typhina subsp.poae conidia and mycelium produced by theseconidia. In this study 36.4% and 33.7% of plantsharbored the endophyte after exposure to conidiaoriginating respectively from leaves or stromatae.These results suggest that conidia may be responsiblefor horizontal dissemination of Epichloe fungi andthat conidia or mycelium derived from conidia aredirectly responsible for infections of host grass.Ascospores also are responsible for horizontal dis-semination. The water-disseminated conidia may beresponsible for local distribution, whereas the wind-disseminated ascospores may function to vector thefungus longer distances. However, stroma productionalso may enhance horizontal dissemination of conidiaover large distances by facilitating insect vectoring(Bultman et al. 1995, 1998). Flying insect vectorscould transport conidia long distances and initiateinfections via tillers or seedling infections.
In a mixed mode of reproduction that is wide-spread in the northern hemisphere (e.g. TABLE I) theinfected plant forms both stroma-bearing tillers andnormal tillers with fertile inflorescences (White 1988,Schardl 2010); the inflorescences produce fullydeveloped and viable seeds that are asymptomaticallycolonized by fungal hyphae that enable verticaltransmission of the fungal endophyte by the seeds.In this mixed reproduction strategy, the fungus oninfected plants may form either sexual stromata withascospores or partial/full asexual stromata, togetherwith epibiotic mycelial networks bearing conidia; thisstrategy may result in dissemination of the fungus andultimately colonization of new plants. Such complexand diverse modes of reproduction in Epichloe spp.may effectively support wide distribution of distinctEpichloe-grass associations in various ecosystems.
TADYCH ET AL.: BIOLOGY OF EPICHLOE 193

EPIBIOTIC CONIDIA—ROLES IN HYBRIDIZATION
Horizontal dissemination involving epibiotic conidiamay be responsible for widespread associations ofasexual Epichloe endophytes with grasses, especially inthe southern hemisphere and northernmost subarcticregions of the northern hemisphere (e.g. Finland,Norway, Sweden). In these regions asymptomaticEpichloe endophytes showing epibiotic conidial stagesare prevalent, and in the subarctic regions of thenorthern hemisphere stromata stages, with or withoutperithecia, have been found occasionally on someendophytically infected grasses (Koponen and Makela1976; Eckblad and Torkelsen 1989; Bazely et al. 2007;Wali et al. 2007; Saikkonen et al. 2010; Iannone et al.2011, 2012a, b). In South America, studies of field andherbarium grass specimens revealed that almost 40 outof more that 50 grass species in the genera Briza,Bromus, Calamagrostis, Festuca, Hordeum, Melica,Phleum and Poa were infected by Epichloe endophytes(Iannone et al. 2011, 2012a, b, c, 2013; Mc Cargo et al.2014). Even though these grasses inhabit a wide rangeof environments, from tropical through subtropicalrainforests to cold deserts in southern Patagonia, aswell as altitudes of up to 3000 m, none of the infectedgrasses exhibited stroma development (Iannone et al.2012a). Similarly, during intensive field studies inFinland, Norway and Sweden, including the northern-most subarctic regions of the countries, stromata of E.festucae also were never found on F. rubra and F. ovina(Bazely et al. 2007, Wali et al. 2007). Because conidiaare produced epibiotically, sexual stromata may not beessential for horizontal transmission of the fungus to anew host. Host jumps have been proposed recently tooccur in some Epichloe-grass associations (Gentile et al.2005, Oberhofer and Leuchtmann 2012, Tadych et al.2012). Host jumps may be linked with the capabilityfor horizontal dissemination by ascospores (Craven2012). However, in many other cases host jumps maybe due to dissemination and infectivity of conidiarather than ascospores.
Forty-three Epichloe taxa endophytically associatedwith grasses have been described (TABLE I); 14 areknown to develop sexual structures with viableascospores, while in the remaining 29 taxa the sexualstate has not been observed (Leuchtmann et al. 2014).More than half (51.2%) of all Epichloe species arehybrid (Moon et al. 2004, Leuchtmann et al. 2014);they are prevalent in certain grass populations(Leuchtmann 2003, Gentile et al. 2005, Iannone etal. 2012a, Oberhofer and Leuchtmann 2013). Multiplecopies of the translation elongation factor 1-a (tefA), b-tubulin (tubB) or other genes that originated frominterspecific hybridization can be isolated from thesehybrid species. Because the versions of these sequences
reflect the lineage of the progenitor species, phyloge-netic analyses of these sequences into distinct cladeshave been used to infer the parental Epichloe species.Interspecific hybridization has been proposed toexplain the loss of the sexual stage in Epichloe speciesknown to be hybrid (Kuldau et al. 1999, Gentile et al.2005). There is one known exception; E. liyangensis, asexually reproducing species from China, is a hybrid(Kang et al. 2011). Hybridization may be a mechanismto circumvent the lack of recombination in asexualstrains by the acquisition of an additional genome.This also may enable the hybrids to explore a broaderrange of ecological niches compared to non-hybrids(Brasier 2000, Schardl and Craven 2003, Giraud et al.2008, Hamilton et al. 2010, Oberhofer and Leucht-mann 2012). How and where hybridization betweenEpichloe species occurs is not yet known, but parasexualreproduction by somatic vegetative fusion (anastomo-sis) of fungal hyphae from Epichloe species has beenproposed (Schardl et al. 1994, 1997; Tsai et al. 1994;Schardl and Craven 2003; Moon et al. 2004, 2007;Schardl 2010). The origin of hybrids from two or moreEpichloe genomes clearly indicates that host jumps arethe likely basis for hybridization (Moon et al. 2004,Gentile et al. 2005, Oberhofer and Leuchtmann 2012,Tadych et al. 2012).
With the discovery that conidia may infect seedlingsand thus may set up primary infections in grasses, it isclear that biologists must begin to examine theoccurrence and frequency of horizontal transmissionof Epichloe spp. There remains a number of questionsthat need to be addressed including: (i) do ascosporesform hyphae directly or do they produce conidia thatare infective, (ii) what range of distances can conidia orascospores travel, (iii) do insects vector conidia thatmay set up infections, (iv) are multiple vectors involvedin the fertilization of stromata, (v) do environmentalfactors such as day length affect stroma development,(vi) could this account for regional differences instromata development, (vii) are ascospores or conidiainvolved in hybridization and (viii) does hybridizationoccur within plants or outside plants? Not only dovarious aspects of the ecology of fungal endophytes ingrasses need to be systematically examined, but there isenormous complexity that will require elucidation bybiochemical and molecular techniques. Clearly, fur-ther unraveling of the strands of this complexity will bea challenge for all investigators conducting research inEpichloe -Pooideae symbioses.
ACKNOWLEDGMENTS
Our research on grass endophytes was supported in part bythe New Jersey Agricultural Experiment Station and theRutgers Turfgrass Science Center.
194 MYCOLOGIA

LITERATURE CITED
Afkhami ME, Rudgers JA. 2008. Symbiosis lost: imperfectvertical transmission of fungal endophytes in grasses.Am Nat 172:405–416, doi:10.1086/589893
Ahlholm JU, Helander M, Lehtimaki S, Wali P, Saikkonen K.2002. Vertically transmitted fungal endophytes: differentresponses of host-parasite systems to environmentalconditions. Oikos 99:173–183, doi:10.1034/j.1600-0706.2002.990118.x
Alderman SC. 2013. Survival, germination and growth ofEpichloe typhina and significance of leaf wounds andinsects in infection of orchardgrass. Plant Dis 97:323–328, doi:10.1094/PDIS-01-12-0075-RE
———, Rao S. 2008. Ascosporic fertilization of Epichloetyphina in Dactylis glomerata seed production fields inOregon and implications for choke management. PlantHealth Prog, doi:10. 1094/PHP-2008-0421-01-BR
Arechevaleta M, Bacon CW, Hoveland CS, Radcliffe DE.1989. Effect of the tall fescue endophyte on plantresponse to environmental stress. Agron J 81:83–90,doi:10.2134/agronj1989.00021962008100010015x
Arroyo Garcıa R, Martınez Zapater JM, Garcıa Criado B,Zabalgogeazcoa I. 2002. Genetic structure of naturalpopulations of the grass endophyte Epichloe festucae insemiarid grasslands. Mol Ecol 11:355–364, doi:10.1046/j.0962-1083.2001.01456.x
Bacon CW, Hinton DM. 1988. Ascosporic iterative germi-nation in Epichloe typhina. Trans Br Mycol Soc 90:563–569, doi:10.1016/S0007-1536(88)80060-3
———, ———. 1991. Microcyclic conidiation cycles in Epichloetyphina. Mycologia 83:743–751, doi:10.2307/3760431
———, Porter JK, Robbins JD, Luttrell ES. 1977. Epichloetyphina from toxic tall fescue grasses. Appl EnvironMicrobiol 34:576–581.
Bazely DR, Ball JP, Vicari M, Tanentzap AJ, Berenger M,Rakocevic T, Koh S. 2007. Broad-scale geographicpatterns in the distribution of vertically transmitted,asexual endophytes in four naturally occurring grassesin Sweden. Ecography 30:367–374, doi:10.1111/j.0906-7590.2007.04985.x
———, Vicari M, Emmerich S, Filip L, Lin D, Inman A. 1997.Interactions between herbivores and endophyte-infectedFestuca rubra from the Scottish islands of St Kilda, Benbeculaand Rum. J Appl Ecol 34:847–860, doi:10.2307/2405276
Belesky DP, Devine OJ, Pallas JE, Stringer WC. 1987.Photosynthetic activity of tall fescue as influenced bya fungal endophyte. Photosynthetica 21:82–87.
Bradshaw AD. 1959. Population differentiation in Agrostistenuis Sibth. II. The incidence and significance ofinfection by Epichloe typhina. New Phytol 58:310–315,doi:10.1111/j.1469-8137.1959.tb05363.x
Brasier C. 2000. The rise of the hybrid fungi. Nature 405:134–135, doi:10.1038/35012193
Brem D. 2001. The role of the transmission mode for grassendophyte evolution [doctoral dissertation]. Zurich:Swiss Federal Institute of Technology. 136 p.
———, Leuchtmann A. 1999. High prevalence of horizon-tal transmission of the fungal endophyte Epichloesylvatica. Bull Geobot Inst ETH 65:3–12.
———, ———. 2003. Molecular evidence for host-adaptedraces of the fungal endophyte Epichloe bromicola afterpresumed host shifts. Evolution 57:37–51.
Bright M, Bulgheresi S. 2010. A complex journey: transmis-sion of microbial symbionts. Nat Rev Microbiol 8:218–230, doi:10.1038/nrmicro2262
Brosi GB, McCulley RL, Bush LP, Nelson JA, Classen AT,Norby RJ. 2011. Effects of multiple climate changefactors on the tall fescue-fungal endophyte symbiosis:infection frequency and tissue chemistry. New Phytol189:797–805, doi:10.1111/j.1469-8137.2010.03532.x
Bryant MK, May KJ, Bryan GT, Scott B. 2007. Functionalanalysis of a b-1,6-glucanase gene from the grassendophytic fungus Epichloe festucae. Fungal Genet Biol44:808–817, doi:10.1016/j.fgb.2006.12.012
Bucheli E, Leuchtmann A. 1996. Evidence for geneticdifferentiation between choke-inducing and asymptomticstrains of the Epichloe grass endophyte from Brachypodiumsylvaticum. Evolution 50:1879–1887, doi:10.2307/2410746
Bultman TL, Aguilera A, Sullivan TJ. 2012. Influence of fungalisolates infecting tall fescue on multitrophic interactions.Fungal Ecol 5:372–378, doi:10.1016/j.funeco.2011.06.004
———, Leuchtmann A. 2003. A test of host specialization byinsect vectors as a mechanism for reproductive isola-tion among entomophilous fungal species. Oikos 103:681–687, doi:10.1034/j.1600-0706.2003.12631.x
———, Welch AM, Boning RA, Bowdish TI. 2000. The costof mutualism in a fly-fungus interaction. Oecologia 124:85–90, doi:10.1007/s004420050027
———, White JF Jr. 1988. ‘‘Pollination’’ of a fungus by a fly.Oecologia 75:317–319, doi:10.1007/BF00378616
———, ———, Bowdish TI, Welch AM, Johnston J. 1995.Mutualistic transfer of Epichloe spermatia by Phorbiaflies. Mycologia 87:182–189, doi:10.2307/3760903
———, ———, ———, ———. 1998. A new kind ofmutualism between fungi and insects. Mycol Res 102:235–238, doi:10.1017/S0953756297004802
Bush LP, Fannin FF, Siegel MR, Dahlman DL, Burton HR.1993. Chemistry, occurrence and biological effectsof saturated pyrrolizidine alkaloids associated withendophyte-grass interactions. Agric Ecosyst Environ44:81–102, doi:10.1016/0167-8809(93)90040-V
———, Wilkinson HH, Schardl CL. 1997. Bioprotectivealkaloids of grass-fungal endophyte symbioses. PlantPhysiol 114:1–7.
Cao M, Koulman A, Johnson LJ, Lane GA. 2008. Advanceddata-mining strategies for the analysis of direct-infusionion trap mass spectrometry data from the association ofLolium perenne with its endophytic fungus Neotypho-dium lolii. Plant Physiol 146:1501–1514, doi:10.1104/pp.107.112458
Cheplick GP, Faeth SH. 2009. Ecology and evolution of thegrass-endophyte symbiosis. New York: Oxford Univ.Press. 241 p.
Christensen MJ, Ball OJP, Bennett R, Schardl CL. 1997.Fungal and host genotype effects on compatibility andvascular colonization by Epichloe festucae. Mycol Res101:493–501, doi:10.1017/S0953756296002833
———, Bennett RJ, Ansari HA, Koga H, Johnson RD, BryanGT, Simpson WR, Koolaard JP, Nickless EM, Voisey CR.
TADYCH ET AL.: BIOLOGY OF EPICHLOE 195

2008. Epichloe endophytes grow by intercalary hyphalextension in elongating grass leaves. Fungal Genet Biol45:84–93, doi:10.1016/j.fgb.2007.07.013
———, ———, Schmid J. 2002. Growth of Epichloe/Neotyphodium and p-endophytes in leaves of Loliumand Festuca grasses. Mycol Res 106:93–106, doi:10.1017/S095375620100510X
———, Saulsbury K, Simpson WR. 2012. Conspicuousepiphytic growth of an interspecific hybrid Neotyphodiumsp. endophyte on distorted host inflorescences. FungalBiol 116:42–48, doi:10.1016/j.funbio.2011.09.007
———, Simpson WR, Al Samarri T. 2000. Infection of tallfescue and perennial ryegrass plants by combinationsof different Neotyphodium endophytes. Mycol Res 104:974–978, doi:10.1017/S0953756299002300
———, Voisey CR. 2009. Tall fescue-endophyte symbiosis.In: Fribourg HA, Hannaway DB, West CP, eds. Tallfescue for the twenty-first century. Agron Monogr 53:ASA, CSSA, SSSA. p 251–272.
Chung K-R, Schardl CL. 1997. Sexual cycle and horizontaltransmission of the grass symbiont, Epichloe typhina.Mycol Res 101:295–301, doi:10.1017/S0953756296002602
Clay K. 1986. Induced vivipary in the sedge Cyperus virensand the transmission of the fungus Balansia cyperi(Clavicipitaceae). Can J Bot 64:2984–2988, doi:10.1139/b86-394
———. 1990. Fungal endophytes of grasses. Annu Rev Ecol Syst21:275–298, doi:10.1146/annurev.es.21.110190.001423
———, Schardl CL. 2002. Evolutionary origins and ecolog-ical consequences of endophyte symbiosis with grasses.Am Nat 160:S99–S127, doi:10.1086/342161
Craven KD. 2012. Population studies of native grass-endophytesymbioses provide clues for the roles of host jumps andhybridization in driving their evolution. Mol Ecol 21:2562–2564, doi:10.1111/j.1365-294X.2012.05564.x
———, Blankenship JD, Leuchtmann A, Hignight K,Schardl CL. 2001. Hybrid fungal endophytes symbioticwith grass Lolium pratense. Sydowia 53:44–73.
Crawford KM, Land JM, Rudgers JA. 2010. Fungal endo-phytes of native grasses decrease insect herbivorepreference and performance. Oecologia 164:431–444,doi:10.1007/s00442-010-1685-2
Czarnoleski M, Olejniczak P, Gorzynska K, Kozłowski J,Lembicz M. 2013. Altered allocation to roots and shootsin the endophyte-infected seedlings of Puccinelliadistans (Poaceae). Plant Biol 15:264–273, doi:10.1111/j.1438-8677.2012.00633.x
———, ———, Mikołajczak P, Lembicz M, Kozłowski J.2010. Fungal endophytes protect grass seedlingsagainst herbivory and allow economical seed produc-tion. Evol Ecol Res 12:769–777.
Dugan FM, Sitton JW, Sullivan RF, White JF Jr. 2002. TheNeotyphodium endophyte of barley (Hordeum brevisu-bulatum subsp. violaceum) grows and sporulates on leafsurfaces of the host. Symbiosis 32:147–159.
Eckblad F-E, Torkelsen A-E. 1989. Epichloe typhina inNorway. Opera Bot 100:51–57.
Eriksson O. 1967. On graminicolous pyrenomycetes fromFennoscandia 2. Phragmosporous and scolecosporousspecies. Arkiv Bot 6:381–441.
Faeth SH. 2002. Are endophytic fungi defensive plantmutualists? Oikos 98:25–36, doi:10.1034/j.1600-0706.2002.980103.x
———. 2009. Asexual fungal symbionts alter reproductiveallocation and herbivory over time in their native perennialgrass hosts. Am Nat 173:554–565, doi:10.1086/597376
———, Saari S. 2012. Fungal grass endophytes andarthropod communities: lessons from plant defencetheory and multitrophic interactions. Fungal Ecol 5:364–371, doi:10.1016/j.funeco.2011.09.003
———, Sullivan TJ. 2003. Mutualistic asexual endophytes ina native grass are usually parasitic. Am Nat 161:310–325, doi:10.1086/345937
Fletcher LR, Harvey IC. 1981. An association of a lolitremendophyte with ryegrass staggers. NZ Vet J 29:185–186,doi:10.1080/00480169.1981.34839
Freeman EM. 1904. The seed fungus of Lolium temulentum,L., the Darnel. Phil Trans R Soc Lond B 196:1–27,doi:10.1098/rstb.1904.0001
Gallagher RT, Hawkes AD, Steyn PS, Vleggaar R. 1984.Tremorgenic neurotoxins from perennial ryegrasscausing ryegrass staggers disorder of livestock: structureelucidation of lolitrem B. J Chem Soc, Chem Commun1984:614–616, doi:10.1039/c39840000614
Gentile A, Rossi MS, Cabral D, Craven KD, Schardl CL. 2005.Origin, divergence and phylogeny of epichloe endo-phytes of native Argentine grasses. Mol Phylogenet Evol35:196–208, doi:10.1016/j.ympev.2005.01.008
Gibert A, Volaire F, Barre P, Hazard L. 2012. A fungalendophyte reinforces population adaptive differentia-tion in its host grass species. New Phytol 194:561–571,doi:10.1111/j.1469-8137.2012.04073.x
Giraud T, Refregier G, Le Gac M, de Vienne DM, Hood ME.2008. Speciation in fungi. Fungal Genet Biol 45:791–802, doi:10.1016/j.fgb.2008.02.001
Glenn AE, Bacon CW, Price R, Hanlin RT. 1996. Molecularphylogeny of Acremonium and its taxonomic implica-tions. Mycologia 88:369–383, doi:10.2307/3760878
Gorzynska K, Lembicz M, Olszanowski Z, Leuchtmann A.2010. An unusual Botanophila–Epichloe association in apopulation of orchardgrass (Dactylis glomerata) inPoland. J Nat Hist 44:2817–2824, doi:10.1080/00222933.2010.521595
———, ———, ———, ———. 2011. Botanophila-Epichloeinteraction in a wild grass, Puccinellia distans, lacksdependence on the fly vector. Ann Entomol Soc Am104:841–846, doi:10.1603/AN11009
Granath G, Vicari M, Bazely DR, Ball JP, Puentes A,Rakocevic T. 2007. Variation in the abundance offungal endophytes in fescue grasses along altitudinaland grazing gradients. Ecography 30:422–430, doi:10.1111/j.0906-7590.2007.05027.x
Groppe K, Steinger T, Sanders I, Schmid B, Wiemken A,Boller T. 1999. Interaction between the endophyticfungus Epichloe bromicola and the grass Bromus erectus:effects of endophyte infection, fungal concentration andenvironment on grass growth and flowering. Mol Ecol 8:1827–1835, doi:10.1046/j.1365-294x.1999.00772.x
———, ———, Schmid B, Baur B, Boller T. 2001. Effects ofhabitat fragmentation on choke disease (Epichloe
196 MYCOLOGIA

bromicola) in the grass Bromus erectus. J Ecol 89:247–255, doi:10.1046/j.1365-2745.2001.00538.x
Gundel PE, Garibaldi LA, Martınez-Ghersa MA, Ghersa CM.2012. Trade-off between seed number and weight:influence of a grass-endophyte symbiosis. Basic ApplEcol 13:32–39, doi:10.1016/j.baae.2011.10.008
———, Helander M, Casas C, Hamilton CE, Faeth SH,Saikkonen K. 2013. Neotyphodium fungal endophyte intall fescue (Schedonorus phoenix): a comparison ofthree northern European wild populations and thecultivar Kentucky-31. Fungal Divers 60:15–24, doi:10.1007/s13225-012-0173-x
Hamilton CE, Bauerle TL. 2012. A new currency formutualism? Fungal endophytes alter antioxidant activ-ity in hosts responding to drought. Fungal Divers 54:39–49, doi:10.1007/s13225-012-0156-y
———, Dowling TE, Faeth SH. 2010. Hybridization inendophyte symbionts alters host response to moistureand nutrient treatments. Microbial Ecol 59:768–775,doi:10.1007/s00248-009-9606-9
———, Gundel PE, Helander M, Saikkonen K. 2012.Endophytic mediation of reactive oxygen species andantioxidant activity in plants: a review. Fungal Divers54:1–10, doi:10.1007/s13225-012-0158-9
Herd S, Christensen MJ, Saunders K, Scott DB, Schmid J.1997. Quantitative assessment of in planta distributionof metabolic activity and gene expression of anendophytic fungus. Microbiology 143:267–275, doi:10.1099/00221287-143-1-267
Hughes WE, Hallidays S. 1980. Puccinellia Parl. In: TutinTG, Heywood VH, Burges NA, Moore DM, ValentineOH, Walter SM, Webb DA, eds. Flora Europaea 5.Alismataceae to Orchidaceae (Monocotyledones).Cambridge, UK: Cambridge Univ. Press.
Iannone LJ, Cabral D, Schardl CL, Rossi MS. 2009.Phylogenetic divergence, morphological and physio-logical differences distinguish a new Neotyphodiumendophyte species in the grass Bromus auleticus fromSouth America. Mycologia 101:336–347, doi:10.3852/08-156
———, Corallo AB, Mc Cargo PD, Lupo S. 2012b. Hostpreliminary register of epichloid endophytyes fromUruguay and other countries of South America. In:Nan Z, Li C, eds. Proceedings of the 8th InternationalSymposium on Fungal Endophytes of Grasses. Lanz-hou, PR China: Lanzhou University. p 31–33.
———, Irizarri JGN, Perez LI, Gundel PE. 2012c. Neotypho-dium endophytes in the native grass Hordeum comosumfrom Patagonia (Argentina). In: Nan Z, Li C, eds.Proceedings of the 8th International Symposium onFungal Endophytes of Grasses. Lanzhou, PR China:Lanzhou University. p 34–37.
———, Mc Cargo PD, Giussani LM, Schardl CL. 2013.Geographic distribution patterns of vertically transmit-ted endophytes in two native grasses in Argentina.Symbiosis 59:99–110, doi:10.1007/s13199-012-0214-y
———, Novas MV, Young CA, De Battista JP, Schardl CL.2012a. Endophytes of native grasses from SouthAmerica: biodiversity and ecology. Fungal Ecol 5:357–363, doi:10.1016/j.funeco.2011.05.007
———, White JF Jr, Giussani LM, Cabral D, Novas MV. 2011.Diversity and distribution of Neotyphodium-infectedgrasses in Argentina. Mycol Prog 10:9–19, doi:10.1007/s11557-010-0669-2
Ingold CT. 1948. The water relations of spore discharge inEpichloe. Trans Br Mycol Soc 31:277–288, doi:10.1016/S0007-1536(48)80011-2
Jackowiak B. 1984. Chorologiczne i synekologiczne aspektyekspansji Puccinellia distans (L.) Parl. w Polsce. BadFizjgraf Polska Zachodnia 35:67–91.
———. 1996. Chorological-ecological model of the spreadof Puccinellia distans (Poaceae) in central Europe.Fragm Florist Geobot 41:551–561.
———, Taraszkiewicz M, Wosicka M. 1991. Wpływ terminuwysiewu ziarniakow na rozwoj fenologiczny Puccinelliadistans (L.) Parl. Sprawozdania-Poznanskie Towar-zystwo Przyjacioł Nauk, Wydział Matematyczno-Przyr-odniczy 108:9–12.
Ji YL, Zhan LH, Kang Y, Sun XH, Yu HS, Wang ZW. 2009. Anew stromata-producing Neotyphodium species symbiot-ic with clonal grass Calamagrostis epigeios (L.) Roth.grown in China. Mycologia 101:200–205, doi:10.3852/08-044
Johnson LJ, Koulman A, Christensen M, Lane GA, Fraser K,et al. 2013. An extracellular siderophore is required tomaintain the mutualistic interaction of Epichloe festucaewith Lolium perenne. PLoS Pathog 9:e1003332. doi:10.1371/journal.ppat.1003332, doi:10.1371/annotation/3e7e6415-fb12-4a87-89e6-f87d2e800ba8
———, Steringa M, Koulman A, Christensen M, JohnsonRD, Voisey CR, Bryan G, Lamont I, Rasmussen S. 2007a.Biosynthesis of an extracellular siderophore is essentialfor maintenance of mutualistic endophyte-grass symbi-osis. In: Popay AJ, Thom ER, eds. Proceedings of the6th International Symposium on Fungal Endophytes ofGrasses. Grassl Res Pract Ser No 13. Christchurch: NewZealand Grass Association. p 177–179.
Johnson R, Voisey C, Johnson L, Pratt J, Fleetwood D, KhanA, Bryan G. 2007b. Distribution of NRPS gene familieswithin the Neotyphodium/Epichloe complex. FungalGenet Biol 44:1180–1190, doi:10.1016/j.fgb.2007.04.009
Ju H-J, Hill NS, Abbott T, Ingram KT. 2006. Temperatureinfluences on endophyte growth in tall fescue. Crop Sci46:404–412, doi:10.2135/cropsci2005.0282
Kang Y, Ji YL, Zhu K, Wang H, Miao H, Wang ZW. 2011. Anew Epichloe species with interspecific hybrid originsfrom Poa pratensis ssp. pratensis in Liyang, China.Mycologia 103:1341–1350, doi:10.3852/10-352
Kirby EJM. 1961. Host-parasite relations in the chokedisease of grasses. Trans Br Mycol Soc 44:493–503,doi:10.1016/S0007-1536(61)80048-X
Kohlmeyer J, Kohlmeyer E. 1974. Distribution of Epichloetyphina (Ascomycetes) and its parasitic fly. Mycologia66:77–86, doi:10.2307/3758455
Koponen H, Makela K. 1976. Phyllachora graminis, P.silvatica, Epichloe typhina and Acrospermum graminumon grasses in Finland. Karstenia 15:46–55.
Koulman A, Lee TV, Fraser K, Johnson L, Arcus V, Lott JS,Rasmussen S, Lane G. 2012. Identification of extracel-lular siderophores and a related peptide from the
TADYCH ET AL.: BIOLOGY OF EPICHLOE 197

endophytic fungus Epichloe festucae in culture andendophyte-infected Lolium perenne. Phytochemistry 75:128–139, doi:10.1016/j.phytochem.2011.11.020
———, Tapper BA, Fraser K, Cao M, Lane GA, RasmussenS. 2007. High throughput direct infusion ion trap massspectrometry: a new method for metabolomics. RapidCommun Mass Spectrom 21:421–428, doi:10.1002/rcm.2854
Kover PX, Clay K. 1998. Trade-off between virulence andvertical transmission and the maintenance of a virulentplant pathogen. Am Nat 152:165–175, doi:10.1086/286159
Kuldau GA, Tsai H-F, Schardl CL. 1999. Genome sizes ofEpichloe species and anamorphic hybrids. Mycologia 91:776–782, doi:10.2307/3761531
Latch GCM, Christensen MJ, Samuels GJ. 1984. Fiveendophytes of Lolium and Festuca in New Zealand.Mycotaxon 20:535–550.
Lehtonen P, Helander M, Saikkonen K. 2005. Are endo-phyte-mediated effects on herbivores conditional onsoil nutrients? Oecologia 142:38–45, doi:10.1007/s00442-004-1701-5
Lembicz M. 1998. Life history of Puccinellia distans (L.)Parl. (Poaceae) in the colonization of anthropogenichabitats. Phytocoenosis 10:1–32.
———, Gorzynska K, Leuchtmann A. 2010. Choke diseasecaused by Epichloe bromicola in the grass Agropyron repensin Poland. Plant Dis 94:1372, doi:10.1094/PDIS-12-09-0810
———, ———, Olejniczak P, Leuchtmann A. 2011.Geographical distribution and effects of choke diseasecaused by Epichloe typhina in populations of the grassPuccinellia distans in Poland. Sydowia 63:35–48.
———, ———, Olszanowski Z, Michelsen V, LeuchtmannA. 2013. The occurrence and preference of Botanophilaflies (Diptera: Anthomyiidae) for particular species ofEpichloe fungi infecting wild grasses. Eur J Entomol110:129–134, doi:10.14411/eje.2013.018
———, Olejniczak P. 2009. The fungus Epichloe typhina inpopulations of a halophyte Puccinellia distans: salinityas a possible inhibitor of infection. Acta Soc Bot Pol 78:81–86, doi:10.5586/asbp.2009.011
———, ———, Olszanowski Z, Gorzynska K, LeuchtmannA. 2009. Man-made habitats—hotspots of evolutionarygame between grass, fungus and fly. Biodiv Res Conserv15:47–52, doi:10.2478/v10119-009-0017-z
Leuchtmann A. 1992. Systematics, distribution and hostspecificity of grass endophytes. Nat Toxins 1:150–162,doi:10.1002/nt.2620010303
———. 2003. Taxonomy and diversity of Epichloe endo-phytes. In: White JF Jr, Bacon CW, Hywel-Jones NL,Spatafora JW, eds. Clavicipitalean fungi: evolutionarybiology, chemistry, biocontrol and cultural impacts.New York: Marcel Dekker. p 169–194.
———. 2007. Botanophila flies on Epichloe host species inEurope and North America: no evidence for coevolution.Entomol Exp Appl 123:13–23, doi:10.1111/j.1570-7458.2006.00518.x
———, Bacon CW, Schardl CL, White JF Jr, Tadych M. 2014.Nomenclatural realignment of Neotyphodium specieswith genus Epichloe. Mycologia 106:202–215.
———, Schardl CL. 1998. Mating compatibility andphylogenetic relationships among two new species ofEpichloe and other congeneric European species. MycolRes 102:1169–1182, doi:10.1017/S0953756298006236
———, ———, Siegel MR. 1994. Sexual compatibility andtaxonomy of a new species of Epichloe symbiotic with finefescue grasses. Mycologia 86:802–812, doi:10.2307/3760595
Leyronas C, Raynal G. 2008a. Nouvelles observations sur lesspores d’Epichloe typhina (Fr. ex Tul.), Clavicipitaceepathogene des graminees. Cryptogam Mycol 29:11–17.
———, ———. 2008b. Role of fungal ascospores in theinfection of orchardgrass (Dactylis glomerata) byEpichloe typhina agent of choke disease. J Plant Pathol90:15–21.
Li HM, Crouch JA, Belanger FC. 2005. Fungal endophyteN-acetylglucosaminidase expression in the infectedhost grass. Mycol Res 109:363–373, doi:10.1017/S0953756204002205
Lindstrom JT, Sun S, Belanger FC. 1993. A novel fungalprotease expressed in endophytic infection of Poaspecies. Plant Physiol 102:645–650.
Lyons PC, Plattner RD, Bacon CW. 1986. Occurrence ofpeptide and clavinet ergot alkaloids in tall fescue grass.Science 232:487–489, doi:10.1126/science.3008328
Malinowski DP, Alloush GA, Belesky DP. 1998. Evidence forchemical changes on the root surface of tall fescue inresponse to infection with the fungal endophyteNeotyphodium coenophialum. Plant Soil 205:1–12,doi:10.1023/A:1004331932018
———, Belesky DP. 2000. Adaptations of endophyte-infectedcool-season grasses to environmental stresses: mecha-nisms of drought and mineral stress tolerance. Crop Sci40:923–940, doi:10.2135/cropsci2000.404923x
May KJ, Bryant MK, Zhang X, Ambrose B, Scott B. 2008.Patterns of expression of a lolitrem biosynthetic genein the Epichloe festucae-perennial ryegrass symbiosis.Mol Plant-Microbe Interact 21:188–197, doi:10.1094/MPMI-21-2-0188
Mc Cargo PD, Iannone LJ, Vignale MV, Schardl CL, RossiMS. 2014. Species diversity of Epichloe symbiotic withtwo grasses from southern Argentinean Patagonia.Mycologia 106:339–352.
Meijer G, Leuchtmann A. 1999. Multistrain infections of thegrass Brachypodium sylvaticum by its fungal endophyteEpichloe sylvatica. New Phytol 141:355–368, doi:10.1046/j.1469-8137.1999.00332.x
———, ———. 2000. The effects of genetic and environ-mental factors on disease expression (stroma forma-tion) and plant growth in Brachypodium sylvaticuminfected by Epichloe sylvatica. Oikos 91:446–458,doi:10.1034/j.1600-0706.2000.910305.x
———, ———. 2001. Fungal genotype controls mutualismand sex in Brachypodium sylvaticum infected by Epichloesylvatica. Acta Biol Hung 52:249–263, doi:10.1556/ABiol.52.2001.2-3.9
Moon CD, Craven KD, Leuchtmann A, Clement SL, SchardlCL. 2004. Prevalence of interspecific hybrids amongasexual fungal endophytes of grasses. Mol Ecol 13:1455–1467, doi:10.1111/j.1365-294X.2004.02138.x
198 MYCOLOGIA

———, Guillaumin J-J, Ravel C, Li C, Craven KD, SchardlCL. 2007. New Neotyphodium endophyte species fromthe grass tribes Stipeae and Meliceae. Mycologia 99:895–905, doi:10.3852/mycologia.99.6.895
———, Miles CO, Jarlfors U, Schardl CL. 2002. Theevolutionary origins of three new Neotyphodium endo-phyte species from grasses indigenous to the southernhemisphere. Mycologia 94:694–711, doi:10.2307/3761720
Moravcova L, Frantık T. 2002. Germination ecology ofPuccinellia distans and P. limosa. Biol Brat 57:441–448.
Moy M, Belanger F, Duncan R, Freehoff A, Leary C, MeyerWR, Sullivan R, White JF Jr. 2000. Identification ofepiphyllous mycelial nets on leaves of grasses infectedby clavicipitaceous endophytes. Symbiosis 28:291–302.
———, Li HM, Sullivan R, White JF Jr, Belanger FC. 2002.Endophytic fungal b-1,6-glucanase expression in theinfected host grass. Plant Physiol 130:1298–1308,doi:10.1104/pp.010108
Novas MV, Collantes M, Cabral D. 2007. Environmentaleffects on grass-endophyte associations in the harshconditions of south Patagonia. FEMS Microbiol Ecol61:164–173, doi:10.1111/j.1574-6941.2007.00319.x
Oberhofer M. 2012. Significance of hybridization in Epichloeendophytes of Hordelymus europaens [doctoraldissertation]. Zurich, Switzerland: ETH Zurich. 131 p.http://dx.doi.org/10.3929/ethz-a-007339468
———, Leuchtmann A. 2012. Genetic diversity in epichloidendophytes of Hordelymus europaeus suggests repeatedhost jumps and interspecific hybridizations. Mol Ecol21:2713–2726, doi:10.1111/j.1365-294X.2012.05459.x
Olejniczak P, Lembicz M. 2007. The fungus Epichloe typhinain populations of a halophyte Puccinellia distans:salinity as a possible inhibitor of infection. Oecologia152:485–494, doi:10.1007/s00442-007-0660-z
Panaccione DG, Johnson RD, Wang J, Young CA, Dam-rongkool P, Scott B, Schardl CL. 2001. Elimination ofergovaline from a grass-Neotyphodium endophyte sym-biosis by genetic modification of the endophyte. ProcNatl Acad Sci USA 98:12820–12825, doi:10.1073/pnas.221198698
Parker L, Bultman TL. 1991. Ecological cost in a fly-fungusmutualism. Am Midl Nat 126:208–211, doi:10.2307/2426166
Philipson MN, Christey MC. 1986. The relationship of hostand endophyte during flowering, seed formation andgermination of Lolium perenne. NZ J Bot 24:125–134,doi:10.1080/0028825X.1986.10409724
Qawasmeh A, Obied HK, Raman A, Wheatley W. 2012a.Influence of fungal endophyte infection on phenoliccontent and antioxidant activity in grasses: interactionbetween Lolium perenne and different strains ofNeotyphodium lolii. J Agric Food Chem 60:3381–3388,doi:10.1021/jf204105k
———, Raman A, Wheatley W, Nicol H. 2012b. Antiox-idative capacity of phenolic compounds extracted fromLolium perenne and Lolium arundinaceum infected withNeotyphodium (Hypocreales: Clavicipitaceae). Acta Phy-siol Plant 34:827–833, doi:10.1007/s11738-011-0878-6
Rao S, Baumann D. 2004. The interaction of a Botanophilafly species with an exotic Epichloe fungus in a cultivatedgrass: fungivore or mutualist? Entomol Exp Appl 112:99–105, doi:10.1111/j.0013-8703.2004.00189.x
———, Alderman S, Kaser J, Hoffman G. 2010. Bugs, slugsand exotic Epichloe typhina: close encounters of theOregon kind. In: Abstracts from the Joint Meeting ofthe MSA and International Symposium on FungalEndophytes of Grasses (ISFEG). Lexington, Kentucky,28 Jun–1 Jul 2010. Inoculum 61:69.
———, ———, ———, ———. 2012. Fertilization ofEpichloe typhina in cultivated Dactylis glomerata byfactors besides Botanophila flies. In: Young CA, AikenGE, McCulley RL, Strickland JR, Schardl CL, eds.Epichloae, endophytes of cool season grasses: implica-tions, utilization and biology. Ardmore, Oklahoma:The Samuel Roberts Noble Foundation. p 122–126.
Rasmussen S, Parsons AJ, Fraser K, Xue H, Newman JA.2008. Metabolic profiles of Lolium perenne are differ-entially affected by nitrogen supply, carbohydratecontent and fungal endophyte infection. Plant Physiol146:1440–1453, doi:10.1104/pp.107.111898
———, ———, Newman JA. 2009. Metabolic analysis of theLolium perenne-Neotyphodium lolii symbiosis: more thanjust alkaloids? Phytochem Rev 8:535–550, doi:10.1007/s11101-009-9136-6
Raynal G. 1991a. Liberation des ascospores d’Epichloetyphina, agent de la quenouille du dactyle. Conse-quences pour l’epidemiologie et la lute. Fourrages 127:345–358.
———. 1991b. Les Acremonium, champignons endophytesdes graminees prairiales. Des symbiotes empoisonnantsmais captivants. Phytoma LdV 428:35–40.
Rodriguez RJ, White JF Jr, Arnold AE, Redman RS. 2009. Fungalendophytes: diversity and functional roles. New Phytol 182:314–330, doi:10.1111/j.1469-8137.2009.02773.x
Rowan DD, Gaynor DL. 1986. Isolation of feeding deter-rents against Argentine stem weevil from ryegrassinfected with the endophyte Acremonium loliae. J ChemEcol 12:647–658, doi:10.1007/BF01012099
Rudgers JA, Clay K. 2008. An invasive plant-fungal mutual-ism reduces arthropod diversity. Ecol Lett 11:831–840,doi:10.1111/j.1461-0248.2008.01201.x
Saikkonen K, Ahlholm J, Helander M, Lehtimaki S,Niemelainen O. 2000. Endophytic fungi in wild andcultivated grasses in Finland. Ecography 23:360–366,doi:10.1111/j.1600-0587.2000.tb00292.x
———, Lehtonen P, Helander M, Koricheva J, Faeth SH. 2006.Model systems in ecology: dissecting the endophyte-grassliterature. Trends Plant Sci 11:428–433, doi:10.1016/j.tplants.2006.07.001
———, Wali P, Helander M. 2010. Genetic compatibilitydetermines endophyte-grass combinations. PLoS ONE5:e11395, doi:10.1371/journal.pone.0011395
———, ———, ———, Faeth S. 2004. Evolution ofendophyte-plant symbioses. Trends Plant Sci 9:275–280, doi:10.1016/j.tplants.2004.04.005
Sampson K. 1933. The systemic infection of grasses byEpichloe typhina (Pers.) Tul. Trans Br Mycol Soc 18:30–47, doi:10.1016/S0007-1536(33)80025-8
TADYCH ET AL.: BIOLOGY OF EPICHLOE 199

Schardl CL. 2010. The epichloae, symbionts of the grasssubfamily Pooideae. Ann Missouri Bot Gard 97:646–665, doi:10.3417/2009144
———, Craven KD. 2003. Interspecific hybridization inplant-associated fungi and oomycetes: a review. MolEcol 12:2861–2873, doi:10.1046/j.1365-294X.2003.01965.x
———, Leuchtmann A. 2005. The Epichloe endophytes ofgrasses and the symbiotic continuum. In: Dighton J,White JF Jr, Oudemans P, eds. The fungal community:its organization and role in the ecosystem. Boca Raton,Florida: CRS Press, Taylor & Francis Group. p 475–503.
———, ———, Chung K-R, Penny D, Siegel MR. 1997.Coevolution by common descent of fungal symbionts(Epichloe spp.) and grass hosts. Mol Biol Evol 14:133–143, doi:10.1093/oxfordjournals.molbev.a025746
———, ———, Spiering MJ. 2004. Symbioses of grass withseedborne fungal endophytes. Annu Rev Plant Biol 55:315–340, doi:10.1146/annurev.arplant.55.031903.141735
———, ———, Tsai H-F, Collett MA, Watt DM, Scott DB.1994. Origin of a fungal symbiont of perennial ryegrassby interspecific hybridization of a mutualist with theryegrass choke pathogen, Epichloe typhina. Genetics136:1307–1317.
———, Moon CD. 2003. Processes of species evolution inEpichloe/Neotyphodium endophytes of grasses. In:White JF Jr, Bacon CW, Hywel-Jones NL, SpataforaJW, eds. Clavicipitalean fungi: evolutionary biology,chemistry, biocontrol and cultural impacts. New York:Marcel Dekker. p 273–310.
———, Scott B, Florea S, Zhang D. 2009. Epichloeendophytes: clavicipitaceous symbionts of grasses. In:Deising HB, ed. The Mycota V: plant relationships. 2nded. Berlin: Springer-Verlag. p 275–306.
———, Young CA, Faulkner JR, Florea S, Pan J. 2012.Chemotypic diversity of epichloae, fungal symbionts ofgrasses. Fungal Ecol 5:331–344, doi:10.1016/j.funeco.2011.04.005
———, ———, Hesse U, Amyotte SG, Andreeva K, et al.2013. Plant-symbiotic fungi as chemical engineers:multi-genome analysis of the Clavicipitaceae revealsdynamics of alkaloid loci. PLoS Genetics 9:e1003323,doi:10.1371/journal.pgen.1003323
Schiestl FP, Steinebrunner F, Schulz C, von Reuß S, FranckeW, Weymuth C, Leuchtmann A. 2006. Evolution of‘pollinator’-attracting signals in fungi. Biol Lett 2:401–404, doi:10.1098/rsbl.2006.0479
Schmidt SP, Hoveland CS, Clark EM, Davis ND, Smith LA,Grimes HW, Holliman JL. 1982. Association of endo-phytic fungi with fescue toxicity in steers fed Kentucky-31 tall fescue seed or hay. J Anim Sci 55:1259–1263.
Siegel MR, Johnson MC, Varney DR, Nesmith WC, BucknerRC, Bush LP, Burrus PB II, Jones TA, Boling JA. 1984. Afungal endophyte in tall fescue: incidence and dissem-ination. Phytopathology 74:932–937, doi:10.1094/Phyto-74-932
Spatafora JW, Sung G-H, Sung J-M, Hywel-Jones NL, WhiteJF Jr. 2007. Phylogenetic evidence for an animalpathogen origin of ergot and the grass endophytes.
Mol Ecol 16:1701–1711, doi:10.1111/j.1365-294X.2007.03225.x
Steinebrunner F, Twele R, Francke W, Leuchtmann A,Schiestl FP. 2008a. Role of odor compounds in theattraction of gamete vectors in endophytic Epichloe fungi.New Phytol 178:401–411, doi:10.1111/j.1469-8137.2007.02347.x
———, Schiestl FP, Leuchtmann A. 2008b. Variation ofinsect attracting odor in endophytic Epichloe fungi:phylogenetic constraints versus host influence. J ChemEcol 34:772–782, doi:10.1007/s10886-008-9476-5
———, ———, ———. 2008c. Ecological role of volatilesproduced by Epichloe: differences in antifungal toxicity.FEMS Microb Ecol 64:307–316, doi:10.1111/j.1574-6941.2008.00452.x
Sun S, Clarke BB, Funk CR. 1990. Effect of fertilizer andfungicide applications on choke expression and endo-phyte transmission in chewings fescue. In: QuisemberrySS, Joost RE, eds. Proceedings of the InternationalSymposium on Acremonium/Grass Interactions. BatonRouge: Louisiana Agricultural Experiment Station.p 62–66.
Tadych M, Ambrose KV, Bergen MS, Belanger FC, White JFJr. 2012. Taxonomic placement of Epichloe poae sp. nov.and horizontal dissemination to seedlings via conidia.Fungal Divers 54:17–131.
———, Bergen M, Dugan FM, White JF Jr. 2007. Evaluationof the potential role of water in spread of conidia of theNeotyphodium endophyte of Poa ampla. Mycol Res 111:466–472, doi:10.1016/j.mycres.2007.02.002
———, White JF Jr. 2007. Ecology of epiphyllous stages ofendophytes and implications for horizontal dissemina-tion. In: Popay AJ, Thom ER, eds. Proceedings of the6th International Symposium on fungal endophytes ofgrasses. Grassl Res Pract Ser No. 13. Christchurch: NewZealand Grass Association. p 157–161.
Takach JE, Mittal S, Swoboda GA, Bright SK, Trammell MA,Hopkins AA, Young CA. 2012. Genotypic and chemo-typic diversity of Neotyphodium endophytes in tallfescue from Greece. Appl Environ Microbiol 78:5501–5510, doi:10.1128/AEM.01084-12
Tan YY, Spiering MJ, Scott V, Lane GA, Christensen MJ,Schmid J. 2001. In planta regulation of extension of anendophytic fungus and maintenance of high metabolicrates in its mycelium in the absence of apical extension.Appl Environ Microb 67:5377–5383, doi:10.1128/AEM.67.12.5377-5383.2001
Tintjer T, Leuchtmann A, Clay K. 2008. Variation inhorizontal and vertical transmission of the endophyteEpichloe elymi infecting the grass Elymus hystrix. NewPhytol 179:236–246, doi:10.1111/j.1469-8137.2008.02441.x
Torres MS, White JF Jr, Zhang X, Hinton DM, Bacon CW.2012. Endophyte-mediated adjustments in host mor-phology and physiology and effects on host fitness traitsin grasses. Fungal Ecol 5:322–330, doi:10.1016/j.funeco.2011.05.006
Tsai H-F, Liu J-S, Staben C, Christensen MJ, Latch GCM,Siegel MR, Schardl CL. 1994. Evolutionary diversifica-
200 MYCOLOGIA

tion of fungal endophytes of tall fescue grass byhybridization with Epichloe species. Proc Natl AcadSci USA 91:2542–2546, doi:10.1073/pnas.91.7.2542
Vare H, Itamies J. 1995. Phorbia phrenione (Seguy) (Diptera:Anthomyiidae) in Finland. Sahlbergia 2:119–124.
Wali PR, Ahlholm JU, Helander M, Saikkonen K. 2007.Occurrence and genetic structure of the systemic grassendophyte Epichloe festucae in fine fescue populations.Microb Ecol 53:20–29, doi:10.1007/s00248-006-9076-2
Webster J, Weber RWS. 2007. Introduction to fungi. 3rd ed.New York: Cambridge Univ. Press. 841 p.
Welch AM, Bultman TL. 1993. Natural release of Epichloetyphina ascospores and its temporal relationship tofly parasitism. Mycologia 85:756–763, doi:10.2307/3760606
Wennstrom A. 1996. The distribution of Epichloe typhina innatural plant populations of the host plant Calamagrostispurpurea. Ecography 19:377–381, doi:10.1111/j.1600-0587.1996.tb00248.x
Western JH, Cavett JJ. 1959. The choke disease of cocksfoot(Dactylis glomerata) caused by Epichloe typhina (Fr.)Tul. Trans Br Mycol Soc 42:298–307, doi:10.1016/S0007-1536(56)80037-5
White JF Jr. 1988. Endophyte-host associations in foragegrasses XI. A proposal concerning origin and evolu-tion. Mycologia 80:442–446, doi:10.2307/3807845
———. 1997. Perithecial ontogeny in the fungal genusEpichloe: an examination of the clavicipitalean cen-trum. Am J Bot 84:170–178, doi:10.2307/2446078
———, Bultman TL. 1987. Endophyte-host associations inforage grasses VIII. Heterothallism in Epichloe typhina.Am J Bot 74:1716–1721, doi:10.2307/2444142
———, Camp C. 1995. Impact of the fungal endophyteEpichloe amarillans on the grass Agrostis hiemalis: astudy of water relations. Symbiosis 18:15–25.
———, Chambless DA. 1991. Endophyte-host associationsin forage grasses XV. Clustering of stromata-bearingindividuals of Agrostis hiemalis infected by Epichloetyphina. Am J Bot 78:527–533, doi:10.2307/2445262
———, Glenn AE, Chandler KF. 1993. Endophyte-hostassociations in grasses XVIII. Moisture relations andinsect herbivory of the emergent stromal leaf ofEpichloe. Mycologia 85:195–202, doi:10.2307/3760456
———, Halisky PM, Sun SC, Morgan-Jones G, Funk CR Jr.1992. Endophyte-host associations in grasses XVI.Patterns of endophyte distribution in species of the tribeAgrostideae. Am J Bot 79:472–477, doi:10.2307/2445162
———, Martin TI, Cabral D. 1996. Endophyte-host associationsin grasses XXII. Conidia formation by Acremoniumendophytes on the phylloplanes of Agrostis hiemalis andPoa rigidifolia. Mycologia 88:174–178, doi:10.2307/3760920
———, Morrow AC, Morgan-Jones G, Chambless DA. 1991.Endophyte-host associations in forage grasses XIV.Primary stromata formation and seed transmission inEpichloe typhina: development and regulatory aspects.Mycologia 83:72–81, doi:10.2307/3759833
———, Torres MS. 2010. Is plant endophyte-mediateddefensive mutualism the result of oxidative stressprotection? Physiol Plant 138:440–446, doi:10.1111/j.1399-3054.2009.01332.x
Yanagida N, Irie T, Tanaka E, Teramoto C, Kuwabara K,Tajimi A. 2005. New choke diseases and their molecularphylogenetic analysis in Agropron ciliare var. minus andAgropyron tsukushiense var. transiens. Mycologia 97:1287–1291, doi:10.3852/mycologia.97.6.1287
TADYCH ET AL.: BIOLOGY OF EPICHLOE 201
Top Related
Copyright © 2022 FDOKUMEN

