







Bahasa
Halaman
Hukum


R. Kishore D.J. Hicklin D.V. Dellaratta U. Golde T. Kageshita B. Seliger S. Ferrone
Key words: LMP; monoclonal: peptide; recombinant protein; TAP
Acknowledgments: The authors wish to acknowledge the excellent secretarial assistance of M r s . Harriett V. Harrison, Mrs. Donna D. James and Miss Kathleen S. Williams and the excellent technical assistance of Ms. Silke Hammers. They would also like to thank Dr. Young Yang , R.W. Johnson Pharmaceutical Research Institute. La Jolla. CA, for providing anti-proteasome mAb and rabbit anti- delta serum. This Investigation was supported by PHS grant CA67108 awarded by the National Cancer Institute. DHHS.
Received 29 July, revised, accepted for publication 1 5 September 1997
Copyfight 0 Munkrgaard 1998 Tissue Antigens. ISSN 0001-2815
Tissue Antigens 1998: 51: 1 2 9 1 4 0 Printed in Denmark. All rights reserved
Development and characterization of mouse anti-human LMP2, LMP7, TAPl and TAP2 monoclonal antibodies
Abstract: Low molecular mass polypeptides &MF) 2 and LMP7 and trans- porter associated with antigen processing (TAP) subunits TAPl and TAP2 play a crucial role in antigen processing and cell surface expression of HLA class I molecules. Since monoclonal antibodies (mAb) to these molecules will facilitate the analysis of their expression, structure and function in normal and transformed cells, in the present study we have developed these reagents. Specifically anti-LMP2 and LMP7 mAb were generated €rom BALB/c mice immunized with specific peptides, and anti-TAP1 and TAP2 mAb from BALB/c mice immunized with respective recombinant proteins. mAb VF101-39F7 and VF101-39G5 were shown to be specific for LMPP, mAb VF103-5D5 and VF103-8C2 for LMP7, mAb VF108-1B3 and VF108-12D6 for TAPl and mAb VF118-1E4 and VF118-2C5 for TAP2, since they reacted specifically with the corresponding immunogens in ELISA and with the corresponding LMP and TAP subunits when tested in Western blotting with human lymphoid cell extracts. Furthermore, the mAb immunoprecipitated components with the characteristic electrophoretic mobility from lymphoid cells. Both anti-LMP and anti-TAP mAb stained keratinocytes and infilt- rating lymphocytes in frozen and formalin-fixed, paraffin embedded sections of normal skin in indirect immunoperoxidase reactions. Furthermore, all the mAb except mAb VF103-5D5 stained the cytoplasm of lymphoid cells in an intracytoplasmic staining reaction. The specificity and reactivity pattern of the mAb we have characterized indicate that they will be valuable reagents to analyze the cellular expression and tissue distribution of LMP and TAP subunits.
HLA class I molecules play a crucial role in the cellular immune re- sponse by presenting 8-9 amino acid long peptides to cytotoxic T lymphocytes (1). These peptides are generated mostly, although not exclusively from endogenous proteins and loaded onto Pz-micro- globulin (P2m) associated HLA class I heavy chains by the compon- ents of the antigen processing machinery (2,3). Among these compon- ents are the low molecular mass polypeptide (LMP) 2 and LMW, and
Authors' amllatlons:
R. Kishore'. D.J. Hicklin'. D.V. Deliaratta'. U. Golde'. T. Kageshita'. 6. Seltge?. S. krrone'
'Department of Microbiology and Immunoiogy, New York Medical College.Valhalla. NY-10595.
'Department of Internal Medicine, Division of HematologyjOncology, Langenbeckstr. 1. Johannes Gutenberg University. Mainz. Germany
Cornrpondsnce to: Dr. Soldano Ferrone Department of Microbiology
and lmmunologv New York Medical College Valhaila, NY 10595 Tel: (914) 594-4175 Fax: (914) 5944176 E-mail: [email protected]
129

Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
transporter associated with antigen processing (TAP) 1 and TAP2 (3, 4). The homologous 21 kD LMP2 and 23 kD LMW proteins are sub- units of the multicatalytic protease complex or 20s proteasome (5- 7). Incorporation of LMp2 and/or LMP7 into the proteasome complex increases the amount of peptides containing carboxyl-terminal basic or hydrophobic amino acid residues which preferentially bind to HLA class I molecules (8-10). The -7OkD TAPl and the -76kD TAP2 subunits form the TAP heterodimer, which in association with the 48kD glycoprotein tapasin (ll), transports antigenic peptides from the cytosol to newly synthesized HLA class I heavy chain-P2m com- plexes in the endoplasmic reticulum (ER) (12-16).
Analysis of the expression and functional properties of LMpZ, LMP7, TAPl and TAP2 molecules in normal and transformed cells has been hampered by the limited availability of the corresponding antibodies. Only a few anti- LMPB, LMP7, TAPl and TAP2 sera have been described in the literature (for review, see 17). They have been developed by immunizing rabbits with synthetic peptides or truncated recombinant proteins. These reagents have been useful to characterize LMP and TAP expression in normal and transformed cells and to analyze the association of TAPl and TAP2 with HLA class I alleles (18). However, like all conventional antisera, these reagents are available in limited amounts and are difficult to stan- dardize because of differences in antibody content among bleedings obtained from an immunized animal and also amongst animals. An additional major limitation of anti-LMPS, anti-LMW, anti-TAP1 and anti-TAP2 xenoantisera is their frequent contamination with anti- bodies to unrelated antigens, mostly of unknown nature. This con- tamination requires the purification of anti-LMP2, LMP7, TAPl and TAP2 antibodies by affinity chromatography on the corresponding antigens in order to increase their specificity. This procedure further limits the amount of antibodies available for binding and immuno- chemical assays. Lastly, because of their polyclonal nature, affinity purified rabbit antibodies are not the reagents of choice for intra- cellular staining of cells.
Although all these limitations can be overcome by the use of mAb, only a small number of these reagents have been developed. To the best of our knowledge, no anti-human LMP2 and LMW mAb have been reported. Only three anti-TAP1 mAb (1%21) and two anti-TAP2 mAb (19, 21) have been reported in the literature. More- over, details about their development and characterization are scanty. The reactivity pattern with frozen tissue sections and with intracellular protein has been described only for one anti-TAP1 mAb (20). No information is available about the ability of mAb to stain formalin-fixed tissue sections. These questions have been addressed in the present study, which describes the development and characterization of a panel of anti-LMP2, anti-LMP7, anti-TAP1 and anti-TAP2 mAb.
Material and methods
Animals
Eight to twelve week old female BALBk mice were purchased from Charles River Breeding Laboratory, Wilmington, MA.
Synthetic peptides
The LIvlP2, LMP7, TAPl and TAP2 specific peptides, which repre- sent C-terminal amino acid residues 207-234, 308-326, 777-795 and 667-704 of LMP2, LMP7, TAPl and TAP2, respectively, were syn- thesized and purified as described previously (17, 22, 23).
Synthetic oligonucleotide primers and recombinant fusion
proteins
The LMP2 forward (1-24) and reverse (623-641) primers and the LMP7 forward (1-21) and reverse (779-800) primers whose nucleotide sequences have been reported elsewhere (22), the TAPl forward
3' and reverse (2433-2465) 5'-CTC TAG AGG TTA TTC TGG AGC ATC TGC AGG AGG-3' primers and the TAP2 forward (975-1006)
reverse (2125-2161) 5'-ATG AAT TCC TTA CTG CTG AAC CAG GCG GGA ATA GAG G-3' primers were synthesized on an Applied Biosystems 392 DNA/RNA synthesizer (Applied Biosystems, Foster City, CA). Numbers in brackets following each primer indicate the corresponding nucleotide numbers for the sequence of LMP2, LMP7, TAPl and TAP2 cDNA starting at the ATG initiation codon. Cloning and expression of Lh4P2 and LMW recombinant proteins have been described elsewhere (22). Truncated TAPl and TAP2 cDNA were synthesized by using Superscript Reverse Transcriptase (Gibco-BRL, Gaithersburg, MD) according to the manufacturer's instructions from mRNA. The latter was isolated from cultured human melanoma cells Colo 38 with a Micro-fast Track kit (Invitrogen Inc., San Diego, CA) following the manufacturer's instructions. Amplification of TAPl and TAP2 cDNA, cloning and expression of truncated TAPl and TAP2 recombinant protein fused to thioredoxin (Trx) were carried out fol- lowing the methodology utilized to clone and express LMP2 and LMP7 recombinant proteins (22).
(1501-1528) 5'-GGG ATC CTG TGA CCA GTG GGG CTG TAA GC-
5'-CGG ATC CGA AGT GCT TCG GGA GAT CCA GGA TG -3' and
Cell lines and tissues
Mouse myeloma cells P3-X63-Ag8.653 and cultured human B lymph- oid cells LG2, Raji and T1 and Burkitt lymphoma cell Namalwa which express LMP2, LMP7, TAPl and TAPZ, and human TXB lympho-
130 Tissue Antigens 1998: 51: 129-140
Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
VF 103-SDS
VF103-8C2 I
mAb F I I
0.00 1.20 2.40
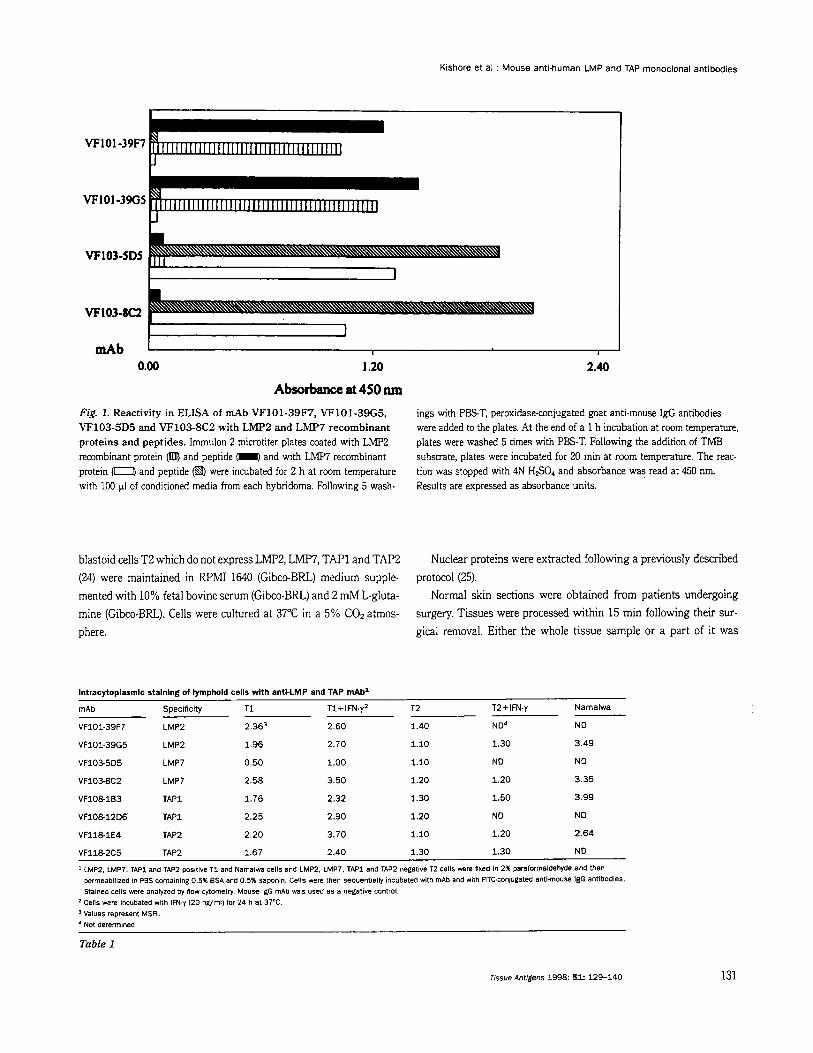
Absorbance at 450 nm Fig. 1. Reactivity in ELISA of mAb VF101-39F7, VF101-39G5, VF103-5D5 and VF103-8C2 with LMP2 and LMP7 recombinant proteins and peptides. Immulon 2 microtiter plates coated with LMP2 recombinant protein Cmm, and peptide (I and with LMP7 recombinant protein were incubated for 2 h at room temperature with 100 p1 of conditioned media from each hybridoma. Following 5 wash-
ings with P E T , peroxidase-conjugated goat anti.mouse IgG antibodies were added to the plates. At the end of a 1 h incubation at room temperature, plates were washed 5 times with PBS-T. Following the addition of TMB substrate, plates were incubated for 20 min at rcmm temperature. The reac- tion was stopped with 4N HSO, and absorbance was read at 450 nm. Results are expressed as absorbance units.
and peptide
blastoid cells T2 which do not express LMF'2, LMp7, TAPl and TAP2 (24) were maintained in WMI 1640 (Gibco-BRL.) medium supple- mented with 10% fetal bovine serum (Gibco-BRL) and 2 mM L-gluta- mine (Gibco-BRL). Cells were cultured at 37°C in a 5% COz atmos- phere.
Nuclear proteins were extracted following a previously described protocol (25).
Normal skin sections were obtained from patients undergoing surgery. Tissues were processed within 15 min following their sur- gical removal. Either the whole tissue sample or a part of it was
lntracytoplasmlc staining of lymphold cells with antl-LMP and TAP mAbl
mAb Specificity T i T I + IFN-y2 T2 T2+IFN-y Namalwa
VF101-39R LMP2 2.363 2.60 1.40 ND4 ND
VFlOl-39G5 LMP2 1.96 2.70 1.10 1.30 3.49
VF1035D5 LMP7 0.50 1.00 1.10 ND ND
VF1038C2 LMP7 2.58 3.50 1.20 1.20 3.35
VFlOElB3 TAPl 1.76 2.32 1.30 1.50 3.99
VFlOE12D6 TAPl 2.25 2.90 1.20 ND ND
VFllElE4 TAP2 2.20 3.70 1.10 1.20 2.64
VFl182C5 TAP2 1.67 2.40 1.30 1.30 ND
LMP2. LMP7. TAPl and TAP2 positive T I and Namalwa cells and LMP2. LMP7, TAPl and TAP2 negative T2 cells were fixed in 2% paraforrnaidehyde and then permeabilized in PBS containing 0.5% BSA and 0.5% saponin. Cells were then sequentially incubated with mAb and with nTCconJugated anti-mOUSe IgG antibodies. Stained cells were analyzed by now cyiometry. Mouse IgG mAb was used as a negative control. Cells were incubated with IFN-y (20 numi) for 24 h at 37°C. Values represent MSFI. Not determined.
Table 1
Tissue Antigens 1998: 5 1 129-140 131
Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
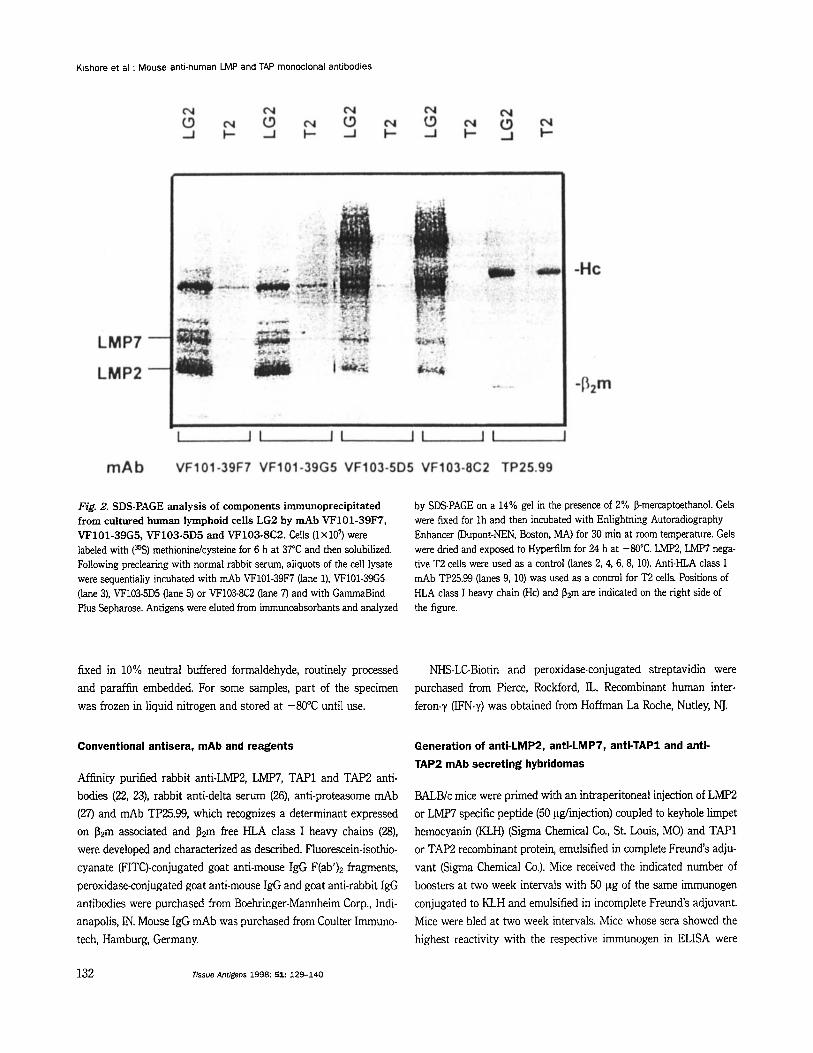
Fig. 2. SDS-PAGE analysis of components irnmunoprecipitated from cultured human lymphoid cells LG2 b y mAb VF101-39F7, VF101-39G5, VF103-5D5 and VF103-8C2. Cells ( lX107) were labeled with (%) methionindcysteine for 6 h at 37°C and then solubilized. Following preclearing with normal rabbit serum, aliquots of the cell lysate were sequentially incubated with mAb VF101-39F7 (lane l), VF101-39G5 (lane 3), VF103-5D5 (lane 5) or VF103-8C2 (lane 7) and with GammaBind Plus Sepharose. Antigens were eluted from immunoabsorbants and analyzed
by SDS-PAGE on a 14% gel in the presence of 2% P-mercaptoethanol. Gels were fixed for Ih and then incubated with Enlightning Autoradiography Enhancer @upont-NEN, Boston, MA) for 30 min at room temperature. Gels were dried and exposed to Hyperfilm for 24 h at -80°C. LMP2, LMW nega- tive T2 cells were used as a control (lanes 2, 4, 6, 8, 10). Anti-KLA class I mAb TP25.99 (lanes 9, 10) was used as a control for T2 cells. Positions of KLA class I heavy chain (Hc) and Pzm are indicated on the right side of the figure.
fixed in 10% neutral buffered formaldehyde, routinely processed NHS-LC-Biotin and peroxidase-conjugated streptavidin were and paraffin embedded. For some samples, part of the specimen purchased from Pierce, Rockford, IL. Recombinant human inter- was frozen in liquid nitrogen and stored at -80°C until use. feron-y (IFN-y) was obtained from Hoffman La Roche, Nutley, NJ.
Conventional antisera, mAb and reagents
Affinity purified rabbit anti-LMF'2, LMP7, TAPl and TAP2 anti- bodies (22, =), rabbit anti-delta serum (26), anti-proteasome mAb (27) and mAb TP25.99, which recognizes a determinant expressed on Pzm associated and pgm free HLA class I heavy chains (B), were developed and characterized as described. Fluorescein-isothio- cyanate (F1TC)-conjugated goat anti-mouse IgG F(ab')z fragments, peroxidase-conjugated goat anti-mouse IgG and goat anti-rabbit IgG antibodies were purchased from Boehringer-Mannheim Corp., Indi- anapolis, IN. Mouse IgG mAb was purchased from Coulter Immuno- tech, Hamburg, Germany.
Generation of anti-LMP2, anti-LMP7, anti-TAP1 and anti-
TAP2 mAb secreting hybridomas
BALBk mice were primed with an intraperitoneal injection of LMP2 or LMP7 specific peptide (50 pglinjection) coupled to keyhole limpet hemocyanin (KLK) (Sigma Chemical Co., St. Louis, MO) and TAPl or TAP2 recombinant protein, emulsified in complete Freund's adju- vant (Sigma Chemical Co.). Mice received the indicated number of boosters at two week intervals with 50 pg of the same immunogen conjugated to KLH and emulsified in incomplete Freund's adjuvant. Mice were bled at two week intervals. Mice whose sera showed the highest reactivity with the respective immunogen in ELISA were
132 Tissue Antigens 1998: 51: 129-140
Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
boosted intravenously with 50 pg of immunogen five days before splenwytes were harvested and fused to mouse myeloma cells P3- X63-Ag8.653. Hybridization and subcloning were performed using standard techniques (29). Subtyping of mAb was performed with the Isostrip kit (Boehringer-Mannheim Corp.) following the manu-
facturer's instructions. mAb were purified from ascites using a Hi Trap Protein G column (Pharmacia Biotech Inc., Piscataway, NJ) following the manufacturer's instructions. mAb were biotinylated using NHS-LC-Biotin (Pierce) following the manufacturer's instruc- tions.
Binding Assays
ELISA was performed in Immulon 2 microtiter plates (Dynatech Laboratories, Chantilly, VA) following the methodology described elsewhere (22, 23). Cross-blocking assays to map antigenic deterrni- nants recognized by mAb were performed by incubating LMP2, LMW, TAP1 and TAP2 recombinant proteins for 2 h at room tem- perature in microtiter plates coated with one of the corresponding mAb (5 pg/well). Following 5 washings with phosphate buffered salind0.05 YO Tween-20 (PBS-T), captured recombinant proteins were incubated for 2 h at room temperature with a biotinylated mAb (50 ng/well). Plates were then washed 5 times with PBS-T and incubated for 1 h at room temperature with peroxidase-conjugated streptavidin diluted 12500 (100 pVwel1). Following 5 additional washings with PEE-T, plates were incubated with 100 pl/well of Tetramethylbemidine peroxidase substrate (Kirkegaard and Perry Labs., Gaithersburg, MD) and H202 solution (1:l). The reaction was stopped with 4N H S 0 4 after a 20 min incubation at room tempera- ture. Absorbance at 450 nm was read with a Microplate autoreader (€30-Tek Instrument Inc., Winooski, VT). Results are expressed as absorbance units.
lntracytoplasmic staining
Intracytoplasmic staining of cells with mAb was performed using a previously described protocol (30). Briefly, cells were fixed in 2% paraformaldehyde for 15 min at 4°C and then permeabilized in PBS containing 0.5% bovine serum albumin (BSA) and 0.5% saponin (Sigma Chemical Co.). Cells (1X106) were incubated with an excess of mAb for 30 min at 4°C. Following two washings with ice cold PBS containing 0.5% BSA (F'BS/BSA), cells were incubated with FITC-conjugated goat anti-mouse I ~ G antibodies in the dark for an additional 30 min at 4°C. Following two washings with PBSIBSA, Cells Were analyzed O n a flow cytometer (Coulter Immunotech, Ham- burg, Germany). Mouse IgG mAb was used as a negative control. Results are expressed as mean specific fluorescence intensity (MSFI), which was calculated by dividing the mean fluorescence intensity of cells stained with test mAb by the mean fluorescence intensity of cells incubated with control IgG mAb.
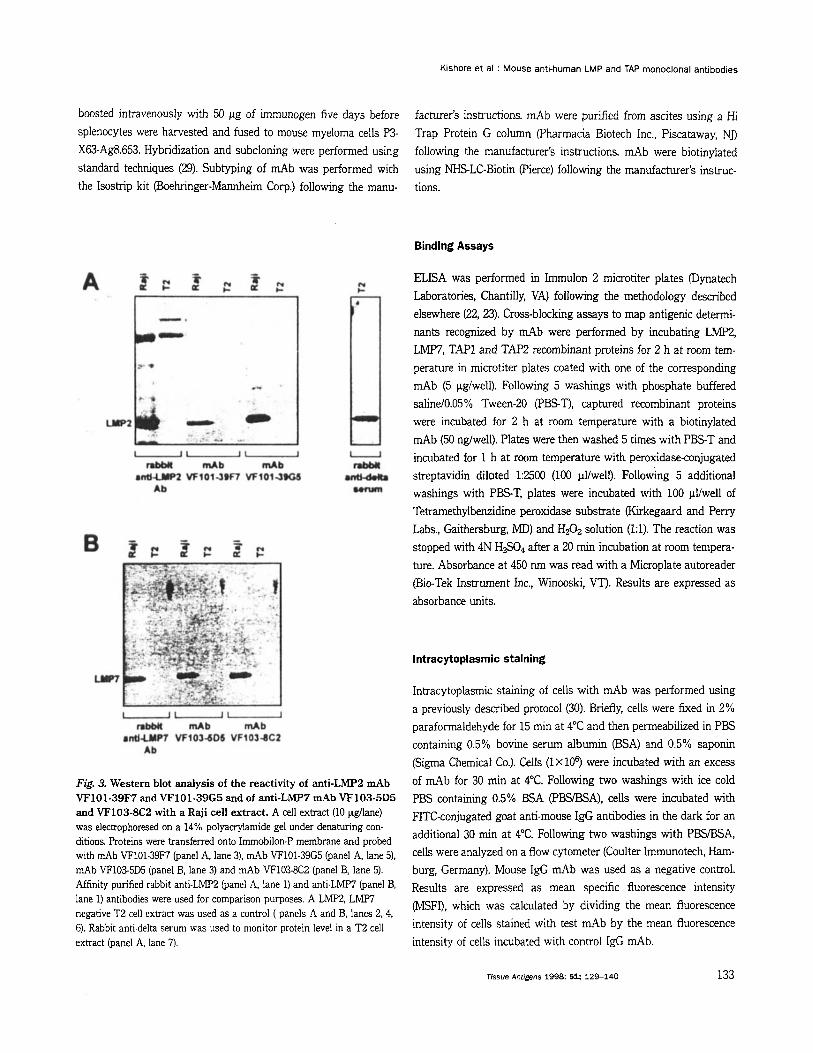
Fig. 3. Western blot analysis of h e reactivity of anti-^^^^ d b VF101-39F7 and VF101-39G5 and of anti-LMP7 mAb VF103-5D5 and VF103-8C2 with a Raji cell extract. A cell extract (10 pdlane) was eleckophoresed on a 14% polyauylamide gel under denaturing con- ditions. Proteins were transferred onto Immobilon-P membrane and probed with d b VF101-39F7 (panel A, lane 3), mAb VF101-39G5 (panel A, lane 5), mAb VF103-5D5 (panel B, lane 3) and mAb VF103-8C2 (panel B, lane 5). Affinity purified rabbit anti-LMF'2 (panel A, lane 1) and anti-~M€'7 (panel lane 1) antibodies were used for comparison purposes. A LMP2, LMF7 negative T2 cell extract was used as a control ( panels A and B, lanes 2.4, 6). Rabbit anti-delta serum was used to monitor protein level in a ~2 cell extract (panel A, lane 7).
Tissue Antigens 1998: 51: 129-140 133
Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
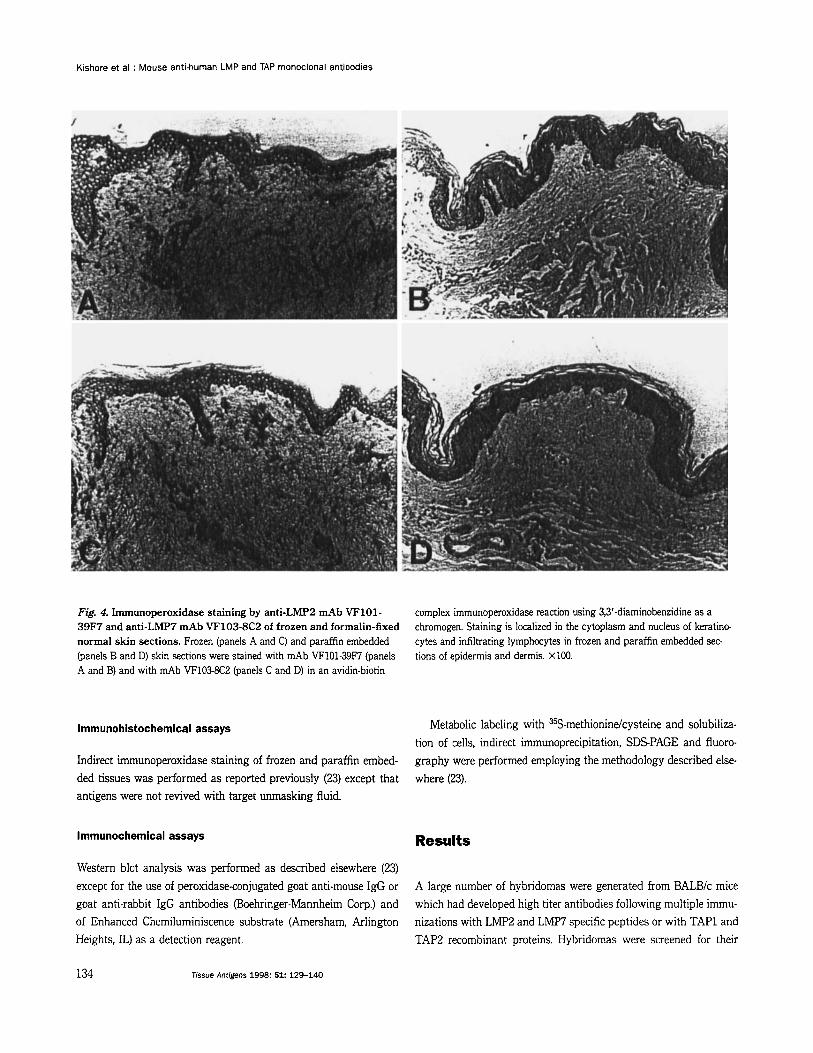
Fig. 4. Immunoperoxidase staining by anti-LMP2 rnAb VF101- 39F7 and anti-LMP7 mAb VF103-8C2 of frozen and formalin-fixed normal skin sections. Frozen (panels A and C) and paraffin embedded (panels B and D) skin sections were stained with mAb VF101-39F7 (panels A and B) and with mAb VF103-8C2 (panels C and D) in an avidin-biotin
complex immunoperoxidase reaction using 3,3'-diaminobenzidine as a chromogen. Staining is localized in the cytoplasm and nucleus of keratino- cytes and infiltrating lymphocytes in frozen and paraffin embedded sec- tions of epidermis and dermis. X100.
lmmunohistochemfcal assays
Indirect immunoperoxidase staining of frozen and paraffin embed- ded tissues was performed as reported previously (23) except that antigens were not revived with target unmasking fluid.
Metabolic labeling with 35S-methionine/cysteine and solubiliza- tion of cells, indirect immunoprecipitation, SDS-PAGE and fluoro- graphy were performed employing the methodology described else- where (23).
lmmunochemical assays Results
Western blot analysis was performed as described elsewhere (23) except for the use of peroxidase-conjugated goat anti-mouse IgG or goat anti-rabbit IgG antibodies (Boehringer-Mannheim Corp.) and of Enhanced Chemiluminiscence substrate (Amersham, Arlington Heights, IL) as a detection reagent.
A large number of hybridomas were generated from BALB/c mice which had developed high titer antibodies following multiple immu- nizations with LMP2 and LMW specific peptides or with TAP1 and TAP2 recombinant proteins. Hybridomas were screened for their
134 Tissue Antigens 1998 51: 129-140
Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
specific reactivity in ELISA with LMp2, LMP7, TAPl and/or TAP2 recombinant proteins.
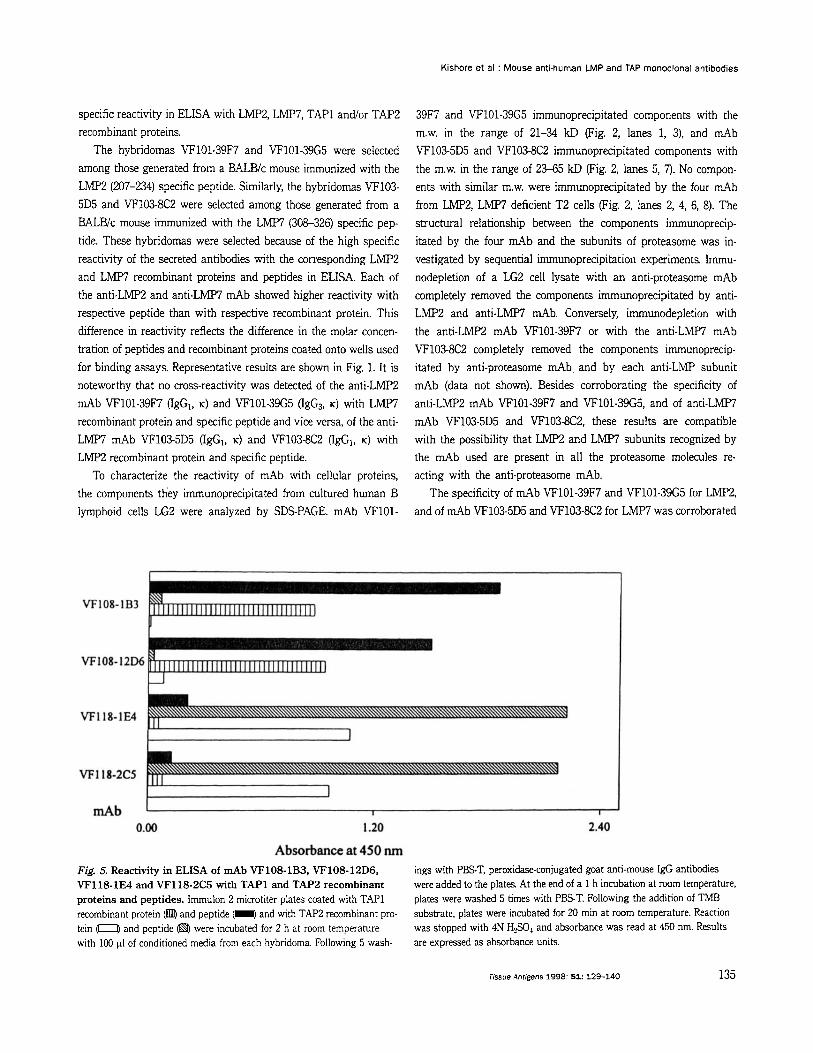
The hybridomas VF101-39F7 and VF101-39G5 were selected among those generated from a BALBlc mouse immunized with the LMp2 (207-234) specific peptide. Similarly, the hybridomas VF103- 5D5 and VF103-8C2 were selected among those generated from a BALBlc mouse immunized with the LMP7 (308-326) specific pep- tide. These hybridomas were selected because of the high specific reactivity of the secreted antibodies with the corresponding LMP2 and LMP7 recombinant proteins and peptides in ELISA. Each of the anti-LMPi! and anti-LMP7 mAb showed higher reactivity with respective peptide than with respective recombinant protein. This difference in reactivity reflects the difference in the molar concen- tration of peptides and recombinant proteins coated onto wells used for binding assays. Representative results are shown in Fig. 1. It is noteworthy that no cross-reactivity was detected of the anti-LMP2 mAb VF101-39F7 (IgGI, K) and VF101-39G5 (IgG3, K) with LMP7 recombinant protein and specific peptide and vice versa, of the anti- LMP7 mAb VF103-5D5 (IgG1, K) and VF103-8C2 (IgG,, K) with LMP2 recombinant protein and specific peptide.
To characterize the reactivity of mAb with cellular proteins, the components they immunoprecipitated from cultured human B lymphoid cells LG2 were analyzed by SDS-PAGE. mAb VF101-
39F7 and VF101-39G5 immunoprecipitated components with the m.w. in the range of 21-34 kD (Fig. 2, lanes 1, 3), and mAb VF103-5D5 and VF103-8C2 immunoprecipitated components with the m.w. in the range of 23-65 kD (Fig. 2, lanes 5, 7). No compon- ents with similar m.w. were irnmunoprecipitated by the four mAb from LMp2, LMP7 deficient T2 cells (Fig. 2, lanes 2, 4, 6, 8). The structural relationship between the components immunoprecip- itated by the four mAb and the subunits of proteasome was in- vestigated by sequential immunoprecipitation experiments. Immu- nodepletion of a LG2 cell lysate with an anti-proteasome mAb completely removed the components immunoprecipitated by anti- LMP2 and anti-LMP7 mAb. Conversely, immunodepletion with the anti-LMP2 mAb VF101-39F7 or with the anti-LMP7 mAb VF103-8C2 completely removed the components immunoprecip- itated by anti-proteasome mAb and by each anti-LMp subunit mAb (data not shown). Besides corroborating the specificity of anti-LMP2 mAb VF101-39F7 and VF101-39G5, and of anti-LMP7 mAb VF103-5D5 and VF103-8C2, these results are compatible with the possibility that LMP2 and LMP7 subunits recognized by the mAb used are present in all the proteasome molecules re- acting with the anti-proteasome mAb.
The specificity of mAb VF101-39F7 and VF101-39G5 for LMp2, and of mAb VF103-5D5 and VF103-8C2 for LMP7 was corroborated
VF 108- 1B3
VF108- 12D6
VF 1 18- 1E4
VF 1 18-2C5 1
mAb I I
0.00 1.20 2.40
Absorbance at 450 nm Fig. 5. Reactivity in ELISA of d b VF108-1B3, VF108-12D6, VF118-1E4 and VF118-2C5 with TAPl and TAP2 recombinant proteins and peptides. Immulon 2 microtiter plates coated with TAP1 recombinant protein (@ and peptide (I and with TAP2 recombinant pro- tein with 100 pl of conditioned media from each hybridoma. Following 5 wash-
ings with PEST, peroxidase-conjugated goat anti-mouse IgG antibodies were added to the plates. At the end of a 1 h incubation at room temperature, plates were washed 5 times with PEST. Following the addition of TMB substrate, plates were incubated for 20 min at room temperature. Reaction was stopped with 4N H2S04 and absorbance was read at 450 nm. Results are expressed as absorbance units.
and peptide @$ were incubated for 2 h at room temperature
Tissue Anrigens 1998: 51: 129-140 135

Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
affinity purified rabbit anti-LMP2 and anti-LMW antibodies with Raji and T2 cell extracts (Fig. 3A and B, lanes 1, 2).
Cross-blocking experiments mapped the antigenic determinants recognized by the two anti-LMP2 and by the two anti-LMF'7 mAb. Each of the two anti-LMP7 mAb could bind to the LMF7 recombin- ant protein captured by the other mAb, but did not bind to the recombinant protein captured by the homologous mAb. These re- sults suggest that the two anti-LMW mAb recognize distinct and spatially distant determinants on the LMW recombinant protein. The binding of each of the two anti-LMP2 mAb to LMP2 recombin- ant protein captured by the other mAb was reduced by about 60%, suggesting that the anti-LIW2 mAb VF101-39M and VF101-39G5 recognize distinct, but spatially close antigenic determinants.
Anti-LMP2 mAb VF101-39F7 and VF101-39G5 stained the cyto- plasm of T1 and Namalwa cells in an intracellular staining reaction. However, out of the two anti-LMF'7 mAb, only mAb VF103-8C2 was able to stain the cytoplasm of T1 cells. The staining is specific, since the three mAb did not stain T2 cells which do not express LMP2 and LMF'7. Furthermore, the staining intensity was enhanced in T1 cells following treatment with JFN-7, but not in T2 cells. Representative results of intracellular staining are shown in Table 1.
In indirect immunoperoxidase reactions, anti-LMP2 as well as anti-LMP7 mAb stained keratinocytes and infiltrating lymphocytes in frozen as well as formalin-fixed, paraffin embedded tissue sec- tions of normal skin. The staining intensity of frozen as well as paraffin embedded tissue sections by each of the four mAb was similar. Staining of frozen sections was predominantly cytoplasmic (Fig. 4, panels A, C), while that of paraffin embedded tissue sections was localized both in cytoplasm and in nucleus of cells (Fig. 4, panels B, D). The presence of and LMp7 in the nuclei was
Fig. 6. Western blot analysis of the reactivity of mAb VF101- 1B3, VF108-12D6, VF118-1E4 and VF118-2C5 with a Raji cell extract. A cell extract (10 pgflane) was electrophoresed on a 12% p01y- aaylamide gel under denaturing conditions. Proteins were transferred onto Immobilon-P membrane and probed with mAb VF108-1B3 (panel A, lane 3), mAb VF10812D6 (panel A, lane 5). mAb VF1181E4 (panel B, lane 3) and mAb VF118-2C5 (panel B, lane 5). Affinity purified rabbit anti-TAP1 and anti-TAP2 antibodies (panels A and B, lane 1) were used for comparison purposes. A TAPI, TAP2 negative T2 cell extract was used as a control. Rabbit anti-delta serum was used to monitor protein level in a T2 cell extract (panel A, lane 7).
by the results of Western blot analysis of their reactivity with a LMP2, LMP7 positive Raji cell extract (Fig. 3 A and B). mAb VF101- 39F7 and VF101-39G5 recognize a single 21 kD protein (Fig. 3A, lanes 3,5) and mAb VF103-5D5 and VF103-8C2 recognize a single 23 kD protein (Fig. 3B, lanes 3, 5) corresponding to LMP2 and LMW, respectively. The reactivity is specific, since the four mAb recognized no component in a T2 cell extract (Fig. 3A and B, lanes 4, 6). The reactivity patterns of the four mAb resemble those of
corroborated by the reactivity in Western blotting of anti-LMP2 and anti-LMP7 mAb with a nuclear extract from Raji cells (data not shown).
The hybridomas VF1081B3 and VF10812D6 were selected among those generated from a BALBlc mouse immunized with TAPl recombinant protein, and the hybridomas VF118-1E4 and VF1182C5 were selected among those generated kom one BALBlc mouse immunized with TAP2 recombinant protein. The hybridomas were selected on the basis of high specific reactivity of the secreted antibodies with TAPl and TAP2 recombinant proteins and peptides in ELISA. Each of anti-TAP1 and anti-TAP2 mAb showed higher reactivity with respective peptide than with respective recombinant protein. This difference in reactivity reflects the difference in the molar concentration of peptides and recombinant proteins coated onto wells used for binding assays. Representative results are shown in Fig. 5.
136 Jissue Antigens 1998: 51: 129-140
Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
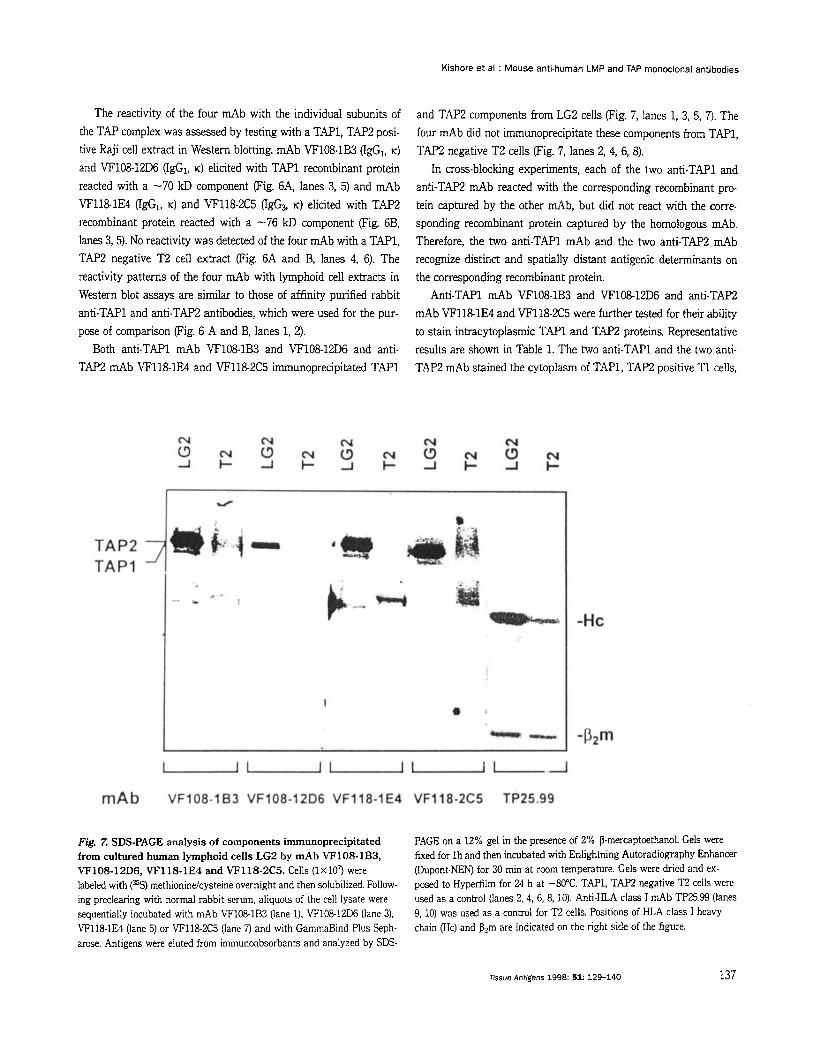
The reactivity of the four mAb with the individual subunits of the TAP complex was assessed by testing with a TAP1, TAP;! posi- tive Raji cell extract in Western blotting. mAb VF108-1B3 (IgGI, K) and VF10812D6 (IgG1, K) elicited with TAPl recombinant protein reacted with a -70 kD component (Fig. 6A, lanes 3, 5) and mAb VF118-1E4 (lgG1, K) and VF118-2C5 0 6 3 , K) elicited with TAP2 recombinant protein reacted with a -76 kD component (Fig. 6B, lanes 3, 5). No reactivity was detected of the four mAb with a TAP1, TAP2 negative T2 cell extract (Fig. 6A and B, lanes 4, 6). The reactivity patterns of the four mAb with lymphoid cell extracts in Western blot assays are similar to those of affinity purified rabbit anti-TAP1 and anti-TAP2 antibodies, which were used for the pur- pose of comparison (Fig. 6 A and B, lanes l, 2).
Both anti-TAP1 mAb VF108-1B3 and VF108-12D6 and anti- TAP2 mAb VF118-1E4 and VF1182C5 immunoprecipitated TAPl
and TAP2 components from LG2 cells (Fig. 7, lanes 1, 3, 5, 7). The four mAb did not immunoprecipitate these components from TAP1, TAP;! negative T2 cells (Fig. 7, lanes 2, 4, 6, 8).
In cross-blocking experiments, each of the two anti-TAP1 and anti-TAP;! mAb reacted with the corresponding recombinant pro- tein captured by the other mAb, but did not react with the corre- sponding recombinant protein captured by the homologous mAb. Therefore, the two anti-TAP1 mAb and the two anti-TAP2 mAb recognize distinct and spatially distant antigenic determinants on the corresponding recombinant protein.
Anti-TAP1 mAb VF108-1B3 and VF108-12D6 and anti-TAP2 mAb VF118-1E4 and VF118-2C5 were further tested for their ability to stain intracytoplasmic TAPl and TAP2 proteins. Representative results are shown in Table 1. The two anti-TAP1 and the two anti- TAP;! mAb stained the cytoplasm of TAPl, TAP;! positive T1 cells,
Fig. Z SDS-PAGE analysis of components immunoprecipitated from cultured human lymphoid cells LG2 by mAb VF108-1B3, VF108-12D6, VF118-1E4 and VF118-2C5. Cells (1x107 were labeled with (%) methionindcysteine overnight and then solubilized. Follow- ing preclearing with normal rabbit serum, aliquots of the cell lysate were sequentially incubated with mAb VF108-1B3 (lane 1). VF108-12D6 (lane 3). VF118-1E4 (lane 5) or VF118-2C5 (lane 7) and with GammaBind Plus Seph- arose. Antigens were eluted from immunoabsorbants and analyzed by SDS-
PAGE on a 12% gel in the presence of 2% P-mercaptoethanol. Gels were fixed for l h and then incubated with Enlighhing Autoradiography Enhancer (Dupont-NEN) for 30 min at rmm temperature. Gels were dried and ex- posed to Hyperfilm for 24 h at -80°C. TAP1, TAP2 negative T2 cells were used as a control (lanes 2, 4, 6, 8, 10). Anti-HLA class I mAb TP25.99 (lanes 9, 10) was used as a control for T2 cells. Positions of HLA class I heavy chain (Hc) and P?rn are indicated on the right side of the figure.
Tissue Antigens 1998: 51: 129-140 137
Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
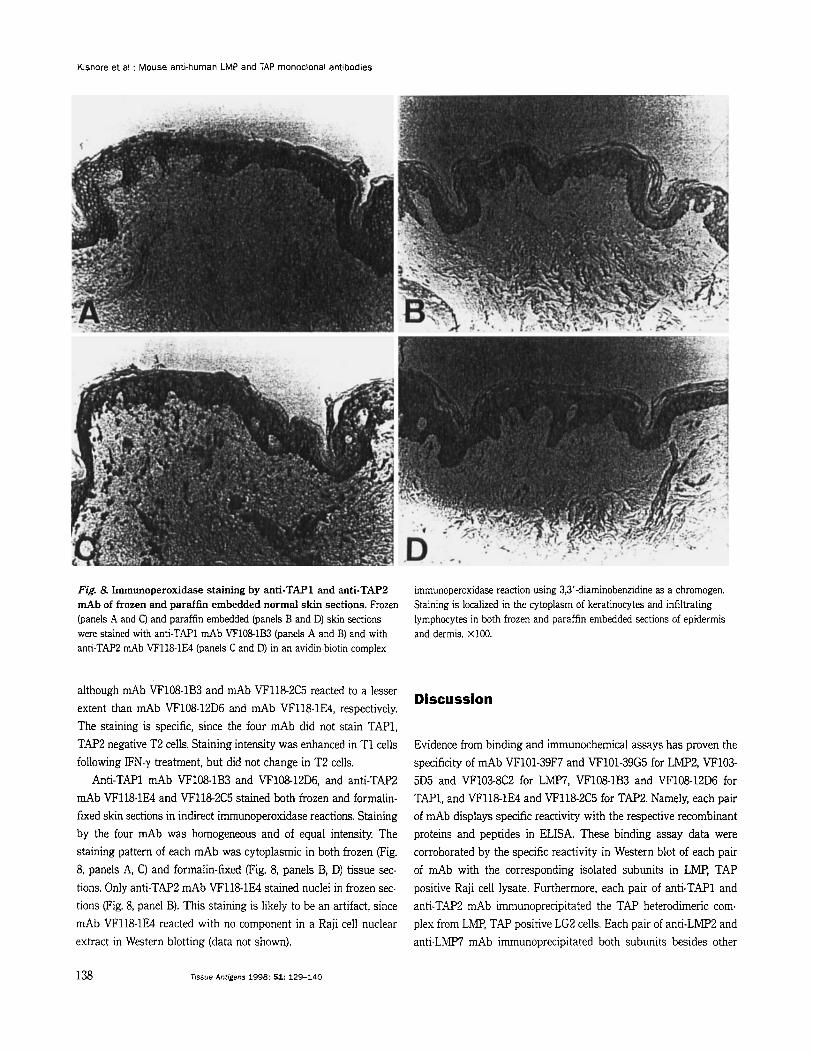
Fig. 8. Immunoperoxidase staining by anti-TAP1 and anti-TAP2 mAb of frozen and paraflin embedded normal skin sections. Frozen (panels A and C) and paraffin embedded (panels B and D) skin sections were stained with anti-TAP1 mAb VF1081B3 (panels A and B) and with anti-TAP2 mAb VF118-1E4 (panels C and D) in an avidin-biotin complex
immunoperoxidase reaction using 3,3’-diaminobenzidine as a chromogen. Staining is localized in the cytoplasm of keratinocytes and infiltrating lymphocytes in both frozen and paraffin embedded sections of epidermis and dermis. X100.
although mAb VF108-1B3 and mAb VF118-2C5 reacted to a lesser extent than mAb VF108-12D6 and mAb VF118-1E4, respectively. Discussion
The staining is specific, since the four mAb did not stain TAP1, TAP2 negative T2 cells. Staining intensity was enhanced in T1 cells following IFN-y treatment, but did not change in T2 cells.
Anti-TAP1 mAb VF108-1B3 and VF10812D6, and anti-TAP2 mAb VF118-1E4 and VF118-2C5 stained both frozen and formalin- fixed skin sections in indirect immunoperoxidase reactions. Staining by the four mAb was homogeneous and of equal intensity. The staining pattern of each mAb was cytoplasmic in both frozen (Fig. 8, panels A, C) and formalin-fixed (Fig. 8, panels B, D) tissue sec- tions. Only anti-TAP2 mAb VF118-1E4 stained nuclei in frozen sec- tions (Fig. 8, panel B). This staining is likely to be an artifact, since mAb VF118-1E4 reacted with no component in a Raji cell nuclear extract in Western blotting (data not shown).
Evidence from binding and immunochemical assays has proven the specificity of mAb VF101-39F7 and VF101-39G5 for LMP2, VF103- 5D5 and VF103-8C2 for LMW, VF108-1B3 and VF108-12D6 for TAP1, and VF118-1E4 and VF118-2C5 for TAP2 Namely, each pair of mAb displays specific reactivity with the respective recombinant proteins and peptides in ELISA. These binding assay data were corroborated by the specific reactivity in Western blot of each pair of mAb with the corresponding isolated subunits in LMP, TAP positive Raji cell lysate. Furthermore, each pair of anti-TAP1 and anti-TAP2 mAb immunoprecipitated the TAP heterodimeric com- plex from LMP, TAP positive LG2 cells. Each pair of anti-LMP2 and anti-LMP7 mAb immunoprecipitated both subunits besides other
138 Tissue Antigens 1998: 51: 129-140

Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
proteasome subunits from LG2 cells. The eight mAb neither reacted with a T2 cell extract in Western blots nor immunoprecipitated any component from LMP, TAP negative T2 cells.
It is noteworthy that anti-LMP2 and anti-LMP7 mAb elicited with respective C-terminal peptides recognize distinct antigenic de- terminants on the corresponding recombinant proteins. These re- sults indicate that at least two immunogenic determinants are ex- pressed on each of the peptides used as immunogens. Furthermore, the two anti- TAPl and the two anti-TAP2 mAb elicited with TAPl and TAP2 recombinant proteins, respectively, react with the corre- sponding C-terminal peptides in ELISA. These results are not unex- pected, since this portion of each TAP subunit is hydrophilic and, therefore, is likely to be most immunogenic.
Anti-LMP2 and anti-LMP7 mAb immunoprecipitated no com- ponents from a LG2 cell lysate immunodepleted with an anti-pro- teasome mAb. Conversely, the latter mAb immunoprecipitated no component from a LG2 cell lysate following immunodepletion with either anti-LMP2 or anti-LMW mAb. These observations suggest that anti-LMP2 and anti-LMP7 mAb co-precipitate other proteasome subunits, and that in B lymphoid cells LG2, most, if not all of the proteasome molecules contain LMP2 and LMW subunits. We have observed in ongoing experiments that LG2 cells express very low amounts of delta and MB1 proteins. This phenotype resembles IFN- y treated melanocytes and fibroblasts (unpublished data) and s u g gests that LG2 cells are in a state of constant activation.
In view of the high degree of homology between LMP2 and LMP7, it is noteworthy that no cross-reactivity was detected, both, in binding and immunochemical assays of anti-LMP2 mAb with LMW recombinant protein and peptide as well as with native pro- tein, and of anti-LMW mAb with LMP2 recombinant protein, pep-
References
1. Townsend AR, Gotch FM, Davey J. Cytotoxic T cells recognize fragments of the influenza nucleoprotein. cell 1985: 42: 45767.
proteasomes and antigen presentation. Nature 1992: 357: 375-9.
3. Glynne R, Powis SH, Beck S, Kelly A, Kerr LA, Trowsdale J. A proteasome-related gene between the two ABC transporter loci in the class 11 region of the human MHC. Nature 1991: 353: 3 5 7 a .
2. Goldberg AL, Rock KL. Proteolysis,
tide and native protein. Anti-LMP2 mAb and anti-LMP7 mAb are not likely to cross-react with delta and MB1 subunits of the pro- teasome, since SDS-PAGE analysis detected no component in the immunoprecipitate from T2 cells. Furthermore, Western blot analy- sis detected no reactivity of the anti-LMPZ and LMp7 mAb with a T2 cell extract. Proteasome subunits delta and MB1, which share 59 and 67% homology with LMP2 and LMW, respectively, have been shown previously to be expressed in T2 cells (4).
In immunohistochemical reactions, each pair of anti-LMP2, LMl'7, TAPl and TAP2 mAb stained both frozen and formalin- fixed, paraffin embedded tissue sections of normal skin. Staining by anti-TAP1 and TAP2 mAb was localized to the cytoplasm of keratinocytes and infiltrating lymphocytes in normal skin sections. These results are in agreement with the previously described local- ization of TAP in the ER and Golgi apparatus (15). In contrast, the two anti-LMP2 and the two anti-LMP7 mAb stained the cytoplasm as well as the nucleus of keratinocytes and infiltrating lymphocytes in normal skin sections. Nuclear expression of LMP2 and LMP7 is not unique to human keratinocytes, since it has also been reported in frozen sections of mouse thymus and liver (31). Moreover, nuclear localization of proteasome has been previously reported in different tissue sections of Halocynthia roretzi (32) and Drosophila melanogus- ter (33, 34) as well as in human hematopoietic cells (35). There is not much information about the tissue distribution of LMP and TAP in adults, their ontogeny and clinical significance. The mAb we have described are likely to facilitate experiments to obtain this information as well as implement retrospective studies to assess the frequency of abnormalities in transformed cells and their clinical significance.
4. Fklich W, Glynne RJ, Senger G, Sheer D, Trowsdale J. Proteasome components with reciprocal expression to that of the MHC- encoded LMl' proteins. Cum Biof 1994: 4: 769-76.
5. Trowsdale J, Hanson I, Mockridge I. Beck S, Townsend A, Kelly A. Sequences encoded in the Class I1 region of the MHC related to the 'ABC' superfamily of transporters. Nature 1990 348: 7414.
6. Yang Y, Waters m, Fruh K, Peterson PA. Proteasoms are regulated by interferon gamma: implications for antigen processing. R o c Natl Acad Sci U S A 1992 89 4928-32.
Trowsdale J, Ting JP Coordinate regulation of the human TAPl and LMP2 genes from a shared bidirectional promoter. Exp Med 1995: 181: 145%71.
7. Wright KL, White LC, Kelly A, Beck S,
lissue Antigens 1998: 51: 129-140 139

Kishore et al : Mouse anti-human LMP and TAP monoclonal antibodies
8. Dick LR, Aldrich C, Jameson Sc et al. Proteolytic processing of ovalbumin and - galactosidase by the proteasome to yield antigenic peptides. J Immunol1994: 152: 3884-94.
9. Momburg F, Roelse J, Howard JC, Butcher GW, Hammerling GJ, Neefjes J. Selectivity of MHC-encoded peptide transporters from human, mouse and rat. Nature 1994: 367 648-51.
10. Schumacher TN, Kantesaria DV, Heemels MT et al. Peptide length and sequence specificity of the mouse TAPlflAP2 translocator. J Exp Med 1994 179 533- 40.
11. Sadasivan B, Lehner PJ. Ortmann B, Spies T, Cresswell P. Roles for calreticulin and a novel glycoprotein, tapasin, in the interaction of MHC class I molecules with TAP. Immunity 1996: 5: 103-14.
12. Deverson EV, Gow IR, Coadwell WJ, Monaco JJ, Butcher GW, Howard JC. MHC class Il region encoding proteins related to the rnultidrug resistance family of transmembrane transporters. Nature 1990: 348 738-41.
protein genes in the rnurine MHC possible implications for antigen processing. Science 1990: 250 1723-6.
14. Spies T, Bresnahan M, Bahram S et al. A gene in the human major histocompatibility complex class II region controlling the class I antigen presentation pathway. Nature 1990: 348 744-7.
15. Kleijmeer MJ, Kelly A, Geuze HJ, Slot w, Townsend A, Trowsdale J. Location of MHC- encoded transporters in the endoplasmic reticulum and cisColgi. Nature 1992 357 342-4.
16. DeMars R. Rudersdorf R, Chang C et al. Mutations that impair a posttransaiptional step in expression of HLA-A and -B antigens. PYOC Nut1 Acad Sci US A 1985
13. Monaco JJ, Cho S, Attaya M. Transport
82: 8183-7. 17. Hicklin DJ, Kageshita T, Dellaratta DV,
Boccaletti V, Ferrone S. Defects in HLA Class I antigen presentation machinery in melanoma cells In: Maio M, ed. Immunology of Human Melanoma. Tumor - Host Interaction and Immunotherafg Amsterdam: 10s Press, 1996: 95411.
18. Neisig A, Wubbolts R, Zang X, Melief C, Neefjes J. Allele-specific differences in the interaction of MHC class I molecules with transporters associated with antigen processing. J Immunol1996: 156 3196- 206.
19. Meyer TH, Van Endert PM, Uebel S, Ehring B, Tampe R. Functional expression and purification of the ABC transporter complex associated with antigen processing (TAP) in insect cells. FEBS Left 1994: 351: 443-7.
Gatter KC, Harris AL. Loss of transporter in antigen processing 1 transport protein and major histocompatibility complex class I molecules in metastatic versus primary breast cancer. Cancer Res 1995: 55: 51914.
21. Nijenhuis M, Schmitt S, Armandola EA, Obst R, Brunner J, Hammerling GJ. Identification of a contact region for peptide on the TAP1 chain of the transporter associated with antigen processing. J Immunol1996: 156 2186-95.
Kageshita T and Ferrone S. Characterization of rabbit antisera elicited with human LMP2 and LMP7 specific peptides and recombinant proteins. Tissue Antzgens. In Press 1997.
23. Hicklin DJ, Kageshita T, Ferrone S. Development and characterization of rabbit antisera to human MHC-linked transporters associated with antigen processing. Tissue Antigens 1996: 48: 33- 46.
24. Salter RD, Alexander J, Levine F, Pious D, Cresswell F! Evidence for two transacting genes regulating HLA class I1 antigen expression. J lmmunol1985: 135: 4235-8.
25. Dignam JD, Lebovitz RM, Roeder RG. Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nuc& Acids Res 1983: 11: 1475-89.
26. Fruh K, Gossen M, Wang K, Bujard H, Peterson PA, Yang Y. Displacement of housekeeping proteasome subunits by MHC-encoded LMps: a newly discovered mechanism for modulating the multicatalytic proteinase complex. EMBO J 1994: 13: 3236-44.
20. Kaklamanis L, Leek R, Koukourakis M,
22. Dellaratta DV, Hicklin DJ, Kishore R,
27. Kaltoft MB, Koch C, Uerkvitz W, Hendil KB. Monoclonal antibodies to the human multicatalytic proteinase (proteasome). Hybridoma 1992 11: 507-17.
28. DUrso CM, Wang 2, Cao Y, Tatake R, Zeff RA, Ferrone S. Lack of HLA Class I antigen expression by cultured melanoma cells FO- 1 due to a defect in B2m gene expression. j Uin Invest 1991: 87: 284-92.
29. Kohler G, Milstein C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975 256 495-7.
30. Marincola FM, Hijazi YM, Fetsch P et al. Analysis of expression of the melanoma- associated antigens MART-1 and gpl00 in metastatic melanoma cell lines and in in situ lesions. j Immunothr 1996: 19 192-205.
31. Frentzel S, Kuhn-Hartmann I, Gernold M, Gott P, Seelig A, Kloetzel PM. The major- histocompatibility-complex-encoded beta- type proteasome subunits LMP2 and LMP7. Evidence that LMP2 and LMP7 are synthesized as pro-proteins and that cellular levels of both mRNA and LMP- containing 20s proteasomes are differentially regulated. Eur J Biochem. 1993: 216 11!3-26.
32. Kawahara H, Yokosawa H. Cell cycle- dependent change of proteasome distribution during embryonic development of the ascidian Halocynthia roretzi. Deu Biol 1992 151: 27-33.
33. Haass C, Pesold Hurt B, Multhaup G, Beyreuther K, Kloetzel PM. The PROS-35 gene encodes the 35 kd protein subunit of Drosophila melanogaster proteasome. EMBO J 1989 8: 2373-9.
34. Klein U, Gernold M, Kloetzel PM. Cell- specific accumulation of Drosophila proteasomes (MCP) during early development. J Cell Bio11990: 11 1: 2275- 82.
Regulation of gene expression of proteasomes (multi-protease complexes) during growth and differentiation of human hematopoetic cells. 1 Biol Chem 1992 : 267 18100-9.
35. Shimbara N, Orino E, Sane S et al.
140 Tissue Antigens 1998: 51: 129-140
Top Related
Copyright © 2022 FDOKUMEN

