







Bahasa
Halaman
Hukum


BULLETIN OF MARINE SCIENCE 44(1) 251-2621989
DECAPOD AND STOMATOPOD COMMUNITIES OFSEAGRASS-COVERED MUD BANKS IN FLORIDA BAYINTER- AND INTRA-BANK HETEROGENEITY WITH
SPECIAL REFERENCE TO ISOLA TED SUBENVIRONMENTS
JeffG Holmquist George V N Powell and Susan M Sogard
ABSTRACTUsing throw trap sampling we compared the decapod and stomatopod fauna of the grass-
covered mud banks in five major subenvironments of Florida Bay and tested for zonationin these fauna across individual banks Fauna was primarily Gulf-Carolinean despite thepresence of a rich Antillean community just outside the bay The two subenvironmentsadjacent to open ocean had high species richness but banks became increasingly depauperatetoward the interior of the bay with the innermost study site supporting a virtual monocultureof the grass shrimp Thor joridanus Species richness was similar across bank tops and exposedand sheltered sides For the majority of species and sites the exposed sides had the lowestabundance of the three transects We suggest that restricted circulation may limit larvalrecruitment to inner portions of the bay (particularly for those species whose adult populationswithin the bay frequently fell to virtually nil) and that physical parameters especially salinitypartially mediate the community structure of organisms that do gain access to the baysisolated areas
Florida Bays unique system of web-like mud banks (Holmquist et aI 1989fig 1) divide the bay into shallow basins less than 3 m deep The shallow banksare covered with dense meadows of Thalassia testudinum Halodule wrightiiSyringodium filiforme with Thalassia by far the most abundant (Zieman et aI1989) Despite comprising only 25 of Florida Bays area the banks exert aconsiderable influence on the bay ecosystem in terms of sediment accumulation(Enos and Perkins 1979) seagrass standing crop and productivity (Zieman et aI1989) and restriction of circulation (Ginsburg 1956 Fleece 1962 Gorsline 1963Taft and Harbaugh 1964 Scholl 1966 Turney and Perkins 1972 Enos andPerkins 1979 Davies 1980 Costello et aI 1986) Because the banks are shallowthe demersal community faces predation from wading birds (Recher and Recher1968) and extremes of turbulence temperature and desiccation not encounteredin deeper seagrass beds (Holmquist et aI 1989) but this stress may be mitigatedby the greater bank seagrass cover
Florida Bays marine fauna is of considerable interest as the bay functions asan interface for Antillean Carolinean and Gulf ecological provinces Despite thekey location the marine invertebrates have been little surveyed the Crustaceaare no exception Tabb et ai (1962b) and Brook (1978) provided a study of theextreme northwestern bay and M Robblee and J Ti1mant are currently pursuingspatial and temporal aspects of decapod community ecology in Johnson Key BasinBesides this work there are investigations of commercial species (pink shrimpstone crab spiny lobster) populations (Higman 1952 Idyll et aI 1964 1966Tabb et aI 1962a Allen and Costello 1966 Costello and Allen 1966 Davisand Dodrill 1979 Bert et aI 1986 Costello et aI 1986 Bert and Stevelyunpublished IDavis and Dodrill 1989) and three species lists from small portions
Bert T M and J M Stevely Population characteristics of the stone crab Menippe mercenaria in Florida Bay and the FloridaKcys Unpublished
251
252 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
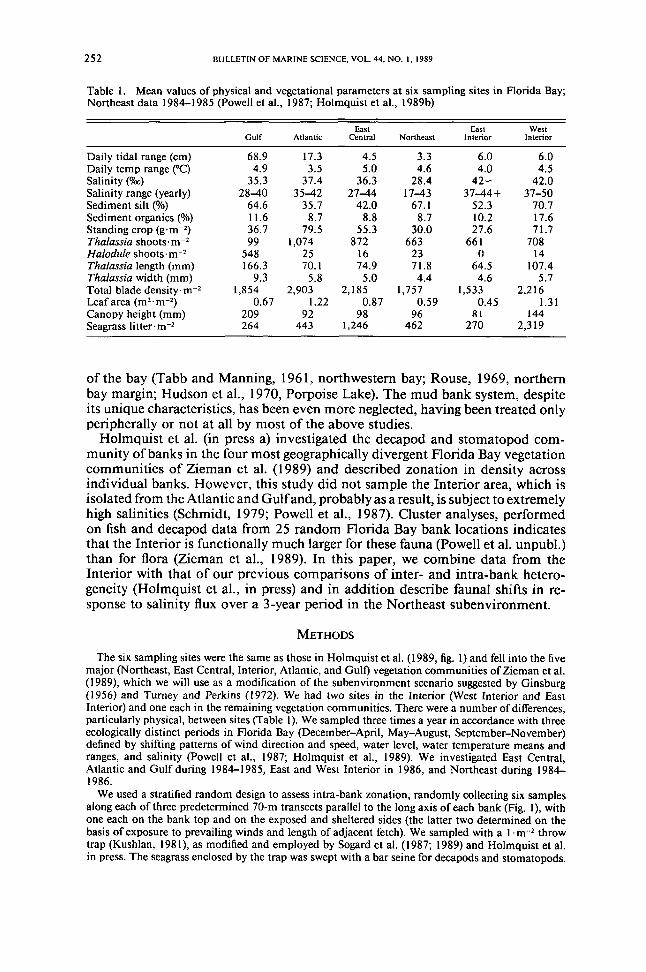
Table 1 Mean values of physical and vegetational parameters at six sampling sites in Horida BayNortheast data 1984-1985 (Powell et aI 1987 Holmquist et aI 1989b)
East East WestGulf Atlantic Central Northeast Interior Interior
Daily tidal range (cm) 689 173 45 33 60 60Daily temp range (C) 49 35 50 46 40 45Salinity (000) 353 374 363 284 42- 420Salinity range (yearly) 28-40 35-42 27-44 17-43 37-44+ 37-50Sediment silt () 646 357 420 671 523 707Sediment organics () 116 87 88 87 102 176Standing crop (gmiddotm-2) 367 795 553 300 276 717Thalassia shootsmiddotm-2 99 1074 872 663 661 708Halodule shootsm-2 548 25 16 23 0 14Thalassia length (mm) 1663 701 749 718 645 1074Thalassia width (mm) 93 58 50 44 46 57Total blade densitymiddotm-2 1854 2903 2185 1757 1533 2216Leaf area (m2bull m-2) 067 122 087 059 045 131Canopy height (mm) 209 92 98 96 81 144Seagrass litterm-2 264 443 1246 462 270 2319
of the bay (Tabb and Manning 1961 northwestern bay Rouse 1969 northernbay margin Hudson et aI 1970 Porpoise Lake) The mud bank system despiteits unique characteristics has been even more neglected having been treated onlyperipherally or not at all by most of the above studies
Holmquist et aI (in press a) investigated the decapod and stomatopod com-munity of banks in the four most geographically divergent Florida Bay vegetationcommunities of Zieman et al (1989) and described zonation in density acrossindividual banks However this study did not sample the Interior area which isisolated from the Atlantic and Gulf and probably as a result is subject to extremelyhigh salinities (Schmidt 1979 Powell et al 1987) Cluster analyses performedon fish and decapod data from 25 random Florida Bay bank locations indicatesthat the Interior is functionally much larger for these fauna (Powell et al unpubl)than for flora (Zieman et al 1989) In this paper we combine data from theInterior with that of our previous comparisons of inter- and intra-bank hetero-geneity (Holmquist et al in press) and in addition describe faunal shifts in re-sponse to salinity flux over a 3-year period in the Northeast subenvironment
METHODS
The six sampling sites were the same as those in Holmquist et al (1989 fig I) and fell into the fivemajor (Northeast East Central Interior Atlantic and Gulf) vegetation communities of Zieman et al(1989) which we will use as a modification of the subenvironment scenario suggested by Ginsburg(1956) and Turney and Perkins (1972) We had two sites in the Interior (West Interior and EastInterior) and one each in the remaining vegetation communities There were a number of differencesparticularly physical between sites (Table I) We sampled three times a year in accordance with threeecologically distinct periods in Horida Bay (December-April May-August September-November)defined by shifting patterns of wind direction and speed water level water temperature means andranges and salinity (Powell et aI 1987 Holmquist et aI 1989) We investigated East CentralAtlantic and Gulf during 1984-1985 East and West Interior in 1986 and Northeast during 1984-1986
We used a stratified random design to assess intra-bank zonation randomly collecting six samplesalong each of three predetermined 70-m transects parallel to the long axis of each bank (Fig I) withone each on the bank top and on the exposed and sheltered sides (the latter two determined on thebasis of exposure to prevailing winds and length of adjacent fetch) We sampled with a Imiddot m-2 throwtrap (Kushlan 1981) as modified and employed by Sogard et al (1987 1989) and Holmquist et alin press The seagrass enclosed by the trap was swept with a bar seine for decapods and stomatopods
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 253
T-7-7-
-I--f-
Figure 1 Bank cross section indicating transect positions crosses indicate the six random samplestransectsampling period
We performed ANOVAs and discriminant function analyses using the SPSSPC+ package (SPSSand Norusis 1986) on log-transformed data Site and transect collections were rarefied to N = 2000individuals and N = 250 individuals respectively using the methods of Heck et al (1975) to com-pensate for differing sampling effort and abundances
RESULTS
We collected 51 decapod and 2 stomatopod species during the study (Table 2)Small caridean shrimp especially Thor jloridanus were extremely abundant the18 caridean species comprised 97 of catch (T jloridanus 89) Of the fiveanomuran species two were numerically important Brachyurans although lack-ing in total abundance were represented by 27 species
Expected number of species was highest at Gulf and Atlantic very low at WestInterior and intermediate at East Central East Interior and Northeast (Table 3)Although expected number of species was roughly similar to the latter three sitesEast Central had twice the number of rare species (here arbitrarily defined as thosewith densities lt01 m-2) found at Northeast and four times the number collectedat East Interior
There were significant differences in density among sites for the 16 most com-mon decapods (ANOVA all P lt 001 Table 4) Gulf had the highest densitiesfor lOaf the 14 most abundant species Alpheus normanni Callinectes ornatusand a variety of rare species were most common at Atlantic (Tables 2 4) EastCentrals fauna was similar to but less species rich than that of Atlantic Eury-panopeus depressus was the only species in Table 4 most common at the depau-perate Northeast site Paguristes tortugae was most abundant at East Interior(Table 4) the only other species with densities greater than lm-2 at this site wereThor jloridanus and Hippolyte pleuracanthus (Table 2) The West Interior siterepresented a virtual monoculture of T jloridanus with a density of 884 individ-ualsmiddotm-2bull Sign tests on densities of the species in Table 4 revealed a significantprogression in density of Gulf gt Atlantic gt East Central gt East Interior gtNortheast = West Interior (P lt 005)
Five significant functions separating the six sites were derived from discriminant
254 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 2 Species of decapods and stomatopodsmiddot densities at six sites in Florida Bay percent of totaland rank (N = total number of throw trap samples at each site - = a individuals caught at site)
Sites
Northmiddot East East Westeast Central Interior Interior Atlantic GulfN- N~ N- N= N= N= of216 216 54 54 216 180 total Rank
Actual No species II 20 9 5 37 34Expected No species 84 130 100 21 198 203
DecapodaPenaeidae
Penaeus duorarum 001 034 009 002 078 808 070 7Palaemonidae
Leander pauensis 037 0Q2 17L tenuicornis 020 017 001 19Paaemon jloridanus 003 000 22Pericimenes americanus am 526 3015 265 2P iridescens 002 000 23P longicaudatus 000 006 1866 140 4
AlpheidaeApheus armillatus 002 000 23A heterochaeis 267 131 043 011 076 717 091 5A normanni 000 008 306 171 037 8
OgyrididaeOgyrides aphaerostris 001 000 23
HippolytidaeHippoyte peuracanthust 322 153 246 067 113 1898 210 3Latreutes fucorum 000 021 390 033 10L parvuus 005 000 22Thor dobkini 014 001 20Thor jloridanus 353 6701 3656 88426 10810 4176 8944 ITozeuma caroinense 02 215 016 12
ProcessidaeAmbidexter symmetricus 013 001 20Processa bermudensis 000 001 000 23
PalinuridaePanuirus argus 001 002 010 001 20
DiogenidaePaguristes anomaus 000 000 23P tortugaej 035 219 104 004 033 II
PaguridaePagurus mcaughlinae 006 104 044 153 710 073 6
PorcellanidaePetrolisthes armatus 000 000 23
CalappidaeCaappa sp sect 002 000 23
MajidaeLibinia dubia 001 003 077 006 15Metoporhaphis cacarata 000 000 23Microphrys bicornutus 001 000 23Mithrax forceps 000 000 23Peia mutica 002 000 23Pitho anisodon 0Q2 004 003 001 20Podochea riisei 000 000 23
PortunidaeCainectes ornatus 044 015 078 000 010 13C sapidus 001 011 004 000 001 001 19Cainectes sppsect 001 001 000 23
HOLMQUIST ET Al FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 255
Table 2 Continued
Sites
North-eastN~216
EastCentralN=216
EastInteriorN=54
WestInteriorN=54
AtlanticN=216
GulfN=180
oftotal Rank
005
4202 7334 4273 88512 12575 1405
Portunus depressifronsP gibbesiiPordwayiP sayiP sebaeP spinimanusPortunus Sppsect
MenippidaeMenippe mercenaria
PanopeidaeDyspanopeus texanusEurypanopeus depressusMicropanope spsectNeopanope packardiiD texanus and N
packardiillP occidentalisPanopeus simpsoniRhithropanopeus harrisii
GoneplacidaeEucratopsis crassimanus
PinnotheridaePinnixa sayana
OcypodidaeUca spsect
StomatopodaSquillidae
Meiosquilla quadridensGonodactylidae
Pseudosquilla ciliata
Total density
048000004
001
022
001
074
am005000
017
020 002
013001000000
007am001
001
171
019019003
000
000
000001
086
182
128000006
022
001
000
001 20000 23000 23000 23000 23001 22000 23
000 23
007 14002 16000 23034 9
0130Q2 18001 21002 18
002 18
000 23
000 23
000 23
000 22
bull Species observed on banks but not collected in study Palaemonetes intermedius Dildo Key Bank (M Robbleeand J Tilmant unpubl)Sicyonia typica Dave Foy Bank Alpheusfloridanus Colton Key Bank Lysmata wurdemanni First National Bank Clibarnarius viuatusCross Bank Cardisoma guanhumi Upper Cross Bank (Powell et al bull unpubl pers obs)t See Gore et al (198 I) for discussion of H pleuracamhuslzoslericola complex OUf P tortugar should be viewed as a species complex pending further systematic work (P McLaughlin peTS comm)sect Unidentifiable juvenilesII Juvenile female N packardii and D lexalJus
function analysis using densities of the 21 most abundant species as discriminatingvariables (Table 5) Ensuing classification indicated that East Central and EastInterior were the least distinct sites with incorrect assignment primarily to eachother and Northeast (Table 6) Atlantic Northeast and West Interior had highcorrect classification rates of approximately 90 Gulf was clearly distinct witha correct classification rate of 989 The high overall correct classification rate(854) was a demonstration of consistently different community structure amongthe various sites
Differences in community structure across individual banks were present thoughnot as striking as between sites At the four outermost sites the sheltered sidehad the highest total biomass and density of all species combined The exposedside had the lowest population densities the top and sheltered side did not differ
256 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 3 Species richness stratified by abundance (abundance determined on the basis of densitiesat the site in question)
West East EastInterior Interior Northeast Central Gulf Atlantic
Common species gt0Imiddotm-2 3 6 5 8 17 ISRare species lt0Imiddotm-2 2 3 6 12 17 22Actual No 5 10 II 20 34 37Expected No 21 100 84 130 203 198
from each other significantly (sign tests on density ranks across transects for 20most common species P lt 005) The exposed side however was not the mostdepauperate at the Interior sites Sign tests on the Interior data failed to showsignificant differences in overall transect densities There was no distinct patternin species richness across transects (Table 7)
We noted significant changes in decapod densities over the 3 years of samplingat Northeast (Table 8) Hippolytids were not taken in January 1984 a singleHippolyte pleuracanthus was collected in May There were 3 H pleuracanthusmiddotm-2 and 01 Thor floridanusmiddotm-2 in September During the next year there wasa steady increase in T floridanus to 170m-2 H pleuracanthus peaked at 20middotm-2 in January 1985 but averaged only 10m-2 in May and September Althoughthe H pleuracanthus population did not change significantly in 1986 T florida nusincreased to 380m-2 by September of that year (Table 8) We also noted a decreasein abundance for two mud crab species Eurypanopeus depressus and Rhithro-panopeus harrisii The abundance of the former dropped sharply in 1986 whereasdensity of the latter fell steadily throughout the study
Table 4 Results of I-way ANOVAs comparing densities across sites Interior data from 1986 allother sites 1985 Sites are listed in increasing order and those bearing the same superscript were notsignificantly different for the species in question (Student-Newman-Keuls multiple range tests a =005) All statistical analyses were performed on log-transformed data All F values were significantat P lt 00 I WI = West Interior EI = East Interior NE = Northeast EC = East Central A = AtlanticG = Gulf
Sites
Lowest HighestF density density
Total decapods 480 NE EI EC A G WIPenaeus duorarum 2960 NE WI EI EC A GPericlimenes americanus 1072 WI NEI EI EC A GP longicaudatus 611 WI NE EI EC A GAlpheus heterochaelis 1192 WI EI A EC NE GA normanni 1817 WI EI NE EC G AHippolyte pleuracanthus 889 WI Al ECI EI NE GLatreutes fucorum 436 WI NE EI EC A GThor floridanus 380 NE EI G EC A WITozeuma carolinense 472 WI NEI EI EC AI GPaguristes tortugae 624 WI NEI G EC A EIPagurus mclaughlinae 1866 WP NP EI Al EC GLibinia dubia 738 WI NE EI EC A GCallinectes ornatus 440 WI NE G EI EC2 A2Neopanope packardii 693 WP NP EI2 EC A GDypanopeus texanus 1217 WI NE EI EC A GEuropanopeus depressus 351 WI Ell ECI A G NE
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 257
Table 5 Discriminant function analysis results for decapod species collected at six sampling sitesduring 1985 and 1986 (all five functions were significant at P lt 000 I and all were included in theclassification process)
Function 4
Eigenvalue 784 241 074 056 022Variance explained () 666 204 63 48 188Scores at group centroids
East Centra] -112 00] -039 -056 -091Northeast -133 -132 057 052 008Atlantic -007 318 046 037 010Gulf 670 -060 -020 -005 004East Interior -144 -016 042 -207 072West Interior -202 002 -251 044 051
DISCUSSION
Most crustacean work in Florida Bay has emphasized the extreme northern andparticularly northwestern bay periphery The fauna described by Tabb and Man-ning (1961) Tabb et a1 (1962a basins channels and banks in the Cape Sable-Sandy Key-Flamingo area) and Rouse (1970 basin just north of Murray Key) isbasically consistent with that of our Gulf site Current studies in Johnson KeyBasin and bordering banks and mangrove islands (M Robblee and J Tilmantunpub1) also indicate a crustacean community similar to that of our Gulf siteHowever Rouses (1970) species list from basin stations straddling our Northeastsite reports a fauna more diverse than we found at the latter A Porpoise Lake(East Central subenvironment) species list (Hudson et aI 1970) reports a faunamuch like that of our Atlantic and East Central sites but some species foundprimarily at our Gulf site are also present (eg Leander paulensis)
Tabb et a1 (1962a) noted that the fauna in the aforementioned Cape Sable-Sandy Key-Flamingo area was more Gulf-Carolinean than Antillean and some-what surprisingly we found this characterization to hold in general for the entirebay Holmquist et a1 (in press) compared the crustacean mud bank fauna tothat of grassbeds in Puerto Rico (Bauer 1985a 1985b 1985c) and Indian River(Florida east coast Gore et aI 1981) and seagrass and other habitats in ApalacheeBay (junction of Florida peninsula and panhandle Hooks et aI 1976) The FloridaBay fauna shared few species with that of Puerto Rico but was similar to theIndian River and Apalachee Bay communities The bays mud bank fauna thusappears to have more temperate than tropical affinities
Table 6 Results of c1assificaiton of individual throw traps using discriminant functions derived fromall traps collected during 1985 and] 986 N = number of traps at each site Diagonal values indicatethe percentage of samples correctly classified at each site Overall correct classification rate = 854
Predicted membenhip ()
East East WestActual group N Central Nonheast Atlantic Gulf Interior Interior
East Central 108 620 167 37 00 167 09Northeast 215 28 916 00 00 14 42Atlantic 108 56 37 898 00 09 00Gulf 90 00 00 11 989 00 00East Interior 54 148 167 00 00 685 00West Interior 54 19 37 00 00 19 926
258 BULLETIN OF MARINE SCIENCE VOL 44 NO 11989
Table 7 Expected and actual number of species by transect and site
Transect West Interior Northeast EaSt Interior East Central AUaotic GulF
ShelteredExpected 14 48 59 71 94 132Actual 2 9 7 19 25 23
TopExpected 11 84 79 92 111 125Actual 4 II 7 14 25 25
ExposedExpected 11 49 58 69 109 136Actual 2 7 8 12 28 23
A trend of decreasing species richness (Table 3) and decreasing occurrence ofGulf and Atlantic suite species (Tables 2 4) is apparent as one progresses fromthe outermost bay sites to West Interior The main chain oflimestone keys to thesouth and east of the bay and the outermost perimeter of mud banks present aformidable circulation barrier (Ginsburg 1956 Tumey and Perkins 1972 Cos-tello et a1 1986) Tumey and Perkins provide a Florida Bay chart indicating anassumed limit oflunar tidal influence which is supported by records oflunar tidalrange at our study sites (Holmquist et a1 in press) We thus hypothesized thatthe mud bank network may constitute a significant obstacle to recruitment of Gulfand Atlantic species into the inner reaches of the bay Costello et aI (1986) havesuggested that the damping of circulation by the keys and mud banks may beresponsible for the paucity of juvenile pink shrimp in the eastern and interiorbay
A more Antillean fauna was found on banks on the Atlantic side of the Keysand along channels leading into Florida Bay (Powell et a1 unpubI) a number ofthese species were also found at the speciose Atlantic site Only occasional in-dividuals ofthe large suite of temperate species present at the Gulf site adjacentto the Gulf of Mexico were found at the Atlantic site and vice versa East Centralclose to but with limited communication with the Atlantic had an intermediatenumber of species including a number found predominantly at Atlantic (Table2) The majority of misclassified Atlantic traps were classified as East Central(Table 6) Northeast and East Interior about 16 and 20 km from open oceanrespectively and separated from the latter by several intervening banks hadsomewhat lower species richness than did East Centra1 With the exception of asingle Panuirus argus at East Interior the occasional Atlantic species were notcollected at these sites West Interior was separated from the southern bay by anadditional major bank system and a virtual wall of islands to the southeast Thisarea has very limited exchange with outlying areas as indicated by salinity dif-ferentials of over 10000 (Schmidt 1979 Powell et a1 unpub1) In tum this wasthe least species rich site with Thor joridanus representing 999 of all individ-uals and an expected number of species of only 21
However Holmquist et aI (in press) point out that restricted circulation isnot of itself an adequate mechanism for explaining the observed progression inspecies richness and suggest that unless a given species suffers a frequent de factoextinction it should eventually be capable of colonizing the bays interior despitepoor circulation These authors suggest cold-induced mortality (Storey and Gudg-er 1936 Storey 1937 Miller 1940 Tabb et a1 1962a Powell et a1 1987) andreproductive strategy (eg pink shrimp migrations from the bay Tabb et aI
HOLMQUIST IT AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 259
Table 8 Geometric means for densities at Northeast during 1984 1985 and 1986 and results ofI-way ANOV As comparing these densities Values with the same superscript were not significantlydifferent at P lt 005 (Student-Newman-Keuls multiple range tests a = 005) ( P lt 005 P lt001)
1984 1985 1986 F
Total decapods 268 2332 9132 1236Alpheus heterochaelis 155 205 233 37Thor jloridanus 009 994 6089 1265Hippolyte pleuracanthus 024 203 173 192Eurypanopeus depressus 026 026 007 56Rhithropanopeus harrisii 016 008 0 85
1962b Costello and Allen 1966) as likely sources of this requisite periodic ex-tirpation
If restricted circulation in conjunction with either cold fronts or other popu-lation reducing mechanisms does in fact limit larval access to the isolated areasof the bay then the degree of tendency to extreme population minima shouldhave been inversely related to penetration into the bay Indeed those species thatwere reduced from sizable populations to very low densities in May (Penaeusduorarum Pericimenes americanus P longicaudatus Latreutes fucorum andTozeuma carolinense Holmquist et a1 in press) demonstrated the least intrusioninto the bay (gt85 of each population found at a single site either Atlantic orGulf Table 2) Conversely those species that maintained substantial populationsthroughout the winter (Alpheus heterochaelis Hippolyte pleuracanthus Thor flor-idanus Paguristes tortugae Pagurus mcaughlinae Callinectes ornatus Neopan-ope packardii Holmquist et a1 in press) were more widespread (only 57-72of a population found at Atlantic or Gulf Table 2) Alpheus normanni appearedto be an intermediate
The fact that species not suffering severe population minima had broad distri-butions again indicates that restricted circulation is of itself an insufficient mech-anism for limiting recruitment to Florida Bay The northwestern Gulf of Mexicocoast provides an example of the inverse relationship that of acute cold butwithout restricted circulation Many species from our Gulf site are also presentat Cedar Key (Marshall 1985) These species suffer severe reductions in densitypossibly due to temperatures averaging 55degC colder than Florida Bay banks inwinter (based on data in Marshall 1985) but rebound during the summer pre-sumably because of unrestricted access of larvae to the site A similar situationis apparent at our Gulf and Atlantic sites where some species were almost extir-pated each year but recolonized the habitat during the summer and fall Thusboth restricted circulation and frequent near extinction are required to producethe most likely explanation for the depauperate Interior and Northeast suben-vironments Because the bay is a nexus for contiguous marine faunal provinces(Antillean and Gulf-Carolinean) and many Atlantic and Gulf species do not pen-etrate past their respective subenvironments Florida Bay has a rather dichoto-mous distribution of decapod fauna
Salinity patterns were probably important in determining patterns of distri-bution as well Our Northeast site had a mean 1983 salinity of about 16000(Whiteand Rosendahl unpubl) followed by a punctuated rise in salinity during our studyfrom a 1984 mean of 24000to means over 30000in 1985 and 1986 (Powell et al1987) Colonization by and an explosive increase of hippolytids at Northeastoccurred in conjunction with this salinity increase (Table 8) (although there were
260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087
252 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 1 Mean values of physical and vegetational parameters at six sampling sites in Horida BayNortheast data 1984-1985 (Powell et aI 1987 Holmquist et aI 1989b)
East East WestGulf Atlantic Central Northeast Interior Interior
Daily tidal range (cm) 689 173 45 33 60 60Daily temp range (C) 49 35 50 46 40 45Salinity (000) 353 374 363 284 42- 420Salinity range (yearly) 28-40 35-42 27-44 17-43 37-44+ 37-50Sediment silt () 646 357 420 671 523 707Sediment organics () 116 87 88 87 102 176Standing crop (gmiddotm-2) 367 795 553 300 276 717Thalassia shootsmiddotm-2 99 1074 872 663 661 708Halodule shootsm-2 548 25 16 23 0 14Thalassia length (mm) 1663 701 749 718 645 1074Thalassia width (mm) 93 58 50 44 46 57Total blade densitymiddotm-2 1854 2903 2185 1757 1533 2216Leaf area (m2bull m-2) 067 122 087 059 045 131Canopy height (mm) 209 92 98 96 81 144Seagrass litterm-2 264 443 1246 462 270 2319
of the bay (Tabb and Manning 1961 northwestern bay Rouse 1969 northernbay margin Hudson et aI 1970 Porpoise Lake) The mud bank system despiteits unique characteristics has been even more neglected having been treated onlyperipherally or not at all by most of the above studies
Holmquist et aI (in press a) investigated the decapod and stomatopod com-munity of banks in the four most geographically divergent Florida Bay vegetationcommunities of Zieman et al (1989) and described zonation in density acrossindividual banks However this study did not sample the Interior area which isisolated from the Atlantic and Gulf and probably as a result is subject to extremelyhigh salinities (Schmidt 1979 Powell et al 1987) Cluster analyses performedon fish and decapod data from 25 random Florida Bay bank locations indicatesthat the Interior is functionally much larger for these fauna (Powell et al unpubl)than for flora (Zieman et al 1989) In this paper we combine data from theInterior with that of our previous comparisons of inter- and intra-bank hetero-geneity (Holmquist et al in press) and in addition describe faunal shifts in re-sponse to salinity flux over a 3-year period in the Northeast subenvironment
METHODS
The six sampling sites were the same as those in Holmquist et al (1989 fig I) and fell into the fivemajor (Northeast East Central Interior Atlantic and Gulf) vegetation communities of Zieman et al(1989) which we will use as a modification of the subenvironment scenario suggested by Ginsburg(1956) and Turney and Perkins (1972) We had two sites in the Interior (West Interior and EastInterior) and one each in the remaining vegetation communities There were a number of differencesparticularly physical between sites (Table I) We sampled three times a year in accordance with threeecologically distinct periods in Horida Bay (December-April May-August September-November)defined by shifting patterns of wind direction and speed water level water temperature means andranges and salinity (Powell et aI 1987 Holmquist et aI 1989) We investigated East CentralAtlantic and Gulf during 1984-1985 East and West Interior in 1986 and Northeast during 1984-1986
We used a stratified random design to assess intra-bank zonation randomly collecting six samplesalong each of three predetermined 70-m transects parallel to the long axis of each bank (Fig I) withone each on the bank top and on the exposed and sheltered sides (the latter two determined on thebasis of exposure to prevailing winds and length of adjacent fetch) We sampled with a Imiddot m-2 throwtrap (Kushlan 1981) as modified and employed by Sogard et al (1987 1989) and Holmquist et alin press The seagrass enclosed by the trap was swept with a bar seine for decapods and stomatopods
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 253
T-7-7-
-I--f-
Figure 1 Bank cross section indicating transect positions crosses indicate the six random samplestransectsampling period
We performed ANOVAs and discriminant function analyses using the SPSSPC+ package (SPSSand Norusis 1986) on log-transformed data Site and transect collections were rarefied to N = 2000individuals and N = 250 individuals respectively using the methods of Heck et al (1975) to com-pensate for differing sampling effort and abundances
RESULTS
We collected 51 decapod and 2 stomatopod species during the study (Table 2)Small caridean shrimp especially Thor jloridanus were extremely abundant the18 caridean species comprised 97 of catch (T jloridanus 89) Of the fiveanomuran species two were numerically important Brachyurans although lack-ing in total abundance were represented by 27 species
Expected number of species was highest at Gulf and Atlantic very low at WestInterior and intermediate at East Central East Interior and Northeast (Table 3)Although expected number of species was roughly similar to the latter three sitesEast Central had twice the number of rare species (here arbitrarily defined as thosewith densities lt01 m-2) found at Northeast and four times the number collectedat East Interior
There were significant differences in density among sites for the 16 most com-mon decapods (ANOVA all P lt 001 Table 4) Gulf had the highest densitiesfor lOaf the 14 most abundant species Alpheus normanni Callinectes ornatusand a variety of rare species were most common at Atlantic (Tables 2 4) EastCentrals fauna was similar to but less species rich than that of Atlantic Eury-panopeus depressus was the only species in Table 4 most common at the depau-perate Northeast site Paguristes tortugae was most abundant at East Interior(Table 4) the only other species with densities greater than lm-2 at this site wereThor jloridanus and Hippolyte pleuracanthus (Table 2) The West Interior siterepresented a virtual monoculture of T jloridanus with a density of 884 individ-ualsmiddotm-2bull Sign tests on densities of the species in Table 4 revealed a significantprogression in density of Gulf gt Atlantic gt East Central gt East Interior gtNortheast = West Interior (P lt 005)
Five significant functions separating the six sites were derived from discriminant
254 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 2 Species of decapods and stomatopodsmiddot densities at six sites in Florida Bay percent of totaland rank (N = total number of throw trap samples at each site - = a individuals caught at site)
Sites
Northmiddot East East Westeast Central Interior Interior Atlantic GulfN- N~ N- N= N= N= of216 216 54 54 216 180 total Rank
Actual No species II 20 9 5 37 34Expected No species 84 130 100 21 198 203
DecapodaPenaeidae
Penaeus duorarum 001 034 009 002 078 808 070 7Palaemonidae
Leander pauensis 037 0Q2 17L tenuicornis 020 017 001 19Paaemon jloridanus 003 000 22Pericimenes americanus am 526 3015 265 2P iridescens 002 000 23P longicaudatus 000 006 1866 140 4
AlpheidaeApheus armillatus 002 000 23A heterochaeis 267 131 043 011 076 717 091 5A normanni 000 008 306 171 037 8
OgyrididaeOgyrides aphaerostris 001 000 23
HippolytidaeHippoyte peuracanthust 322 153 246 067 113 1898 210 3Latreutes fucorum 000 021 390 033 10L parvuus 005 000 22Thor dobkini 014 001 20Thor jloridanus 353 6701 3656 88426 10810 4176 8944 ITozeuma caroinense 02 215 016 12
ProcessidaeAmbidexter symmetricus 013 001 20Processa bermudensis 000 001 000 23
PalinuridaePanuirus argus 001 002 010 001 20
DiogenidaePaguristes anomaus 000 000 23P tortugaej 035 219 104 004 033 II
PaguridaePagurus mcaughlinae 006 104 044 153 710 073 6
PorcellanidaePetrolisthes armatus 000 000 23
CalappidaeCaappa sp sect 002 000 23
MajidaeLibinia dubia 001 003 077 006 15Metoporhaphis cacarata 000 000 23Microphrys bicornutus 001 000 23Mithrax forceps 000 000 23Peia mutica 002 000 23Pitho anisodon 0Q2 004 003 001 20Podochea riisei 000 000 23
PortunidaeCainectes ornatus 044 015 078 000 010 13C sapidus 001 011 004 000 001 001 19Cainectes sppsect 001 001 000 23
HOLMQUIST ET Al FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 255
Table 2 Continued
Sites
North-eastN~216
EastCentralN=216
EastInteriorN=54
WestInteriorN=54
AtlanticN=216
GulfN=180
oftotal Rank
005
4202 7334 4273 88512 12575 1405
Portunus depressifronsP gibbesiiPordwayiP sayiP sebaeP spinimanusPortunus Sppsect
MenippidaeMenippe mercenaria
PanopeidaeDyspanopeus texanusEurypanopeus depressusMicropanope spsectNeopanope packardiiD texanus and N
packardiillP occidentalisPanopeus simpsoniRhithropanopeus harrisii
GoneplacidaeEucratopsis crassimanus
PinnotheridaePinnixa sayana
OcypodidaeUca spsect
StomatopodaSquillidae
Meiosquilla quadridensGonodactylidae
Pseudosquilla ciliata
Total density
048000004
001
022
001
074
am005000
017
020 002
013001000000
007am001
001
171
019019003
000
000
000001
086
182
128000006
022
001
000
001 20000 23000 23000 23000 23001 22000 23
000 23
007 14002 16000 23034 9
0130Q2 18001 21002 18
002 18
000 23
000 23
000 23
000 22
bull Species observed on banks but not collected in study Palaemonetes intermedius Dildo Key Bank (M Robbleeand J Tilmant unpubl)Sicyonia typica Dave Foy Bank Alpheusfloridanus Colton Key Bank Lysmata wurdemanni First National Bank Clibarnarius viuatusCross Bank Cardisoma guanhumi Upper Cross Bank (Powell et al bull unpubl pers obs)t See Gore et al (198 I) for discussion of H pleuracamhuslzoslericola complex OUf P tortugar should be viewed as a species complex pending further systematic work (P McLaughlin peTS comm)sect Unidentifiable juvenilesII Juvenile female N packardii and D lexalJus
function analysis using densities of the 21 most abundant species as discriminatingvariables (Table 5) Ensuing classification indicated that East Central and EastInterior were the least distinct sites with incorrect assignment primarily to eachother and Northeast (Table 6) Atlantic Northeast and West Interior had highcorrect classification rates of approximately 90 Gulf was clearly distinct witha correct classification rate of 989 The high overall correct classification rate(854) was a demonstration of consistently different community structure amongthe various sites
Differences in community structure across individual banks were present thoughnot as striking as between sites At the four outermost sites the sheltered sidehad the highest total biomass and density of all species combined The exposedside had the lowest population densities the top and sheltered side did not differ
256 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 3 Species richness stratified by abundance (abundance determined on the basis of densitiesat the site in question)
West East EastInterior Interior Northeast Central Gulf Atlantic
Common species gt0Imiddotm-2 3 6 5 8 17 ISRare species lt0Imiddotm-2 2 3 6 12 17 22Actual No 5 10 II 20 34 37Expected No 21 100 84 130 203 198
from each other significantly (sign tests on density ranks across transects for 20most common species P lt 005) The exposed side however was not the mostdepauperate at the Interior sites Sign tests on the Interior data failed to showsignificant differences in overall transect densities There was no distinct patternin species richness across transects (Table 7)
We noted significant changes in decapod densities over the 3 years of samplingat Northeast (Table 8) Hippolytids were not taken in January 1984 a singleHippolyte pleuracanthus was collected in May There were 3 H pleuracanthusmiddotm-2 and 01 Thor floridanusmiddotm-2 in September During the next year there wasa steady increase in T floridanus to 170m-2 H pleuracanthus peaked at 20middotm-2 in January 1985 but averaged only 10m-2 in May and September Althoughthe H pleuracanthus population did not change significantly in 1986 T florida nusincreased to 380m-2 by September of that year (Table 8) We also noted a decreasein abundance for two mud crab species Eurypanopeus depressus and Rhithro-panopeus harrisii The abundance of the former dropped sharply in 1986 whereasdensity of the latter fell steadily throughout the study
Table 4 Results of I-way ANOVAs comparing densities across sites Interior data from 1986 allother sites 1985 Sites are listed in increasing order and those bearing the same superscript were notsignificantly different for the species in question (Student-Newman-Keuls multiple range tests a =005) All statistical analyses were performed on log-transformed data All F values were significantat P lt 00 I WI = West Interior EI = East Interior NE = Northeast EC = East Central A = AtlanticG = Gulf
Sites
Lowest HighestF density density
Total decapods 480 NE EI EC A G WIPenaeus duorarum 2960 NE WI EI EC A GPericlimenes americanus 1072 WI NEI EI EC A GP longicaudatus 611 WI NE EI EC A GAlpheus heterochaelis 1192 WI EI A EC NE GA normanni 1817 WI EI NE EC G AHippolyte pleuracanthus 889 WI Al ECI EI NE GLatreutes fucorum 436 WI NE EI EC A GThor floridanus 380 NE EI G EC A WITozeuma carolinense 472 WI NEI EI EC AI GPaguristes tortugae 624 WI NEI G EC A EIPagurus mclaughlinae 1866 WP NP EI Al EC GLibinia dubia 738 WI NE EI EC A GCallinectes ornatus 440 WI NE G EI EC2 A2Neopanope packardii 693 WP NP EI2 EC A GDypanopeus texanus 1217 WI NE EI EC A GEuropanopeus depressus 351 WI Ell ECI A G NE
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 257
Table 5 Discriminant function analysis results for decapod species collected at six sampling sitesduring 1985 and 1986 (all five functions were significant at P lt 000 I and all were included in theclassification process)
Function 4
Eigenvalue 784 241 074 056 022Variance explained () 666 204 63 48 188Scores at group centroids
East Centra] -112 00] -039 -056 -091Northeast -133 -132 057 052 008Atlantic -007 318 046 037 010Gulf 670 -060 -020 -005 004East Interior -144 -016 042 -207 072West Interior -202 002 -251 044 051
DISCUSSION
Most crustacean work in Florida Bay has emphasized the extreme northern andparticularly northwestern bay periphery The fauna described by Tabb and Man-ning (1961) Tabb et a1 (1962a basins channels and banks in the Cape Sable-Sandy Key-Flamingo area) and Rouse (1970 basin just north of Murray Key) isbasically consistent with that of our Gulf site Current studies in Johnson KeyBasin and bordering banks and mangrove islands (M Robblee and J Tilmantunpub1) also indicate a crustacean community similar to that of our Gulf siteHowever Rouses (1970) species list from basin stations straddling our Northeastsite reports a fauna more diverse than we found at the latter A Porpoise Lake(East Central subenvironment) species list (Hudson et aI 1970) reports a faunamuch like that of our Atlantic and East Central sites but some species foundprimarily at our Gulf site are also present (eg Leander paulensis)
Tabb et a1 (1962a) noted that the fauna in the aforementioned Cape Sable-Sandy Key-Flamingo area was more Gulf-Carolinean than Antillean and some-what surprisingly we found this characterization to hold in general for the entirebay Holmquist et a1 (in press) compared the crustacean mud bank fauna tothat of grassbeds in Puerto Rico (Bauer 1985a 1985b 1985c) and Indian River(Florida east coast Gore et aI 1981) and seagrass and other habitats in ApalacheeBay (junction of Florida peninsula and panhandle Hooks et aI 1976) The FloridaBay fauna shared few species with that of Puerto Rico but was similar to theIndian River and Apalachee Bay communities The bays mud bank fauna thusappears to have more temperate than tropical affinities
Table 6 Results of c1assificaiton of individual throw traps using discriminant functions derived fromall traps collected during 1985 and] 986 N = number of traps at each site Diagonal values indicatethe percentage of samples correctly classified at each site Overall correct classification rate = 854
Predicted membenhip ()
East East WestActual group N Central Nonheast Atlantic Gulf Interior Interior
East Central 108 620 167 37 00 167 09Northeast 215 28 916 00 00 14 42Atlantic 108 56 37 898 00 09 00Gulf 90 00 00 11 989 00 00East Interior 54 148 167 00 00 685 00West Interior 54 19 37 00 00 19 926
258 BULLETIN OF MARINE SCIENCE VOL 44 NO 11989
Table 7 Expected and actual number of species by transect and site
Transect West Interior Northeast EaSt Interior East Central AUaotic GulF
ShelteredExpected 14 48 59 71 94 132Actual 2 9 7 19 25 23
TopExpected 11 84 79 92 111 125Actual 4 II 7 14 25 25
ExposedExpected 11 49 58 69 109 136Actual 2 7 8 12 28 23
A trend of decreasing species richness (Table 3) and decreasing occurrence ofGulf and Atlantic suite species (Tables 2 4) is apparent as one progresses fromthe outermost bay sites to West Interior The main chain oflimestone keys to thesouth and east of the bay and the outermost perimeter of mud banks present aformidable circulation barrier (Ginsburg 1956 Tumey and Perkins 1972 Cos-tello et a1 1986) Tumey and Perkins provide a Florida Bay chart indicating anassumed limit oflunar tidal influence which is supported by records oflunar tidalrange at our study sites (Holmquist et a1 in press) We thus hypothesized thatthe mud bank network may constitute a significant obstacle to recruitment of Gulfand Atlantic species into the inner reaches of the bay Costello et aI (1986) havesuggested that the damping of circulation by the keys and mud banks may beresponsible for the paucity of juvenile pink shrimp in the eastern and interiorbay
A more Antillean fauna was found on banks on the Atlantic side of the Keysand along channels leading into Florida Bay (Powell et a1 unpubI) a number ofthese species were also found at the speciose Atlantic site Only occasional in-dividuals ofthe large suite of temperate species present at the Gulf site adjacentto the Gulf of Mexico were found at the Atlantic site and vice versa East Centralclose to but with limited communication with the Atlantic had an intermediatenumber of species including a number found predominantly at Atlantic (Table2) The majority of misclassified Atlantic traps were classified as East Central(Table 6) Northeast and East Interior about 16 and 20 km from open oceanrespectively and separated from the latter by several intervening banks hadsomewhat lower species richness than did East Centra1 With the exception of asingle Panuirus argus at East Interior the occasional Atlantic species were notcollected at these sites West Interior was separated from the southern bay by anadditional major bank system and a virtual wall of islands to the southeast Thisarea has very limited exchange with outlying areas as indicated by salinity dif-ferentials of over 10000 (Schmidt 1979 Powell et a1 unpub1) In tum this wasthe least species rich site with Thor joridanus representing 999 of all individ-uals and an expected number of species of only 21
However Holmquist et aI (in press) point out that restricted circulation isnot of itself an adequate mechanism for explaining the observed progression inspecies richness and suggest that unless a given species suffers a frequent de factoextinction it should eventually be capable of colonizing the bays interior despitepoor circulation These authors suggest cold-induced mortality (Storey and Gudg-er 1936 Storey 1937 Miller 1940 Tabb et a1 1962a Powell et a1 1987) andreproductive strategy (eg pink shrimp migrations from the bay Tabb et aI
HOLMQUIST IT AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 259
Table 8 Geometric means for densities at Northeast during 1984 1985 and 1986 and results ofI-way ANOV As comparing these densities Values with the same superscript were not significantlydifferent at P lt 005 (Student-Newman-Keuls multiple range tests a = 005) ( P lt 005 P lt001)
1984 1985 1986 F
Total decapods 268 2332 9132 1236Alpheus heterochaelis 155 205 233 37Thor jloridanus 009 994 6089 1265Hippolyte pleuracanthus 024 203 173 192Eurypanopeus depressus 026 026 007 56Rhithropanopeus harrisii 016 008 0 85
1962b Costello and Allen 1966) as likely sources of this requisite periodic ex-tirpation
If restricted circulation in conjunction with either cold fronts or other popu-lation reducing mechanisms does in fact limit larval access to the isolated areasof the bay then the degree of tendency to extreme population minima shouldhave been inversely related to penetration into the bay Indeed those species thatwere reduced from sizable populations to very low densities in May (Penaeusduorarum Pericimenes americanus P longicaudatus Latreutes fucorum andTozeuma carolinense Holmquist et a1 in press) demonstrated the least intrusioninto the bay (gt85 of each population found at a single site either Atlantic orGulf Table 2) Conversely those species that maintained substantial populationsthroughout the winter (Alpheus heterochaelis Hippolyte pleuracanthus Thor flor-idanus Paguristes tortugae Pagurus mcaughlinae Callinectes ornatus Neopan-ope packardii Holmquist et a1 in press) were more widespread (only 57-72of a population found at Atlantic or Gulf Table 2) Alpheus normanni appearedto be an intermediate
The fact that species not suffering severe population minima had broad distri-butions again indicates that restricted circulation is of itself an insufficient mech-anism for limiting recruitment to Florida Bay The northwestern Gulf of Mexicocoast provides an example of the inverse relationship that of acute cold butwithout restricted circulation Many species from our Gulf site are also presentat Cedar Key (Marshall 1985) These species suffer severe reductions in densitypossibly due to temperatures averaging 55degC colder than Florida Bay banks inwinter (based on data in Marshall 1985) but rebound during the summer pre-sumably because of unrestricted access of larvae to the site A similar situationis apparent at our Gulf and Atlantic sites where some species were almost extir-pated each year but recolonized the habitat during the summer and fall Thusboth restricted circulation and frequent near extinction are required to producethe most likely explanation for the depauperate Interior and Northeast suben-vironments Because the bay is a nexus for contiguous marine faunal provinces(Antillean and Gulf-Carolinean) and many Atlantic and Gulf species do not pen-etrate past their respective subenvironments Florida Bay has a rather dichoto-mous distribution of decapod fauna
Salinity patterns were probably important in determining patterns of distri-bution as well Our Northeast site had a mean 1983 salinity of about 16000(Whiteand Rosendahl unpubl) followed by a punctuated rise in salinity during our studyfrom a 1984 mean of 24000to means over 30000in 1985 and 1986 (Powell et al1987) Colonization by and an explosive increase of hippolytids at Northeastoccurred in conjunction with this salinity increase (Table 8) (although there were
260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087

HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 253
T-7-7-
-I--f-
Figure 1 Bank cross section indicating transect positions crosses indicate the six random samplestransectsampling period
We performed ANOVAs and discriminant function analyses using the SPSSPC+ package (SPSSand Norusis 1986) on log-transformed data Site and transect collections were rarefied to N = 2000individuals and N = 250 individuals respectively using the methods of Heck et al (1975) to com-pensate for differing sampling effort and abundances
RESULTS
We collected 51 decapod and 2 stomatopod species during the study (Table 2)Small caridean shrimp especially Thor jloridanus were extremely abundant the18 caridean species comprised 97 of catch (T jloridanus 89) Of the fiveanomuran species two were numerically important Brachyurans although lack-ing in total abundance were represented by 27 species
Expected number of species was highest at Gulf and Atlantic very low at WestInterior and intermediate at East Central East Interior and Northeast (Table 3)Although expected number of species was roughly similar to the latter three sitesEast Central had twice the number of rare species (here arbitrarily defined as thosewith densities lt01 m-2) found at Northeast and four times the number collectedat East Interior
There were significant differences in density among sites for the 16 most com-mon decapods (ANOVA all P lt 001 Table 4) Gulf had the highest densitiesfor lOaf the 14 most abundant species Alpheus normanni Callinectes ornatusand a variety of rare species were most common at Atlantic (Tables 2 4) EastCentrals fauna was similar to but less species rich than that of Atlantic Eury-panopeus depressus was the only species in Table 4 most common at the depau-perate Northeast site Paguristes tortugae was most abundant at East Interior(Table 4) the only other species with densities greater than lm-2 at this site wereThor jloridanus and Hippolyte pleuracanthus (Table 2) The West Interior siterepresented a virtual monoculture of T jloridanus with a density of 884 individ-ualsmiddotm-2bull Sign tests on densities of the species in Table 4 revealed a significantprogression in density of Gulf gt Atlantic gt East Central gt East Interior gtNortheast = West Interior (P lt 005)
Five significant functions separating the six sites were derived from discriminant
254 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 2 Species of decapods and stomatopodsmiddot densities at six sites in Florida Bay percent of totaland rank (N = total number of throw trap samples at each site - = a individuals caught at site)
Sites
Northmiddot East East Westeast Central Interior Interior Atlantic GulfN- N~ N- N= N= N= of216 216 54 54 216 180 total Rank
Actual No species II 20 9 5 37 34Expected No species 84 130 100 21 198 203
DecapodaPenaeidae
Penaeus duorarum 001 034 009 002 078 808 070 7Palaemonidae
Leander pauensis 037 0Q2 17L tenuicornis 020 017 001 19Paaemon jloridanus 003 000 22Pericimenes americanus am 526 3015 265 2P iridescens 002 000 23P longicaudatus 000 006 1866 140 4
AlpheidaeApheus armillatus 002 000 23A heterochaeis 267 131 043 011 076 717 091 5A normanni 000 008 306 171 037 8
OgyrididaeOgyrides aphaerostris 001 000 23
HippolytidaeHippoyte peuracanthust 322 153 246 067 113 1898 210 3Latreutes fucorum 000 021 390 033 10L parvuus 005 000 22Thor dobkini 014 001 20Thor jloridanus 353 6701 3656 88426 10810 4176 8944 ITozeuma caroinense 02 215 016 12
ProcessidaeAmbidexter symmetricus 013 001 20Processa bermudensis 000 001 000 23
PalinuridaePanuirus argus 001 002 010 001 20
DiogenidaePaguristes anomaus 000 000 23P tortugaej 035 219 104 004 033 II
PaguridaePagurus mcaughlinae 006 104 044 153 710 073 6
PorcellanidaePetrolisthes armatus 000 000 23
CalappidaeCaappa sp sect 002 000 23
MajidaeLibinia dubia 001 003 077 006 15Metoporhaphis cacarata 000 000 23Microphrys bicornutus 001 000 23Mithrax forceps 000 000 23Peia mutica 002 000 23Pitho anisodon 0Q2 004 003 001 20Podochea riisei 000 000 23
PortunidaeCainectes ornatus 044 015 078 000 010 13C sapidus 001 011 004 000 001 001 19Cainectes sppsect 001 001 000 23
HOLMQUIST ET Al FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 255
Table 2 Continued
Sites
North-eastN~216
EastCentralN=216
EastInteriorN=54
WestInteriorN=54
AtlanticN=216
GulfN=180
oftotal Rank
005
4202 7334 4273 88512 12575 1405
Portunus depressifronsP gibbesiiPordwayiP sayiP sebaeP spinimanusPortunus Sppsect
MenippidaeMenippe mercenaria
PanopeidaeDyspanopeus texanusEurypanopeus depressusMicropanope spsectNeopanope packardiiD texanus and N
packardiillP occidentalisPanopeus simpsoniRhithropanopeus harrisii
GoneplacidaeEucratopsis crassimanus
PinnotheridaePinnixa sayana
OcypodidaeUca spsect
StomatopodaSquillidae
Meiosquilla quadridensGonodactylidae
Pseudosquilla ciliata
Total density
048000004
001
022
001
074
am005000
017
020 002
013001000000
007am001
001
171
019019003
000
000
000001
086
182
128000006
022
001
000
001 20000 23000 23000 23000 23001 22000 23
000 23
007 14002 16000 23034 9
0130Q2 18001 21002 18
002 18
000 23
000 23
000 23
000 22
bull Species observed on banks but not collected in study Palaemonetes intermedius Dildo Key Bank (M Robbleeand J Tilmant unpubl)Sicyonia typica Dave Foy Bank Alpheusfloridanus Colton Key Bank Lysmata wurdemanni First National Bank Clibarnarius viuatusCross Bank Cardisoma guanhumi Upper Cross Bank (Powell et al bull unpubl pers obs)t See Gore et al (198 I) for discussion of H pleuracamhuslzoslericola complex OUf P tortugar should be viewed as a species complex pending further systematic work (P McLaughlin peTS comm)sect Unidentifiable juvenilesII Juvenile female N packardii and D lexalJus
function analysis using densities of the 21 most abundant species as discriminatingvariables (Table 5) Ensuing classification indicated that East Central and EastInterior were the least distinct sites with incorrect assignment primarily to eachother and Northeast (Table 6) Atlantic Northeast and West Interior had highcorrect classification rates of approximately 90 Gulf was clearly distinct witha correct classification rate of 989 The high overall correct classification rate(854) was a demonstration of consistently different community structure amongthe various sites
Differences in community structure across individual banks were present thoughnot as striking as between sites At the four outermost sites the sheltered sidehad the highest total biomass and density of all species combined The exposedside had the lowest population densities the top and sheltered side did not differ
256 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 3 Species richness stratified by abundance (abundance determined on the basis of densitiesat the site in question)
West East EastInterior Interior Northeast Central Gulf Atlantic
Common species gt0Imiddotm-2 3 6 5 8 17 ISRare species lt0Imiddotm-2 2 3 6 12 17 22Actual No 5 10 II 20 34 37Expected No 21 100 84 130 203 198
from each other significantly (sign tests on density ranks across transects for 20most common species P lt 005) The exposed side however was not the mostdepauperate at the Interior sites Sign tests on the Interior data failed to showsignificant differences in overall transect densities There was no distinct patternin species richness across transects (Table 7)
We noted significant changes in decapod densities over the 3 years of samplingat Northeast (Table 8) Hippolytids were not taken in January 1984 a singleHippolyte pleuracanthus was collected in May There were 3 H pleuracanthusmiddotm-2 and 01 Thor floridanusmiddotm-2 in September During the next year there wasa steady increase in T floridanus to 170m-2 H pleuracanthus peaked at 20middotm-2 in January 1985 but averaged only 10m-2 in May and September Althoughthe H pleuracanthus population did not change significantly in 1986 T florida nusincreased to 380m-2 by September of that year (Table 8) We also noted a decreasein abundance for two mud crab species Eurypanopeus depressus and Rhithro-panopeus harrisii The abundance of the former dropped sharply in 1986 whereasdensity of the latter fell steadily throughout the study
Table 4 Results of I-way ANOVAs comparing densities across sites Interior data from 1986 allother sites 1985 Sites are listed in increasing order and those bearing the same superscript were notsignificantly different for the species in question (Student-Newman-Keuls multiple range tests a =005) All statistical analyses were performed on log-transformed data All F values were significantat P lt 00 I WI = West Interior EI = East Interior NE = Northeast EC = East Central A = AtlanticG = Gulf
Sites
Lowest HighestF density density
Total decapods 480 NE EI EC A G WIPenaeus duorarum 2960 NE WI EI EC A GPericlimenes americanus 1072 WI NEI EI EC A GP longicaudatus 611 WI NE EI EC A GAlpheus heterochaelis 1192 WI EI A EC NE GA normanni 1817 WI EI NE EC G AHippolyte pleuracanthus 889 WI Al ECI EI NE GLatreutes fucorum 436 WI NE EI EC A GThor floridanus 380 NE EI G EC A WITozeuma carolinense 472 WI NEI EI EC AI GPaguristes tortugae 624 WI NEI G EC A EIPagurus mclaughlinae 1866 WP NP EI Al EC GLibinia dubia 738 WI NE EI EC A GCallinectes ornatus 440 WI NE G EI EC2 A2Neopanope packardii 693 WP NP EI2 EC A GDypanopeus texanus 1217 WI NE EI EC A GEuropanopeus depressus 351 WI Ell ECI A G NE
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 257
Table 5 Discriminant function analysis results for decapod species collected at six sampling sitesduring 1985 and 1986 (all five functions were significant at P lt 000 I and all were included in theclassification process)
Function 4
Eigenvalue 784 241 074 056 022Variance explained () 666 204 63 48 188Scores at group centroids
East Centra] -112 00] -039 -056 -091Northeast -133 -132 057 052 008Atlantic -007 318 046 037 010Gulf 670 -060 -020 -005 004East Interior -144 -016 042 -207 072West Interior -202 002 -251 044 051
DISCUSSION
Most crustacean work in Florida Bay has emphasized the extreme northern andparticularly northwestern bay periphery The fauna described by Tabb and Man-ning (1961) Tabb et a1 (1962a basins channels and banks in the Cape Sable-Sandy Key-Flamingo area) and Rouse (1970 basin just north of Murray Key) isbasically consistent with that of our Gulf site Current studies in Johnson KeyBasin and bordering banks and mangrove islands (M Robblee and J Tilmantunpub1) also indicate a crustacean community similar to that of our Gulf siteHowever Rouses (1970) species list from basin stations straddling our Northeastsite reports a fauna more diverse than we found at the latter A Porpoise Lake(East Central subenvironment) species list (Hudson et aI 1970) reports a faunamuch like that of our Atlantic and East Central sites but some species foundprimarily at our Gulf site are also present (eg Leander paulensis)
Tabb et a1 (1962a) noted that the fauna in the aforementioned Cape Sable-Sandy Key-Flamingo area was more Gulf-Carolinean than Antillean and some-what surprisingly we found this characterization to hold in general for the entirebay Holmquist et a1 (in press) compared the crustacean mud bank fauna tothat of grassbeds in Puerto Rico (Bauer 1985a 1985b 1985c) and Indian River(Florida east coast Gore et aI 1981) and seagrass and other habitats in ApalacheeBay (junction of Florida peninsula and panhandle Hooks et aI 1976) The FloridaBay fauna shared few species with that of Puerto Rico but was similar to theIndian River and Apalachee Bay communities The bays mud bank fauna thusappears to have more temperate than tropical affinities
Table 6 Results of c1assificaiton of individual throw traps using discriminant functions derived fromall traps collected during 1985 and] 986 N = number of traps at each site Diagonal values indicatethe percentage of samples correctly classified at each site Overall correct classification rate = 854
Predicted membenhip ()
East East WestActual group N Central Nonheast Atlantic Gulf Interior Interior
East Central 108 620 167 37 00 167 09Northeast 215 28 916 00 00 14 42Atlantic 108 56 37 898 00 09 00Gulf 90 00 00 11 989 00 00East Interior 54 148 167 00 00 685 00West Interior 54 19 37 00 00 19 926
258 BULLETIN OF MARINE SCIENCE VOL 44 NO 11989
Table 7 Expected and actual number of species by transect and site
Transect West Interior Northeast EaSt Interior East Central AUaotic GulF
ShelteredExpected 14 48 59 71 94 132Actual 2 9 7 19 25 23
TopExpected 11 84 79 92 111 125Actual 4 II 7 14 25 25
ExposedExpected 11 49 58 69 109 136Actual 2 7 8 12 28 23
A trend of decreasing species richness (Table 3) and decreasing occurrence ofGulf and Atlantic suite species (Tables 2 4) is apparent as one progresses fromthe outermost bay sites to West Interior The main chain oflimestone keys to thesouth and east of the bay and the outermost perimeter of mud banks present aformidable circulation barrier (Ginsburg 1956 Tumey and Perkins 1972 Cos-tello et a1 1986) Tumey and Perkins provide a Florida Bay chart indicating anassumed limit oflunar tidal influence which is supported by records oflunar tidalrange at our study sites (Holmquist et a1 in press) We thus hypothesized thatthe mud bank network may constitute a significant obstacle to recruitment of Gulfand Atlantic species into the inner reaches of the bay Costello et aI (1986) havesuggested that the damping of circulation by the keys and mud banks may beresponsible for the paucity of juvenile pink shrimp in the eastern and interiorbay
A more Antillean fauna was found on banks on the Atlantic side of the Keysand along channels leading into Florida Bay (Powell et a1 unpubI) a number ofthese species were also found at the speciose Atlantic site Only occasional in-dividuals ofthe large suite of temperate species present at the Gulf site adjacentto the Gulf of Mexico were found at the Atlantic site and vice versa East Centralclose to but with limited communication with the Atlantic had an intermediatenumber of species including a number found predominantly at Atlantic (Table2) The majority of misclassified Atlantic traps were classified as East Central(Table 6) Northeast and East Interior about 16 and 20 km from open oceanrespectively and separated from the latter by several intervening banks hadsomewhat lower species richness than did East Centra1 With the exception of asingle Panuirus argus at East Interior the occasional Atlantic species were notcollected at these sites West Interior was separated from the southern bay by anadditional major bank system and a virtual wall of islands to the southeast Thisarea has very limited exchange with outlying areas as indicated by salinity dif-ferentials of over 10000 (Schmidt 1979 Powell et a1 unpub1) In tum this wasthe least species rich site with Thor joridanus representing 999 of all individ-uals and an expected number of species of only 21
However Holmquist et aI (in press) point out that restricted circulation isnot of itself an adequate mechanism for explaining the observed progression inspecies richness and suggest that unless a given species suffers a frequent de factoextinction it should eventually be capable of colonizing the bays interior despitepoor circulation These authors suggest cold-induced mortality (Storey and Gudg-er 1936 Storey 1937 Miller 1940 Tabb et a1 1962a Powell et a1 1987) andreproductive strategy (eg pink shrimp migrations from the bay Tabb et aI
HOLMQUIST IT AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 259
Table 8 Geometric means for densities at Northeast during 1984 1985 and 1986 and results ofI-way ANOV As comparing these densities Values with the same superscript were not significantlydifferent at P lt 005 (Student-Newman-Keuls multiple range tests a = 005) ( P lt 005 P lt001)
1984 1985 1986 F
Total decapods 268 2332 9132 1236Alpheus heterochaelis 155 205 233 37Thor jloridanus 009 994 6089 1265Hippolyte pleuracanthus 024 203 173 192Eurypanopeus depressus 026 026 007 56Rhithropanopeus harrisii 016 008 0 85
1962b Costello and Allen 1966) as likely sources of this requisite periodic ex-tirpation
If restricted circulation in conjunction with either cold fronts or other popu-lation reducing mechanisms does in fact limit larval access to the isolated areasof the bay then the degree of tendency to extreme population minima shouldhave been inversely related to penetration into the bay Indeed those species thatwere reduced from sizable populations to very low densities in May (Penaeusduorarum Pericimenes americanus P longicaudatus Latreutes fucorum andTozeuma carolinense Holmquist et a1 in press) demonstrated the least intrusioninto the bay (gt85 of each population found at a single site either Atlantic orGulf Table 2) Conversely those species that maintained substantial populationsthroughout the winter (Alpheus heterochaelis Hippolyte pleuracanthus Thor flor-idanus Paguristes tortugae Pagurus mcaughlinae Callinectes ornatus Neopan-ope packardii Holmquist et a1 in press) were more widespread (only 57-72of a population found at Atlantic or Gulf Table 2) Alpheus normanni appearedto be an intermediate
The fact that species not suffering severe population minima had broad distri-butions again indicates that restricted circulation is of itself an insufficient mech-anism for limiting recruitment to Florida Bay The northwestern Gulf of Mexicocoast provides an example of the inverse relationship that of acute cold butwithout restricted circulation Many species from our Gulf site are also presentat Cedar Key (Marshall 1985) These species suffer severe reductions in densitypossibly due to temperatures averaging 55degC colder than Florida Bay banks inwinter (based on data in Marshall 1985) but rebound during the summer pre-sumably because of unrestricted access of larvae to the site A similar situationis apparent at our Gulf and Atlantic sites where some species were almost extir-pated each year but recolonized the habitat during the summer and fall Thusboth restricted circulation and frequent near extinction are required to producethe most likely explanation for the depauperate Interior and Northeast suben-vironments Because the bay is a nexus for contiguous marine faunal provinces(Antillean and Gulf-Carolinean) and many Atlantic and Gulf species do not pen-etrate past their respective subenvironments Florida Bay has a rather dichoto-mous distribution of decapod fauna
Salinity patterns were probably important in determining patterns of distri-bution as well Our Northeast site had a mean 1983 salinity of about 16000(Whiteand Rosendahl unpubl) followed by a punctuated rise in salinity during our studyfrom a 1984 mean of 24000to means over 30000in 1985 and 1986 (Powell et al1987) Colonization by and an explosive increase of hippolytids at Northeastoccurred in conjunction with this salinity increase (Table 8) (although there were
260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087
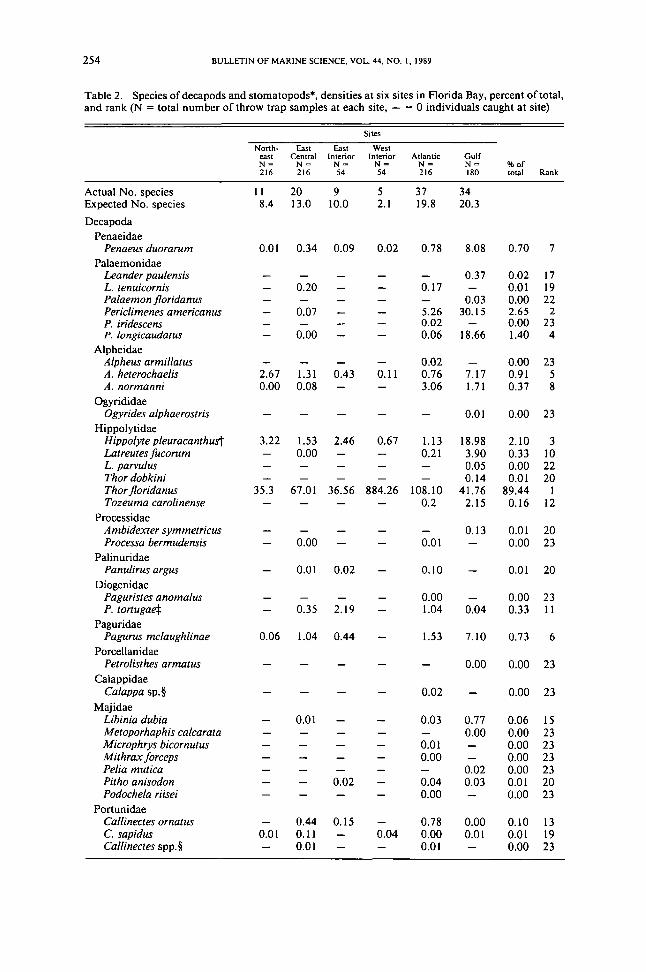
254 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 2 Species of decapods and stomatopodsmiddot densities at six sites in Florida Bay percent of totaland rank (N = total number of throw trap samples at each site - = a individuals caught at site)
Sites
Northmiddot East East Westeast Central Interior Interior Atlantic GulfN- N~ N- N= N= N= of216 216 54 54 216 180 total Rank
Actual No species II 20 9 5 37 34Expected No species 84 130 100 21 198 203
DecapodaPenaeidae
Penaeus duorarum 001 034 009 002 078 808 070 7Palaemonidae
Leander pauensis 037 0Q2 17L tenuicornis 020 017 001 19Paaemon jloridanus 003 000 22Pericimenes americanus am 526 3015 265 2P iridescens 002 000 23P longicaudatus 000 006 1866 140 4
AlpheidaeApheus armillatus 002 000 23A heterochaeis 267 131 043 011 076 717 091 5A normanni 000 008 306 171 037 8
OgyrididaeOgyrides aphaerostris 001 000 23
HippolytidaeHippoyte peuracanthust 322 153 246 067 113 1898 210 3Latreutes fucorum 000 021 390 033 10L parvuus 005 000 22Thor dobkini 014 001 20Thor jloridanus 353 6701 3656 88426 10810 4176 8944 ITozeuma caroinense 02 215 016 12
ProcessidaeAmbidexter symmetricus 013 001 20Processa bermudensis 000 001 000 23
PalinuridaePanuirus argus 001 002 010 001 20
DiogenidaePaguristes anomaus 000 000 23P tortugaej 035 219 104 004 033 II
PaguridaePagurus mcaughlinae 006 104 044 153 710 073 6
PorcellanidaePetrolisthes armatus 000 000 23
CalappidaeCaappa sp sect 002 000 23
MajidaeLibinia dubia 001 003 077 006 15Metoporhaphis cacarata 000 000 23Microphrys bicornutus 001 000 23Mithrax forceps 000 000 23Peia mutica 002 000 23Pitho anisodon 0Q2 004 003 001 20Podochea riisei 000 000 23
PortunidaeCainectes ornatus 044 015 078 000 010 13C sapidus 001 011 004 000 001 001 19Cainectes sppsect 001 001 000 23
HOLMQUIST ET Al FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 255
Table 2 Continued
Sites
North-eastN~216
EastCentralN=216
EastInteriorN=54
WestInteriorN=54
AtlanticN=216
GulfN=180
oftotal Rank
005
4202 7334 4273 88512 12575 1405
Portunus depressifronsP gibbesiiPordwayiP sayiP sebaeP spinimanusPortunus Sppsect
MenippidaeMenippe mercenaria
PanopeidaeDyspanopeus texanusEurypanopeus depressusMicropanope spsectNeopanope packardiiD texanus and N
packardiillP occidentalisPanopeus simpsoniRhithropanopeus harrisii
GoneplacidaeEucratopsis crassimanus
PinnotheridaePinnixa sayana
OcypodidaeUca spsect
StomatopodaSquillidae
Meiosquilla quadridensGonodactylidae
Pseudosquilla ciliata
Total density
048000004
001
022
001
074
am005000
017
020 002
013001000000
007am001
001
171
019019003
000
000
000001
086
182
128000006
022
001
000
001 20000 23000 23000 23000 23001 22000 23
000 23
007 14002 16000 23034 9
0130Q2 18001 21002 18
002 18
000 23
000 23
000 23
000 22
bull Species observed on banks but not collected in study Palaemonetes intermedius Dildo Key Bank (M Robbleeand J Tilmant unpubl)Sicyonia typica Dave Foy Bank Alpheusfloridanus Colton Key Bank Lysmata wurdemanni First National Bank Clibarnarius viuatusCross Bank Cardisoma guanhumi Upper Cross Bank (Powell et al bull unpubl pers obs)t See Gore et al (198 I) for discussion of H pleuracamhuslzoslericola complex OUf P tortugar should be viewed as a species complex pending further systematic work (P McLaughlin peTS comm)sect Unidentifiable juvenilesII Juvenile female N packardii and D lexalJus
function analysis using densities of the 21 most abundant species as discriminatingvariables (Table 5) Ensuing classification indicated that East Central and EastInterior were the least distinct sites with incorrect assignment primarily to eachother and Northeast (Table 6) Atlantic Northeast and West Interior had highcorrect classification rates of approximately 90 Gulf was clearly distinct witha correct classification rate of 989 The high overall correct classification rate(854) was a demonstration of consistently different community structure amongthe various sites
Differences in community structure across individual banks were present thoughnot as striking as between sites At the four outermost sites the sheltered sidehad the highest total biomass and density of all species combined The exposedside had the lowest population densities the top and sheltered side did not differ
256 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 3 Species richness stratified by abundance (abundance determined on the basis of densitiesat the site in question)
West East EastInterior Interior Northeast Central Gulf Atlantic
Common species gt0Imiddotm-2 3 6 5 8 17 ISRare species lt0Imiddotm-2 2 3 6 12 17 22Actual No 5 10 II 20 34 37Expected No 21 100 84 130 203 198
from each other significantly (sign tests on density ranks across transects for 20most common species P lt 005) The exposed side however was not the mostdepauperate at the Interior sites Sign tests on the Interior data failed to showsignificant differences in overall transect densities There was no distinct patternin species richness across transects (Table 7)
We noted significant changes in decapod densities over the 3 years of samplingat Northeast (Table 8) Hippolytids were not taken in January 1984 a singleHippolyte pleuracanthus was collected in May There were 3 H pleuracanthusmiddotm-2 and 01 Thor floridanusmiddotm-2 in September During the next year there wasa steady increase in T floridanus to 170m-2 H pleuracanthus peaked at 20middotm-2 in January 1985 but averaged only 10m-2 in May and September Althoughthe H pleuracanthus population did not change significantly in 1986 T florida nusincreased to 380m-2 by September of that year (Table 8) We also noted a decreasein abundance for two mud crab species Eurypanopeus depressus and Rhithro-panopeus harrisii The abundance of the former dropped sharply in 1986 whereasdensity of the latter fell steadily throughout the study
Table 4 Results of I-way ANOVAs comparing densities across sites Interior data from 1986 allother sites 1985 Sites are listed in increasing order and those bearing the same superscript were notsignificantly different for the species in question (Student-Newman-Keuls multiple range tests a =005) All statistical analyses were performed on log-transformed data All F values were significantat P lt 00 I WI = West Interior EI = East Interior NE = Northeast EC = East Central A = AtlanticG = Gulf
Sites
Lowest HighestF density density
Total decapods 480 NE EI EC A G WIPenaeus duorarum 2960 NE WI EI EC A GPericlimenes americanus 1072 WI NEI EI EC A GP longicaudatus 611 WI NE EI EC A GAlpheus heterochaelis 1192 WI EI A EC NE GA normanni 1817 WI EI NE EC G AHippolyte pleuracanthus 889 WI Al ECI EI NE GLatreutes fucorum 436 WI NE EI EC A GThor floridanus 380 NE EI G EC A WITozeuma carolinense 472 WI NEI EI EC AI GPaguristes tortugae 624 WI NEI G EC A EIPagurus mclaughlinae 1866 WP NP EI Al EC GLibinia dubia 738 WI NE EI EC A GCallinectes ornatus 440 WI NE G EI EC2 A2Neopanope packardii 693 WP NP EI2 EC A GDypanopeus texanus 1217 WI NE EI EC A GEuropanopeus depressus 351 WI Ell ECI A G NE
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 257
Table 5 Discriminant function analysis results for decapod species collected at six sampling sitesduring 1985 and 1986 (all five functions were significant at P lt 000 I and all were included in theclassification process)
Function 4
Eigenvalue 784 241 074 056 022Variance explained () 666 204 63 48 188Scores at group centroids
East Centra] -112 00] -039 -056 -091Northeast -133 -132 057 052 008Atlantic -007 318 046 037 010Gulf 670 -060 -020 -005 004East Interior -144 -016 042 -207 072West Interior -202 002 -251 044 051
DISCUSSION
Most crustacean work in Florida Bay has emphasized the extreme northern andparticularly northwestern bay periphery The fauna described by Tabb and Man-ning (1961) Tabb et a1 (1962a basins channels and banks in the Cape Sable-Sandy Key-Flamingo area) and Rouse (1970 basin just north of Murray Key) isbasically consistent with that of our Gulf site Current studies in Johnson KeyBasin and bordering banks and mangrove islands (M Robblee and J Tilmantunpub1) also indicate a crustacean community similar to that of our Gulf siteHowever Rouses (1970) species list from basin stations straddling our Northeastsite reports a fauna more diverse than we found at the latter A Porpoise Lake(East Central subenvironment) species list (Hudson et aI 1970) reports a faunamuch like that of our Atlantic and East Central sites but some species foundprimarily at our Gulf site are also present (eg Leander paulensis)
Tabb et a1 (1962a) noted that the fauna in the aforementioned Cape Sable-Sandy Key-Flamingo area was more Gulf-Carolinean than Antillean and some-what surprisingly we found this characterization to hold in general for the entirebay Holmquist et a1 (in press) compared the crustacean mud bank fauna tothat of grassbeds in Puerto Rico (Bauer 1985a 1985b 1985c) and Indian River(Florida east coast Gore et aI 1981) and seagrass and other habitats in ApalacheeBay (junction of Florida peninsula and panhandle Hooks et aI 1976) The FloridaBay fauna shared few species with that of Puerto Rico but was similar to theIndian River and Apalachee Bay communities The bays mud bank fauna thusappears to have more temperate than tropical affinities
Table 6 Results of c1assificaiton of individual throw traps using discriminant functions derived fromall traps collected during 1985 and] 986 N = number of traps at each site Diagonal values indicatethe percentage of samples correctly classified at each site Overall correct classification rate = 854
Predicted membenhip ()
East East WestActual group N Central Nonheast Atlantic Gulf Interior Interior
East Central 108 620 167 37 00 167 09Northeast 215 28 916 00 00 14 42Atlantic 108 56 37 898 00 09 00Gulf 90 00 00 11 989 00 00East Interior 54 148 167 00 00 685 00West Interior 54 19 37 00 00 19 926
258 BULLETIN OF MARINE SCIENCE VOL 44 NO 11989
Table 7 Expected and actual number of species by transect and site
Transect West Interior Northeast EaSt Interior East Central AUaotic GulF
ShelteredExpected 14 48 59 71 94 132Actual 2 9 7 19 25 23
TopExpected 11 84 79 92 111 125Actual 4 II 7 14 25 25
ExposedExpected 11 49 58 69 109 136Actual 2 7 8 12 28 23
A trend of decreasing species richness (Table 3) and decreasing occurrence ofGulf and Atlantic suite species (Tables 2 4) is apparent as one progresses fromthe outermost bay sites to West Interior The main chain oflimestone keys to thesouth and east of the bay and the outermost perimeter of mud banks present aformidable circulation barrier (Ginsburg 1956 Tumey and Perkins 1972 Cos-tello et a1 1986) Tumey and Perkins provide a Florida Bay chart indicating anassumed limit oflunar tidal influence which is supported by records oflunar tidalrange at our study sites (Holmquist et a1 in press) We thus hypothesized thatthe mud bank network may constitute a significant obstacle to recruitment of Gulfand Atlantic species into the inner reaches of the bay Costello et aI (1986) havesuggested that the damping of circulation by the keys and mud banks may beresponsible for the paucity of juvenile pink shrimp in the eastern and interiorbay
A more Antillean fauna was found on banks on the Atlantic side of the Keysand along channels leading into Florida Bay (Powell et a1 unpubI) a number ofthese species were also found at the speciose Atlantic site Only occasional in-dividuals ofthe large suite of temperate species present at the Gulf site adjacentto the Gulf of Mexico were found at the Atlantic site and vice versa East Centralclose to but with limited communication with the Atlantic had an intermediatenumber of species including a number found predominantly at Atlantic (Table2) The majority of misclassified Atlantic traps were classified as East Central(Table 6) Northeast and East Interior about 16 and 20 km from open oceanrespectively and separated from the latter by several intervening banks hadsomewhat lower species richness than did East Centra1 With the exception of asingle Panuirus argus at East Interior the occasional Atlantic species were notcollected at these sites West Interior was separated from the southern bay by anadditional major bank system and a virtual wall of islands to the southeast Thisarea has very limited exchange with outlying areas as indicated by salinity dif-ferentials of over 10000 (Schmidt 1979 Powell et a1 unpub1) In tum this wasthe least species rich site with Thor joridanus representing 999 of all individ-uals and an expected number of species of only 21
However Holmquist et aI (in press) point out that restricted circulation isnot of itself an adequate mechanism for explaining the observed progression inspecies richness and suggest that unless a given species suffers a frequent de factoextinction it should eventually be capable of colonizing the bays interior despitepoor circulation These authors suggest cold-induced mortality (Storey and Gudg-er 1936 Storey 1937 Miller 1940 Tabb et a1 1962a Powell et a1 1987) andreproductive strategy (eg pink shrimp migrations from the bay Tabb et aI
HOLMQUIST IT AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 259
Table 8 Geometric means for densities at Northeast during 1984 1985 and 1986 and results ofI-way ANOV As comparing these densities Values with the same superscript were not significantlydifferent at P lt 005 (Student-Newman-Keuls multiple range tests a = 005) ( P lt 005 P lt001)
1984 1985 1986 F
Total decapods 268 2332 9132 1236Alpheus heterochaelis 155 205 233 37Thor jloridanus 009 994 6089 1265Hippolyte pleuracanthus 024 203 173 192Eurypanopeus depressus 026 026 007 56Rhithropanopeus harrisii 016 008 0 85
1962b Costello and Allen 1966) as likely sources of this requisite periodic ex-tirpation
If restricted circulation in conjunction with either cold fronts or other popu-lation reducing mechanisms does in fact limit larval access to the isolated areasof the bay then the degree of tendency to extreme population minima shouldhave been inversely related to penetration into the bay Indeed those species thatwere reduced from sizable populations to very low densities in May (Penaeusduorarum Pericimenes americanus P longicaudatus Latreutes fucorum andTozeuma carolinense Holmquist et a1 in press) demonstrated the least intrusioninto the bay (gt85 of each population found at a single site either Atlantic orGulf Table 2) Conversely those species that maintained substantial populationsthroughout the winter (Alpheus heterochaelis Hippolyte pleuracanthus Thor flor-idanus Paguristes tortugae Pagurus mcaughlinae Callinectes ornatus Neopan-ope packardii Holmquist et a1 in press) were more widespread (only 57-72of a population found at Atlantic or Gulf Table 2) Alpheus normanni appearedto be an intermediate
The fact that species not suffering severe population minima had broad distri-butions again indicates that restricted circulation is of itself an insufficient mech-anism for limiting recruitment to Florida Bay The northwestern Gulf of Mexicocoast provides an example of the inverse relationship that of acute cold butwithout restricted circulation Many species from our Gulf site are also presentat Cedar Key (Marshall 1985) These species suffer severe reductions in densitypossibly due to temperatures averaging 55degC colder than Florida Bay banks inwinter (based on data in Marshall 1985) but rebound during the summer pre-sumably because of unrestricted access of larvae to the site A similar situationis apparent at our Gulf and Atlantic sites where some species were almost extir-pated each year but recolonized the habitat during the summer and fall Thusboth restricted circulation and frequent near extinction are required to producethe most likely explanation for the depauperate Interior and Northeast suben-vironments Because the bay is a nexus for contiguous marine faunal provinces(Antillean and Gulf-Carolinean) and many Atlantic and Gulf species do not pen-etrate past their respective subenvironments Florida Bay has a rather dichoto-mous distribution of decapod fauna
Salinity patterns were probably important in determining patterns of distri-bution as well Our Northeast site had a mean 1983 salinity of about 16000(Whiteand Rosendahl unpubl) followed by a punctuated rise in salinity during our studyfrom a 1984 mean of 24000to means over 30000in 1985 and 1986 (Powell et al1987) Colonization by and an explosive increase of hippolytids at Northeastoccurred in conjunction with this salinity increase (Table 8) (although there were
260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087
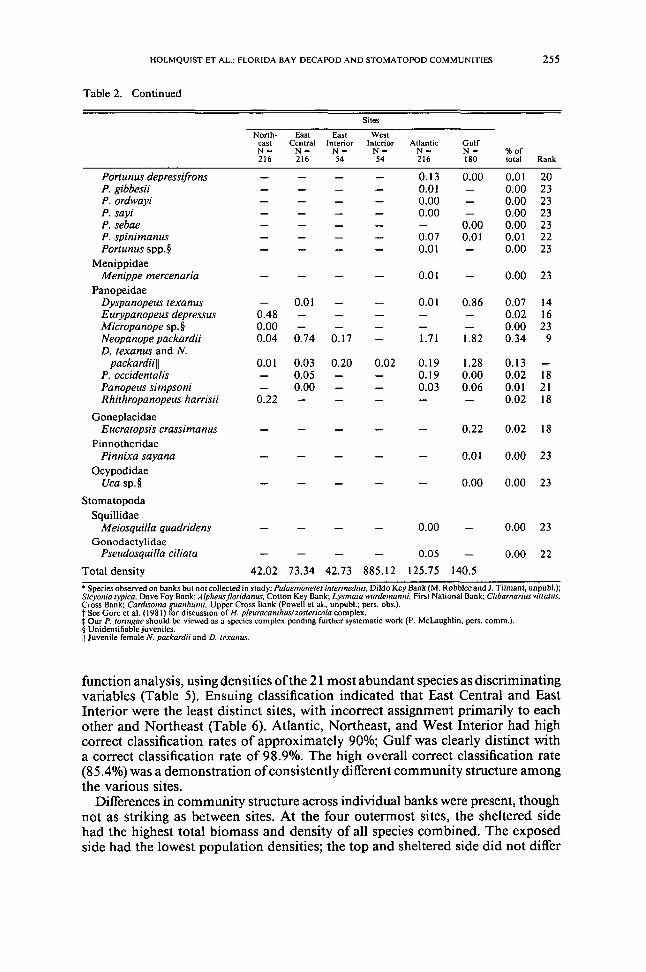
HOLMQUIST ET Al FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 255
Table 2 Continued
Sites
North-eastN~216
EastCentralN=216
EastInteriorN=54
WestInteriorN=54
AtlanticN=216
GulfN=180
oftotal Rank
005
4202 7334 4273 88512 12575 1405
Portunus depressifronsP gibbesiiPordwayiP sayiP sebaeP spinimanusPortunus Sppsect
MenippidaeMenippe mercenaria
PanopeidaeDyspanopeus texanusEurypanopeus depressusMicropanope spsectNeopanope packardiiD texanus and N
packardiillP occidentalisPanopeus simpsoniRhithropanopeus harrisii
GoneplacidaeEucratopsis crassimanus
PinnotheridaePinnixa sayana
OcypodidaeUca spsect
StomatopodaSquillidae
Meiosquilla quadridensGonodactylidae
Pseudosquilla ciliata
Total density
048000004
001
022
001
074
am005000
017
020 002
013001000000
007am001
001
171
019019003
000
000
000001
086
182
128000006
022
001
000
001 20000 23000 23000 23000 23001 22000 23
000 23
007 14002 16000 23034 9
0130Q2 18001 21002 18
002 18
000 23
000 23
000 23
000 22
bull Species observed on banks but not collected in study Palaemonetes intermedius Dildo Key Bank (M Robbleeand J Tilmant unpubl)Sicyonia typica Dave Foy Bank Alpheusfloridanus Colton Key Bank Lysmata wurdemanni First National Bank Clibarnarius viuatusCross Bank Cardisoma guanhumi Upper Cross Bank (Powell et al bull unpubl pers obs)t See Gore et al (198 I) for discussion of H pleuracamhuslzoslericola complex OUf P tortugar should be viewed as a species complex pending further systematic work (P McLaughlin peTS comm)sect Unidentifiable juvenilesII Juvenile female N packardii and D lexalJus
function analysis using densities of the 21 most abundant species as discriminatingvariables (Table 5) Ensuing classification indicated that East Central and EastInterior were the least distinct sites with incorrect assignment primarily to eachother and Northeast (Table 6) Atlantic Northeast and West Interior had highcorrect classification rates of approximately 90 Gulf was clearly distinct witha correct classification rate of 989 The high overall correct classification rate(854) was a demonstration of consistently different community structure amongthe various sites
Differences in community structure across individual banks were present thoughnot as striking as between sites At the four outermost sites the sheltered sidehad the highest total biomass and density of all species combined The exposedside had the lowest population densities the top and sheltered side did not differ
256 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 3 Species richness stratified by abundance (abundance determined on the basis of densitiesat the site in question)
West East EastInterior Interior Northeast Central Gulf Atlantic
Common species gt0Imiddotm-2 3 6 5 8 17 ISRare species lt0Imiddotm-2 2 3 6 12 17 22Actual No 5 10 II 20 34 37Expected No 21 100 84 130 203 198
from each other significantly (sign tests on density ranks across transects for 20most common species P lt 005) The exposed side however was not the mostdepauperate at the Interior sites Sign tests on the Interior data failed to showsignificant differences in overall transect densities There was no distinct patternin species richness across transects (Table 7)
We noted significant changes in decapod densities over the 3 years of samplingat Northeast (Table 8) Hippolytids were not taken in January 1984 a singleHippolyte pleuracanthus was collected in May There were 3 H pleuracanthusmiddotm-2 and 01 Thor floridanusmiddotm-2 in September During the next year there wasa steady increase in T floridanus to 170m-2 H pleuracanthus peaked at 20middotm-2 in January 1985 but averaged only 10m-2 in May and September Althoughthe H pleuracanthus population did not change significantly in 1986 T florida nusincreased to 380m-2 by September of that year (Table 8) We also noted a decreasein abundance for two mud crab species Eurypanopeus depressus and Rhithro-panopeus harrisii The abundance of the former dropped sharply in 1986 whereasdensity of the latter fell steadily throughout the study
Table 4 Results of I-way ANOVAs comparing densities across sites Interior data from 1986 allother sites 1985 Sites are listed in increasing order and those bearing the same superscript were notsignificantly different for the species in question (Student-Newman-Keuls multiple range tests a =005) All statistical analyses were performed on log-transformed data All F values were significantat P lt 00 I WI = West Interior EI = East Interior NE = Northeast EC = East Central A = AtlanticG = Gulf
Sites
Lowest HighestF density density
Total decapods 480 NE EI EC A G WIPenaeus duorarum 2960 NE WI EI EC A GPericlimenes americanus 1072 WI NEI EI EC A GP longicaudatus 611 WI NE EI EC A GAlpheus heterochaelis 1192 WI EI A EC NE GA normanni 1817 WI EI NE EC G AHippolyte pleuracanthus 889 WI Al ECI EI NE GLatreutes fucorum 436 WI NE EI EC A GThor floridanus 380 NE EI G EC A WITozeuma carolinense 472 WI NEI EI EC AI GPaguristes tortugae 624 WI NEI G EC A EIPagurus mclaughlinae 1866 WP NP EI Al EC GLibinia dubia 738 WI NE EI EC A GCallinectes ornatus 440 WI NE G EI EC2 A2Neopanope packardii 693 WP NP EI2 EC A GDypanopeus texanus 1217 WI NE EI EC A GEuropanopeus depressus 351 WI Ell ECI A G NE
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 257
Table 5 Discriminant function analysis results for decapod species collected at six sampling sitesduring 1985 and 1986 (all five functions were significant at P lt 000 I and all were included in theclassification process)
Function 4
Eigenvalue 784 241 074 056 022Variance explained () 666 204 63 48 188Scores at group centroids
East Centra] -112 00] -039 -056 -091Northeast -133 -132 057 052 008Atlantic -007 318 046 037 010Gulf 670 -060 -020 -005 004East Interior -144 -016 042 -207 072West Interior -202 002 -251 044 051
DISCUSSION
Most crustacean work in Florida Bay has emphasized the extreme northern andparticularly northwestern bay periphery The fauna described by Tabb and Man-ning (1961) Tabb et a1 (1962a basins channels and banks in the Cape Sable-Sandy Key-Flamingo area) and Rouse (1970 basin just north of Murray Key) isbasically consistent with that of our Gulf site Current studies in Johnson KeyBasin and bordering banks and mangrove islands (M Robblee and J Tilmantunpub1) also indicate a crustacean community similar to that of our Gulf siteHowever Rouses (1970) species list from basin stations straddling our Northeastsite reports a fauna more diverse than we found at the latter A Porpoise Lake(East Central subenvironment) species list (Hudson et aI 1970) reports a faunamuch like that of our Atlantic and East Central sites but some species foundprimarily at our Gulf site are also present (eg Leander paulensis)
Tabb et a1 (1962a) noted that the fauna in the aforementioned Cape Sable-Sandy Key-Flamingo area was more Gulf-Carolinean than Antillean and some-what surprisingly we found this characterization to hold in general for the entirebay Holmquist et a1 (in press) compared the crustacean mud bank fauna tothat of grassbeds in Puerto Rico (Bauer 1985a 1985b 1985c) and Indian River(Florida east coast Gore et aI 1981) and seagrass and other habitats in ApalacheeBay (junction of Florida peninsula and panhandle Hooks et aI 1976) The FloridaBay fauna shared few species with that of Puerto Rico but was similar to theIndian River and Apalachee Bay communities The bays mud bank fauna thusappears to have more temperate than tropical affinities
Table 6 Results of c1assificaiton of individual throw traps using discriminant functions derived fromall traps collected during 1985 and] 986 N = number of traps at each site Diagonal values indicatethe percentage of samples correctly classified at each site Overall correct classification rate = 854
Predicted membenhip ()
East East WestActual group N Central Nonheast Atlantic Gulf Interior Interior
East Central 108 620 167 37 00 167 09Northeast 215 28 916 00 00 14 42Atlantic 108 56 37 898 00 09 00Gulf 90 00 00 11 989 00 00East Interior 54 148 167 00 00 685 00West Interior 54 19 37 00 00 19 926
258 BULLETIN OF MARINE SCIENCE VOL 44 NO 11989
Table 7 Expected and actual number of species by transect and site
Transect West Interior Northeast EaSt Interior East Central AUaotic GulF
ShelteredExpected 14 48 59 71 94 132Actual 2 9 7 19 25 23
TopExpected 11 84 79 92 111 125Actual 4 II 7 14 25 25
ExposedExpected 11 49 58 69 109 136Actual 2 7 8 12 28 23
A trend of decreasing species richness (Table 3) and decreasing occurrence ofGulf and Atlantic suite species (Tables 2 4) is apparent as one progresses fromthe outermost bay sites to West Interior The main chain oflimestone keys to thesouth and east of the bay and the outermost perimeter of mud banks present aformidable circulation barrier (Ginsburg 1956 Tumey and Perkins 1972 Cos-tello et a1 1986) Tumey and Perkins provide a Florida Bay chart indicating anassumed limit oflunar tidal influence which is supported by records oflunar tidalrange at our study sites (Holmquist et a1 in press) We thus hypothesized thatthe mud bank network may constitute a significant obstacle to recruitment of Gulfand Atlantic species into the inner reaches of the bay Costello et aI (1986) havesuggested that the damping of circulation by the keys and mud banks may beresponsible for the paucity of juvenile pink shrimp in the eastern and interiorbay
A more Antillean fauna was found on banks on the Atlantic side of the Keysand along channels leading into Florida Bay (Powell et a1 unpubI) a number ofthese species were also found at the speciose Atlantic site Only occasional in-dividuals ofthe large suite of temperate species present at the Gulf site adjacentto the Gulf of Mexico were found at the Atlantic site and vice versa East Centralclose to but with limited communication with the Atlantic had an intermediatenumber of species including a number found predominantly at Atlantic (Table2) The majority of misclassified Atlantic traps were classified as East Central(Table 6) Northeast and East Interior about 16 and 20 km from open oceanrespectively and separated from the latter by several intervening banks hadsomewhat lower species richness than did East Centra1 With the exception of asingle Panuirus argus at East Interior the occasional Atlantic species were notcollected at these sites West Interior was separated from the southern bay by anadditional major bank system and a virtual wall of islands to the southeast Thisarea has very limited exchange with outlying areas as indicated by salinity dif-ferentials of over 10000 (Schmidt 1979 Powell et a1 unpub1) In tum this wasthe least species rich site with Thor joridanus representing 999 of all individ-uals and an expected number of species of only 21
However Holmquist et aI (in press) point out that restricted circulation isnot of itself an adequate mechanism for explaining the observed progression inspecies richness and suggest that unless a given species suffers a frequent de factoextinction it should eventually be capable of colonizing the bays interior despitepoor circulation These authors suggest cold-induced mortality (Storey and Gudg-er 1936 Storey 1937 Miller 1940 Tabb et a1 1962a Powell et a1 1987) andreproductive strategy (eg pink shrimp migrations from the bay Tabb et aI
HOLMQUIST IT AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 259
Table 8 Geometric means for densities at Northeast during 1984 1985 and 1986 and results ofI-way ANOV As comparing these densities Values with the same superscript were not significantlydifferent at P lt 005 (Student-Newman-Keuls multiple range tests a = 005) ( P lt 005 P lt001)
1984 1985 1986 F
Total decapods 268 2332 9132 1236Alpheus heterochaelis 155 205 233 37Thor jloridanus 009 994 6089 1265Hippolyte pleuracanthus 024 203 173 192Eurypanopeus depressus 026 026 007 56Rhithropanopeus harrisii 016 008 0 85
1962b Costello and Allen 1966) as likely sources of this requisite periodic ex-tirpation
If restricted circulation in conjunction with either cold fronts or other popu-lation reducing mechanisms does in fact limit larval access to the isolated areasof the bay then the degree of tendency to extreme population minima shouldhave been inversely related to penetration into the bay Indeed those species thatwere reduced from sizable populations to very low densities in May (Penaeusduorarum Pericimenes americanus P longicaudatus Latreutes fucorum andTozeuma carolinense Holmquist et a1 in press) demonstrated the least intrusioninto the bay (gt85 of each population found at a single site either Atlantic orGulf Table 2) Conversely those species that maintained substantial populationsthroughout the winter (Alpheus heterochaelis Hippolyte pleuracanthus Thor flor-idanus Paguristes tortugae Pagurus mcaughlinae Callinectes ornatus Neopan-ope packardii Holmquist et a1 in press) were more widespread (only 57-72of a population found at Atlantic or Gulf Table 2) Alpheus normanni appearedto be an intermediate
The fact that species not suffering severe population minima had broad distri-butions again indicates that restricted circulation is of itself an insufficient mech-anism for limiting recruitment to Florida Bay The northwestern Gulf of Mexicocoast provides an example of the inverse relationship that of acute cold butwithout restricted circulation Many species from our Gulf site are also presentat Cedar Key (Marshall 1985) These species suffer severe reductions in densitypossibly due to temperatures averaging 55degC colder than Florida Bay banks inwinter (based on data in Marshall 1985) but rebound during the summer pre-sumably because of unrestricted access of larvae to the site A similar situationis apparent at our Gulf and Atlantic sites where some species were almost extir-pated each year but recolonized the habitat during the summer and fall Thusboth restricted circulation and frequent near extinction are required to producethe most likely explanation for the depauperate Interior and Northeast suben-vironments Because the bay is a nexus for contiguous marine faunal provinces(Antillean and Gulf-Carolinean) and many Atlantic and Gulf species do not pen-etrate past their respective subenvironments Florida Bay has a rather dichoto-mous distribution of decapod fauna
Salinity patterns were probably important in determining patterns of distri-bution as well Our Northeast site had a mean 1983 salinity of about 16000(Whiteand Rosendahl unpubl) followed by a punctuated rise in salinity during our studyfrom a 1984 mean of 24000to means over 30000in 1985 and 1986 (Powell et al1987) Colonization by and an explosive increase of hippolytids at Northeastoccurred in conjunction with this salinity increase (Table 8) (although there were
260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087
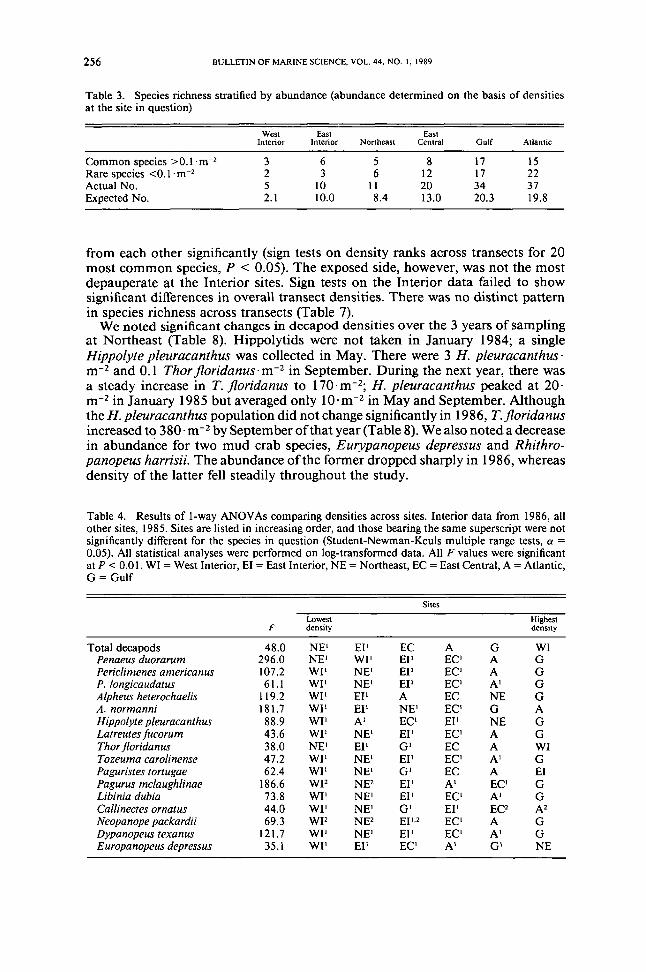
256 BULLETIN OF MARINE SCIENCE VOL 44 NO I 1989
Table 3 Species richness stratified by abundance (abundance determined on the basis of densitiesat the site in question)
West East EastInterior Interior Northeast Central Gulf Atlantic
Common species gt0Imiddotm-2 3 6 5 8 17 ISRare species lt0Imiddotm-2 2 3 6 12 17 22Actual No 5 10 II 20 34 37Expected No 21 100 84 130 203 198
from each other significantly (sign tests on density ranks across transects for 20most common species P lt 005) The exposed side however was not the mostdepauperate at the Interior sites Sign tests on the Interior data failed to showsignificant differences in overall transect densities There was no distinct patternin species richness across transects (Table 7)
We noted significant changes in decapod densities over the 3 years of samplingat Northeast (Table 8) Hippolytids were not taken in January 1984 a singleHippolyte pleuracanthus was collected in May There were 3 H pleuracanthusmiddotm-2 and 01 Thor floridanusmiddotm-2 in September During the next year there wasa steady increase in T floridanus to 170m-2 H pleuracanthus peaked at 20middotm-2 in January 1985 but averaged only 10m-2 in May and September Althoughthe H pleuracanthus population did not change significantly in 1986 T florida nusincreased to 380m-2 by September of that year (Table 8) We also noted a decreasein abundance for two mud crab species Eurypanopeus depressus and Rhithro-panopeus harrisii The abundance of the former dropped sharply in 1986 whereasdensity of the latter fell steadily throughout the study
Table 4 Results of I-way ANOVAs comparing densities across sites Interior data from 1986 allother sites 1985 Sites are listed in increasing order and those bearing the same superscript were notsignificantly different for the species in question (Student-Newman-Keuls multiple range tests a =005) All statistical analyses were performed on log-transformed data All F values were significantat P lt 00 I WI = West Interior EI = East Interior NE = Northeast EC = East Central A = AtlanticG = Gulf
Sites
Lowest HighestF density density
Total decapods 480 NE EI EC A G WIPenaeus duorarum 2960 NE WI EI EC A GPericlimenes americanus 1072 WI NEI EI EC A GP longicaudatus 611 WI NE EI EC A GAlpheus heterochaelis 1192 WI EI A EC NE GA normanni 1817 WI EI NE EC G AHippolyte pleuracanthus 889 WI Al ECI EI NE GLatreutes fucorum 436 WI NE EI EC A GThor floridanus 380 NE EI G EC A WITozeuma carolinense 472 WI NEI EI EC AI GPaguristes tortugae 624 WI NEI G EC A EIPagurus mclaughlinae 1866 WP NP EI Al EC GLibinia dubia 738 WI NE EI EC A GCallinectes ornatus 440 WI NE G EI EC2 A2Neopanope packardii 693 WP NP EI2 EC A GDypanopeus texanus 1217 WI NE EI EC A GEuropanopeus depressus 351 WI Ell ECI A G NE
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 257
Table 5 Discriminant function analysis results for decapod species collected at six sampling sitesduring 1985 and 1986 (all five functions were significant at P lt 000 I and all were included in theclassification process)
Function 4
Eigenvalue 784 241 074 056 022Variance explained () 666 204 63 48 188Scores at group centroids
East Centra] -112 00] -039 -056 -091Northeast -133 -132 057 052 008Atlantic -007 318 046 037 010Gulf 670 -060 -020 -005 004East Interior -144 -016 042 -207 072West Interior -202 002 -251 044 051
DISCUSSION
Most crustacean work in Florida Bay has emphasized the extreme northern andparticularly northwestern bay periphery The fauna described by Tabb and Man-ning (1961) Tabb et a1 (1962a basins channels and banks in the Cape Sable-Sandy Key-Flamingo area) and Rouse (1970 basin just north of Murray Key) isbasically consistent with that of our Gulf site Current studies in Johnson KeyBasin and bordering banks and mangrove islands (M Robblee and J Tilmantunpub1) also indicate a crustacean community similar to that of our Gulf siteHowever Rouses (1970) species list from basin stations straddling our Northeastsite reports a fauna more diverse than we found at the latter A Porpoise Lake(East Central subenvironment) species list (Hudson et aI 1970) reports a faunamuch like that of our Atlantic and East Central sites but some species foundprimarily at our Gulf site are also present (eg Leander paulensis)
Tabb et a1 (1962a) noted that the fauna in the aforementioned Cape Sable-Sandy Key-Flamingo area was more Gulf-Carolinean than Antillean and some-what surprisingly we found this characterization to hold in general for the entirebay Holmquist et a1 (in press) compared the crustacean mud bank fauna tothat of grassbeds in Puerto Rico (Bauer 1985a 1985b 1985c) and Indian River(Florida east coast Gore et aI 1981) and seagrass and other habitats in ApalacheeBay (junction of Florida peninsula and panhandle Hooks et aI 1976) The FloridaBay fauna shared few species with that of Puerto Rico but was similar to theIndian River and Apalachee Bay communities The bays mud bank fauna thusappears to have more temperate than tropical affinities
Table 6 Results of c1assificaiton of individual throw traps using discriminant functions derived fromall traps collected during 1985 and] 986 N = number of traps at each site Diagonal values indicatethe percentage of samples correctly classified at each site Overall correct classification rate = 854
Predicted membenhip ()
East East WestActual group N Central Nonheast Atlantic Gulf Interior Interior
East Central 108 620 167 37 00 167 09Northeast 215 28 916 00 00 14 42Atlantic 108 56 37 898 00 09 00Gulf 90 00 00 11 989 00 00East Interior 54 148 167 00 00 685 00West Interior 54 19 37 00 00 19 926
258 BULLETIN OF MARINE SCIENCE VOL 44 NO 11989
Table 7 Expected and actual number of species by transect and site
Transect West Interior Northeast EaSt Interior East Central AUaotic GulF
ShelteredExpected 14 48 59 71 94 132Actual 2 9 7 19 25 23
TopExpected 11 84 79 92 111 125Actual 4 II 7 14 25 25
ExposedExpected 11 49 58 69 109 136Actual 2 7 8 12 28 23
A trend of decreasing species richness (Table 3) and decreasing occurrence ofGulf and Atlantic suite species (Tables 2 4) is apparent as one progresses fromthe outermost bay sites to West Interior The main chain oflimestone keys to thesouth and east of the bay and the outermost perimeter of mud banks present aformidable circulation barrier (Ginsburg 1956 Tumey and Perkins 1972 Cos-tello et a1 1986) Tumey and Perkins provide a Florida Bay chart indicating anassumed limit oflunar tidal influence which is supported by records oflunar tidalrange at our study sites (Holmquist et a1 in press) We thus hypothesized thatthe mud bank network may constitute a significant obstacle to recruitment of Gulfand Atlantic species into the inner reaches of the bay Costello et aI (1986) havesuggested that the damping of circulation by the keys and mud banks may beresponsible for the paucity of juvenile pink shrimp in the eastern and interiorbay
A more Antillean fauna was found on banks on the Atlantic side of the Keysand along channels leading into Florida Bay (Powell et a1 unpubI) a number ofthese species were also found at the speciose Atlantic site Only occasional in-dividuals ofthe large suite of temperate species present at the Gulf site adjacentto the Gulf of Mexico were found at the Atlantic site and vice versa East Centralclose to but with limited communication with the Atlantic had an intermediatenumber of species including a number found predominantly at Atlantic (Table2) The majority of misclassified Atlantic traps were classified as East Central(Table 6) Northeast and East Interior about 16 and 20 km from open oceanrespectively and separated from the latter by several intervening banks hadsomewhat lower species richness than did East Centra1 With the exception of asingle Panuirus argus at East Interior the occasional Atlantic species were notcollected at these sites West Interior was separated from the southern bay by anadditional major bank system and a virtual wall of islands to the southeast Thisarea has very limited exchange with outlying areas as indicated by salinity dif-ferentials of over 10000 (Schmidt 1979 Powell et a1 unpub1) In tum this wasthe least species rich site with Thor joridanus representing 999 of all individ-uals and an expected number of species of only 21
However Holmquist et aI (in press) point out that restricted circulation isnot of itself an adequate mechanism for explaining the observed progression inspecies richness and suggest that unless a given species suffers a frequent de factoextinction it should eventually be capable of colonizing the bays interior despitepoor circulation These authors suggest cold-induced mortality (Storey and Gudg-er 1936 Storey 1937 Miller 1940 Tabb et a1 1962a Powell et a1 1987) andreproductive strategy (eg pink shrimp migrations from the bay Tabb et aI
HOLMQUIST IT AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 259
Table 8 Geometric means for densities at Northeast during 1984 1985 and 1986 and results ofI-way ANOV As comparing these densities Values with the same superscript were not significantlydifferent at P lt 005 (Student-Newman-Keuls multiple range tests a = 005) ( P lt 005 P lt001)
1984 1985 1986 F
Total decapods 268 2332 9132 1236Alpheus heterochaelis 155 205 233 37Thor jloridanus 009 994 6089 1265Hippolyte pleuracanthus 024 203 173 192Eurypanopeus depressus 026 026 007 56Rhithropanopeus harrisii 016 008 0 85
1962b Costello and Allen 1966) as likely sources of this requisite periodic ex-tirpation
If restricted circulation in conjunction with either cold fronts or other popu-lation reducing mechanisms does in fact limit larval access to the isolated areasof the bay then the degree of tendency to extreme population minima shouldhave been inversely related to penetration into the bay Indeed those species thatwere reduced from sizable populations to very low densities in May (Penaeusduorarum Pericimenes americanus P longicaudatus Latreutes fucorum andTozeuma carolinense Holmquist et a1 in press) demonstrated the least intrusioninto the bay (gt85 of each population found at a single site either Atlantic orGulf Table 2) Conversely those species that maintained substantial populationsthroughout the winter (Alpheus heterochaelis Hippolyte pleuracanthus Thor flor-idanus Paguristes tortugae Pagurus mcaughlinae Callinectes ornatus Neopan-ope packardii Holmquist et a1 in press) were more widespread (only 57-72of a population found at Atlantic or Gulf Table 2) Alpheus normanni appearedto be an intermediate
The fact that species not suffering severe population minima had broad distri-butions again indicates that restricted circulation is of itself an insufficient mech-anism for limiting recruitment to Florida Bay The northwestern Gulf of Mexicocoast provides an example of the inverse relationship that of acute cold butwithout restricted circulation Many species from our Gulf site are also presentat Cedar Key (Marshall 1985) These species suffer severe reductions in densitypossibly due to temperatures averaging 55degC colder than Florida Bay banks inwinter (based on data in Marshall 1985) but rebound during the summer pre-sumably because of unrestricted access of larvae to the site A similar situationis apparent at our Gulf and Atlantic sites where some species were almost extir-pated each year but recolonized the habitat during the summer and fall Thusboth restricted circulation and frequent near extinction are required to producethe most likely explanation for the depauperate Interior and Northeast suben-vironments Because the bay is a nexus for contiguous marine faunal provinces(Antillean and Gulf-Carolinean) and many Atlantic and Gulf species do not pen-etrate past their respective subenvironments Florida Bay has a rather dichoto-mous distribution of decapod fauna
Salinity patterns were probably important in determining patterns of distri-bution as well Our Northeast site had a mean 1983 salinity of about 16000(Whiteand Rosendahl unpubl) followed by a punctuated rise in salinity during our studyfrom a 1984 mean of 24000to means over 30000in 1985 and 1986 (Powell et al1987) Colonization by and an explosive increase of hippolytids at Northeastoccurred in conjunction with this salinity increase (Table 8) (although there were
260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087
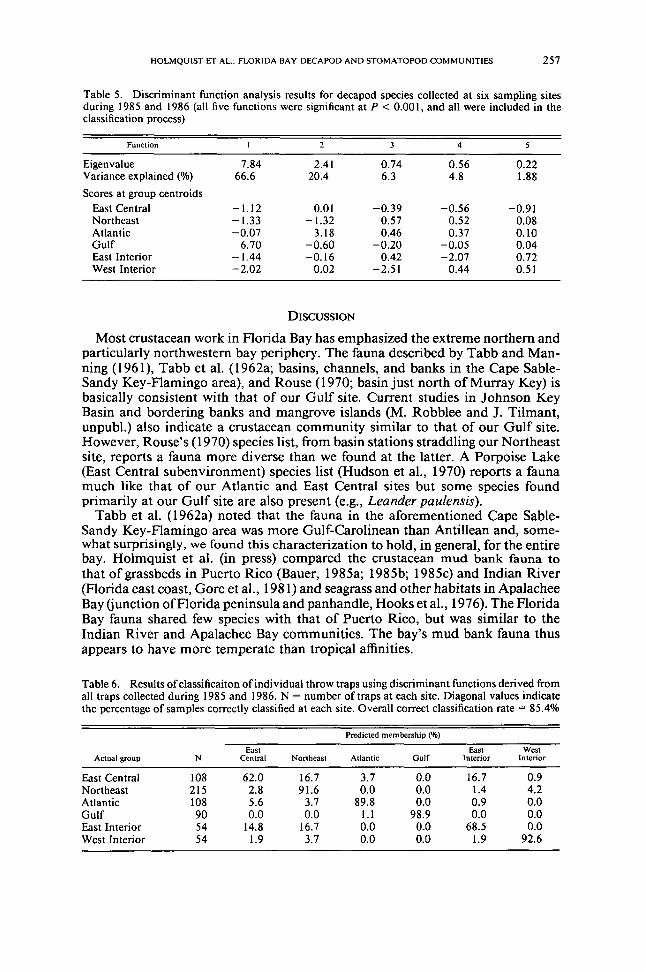
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 257
Table 5 Discriminant function analysis results for decapod species collected at six sampling sitesduring 1985 and 1986 (all five functions were significant at P lt 000 I and all were included in theclassification process)
Function 4
Eigenvalue 784 241 074 056 022Variance explained () 666 204 63 48 188Scores at group centroids
East Centra] -112 00] -039 -056 -091Northeast -133 -132 057 052 008Atlantic -007 318 046 037 010Gulf 670 -060 -020 -005 004East Interior -144 -016 042 -207 072West Interior -202 002 -251 044 051
DISCUSSION
Most crustacean work in Florida Bay has emphasized the extreme northern andparticularly northwestern bay periphery The fauna described by Tabb and Man-ning (1961) Tabb et a1 (1962a basins channels and banks in the Cape Sable-Sandy Key-Flamingo area) and Rouse (1970 basin just north of Murray Key) isbasically consistent with that of our Gulf site Current studies in Johnson KeyBasin and bordering banks and mangrove islands (M Robblee and J Tilmantunpub1) also indicate a crustacean community similar to that of our Gulf siteHowever Rouses (1970) species list from basin stations straddling our Northeastsite reports a fauna more diverse than we found at the latter A Porpoise Lake(East Central subenvironment) species list (Hudson et aI 1970) reports a faunamuch like that of our Atlantic and East Central sites but some species foundprimarily at our Gulf site are also present (eg Leander paulensis)
Tabb et a1 (1962a) noted that the fauna in the aforementioned Cape Sable-Sandy Key-Flamingo area was more Gulf-Carolinean than Antillean and some-what surprisingly we found this characterization to hold in general for the entirebay Holmquist et a1 (in press) compared the crustacean mud bank fauna tothat of grassbeds in Puerto Rico (Bauer 1985a 1985b 1985c) and Indian River(Florida east coast Gore et aI 1981) and seagrass and other habitats in ApalacheeBay (junction of Florida peninsula and panhandle Hooks et aI 1976) The FloridaBay fauna shared few species with that of Puerto Rico but was similar to theIndian River and Apalachee Bay communities The bays mud bank fauna thusappears to have more temperate than tropical affinities
Table 6 Results of c1assificaiton of individual throw traps using discriminant functions derived fromall traps collected during 1985 and] 986 N = number of traps at each site Diagonal values indicatethe percentage of samples correctly classified at each site Overall correct classification rate = 854
Predicted membenhip ()
East East WestActual group N Central Nonheast Atlantic Gulf Interior Interior
East Central 108 620 167 37 00 167 09Northeast 215 28 916 00 00 14 42Atlantic 108 56 37 898 00 09 00Gulf 90 00 00 11 989 00 00East Interior 54 148 167 00 00 685 00West Interior 54 19 37 00 00 19 926
258 BULLETIN OF MARINE SCIENCE VOL 44 NO 11989
Table 7 Expected and actual number of species by transect and site
Transect West Interior Northeast EaSt Interior East Central AUaotic GulF
ShelteredExpected 14 48 59 71 94 132Actual 2 9 7 19 25 23
TopExpected 11 84 79 92 111 125Actual 4 II 7 14 25 25
ExposedExpected 11 49 58 69 109 136Actual 2 7 8 12 28 23
A trend of decreasing species richness (Table 3) and decreasing occurrence ofGulf and Atlantic suite species (Tables 2 4) is apparent as one progresses fromthe outermost bay sites to West Interior The main chain oflimestone keys to thesouth and east of the bay and the outermost perimeter of mud banks present aformidable circulation barrier (Ginsburg 1956 Tumey and Perkins 1972 Cos-tello et a1 1986) Tumey and Perkins provide a Florida Bay chart indicating anassumed limit oflunar tidal influence which is supported by records oflunar tidalrange at our study sites (Holmquist et a1 in press) We thus hypothesized thatthe mud bank network may constitute a significant obstacle to recruitment of Gulfand Atlantic species into the inner reaches of the bay Costello et aI (1986) havesuggested that the damping of circulation by the keys and mud banks may beresponsible for the paucity of juvenile pink shrimp in the eastern and interiorbay
A more Antillean fauna was found on banks on the Atlantic side of the Keysand along channels leading into Florida Bay (Powell et a1 unpubI) a number ofthese species were also found at the speciose Atlantic site Only occasional in-dividuals ofthe large suite of temperate species present at the Gulf site adjacentto the Gulf of Mexico were found at the Atlantic site and vice versa East Centralclose to but with limited communication with the Atlantic had an intermediatenumber of species including a number found predominantly at Atlantic (Table2) The majority of misclassified Atlantic traps were classified as East Central(Table 6) Northeast and East Interior about 16 and 20 km from open oceanrespectively and separated from the latter by several intervening banks hadsomewhat lower species richness than did East Centra1 With the exception of asingle Panuirus argus at East Interior the occasional Atlantic species were notcollected at these sites West Interior was separated from the southern bay by anadditional major bank system and a virtual wall of islands to the southeast Thisarea has very limited exchange with outlying areas as indicated by salinity dif-ferentials of over 10000 (Schmidt 1979 Powell et a1 unpub1) In tum this wasthe least species rich site with Thor joridanus representing 999 of all individ-uals and an expected number of species of only 21
However Holmquist et aI (in press) point out that restricted circulation isnot of itself an adequate mechanism for explaining the observed progression inspecies richness and suggest that unless a given species suffers a frequent de factoextinction it should eventually be capable of colonizing the bays interior despitepoor circulation These authors suggest cold-induced mortality (Storey and Gudg-er 1936 Storey 1937 Miller 1940 Tabb et a1 1962a Powell et a1 1987) andreproductive strategy (eg pink shrimp migrations from the bay Tabb et aI
HOLMQUIST IT AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 259
Table 8 Geometric means for densities at Northeast during 1984 1985 and 1986 and results ofI-way ANOV As comparing these densities Values with the same superscript were not significantlydifferent at P lt 005 (Student-Newman-Keuls multiple range tests a = 005) ( P lt 005 P lt001)
1984 1985 1986 F
Total decapods 268 2332 9132 1236Alpheus heterochaelis 155 205 233 37Thor jloridanus 009 994 6089 1265Hippolyte pleuracanthus 024 203 173 192Eurypanopeus depressus 026 026 007 56Rhithropanopeus harrisii 016 008 0 85
1962b Costello and Allen 1966) as likely sources of this requisite periodic ex-tirpation
If restricted circulation in conjunction with either cold fronts or other popu-lation reducing mechanisms does in fact limit larval access to the isolated areasof the bay then the degree of tendency to extreme population minima shouldhave been inversely related to penetration into the bay Indeed those species thatwere reduced from sizable populations to very low densities in May (Penaeusduorarum Pericimenes americanus P longicaudatus Latreutes fucorum andTozeuma carolinense Holmquist et a1 in press) demonstrated the least intrusioninto the bay (gt85 of each population found at a single site either Atlantic orGulf Table 2) Conversely those species that maintained substantial populationsthroughout the winter (Alpheus heterochaelis Hippolyte pleuracanthus Thor flor-idanus Paguristes tortugae Pagurus mcaughlinae Callinectes ornatus Neopan-ope packardii Holmquist et a1 in press) were more widespread (only 57-72of a population found at Atlantic or Gulf Table 2) Alpheus normanni appearedto be an intermediate
The fact that species not suffering severe population minima had broad distri-butions again indicates that restricted circulation is of itself an insufficient mech-anism for limiting recruitment to Florida Bay The northwestern Gulf of Mexicocoast provides an example of the inverse relationship that of acute cold butwithout restricted circulation Many species from our Gulf site are also presentat Cedar Key (Marshall 1985) These species suffer severe reductions in densitypossibly due to temperatures averaging 55degC colder than Florida Bay banks inwinter (based on data in Marshall 1985) but rebound during the summer pre-sumably because of unrestricted access of larvae to the site A similar situationis apparent at our Gulf and Atlantic sites where some species were almost extir-pated each year but recolonized the habitat during the summer and fall Thusboth restricted circulation and frequent near extinction are required to producethe most likely explanation for the depauperate Interior and Northeast suben-vironments Because the bay is a nexus for contiguous marine faunal provinces(Antillean and Gulf-Carolinean) and many Atlantic and Gulf species do not pen-etrate past their respective subenvironments Florida Bay has a rather dichoto-mous distribution of decapod fauna
Salinity patterns were probably important in determining patterns of distri-bution as well Our Northeast site had a mean 1983 salinity of about 16000(Whiteand Rosendahl unpubl) followed by a punctuated rise in salinity during our studyfrom a 1984 mean of 24000to means over 30000in 1985 and 1986 (Powell et al1987) Colonization by and an explosive increase of hippolytids at Northeastoccurred in conjunction with this salinity increase (Table 8) (although there were
260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087
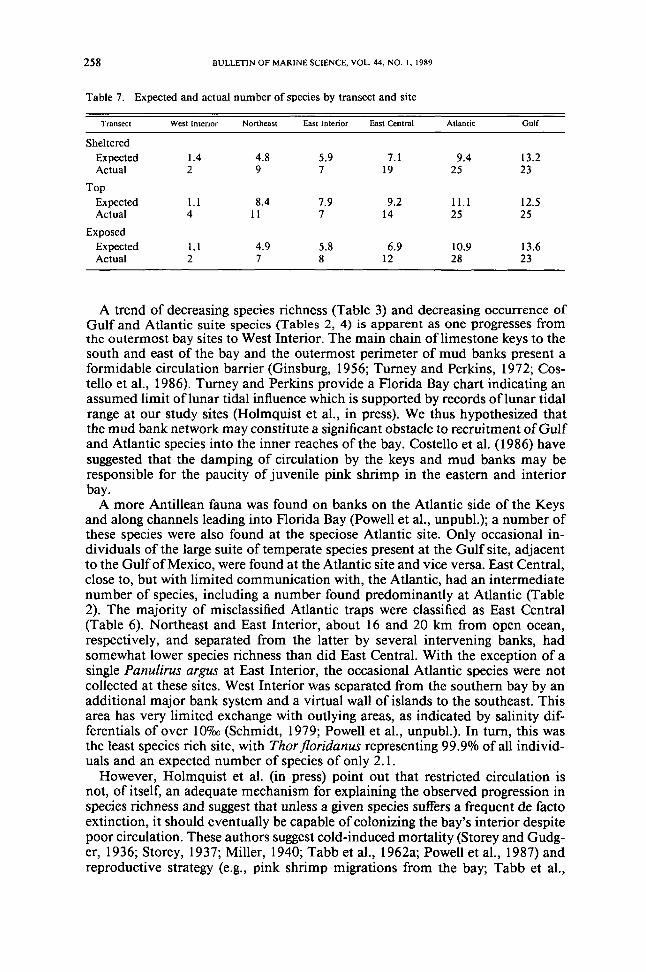
258 BULLETIN OF MARINE SCIENCE VOL 44 NO 11989
Table 7 Expected and actual number of species by transect and site
Transect West Interior Northeast EaSt Interior East Central AUaotic GulF
ShelteredExpected 14 48 59 71 94 132Actual 2 9 7 19 25 23
TopExpected 11 84 79 92 111 125Actual 4 II 7 14 25 25
ExposedExpected 11 49 58 69 109 136Actual 2 7 8 12 28 23
A trend of decreasing species richness (Table 3) and decreasing occurrence ofGulf and Atlantic suite species (Tables 2 4) is apparent as one progresses fromthe outermost bay sites to West Interior The main chain oflimestone keys to thesouth and east of the bay and the outermost perimeter of mud banks present aformidable circulation barrier (Ginsburg 1956 Tumey and Perkins 1972 Cos-tello et a1 1986) Tumey and Perkins provide a Florida Bay chart indicating anassumed limit oflunar tidal influence which is supported by records oflunar tidalrange at our study sites (Holmquist et a1 in press) We thus hypothesized thatthe mud bank network may constitute a significant obstacle to recruitment of Gulfand Atlantic species into the inner reaches of the bay Costello et aI (1986) havesuggested that the damping of circulation by the keys and mud banks may beresponsible for the paucity of juvenile pink shrimp in the eastern and interiorbay
A more Antillean fauna was found on banks on the Atlantic side of the Keysand along channels leading into Florida Bay (Powell et a1 unpubI) a number ofthese species were also found at the speciose Atlantic site Only occasional in-dividuals ofthe large suite of temperate species present at the Gulf site adjacentto the Gulf of Mexico were found at the Atlantic site and vice versa East Centralclose to but with limited communication with the Atlantic had an intermediatenumber of species including a number found predominantly at Atlantic (Table2) The majority of misclassified Atlantic traps were classified as East Central(Table 6) Northeast and East Interior about 16 and 20 km from open oceanrespectively and separated from the latter by several intervening banks hadsomewhat lower species richness than did East Centra1 With the exception of asingle Panuirus argus at East Interior the occasional Atlantic species were notcollected at these sites West Interior was separated from the southern bay by anadditional major bank system and a virtual wall of islands to the southeast Thisarea has very limited exchange with outlying areas as indicated by salinity dif-ferentials of over 10000 (Schmidt 1979 Powell et a1 unpub1) In tum this wasthe least species rich site with Thor joridanus representing 999 of all individ-uals and an expected number of species of only 21
However Holmquist et aI (in press) point out that restricted circulation isnot of itself an adequate mechanism for explaining the observed progression inspecies richness and suggest that unless a given species suffers a frequent de factoextinction it should eventually be capable of colonizing the bays interior despitepoor circulation These authors suggest cold-induced mortality (Storey and Gudg-er 1936 Storey 1937 Miller 1940 Tabb et a1 1962a Powell et a1 1987) andreproductive strategy (eg pink shrimp migrations from the bay Tabb et aI
HOLMQUIST IT AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 259
Table 8 Geometric means for densities at Northeast during 1984 1985 and 1986 and results ofI-way ANOV As comparing these densities Values with the same superscript were not significantlydifferent at P lt 005 (Student-Newman-Keuls multiple range tests a = 005) ( P lt 005 P lt001)
1984 1985 1986 F
Total decapods 268 2332 9132 1236Alpheus heterochaelis 155 205 233 37Thor jloridanus 009 994 6089 1265Hippolyte pleuracanthus 024 203 173 192Eurypanopeus depressus 026 026 007 56Rhithropanopeus harrisii 016 008 0 85
1962b Costello and Allen 1966) as likely sources of this requisite periodic ex-tirpation
If restricted circulation in conjunction with either cold fronts or other popu-lation reducing mechanisms does in fact limit larval access to the isolated areasof the bay then the degree of tendency to extreme population minima shouldhave been inversely related to penetration into the bay Indeed those species thatwere reduced from sizable populations to very low densities in May (Penaeusduorarum Pericimenes americanus P longicaudatus Latreutes fucorum andTozeuma carolinense Holmquist et a1 in press) demonstrated the least intrusioninto the bay (gt85 of each population found at a single site either Atlantic orGulf Table 2) Conversely those species that maintained substantial populationsthroughout the winter (Alpheus heterochaelis Hippolyte pleuracanthus Thor flor-idanus Paguristes tortugae Pagurus mcaughlinae Callinectes ornatus Neopan-ope packardii Holmquist et a1 in press) were more widespread (only 57-72of a population found at Atlantic or Gulf Table 2) Alpheus normanni appearedto be an intermediate
The fact that species not suffering severe population minima had broad distri-butions again indicates that restricted circulation is of itself an insufficient mech-anism for limiting recruitment to Florida Bay The northwestern Gulf of Mexicocoast provides an example of the inverse relationship that of acute cold butwithout restricted circulation Many species from our Gulf site are also presentat Cedar Key (Marshall 1985) These species suffer severe reductions in densitypossibly due to temperatures averaging 55degC colder than Florida Bay banks inwinter (based on data in Marshall 1985) but rebound during the summer pre-sumably because of unrestricted access of larvae to the site A similar situationis apparent at our Gulf and Atlantic sites where some species were almost extir-pated each year but recolonized the habitat during the summer and fall Thusboth restricted circulation and frequent near extinction are required to producethe most likely explanation for the depauperate Interior and Northeast suben-vironments Because the bay is a nexus for contiguous marine faunal provinces(Antillean and Gulf-Carolinean) and many Atlantic and Gulf species do not pen-etrate past their respective subenvironments Florida Bay has a rather dichoto-mous distribution of decapod fauna
Salinity patterns were probably important in determining patterns of distri-bution as well Our Northeast site had a mean 1983 salinity of about 16000(Whiteand Rosendahl unpubl) followed by a punctuated rise in salinity during our studyfrom a 1984 mean of 24000to means over 30000in 1985 and 1986 (Powell et al1987) Colonization by and an explosive increase of hippolytids at Northeastoccurred in conjunction with this salinity increase (Table 8) (although there were
260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087
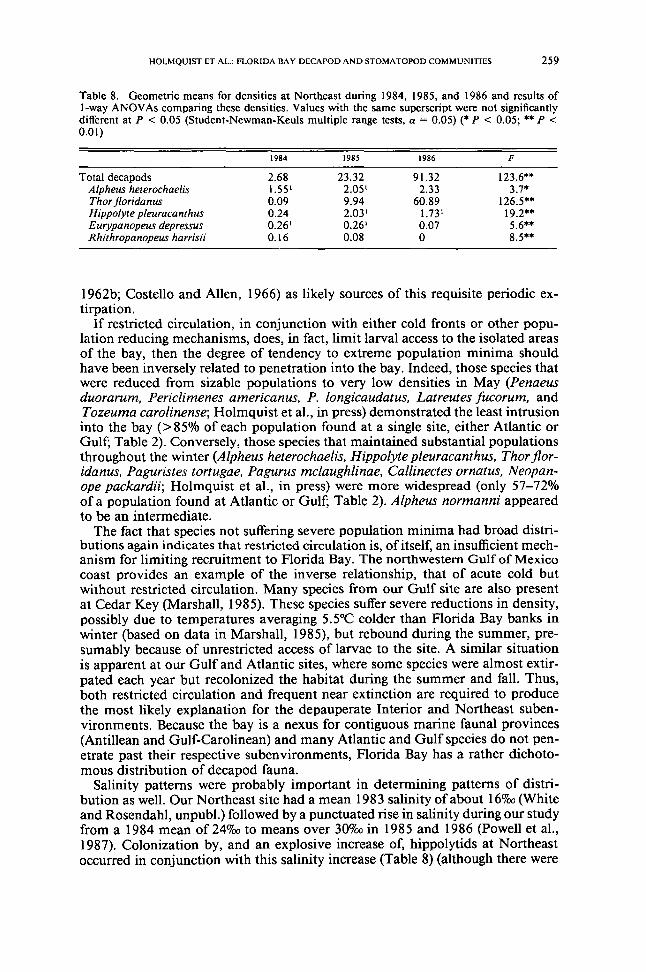
HOLMQUIST IT AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 259
Table 8 Geometric means for densities at Northeast during 1984 1985 and 1986 and results ofI-way ANOV As comparing these densities Values with the same superscript were not significantlydifferent at P lt 005 (Student-Newman-Keuls multiple range tests a = 005) ( P lt 005 P lt001)
1984 1985 1986 F
Total decapods 268 2332 9132 1236Alpheus heterochaelis 155 205 233 37Thor jloridanus 009 994 6089 1265Hippolyte pleuracanthus 024 203 173 192Eurypanopeus depressus 026 026 007 56Rhithropanopeus harrisii 016 008 0 85
1962b Costello and Allen 1966) as likely sources of this requisite periodic ex-tirpation
If restricted circulation in conjunction with either cold fronts or other popu-lation reducing mechanisms does in fact limit larval access to the isolated areasof the bay then the degree of tendency to extreme population minima shouldhave been inversely related to penetration into the bay Indeed those species thatwere reduced from sizable populations to very low densities in May (Penaeusduorarum Pericimenes americanus P longicaudatus Latreutes fucorum andTozeuma carolinense Holmquist et a1 in press) demonstrated the least intrusioninto the bay (gt85 of each population found at a single site either Atlantic orGulf Table 2) Conversely those species that maintained substantial populationsthroughout the winter (Alpheus heterochaelis Hippolyte pleuracanthus Thor flor-idanus Paguristes tortugae Pagurus mcaughlinae Callinectes ornatus Neopan-ope packardii Holmquist et a1 in press) were more widespread (only 57-72of a population found at Atlantic or Gulf Table 2) Alpheus normanni appearedto be an intermediate
The fact that species not suffering severe population minima had broad distri-butions again indicates that restricted circulation is of itself an insufficient mech-anism for limiting recruitment to Florida Bay The northwestern Gulf of Mexicocoast provides an example of the inverse relationship that of acute cold butwithout restricted circulation Many species from our Gulf site are also presentat Cedar Key (Marshall 1985) These species suffer severe reductions in densitypossibly due to temperatures averaging 55degC colder than Florida Bay banks inwinter (based on data in Marshall 1985) but rebound during the summer pre-sumably because of unrestricted access of larvae to the site A similar situationis apparent at our Gulf and Atlantic sites where some species were almost extir-pated each year but recolonized the habitat during the summer and fall Thusboth restricted circulation and frequent near extinction are required to producethe most likely explanation for the depauperate Interior and Northeast suben-vironments Because the bay is a nexus for contiguous marine faunal provinces(Antillean and Gulf-Carolinean) and many Atlantic and Gulf species do not pen-etrate past their respective subenvironments Florida Bay has a rather dichoto-mous distribution of decapod fauna
Salinity patterns were probably important in determining patterns of distri-bution as well Our Northeast site had a mean 1983 salinity of about 16000(Whiteand Rosendahl unpubl) followed by a punctuated rise in salinity during our studyfrom a 1984 mean of 24000to means over 30000in 1985 and 1986 (Powell et al1987) Colonization by and an explosive increase of hippolytids at Northeastoccurred in conjunction with this salinity increase (Table 8) (although there were
260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087

260 BULLETIN OF MARINE SCIENCE VOL 44 NO1 L989
simultaneous but less dramatic increases of Thor florida nus at some of the othersites as well) The brackish-water Eurypanopeus depressus and Rhithropanopeusharrisii (both typically found in 15-20000in Florida Bay Tabb et al 1962a) weresimultaneously virtually eliminated at this site Conversely the somewhat highersalinity at the Interior sites (x = 42000Table 1) may have contributed to thepaucity of species at West Interior However despite the apparent importance ofsalinity in mediation of bank crustacean assemblages the direct relationship be-tween salinity and species richness observed in many other estuarine studies(Remane 1934 Wells 1961 Remane and Schlieper 1971 Gainey and Greenberg1977) is not operative to the exclusion of other factors among our study sites(Tables 2 4) Our least speciose site (West Interior) did not have the lowest salinityof the sites but instead had higher than oceanic salinity Atlantic with virtuallyidentical salinities to those of East Central had much greater species richness thanthe latter site
Various aspects of habitat quality (including gradients in vegetation sedimentand physical factors) are also important in determining decapod character in thebay (Costello et al 1986 Holmquist et aI in press) and may contribute todistribution at the subenvironment level Stepwise multiple regression of a varietyof habitat variables on species richness and population densities indicates thatphysical characteristics particularly tidal range exert more influence on the deca-pod community than do vegetational parameters (Holmquist et aI in press)The importance of tidal range which seems to be closely related to amount ofwater exchange with the Atlantic and Gulf (Ginsburg 1956 Gorsline 1963Scholl 1966 Turney and Perkins 1972) lends further support to our hypothesisof circulation as a major structuring factor Again on a larger scale Atlantic andEast Central with very similar hydrological sediment and seagrass parameters(Table I) and differing primarily in access to tropical Atlantic waters (Fig 1) werestrikingly different with regard to species richness Degree of circulation wasprobably a more important determinant of community structure than were localhabitat characteristics
Most species were least abundant on the exposed sides of most banks presum-ably because of high turbulence and poorly-developed seagrass canopies (Powellet al 1987) on this transect The exposed side however did not have the lowestdensities at East Interior and West Interior These two sites did not have theprogression in seagrass richness sediment structure and organic content foundat most of the other sites (Powell et al 1987 Holmquist et al 1989) The lackof intra-site differences in abundance at these two sites may be the result
Gore et al (I 981) Bauer (I 985a) Howard (I 985) and Marshall (I 985) recordedcaridean densities (69 49 gt 50 and 218 carideansmiddotm-2 respectively) much higherthan those of most previous studies Carideans were numerous on Florida Baysbanks as well with an overall mean of 213 individualsmiddotm-2 Thor joridanusaccounted for 196middotm-2bull At West Interior Tjoridanus had a density of884middotm-2bull
Caridean abundances also appear to be very high in the bays basins (M Robbleeand J Tilmant unpubl) Although we believe that this abundance is in part afunction of the efficiency of the throw trap in this environment (Kushlan 1981Sogard et al 1987) it is clear that Florida Bay mud banks provide an exceptionalhabitat for grass shrimp despite the harsh physical characteristics of the mud banksystem
ACKNOWLEDGMENTS
This project was very labor-intensive ATP expenditure courtesy of L Taylor B McKelvy JNegyesi J Fourqurean P Racsko 1 Prentice P Hinchcliff and 41 volunteers from Earthwatch andthe Center for Field Research We are grateful to F Chace J Garcia-Gomez R Lemaitre and A
HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087

HOLMQUIST ET AL FLORIDA BAY DECAPOD AND STOMATOPOD COMMUNITIES 261
Williams for kindly identifYing representative specimens of some species to J Zieman B FelgenhauerD Hoss and B Hettler for generously loaning equipment and to K Hainge-Holmquist and R Darrahfor other assistance Discussions with S Berkeley M Fonseca J Kenworthy M Robblee D Rob-ertson D Tabb G Thayer J Tilmant and J Zieman were very helpful as were the anonymousreviewers N Paul was our tireless typist Special thanks to H Powell for her constant support Wewere supported with grants from the South Florida Research Center (National Park Service CX5280-3-2339) the Everglades Protection Association and the Center for Field Research
LITERATURE CITEDAllen D M and T J Costello 1966 Releases and recoveries of marked pink shrimp Penaeus
duorarum Burkenroad in south Florida waters 1958-64 Data Rep Fish Wild Servo US 112microfiches
Bauer R T 1985a Diel and seasonal variation in species composition and abundance of carideanshrimps (Crustacea Decapoda) from seagrass meadows on the north coast of Puerto Rico BullMar Sci 36 150-162
--- 1985b Penaeoid shrimp fauna from tropical seagrass meadows species composition diurnaland seasonal variation in abundance Proc BioI Soc Wash 98 177-190
--- 1985c Hermit crab fauna from seagrass meadows in Puerto Rico species composition dieland seasonal variation in abundance J Crust BioI 5 249-257
Bert T M J Tilmant J W Dodrill and G E Davis 1986 Aspects of the population dynamicsand biology of the stone crab (Menippe mercenaria) in Everglades and Biscayne National Parksas determined by trapping South Florida Research Center Report SFRC-8604 77 PP
Brook I M 1978 Comparative macrofaunal abundance in turtlegrass (Thalassia testudinum) com-munities in south Florida characterized by high blade density Bull Mar Sci 28 212-217
Costello T J and D M Allen 1966 Migrations and geographic distribution of pink shrimpPenaeus duorarum of the Tortugas and Sanibel grounds Florida Fish Bull 65 449-459
--- --- and 1 H Hudson 1986 Distribution seasonal abundance and ecology of juvenilenorthern pink shrimp Penaeus duorarum in the Florida Bay area NOAA Technical Memoran-dum NMFS-SEFC-161 1-84
Davis T D 1980 Peat formation in Florida Bay and its significance in interpreting the recentvegetational and geological history of the bay area PhD Dissertation Pennsylvania State Uni-versity State College Pennsylvania 316 pp
Davis G E and J W Dodrill 1979 Marine parks and sanctuaries for spiny lobster fisheriesmanagement Proc Gulf Carib Fish Inst 32 194-207
--- and --- 1989 Recreational fishery and population dynamics of spiny lobsters Panulirusargus in Florida Bay Everglades National Park 1977-80 Bull Mar Sci 44 78-88
Enos P and R D Perkins 1979 Evolution of Florida Bay from island stratigraphy Geo SocAmer Bull Pt I 90 59-83
Fleece J B 1962 The carbonate geochemistry and sedimentology of the keys ofF1orida Bay F1oridaMS Thesis Florida State University Tallahassee Florida 112 pp
Gainey L F and M 1 Greenberg 1977 Physiological basis of the species abundance-salinityrelationship in molluscs a speculation Mar Bio 40 41-49
Ginsburg R N 1956 Environmental relationships of grain size and constituent particles in somesouth Florida carbonate sediments Bull Amer Assoc Petrol Geo 40 2384-2427
Gore R H E E Gallaher L E Scotto and K A Wilson 1981 Studies on decapod Crustaceafrom the Indian River region of Florida XI Community composition structure biomass andspecies-areal relationships of seagrass and drift algae-associated macrocrustaceans Estuar CoastShelf Sci 12 485-508
Gorsline D S 1963 Environments of carbonate deposition in Florida Bay and the Florida StraitsPages 130-143 in R O Bass and S L Sharps eds Symposium on shelf carbonates of the ParadoxBasin Four Corners Geological Society Denver Colorado
Heck K L Jr G van Belle and D Simberloff 1975 Explicit calculation of the rarefaction diversitymeasurement and the determination of sufficient sample size Ecology 56 1459-1461
Higman 1 B 1952 Preliminary investigation of the live bait shrimp fishery of Florida Bay and thekeys Mar Lab Univ Miami Coral Gables Fla Report to Fla State Bd Conserv ML 37898 pp
Holmquist J G G V N Powell and S M Sogard In press Decapod and stomatopod assemblageson an unusual system of seagrass-covered mud-banks in Florida Bay Bull Mar BioI
--- --- and --- 1989 Sediment water level and water temperature characteristics ofFlorida Bays grass-covered mudbanks Bull Mar Sci 44 348-364
Hooks T A K L Heck Jr and R J Livingston 1976 An inshore marine invertebrate communitystructure and habitat associations in the northeastern Gulf of Mexico Bull Mar Sci 26 99-109
Howard R K 1985 The trophic ecology of caridean shrimps in an eelgrass community AquatBot 18 155-174
262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087

262 BULLETINOFMARINESCIENCEVOL44 NOI 1989
Hudson J H D M Allen and T J Costello 1970 The flora and fauna ofa basin in central FloridaBay US Fish Wild Servo Spec Sci Rpt-Fish 604 1-16
Idyll C P E S Iverson and B Yokel 1964 Movements of juvenile pink shrimp in the EvergladesNational Park Florida (Abstract) Proc GulfCaribb Fish Inst 16 51-52
-- -- and -- 1966 Abundance of juvenile pink shrimp on the Everglades NationalPark nursery grounds Circ Fish Wildl Serv Wash 246 19-20
Kushlan J A 1981 Sampling characteristics of enclosure throw traps Trans Amer Fish Soc 110557-562
Marshall M J 1985 Stability in an assemblage of caridean and penaeid shrimps inhabiting anintertidal Thalassia bed PhD Thesis University of Florida Gainesville Florida 138 pp
Miller E M 1940 Mortality of fishes due to cold on the southeast Florida coast 1940 Ecology21 420-421
Powell G V N S M Sogard and J G Holmquist 1987 Ecology of shallow-water habitats inFlorida Bay Final Report NPS contract CX5280-3-2339 South Florida Research Center Home-stead Florida 405 pp
Recher H F and J A Recher 1968 Comments on the escape of prey from avian predators Ecology49 560-562
Remane A 1934 Die Brackwasserfauna Verh Dt Zool Ges 7 34-74-- and C Schlieper 1971 Biology of brackish water Wiley-Interscience New York 372 ppRouse W L 1969 (1970) Littoral Crustacea from southwest Florida Fla Sci 32 127-152Schmidt T W 1979 Ecological study of fishes and the water quality characteristics of Florida Bay
Everglades National Park Florida Project Report RSP-EVER N-36 South Florida ResearchCenter Homestead Florida 145 pp
Scholl D W 1966 Florida Bay a modem site of limestone formation Pages 282-288 in R WFairbridge ed The encyclopedia of oceanography Reinhold New York
Sogard S M G V N Powell and J G Holmquist 1987 Epibenthic fish communities on FloridaBay banks relations with physical parameters and seagrass cover Mar Ecol Prog Ser 4025-39
-- -- and -- 1989 Spatial distribution and trends in abundance of fishes residing inseagrass meadows on Florida Bay mudbanks Bull Mar Sci 44 179-199
SPSS and M J Norusis 1986 SPSSPC+ SPSS Inc Chicago 643 ppStorey M 1937 The relation between normal range and mortality of fishes due to cold at Sanibel
Island Florida Ecology 18 10-26-- and E W Gudger 1936 Mortality of fishes due to cold at Sanibel Island Florida 1886-
1936 Ecology 17 640-648-- and R B Manning 1961 A checklist of the flora and fauna of northern Florida Bay and
adjacent brackish waters of the Florida mainland collected during the period July 1957-Septem-ber 1960 Bull Mar Sci II 552-649
Tabb D C D L Dubrow and A E Jones 1962a Studies on the biology of the pink shrimpPenaeus duorarum Burkenroad in Everglades National Park Florida Fla State Bd ConservTech Ser 37 1-30
-- -- and R B Manning 1962b The ecology of northern Florida Bay and adjacentestuaries Fla State Bd Conserv Tech Ser 39 1-79
Taft W H and J W Harbaugh 1964 Modem carbonate sediments of southern Florida Bahamasand Espirito Santo Island Baja California a comparison of their mineralogy and chemistryStanford Univ Publ Geol Sci 8 1-133
Tumey W 1 and B F Perkins 1972 Molluscan distribution in Florida Bay University of MiamiRosenstiel School of Marine and Atmospheric Science Miami Florida Sedimenta III 37 pp
Wells H W 1961 The fauna of oyster beds with special reference to the salinity factor EcolMonogr 31 239-266
Zieman J C J W Fourqurean and R L Iverson 1989 Distribution abundance and productivityof seagrasses and macroalgae in Florida Bay Bull Mar Sci 44 292-311
DATEACCEPTED February 22 1988
AFFIuATIONNational Audubon Society Research Department 115 Indian Mound Trail TavernierFlorida 33070
ADDRESSES (JGH) Department of Biological Science Florida State University Tallahassee Florida32306 (G VNP) National Audubon Society Research Department 115 Indian Mound Trail Taver-nier Florida 33070 (SMS) Rutgers Marine Field Station PO Box 278 Tuckerton New Jersey08087
Top Related
Copyright © 2022 FDOKUMEN

