







Bahasa
Halaman
Hukum



"STUDIES ON THE CHROMOSOM'ES OF C·ERTAIN INSECTS WITH SPECIAL REFERENCE
TO THE EFFECT OF DRUGS'"
A THESIS FOR THE PH. D. DEGREE CF'
BHOPAL UNIVERSITY, 8HClPAL 1975
BY
MISS PUS UP A SURlY ASTAV
JR. RESEARCH FELLOW, U. G. C.
MOTIJ.AL VIGYAN MAHAVIDYALAYA
BHOPAL INDIA

I I I I
! ,
I
i i
f
Department of Zoology Motilal Vigyan Mahavidyalaya
Bhopal (M.P.)
Oertified that Miss Pushpa Shrivastav, Junior
Research Fellow, U.G.C., has worked under my guidance on
the "Studies on the chromosomes of certain insects with
special reference to the effect of drugs" for the Ph.D. ,
Degree of Bhopal Unlversity. She worked in the researoh
laboratory of the Department of Zoology, Motllal Vigyan
Mahavidyalaya, Bhopal, for more than 200 days. This
thesis represents her original findings and has not been
submitted either partly or fully, for any other degree
in any other University.
Miss Shrivastav joined the leoturership in
Zoology, Oollegiate Branoh of Eduoation Department,
Government of M.P., late in 1974 and is submitting the
thesis while in the same oadre. The thesis is in a
presentable form and is ready for examination. It ful
fils the requirements presoribed by the Bhopal University,
Bhopal.
( P. Jaitly )
Assistant Prof. of Zoology
Motilal Vigyan Mahavidyalaya,
Bhopal (M.P.).

ACKNOWLEDGEMENTS
I gratefully aoknowledge the oompetent
supervision of Dr.(Miss) P.Jaitly. who has been a
oonstant souroe of inspiration to me. Her keen
interest in my progress rejuvenated me in my days of
desperation. I am thankful to Dr.R.C.Chourasia, the
Head of the Department of Zoology and the then
Prinoipal, Dr.S,N,Kaveeshwar for permitting me to
work in the Department. The author is also thankful
to Dr.K. C.Mehra, Head of the Anatomy' Department,
Medioal College, Bhopal, for permitting photography
in his Department.
The author is also thankful to the members
of the staff of the Zoology Department, Motilal Soience
College, for their cooperation, and to DR.G.P.Mukerji.,
now the Head of the Zoology Department, Government
Narbada College, Hoshangabad, M.P., for his suggestions
in the rearing of the grasshoppers. Thanks are also due
to my research oolleague, Mias Shail Saxena, for her
help during the oompilation of the thesis.
The author is especially thankful to
Prof. S.P.Ray-Chaudhuri for his suggestion to inolude
the banding pattern in the oaryologioal study. His
student Dr. Lalji Singh helped too by lending me the
literature on banding pattern and giving his valuable
suggestions from time to time. The study of banding
pattern was carried out at the Cytogenetios Laboratary

of Benaras Hindu University, for which the permission.
was kindly given by Dr.T.Sharma and friendly help
extended by his research students. All this is
gratefully acknowledged.
Thanks are due to the British Museum ot
Natural History for getting the specimens identified.
I am much beholden to my father, Prof.P.N.Shr1vastav,
who has taken keen interest in my academic career and
has always been a source of inspiration to me.
Lastly, the financial assistance of
University Grants Commission, New Delhi, is grateful17"
acknowledged for awarding the Junior Researoh Fellowship
to me.
( P.SHRIVASTAV )

CONTENTS
I Introduction.
11 Literature Survey.
II! Material.
IV Experimental Procedure and Methods.
A. Germinal Tissue (testes).
B. Somatic Tissue (gastrio caeca).
C. Drugs.
D. Division-Cycle and Drug-treatment.
E. Banding Pattern.
V Observations.
A. Caryologioal study :
1. Trilophidia annulata Thunberg.
a. Meiosis.
b. Abnormalities in the
Natural Population.
(i) Extra fragment.
(ii) Polyploid oells.
(iii) Anaphase bridge.
(iv) Chromosome breakage
o. Mitosis.
2. ChrotogonuB trachypterus Blanchard.
a. Meiosis.
b. Abnormalities in the Natural Population.
(i) 'Supernumerary Chromosomes.
(ii) Polyploidy.
1 - 6.
1 - 23.
24 - 28.
29 - 34.
29
29 - 30.
30 - 31.
31 - 33.
33 - 34.
35 - 93.
35 - 67.
35 - 43.
35 - 4;).
41 - 42.
43.
43 - 53.
43 - 44.
44 - 53.

(iii) Non-disjunction.
(iv) Understained regions.
(v) Chromatid breakage.
(vi) Extra fragment.
c. Mitosis.
3. Leva indica Bo1iver.
a. Meiosis.
b. Abnormalities in the Natura.l Population.
(i) Extra. fragment.
(ii) Polyploid cells.
(iii) Abnormal spindle.
(1v) Breakage.
c. Mitosis.
4. Attraotomorpha crenulate. 'abrlcius.
a. Abnorme.lities in the Natural Population.
(i) Chromatid breakage.
(1i) Chromatid separation.
b. Mitosis.
5. Poekilocerus ~ictus Fabrioius.
a. Mitosis
b. Banding pattern
B. Effeot of Drugs :
1. Quantitative Study: Caffeine.
2. Qualitative Study: Caffeine.
a. Control Series.
b. Treated Series
53.
53 - 58.
55.
55 - 57.
57 - 58.
53 - 61.
60 - 61.
61.
61 - 67.
62 - 63.
63 - 67.
67 - 92.
67 -78.
78 - 82.
79.
79 - 82.

(i) Mitotio ohanges. 79.
(ii) Ohanges in the Meiotic prophase stages. 79 - 80.
1) High condensation.
2) Lesions.
(iii) Changes at metaphase I ill. 80 - 82.
1) Polyploidy.
2) Olumping.
3) Spindle abnormality.
(iv) Changes at first and second post-metaphase stages.
1) Breakage.
3. Qualitative study : Enteromyoetin.
(i) Mitotio ohanges.
(ii) Changes in the meiotio prophase stages.
1) Breakage.
2) Polyploidy.
82.
82 - 86.
82 - 83.
(iii) Ohanges at metaphase I ·1 11. 83 - 84.
1) Fragmentation.
(iv) Ohanges at first and second post-metaphase stages.
1) Stiokiness.
2) Spindle abnormality.
4. Qualitative Study: Formalin.
(i) Mitotio ohanges.
(ii) Changes in the meiotio prophase stages.
84 - 66.
86 - 90.
86 - 87.

1) Deohromatization and breakage.
2} Interbivalent oonneotion.
3) Breakage.
4) Clumping.
(iii) Changes at metaphase I ~ 11.
1) Inter-bivalent oonneotion.
2) Destruotion of morphology.
3) Lesions.
4) stretohed oentromsre and dechromatization.
5) star-shaped oonfigurations.
(iv) Changes at first and second post-metaphase stages.
5. Qualitative study: Niootine
(i) Mitotio changes.
1) Fragmentation.
(ii) Changes in the meiotio prophase stages.
1) Breakage.
2) Clumping.
(iii) Changes at metaphase I & 11
(iv) Changes at first and seoond post-metaphase stages.
VI Results and Discussion.
A. Meiosis and Mitosis.
B. Banding Pattern of P. Eictus.
C. Abnormalities in the ~atural Population.
D. Effect of Drugs.
87 - 90.
90.
90 - 92.
90.
91.
92.
92.
93 - 125.
93 - 104.
104 - 108.
108 - 117.
117 - 125.

VII Summary. 126 - 132.
A. Caryological study 126 129.
B. Abnormalities in the Natural Population. 129 - 130.
c. Banding Pattern of P. ,Eiotus. 130 - 131
D. Effeot of Drugs. 131 - 132.
VIII Bibliography. 133 - 163.
IX Explanation of Figures. 164 - 132.
X Abbreviations. 183 - 1135.

I
INTRODUCTION

-1-
INTRODUCTION
The chromosomes of grasshoppers have always
been popular with the nuolear cytologists. They enjoy the
topmost position with respeot to the suitability of mate
rial among the invertebrates. They have large germ and
somatio cells with correspondingly large chromosomes.
Moreover, the extreme olarity of preparation and readily
available divisional stages in their testes. make the
grasshoppers a olassical material for cytological studies.
The faot that India is very rioh in grasshopper
fauna is a boon to our oytologists. Considerable work has
been done on this fauna by various Indian soholars. still,
the studies in India oannot be regarded as very extensive
when we take into consideration other parts of the world,
especially U.S.A., Japan and Australia.
A survey of studies on grasshopper chromosomes
reveals that research in morphological aspeots of chromo
somes is waning beoause of the current interest in mole
cular cyto-chemistry. The latter branch has become rich
with information in the last deoade. still, the morpholo
gical stUdies cannot be regarded as orthodox owing to
their relation to the modern systematios.
In faot, the modern systematists regard the
cytogenetioal information as an essential feature because
the speoies are considered to be the objective reality of
some particular genetic continuity. In a good many cases,

where the species could not be distinguished on purely
morphological grounds, help was sought from the fields of
cytology, genetice, ecology, embryology and biochemistry.
The works of Hughes Schrader (1948), Patterson and stone
(1952), White (1954),Darlington (1956), Makino (1956),
Swanson (1958), Lewio and John (1963), Hew1tt (1964), John
and Lewis (1965), Manna and Mazumder (1961,s), Hsu and
Benirsohke (1967, 1968), John and Hewitt (1968), White
(1968, 1970) and others, advocate the usefulness of cytology
in the taxonomy of plants and animals. The various cytolo
gical 'toolst, with the help of which systematists establish
homology and analogy between allied species and groups, are
- analysis of caryotype, diploid number, metrical data of
metaphase chromosomes, existence of specialized chromosomes,
behaviour and qrrangement of chromosomes during cell division,
speciality of some oytoplasmic and nuclear bodies and hybri
dization results, etc. (Manna, 1969). Thus a nuclear cyto
logist provides an independent set of data for establishing
the inter-relationships of different groups. And the study
of cBryotypes is not limited merely to the counting of
chromosome number and morphology. but also helps in under
standing the cytogenetic basis of speciation.
The cytological data of Indian grasshoppers are
very limited. In many cases the in~ormation is incomplete
and unevenly scored. The most neglected aspect is the somatic
oaryologioal st~dies. It was, therefore, the author's
earnest desire to take a plunge in this indifferently
oharted ooean. The work includes the morphological studies

-3-
of both germiD!:ll Dnd s01llc1.tic chromosomes.
An attempt is here made to study the effect of
a number of chemicals also, viz., caffeine, nicotine,
formaline and enterornycetln. The importance of testing for
mutagenic effects of substances that are used as preserva
tives or are coneti tuents of hump...ll food/medicine has been
stressed by Bald.une (1956). All the four chemicals are
important from this point of view. Caffeine is used for
both cualitetive a.nd quantitc.tive studies, while formaline,
enteromycetin a.nd nicotine are used for qualitative studies
only.
The study of chromosomal p.berra.tions by chemi
cals has a special sienificapce and immense possibilities.
First, chromosome fragmentation followed by translocation
of some fragments may give rise to a new pattern of ohromo
some segments re-su.1ting in heritable phenotypio difference.
The latter is oov1ously i,ll.portant in the evolution of
new speoies and desirable varieties. Seoondly, the study
of causes and consequenoes of aberrations is fundamental
to our understanding of many important biological problems.
Chromosome breakage is perhaps the most important cause
of the death of cancer cells following irradiation. The
chromosome-breaking property of chemicals has an important
bearing on the ohemotherapy of cancer too. Here it is
pertinent to mention that the effects of not all the
chemicals used in the present work have been reported
in higher vertebrates. However, there is a basic similarity
in the mechanism of replication of the genetic material in

-4-
all forms of life. The same faot allows one to draw para
llel conolusions to a great extent, irrespective of the
animal type experimented upon. Lastly, the study of
chromosomal aberrations by means of chemioals also helps
in bringing about the differential nature of ohromosome
segments.
The above-mentioned aspeots need a thorough and
systematio effort in investigation. A survey of the availa
ble literature makes it quite apparent that though a host
of workers have been practising the induction of ohromoso
mal aberrations by chemical agents, very few tried to do a
thorough and systematic investigation. The author tried
her beat, therefore, to state the facts as systematically
as possible and hopes to bridge a few gaps in the archi
tecture of relating information.
Though the study on the morphology is being made
and effect of ohemicals Observed still with enthusiasm,
another trend has been set in during the more reoent years.
Cytologists have been trying to identify single ohromosomes
in animals and plants. Their eff'orts resulted in a variety
of chromosome identification methods and their applications.
Unfortunately the investigators could not keep in touch
with each others' works due to the delay in publioation.
This brought about a lag in the organization of further
research and also often duplication of work. To overcome
these difficulties the various investigators gathered toge
ther in the 23rd Nobel symposium held on 25-27 September,
1912. The proceedings of this meeting are beautifully compile~

-5-
by Caspersson and Zeoh (1973) in their book -
"Chromosome Identifioation : Teohnique and Applioations
in Biology and Medicine".
In this new era, the various workers oonoentrated
on human ohromosome studies with the intention of linking ~)
speoific dideases with distinct sites on chromosomes.
Entire populations were screened through automatic maDns to
detect or prediot inherited abnormalities. The honour of
first outstanding discovery goes to Caspersson !1 al (1968)
who observed a peculiar pattern of fluofescence in the
chromosollles of Vici~ faba and Trillium erectum following
quinacrine mustard staining. The chromosomes when viewed
under ultraviolet light, fluoresced or glowed in alternate
light and dark bands, named as Q-bands. Later on, the
same method was applied to human and animal chromosomes
as well. The 'banding pattern' studies have now become a
'vogue' in nuclear oytology and have triggered a rush on ch
romatid firle structure analysis all over the world.
Now a greater range of methods is available
( Hsu, 1972 ). A number of chemicals, other than quinacri
ne and its derivatives, oan also produce bands. These bands
are either identioal with or different from the fluore-
scent ones and are named as - C-, G-, wld R-bands.
The banding pattern study has been undertaken
in grasshopper chromosomes also but the reports are very
scanty. An attempt is made to contribute in this field
with whatever means available. This work, thus, touches

-6-
a variety of fields, viz., morphological studies of
germinal and somatio ohro:nosomes, effect of ohemicals
and the banding pattern.
The unlirr;ited hardships un.dergone to complete
the work will be rewarded. if it is found to have some
value to the disoerning.
))J))J)))J
)3°

11
LITERATURE SURVEY

-7-
LITERATURE SURVEY
The Orthoptera are frequently quoted as one of
the classic examples of karyotype stability. Cytologically
they have been extensively studied and each major group
exhibits a chracteristic pattern. Amongst the two
taxonomical groupings of Acrididae, Cryptosaoci have typi
oally 2n=23 and Chasmosaooi 2n=19 aorooentric chromosomes.
Cyto-taxonomical oharacterizations of the different
subfamilies or genera of shorthorned grasshoppers have
great limitations beoause of the orthodox ohromosome
number and meiotio pattern.
A good number of scientists have reported on
grasshopper chromosomes from different corners of the world.
The available literature relating to this work is surveyed
by the author under a number of headings, namely, general
( orthodox ) oaryological study; unorthodox ohromosome
behaviour including (a) structural heterozygosity for
unequal and asymmetrical bivalents, (b) supernumerary
chromosomes, (0) heterochromatization leading to polymor
phism, (d) polymorphism due to change in the charaoteristic
spermatogonial number, (e) neo-XY sex chromosomes, and
(t) spontaneous meiotic aberrations; parthenogenesis;
ohemioally induoed aberrations; telocentricity and finally,
the banding pattern.
The orthodox pattern in the oaryology of Aori
doidea has been reported by a host of workers. The Indian
fauna was paid attention to by - Asana (1934), who published

,/
-8-
a series of papers on the chromosomes of IndiBn grassho
ppers; Rao (1937) oompared the oaryo1ogy of eight genera
of Pyrgomorphini; Asana ~ 81 ( 1938, 1939 ) made a
chromosomal survey of Indian speoies belonging to Pami1y
Aorididae. A oomparative study was undertaken by Ray
Chaudhuri and Dutt (1947) in three genera. A cytologioal
ingestigation of Ph10eba sp., Tristis Eu1vinat~ and
Chrotogonus sp. was done by Dutt (1948 s,b; 1950). Manna
gave a oomprehensive aocount of meiosis in a number of
oriental speoies ( Manna, 1954, 1967 a, 1969 ). Srivastava
( 1954, 1956 ) reported the ohromosome behaviour in
Chroto~onus inoertus and Dissosteira oarolina. Sharma!!!±
,I (1963e) reported the variation in the mean ohiasma --' frequenoy per nuoleus in the wild populations of Chrotogonus
traohyPterus. Ohatterjee (1971) reported the orthodox
meiotic pattern of 0ela uvarovi and Xenocatant09s humi1is
humi1~s and again of Euprepocnemis roseus, Ceracris
def1orata, Aulacobothrus 1uteioeps and Hieroglyphus banian
(Ohatterjee et aI, 1971). --Orthodox pattern has been reported in Aoridoidea
from other parts of the world also. McClung (1902, 1905,
1914 ) conoentrated on the behaviour of the spermatocyte
ohromosomes, but his aocount was somewhat unsatisfactory
in the se~se that he oould not state the multiple sex
ohromosome meohanism distinotly. Metrioal ?~alysis of
ohromosomes, chiasma frequency study, etc. ( Ooleman,
1943 ) yielded supportive evidences which helped in

-9-
establishing the relationship of some species. A detailed
account of the cytology of Oxalis disper was reported by
Marks (1957). A comparative study of male and female
meiosis in the grasshopper Stethophyma grossum showed that
both the frequency and distribution of chiasmata in the
oocytes were Quite different from the male situation (Perry
and Jones, 1974). Besides these individual efforts, groups
of scientists scanned the populations of particular regions.
The British population was through1y investigated by Hewitt
(1964, 1965), John and Hewitt (1966, 1910) and Hewitt and
John (1968, 1972). The grasshopper fauna of Japan was
studied by Asana, Makino and Niyama (1938, 1939), Hareyama
(1941), Makino and Momma (1950) and Makino (1956). White
and co-workers paid attention to the African, American
and Australian populations and released a series of exce
llent pob1ioations ( White, 1940, 1941b, 1953, 1954, 1956,
1957 a,b,o, 1961, 1963, 1965, 1966, 1968, 1970; White, Mesa
and Mesa, 1967; ~~ite, Cheyney and Key, 1963; White, Carson
and Cheyney, 1964; White and Cheyney, 1966; White and Key
1957d; "~ite and Andrew, 1960, 1962; White, Lewontin and
Andrew, 1963; White, B1aokith, Blackith and Cheyney, 1967,
and Atchley, 1974).
The various studies were facilitated by an
improvement in the methods of handling the hereditary
material ( Darlington, 1960, Sharma and Sharma, 1972 ) ."
and an understanding of the mode of evolution of chromo-
) somes in the Orthoptera ( White, 1941 b, 1954; Ray-............... , ..
Choudhuri and Manna, 1951; John and Hewitt, 1968).

-10-
Most of these reports oentred around morphome
trio data, oomparative study, behaviour of autosomes and
sex ohromosomes. meiosis and ohiasma formation, eto.
Recently an interesting report had been given
by Nur (1973) in his work on the random arrangement of
ohromosomes in the radial metaphase oonfiguration of
Melanoplus femurrubrum. The report offers several advan
tages for studying ohromosome assooiations. Any treatment
whioh ohanges the position of the chromosomes, is also
expected to destroy the radial oonfiguration. Thus, it is
possible to ohoose for study only unaffeoted oells.
In the animal world, natural heterozygotes have
always been attraoting the attention of oytologists. They
play a major role in oaryotypic evolution. The structural
rearrangements, whioh are oonsidered as ohromosomal mutation
are generally detectable oytologically. A l::;rge fraction
of them remains unaocounted because they are likely to
produce inviability and sterility. Natural seleotion plays
its part in seleoting the superior adaptive ones from the
structural heterozygotes and gives rise to a polymorphio
population. Chromosomal polymorphisma h:_ve been studied
with regard to para- and peri-oentrio invession. translo
oation, deletion, duplioation and non-disjunotion. Such
studies mainly oentred around the salivary gland ohromoso
mes of dipterans ( Drosophila and Rhynohosoiara ) in the
beginning. But, since the germinal ohromosoH.es of
Drosophila are unsuit8ble for meiotic study, attention was

-11-
diverted later to the germinal chromosomes of grasshoppers.
The nutural heterozygotes in grasshoppers have been very
efficiently reviewed by White ( 1954, 1956. 1951 a,b,o,
1961, 1963, 1966; White et aI, 1960, 1962, 1963, 1966, 1961).
Several instances of unorthodox ohromosome behaviour have
been reported in about forty speoies of grasshoppers by
Manna (1969).
The oocurrence of supernumerary chromoso~es was
reported in Aiolopus sp. ( Ray-Chaudhuri and Manna, 1951;
Ray-Chaudhuri and Guha, 1955 ). in Tagarta indica (Manna,
1954 ), in Myrmeleotettix maoulatus ( John and Hewitt,
1965 a,b; Barkar, 1966; Hewitt and John. 1967 ). in
Chrotogonus traohypterus ( Sharma,. Parshad and Gupta, 1965)
in Loousts migratoria ( Rahiman and Rajasekarasetty, 1967a)
in Chorthippus paral1e1u~ ( Hewitt and John, 1968 ), in
Aorotylus humbertianu6 ( Gururaj and Rajasekarasetty, 1971)
and in Aorida lata and Oedaleus infernalis ( Takizawa and
Narasawa, 1971 ). It was established earlier ( John and
Hewitt, 1965a and b) that British populations of the mottled
grasshopper Myrme1eotettix maculatus were polymorphic with
respeot to the presence of B-ohromosomes. In one population
10-70 ~ of the individuals possesed 1-3 supernumerary
elements while none in the other population. This condition
results in the occurrenoe of four caryotypes, namely,
S ( standard diploid oomplement ), 5+1, S+2 and S+3. The
cytogenetio systems of grasshoppers and loousts were
reviewed by John (1913), giving speoial referenoe tothe

-12-
origin and evolution of supernamerary segments. The namber
of B-chromosomes varied from 0-7 in the Japanese population
of Attractomorpha bedeli ( Sannomiya, 1973 ). The effect I
of supernamerary heterochromatilU'iwas observed in three I
species by Schroeter and Godfrey (1974).
The deviation in the characteristio spermatogo
nial number towards the lower ones due to the centric
fusion of two aorocentrio elements had been enoountered by
a number of workers. Besides, studies on the sex chromosome
mechanisms of the Aoridoidea revealed the existenoe of
many speoies in whioh the original XO : XX mechanism has
been replaoed by an XY IXX one, beoause of a centrio fusion
between the original X and an acrocentric autosome ( V'I'hi te,
1940, 1941b; King, 1950; RaY-Ch+udhuri and Guha, 1952; I
Manna and Chatterjee, 1963; White and Cheyney, 1966;
Manna, 1967a; White, Mesa and Mesa, 1961; Mesa and Mesa,
1961; Hewitt and John, 1972).
Newly arisen XI meohanisms are of considerable
genetic interest. One may expect to find in them a series
of genetio transformations, namely, evolutionary hetero
chromatization of ohromosome regions, acquisition of new
differential segment and perhaps, development of new
mechanisms of dosage compensation. It is evident from the
available report that the newly arisen XI mechanisms are
not a long :1term evolutionary success. Instead, they tend
to oocur in isolated species or small groups of closely
re la ted speoies. There is not a single genus in which 8.11

/'
-13-
the members have neo-XY sex chromosome mechanisms.
The X1X2Y : XI X1X2X2 systems have also been
reported in a few short-horned grasshoppers ( King and
Beams, 1938; White, 1940, 1941b, 1953; White and Cheyney,
1966; Mesa and Mesa, 1967 ). This system is an advanoe
over the XI : XX one and has arisen thro~gh an additional
Y-autosome fusion. It is also s short-lived failure from
the evolutionary view-point. On the other hand, the
X1X2Y system of praying mantids. is a oonspicuous evolu
tionary suooess, and has arisen through a mutual translo
oation in an XO form without passing through a neo-XY stage
( White, 1940, 1941a, 1965; Hughes-Sohrader, 1950, 1953 )
Among the Tettigonoides, five speoies have so
far been reported to possess the multiple sex chromosome
mechanism. One is the "Mormon Cricket" - Anabrus simplex
( Decticinae ), reported by McClung ( 1902, 1905, 1914 ).
The other two are an unidentified species of Isopsera --( Phaneropterinae ) with XY males and Letana atomifera
( Tettigoninae ) with X1X2Y males. These two belong to
India and were reported by Dave ( 1965 ). Another two,
Theodoria me1anocnemis ( Phaneropterinae ) and Yorkiella
piots ( Listrosoelinae ) were reported by White, Mesa and
Mesa (1966).
-----Polyploidy in the natural oondition is not an
infrequent phenomenon. Meiosis of the diploid and
tetraploid spermatooytes was studied by Ray-Chaudhuri
and Bose (1948) in Attractomorpha speoies. John and

-14-
Henderson ( 1962 ) reported polyploidy in ~sto~erca
Earane~.Sharma, Prasad and Gupta ( 1965 ) found
scattered polyploid cells in four individuals of
ghroto~onus trachg,pterus. Tetraploidy was reported in
PyreomorEha bispinosa by Nirmala and Rajasekarasetty (1971) ,
while the chromosome complement of Plrgomorpha kraussi
was found to be 22-ploid ( Lewis and John, 1959 ).
Parthenogenesis, as is apparent from the acanty literature,
is not as frequent a phenomenon. White, Cheyney and Key
( 1963 ) reported a case with complex structural hetero
zygosity.
Literature on spontaneous meiotic aberrations
in grasshoppers is quite rich. The association between
two nonhornologous chromosomes in Gesonia punctifrons
was reported by Ray-Choudhuri and Manna ( 1950 ). In
the Bryodema species White ( 1954a ) noticed an extreme
form of chiasma localization. Anomalous asoooiations and
the behaviour of pseudo-bivalents was studied in the
meiosis of two inter - specific hybrids of Bromus
( WaIters, 1954 ), where the chromosomes seemed to be held
together merely by chromosome matrix connections. White
( 1951c ), while releasing a series of papers on the
cytogenetics of Moraba scurra, reported heterozygosity
for "elastic constrictions" in that species. Srivastava
( 1959 ) came aoross Et number of spontaneous meiotio
aberrations while studying the spermatogenesis of
Poeki1ocera Eiate. Kayano and Nakamura (1960) in their

-15-
ohiasma study, reported a heterozygote for El translooa
tion in Acridt! lata. An inversion heterozygote was obser
ved by Lewontin and White ( 1960 ) in Moraba sourra.
Calliptamus palaestinensis possessed an unequal bivalent
( Nur, 1961 ). Ray-Choudhuri and Sherma , 1962 } observed
dioentric bridge and cleavage in certain grasshoppers. A
large number of aberrations were noted by Sharma, Pr6sad
and Bedi ( 1962, 1963 ) in their study on the populations
of Chrotogonus traohypterus. They reported suppression of
paChyten~airing, anaphase bridges, anaphase laggards,
polyploidy, chromosomal variability, unequal bivalents and
translooation heterozygotes. and discussed various aspects
of the breakdown of mitotic and meiotic stability. Adap
tive properties of inversion were reviewed in Moraba
sourra ( White, Lewontin and Andrew, 1963 ). A spontane
ous interchange was marked in Chorthippus brunneus with
extensive ohiasma formation in an interstitial segment
( John and Hewitt, 1963 ). A spontaneus chromosome brea
kage was reported in a grasshcpper of British population,
and in an undescribed species of Morsbinee; and its meio
tio oonsequences were studied by Lewis and John ( 1966 )
and White ( 1966 ) respeotively. l!;volutionary implioations
of pericentric inversions in Austrioce-tes interioris were
disoussed by Nrulkivell ( 1967 ). In Metrioptera brachyptera
a prolonged non-homologous association resulted in the
formation of a pseudomultiple ( Southern, 1961a ). A -'-' survey of 'vie,tiQa' group of morabine grasshoppers revealed

-16-
the occurrence of a number of spontaneous meiotic aherra
tions ( White, Blackith, Blackith and Oheyney, 1967 ). A
male grasshopper, Oamnula pellucida, was found to be
heterozygous for a paracentric inversion ( Nur, 1968 ).
Nirmala and Rajasekarasetty ( 1971 ) reported a number of
chromosome abnormalities in the wild populations of
Plrgomorpha bispinosa.
All the above referred casea of polymorphism
have been found to be playing a key role in the evolution
of caryotypes in animal speciation. Their role in the
cellular adaptability has also been proved to be extremely
important for the preparation of in vitro cell lines as
well as with regard to prognosis of cancer.
Since the frequency of spontaneous aberrations
is very low, the meohanism of structural roarrangements
and chromosomal evolution is understood better from the
indu.ced aberration studies. ;tIuch of our information regar
ding the nature, mechanism and extent of induced changes
in the chromosomes has co~e from the study of pl~~t
material. It h1=Js now gradually become apparent that there
are certain important differences betWeen the reactions
of the plant and animal chromosomes, a-t least so far as
ohemical treatments are concerned. For example, very many
different chemicals can induce ohromosome breakages in the
former ( Sharma and Sha~~a, 1960 ) whereas there are only
a few chemicals which call produce trlle ohromosome breaka
ges in animal chromosomes as distinct from stickiness and
its consequences. A part of' this dif'f'srence in behaviour

i I f
-17-
may be due to metabolic differences in beheviour in the
cell and nuolear membranes of anima.l and plants, but
nothing definite is known. Therefore it seems desirable
to have more data of a quantitative kind on the induced
chromosome changes seen in animal cell divisions immedia-
tely following treatment.
The neoessary impetus for research on the effects
of outside agencies on chromosomes was provided by the
discovery of X-ray induced mutation in Drosophila by
Muller ( 1921 ), In the meantime, the polyploidising
action of oolchicine and its effective application on
plants had been disclosed through the works of Blakeslee
and Avery ( 1937 land their collaborators. The result of
this Ilenthusiasm led to the disoovery of mutagenic action
of nitrogen mustard in Drosophila,. Since then various che
ffiicals and drugs have been reviewed from time to time.
Work on grasshopper chromosomes was very limited in the
sixties ( Gaulden and Carlson, 1951; Dutta. Gupta and
DivakEr8n, 1959; Rao; 1960 and others ). With the ad~ent ,
of the. seventies, the literature gradually became recher
and richer with the contributions of Manna and co-workers, (\.,
and few others - Carlson and Gaulden ( 1964 ), Manna and
Mazumder ( 1964 ), Manna and Roy ( 1964 ), Manna and
Purida ( 1965 a,b ), Desai, Sirsi, Shankarf;lppa and Kasturi
Bai ( 1966 ); Manna and Lahiri ( 1966 ); Manna and
Mukherjee ( 1966 ); Jain and Singh ( 1967 ); Manna (1967a); -'---Manna and Parida ( 1967 ); Wilson and Friedkin ( 1967 );

-18-
r;~anna and Perida ( 1968 ) ; Klassen, Chang and Eide (1969);
!VIanna and Parida ( 1970 ) ; Bhunya and Manna ( 1971 ) ;
Manna Fmd Bhunya ( 1971 ) ; Manna and Parida ( 1971 ) ;
Mue11er, Gaulden and Drane ( 1911 ); Manna and Bhunya
( 1972 ); Bhattacharya ( 1973 ); Bhunya and Acharya (1973);
Bhunya and Das ( 1973 ); Nirmala and Rajasekarasetty
( 1973 ); Pati and Bhunya ( 19'73 ); Saha ( 1973 ) ;
Saha and Chatterjee ( 1973 ); Bhattacharya and Dab
( 1914 ); Bhunya, Farida and Ghosh ( 1974 ); Saha and
Chatterjee ( 1914 ); and Saha and Khudabakeh ( 1974 ).
The varipus chemicals tested by these soientists
were - (1) antibiotios - erythromycin, oh1oram.pheniool,
bTiSeofu1vin, novobiocin, st~eptomyoin, tetracyoline eto.;
(2) hormones - corline, insulin, ovocyo1ine, testoveron
eto.; (3) phenols - dihydroxy-oha1oone, gallio aoid,
parabenzoquinone eto.; (4) ohelating agents - oupferron,
EDTA, versene etc.; (5) a1oohos - ethyl aloohol, methyl
aloohol, t-butyl a1oohol; (6) nuoleic aoid analogues -
BUDR, IUDR, maleio hydrazide; ('7) metal1io salts -,
aluminium oh1oride, oaloium chloride, merourio iodide,
merourio nitrate, potassium cyanide; (8) alkaloids -
oolohioine, a.trophine; (9) tranquiliser - largaoti1;
(10) formaline; (11) arginine; (12) semioarbazide;
(13) apho1ate; (14) urethane, etc.
A survey of the findings obtained after using
the above-mentioned ohemioals revealed an interesting
oolleotion of generalizations - (1) feeding method of

-19-
administration was found to be quantitatively more effective
than the injection method, as evidenced from the comparative
aberration frequency study. Qualitatively, however, the
effects were the same in both cases and they were (a) in the
spindle appara.tus - manifested by fusion of cells, polyploi
dy, failure of spindle formation or the destruction of the
spindle, and (b) in the chromosome - manifested by failure
of synapsis, reduction of chiasma, C-meiotic metaphase, star
or ball configurations, lagging chromosomes, failure of ori
entation, sticky effect, etc. (2) The meiotic chromosomes of
grasshoppers did not have equal responses to some particular
chemicals. (3) Some chemicals affected the X-chromosome more
than the autosomes, while no preferential effect had been
observed with other chemicals. (4) The effect of most of the
chemicals on the chromosomes of different stages of division
appeared to be non-specific while corline had been found to
be stage sensitive since it damaged more specifically the
anaphase chromosomes. (5) The breaking point in the X-chro
mosome followed more clearly at diplotene-diakinesis stages
indicated that breaks were more common to the 2/5 and 4/5
segments. (6) There is inter-specific sensitivity with re~a
rd to X-rays as well as chemicals. (7) The aberration fre
quency data of the X-chromosome indicated some cumulative
effect when the specimens were treated with a mixture of
maleic hydrazide and aluminium chloride and the results were
compared with those of separately treated ones. The results
sug;ested thDt they might be locus specific in action.

-20-
(8) Some chemicals tested for the dosage-frequency relation
ship did not yield positive result. (9) The time of first
appearance of aberration va.ried considerable. This is
expected since the mode of action of various chemicals is
different.
The above stu.dies have yielded u.seful information
on the architecture and the mode of action of the genes,
chromosomes and cells. Chromosome breakages perhaps lie
at the basis of the chain of reactions which follow whole
body irr'adiation leading to the death of experimental
animals. In the present age, when we intend to exploit
atomic energy -are ,essential for an intelligent planning
for the protection of the hereditary material from the
hazards of radiation ( Ray-Choudhuri, 1961 ).
There has always been a confusion regarding the
position of the centromere in grasshopper chromosomes.
The strongly preva.iling view was that chromosomes with
strictly terminaloontromeres could not survive in nature.
Probably the first person to suggest and adopt this view
was Ne.vashin (1916). After him this remained the generally
accepted view. The school of thought which rejected the
concept of telocentricity argued that all stable rod
chromosomes were acrooentrics. It was believed that the
minute heads situated at the centric ends of rod chromo
somes clearly demonstrated the existence of short arms
( White, 1935; Coleman, 1943). Thus,the occasional formation
of ditactic biva.lents ( Dgrlington, 1936 ) could then be

-21-
explained only by the association of these very small arms.
It was also pointed out that when telocentrios either
arose spontaneously ( Upcott, 1937 ) or following X-irradia
tion ( Rhoades, 1936 ) through the fraoture of an inter
stitial centromere, their unstable behaviour usually led to
their rapid elimination.
However, as the techniques of chromosome analysis
improved over the years, there has been a gradual rise in
the data which supported another school of thought. The
latter considered that not all rod chromosomes are
acrocentrics. Works of Schrader (1939), Ostergren (1947)
and Lima-De-Faria (1956) made it clear with some degree
of certainty that for many rod chromosomes the so-called
"small arms" represent fused centromeric chromosomes.
There are a number of exmples of derived telocentrics in , I
plants ( ~arlington and La Court 1950; Marks, 1957a;
Strid, 1968 ). The various confusions regarding the
nomenclature of centromeric position were skilfully dealt
with by Levant Fredga and Sandberg (1964 ). John and
Hewitt ( 1966 ) suggested that the production of ditac
tic bivalents could be the result of a chiasma actually
forming within the terminal centromeres of homologous
rods.
White and some of his co-workers claim that in
animals the prevailing rod choromosome is an acrocentric.
They are of the opinion that misdivision leading to the
establishment of stable telocentrics h.as not contributed

-22-
in any significant way to the evolution of animal chro-
mosomes. John and colleagues, however, repeatedly ques-
tione4 such beliefs, because their own studies on
Orthopteran species have clearly revealed the existence
of telocentrics ( Southern, 19618.; John and Hewitt. 1966,
1968 ). Nur ( 1968 ) also reported stable telocentric
chromosomes in Myrmeleotettix maculatus. The only
debatable point concerns their origin. Unlike White,
these workers have not ruled out centric fission as a
possible pathway.
still there are scholars who are unwilling to
8.ccept that stable telocentrics may exist in the normal
chromosome complement in certain species. The probable
reason for this attitude is that it would demand a
radical change in their outlook concerning caryotype
evolution. It would particularly necessitate a reapp
I'a.isal of the role that centric fission may have had in
this process.
Lastly, coming to the banding pattern, its
discovery not only gave new impetus to research but has
actually rendered all previous findings obsolete. The
present developments in the chromosome technology make
the old preparations look like a palaeolithic research
tool.
The techniques are immensely important for the
fundamental approach to the organization of the ;gene
systems and on its mode of work in cell differenciation

-23- •
and oell funotion.
The banding teohniques not only make an invaluable
oontribution to the fields of medicine and biology, but
also to evolution ( Grouohy !!!!!.' 1913, and Pearson, 1973).
The information available so far mainly centres around
mammalian subjects and has been compiled, as previously
stated, by Caspersson and Zeoh ( 1973 ). One paper,
however, deals with the banding pattern of heterochromatio
B-chromosomes of M,p:meleotettix maculetus ( Galaghar,
Hewitt and Gibson, 1973 ). Lateral subunits in the C-bands
of unreplioated ohromosomes of Dis:;;,osteira carolina were
studied by Bregman ( 1913 ). The location, structure and
behaviour of C-heterochromatin was reported in Dichroplus
silveira~idoi (Cardoso et aI, 1914 ). An unusual
behaviour of the heteroohromatio blocks looated in the
so-oalled synaptio region of the sex bivalent ( Neo Y-
Neo X ) was found to remain paired from early prophase
through metaphase I. Brown and Wilmore ( 1974 ) reported
the looation of repetitious DNA in the chromosomes of the
desert looust Schistooeroa gregaria using a modified giemsa
staining teohnique. Klaste.rska., ~atarajan and Ramel (1974)
utilized Q-banding and C-banding techniques to olarify
the relationship between heteroohromatin, centromere and
ohiasma localization in Bryodema tuberculata. The author
has also made a hesitant attempt to step in this territory
and hopes that the present study will add a little bit to
the existing knowledge on the subject.

III
MATERIAL

· M/M : rmmnn !
19 .1
M/M i rmmnn ~
F1g.2
"nTmmn j
Fi g . 3

-24-
MATERIAL
A. SPECIES
The following species of short-horned grasshoppers
belonging to the family Acrididae ( Orthoptera : Acridoidea)
constitute the material for the present study:
(l) Tril0;Ehidia annulate Thunberg.
(2) Chrotos:onus trachypterus Blanchard.
( 3) Leva indica Boliver.
(4) AttraotomorEha crenulata Fabr1oius.
(5) Poekilooerus Eictus Fabric1us.
TriloEhid1a annulata I
( Fig. 1 ) The specimen belongs to the sub -
family Oedipodinae. In winter, the adults invariably gather
around the carrot plants, while the nymphs conceal them
selves in between the branohes. In summer and rainy
eeasons they are found scattered in the grass. Adults are
capable of short flights.
Chrotogonu6 trachypterus I
( Fig. 2 ) Commonly oalled sand-hopper, this
specimen belongs to the sub - family pyrgomorphinae.
The males can easily be distinguished from the muoh stouter
females. The males can flyover short distances but the
females only hop because of their heavy bodies.
Leva indica z
( Fig. 3 ) This specimen belongs to the sub
family Gomphocerinae. Both the male and female adults are
fast fliers. The nymphs are aotive too. The adult males

f'"
-25-
make a oharaoteristic 'ohirping' sound with their hind
legs.
Attraotomorpha orenulata :
( Fig. 4 ) This slender grasshopper belongs to
the sub-family Pyrgomorphinae. The adults are not very
aotive and oan hop over short distanoes. A oharaoteristio
oolour ohange is observed in this speoimen. During the
hotter months it aquires a dull, buff - brown oolour,
while during the oold and moist weather it has a fresh
green oolour. Suoh oolour polymorphism is not a rare
phenomenon in the inseot world ruld is directly related to
temperature changes.
Poekilooerus piotus :
( Fig. 5 ) This large speoimen measuring about
c' 5-7 mm. belongs to the sub-family pyrgomorphinae. The
pigment is red, yellow and green in the nymphal stages
and turns to prominent green patches on yellow baokground
in the adults. When irritated, the insect seoretes an ill
smelling liquid.
B. COLLECTION
All the speoimens were oollected from the bota
nical garden of Motilal Scienoe College,Bhopal except
Poekilocerus piotus. The latter was oollected from the
Gwalior University oampus. One population of Chrotogonus
traohypterus was also available at the Arera Colony locality
of Bhopal from where individuals were ocoasionally oolleoted
for examination and oomparison with the Science College
population. Both the populations are widely separated.

M/M r
fTlllllTTTl ~
Fig. 4
\
\
/
~ o 10 10 ]0
Fig. 5

-26-
Since the activity of the grasshoppers varies
directly with the temperature, collecting was easiest
when the day warmed enough so that they could easily be
flushed out of the grass, but when it was hot they began
taking flight too quickly to be caught with a net. Nymphs
could easily be caught by hand more easily than by net.
Poekilocerus pictus was available from May to
August. Rest of the grasshoppers were available through
out the year.
C. REARING
The grasshopper Chrotogonus traohypterus was
reared by the ~ethod of Carlson and Gaulden (1964). No
long - term rearicg was required for the rest of the looal
grasshoppers. Adults and last instar nymphs were colleoted
directly from the field and kept in the cages until requi
red for examination. As Poekilocerus piotus ({id not belong
to the local inseot - fauna, it had to be reared from the
early nymphal inste,rs. Every possible attempt was made
to provide natural surroundings to the insects.
D. C.flGES
The cages used were similar to the type descri
bed by Carlson and Gau1den ( 1964 ), me2suring 23 om. high
x 18 om. wide x 15 cm. deep ( Fig.6 ). For Poekiloc~
pictus larger oages were provided, measuring 15 om. high
x )0 om. wide x 25 cm. deep. (lig.7). About 10 - 15
adults were plaoed in one oage.

F1g.6
Fig. 7

-21-
E. FOOD
The nymphs and adults of Poekilocerus pictus
were fed with the leaves of the lutex - seoreting plant
Calotropis of the family Asclepidiaceae. Other grassho
ppers were given a variety of food, including, wheat
seedling, carrot tops and fresh grass soaked in water.
The grass was placed inside the cage in a ata.ining jar
filled with water. The grasshoppers were often seen to
ingest drops of water from the grass.
F. TISSUE
A review of' ava.ila.ble literature makes it
obvious that much attention has been paid to the study
of ohromosomes of the germinal tissue, and that too of
the male grasshoppers. The caryology of the somatic tisDue
has been largely ignored. However, about a deoade ago,
an excellent study was undertaken by Carlson and Gaulden
( 1964 ). They utilized to good advantage the large
neuroblasts in an early embryonic stage of the grazshopper
Chrotophaga viridifasoiata. There are, however, oertain
limitations of the neuroblast teohnique. In order to have
a oontinuous supply of young embryos throughout the year,
it is neoessary to maintain a healthy stock of individuals
in oages under controlled conditions. Besides, the oulture
teohnique itself is very elaborate and involves the use of
various delicate and expensive equ1pments.
Fox ( 1969, 1970 ) used abdominal fit body and

-28-
malpighian tubules in his report on the DNA va~ues in
somatic fiscues. These tissues, are, however, extremely r""
delicate and need to be handled by leaving it attached to
the abdominal sternite and the gut, respectively, during
fixation and staining.
In the present study gastric caeca huve been
used for SOIru:itic caryologioal study. Though a very useful
tissue, its use seems to be rather limited. Nankivell
( 196'1 ) used the gastric caeca of the females of
Austriocetes interioris to locate polymorphism for
perioentric inversions in the sex chromosomes. Nanda
( unpublished ) used the same tissue in her study on the
somatic chromosomea of a cockroach. The author used the
tissue in her report on the caryology of three speoies of
short horned grasshoppera ( ShrivastCi.v, 1975 ).
The testes of either lest instar n~ph or young
adul t were taken for the study of meiosis and bfUldirig
pattern.
.)-0-0-0-0-0-0 0-0-0
o

IV
EXPERIMENTAL PROCEDURE
AND METHODS

-29-
EXPERINTENTAL PROCEDURE A;':D M.F~THODS
A. Germinal Tissue (teste~) :
Testes of the pr-e-hdul t and adult male,:;rasGho
ppers were dissected out in invertebrate saline and kept
in 1% Sodium citrate for 15 - 20 minutes. Aceto-alcohol
( 1 : 3 ) was used as fixative. Staining was done accor
ding to the schedule followed by Mangalangi and su~ra.manium
( 1963 ), using 0.5% aqueous Heidenhain's haematoxylin.
Temporary slides were sealed with vcseline for observation.
The schedule was sli~htly modified for making the slides
permanent. After removing the seal with xylene, the slides
were inverted in a large petri - dish containing absolute
butanol and a piece of glass rod. The whole set-up was
covered with another petri - dish of lar5er diameter.
After the separation of the cover - ;~lass ( which required
15 minutes to 24 hours ), mounting was done in D.P.X.
B. Somatic Tissue ( gastric caeca ) :
The orthodox squash techni<1ue proved inadequate
for the routine analysis of the somatic tissue and, there
fore, an air - drying technique ( Crozier, 1970 ) was
adopted after some alterations. The technique may be
summarised as follows :-
1. Actively feeding nymphs were injected with J.2 ml. of
0.1% colchicine prepared in Ringer Solution 'A'.
2. The nymphs ware dissected in 1% Sodium citrate after
4 hours. The gastric caeca were removed while still
................ -----------------------

-JJ-
attached to a portion of the gut and were kept in fresh
hypotonic solution for 10 - 20 minutes.
3. The tissue was fixed in aceta-methanol ( 1 : 3 ) for
30 minutes.
4. A small piece of tissue was transferred to a drop of
60% acetic acid ( aqueous ) on a clean warmed slide. This
brought about a rapid dissociation of the cell.
5. A drop of aceta - methanol was added to the preparation
and the slide was tilted in HII directions to achieve a
maximum spread of the li(~uid.
6. The slide was stained in 2% aceto-c8rmine, mounted, and
placed in a covered container inside an oven. It was left
for 3 hours or until microscopic81 exmination revealed a
satisfactory staining.
7. The slides were placed vertically in acetic-ethanol
to loosen the cover-glasses.
8. After dehydration in 95% and absolute ethanol, and
clearing in xylene, the slides were mounted in D.P.X.
Originally devised for ants ( Order Hymenoptera ),
this method reC'luires very small amount of lnateriCiI. A regular
and complete fla.ttening of a dispersion of cells is obt&ined
without the use of mechanical means. The slides can be
m~1de permanent with rninimel cell loss.
c. Dru'Is:
Following concentratioi:Js were prepared in
invertebrate saline fit the time of treatment :-
1. Caffeine :-:).1%, :).2-', :).3%, 0.4%, ·).5fo, ).61', J. 7~t

-31-
J.d>b t ;).9%, and 1. (w/v).
2. Formaline:- 3.0,0 (v/v).
3. Enteroll2l.oetin ( ohlorbmphellicol cl3psules ) :- O.25}b (w/v).
4. Niootine:- J.5% ( v/V).
The drugs were introduced into the body of the
insect by injection. The needle was inserted through the
coxal cavity of the meta-thorasic leg, arId ,).2 ml. solution
was allowed to escape the syringe.
D. Division - Cycle and Drug - treatment :
To seoure the effect of chemicals on the chromo
somes, various artificial means are adopted to secure
maximum availability of metaphases - t~ stage at whioh the
detection of aberrations is most convenient. The alkaloid
~olchicine, e.g., and its derivative colcemid are commonly
used to arrest the division at metaphase. Ap(~trt from the
advan tages, hO'Never, these agen ta can induoe a number of
aberrations either independently or in combination with
the chemical, the effect of which is to be investigated.
It is difficult, if not impossible, to deiect and differe
ntiate the 'independent' effect from the 'combination'
effect. Moreover, the use of such means would complicate
the quantitative analysis of the effect of the ohemical.
No metaphase - arresting agent was therefore, used. along
with the chemic21s.
In the present investigation, the effect was
studied on the IDcle germ cells of Chrotogonus trachl~terus
because sperme.togenesis in the testes tubules of this

-32-
species oocurs in an orderly menner. The large chr'omosomes
Hre also well - suited for analysis, The approximate
duration of various divisional Gtages was determined in the
schedule of spermatogenesis ( Fig. a ) according to the
instrnctions given by Subramanium ( 1971 ). This schedule
allowed tentatively to deduce the time during the pre -
meiotic cell - stage when the chrom080meu were exposed to
the chemical.
Thirty - five days old gras.:.::;hoppers were chosen
for experiment. In the first lot, a sst of five grl.:isshoppers
each, were treated with the ten concentrations of ca.ffeine.
Controls were maintained for eaoh set. This experiment helped
to seleot that concentration which was most effective. This
effective concentration of caffeine was then used in the
second lot to study its rospo.ase to the increase of tre::: t
ment time. For this experiment, the testes were fixed at
4, Id, 48 !lnd 72 hours and 6, 7, 8, 14 and 21 dbYs.
In the third, fourth and fifth lots, the selected
concentrations of formaline, rticotine and enteromycetin
were uc'ied Eind the testes were fixed at the above - mentioned
:timings. The tisGue was proceeDed &5 described e'· rlier for
rsermint·d tissue. bU.t the hypotonic tre8.tment was given
for one hour.
Chromosome figures were se:lected for analysis
under low ma~lification ( 10 X objective J.Only those
figures v.ere chosen that could be reliably analysod by
using oil immersion. The cell stages analysed were diakinesis.

-- --26 DAYS------
i PI?IMORDIAL MI SPERMATlDS
I SPERMS
I SPERMATOGONIA PREMEIOTIC PROPHASE I! SPERMATO
I SPERMATOGON IAL GENES:JS DIVISIONS
lE 8DAYS I DAY ·9DAYS
SCHEQULE OF SPERMATOGENSIS IN
Fig.8
DIAGRAMMATIC REPRESENTATION OF THE METHOD OF COUNTING METAPHASES IN THE SQUASH OF
A TESTIS-FOLLICLE
Stages other than metaphase.
Metaphases.
Path follo",e d by the ocular micr·;meter (by moving the stage of the microscooe).
1. Slide. 2. Cover - glass.
3. Area covered by the checKered ocular micrometer at a pertlcular focus.
4. Initial position of ocular micrometer. 5. Ultimate position of ocular m!crQmeter~
Fig. 9

-33-
metaphl:;sc I, and anaphGse 1. Chromosomal abnormalities
observed in spermc.togonial metaprtases are not reported
pecBuse of uncertainty about the exact stage of the mitotic
cell - cycle during which treatment occurred. The spermato
gonial mitoses require only 1-2 days hod the srunpling in
this investigation was too infrequent to give a reliable
sensitivity of ohromosomes throughout the mitotic cycle.
The meiotic divisions, on the other hand, take only a
short time ( 1 to 2 days ) to complete. Therefore, the data
from these stages of' meiosis were pooled and the two daughter
oells of enaphase I were counted as one cell in the tabula
tion.
The cells were oounted with the help of' a cheque
red ocular miorometer n.nd the slide wa.s thorouehly scanned
by a method represented diagrammatically in Fig. 9.
E. Banding Pattern
The testes of Pocki1ocerus pictus were fixed in
acetic-ethanol ( 1 I 3 ) for 1 hour. The tissue was then
transferred to 60% acetic acid for About 15 - 20 minutes.
This brought about breaking down of the tissue. Aggregates
of cells settled at the bottom of the tube. The tissue was
spun at 1)00 r.p.m. for 5 minutes. Afterwards the super
natant was drained out with the help of a pipette. Freshly
prepared fixative was added to the tube and again the tissue
was spun at 100) r.p.m. for 5 minutes. The supernatant was
removed and the air- dried prepp,rations were made with the
rem2ining fluid.

-34-
The slides were stored for 24 hours and then the
procedure followed wee according to Sumnerts (1912)
technique with a few altera.tions :-
(i) hydrolysis in 0.2 NHol was carried out for 30 minutes
instead of an hour;
(ii) the time of treatment with 5% barium hydroxide ranged
from 3 to 7 minutes, instead of 5 to 15 minutes;
(iii) incubation in 2xSSC was carried out for 2 hours
instead of one.
The camera luoida diagrams were made using an
oil - immersion objective and a 15 x eye-piece. The
photomicrographs were taken using an oil-immersion or 40 x c
obje.tive and an 8 x eye-piece. Thus the final magnifications ,. of the photographs are x 800 and x 320, respectively.
*=*=*=*=""=*=*

V
OBSERVATIONS

-35-
OBSERVATIONS
A. Oaryo1ogice1 Study
The caryo1ogy of Trilophidia annulate is reported in
detail. As far as the author is aware, this species has
not been reported in the past. Two populations of Chroto
gonus traohYEterus were scanned by Sharma and oo-workers
in a series of papers ( 1962, 1963, 1964, 1965 and 1967 ).
Their findings are compared and oontrasted with those of
the two local popu1ations of the same speoies. Also the
details of the morphometrio analysis, which were not
available in the above-mentioned papers, are inoluded in
this work.
A brief report of the meiosis and mitosis of
Leva indioa is given. The meiosis of the diploid and te~ra
ploid spermatocytes of Attraotomorpha speoies has already
been reported by Ray-Chaudhuri and Bose ( 1948 ) and,
therefore, only morphometric data and mitotic study is
undertaken here. The spermatogenesis of Peekilocera picts
was worked out by Srivastsva (1959) and hence in this case
too, only mitotic oaryological study is reported, along
with the banding pattern.
1. !ri1ophidia annulata Thunberg.
a. Meiosis I The diploid number of ohromosomes in the
male germ cells of T. annulata is 23 ( Fig. 10 ). Measure
ment of chromosomes was made from twenty clear well-plaoed
metaphase plates ( Table 1 ). The chromosomes oan be

• \
•
• o
• • •
Fig.ID
2 3 4 5 6
(\~ nn 1\0 n" 1\1\ 1\/\ (X'~
7 8 9 10 11 'x'
f\A I\A AA AI\ " .. A , IO? j
Fi g.ll

-36-
classified into three long, seven medium and two short
chromosomes ( Fig. 11 ). The sex chromosome ( X ) is the
third in order of length. The minute second arm is not
visible in ~y chromosome. The sex chromosome shows slight
heteropyonosis at the spermatogonial metaphase and can
be easily demarcated from the autosomes ( Fig. 12 ). The
general course of meiosis is of the classical type.
In the prophase stage, the pachytene nuclei
reveal the X as a strongly heteropycnotic body (Fig. 13 t 14).
in tne pachytene nualei. The eleven autosomal bivalents
show a clear sp11 t down the middle '( Fig. 14 ), which the
later stages show to be reductional, i.e. between the two
synapsed chromosomes, rather than between the chromatids
of a single chromosome. There is an indication of the
presence of small arms ( marked by arrow ) in one of the
bivalents ( Fig. 14 }.There is a distinot tendency to
heteropycnosis at one end of several of the autosomal
bivalents. The heteropycnotio end is probably the proximal ,
one, as believed by White ( 196i'). Small blocks of hetero-'
chromatin are present interstitially in the bivalents
( Fig. 15 ).
At diplotene, the bivalents show a greater affi
nity for stain and exhibit the lampbrush fibres so chara
cteristic of grasshopper spermatocytes. This characteristic
can be seen at pachytene also ( Fig. 14 ). A single split
can be observed separating the two homologous chromosomes
of which the bivalent is composed. The split is thus

-37-
Table 1 : Mean length in miora of the spermatogonial
metaphase ohromosomes.
I Chromosome No. Length ~ S.D.
1 1.38 + 0.0101
2 6.16 + 0.0226 -3 5.87 .:!: 0.0001 I
I
4 5.41 + 0.0001 -5 4.75 + 0.0001 -6 4.56 + J.OOOl
-7 4.03 .:!: 0.0001
8 3.62 + 0.0002 -9 2.89 + 0.0002 -
10 1.91 :!: 0.0001
11 1.37! O.lJOO
• X' 6.34 + 0.0001 -

. ''''-"
, :1. ,
. "" ,.-., .. ~ I t , ,-
\ .i.i~ . ;
.
h.b _. __ ..
'\l
~\J " .. " l.'-X
)"> •
"
"'\' ~ A
Fi g .l:-2
$ ' h.4b
~ 1,.-- ' ., .. ' " ' .. "
~ -.. "
Fig.l3
h.b--:---
Fig.14
-, ,-"
~ ,
'. ,
. ~ .
t. . J

"0
I' • ~
Fig.15
.'
F1g.16
F1g.17
- ..... \ -' I 0( .. . . ,.

-38-
Table 2 : Chiasma frequency at different stages of
meiosis
-Number or Stages of Total Mean chiasma frequenoy
Nuo1ei Meiosis Chiasmata per Nuo1eus
10 Diplotene 160 16.00 + 0.02 -15 Diakinesis 172 11.46 + 1.24 -20 Metaphase I 227 11.35 + 1.26 -
reductional and not an equational one. In this stage the
X chromosome shows a pronounoed negative heteropycnosis.
The latter is more evidect in the understained nuolei. Where
the stain is dark, the X chromosome looks just like the
8utosomae bivalents ( Fig. 17 ). Each of the two long
autosomal bivalents invariably contains at least two chias
mata, and sometimes even three ( Fig. 11 ). The number of d
chiasmata never excees two in the medium bivalents and one 1\
in the small bivalents. Mean ohiasmatfrequenoy per nucleus
is 16 + 0.02 ( ~able 2 ). -In the diakinesis stage, the 'woolly' appearanoe
is lost to a great extent and the bivalents aquire an
almost smooth outline. A variety of shapes of the bivalents
is found in this stage ( Fig. 31 ). The two long bivalents

-39-
exhibit either 'diamond' or 'ring' shapes, the former
when the interstitial chiamata are at the end point of
terminalization, and the latter when the terminalization
is complete. Three or four of the medium sized bivalents I ~
r
r
look 'oross' - shaped owing to the presenoe of one
interstitial ohiasma, which has not been terminalized. Rest
of the medium bivalents look either ·v· or'rodt shaped j'.,-
because of the terminalization of the interstitial ohias-
mata. Eaoh of the two small bivalents looks like two
adjaoent dots. Mean ohi8sma fre~uency per nacleus is
11.46 .:t 1.24.
Maximum condensation oCourrs at metaphase I and
no heteroohromatio ohromosome or chromosome region can be
Observed. In the polar view ( Fig. 16 ) the eleven
bivalents are seen to form an irregular oircle. The large
and medium bivalents ocouPY the peripheral position while
the small ones remain in the centre. The X ohromosome can
easily be distinguished from the others because it always
lingers outside the circle, either 'above' or 'below' the
equatorial plate ( Fig. 18 ). The proximal ends of the
chromosomes are distinctly pulled apart at first metaphase,
but only over a short distance. It may 'be taken as an indi
cation of a 'premetaphase-atretch' during spindle formation,
but this is nothing like as strongly developed as it is in
many mantids and phssmids ( Schrader. 1944; Hughes-Schrader,
1947 ). Mean chiasma freQuency per nucleus is 11.35 + 1.26. -Thus the loss of chiasmata is more between diplotene and
1\ / .

[ 1
• • .. ......
", --x 4' . , "
If,. + J .," J , .~. r'- ~ ·
-"" ..... - ' . ' "
_________________ ~~ _______ !~I1J~~~\~~~ __ ._J
I ~
Fig.18
Fig.19
Fig. 20
J \

-40-
diakinesis than between diakinesis and Metaphase I.
Anaphase I oonsists simply in pulling asunder of
the two ohromoBomes of eaoh bivalent ( Fig. 20 ). Sometimes,
the bivalents exhibit the 'pseudo-ohiasmata' ( Fig. 19 ),
when the terminal points of the separating bivalents stiok
together for a oonsiderable time. A,distinot split is
visible between the ohromatids of whioh each of the
separating chromosome is composed. Thus in early anaphase
I the daughter aorooentrios appear 'V'-shaped as is normal
in the ohiasmate meiosis. The univalent X ohromosome also
shows a split between its ohromatids in the late ( Fig. 21 )
if not in the early anaphase ( Fig. 22 ). In the normal
oourse no thread-like oonneotion is seen to conneot the
separating daughter ohromosomes.
At telophase If the spindle fibres conneoting the
separating daughter ohromosomes stain quite deeply and
are clearly visible ( Fig. 25 ). Deoondens.ation sets in
at late telophase and the daughter nuclei look like loosely
spun balls of chromatin fibres.
The seoond meiotio division follows the usual
oourse. At metaphase 11 the chromosomes look rounded and
condensed ( Fig. 23 and 24 ), but are smaller in size and
closer together as oompared to metaphase I chromosomes.
At anaphase two, the ohromosomes divide equationally. The
_ J 'rod' shaped 'chromatids' or the daughter chromosomes move ! I'" t'- towards the opposite poles ( Fig. 23 and 24 ). The 'rods'
assume a star-like appearanoe at the end of telophase 11
( Fig. 26 ).

-~
-?fr-
~ .,.
'I
... " . "
"'~
... ,'" , l".
.. .. 't. ."
. j,;
Fig. 21
~ 'X'-( "," ,V
", i • ... t". . 11 , ., "" o 7 '
Fig. 22
/),
J ' 6 ? f (. ..
:/' I... •
I, '"
~
. .. . '{i ~
.~
~
Fig.23
\ ) /~ \,
',,~ ~'. ~
!i' .......
-:-.. , - ( ~,

•
, .. ~~:. .-,. ·11 ~. _ d", .'
~ J£ . ~ ~ :
• -::.t
• •
Fi g . 24
~- -:.;'
"liW .. .
Fi g . 25
Fig. 26
-
ii' .;,' '.
, ~
•
., .' ~.
•
. J~

1'. I
-41-
b. Abnormalities in the Natural Population :-
The frequency of individuals showing aberrations
in the natural population is very low ( T8ble 3 ). One
hundred and six individuals were scored in all. The aberra-
tions can be analyzed under the following headings :-
(i) Extra frasment : In a single individual, three
metaphase I cells out of 35 and one anaphase r cell out of
50, have been found withhan extra element. The latter is i
round in shape and lies free ( Fig. 21 and 28 ). The
fragment is smaller than the smallest autosomal bivalent
and does not show negative heteropycnosis.
(ii) Polyploid cells: In a single follicle of an
individual, 13 cells in diakinesis are found to be
polyploid ( Fig. 29, 30 and 31 ). All the polyploid cells
observed are tetraploid in nature. The two XS have never
been found to pair, though they may lie close to each
other. No multivalents are found in the cells. In some of
the other follicles of the same individual, a few scattered
cells are found.
(iii) Anaphase bridge: Only one out of a total of 99
cells in anaphase It is found to possess s.n anaphase bridge.
A fragment of considerable stze accompanies the dicentric
bridge ( Pig. 32 ). Manna ( 1954 ) reported the occurI'ence
of dicentric gridge without any accompanying fragment in
T. arululata. The dicentric bridge involves an intricate
terminal association of only one of the daughter chrometids
of a homologue with the corresponding one of the other.

~ - ... \~ ,,"- • ~r .... ,\.'" .. • , ex ,f '" .' 0' " \ . • •
. f .. "', , .
" . .. ,. -0 -, ..
, ." .-• , F1g.2 '7
-i"'" -~,
\' eX , f ',"
i.
•
F1g.28
, ~ ';'1 t-... .,.. -¥ , ... r r •
'~ .~~\ I • "~I ~ . ' -' \ ~ .. -
• - .... ~ y~. .~
? ~:/~- ~ c..
~ . -"" ... 1r., , '" .I..,,'"
~ ~'';'
, "
"- " '~ ~ .§.'
"'J .~:-
Fig. 29

-r
,,J( x..,,' ,'-
I
- • - , •
~ " W
~ t. p ,c.
\
~~ I ~ 'f , f)
• ..... ~ .~
.... .,
~
, .
...
~ ,.
-i
/ tp ,c
t..' . , .. ' ~, . . <.
~ ~ "..0}
• .. ~~;
~ ~ 1,':
,'.- ~:-
F1g.30
-I
~ ... • + I -Fig. 31

-42-
The accompanying fragment is a small body lying at the
equator of the spindle. The bridge is stretched between
the two poles of the spindle, the other daughter half of
each homologue lying free, giving it a characteristic
appearance. The two limbs which form the bridge are asso
ciatedat their terminal ends. The fragment seems apparen
tly acentric in nature.
(iv) Chromosome breakage: In a single individual, one
cell at early anaphase It shows a breakage in the sex
chromosome (Fig. 33 ). The breakage is at the chromatid
level and is confined to the distal region. The fragment
lies close to the arm and there is no visible connection
between the two.
Table 3 : Frequency distribution of abnormalities with
their percentages.
Types of' Stage of Total Cells Total of Abnormality Meiosis Studied Abnormal Cells
Extra Metaphase
I Fragment I 35 3
I
Percen-tage of Aberration
a.51 %
I1 Diakinesis I Polyploid
I 64·.00 % Cells 25 I
16 ~'-'
Anaphase Anaphase Bridge I 99 1 l.vl %
Chromosome Anaphase Breakage I 71 1 1.40 %

-I I
~~~ fr. ch
r~ / r . frj aS5o .ch- e-
\ b \J
l: '(i: • (
- -I Fig. 32
.. " A :>, •
~ ~ .,,, ' br I! , .. ,. . .
. ~ ~;... ,:
Fig. 33

-43-
o. Mitosis: At prophase, no ohromatin 'threads' are visible.
The whole nuolear volume is filled up with heterogenous
material. In the male tissue, the single X-chromosome remains
isopycnotic during interphase and hence no Barr body oan
be seen ( Fig. 34 and 35 ). In the female tissue, however,
one of the two X-chromosomes becomes heteropycnotio. Thus
the behaviour of the X-ohromosomes in somatic cells is exa
otly opposite to that Observed in germ oells ( Mukherjee,
1965 ). At metaphase, the chromosomes beoome visible as long
bodies with a distinot split between their chromatids. The
degree of oondensation is less as oompared to the germinal
chromosomes. A comparison of the lengths of germinal
( spermatogonial metaphase ) ohromosomes and that of the
somatio ohromosomes ( Fig. 36 and 37 ) makes it olear that
the latter are approximately one micron longer than the
former. There is a slight indioation of the presence of
small arms in some of the longer chromosomes ( Fig. 34 and 35,
arrow marks ). No anaphase and telophase oould be found.
2. Chrotogonus traohypterus Blanohard.
The diploid number of chromosomes in the ma.le germ
oells of Chroto8onus traohypteru8 is nineteen (Sharma,
Parshad and Bedi, 1962 ). Measurements of ohromosomes
were made froml twettty olear well-plaoed spermatogonial
metaphase plates ( T8ble 4 ). The chromosomes oould be
classified into three long, six medium and one short
chromosomes ( Fig. 38 ). The sex chromosome i~ the longest.
a.. Meiosis: The meiosis hCis already been worked out

Fig. 34
Fig. 35

7
'" 6 '-u
L 5 .<: 4
~ 3 t::n c Cl>
.....J
'"
8
7
2
~ 6 L 5 c .- 4
..r:::: ~
~3 Cl>
...J 2
2'1
2 3 4 5 6 7 8 9 10 11 X C hro mo so m e Number
Flg.36
L.... L- I- L- LL I-L- ...... L-I-I- I-
1 2 3 4 5 6 7 8 9 10 11 ·x' Chromosome Number
Fig. 37

-44-
Table 4 I Mean length in micra of the spermatogonial metapha.se chromosomes.
Chromosome No. Length + s. D.
1 4.90 + 0.65 -2 4.53 + 0.20
I
-3 3.90 + 0.13 -
I I
I I
4 3.60 + 0.08
5 3.43 + 0.08 -6 3.19 .! 0.19
7 2.95 + 0.11 -8 2.59 + 0.13 -9 1.75 + 0.08 -
I ' X' 5.85 + 0.14 I -
hence it is omitted by the author here. A few points of
interest are, however, being mentioned. The 'megameric'
bivalent, which is a diagnostio feature of the Ottu (Punjab)
population (Sharma, Parshad and Gupta, 1965) and is absent in
the Delhi population (Sharma, Parshad and Bedi, 1962), is
absent in the local popu12tion of Bhopal too. In the early
prophases, two heteropycnotic bodies oan be observed - one
large and one small (Fig. 39). The former is obviously the

2 3 4
6 7 8 9
AA n .. A" ....
\ top,
Fig. 38
_____ - C.C
F1g.39
~ O-p.c \/,..0 • \ ,;J
... ~'X'
Fig. 40
5
a

-45-
Table 5 Chiasma frequency at different stages of meiosis in the Bhopa1 population of Chrotogo!lUS trachYl;lterus.
Number of stages of Total Mean chiasma frequency Nuclei Meiosis Chiasmata per nucleus
10 Diplotene 155 15.5 .:!: 1.21
13 Diakinesis 146 11.23 + 1.04 -15 Metaphase I 160 10.66 + 0.25 -
Table 6 s Mean chiasma frequency per nucleus in the three popu1ations of Chrotogonus trachypterus.
Mean chiasma frequency per nucleus Population
Diplotene Dakinesis Metaphase I
Delhi 15.25 + 0.12 12.15 + 0.31 10.95 + 0.94
Ottu(Punjab) 15.86 + - 11.93 + - 11.20 + --Bhopa1 15.5 + 1.27 11.23 + 1.04 10.66 + 0.25 -
-
sex chromatin 2nd the latter probably belongs to the supernu
merary elements. At anaphase I, the formation of 'pseudo
chiasmata' is found to be a regular feature (Fig. 40, 41, 42
and 43). In the other popu1ations, reported earlier, this
characteristic is not so prominent.
In the present study the loss of chiasmata is
str.onger between diplotene and diakinesis (Table 5) and mean
chiasma frequency per nucleus differs from that of the Delhi

• .. ( ,
"::>- X
,. -
~ 0 • •
• p.c-"
• I
Fig. 41
Fig.42
Fig.43
( , X I , .. f.-
•. ~ , -•
# •

-46-
popula.tion but shows a distinct similarity to tha.t of the
Ottu population ( Table 6 ). ,
b. Abnormalities in the natural E~Eulation :-
A total of eighty-six individuals were investigated
to locate the aberrations. A list of various typys of
aberrations in the Delhi and Ottu population and their
presence or absence and frequency in the Bhopal population
is given in Table 7.
(i) Supernumerary Chromosomes: Only one individua.l
revealed extra elements in its cells, which are smaller than
the smallest autosome and are positively heteropycnotic
during diplotene and early diakinesis. Extra elements with
such oharaoteristics have been designated as supernumerary
chromosomes by a number of scholars ( Carothers, 1917;
Carrol1, 1920; White, 1949; Rothfels, 1950; Rees and Jamieson,
1954; Srivastava, 1954; Evans, 1954; Ray-Choudhuri and Guha,
1955 a.nd Nur, 1963 ). Extra chromosomes with similar bahaviour
have been described as 'B' chromosomes by Lewis and John(1959).
The number of supernumerary chromosomes in the
meiotio cells of indiVidual No. lA varied from one to six.
which shows that they are mitotically unstable. resulting
in their accumulation during the pre-meiotic division. In
the indi.vidua1 under consideration. 68 cells out of a te.tal
of 91, carry two supernumerary chromo30mes ( Tables 8 & 9 ).
Sharma et sI ( 1965 ) reported the occurrence of 1 - 7
supernumerHrY elements in the Ottu population of Chroto~onus
trachypterus.

-41-
Table 1 I Various aberrations and their distribution in the three populations of Chrotogonus traohlEterus.
I Nature of aberration Presenoe (+)/Absence (-) Number of in the population of Individuals
Delhi I Ottu Bhopal (Bhopa.l)
1. Supernumerary Chromosomes (-) ( +) ( +) 1
2. Aneuploidy ( +) (+) ( +) -33. Polyploid Cells (+) ( +) ( +) 3
4. Suppression of paohytene pairing ( +) (-) (-) -
5. Anaphase laggards ( +) ( +) (-) -6. Anaphase bridges (+) ( +) (-) -
--7. Non-disjunotion (-) I
(+ ) (+ ) 1
8. Understained Regions (-) ( +) ( +) 3
9. Chromatid bree,kage (-) (+ ) (+ ) 1
10. Extra fragment (-) ( +) ( +) 1
11. Unequal bivalent ( +) (-) (-) -
n

-48-
Table 8 : Frequenoy distribution of the two supernumerary ohromosomes at various stages of meiosis.
stage of meiosis Number of Cells with Peroentage Cells S + 2s
l1li1 --Diptotene 20 13 65 %
Diakinesis 52 4,3 82.69 %
Anaphase I 4 3 75.00 ~
Meta.phase I 15 9 60.00 ~
Column Total J 91 68 74.70 ~
Table 9 : Frequenoy distribution of the supernumerary ohromosomes in the meiotio oells ( pooled data ).
Chromosome
I
Complement S + 1s S + 28 I s + 3s s + 4s s + 58 S + 68
I
Number of Cells 12 68 3 5 ;fl 2
Percentage ~.18; - -
74.70% 3.29% 5.49% 1.09'?, 2.19%

-49-
The supernumerary element. when single t
( unvalent ), orientates normally at the equator. At anaphase 1\
I and 11 it moves undivided to either of the poles along
with the autosomes showing a double reduction. In a long
cell st teloph~se II~ the univalent is found to l~g
behind ( Fig. 44 ).
~ben represented more than once, the supernume
rary chromosomes usually form bivalents, each with a
single terminal chi~sma. The single bivalent in the n + 18a
condition 1£ found to divide reductionally at anaphase 11
( Fig. 45 ). In the triplioate oondition, one bivalent i~
accompanied by a univalent. In a sine~e cell out of three,
the bivalents exhibit non-disjunction at anaphase 11 and
pass to the same pole, whila the single univalent passes
to the other pole ( Fig. 45 ).
In the n + 4s condition, all the five cells
have the supernumeraries in the form of two bivalents. The
latter show double reduction and pass to the opposite poles
in an undivided state ( Fig. 46 ). This behaviour is not
noticed 1n the Ottu population. In the pentapl1cate condi
tion one bivalent is found at the other pole at anaphase
11 ( Fig. 46 ). The behaviour of the pentaplicate is
undetermined in the Ottu population. When. present in
hexaplioate, the supernumeraries form two bivalents and two
univalents ( Fig. 45 ). Their behaviour at anaphase could
not be determined as both the oells possessing them were at
early anaphase II ( polar view). The hexaplioate condition

~ 1\ • s_ •
it ~
,'t-~
"-,,,,,-~ ~ ., ~
-~
..1-4["
-:...\ 11 - ,
\ ... ~-\' \" I
.... ;'-- -ss I s
. --.
.. • •
t ...
-......
.4 Fig. 44
Fi g .45
Fig.46
~ .,~
~
.1Jj-
'1-
• . 1
" . .

-50-
ie not reported in the Ottu population, while the heptapli
oate oondi tj.on is not found in the Bhopal population.
The behaviour of supernumeraries during anaphase
I was similRr to that of the Ottu ( Punjab ) population.
(ii) Polyplo~.dy:
Soattered polyploid oells were obssrved in three
individuals, oa,rrying otherwise the standard oomplement in
the meiotio I ( 9 AA + X ) and meiotio II ( 9A + X I and (9A)
cells. The frequenoy distribution of the polyploid cells is
given in Table 10.
The two tetraploid oells at diakinesis are ohara
oterized by the absenoe of any multivalent ( Fig. 47 ).
The two sex chromosomes lie away from eaoh other. The lone
tetraploid oell at metaphase II oontains 36dA + 2dX
oomplement ( Fig. 48 ). The daughter sex chromosomes show
a different degree of condensation and the autosomal
univalents reveal a olear split between the daughter
autosomes. In the third case of polyploidy, the first
spermatooyte nuclei are contained within a OOIIl1!lOn oell wall
( binuoleate condition) and at anaphase 11 divide into
9dA + dX - 9da + dX and 9dA - 9dA ( Fig. 49 ). The latter
again remain within the oommon oell wall, resulting in a
tetraploid condition.
In the Delhi and Ottu populutions, a low frequenoy
of polyploidy was noted in the spermatogonial metaphase and
first meiotio prophase and metaphase.
(iii) BPn-disjunotion: In a single individual 3 oells,

.. ~
" ,., . ''\; --:::
• . ~.
c.c..w--:-
, .-:
• . --'/ . /, . r (x •
Fi g. 47
Fig. 48
, , .. , ' . -- I~ ~
'" , --I "
~ '" - --\ , , - .. • Fig. 49
-

-51-
Table 10 : Frequency distribution of polyploid cells during various stages of meiosis.
stage of Iotal cells Cells 'l-lith Perc:entage Meiosis counted polyploidy aberration
-- .. _- -. .-Diakinesis 29 2 6.89 %
-_._"---._-------- .-Metaphase II 11 1 9.09 %
Anaphase II 32 5 15.62 % - ._---_ .. _-- ---
out of a total of 19 ( 15.78 % ), eDlibit non-disjunction
during anaphase II. The chromosomal configuration of two
cells is 8dA and 10da, where two daughter autosomes of a
univalent pass to one and the same pole ( Fig. 50 and 51 ).
In the tbird case, non-disjunction mi&~t have occurred at
anaphase I with "lA + 2AA + X at one pole and 7A at the other.
The latter again underwent non-disjunction at anaphase II,
when the two daughter 8utosome8 of a univalent passed to
one and the same pole. This resulted in cA and 8A oon£igu
rations ( Fig. 52 ).
(i v) Unde!:~!§:!.l!:~.9:)i~;$!.~,!:
Three ind:l. viduals possessed understained regions
in certain autosomes during anaphase I and 11. These are
invariably situated sub-terminally ( Fig. 53, 54 and 55 )
and affect only one member of a pair of homologues except

, \,
• \ non, di5 " .-/
I't~. 1'~1
Fig. 50
loo , "
, :.c ~ 'if
~ ,
* :/<' " .,
Fig. 51
, Y\OY'l. di5 __ .. , :
,) ~" ,~
Fig.52

,
Fig.53
•
Fig.54
/'f I \) - /" dX
~ I ~~)~ u . s . r
Fig. 55

-52-
in one oase ( Fig. 53 ). A small chromatic connection can
be seen between two yarts of the chromosome arm. The
frequenoy of oells carrying understained regions in the three
individuals is given in Table 11.
Table 11 : Frequency distribution of understained regions during different stages of meiosis.
Individual stage of Total Cells Cells with Peroen Number Meiosis oounted I understai- tage
i nad region I i
1. Anaphase I 15 4 26.66%
---2. Anaphase 11 22 8 )6.36%
...
3. Anaphase 11 19 6 31.51%
(v) Chromatid Breakase :
In a single individual. out of 21 cells at
anaphase 11, one oe11 carried a breakage. The latter is at
the proximal end of a daughter chromosome ( Fig. 56 ). The
fragment is not associated with the other part and lies a
little away from it, at a different angle.
(vi) Extra Fragment :
Out of 10 oells, studied during metaphase 11, two
cells h~ve been found with an extra fragment ( Fig. 51 ).
The fragment lies free at the periphery of the spindle, out

•
• I I,
(I J\"
"." /" "
f 1-' . br
Fig . 56
.-
• r.:., /
~~ ~ • • ~
".!.. ... ! , - \ , .. ' -
"*'* ,~ I
- .('
Fig. 51
int .nu
Fig.58

-53-
side the circle of metaphase 11 chromosomes. It is round
in shape and smaller than the supernumerary chromosomes.
c. Mitosis:
The prophases do not show any heteropycnotic body
in the male somatic tissue' Fig. 5d ). At metaphase, a
clear split is visible in all the chromosomes. The sex
chromosome can not be differenciated from the autosomes on
the basis of degree of condensation which is uniform in
both, but it can be differentiated on the basis of length.
No telophases and anaphases could be found.
A comparison of the lengths of spermatogonial and
somatic ( gastric caeca ) metaphase chromosomes, reveals
that the former are much more condensed than the latter.
The histogram of spermatogonial metaphase chromosomes
( Fig. 59 ), based on the data of Table 4, shows that the
longest chromosome measures about 6 micra. while the same
chromosome measures about 15 micra in the somatic tissue
( Fig. 6J ).
In the rod-shaped chromosomes, though reported
to be 'acrocentric' , no second arm could be seen even in
the somatic chromosomes which are loosely coiled.
3. Leva indica Boliver.
The diploid number of chromosomes in the male germ
cells of Leva indica is twenty-three ( Fig. 61 ). Measurements
of chromosomes were mDde from twenty well-placed sperma
togonial metaphase plates ( Table 12 ). The chromosomes can
be classified into three long, six medium and three short

5 ~ L-L)
4
L 3 c:
2 -C. ...... = c: 4J
.....J
2 3 4 5 6 7 8 9 'X'
Chromosome Number
Fig.59
15
12
'" L-U
9 :i c: ._. ~ E, ... C7' c: QJ
...J 3
2 3 4 5 6 7 8 9 ·x' Chromosome Number
Fig. 60

-54-
Table 12 : Mean length in micra of the spermatogonial metaphase chromosomes.
Chromosome No. Length::!: S.D.
1 4.86 + 0.40 -2 4.48 + 0.48 -3 3.55 ! 0.54
4 3.09 + 0.44 -5 2.69 + 0.33 -
I ! 6 2.36 + 0.20 -7 2.14 + 0.30 -.--8
I 1.75 + 0.20 -
I
9 1.34 + - 0.09
10 1.18 + 0.30 -11 1.10 ! 0.31
• X' 5.40 + 0.47 -
chromosomes. ( Fig. 62 ). The sex chromosome is the
longest.

Fig. 61
2 3 5 6
7 8 9 10 11 'X'
n6 •• .... .... .. .. 0 10fL
I
Fl g .62

-55-
a. Meiosis:
The general course of meiosis is of the classical
type. During interphase stage, the sex chromosome appears
as an intensely positive heteropycnotic body. This behavi
our is maintained till diplotene. At late spermatogonial
metaphase, the sex chromosome oan be distinguished from the
autosomes ( Fig. 61 ) due to its length. The arrangement of
the first spermatocyte metaphase chromosomes is normal and
regular. Bivalents of various eh&pe~ and sizes are arranged
equ1distally from the two poles. The sex ohromosome lies
either 'above' or 'below' the equatorial plate. The
'pseudo-ohiasmata' have not been observed at anaphase I.
The long univalents are arranged at random on the radial
metaphase 11 oonfiguration. No asynchronous behaviour has
been noticed at anaphase 11.
A study of ohiasma frequency reveals that the loss
of ohiasmata is stronger between diakin.esis and metaphase
I ( Table 13 ). The long bivalents possess 2-3 chiasmata,
while the medium ones never have more than two chiasmata.
The short bivalents possess one chiesme each.
b. Abnormalities in the natural populatio~ ,
A total of fifty-two individuals were studiea.
The freouenoy of aberrations is very low ( Table 14 ). The
aberrations can be analyzed under the following headings :
(i) Extra fragment: In a single individual, 4 cells out
of 54 at diakinesis, have been found with an extra fragment.
The latter is round in shape and smaller than the smallest

-56-
autosome ( Fig. 63 ).
(ii) Polyploid cells : In a sin~;le follicle of an
individual 11 cells out of 48 at anaphase 11 are tetraploid
in nature. Forty-four daughter chromosomes were contained
within a common cell wall ( Fig. 64 ). Similar type of
tetraploidy has been found in C. trachypterus by the
author. But in the present case, the origin of polyploidy
seems to be different. Here the complement includes 44dA
instead ot the expected 44d.A + 2dx.
Table 13 : Chiasma frequency at different stages of meiosis.
Number of stage of Total Mean ohiasma frequency nuclei meiosis Chiasmata per nucleus
-10 Diplotene 136 13.6 .! 1.49
12 Diakinesis 144 12.;)0 + 1.12
1 Metaphase I 68 9.71 + 1.11 -
(i1i) Abnormal Spindle: At anaphase 11, 30 cells out of
a total of 90, possess an abnormal spindle structure. One
pole is normal with 11/12 daughter chromosomes, while the
other pole is 0ifid ( Fig. 65 ). At this pole the daughter
chromosomes are arranged in two groups. A wide gap is
present between the two groups of chromosomes.

... I r
[.~ ~ l eJ
" I ~ .. , ...... (J •
~ 1 J -- -
Fig.63
-c.W
Fig.64

-51-
Table 14 : Frequenoy distribution of abnormalities in the natural population of Leva indioa.
--.
Types of I Total oells I Stage of Cells with Peroentage a.bnormali ty I meiosis studied tberratiOn
of aberration
.- -Extra Diakinesis 54 4 1.40 % fra.gment
I
AnBPhBse-t +17 •
"' . Polyploid 48 35.41 %
i Cells
I I I
III i Abnormal I Anaphase 90 30 33.33 % ! Spindle
Breakage Spermatogo- 11 5 29.41 % nial metaphase
--
(iv) Bree,kage: At late spermatogonial metaphase, 5 cells
out of a total of 17. possess breaks in the long ohromosomes.
Usually the two pairs of long autosomes are affected and the
sex ohromosome remains unaffected. In Ol1e oase, only one
ohromosome of a pair is affeoted ( Fig. bb ) t but usu~1l1y
the breakage is found in more than one chromosome ( Fig. 6'1 ).
In &11 the ceses, both chromatids of the affected chromosome,
possess the breakage at the same locus.
c. Mitosis:
The prophases do not show any hetropyonotic mass

c..W
~
Fig.65
Fig. 66

Fig.67
.. , ) . , . ,
." . ... I · . ~. 'x'
o ''' :~ ~ . ~ , . " , • h.b
Fig. 68

-58-
in the male somatio tissue. But the cells of the femele
tissue do possess a single indistinct hetropyonotio mass
( Fig. 68 ). At early metaphase the chromosomes attain a 1
leaser degree of condensation as compared to the germinal
tissue, and are long and slender. A distinct split is
present between the chromatids. The homologous chromosomes
lie apart. The sex chromosomes can not be distinguished
from the autosomes on the basis of condensation. Being the
longest in the complement, their identification is possible
by comparative measurements ( Fig. 68 ). At ll:ite metaphase
a greater degree of condensation is achieved and the split
between the chromatids does not look very distinct.
No asynchronous movement has been noticed at
anaphase. The short chromosomes reach the poles first. At
telophase, the daughter chromosomes at the poles can not
be oounted easily beoause of a olose grouping and the onset
of deoondensation. The two daughter cells get separated by
a constriction at the equatorial region. A smooth outline
/' is squired by the daughter nuclei at interphase.
A comparison of the lengths of germinal (spermE,to
gonial ) and somatic ( gastric caeca ) chromosomes reveals
that the difference is not much. The sex chromosome in the
former measures about 5.50 micra, while in the latter about
6.75 micra. ( Fig. 69 and 10 ).
4. Attraotomoryha crenulata Fabricius.
The diploid number of chromosomes in the m81e
germ cells of Attractomorpha crenulata is nineteen ( Fig. 11).
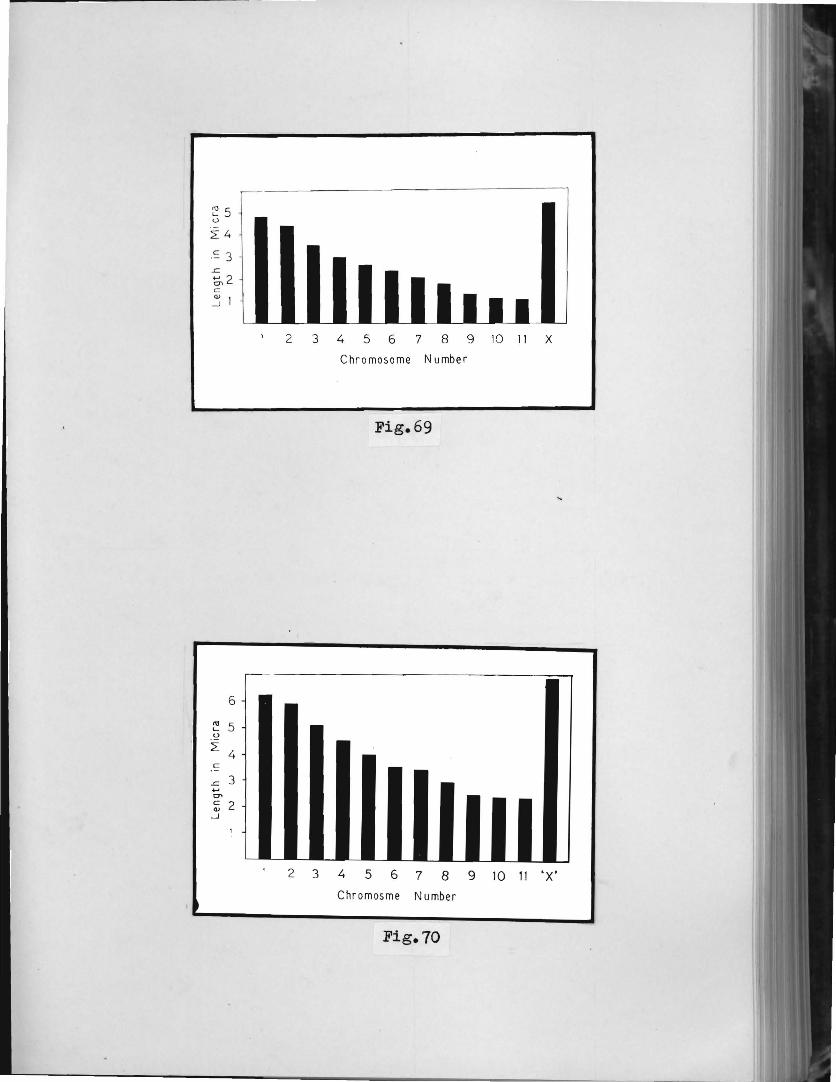
~ 5 <J
== 3
6
'" 5 '-.9 2:
4 c:
..c 3 .., eT> c: 2 '" -l
2 3 4 5 6 7 8 9 10 11 X
Chromosome Number
Fig.69
2 3 4 5 6 7 8 9 10 11 'X'
Chromosme Number
Fig. 70

-59-
Meaauremellts of chromosomes were made from twenty well-spread
spermG.togonial metaphase plates ( Table 15 ). The chromosomes
C&ll be classified into three long. six mediUlll and one short
chromosomes ( Fig. 72 ). The sex chro~osome is the largest
and can be distingu,ished easily i'rom the autosomes ( Fig. 71)
because of its length.
Table 15 : Y..ean length in micra of the sperme.togonial meta.phase chro:n~so~E's.
ChromOSOm~~No. I
I Length .:!: S. D.
i
1 7.78 + 0.96 -2 7.16 + 0.48 -3 6.14 + 0.93 -4
I 5.30 0.42 + -
5 4.98 + 0.41 ,
--_. 6 4.84 + 0.47 -7 4.37 + 0.50 -8 3.67 + 0.41 -9 2.55 + 0.17 -
'X' 8.64 + 1.05 -

•
•
1
6
~A
, ,
2
7
Fig.71
3
8
I'A •• 10)1-, ,
Fig . 72 . '
(
4 5
9 ·X'
•• 0

-6·)-
Meiosis ia not being dealt with by the author except for
a few points of interest.
a. t\b~~!:llalities in the natural population :-
In the natural population of Attraotomorpha orenu
~, polyploidy is a oommon feature. The behaviour of
diploid Rnd tetraploid spermatocytes was reported by
Ray-Choudhuri and Bose , 1943 ). In the present aocount,
therefore, varir:Jtions other than polyploidy 8I'e being
reported.
Only four individuals out of a t~tal of thirty-
five, possessed abnormal cells. The frequency of aberrations
is quite low ( Table 16 ).
Table 16 : Frequency distribution of abnormalities in the na.tural population of Attract?Dlorpha orenulata
I Types of stage of Total oellS~lle with peroentagel of
I
aberration meiosis studied aberra.tion aberration
-AnaphaS~
~ . .. , . . Chromatid I 3 3.33 % Breakcge
-Chroma.tid Anaphase I 90 35 38.88 % Separation
--,---
(1) Chromatid breakase I At anaphase I. the largest
ohromosome, that is the sex ohromosome, possesses breakage

" " t!., (
br f'
. rO~ ,,~~~X' . \) "Vt..'
t' ",. -
Fi g . 73
Fig. 74

... I
1'\
-61-
in one ohromatid ( Fig. 73 )." The break ooours in the distal
region. The fragment lies a little filway from the heteromor
phio univalent and no oonneotion is found between the two.
The broken pieoe does not lag in any of the cells.
(ii) Chromatid Separation : The separation of the two
daughter halves is found in 38.8 % oells at anaphase I
( Fig. 74 ). Thus there is a olear indioation of preoocious /\
behaviour. The daughter halves do not drift away from each
other. The separated ohromatids invariably pass to the same
pole after anaphase I in all the instances noted by the
author.
b. Mitosis I
The prophases do not possess any heteropycnotio
maSf:3 in the me.le tissue. At pro-metaphase the ohromatin gets
resolved into olosely paoked blooks inside the nuolear wall.
At metaphase, the ohromosomes look thioker and longer
( Fig. 75 and 76 ) than the spermatogonial metaphase
ohromosomes. The size differenoiation between the two is
appreoiable. The sex ohromosome measures 8.64 miora in the
germinal tissue and 10.69 miora in the som2tic tissue.
Similarly the smallest autosome me8.Sv.res 2.55 micra in the
germinal and 3.72 miora in the somatic tissue. The centromere
is grea.tly stretohed in some of the ohromosomes, revealing
a lightly stained gap between the two chromatids •• Anaphase
and telophase stages were not available.
5. Poekilooerus piotus Fabrioius
Spermatogonial metaphase plates show the presenoe

Fig. 75
F1g.76

~
8
7
6
t; 5
2: 4 c:
'': 3 ..... ~2 Cl.>
-11
<0 c.... <.>
~
c::
~ ..... t:7\ c:: cv -1
10
9 8 7 6 5 4
3 2.
23456 7 8 9 X Chromosome Number
Fig. 77
2 3 4 5 6 7 8 9 'X'
Chromosome Number
Fig. 78

-62-
of nineteen rod - shaped chromosomes ( 18A + x ) whioh can
be divided into three long, six medium 8nd one short
ohromosomes.
The meiosis of this speoies has been investigat"ed
by a number of soholars ( Asana and Makino, 1934; Rao, 1937;
Dutt, 1955; Srivastava, 1960, and others ). White ( 1940 )
reported that the negative heteropyonosis of the X-chromo
some was either less pronounced or even absent in the last
spermatogonial. division. The small arms, though reported to
be present in all the chromosomes by Dutt ( 1955 ). could
be observed in rare instances. Their presenoe, however, is
said to be well-marked in the smallest autosome by Darlington
( 1936 ) and Coleman ( 1943 ). The smallest autosome shows
positive heteropyonosis at diakinesis, along with the
X- ohromosome ( Mangalangi and Subramanium, 1963 ) though
the latter stains relatively lightly.
The present aocount includes only mitotic oaryo
logioal. study and the banding pattern.
a. Mitosis: The pattern of mitosis is similar to that
reported by the author in other grasshoppers. At ee.rly
metaphase, however, distinct second arms are visible in one
of the smallest autosomes and in one modium f:'}utosome
( Fig. 79 ). Darlington ( 1936 ) and ColE)IDhn ( 1943 )
reported the oocurrence of second arms in the smt.311est
autosome only.
The length of the minute arm, in the present
investigation, is about 0.9 to 1.00 micron. A comparison

]'1g.19
-

• 14
12. I'\l L.. U
10 ~
c: 8 .c .... 6 C'l c: ( . ' .. QJ
.-J 4
2. ') .' I ..
2. 3 4 5 6 7 8 9 ·x' Chromosome Number
Fi g .BO
20
."
~15 ~
c ,-10 .c
...... t:n C C1.I
....J 5
, 1
2 3 4 5 6 7 8 9 ·x' Chromosome Number
Fig .al

-63-
of the lengths of somatio metaphase ohromosomes with that
of the spermatogonial metaphase ohromosomes, makes it olear
that the former is almost double that of the latter ( Fig.
80 and 81 ).
The degree of condensation is uniform in the
autosomes and the sex ohromosome, henoe the latter oannot
be distinguished from the long autosomes easily. The well
separated, thread-like ohromatids ooil around eaoh other at
early ~etaphase and resemble the oOnIigurations aoquired
by the bivalents at diplotene in meiosis ( Fig. 79. arrow
marks ). \
A greater degree of oondensationis achieved at
late metaphase. The maximum reduotion in length is observed
at anaphase. No asynohronous behaviour has been notioed
during anaphase and telophase.
b. Banding pattern c The nature of oentromere has been
observed in all the stages of sperma toger~esis with the help
of Giemsa-staining ( Sumner. 1972 ). The observations not
only differ in many respeots from the routine analysis of
oaryotype. but also oOIJ.tradiot a few points. The various
stages are being analysed systematioally :-
spermatosonial metaphase: As stated earlier, the ohromoso
mes of P. :piotua oan be divided into three oa.tegories, viz.,
long, medium and short. For the sake of oonvenience, there
fore t eaoh ohromosome is given a nomeno18.ture aocording to
ita oategory. Fig. 82 shows all the ohromosomes of the ~ diploid oonpliment with their respt:lotive numbero 8.l1d .A
oategories. There are two pairs of long chromosomes (L1 and L2 )

.1
Fig.82
Fig.83
. •

-64-
six pairs of medium chromosomes (Ml to 1V16 ), and one pair of
short ohromosomes (SI). Besides these eighteen autosomes,
the single X-chromosome, long(:st in the oomplement, is e8sily
distinguishable. Giemsa stain has demaroated a de.rk band of
repetitive DNA sequences in the centromerio region of all the
ohromosomes. This band of heterochromatin, after a closer
observation, is seen to be oomposed of four subunits ( Fig. 82,
X and M4 ). Besides the centromerio heterochromatin, a less
porminent band is present at the tolomerio end of the X
ohromosome. The smallest autosome. (Fig. 82, SI) is the most
distinguished of all because of the presence of dark blobs
of heteroohromatin at the telomeres.
Leptotene : The darkly stained m~~ss at the left hand end of
the leptotene-nucleus ( Fig. 83 ) can be e8sily mistaken
for the positively heteropyonotic sex-chromatin. But the
latter is actually at the opposite end ( Fig. 83, h.b. ).
The dark mass is the outoome of the 'bouquet' arrangement,
so characteristio of leptotene staee, being formed by the
polarization of centromeres and telomeres. The ooourrenoe
and orientation of the loops ( Fig. 83, Ips. ) indicates
that the ohromosomes have doubled on themselves and both
of their ends are polarized towards, and oompose, the
'dark pole' ( Fig. 83, dk. pI. ) simultaneously. The four
sub-units of eentromerio heterochromatin are well-marked
in one of the ohromosomes ( Fig. 83, arrow-mHrk ).
Pachytene: (Fig. 84 and 85 ) Late paohytene nuolei reveal
more details about the oonstitutive heteroohromatin. The

Fig.84
Fi g . 85

-65-
latter is separated from the main chromosome by :3 constric
tion ( Fig. 84, a.rrow mark ). The darkly staining centromeric
chromomeres a..ppear like the t horns of 8n8il' and may easily
be mistaken for the ·second arms'. The smallest autosomal
bivalent can be distinguished without any difficulty because
of both its size and the prominent telomeric band of hetero
chromatin ( Fig. 84 ). The sex chromatin hp..s acquired et
peculiar, geometrioal configura.tion in the interphase
nuoleus ( Fig. 85, arrow mark ). At late pachytene, the sex
chromatin appears dumb-bell shaped and eta,ins lighter than
the constitutive heteroohromatin.
Two more oells at pachytene ( Fig. 86 and81 )
show that the cODst1 ttltive heterochromatin also includes the
proximal part of chromosome arms besides the centromeric
heterochromatin, in almost all the bivalents. The SI
autosomal bivalent appears uniformly dnrk because of the
approximation of centromeric tUld telomeric bands of
heterochromatin ( Fig. 81, SI ).
The band of oonstitutive heterochromatin at the
proximal region of chromosomes is more clearly revealed in
Fig. 83 ( arrow marks ). Most remarkable is the 51 bivalent,
which possesses two distinct, close bands in the distal
region ( Fig. 88, dis. bl and die. b2 ). The M6 bivalent
also possesses bands in its dietal region, each ~~ivalent
having a single band situated sub-terminally. The stain of
these bands is lighter than the other bands.
The bivalents at diplotene Gnd diakinesis normally

"'- pl"O . b. het "'-t .e
"- ,/
F1g . 86
-
F1g . 81

-66-
show a greater affinity for dyes. With the use of Giemsa,
however, only those regions take the stain which possess
highly repetitive DNA sequences. Rest of the portions take
a lighter stain ( Fig. 09 ). The SI hi~alent, the single
intersti tial ohic-:lsma being ter:ainalized. is darker than the
other bivalents, espeoially a't the proximel and distal
regi0ns ( Fig. 89, SI ). The u.n1va1en~ 'X' lies coiled upon
itself in the middle of the plate ( Fig. 89, X ), and looks
darker than expected because of the overlapping.
At diakinesis and metaphase It the bands beoome
almost indistinguishable because of the contraotion of the
ohromosomes. still, the heterochromatin can be loce,ted at
the oentromeric end ( Fig. 90, cen. het. ). The arrangement
of bivalents at metaphase I is very clear in Fig. 90, where
the sex ohromosome and the SI autosomal bivalent lie in the
centre of the spindle, while Ll , L2 and Ml to M6 bivalents
occuVy the peripheral region with their darkly stained
centromeric ends directed towards the oentre of the spindle.
The 'X' possessei:3 a distinct centromeric band ( Fig. 90,bd ).
Another view of metaphase I in profile is
illustrated in Fig. 91. Here the bands oarmot be demarkated
in the bivalerlts. The univalents of SI p..nd M6 pairs have
already started moving towards the opposite poles ( Fig. 91,
SI and Si ; M6 and M· 6 ). The sex chromosome shows a
split between its ohromatids a.nd dark regions at both the
liistal and proximal ends.
In the next stage, i.e., anaphase It the slightly

Fig.88
Fig.89

cent . het
Fig . 90
Fig . 91
Lt
~Cent . hel:

~ ...... ---------------61-
heteropyonotic 'X' can be seen at the t1ower' pole ( Fig.
92 ). Its oentromeric band looks darker than the telomeric
ones of the separated chromatids. While all the M-univalents
and L2-univa1ents have reached their respective poles,
Ll-univalents have just departed from each other at the
equator ( Fig. 92, L1 ). The oentromeric heterochromatin
in the L- and M- un1va1ents appears like two dots, lying
side by side. The Sl -univalents possess Cl. comparatively
light region separating the two d::)rk bands a.t the two ends
( Fig. 92, bdl , bd2 ). The heteropycnotic behavioQr of the
'X' persists only upto anaphase 11. Afterwards. the degree
of condensation is found to be uniform in all the chromo-
somes.
At ane.phase 11, the division of the centromere
is superbly revealed by the separate 'daughter' bands at
the proximal end of the divided chromatids ( daughter
chromosomes ) in Fig. 93. The dc;ughter chromosomes of M4,
M5 and M6 have shifted apart while those of Sl lie overla-
pped ( Fig. 93 ). The sex chromosome shows a gree of
condensation equivalent to that of the autosomes. The
orientation of Llt L2, Ml , M2 and M3 suggests that the
centromeres are directed towards the centre of the spindle.
B. Effeot of Drugs
1. Quanti tative stu~l : Caff'eine
Caffeine is extensively used as a therapeutic
agent and because of the widespread consumption of

Fig.92
M2 '--1- ( Mc;.
. . Ml
c.ent .het . x/ tel . he.\:
" Mo\:
Fi g .93

-63-
oaffeinated beverages a major part of the human raoe is
ohronioally exposed to this compound.
In the field of oaffeine mutagenesis there is
some oonfusion due to several contradictory reports. A
survey of available reports reveals interesting findings
on the nature of this methyl xanthine. Kihlman and Levan
( 194d ) observed two main features in the action of
caffeine - (1) the formation of cell wall after mitosis
was impeded although the spindle functioned normally
(2) chromosome bridge formation at anaphase started
appearing after a few hours of treatment. Most of the
work that followed contained genetical and biochemical
results.
Axelrod and Reichanthal ( 1952 ) studied the
fate of caffeine in human subjects. Some of the important
findings were - (1) caffeine was rapidly and essentio.lly
completely absorbed from the gastro-intestinal tract; (2)
caffeine was almost entirely transformed in ms.n, only
about 1% being excreted in urine; (3) the drug pasiSed
rapidly into the central nervous system ; and (4) there
was no day-to-day acculllulation of the drug.
Goldstein and ~arren ( 1961 ) cQnsidered
mutagenecity of caffeine in mammals on indirect ground.
It was stated in their work that since caffeine was known
to be mutagenic to bacteria, fungi and Drosophila, it
could be mutagenic in man because of the bpsic similarity
of genetio material in all forms of life. Their experiments

J
-69-
on pregnant hum2D subjects clea1'ly demonstrated tfH:;t
caffeine equilibrates fret.ly between maternal plasma t:1.L:.d
1-8 week foetus and the exposure misbt be hC;izardous at this
stage - when the segregatiJn and prolifE:iration of germ cells
takes place.
Cattanach ( 1962 ) found caffeine weakly mutage
nic in mice, for only two possible translocatioll8 were
detected. Ostertag et al ( 1965 ) found the drug mu.tagenic
in man as it could induce breakage in the human ( HeLa )
cells in tissue culture.
Clerk and Clark ( 1968 ) reported that caffeine
could induce ohromosome loss in both male and female
Drosophil~ B.nd suggested that this drue might be a oontri
butory faotor in the induction of Turner's syndrome ( XO )
in man. Kuhlman et al ( 1968 ) in their experimEmt u.pon
human oells in cu.l ture, ±'ound that the action of oaffein
is analogous to that of radiation.
A reoent report by Slizynski ( 1911 ) is based
on direot cytological and chromosolUtil observation on the
rat testis. The tissue was, however. maints.ined outside the
body in a short-term culture. The results obtained were
both positive ( clUt'1lping, stickilless and ohromatin extru
sion from the {luolei ) a.nd r.:eg~i.,.tve ( gap formation,
fra.gmentation and v~ouolar extrusion of material from the
nuclei ).
Most of the reports are not purely oytolo;gical
and give an idea of the nature of the drug itself. Among

-70-
the direct cytological studies, that of Kihlman nnd Leva,n
( 1943 ) on plcmt chromosomes is worth mentioning. In the
case of animals the tissue was invariably treated outside
the body in a culture. ( Ostertag et 81, 1965; Kuhl.ITV n
et aI, 1968 and Slizynski, 1971 ).
In the present work, the tioi.,ue wes trel:1.ted in
a natural state, still at·taohed to tlie body and the division
oycle was also taken into considerati0n for treatment.
In order to make a. comparative study of the
effeot of known concentrationq of caffeine upon the testes
cells of o. trachy;eterus, the frequenc.y of three types of
abnormalities, e.g. fragmentation, clumping end polyploidy,
has been determined. In this experiment, mortality occurred
in the higher dosage range ( 0.5 ,%, ·).6 ;1&,·).'7 %, 0.:3 ~t
0.9 10 and 1 % ). Details of the experin,e:nt are given in
Table 11.
The frequency is scored et d1plotene, diakinesis
a...'1d metaphnse I .i;or each type of e,bnor:n:.-.li ty. The dl:it:::~ r-lre
po~led and averaged according to the number of s.rv1ved
specimen for each concE':ntration of caffeine. f,ll the
ceoncentrations were introduoed at prophase and the effect
waa studied after six dpys, when, according to the division
oycle ( Fig. 3 ) the cells are at matepna.se r. An. analysis of the ~bIJorm81i ty data reve~ds some
interesting facts about the relation of tbe varying concen
tration to the fixed time of tr~at~e~t. The frequency
distribution of the three abnormalities for 0.1 %

-71-
Table 11 : Details of experiments of the control and treated specimens of c. trachypterus.
Concentrations Number of specimen Number of specimen of Caffeine used in Experiment survived after
treatrr.ent
"--, Jcontrol Tref1ted Control Treated
0.1 .~ I 5 5 5 5
0.2 ~ I
5 5 5 5 i i
- i
0.3 % I 5 I 5 5
I 5 i
I !
0.4 % i 5 5 5 5 -.- I
0.5 % ! I 5 5 5 4 I
0.6 to I 5 5 5 I 4
I
0.7 '% I 5 I 5 5 2 , ,
0.8 % I 5 5 5 1 ~- -
0.9 to 5 5 5 0 !
~ : ---_.-_.
>: r-1.0 'fo 5 5 0
- ..
caffeine is given in Table 13. Vfuile the occurrence of
£ragmentation is nomin~l ( 4.44 ~ ), the other two
Total apeci-men sur-vived
10
10
10
10
9
9
7
6
5
5

-72-
abnormalities are quite significant, clumping being 16.44 %
and polyploidy 20 %. The fragmentation involves the chroma
tids only and all the stages are easily recognizable. The
olumps are formed by the stioking together of two or three
chromosomes. Polyploidy is at the level of tetraploidy. The
latter state is devoid of multivalent configurations and
non-assooiation of the sex chromosomes. A comparison of the
treated series with the control series reveals that there
is a slight depression in the rate of progression of
prophase into metaphase. The latter stage is numerioally
higher in the oontrol series. The cell-wall is intact in
all the stages.
Table 18 I Frequenoy distributions with their percentages of some types of abnormalities produced by 0.1 % caffeine ( data. pooled and averaged for the five survived speoimens ).
Types of Total cells Total Peroentage abnormality studied abnormal cells frequenoy
Fragmentation 225 10 4.44 ~
Clumping 225 37 16.44 " Polyploidy 225 45 20.00 %
The concentrations, 0.2 %. 0.3 % and 0.4 % show
a gradual rise in the percentage frequencies of clumping

-13-
Table 19 : Frequenoy distributions with their peroentages of some types of abnormalities produoed by
0.2 % oaffeine ( data pooled and averaged for the five survived speoimens ).
Types of Total oells Total percentage abnormality studied abnormal oells frequenoy
Fragmentation 206 12 5.82 %
Clumping 206 45 2l.84 to
Polyploidy 206 68 33.00 ~
and polyploidy ( Tables 19, 20 and 21, respeotively ).
But the frequency of the former is always lower than the
latter. The inorease in the frequency of fragmentation is
gradual from 0.1 ~ to 0.2 % , but at 0.3 % it zooms up,
being almost double the frequenoy at 0.2 %. Its inorease
is steady and signifioant at 0.4 % also. upto the latter
oonoentration, no mortality ooours and oell-wall formation
has impeded notioeably at 0.4 % oonoentration, where the
degree of ploidy has inoreased to 52.43 %, that of olumping
being 49.26 %. Mortality ooours in the higher dosage range. In
0.5 % oaffeine - treated series, a single individual died
after five days of treatment. Other individuals reveal that
the fragmentation involves more oells though at a slower

-14-
Table 20 : Frequency distributions with their peroentages of some types of abnormalities produoed by 0.3 % caffeine ( data pooled and averaged for the five survived specimens ) •
. --Types of Total cells Total Peroentage abnormality studied abnormal oells frequenoy
Fragmentation 300 35 11.66 %
Clumping 300 121 40.33 %
Polyploidy 300 125 41.66 %
Table 21 I Frequency distributions with their percentages of some types of abnormalities produced by 0.4 % caffeine ( data pooled and averaged for the five survived specimens ).
Types of Total cells Total Percentage abnormality studied abnormal cells frequency
Fragmentation 410 103 25.12 'to
Clumping 410 198 49.26 %
Polyploidy 410 215 52.43 %

-75-
Table 22 I Frequency distributions with their percentages of some types of abnormalities produced by 0.5 % caffeine ( data pooled and averaged for the four survived specimens ).
Types of Total cells Total Percentage abnormality studied abnormal cells frequency
Fragmentation 360 101 28.05 %
Clumping 360 190 52.17 %
Polyploidy 360 212 58.88 %
Table 23 I Frequency distributions with their percentages of some types of abnormalities produced by
0.6 % caffeine ( da.ta pooled and averaged for the four survived specimens ).
Types of Total cells Total Percentage abnormality studied abnormal cells frequency
Fragmentation 350 98 28.00 %
Clumping 350 195 55.11 %
Polyploidy 350 210 60.0J %

-16-
rate ( Table 22 ). The other two abnormalities have, how
ever, inoreased significantly, olwmpimg being lesser than
polyploidy. In this series, the de.:?;ree of ploidy could not
be distinguisLod as the groups of chromO[lomes were closely
approximated. Clumping usually involves all the chromosol{,es
of a set, so that the individuul indentity of the chromo
somes becomes impossible. Moreover. the heteropycnotic
behaviour of the sex chromosomes is not recognizable due
to general over-condensation.
In 0.6 % caffeine treated series the mortality
is similar to the 0.5 % caffeine-treated series but the
aberration has obviously increased at least in the cases
of clumping and polyploidy. The percentage-frequency of
fragmentation is, however, equal to that found in :).5 %
series ( Table 23 ). In 0.1 % series, all the three abnor
maliti~s show an increase in percentage-frequency
( Table 24 ). Only two ~peoimens survived after treatment,
in this series. The 0.8 % solution of caffeine see::ms to
be significantly toxic, as here 80 % mortality oocurred.
The only surviving individual possessed the freouency
distribution given in Table 25. The percentage-frequency
of fragmentation has gone down in this case, while that of
clumping a.nd polyploidy han risen to 8 striking level. The
cell-wall fOrnl:.:.,tion hbs been found to be completely
impeded in this series. The recognition of cell-stages was
very diffioult due to high degree of ploidy and stiokiness
involving a number of nuclei.

-'77-
Table 24 : Frequency distributions with their percentages of some types of abnormalities produced by 0.7 % ca~feine ( data pooled and averaged for the two survived specimens ).
<- -Types of Total cells Total Percentage abnormality • studied abnormal cells frequency
Fragmentation 200 65 32.50 % -
Clumping 200 138 69.00 %
Polyploidy 200 149 74.50 %
Table 25: Frequency distributions with their percentages of some types of abnormalities produced by 0.8 % caffei.oe ( data. pooled for the only survived specimen ).
ITot~ cells -- --
Types of Total Percentage abnormality studied abnormal cells frequency
Fragmentation 90 22 24.44 j,
Clumping 90 65 72.22 % -
Polyploidy 9J 70 77.77 % --

-78-
A systemCltic quantitative am::lysis 08!l1l0t be
oarried out with the series of the lEst two concentrations,
where absolute mortality oocurred • In both the oases, the
speoimens died within one Or 'two d;;:ys of treatment. However,
an examination of the tissue of tbe dying specimen throws
some light on the state of the tissue. It is found that
the oells either do not progress beyond early prophases,
or if they do, their reoo~~ni tion is not posoible. The
ohromatin extrudes out from the nuclei, giving them a
'notohed' uppearanoe. Similar unusual appearance has been
reported by Slizynski ( 1970 ) in mouse ohromosomes after
caffeine-treatment. Clumping is a generF,l feature and
sometimes club-shaped bodies protrude out from the clumped
masses. Pachytene ohromosomes, whenever reoognizable,
possess several ga.ps. It is found that the effect of
caffeine, though highly toxic in the 0.9 fo and 1.0 %
oonoentrations, does not interfere with pairing of
chromosomes at pe_ohy-tene. Frequently the p[;chytene
ohromosomes disintegrate into numerous stron:rly staining
fragments of different sizes.
2. Qualitative studl : Caffein~
The seoond lot of' the 35 - days old males of
C. traohyoterus was used for this analysis. It is observed
in the quantitative analysis that the 0.4 % conoentration of
oaffeine is cuite potent. Moreover, it does not oause
mortality. But with 0.5 % ooncentration onwards, the percen
tage of mortality increases. Beopuse of these two reasons,

-79-
0.4 10 caffeine has been seleoted to study the effect w.nen
the treatment time is being altered. Testes from five males
of the oontrol- and treated-groups were removed at the follo
wing intervals: 4, Id t 43, and 72 hours Emd 6, '7, 8, 14,
and 21 dDYs post-injection. As is clear froIn Fig. 9,
these intervels involve 8,11 thE) stages of spermHtogenesis.
The abnormalities can conveniently be put into four
categories -
(i) mitotic changes;
(ii) changes in the meiotio prophase stages;
(iii) changes at metaphase I and 11;
(iv) changes at first and second post-metaphase stages.
u. Control Series :
In this series, except 'pseudo-chiasmata', no
perceptible ohanges from the normal meiotic behaviour of
the chromosomes h~.s been noticed. There is some general
effect, ho~ever, in comparison to normal individuals. The
disposition of chromosomes in the control series is
relatively clear with some shortening effect.
b. Treated Series :
(i) Mitotic ohanges : not found.
(ii) Changes in the meiotic prophese stages :-
1) High condensation: This abnorm::;l! ty
generelly afieots all the bivalents at
diakinesis. The chromosomes :c;re so thick
that no hollow, oentr~d sp;;l.ce CBn be

-dO-
observed in tho.3e bivc.l(;nts where the two
interstitial chi&sm~ta huvs termin~lized.
The ·l·~.mp-brush' 8flpearance is COIDI)letely
miss.ing and the po;::;i ti vc heteropycnosis of
the sex-chromosome is not distinguishable
( Fig. 94 ).
2) Lesions J Erosion of chromatin occurs at
diakinesis ( Fig. 95 ). About three biva
lents of each plate possess this abnormality.
Lesions a.ppear as stain-less notches or
bands along the arms of the chromosomes.
Sometimes the outline of the chromosomes
becomes wavy or irregular due to the
dechromatization.
(iii) Changes at metaphase I and II :-
These stages are most vulnerable to the effect
of caffeine. The chromosomes ore relatively
contracted. The abnormalities app::.rently
depend u.pon the duration of treatment. with
the increase in treatment-time, more and more
cells ere victimized.
1) P?lXEloidl : Most of the cells at metaphase
I1 possess tetreploid complement. The latter
has the chromosomal configu.ration
36 A + 2 X. It is not possible, however, to
distinguish the sex chromosomes ( }'ig. 96 )
because all the chromosomes 8re over-

L~~ __ _
e; ~ ~ , ,
~
I ,. •
• ::. -/ l
,
F1g.94
'. -" " .. \ ..
•
.. ) ( ___ e
~*- "I /. . )
Q ' "- ,~
..... Q '1
( .., . e.. . . . ;.
Fig.95
Fig.96
v
,~~ , •
•
o " -
I

-81-
condensed end stain darkly. It is clear
from the disposition of the ohromosomes
that the spindle is um~ffeoted, though it
might h~1ve failed at anaphase 1. The
chromatids ( daughter autosomes ) of the
univalents divide and lie parallel to each
other. Sometimes the p.artners m<~iy drift
apart. The cell wall is fOund to be intact
in most of the oaoes.
2) Q-1~.E~nt$: This is the most frecuently
occurring abnormality, involving more and
more cells with the increa.se in treatment
tirr:.e. Ini-tially, only three or four chromo
some. of the metc,phflse 11 plates 8re involved
( t'1g. 97 ). Later on, the clwnping is seen
to involve all the chromosomes ( Fig. 98 ).
The i:ient1ficctioll of the stage ar:rd chromo
somes is, however, possible und a few
norm~?l cells C[,;ln ca observed in the vicinity
of the affected cells ( Fig. 90 ). After a
further increase in the troatment-time, the
clumps become more intense. The identifica
tion of the stage is possible but the
indi vidu~Jl iden"ti ty of the chromosomes is
extremely diffioult. Moreover, the cell walls
cannot be locGted ( i'ig. 99 ).
3) Spindle abnormfci.li t.z: Irwo 3ub-typ€;s of this

, ~ .. ,~f "t. k ..r ':, ... { .. ...... • ~.
• r
.," .. ~ ... J:Oll-~ ""
~, ... ~
~r-
~ ,~
6 "0-
~ t"*
c • ~ /1/ \\\1 • ~~ t ~.
C • ~-
If ...
tr ":" \ • ,. •
#
~ '" • • F1g.91
- I 1-
~ • 'I:'
IIj * ~'t
,~ t;r ~~ -e
-I Iv -Fig.98
-
I -I , Fig.99

-82-
abnorm:.31i t,y ere met VIii th in the prosent
study. In one sub-type, t~e spindle is still
intact but the irr·egul~.lri ty in the arrange
ment of metaphase 11 - chromosomes reveals
that the fibres of spindle are not in align
ment ( Fig. 100 ). In the other type, the
disruption of the whole spinde apparatus has
occurred ( Fig. 101 ). The univalents with
their tii vided chromatids lie haph"lzardly t
usually beyond the norma.l dimensions.
(iv) Changes at first and seoond post-metaphase stages :-
1) Breakage: This abnorIDslity is not very
frequent and usue:lly the smallest, and
sometimes one medium, Butosomes are involved.
The broken piece, though well-separated from
the heteromorphic univalent, passes to the
same pole ( Fig. 102 ).
3. Qualitative studl : Enteromyoetin
(1) Mitotio changes
1) Lesions I This abnormality affects the longer
chromosomes of the complement, especially the
sex-chro~some ( Fig. 133 ). In the latter
case, the lesion occurs in both arms at the
same locus, murked off cler:~rly by stain-less
bands. The fact tl1at the same region is
most susceptible to the effect of enteromyoetil'

.. •
•
~ . •
,. ,..
Fig.lOO
\
Fig.lOl
•
,
•
Fig.102

-33-
is ms·de clef:r in ~ig. 104. This figure also
ShORS the lfurgest autosome with lesions in
both chromatids at the same locus.
(ii) Changes in the meiotic prophase stages :-
1) :area.ka~e : The antibiotic produoed breakage
at the ohromosome-level at diplotene. The
broken piece lies well np~rt from the
heteromorphic bivalent ( Fig. 105 ). ~o
chromatic connection can be seen between
the two. This abnormality usually involves
2)
the longer bi valen ts 8.nd the number of
breaks per cell never exceeds one.
EollEloidZ : A good number of cells tit
diakinesis are tetraploid in nature (Fig.106).
Such cells aoquire greater dimen~1ons and
the bivalents lie haphazardly within. The
chromosomal configurations are.
4n = 38 ( 36 A + 2 X ). No multivalent confi
gurations [;ire met wi th. The sex chromosomes
lie ~part, never showing any tendency to
synepse. The degree of heteropycnosis is
relatively less marked in the sex chromoso
mes, which stain almost (:l.S intensely as the
autosomal bivalents.
(iii) Changes at metaphase I and 11 :-
1) FraiZIDentation : Metaph£cse 11 is found to be
most vulnerable to the effects of this

•
·'1. .. , ~. '"
Fig.lD3
-Fig.1Q4
') " c
,
Fig.105
r~ ...
;

-84-
antibiotic. A study of the victimized cells
shoV'.s that this effect shows no response to
the increase in treatment time. The gradual
steps of fragmentation can be seen ill
Fig. 101. Here, the first step comprises
'notches',. The latter provide a pseudo
appe;::,rance o:f • ea.telli te' to the daughter
chromosomes. In the next step, the fra~nent
shows the signs of detachment because an
achromatic gap appears ( Fig. 107, arrow
mark ). In the lest step, the fragment lies
freely after detachment. Four or more
dt:ughter chromosomes show this type of
fragmentation. Another pl[lte at the rig.t.t haud
side ( Fig. 101 ) shows a number of frag
ments.This type of abnormality comes under
the delayed effect of the antibiotic.
(iv) Changes at first and second post-metaphase stages :-
1) Stickiness: This abnormality he.s been found
in combination with polyploidy at (::naphase
I. The latter stage possesses the tetraplOid
complement. It is seen tbat at the beginning
of anaphase It the bivalents get arranged
on the eoubtor, but are unable to separate
( Fig. lC)8 ). All the chroi'tosomes appear
highly condensed so that no separetion CCln

~ .
1'\
" " 1 -t.~.c..
<.~ -: +:
• ,.' c. ••
,. ~ c: .. • c.. .,..~
~ 1'\ ~\ , - .. '" -J.j
, e -', t\
~
~ {
F1g.106
r
•
Fig.107
/"'~,i~' sep . univ
i" Fig. 108
, '

-d5-
be observed in between the chromatids. The
morphology can be mistaken for the metapha.se
11 except for the orientation of the bivalents,
The latter show the polarity characteristic
of anaphase I stages. Similar tipe of
stickine::;s h,3s been reported by Manna and
Bhunya ( 1971 ) in their study on the effects
of tetracycline on the chromosomes of
Spathosternum praain1ferum.
2) Spindle abnormalit~: This a.bnormality is
most common in anaphase I cells. When the
complement divides normally, the two poles
of the spindle receive 9A + X and 9A
complements. In the affected cells, the
9A + X- pole is always found to be normal
( Fig. 109 ) but the other pole is bifid.
Such cells acquire a tripolar appefirance. e.
The bifid pole usually shows some spcificity 1\
about the distribution of chromosomes. The
smaller group contains three, and the bigger
six, Butosomes ( Fig. 109 ). Moreover, the
chromosomes of the bifid pole enter telopha-
se earlier than the chromosomes of the normal
pole ( Fig. 109, right cell ). This type of
spindle abnorml:\li ty ha.s not been reported
so far.

.. <,
·~ •
, bN-. , bi f. pl ---
bif .pl ___
Fig.109
'~'---- ·chd . br
Fig.110
Fi g. III

-36-
4. Qualitative study: Formaline
(i) Mitotic ch8nges : not found.
(ii) Chani:~es in the meiotio prophase stages :-
1) Dechromatization and breakase : This non
delayed effect projects a number of achro
matic regions in the diplotene-chrolliosomes.
The lat-ter weaken the Etrchi tecture of the
chromosomes ruld usually caU8e breakage at
the level of chromosom.es ( Fig. 110, arrow
mark ). The bi vEtlents usually lie far apart
indicating that the cell-wall has been
destroyed by the chemical. The oharacteristic
'lamp-brush' appearance is also reduced or
missing from the diplotene stages.
2) Interbivalent connection : This is also a
non-delayed effect, in which two or three
biva.lents get conneoted ( Fie. III ). The
connection is revealed by a clrromatin-
thread which stains lighter than the bivalents,
Manna and Parida ( 1965 ) have reported
similar type of connections in their study
on the differentially administered colchi
eine effects upon the testis cells of
Spathosternum prasiniferum.
3) Breaka~e : This effect, though non-delayed,
is of lesD frequent occurrencey The break is
at the chromutid level ( Fig. 112 ) and is

Fig.112
Fig.113
lI" o/COnbV~
~ . ·fA
Fig.114

-'37-
found in one of the medium bivalents. The
sex-chrowosome remtins unaffeoted. Forma
line does not influence the frequency of
chiasma at diplotene sta.ge.
4) ClUlllpintS : The abnormality is found mostly
in the cells at diakinesis. The bivalents
thicken and stain darkly. Besides, the sex
chromosome ls identifiable on tee basis of
morphology alone and not because of its
heteropycnosis. It stains 8S da.rk bS the
autosomal bivalents, The latter form small
'clumps'. The intensity of clumping increa
ses with the time of treatment. In the
e;-;!rlier stages two or three bi valer.. ts stick
together ( Fig. 113 ). Their morphology is,
however, quite distinct.
(iii) Changes at metaphHse I and 11 :-
1) Inter-bivalent connections : This effect is
similrir to the one desoribed earlier for
meiotic proph::;!se stages. But here five. and
sometimes six, bivalents are connected in a
linear fashion ( Fig. 114 and 115 ). Besides
the signs of erosion of chromatin-ma.terial
are also there. The 'lamp-brush fibres' are
obviously les8 prominent. This trpe of
appecJI'ance in the prometaphafJe stage has been
reported by Manna and Parida ( 1965 ) in the

LO .-,.
-. F1g.115
F1g.116
F1g.117

-88-
spermatocytes of Spathosternum~r~~1n1fer~ /",
Fig. 115 shows the occurrence of breaks
besides the ohain of three bivalents.
2) Destruction of morphologz: The chromosomes
of metaphase 11 stage are most affected by
formalin and possess a number of abnorma-
lities. It is observed that the chromosomes
appear thicker than usual and are a.bnormal
in their orientation ( Fig. 116 ). Their
outline is fuzzy and irregular. Usually two
to-three chromosomes stick together to form
small groups. In one cell, four such groups
have been found at ec~ual distances in a
ouadrangle ( Fig. 116, upper cell ). It
gives a 'pseudo-tetranucleate' appearance
to the oell. The stage can be identified,
however, because one or two of the chromo
somes maintain their identity. A pachyten,e
nucleus in Fig. 116, exhibits the presence
of 'snail-horn'-like centromerio-chromomeres
in the synapsed homologous chromosomes.
3) Lesions: This abnormality is a common
feature in the metaphase II chromosomes. It
shows a speoifio affinity towards the sex
chromosome. The latter possesses either one
( Fig. 117 ) or two ( Fig. 118 ) lesions.
Manna (1969) has reported that chemicals

•
•
•
•
F1g.118
". "'-u.s .r , • ,I * .,. J ~ slr."'n
,.
• . - .
I

-89-
like alcohol, 108d nitrate, terramycin, maleic
hydrazide, BUDR. corline and formaline, etc.
affect the X chromosome more than the auto-
somes. The present observation confirms this
specific nature of formaline. The autosomes,
though devoid of lesions. show tUl aberrant
morphology ( Fig. 111 and 118 ). Their
orientation is, however, normal.
4) stretched centromere and dechromatization :
These two abnormalities are also confined to
metaphase 11 chromosomes. The erosion of
chromatin depends upon the time of treatment.
In the beginning it is less pronounced,
oocurring in onl~ some of the chromosomes
( Fig. 119 ). Large achromatic gnps can be
seen in a few of the chromosomes. The
stretching of oentromere is confined to the
longer ohromosomes ( Fig. 119 ). With the
inorease in treatment-time, all the chromo
somes look wiry and lightly-stained (Fig.120).
Moat noticeable is the X-chromosome with
m[J.ximum dechromatization. The oocurrence of
darkly staixling blobs at the telomerio ends
indioates that perhaps the process of
dechromatization proceeds from centromere
tow6rds telomeres ( Fig. 120 ).
5) star-shaped .confi~rati~ : The metaphase 11

-90-
chromosomes 2ssume 'ball' or 'star' shaped
configurations ( Fig. 121 ). In this
abnormali ty the proxirn5.1 ( centromeric )
end~3 of the chromosomes are direoted towards
a sinele pOint in the cell. This polarity
along with the olose approximation of the
ohromosomes results in the darkly staining
• start oonfiguration. The stage oan be
identified because of some of the chromo
somes which have not contributed to the
star-oonfiguration ( l'ig. 121, arrow mGrks).
Such type of abnormality has been reported
to be caused by oolohicine ( Manna and
Parida, 1965 ). Fig. 121 8180 shows another
metaphase 11 cell ( left-hand side) with
aberrant morphology of the chromosomes.
(iv) Changes at first and second post-metaphase
stages: not found.
5. Qualitative Study: Nicotine
This is found to be the most potent drug and
resulted in mortality if the treatment-time exceeded one
week.
(i) Mitotic changes :-
1) Fragmentat.!2!! : The drug inflicts heo.vy
fragmentation in the gonial metaphases.
The breaks are entirely of the chromatid
type ( Fig. 122 ), and do not show any

•
•
•
st . confg~
.... ,. F1g.121
F1g.122
br . p I • -f
"I
I br . p- br
Fig.123
•

-91-
reunions. The latter observation is based on
a large number of cells having numerous
fra,gments. No ring-like structures and no
exchange type configurE'.tions have been found.
The broken ends thus appear not to have the
oapao! ty to reunite, f.l.or probably to resti
tute. Similar type of effect has been repor
ted to be caused by Actinomycin-D on the
chromosomes of Sohistocerca gresaria ( Jain
and Singh, 1961 ). The sex-chromosome, in
the present investigation, remains unaffeo
ted and shows its oharfloteristic negative
heteropycnosis ( Fig. 122 ).
(ii) Ch2n,zes in the meiotic proI?hase st~5es :-
1) Brea.kage : This abnormality is observed in
a large number of diplotene stages. The
breaks oocur in two or three of the bivalents.
The broken pieces lie far away from the main
part ( Fig. 123 ) and are apparently
acentric in nature.
2) ClumI!in~ : It ha.s been observed in diplotene
and diakinesis stages. The bivalents appear
muoh thickened and close together forming
f bf.llls t ( Fig. 124 ). The stages can be
identified, however, because of single
strr,y bivalents lying outside the clumps.

I
Fig.124
i ,
x--
" 1
Fig.125
,~"
I )
••• ,~ : .. , , ~ -•
le-{~ • • t".
) (,
I -11
Fig.126

-92-
(ii1) Cha.nges at metaphase I and 11 :-
1) Clumping: This abnormality is very common
in meta.phase I ste.ges. The intensity of
clumping is not much, however. Usually two
or three bivalents stick together forming
one or two ,groups ( Fig. 125 ). The
orientation of the bivalents remairls
unaffected. The sex chromosome, in most of
the oases, does not include itself in the
olumps.
2) Lesions : This effect is most frl:quent in
metaphase I1 oells. All the ohromosomes,
except the smaller ones, possess lesions.
Fig. 126 shows a diploid cell with lesions
in the chromosomes. The latter have either
wavy or notched outline. Some of the chromo
somes h8ve divided cmd the daughter
chromosomes have drifted ap2rt. The sex
chromosolLe carl not be identified. The
disposition and orientation of ohromosomos
remains normal.
(iv) Changes a.t first and second post-metaphase
stages: not found.
::.::*=*:::*=*:::*=*=*= =*=*=*=
=*=

VI
RESULTS AND DISCUSSION

-93-
RESULTS AND DISCUSSION
A. Meiosis and Mitosis
The speoies of grasshoppers included in the present
investigation can be split into two major groups - 23 -
chromosomes types and 19- chromosomes types. The former
group includes TriloEhidia annulata and Leva indic~. The
latter group includes Chroto5?nus trachypterus,
Attractomorpha orenulata end Poekilocerus pictu~. The analysis
of caryotypes in ::ill the species includes a straight compa
rison of the dimensions and morphology of meiotic and
mitotio complements. Any establishment of relationship of
caryotypes in the speciel2>, is not the Elim of this work. The
main interest of the auther lies in a critical review of the
orthodox beliefs about the caryotypic stability and nature
of centromere in the Acrididae.
The oryptosaccine grasshoppers, T. annulata and
L. indica, possess 22 autosomes aIld single X chromosome ill /',
their male diploid complements. The chromosome behaviour
during the spermEtocyte-divisions is orthodox and is in no
way differen~ from the other species of grasshoppers with
23 chromosomes. The two apeoiBs differ in t.he length of the
X chromosome. The latter, in ~~ annulata, stands seoond in
, order of length, while in L. indica, it 1sI"longeat in the t complement. The lengthwise grouping of the chromosomes is
also different in theSE} species. T. annul~;lta possesses 3
long , 7 medium and 2 short chromosomes ( sub-family

-94-
Oedipodin[:e ) and ~. indics possesses 3 long, 6 medium. and
3 short ohromosomes ( sub-family Gomphocerinae ). A oompari
son of the lengths of' both the serminal and somatio ohromo
somes reveals that the ohromosomes of L. indioa are smaller
than those of T. annulata. The dimensions of the longest and
smallest chromosomes in the germinal tissue of' T. annulata
and L. indics are 7.38 miora/ 5.40 micra and 1.37 micra/
1.10 miora, respeotively. Likewise, in the somatic tissue,
the dimensions are 8.39 miora! 6.<57 miora and 2.47 micra/
2.41 micra.
It is obvious f'rom the oomparison of' lengths that
in both the species, the somatio chromosomes condense to a
lesser degree at metaphase than the gonial metaphase chromo
somes. The behaviour of the sex chromatin differs in the
germinal and somatic tissue. In the former, it squires a /',.
distinot 'horse-shoe' shape during meiotio prophase I. No
heteropyonotio tissue can be found in the somatic tissue of'
male grasshopper. In the females, however. an indistinct
heteropycnotic mass is detectable. A related disoussion
follows in the foregoing account.
A study of chiasma frequency in T. annulate and
L.indica reveals that the loss of chiasmata is more between
diplotene and diakinesis in the former, while in the latter,
it is stronger between diakinesis and metaphec,se I. The number
of chiasmata in both the~ecies never exceeds three in the
long bivalents Hnd two in the medium bivalents. The chhsma
frequency of T. s.nnu18.ta is a little higher than that

-95-
of L.indica.
All attempts to locate the second arms, especialll
1n the less condensed metaphases of the somatic tissue,
proved fruitless. It 1s most tempting to regard the tsnail
horn~- like appearance at paahytene stages of T.annulata
( Fig. 34, 35 ) as second arms. Such structures are,
however, possessed by the centric chromomeres ( Marks,1957),
which become very prominent when sister centromeres are
in a state of repulsion.
The chasmosaccine grasshoppers, c. trachypterus,
A. crenulata and P. pictus, possess 18 autosomes and single
X chromosome in their male diploid complements. The behaviour
of the chromosomes during the spermatocytic divisions is
orthodox and is in no way different from other species of
grasshoppers with nineteen chromosomes. The caryotypes of
c. trachypterus and A. crenulata differ in the length of the
X chromosome. The latter, in c. trachypterus, 1s the longest
in the complement, while in A. crenulata it is second in
order of length. The lengthwise categorisation is,however.
similar in all the three species( sub-family Pyrgomorphinae),
consisting of 3 long, 6 medium and 1 small chromosomes. A
comparison of lengths of chromosomes in the three species
shows that the chromosomes of c. trachypterus are smallest
in size. The longest chromosome in the germinal tissue of
c. trachypterus, A. crenulata and P. pictus measures
5.85/8.64/9.11 micra, respectively. The smallest chromosome
in the germinal tissue measures 1.15/2.55/3.14 miora in the

-96-
same order. The data from the somatic tissue are 15.64/10.69
/14.16 micra for the longest chromosome and 3.88/3.12/5.64
micra for the smallest chromosome.
The above oomparison makes it clear that in all
the species, the somatic chromosomes condense to a less
degree at metaphase than the gOnial metaphase chromosomes.
c. traohlPteru8 and A. crenulata show some inter-individual
differences. The lengths of germinul chromosomes of the
latter species are higher than those of the former speoies.
But the order is reversed for the somatic tissue, where
the chromosomes of c. traohypterus are longer than those of
A. crenulata. An apparent reason for this behaviour seems
to be tha,t dehydration at late prophase, which results in
the gradual condensation of ohromosomes, is less in the
somatic tissue of A. crenula,ta than in c. trach,ypterus.
The somatio-metaphase ohromosomes of P. pictus are remark
able in a.oquiring a length almost double that of their
germinal oounter-parts.
A study of chiasma frequency is worked out for the
local ( Bhopa.1 ) population of c. trachlEterus and is
oompared with the populations of the same species belonging
to Delhi and Ottu ( Punjab ). In the Bhopal - population
the loss of ohiasmata between diplotene and diakinesis is
stronger, as in the other two populations, but the mean
chiasma frequency per nucleus differs slightly.It is less
than the Delhi-population, and almost similar to that 0' the Ottu ( Punjab ) - population.

-97-
In all the three species, the sex - chromosome
does not show posi~1ve-heteropycnosis in the prophases of
male somatio tissue. At somatic metaphase, the degree of
oondensation is uniform in the autosomes and the sex
ohromosome. The identification of the latter, therefore,
depends solely on the morphometric data of the somatic
ohromosomes after a oomparison with the germinal chromo
somes.
All attempts to locate the seoond arms are made
futile in C. trachypterus and A. orenulata. The former
speoies possesses the 'snail-horn'-like structures in its
pachytene nuolei ( Fig. 116 ) oomparable to similar struo
tures of !. annulate. In P. pictus, however, the smallest
autosome, along with one of the medium autosomes, displays
distinct second arms and makes the term 'acrooentrio'
partly justifiable. But there is some oontroversy about the
oocurrenoe of seoond arms in this speoies and a variety
of opinions is available. While Darlington ( 1936 ) and
Coleman ( 1943 ) agree about the • true' acrocentric nature
of the smallest autosome, Dutt ( 1955 ) olaims that in rare
instances, the seoond arms are visible in all the chromosomes.
In the present study, however, seoond arms OanllOt be located
in all the chromosomes.
The heteropycnotio behaviour of the sex ohromoso
me, in the prophases of somatio cells, is a oontroversial
matter and has been reviewed by a number of scholars from
time to time. The mysteries involving the funotional and

-98-
morphologio differentiation of the X-chromosome have begun
to unravel only in the earlier decades of this century.
The oytologio evidenoe for its peculiarities of behaviour
was observed even before its role in sex - determination
could be reoognized. Henking in 1891 ( Wilson, 1925 )
first observed a "peouliar ohromatio - element" in the
gonadal oells of Plrrhocoris apterus which during sperma
tocyte-diVision became greatly condensed, divided equationa
lly in the first division but moved undivided to one pole
during the seoond anaphase. Aooording to McClung ( 1902 ),
"Montogomery ( '98 ) deserves the credit for observing that
this body is merely a chromosome of the spermatogonia that
pursues a somewhat different course from the others".
The unusual heteropyonotio behaviour of the
X-ohromosome which Henking first observed in the spermato
oytes of Pyrrhoooris is now known to be a widespread
charaoteristio of the X-chromosome of many speoies of inseots
and other animals ( Wilson, 1925 ). sttldies on female
germinal cells reoeived co~paratively little attention but
oooytes from various Hemiptera failed to show any intensely
staining ohromosomes that were so characteristic of the
spermatooytes ( Wilson, 1906 ).
Conspiouous heteropyonosis of the X-chromosome
was observed not only in gonadal cells but also in somatio
ntlolei. Geitler in 1937 ( Mittwooh, 1964 ) found that in
somatio interphase ntlclei from both heterogametic ( male )
and homogametic ( female sexes of insects, the X chromosome

-99-
remains heteropyonotic. The oells from the female show two
heteropyonotic bodies and from the males only one. Later
numerous investigations on the chromosome constitution and
sex ohromatin of mammals revealed that the number of sex
ohromatin bodies in diploid oells is always one less than
the number of X-ohromosomes ( Mukherjee t 1965 ). It
gradually became very olear that the behaviour o£ the
x- chromosomes in somatio oells is exaotly opposite to that
observed in germ cells. The single X- chromosome in
heterogametic males becomes heteropycnotic in spermatocytes
but remains isopycnotic in the somatic cells, whereas both
X-chromosomes in homogametic females remain isopycnotic in
oocytes, but one of them becomes heteropyonotio in the
somatio cells. The latter faot applies to the present
investigation also.
An attempt was made to explain this behaviour of
X-chromosome in the female somatic cells through the
"single active X hypothesis" ( Lyon, 1961; Russell, 1961 ).
The hypothesis states that the heteropycnotic X-chromosome
of the female somatic oells is genetioally inaotive. Suoh
an inactive X oan be either paternally or maternally
derived in different oells of the same animal. This
differentiation of the X-chromosome takes plaoe early in
embryonic development a.nd remains oonstant f'or all
desoendants of the differentiated oell. Further observations
showed that the sex chromatin mass in the interphase nuolei
of females is late replioating ( Atkins et al, 1962; --

-100-
Mukherjee and Sinha, 1963 ). The present status of the
hypothesis also inoludes two more statements that " more
probably, heteropyonosis alternates between the two XSs
in a female somatio nuoleus regardless of their parental
derivation tI ( Ohno and Hauschka, 1960 ) and that the
differenciation of the inactive X takes place some time
during the early embryonio life ( Lyon, 1961, 1962 ).
The nature of the centromere is the most contro
versial aspeot of the Acrididae. Two oonflioting views
relating to the rod-shaped ohromosomes exist. Aooording to
McClung ( 1914 ) and Makino and Mamma ( 1950 ) some, at
/' least, of these ohromosomes are genuinhely telocentric.
White ( 1914 ), Darlington ( 1936 ) and Coleman ( 1943 ),
on the other hand, have all argued that these rods are
acrocentrio but that faulty fixation and unfavourable
staining frequently obscure their real struoture. The
latter opinion was based on two lines of evidenoe :-
(i) Small heads are frequently visible at
metaphase both in spermatogonial and at
seoond meiotio division.
(ii) So-oalled ditaotio bivalents are sometimes
produoed, in whioh assooiation is maintained
exolusively by what is interpreted to be
the short arms.
It id worthwhile, at this pOint, to review the
oriteria chosen for distinguishing objeotively between
telocentrio and aorooentrio ohromosomes :-

-101-
(i) The oocurrenoe of small granules at the
oentrio end of rod ohromosomes is not an
adequate indication of short arms sinoe
olearly these oould represent oentromerio
ohromomeres, as was believed by Makino and
Momma ( 1950 ) also. Indeed in terms of the
known struoture of metaoentrics ( Lim~-de
Feria, 1956 ), a telooentric ohromosome is
expected to contain centric chromomeres and
there oan be little doubt that these have
frequently been taken to represent a seoond
arm. It is on the basis of such granules that
White has repeatedly argued for the occurrenoe
of short arms in orthopteran rods.
(ii) As Ostergren, Mole-Bajer and Ba.jer ( 1960 )
po:l,nt out it is possible in fixed and stained
preparations "to see a specieol fibrillar
material that is attaohed to 6ach oentromere
and extends from it in the direotion of the
spindle pole toward whioh the oentromere is
oriented".
(iii) Sinoe oentromeres are reletively unooiled at
metaphase of mitosis they appear both thinner
~nd less deeply stained than the main
chromosome arm. Moreover, as Jones and Colden
( 1968 ) point out, a terminal centromere oan
be expeoted to show enhanced cle.ri ty of

-102-
structure compared with that of metacentrios,
beoause unlike the metacentric, its centromere
region is not subjected to the effect of arm
coiling on both sides.
(iv) At anaphase short arms sbould flex as do long
ones. Telocentrios on the other hanl, are
expeoted to move with their centric ends
pointing directly to the poles ( Levan, Fredga
and Sandberg, 1964 ).
That these oriteria are justifiable is confirmed
by Marks' ( 1957 ) observation on Oxalis dispar. Here two
small stained bodies are visible at the centric end of the
telocentric members. When sister chromatids are in a state
of repulsion they produce a structure "reminiscent of the
horns of a snail". When repulsion is not evident the terminal
oentromeres appear as small, faintly stained. tapering ends.
The present study oonfirms these points.
White ( 1957 ) has argued that "failure to observe
the minute seoond arms should not neocessarily be regarded
as proof that they do not exist, since even in those forms
in whioh they undoubtedly oocur they oan only be seen in
exceptionally well-fixed oells." On the basis of the oriteria
listed above one may feel disinclined to believe in invisible
short arms. The demonstration of absbnce of short arms is
unlikely to convinoe those prep8red to believe in the
invisible. Beoause of the same reason, one cannot accept
White's seoond rule for it is abundantly clear that in many

./
-103-
species the centromere does not lie in a heterochromatio
region.
In many telocentrics the struoture of the centro
mere appears to be equivalent to a half, or at least less
than the whole, of a metaoentrio ( John and Hewitt, 1966b ).
This suggests either an origin by misdivision of a metaoen
tric type or else the production of the metacentrio by the
fusion of two telooentrio centromeres. In other oases the
telo-oentromere seems to be essentially identical in
structure with that of the metaoentrics found in other
species ( Tjio and Levan, 1954; Johes and Colden, 1968 ).
This oould be taken to imply either that the telooentrics
have arisen not by misdivision but rather by whole arm
erosion following breakage proximal to the centromere. Such
an event might be expected to oocur more readily in acro
rather than meta-centric since the extent of material lost
would be small.
Actually the ideas concerning relation-..:,ship
between the rod-shaped and V-shaped chromosomes is clearly
dominated by the belief that strictly telocentric chromosomes
either do not exist ( Nawaschin, 1916; Lewitsky, 1931;
Muller, 1940 ), or if they exist, are unstable ( Rhoades,
1940.)' or if stable they are rare ( white, 1957, 1959 ).
There is, therefore, a long and unfortunate histary to the
whole issue and one which still prejudices contemporary
thinking on the subject.
As is so often the case in science the truth may

-100-
well lie between the two viewpoints discussed earlier. The
author feels more inclined to regard the 'snail-horn' like
structures as oentrio ohromomeres and not the seoond arms.
This clearly demaroates the chromosomes of T. annulata , . c. tra~ypterus, L. indioa and A. orenulata as telooentrio
in nature. This assumption includes the positive evidence
provided by the chromosomes of the somatio tissue ( gastrio
oaeca ). The ohromosomes of P. pictus, however, apparently
form an asymmetrical oaryotype, because the seoond arms
are visible in two ohromosomes only. The details of the
nature of oentromere in this speoies are discussed in the
section of 'banding pattern-.
B. Banding Pattern of P. piotus
In the present investigation, the germinal
chromosomes were subjeoted to Giemsa staining (sumner,
1972 ). The regions with oonstitutive heteroohromatin
stain heavily with Giemsa and a general pioture of the
banding pattern oan be drawn.
From the observation of the behaviour of the
X-ohromo some , it beoomes very clear that the condensed
stat~ is not responsible for the staining reaotions.
Normal staining reactions distinguish X as negatively
heteropycnotio at gonial metaphase, meiotic metaphase I
and anaphase I; and positively heteropyonotio at meiotic
prophase I in P. pictus. But after the treatment for

-105-
C-banding, the X ohromosome stains poorly except for the
centromeric and telomerio bands of heterochromatin. Its
general state of condensation is, however, distinguishable
from the autosomes after applying this technique. Besides,
the teohnique is specifioally reliable for the location
of constitutive heterochromatin.
The characteristic 'bouquet stage' oan be demonstra
ted with this staining method because the orientation of
the prOximal and distal ends with the dark bands becomes
very olear ( Fig. 83 ). Another interesting point noted
by the author is that the sex chromatin, which stains
lighter than the euohromatin, occupies a position opposite
to the • dark pole' B,t leptotene. The • dark pole t is a
oonsequence of bouquet arrangement, formed by the approxi
mation of centromeric and telomeric ends. Since all the
centromerio ends bear darkly staining bands, their
approximation results in a prominent body, referred by the
author as 'dark pole'.
The differentiated oonstitutive heterochromatin
comprises the oentrio ohromomeres and the heterochromatio
region of the proximal part of the chromosomes ( Figs. 84,
85, 86, 87, 88 and 89 ). This oan explain the four SUb-units
of the C-band visible in the X-chromosome ( ~ig. 82 ), out
of which two proximal ones belong to the centrio chromomeres
and the two distal one:::: to the proximal-heterochromatio
bands of the two chromatids. The sub-units come to lie
together when the ohromosomes are comparatively oondensed,

,/
-106-
as during gonial metaphase, diakinesis, metaphase I and
anaphase I, and metaphase 11 and anaphase 11. On the other ,<
hand, the sub-units beoome separated when the ohromoaomes
are less oondensed as during leptotene ( Fig. 83 ),
zygotene, paohytene ( Fige. 04-88 ) and diplotene ( Fig. 89).
That the bands oomprise four sub-units, is further confir
med at metaphase 11 where eaoh ohromatid ( daughter
ohromosome ) contains only two sub-units ( Fig. 93 ). The
latter look like one due to condensation of the chromosomes,
exoept in M4• The oocurrence of four sub-units of
C-bands was reported in the metaoel1trio ohromosomes of
Mz!meleotettix maoulatu:! by Gallaghar, Hewitt and Gibson
( 1973 ). They made no mention of the sub-units in the
telooentrio ohromosomes of the same species. It is felt by
the author that the 'four' sub-units of the metacentric
ohromosomes of M. maoulatus ( where no separation of the
ohromatids was visible ), would have resolved into 'six'
sub-units if the ohromatids were separated. < Thts demons
tration is possible in the lesser condensed stages. On the
other hand, if it is preferred to count the individucd
sub-units 9f the two ohromatids as one, then the four
sub-units found in the present study, may be counted as
three. Out of these three sub-units of C-band, -two belong
to the oentrio ohromomeres and one to the proximal
heteroohromatin of the chromosome.
It is possible to locate the heterochromatin at
all stages of meiosis. At diplotene, the chiasma is found

;' !
-107-
to oocur dietsl to the centric ohromomeres .and proximal to
the proximal heterochromatin of the chromosome. Similar
("'V '" behaviour was noted in Brlodema tuberculcta Klaaterska,
\\n·,
Natarajan and Ramel, n in son::e of the chromosomes.
The size oft the densely staining C-band.::! aeE::.JlS
to be uniform in all the chromosomes. The Giemsa-staining
of the rest of the genome, apart from the constitutive
heterochromatin, is generally quite diffuse. Faint telomeric
blobs of stain t:1.re visible particularly on the X chromo-, some. These have no obvious significance other than the
demonstration of repetitive DNA-sequences. Similer blobs
have been reported in Drosophila ( Hennig, Hennig and
Stein, 1970; Gall, Cohen and Polan, 19'71 ).
The Sl chromosome shows a pattern quite different
from the other chromosomes at all stages of division. Besides
the dark C-band, rest of portion also acquires an equally
dark stain, except for a narrow stain-less region distsl
to the C-band ( Figs. 84, aa and 89 ). The heterochromatin
of the chromosome arm of SI forms two bands ( Fig. a8 ).
Thus Sl is the only autosome in the genome which oontains
oomparatively maximum heterochromatin.
As regards the 'second arms', the present inves-
,-' tigation denies their ocourrenoe. 0rourse. if the centric
ohromomeres are to be regarded as second arms, as White and
followers do, then all the chromosomes are acrocentric in
3ensu stric'tu. The author, however, feels disinclined to
believe in the view. A more intensive investigation is

-108-
needed for a. definite statement about the true nature of the
chromosome.
The abnormalities reported in the present study
inolude :- extra fragment, polyploidy, breakage, anaphase
bridge, abnormal spindle, supernumerary chromosome, non
disjunotion and chromatid-separation.
The extra fragment is found in ~. annulatu,
L. indica and c. t~a~hypterus. In 1_ annul~ta only one
individual out of hundred and six possesses it in 8.57 %
cells at metaphase I. In L. indioa one individual out of
fifty-two possessed the extra fragment in 7.4% cells at
diakinesis. In c. trachlEteru~ one individual out of
eighty-six possesses it in 2.85 % oells at metaphHse 11.
As is apparent from the data, the percentage of aberration
is not very si~lificant in all the three species. The
fragment, in all cases, is the smallest element, in the
oomplement. It is never found to leg behind during anaphase
a!ld thus a.pparently does not interfere wi lih spindle elonga
tion and consequently with cleavage.
The frequency of polyploid cells is also quite
low. In T. annulata 1/106 individual possesses thirteen -tetraploid cells at diakinesis in a single follicle. In
L. indica 1/50 individual possesses 35.41 % tetraploid cells
clt metaphase 11. In C. trachypterus, the polyploid cells are

-109-
observed at diakinesis, metaphase 11 and anaphase 11. The
pooled data show the frequenoy to be 11.10 ~. It is stated
by Rothfels ( 1950 ) and White ( 1951 ) that the
"polyploid oells, in partioular, are not unoommon in
aoridoids". Moreover, ohromosome variations up to 5 ~ are
expeoted in all the individuals from whioh large oell
samples are taken.
The ooourrenoe of polyploidy oan be traoed along
two lines. If the spindle fails to elongate during the
mitotio and meiotio divisions, the tetraploid oells
originate. The latter are quite frequent in the populations
of grasshoppers. Seoondly, if there is a failure of oleavage
during mitosis, the binuoleate oells are formed. The
latter enter prophases separately during meiosis. On
approaohing metaphase, as their nuolear membranes dis
appear, the ohromosomes of both the nuclei come to lie on
a single segment, so that the resulting metaphase is
tetraploid. It is noteworthy that if polyploidy ooours
due to the first reason, the tetraploid oells entering
meiosis oarry a varying number of multivalents when the
spindle fails in one of the mitotio divisions. In
c. traohypterus the tetraploid metaphe.se 11 oell ( Fig. 48)
may have arisen aocording to the given sohedule :-
Step (i)
step {ii}
9 AA + x
9 A + x ; 9 A at anaphase I, due to
failure of spindle elonga.tion.

step (111)
-110-
9 dA + dx ; 9 dA
9 dA + dx ; 9 dA at metaphase 11.
Thus step (iil) gives rise to 36 dA + 2 dx oonfiguration
at metaphase 11 ( Fig. 48 ).
The tetraploid oell at anaphase 11 ( Fig. 49 )
seems to have arisen in a somewhat different manner :-
step (i)
step (li)
Step (iii)
9 AA + x
9 A + x , 9 A • The daughter nuolei
separate after 8,naphase I, but remain wl thin the
same oell-wall due to failure of oytokinesis.
9 ciA + dx 9 dA
========- -- The daughter nuolei 9dA+dx 9dA
of step (1i) divide on
separate spindlesat anaphase 11. This aooounts
for the different orientations of the splndles.
In L. indioa the polyploid oell at anaphase 11 .... , . oontains 44 daughter ohromosomes. The origin of this oonfi
guration can be traoed in the following way :-
Step (1) 11 AA + x ; 11 AA + x
Step (ii) 11 A + x + 11 A .' 11 A + 11 A + x ,
at anaphase I.

Step (iil)
step (iv)
11.A ,
11 dA
11 dA
-111-
11A due to fusion of cells.
11 d.A
11 dA = 44 dA at anaphase 11.
In none of the cases, the polyploid condition
originates due to failure of spindle elongation at mitotio
divisions. Such cases carry multivalents which are not
found in the present study.
The breakage is found in T. annulat~ in 1/106
individual in 1.4 % oells at anaphase I. It Is at the
chroma.tid-level and the fragment lies close to the arm. In
L. indica this abnormality is found in 29.41 % oells at
spermatogonial metaphase and invariably affects the longer
chromosomes only. Both the chromatids possess the breakage
at the same locus. Here also the fragments do not lin6er
away from the arms. In C. trachypterus 4.76 ~ oells at
anaphase 11, and ill A. crenulata 3.33 % cells at anaphase I,
possess breakage at the chromatid - level. In these two
instanoes, the fragments not only form an angle with the
main arm but also lie away from it.
The question whether the breakage is • complete'
Is a difficult one to answer. In T. annulate and L. indica
the broken ends are still quite close together and one
might conclude that no real fragmentation has oocurred.

-112-
White ( 1951 0 ) has desoribed "elastio oonstriotions" in
some speoies of morabine grasshoppers where the two portions
of a ohromosome may sometimes be pulled out to enormous
lengths at first metaphase and anaphase. But in suoh oases
it was oono1uded that the two portions of the ohromosomes
were still connected by a fine thread. In the present case
no thread could be seen in any case, but an invisible
oonneotion may still exist. This seems to be true for
C. traohlEteruB and ~orenulata, beoause the fragment is
never found to lag behind on the equator, neither does it
move to the opposite pole.
The dicentric bridge is found in a single indivi
dual of ~. annula,tl!- , in one cell out of a total of ninety
nine. Similar bridge has been reported in Chr~to~onus
trachlpteru!!, ( Sharma, Parshad and :Eedi, 1962 ). A number
of explanations have been put forth to account for the origin
of dicentric bridge. The latter with or without 1:1 fragment
may occur spontaneously ( Darlington and Upcott, 1941 ;
Rees, 1953; and Manna, 1954 ), or may be induced either by
irradiation ( LaCour and 'Rutisha.user, 1954 ; and Ray
Ch~udhuri, 1961 ) or by chemicals ( De.rlington and Koller,
1941 ; Dar1ington and McLeish, 1951 ; and Steffensen, 1955 ).
The bridges without any fragment, found during
the first and seoond meiotic divisions of Aio1opus ap. and
Tri10phidia annu1r;ta ( Manna, 1954 ), are formed just out of
stickiness in the separating elements. In the present
study t the dioentric bridge of T. annu1~:1ta is accompanied

r
-113-
by a. fragment of oonsiderable size. The bridge does not
appear to be the outoome of B mere stiokiness. It has been
found that the bridge oocurring 8t the first meiotic
division, may arise because of the following reasons :-
(i) Inter/intra chromosome crossing over in re13tively
inverted segments.
(ii) Inverted crossing over.
(iii) Chromosome breakage with sister ohromatid reunion.
(iv) P~.rtial reciprocal or inverted crossing over
between homologues.
(v) Localized a.dhesion between sister or non-sister
chromatids with or without breakage.
(vi) Interlocking of plectonemically ooi1ed stra.nds.
In the first three causes, the bridge is
aooompanied by a fragment ( John and co-workers, 1960 ).
The ooourrence of"" dicentric bridge in the present investi
gation, can only be attributed to second and third causes
because the bridge is not formed by an asymmetrical bivalent
and it has no lobe in the middle. The first cause is not
applicable because the inversion in heterozygous condition
must be represented uniformly in the entire testis or at
1e~st in a port of the testis. In the case under consi-
deration, however, the bridge is found in a single cell of
a follicle. The cause of bridge-formation can, thus, "be
attributed to either the inverted crossing over or ohromo
some breakuse with sister chromatid reunion.
The abnormal spindle is found in L. indica in

-114-
33.33 ~ cells at anaphase 11 in one individu8.1 out of C:i
total of fifty-two. The only probable cause seems to be the
use of colchicine for chromatid separation. But the author
has not came across a similar insta.nce of bifurcation of
one pole of spindle attributed to the use of colchicine.
The occurrence of supernumerary chromosomes is
an interesting feature in the caryology of c. trachlpterus.
The Delhi-population of this species contains 1-7 super
numerary elem8nts ( SharmB, Parshad and Bedi, 1962 ) and
the Ottu ( Punjab ) - population contains the same number
of' supernumeraries along with a megamaric bivalent. In the
present investigation, the Bhopal-population of this
speoies is found to contain 1-6 supernumeraries and no
\ megamerio bivalent. Only one out of a total of eighty-six
\individuals investigated. In the individual under conside
ration the S + 2s condition is most frequent ( 74.70 ~ ).
Rest of the configurations are of lesser frequency. The
S + 6s oonfiguration ienot found in the Ottu ( Punjab)
population, while the S + 1s configuration of the latter
popUlation is not found in the Bhopal-population.
The supernumerary elements show positive hetero
pycnosis at early prophase I ( Fig. 39 ). In the later
stages they become indistinguishable in their staining
intensity with other autosomes and can be differentiated only
due to their size and behaviour. They do not pair with the
X-chromosome or any other autosome of the complement, but
pair with each other. The supernumerary bivalents possess

-115-
single terminal chiasmata.
White ( 1954 ) has classified the supernumerary
chromosomes into mitotically stable and unstable types. In
the former category, all the cells of the germ line contein
the same number of supernumeraries, while in the latter,
there are intra-individual variations between different
follicles, but not within the follicles themselves. The
present material belongs to the latter category.
As regards the oriein of supernumerary elements,
there are two schools of thought. One believes in the
origin of the supernumerary from the X-chromosome
( Robertson, 1917 ; Rothfels, 1950, and others ) on the
bs,sis of tendency of the former to syna.pse with the latter.
The other school believes in the origin of the supernumery
from an autosome ( Carroll, 1920; h~ans, 1954; Ray-
" Ch2~dhuri and Guha, 1955 ). The former sohoo1 of thought
is strongly opposed by Ray-Choudhurl and Guha ( 1955 ) on
the ground that the supernumerary does not exhibit
negative heteropycnosis at early spermatogonial metaphase
( as oonfirmed in the present investigation also ) which
is a oharacteristio feature of X-chromosome in Acrididae.
In the Ottu ( Punjab ) - population of
~ trachY.:Eterus , the origin of super::umeraries is attributed
to the megamerio chromosomes. Since the Bhopa1-popu1ation
lacks the megamerio ohromosomes, the only possible donor
seems to be extra autosomes, included in the standard
complement tl~ough non-disjunction. Suoh autosomes form

-116-
heteromorphio bivalents due to deletion or defioiency and
give rise to supernumeraries. Similar instances have been
reported earlier in Orthoptera - ( Robertson, 1915 ;
Darlington, 1936, and Sharma Parshad and Bedi, 1962 ).
The supernumAraries do not induoe any exophenoty
pie variations with whioh one oan easily study their adaptive
significance. The speeding up of ecrmination hL~f:'l bSE.:n
attributed to their OC(1)xrencs. In the absence of benefioial
effects, the supernumeraries can perstst in the popu1<::ltion
only by oertain acoumulation meohB.uisms ( Nur, 1963, 1969 ).
At th.e same time there are eliminb.tion mechanisms also
t:.D..rough which selection pressu.re is brought on the supor
nu..merariE;s. By the irrterHction of these two opposing foross
the supernumeraries are maintained b.t alj optir.a.wll level in
tae population.
The phenomenon of 11on-disjunctiol:! opel'ates in
the standard complement gl~o besides occurring in the
supernumeraries. Thus in c. traohl12teru§. 15.78 ;J; cells at
anaphase 11 possess variable or-.xomoBomal configurations.
In one instance the two pol(;;;s at anaphase 11 contain 8 dA
and 10 dA instead of 9 dA each ( Fig. 5), 51 ). Another
instance shows 6 dA and 8 dA configurations ( Fig. 52 ).
The latter condition has (Jome into being through the
following steps:-
step (i) ( 9 AA+ x ) at Illt';tHphHse I.
Step (ii) ( 2 AA + 1 A + x ) ; ( 1 A ) E..t anaphase I
instead of ( 9 A + x ) and ( 9 A ) , due to

-117-
non-disjunction of two bivalents.
step (iil) Splitting of ( 7A ) configuration of step (ii)
into :- ( 6 dA ) and (8 dA ) instead of
( 7 dA ) and ( 7 dA ) due to the non-disjunction
of one univalent.
The separation of chromatids has been observed
in A. crenulata in 38.8 % oells at anaphase I ( Fig. 74 ).
The separated chromatids s~ow a precocious behaviour but
do not pass to the opposite poles, till anaphase 11.
A review of abnormalities in the natural popula
tions df the selected grasshoppers makes it olear th8t
maximum abnormalities occur in c. trachypterus. The reason
may be attributed to the inability of these hoppers to
cover long distances. The females virtually do not take fli
ghts beoause of their heavy bodies and rem:7in within li.mi ted
artSs. The meles, though comparatively more active. are
not fast fliers and usually hop, rather than fly, in their
surroundings. Thus, these grasshoppers live as small
olosely breeding populb.tions more or less in se;rregation.
The aberrations, once originated, oontinue to exist in the
germ-line.
D. Effect of Drugs
The O,uanti tative study of efl'ects of caffeine
includes those freauently occurring aberrations which are
obtained when the treatment-time is kept constant and the

-118-
ooncentration of the drug is changed. The aberrations
selected for quantitative analysis, include fragmentation,
clumping and polyploidy. The data have been scored at
diplotene, diakinesis and metaphase I, and pooled and
averaged for the total number of surviving speoimen.
Out of the ten concentrations of caffeine,first
four ( ~.l % to 0.4 % ) do not cause any casualty. From
0.5 % onwards, the percentage of casualties increases.
Twenty percent specimens die after treatment with 0.5 % and 0.6 % caffeine. The peroentage of mortality with 0.7%
and 0.8 % is 60 % and 80 %, respectively. Absolute
mortality inour. with 0.9 % and 1.0 % caffeine. In the
last two ooncentrations, the mortality does not ocour
immediately but after 1-3 days of treatment. The speoimens,
therefore, do not reach the desired stage of division and
~ have to be studied before the scheduled time. Thus a r ~
systematic quantitative analysis is not possible at the
0.9 % and 1.0 % ooncentrations.
A survey of the data of fragmentation at the
different concentrations reveals that it is not easy to
establish a relationship between the percentage frequency
of aberration and the increasing concentration. There is a
grad.ual inorease in the beginning - 4.44 % with 0.1 %
caffeine and 5.82 % with 0.2 % oaffeine. This is followed
by an irregular increase - 11.66 % with 0.3 % oaffeine and
25.12 % with 0.4 % caffeine. With 0.5 % and 0.6 %, the
'peroentage rema.ins the same - 28.05 % and 28.0 %.respectively.

-119-
A further increase is noticed with 0.7 % , i.e. , 32.50 %. With 0.8 ~ oaffeine, however, only 24.44 % aberration is
obtained. In this case the mortality was 80 ;, and the data
were soored from the only surviving specimen. It can be
assumed that the peroentage frequency could have been
higher, if the dAta could be scored from all the treated
specimens. A oonfirmative evidence to this effect is
provided by the tissue of the :).9 % and 1.0 % caffeine
treated specimens, which were fixed before the scheduled
time. Here the divisional stages either do not progress
beyond early prophases, or if they progress their recogni
tion is not possible. The pachytene stages reveal heavy
fragmentation in almost all the chromosomes. The fragments
are of different sizes and stain dGrkly. Where not fragmented
the chromosomes bear 'gape' at short intervals, which are
considered the forerunners of fragmentation. In such nuclei,
the pairing is found to be normal. Thus, caffeine primarily
does not affect the behaviour of the chromosomes but shows
a negative effect on the chromatin material by inducing
gaps and fragmentation. Slizynski ( 1970 ) had obtained
similar type of fragmenta.tion at pachytene stage in the
mouse - testes cells, after usirtg saturated caffeine
solution.
The other two abnormalities - clumping and
polypl.idy, show a more defined relationship with the
increasing concentrations of caffeine. More and more cells
are viotimized as the concentration is increased. starting

-120-
from 16.44 % with 0.1 % oaffeine, the peroentage frequency
of cl~~ping inoreases gradually to 21.84 % ~ith 0.2 % ,
40.33 % with J.3 " , 49.26 % with J.4 fo , 52.1'1 % with
J.5 % , 55.11 % with 0.6 % , 69.00 % with 0.7 % , and
72.22 % with 0.8 % oaffeine. Simile.rly, the d8ta for
polyploidy are 20.0 % ( 0.1 % ), 33.0 % ( 0.2 % ), 41.66 %
( 0.3 ~ ), 52.45 % ( 0.4 % ), 58.88 % ( 0.5 % ), 60.0 % ( o. 6 % ), 14.50 % ( 0.1 % ), and 77.17 % ( 0.8 % ).
Thus, at eaoh oonoentration, the percentage frequency of
polyploidy is higher than that of clumping.
Clumping results primarily due to the aooumulation
of exoessive ohromatin on the chromosomes. The faot that
olumping may second1:1rily be induced by the olose approxima
tion of chromosomes in the highly polyploid cells , is
evident from the data of percentage frequencies of
clumping and polyploidy. Clumping inoreases in accOrdatloe
with the inorease in polyploidy and is always numerioally
lower than the latter. This is expected beoause a close
approximation of ohromosomes along with an acoumulation
of exoessive ohromatin is destined to result in heavy
clumping.
The qualitative analysis is carried out with
0.4 % caffeine, a conoentration, which besides being
effective, does not result in mortality. keeping the
concentration constant, the time is varied at the given
intervals - 4, 18, 48 and '12 hours and 6, 1, 8, 14 and 21
days post - injeotion. These timings involve all the

-121-
divisional stages of spermatogenesis. The gradual steps of
mode of action and intensity of effect of caffeine can be
studied by the present method. The earliest signs of
olumping appear at late prophase ( Fie. 94 ) when tha
chromosomes look thicker than usual.Clwnping inoreases after
metaphase 11 and gradually victimizes all the chromosomes
of a complement ( Figs. 9'7 t 98 and 99 ), so that the
individual identity of the chromosomes is lost ultimately.
The negative effect of caffeine first appea.rs
at late prophase in the form of lesions ( Fig. 95 ) and
culminates in definite breaks in the l8ter staees
( Fig. 102 ). It appears that the breakage caused by
caffeine is not reparable. Cattanach ( 1962 ) has disoussed
this possibility in the mouse chromosomes. while Kuhlman
et sI ( 1968 ) report the fftot for mammalian cells that
ohromosome - breaks cHused by caffeine hr::3.ve lost the
oapaoity to rejoin. Oocurrence of spindle - abnormality
and C- mitotic effect indicate that caffeine is similar in
its action to colchicine, in this respect.
The positive effect of caffeine depends on the
existence of extra chromosomal DNA. The latter may come
from the nuclear sap, where these is DNA in the amount
comparable to that of the chromosomes ( Izawa et aI, 1968 )
or it ma.y be a by-product or a surplus from an unscheduled
synthesis. The present investigation indicates that the
positive and negative actions of caffeine can proceEd
simul taneously in tIle same nucleus. The negative changes t

-122-
in which the chromatin extrudes from the chromosomes, may
lead to chromosome breaks by influencing certain nucleo
tides. The ohoice of the effect may depend on the state of
the nucleus and on the region81 characteristics of the
chromosomes.
The effects of the antibiotic Enteromycetin
( Chloramphenicol - Dey's ) are manifested in the chromo
somes by lesion, breakage, fre,gmentation and stickiness,
and in the spindle apparatus by polyploidy and spindle
destruotion. An analysis of the effects reveals that in its
negative action, enteromycetin involves particular sites
of the larger chromosomes, espGcially that of the sex
chromosome ( Figs. 103 and 104 ). This effect is more
specific in the first part of meiosis. During the second
part, however, the autosomes also get victimized. Here the
break is not preceded by lesion. Instead, notches appear
at the terminal end and become deeper gradually, thus,
constricting off sme!ll fragments ( Fig. 101 ). This effect
is obtained when the treatment exceeds 8 days,
The chromosomes acquire a sticky quality, most
probobly due to some physiological changes. At anaphase I,
the chromosomes orientate normally but are unable to move
towards the two poles, because their distal ends stick to
each other ( Fig. 108 ). In the longer bivalents, the
stickiness involves the sub-terminal parts as well, so tht=Jt
a peculiar tri-rediale appear~.::Jl1ce is acquired by the
chromosomes. Tetracycline has been reported to cause

-123-
simil:':ir effecte on the chromosomes of Spathosternum
pre.siniferum ( Manna and Bhunya, 1971 ) • The e.ction of
enteromycetin differs from tetracycline in showinS I:m affi
nity for the X-chromosome.
In its mild polyploidj.zing e.ction, ecteromycetin
is similar to colchicine, but the type of spindle destruc
tion obtained seems to be unique. The bifid pole possesses
the haploid set minus X, in two groups of 6 and 3 univalents
No probable reason for this behrviour could be found Oltt.
The tOxicity of this antibiotic becomes more
obvious if the treatment is continued for a long time.
when it starts fragmenting the chromosomes.
The chemical- formaline incurrs a variety of
aberrations, mostly mar.lifested in the chromosomes. The
erosion of chromatin from the nuclei is one of the IDfJ.jor
effects. At late prophase, the bivalents show a number of
regions which do not take the stain ( Fig. 110 ). Such
regions result in breakage in the same stage. There is an
indication of recov{~ry from this effect, 8S at metaphase 11
only lesions ( Figs 117 and 118 ) and no breakage are
obtained. The forerunner of lesions, the dechromatization,
brings about a chB.nge in the morphology of the chromosomes
( Fig. 116 ).
Another interesting abnormality obtained with
formaline. is the inter-bivalent connection. The latter
starts appe~::ring at diplotene ( Fig. III ), where usually
two or three bivalents get connected. Later on, more

-124-
bivalents are involved ( Figs. 114 and 115 ) forming a
long ohain. Thin, lightly staining conneotions oan be seen
at the linking points. Similar behaviour has been obtained
with oolohicine in the pro-metaphases of SEsthosternum
prasiniferum ( Manna and Parida, 1965 ). Suoh oonneotions
seem to be of temparary nature beoause later stages are
devoid of them.
Though olumping is obtained with formaline at
late prophase, it is not very signifioant. The bivalents
appear thiokened due to the acoumulation of excessive
DNA, and the sex ohromosome loses its heteropyonotio
behaviour and stains as intensely as the autosomes ( Fig.
113 ). The oel1a reoover from this positive effect of
formaline in the later stages which do not show olumping.
Formaline sholf,s a definite effini ty for metaphase
11. Moreover, this investigation oonfirms the earlier
report that this ohemical affeots the sex ohromosome speoi
fioal1y ( Manna, 1969 ). The oentromere of the sex
ohromosome is greatly stretched at metaphase 11 ( Figs.
119 and 120 ). The ohromosomes appear very thin and
stain lightly due to the erosion of chromatin. Aohromatio
gaps ooour in all the ohromosomes. The telomerio ends are ,
however, the darkest part of ohromosomes, indicating that
dechromatization commences from the proximal end and
proceeds towards the distal end. Thus, nost of the effeots
of forma1ine are of the negative type, originating due to
the produotion of antimetabolites.

-125-
In one instaLce, fo~aline is obsGrved to effeot
the spindle also resulting in 'star'- oOLfigurations
( Fig. 121 ). Suoh aberrations are oharacteristic of the
alkaloid oolchicine ( Manna and Parida, 1965 ).
A preliminary observation on the effect of
nicotine reveals that it is the most potent of the chemicals
used in this investigation. The effects Bre mostly non
delayed. Niootine enflicts heavy frac;mentation at gonial
metaphsses ( Fig. 122 ). In the absence of exchange - type
configurations and ring-like structures, it can be stated
that the broken ends I::!.re uns.ble to reunite. The sex
chromosome remains unaffeoted. Some other abnormali~ties of
lesser intensity occur at diplotene ( Fig. 123 ), metaphase
I ( Fig. 125 ) and metaphase I1 ( Fig. 126 ). TheSe include
the positive effect ( clumping) a."ld the negative effect
( lesions, breaksge ).
The effects of the four chemicals used in this
investigation provide sufficient evidence to conclude that
ttey affect synthetic activities at one stage or the other.
Th.e binding of the chemioals with DNA in the ohromosollles is
expeoted to bring about structural alterations, whioh may
result in the disruption of chromatin material. In all the
instances of break;3ge. reunion is absent. This interpretation
suggests that the break£::ge. and the subsequent failure of
repair mechanism, both result from a block in some essential
synthetic activity, probably both of protein and DNA.

I VII I ~~

-126-
StThfEARY
A. Caryo1ogica1 study :
This study includes the mitosis and meiosis of
five species of grosshoppers belonging to the family
Aorididae ( Order : Orthoptera ) t namely, Tri10phidia
annulata ( sub-fsmily : Oedipodinae ), Ch~otogonus
trachypterus ( sub-family: Pyrgomorphinae ), Leva indica
( sub-family : Gomphooerinae ) t ~ttractomor:eh~ crenulata
( sub-family • Pyrgomorphina.e ) and Poekilocerus pictus • -( sub-family • Pyrgomorphinse ) . •
The diploid number of chrom.osomes in the male
germ cells of T. annu1ata is twenty-three. The chromosomes
can be classified into three long, seven medium and two
short chromosomes. The sex chromosome is the third in order
of length. Meiosis follows the classical pattern. Paohytene
ohromosomes possess 'snai1-horn'-like centric chroiIlomeres,
which give the impression of second ar:ns. The later stages
of meiosis do not show any indication of presence of
second ar'-IlS. ~I!ean chiasma fre::uency per nucleus at
diplotene, diakinesis &.nd metaphase I is 16.00 ! 0.02,
11.46 ! 1.24 and 11.35 ! 1.26, respectively. The
• pseudo-chiasmfita' occur at eHrly anaphase I. During
mi tosis the single 'X' chromosome reml.::.ins isopycnotic at
prophase. The longer Butosomes indicate the presence of
small arms in the somatio tissue ( gastric caeca ). A
comparison of lengths of germina1- and somatic-

-12'1-
ohromosomes makes it olea.r tl'.lEit the lu.tter are about one
micron longer tllan the former at metaphase.
The diploid number of chron:,08omes in the male
germ cells of Chroto$onus trachlP~s is nineteen. The
chromosomes can be classified into three long, six medium
and one short chromosomes. The meiosis of this species
has been worked out in the populations of Delhi state
and Ottu ( Haryana state ) of India. The present accowlt
includes the morphometric data a!jd a few points of interest
in the meiosis of the local ( Bhopal ) populgtion of M.P.
state. The results are compared and contrasted with
those of the other two reported populations. Mean chiasma
frequency per nucleus in the Delhi-, Ottu- and Bhopal
- populations, respectively, is , 15.25 ! 0.'12 / 15.86 /
15.50 + 1.27 at diplotene, 12.'15 ! a.dl / 11.93 / -11.23 + 1.04 at diakinesis, and 10.95 ! 0.94 / 11.20 / -10.66 + 0.25 at metaphase I. No second a.rms are visible -e.t a.ny stage. The pachytene nuclei possess t snail-horn'
like centric chromomeres. During mitosis. the prophases
do not show any heteropycnotic body in tbe m:::lle somatio
tissue. The sex chromosome can be distinguished only on
the basis of its length. Second arms ~,re not visible in
any chromosome • A comparison of the lengths of germinal
and somatic- metaphase chromosomes reveals that the latter
are two times longer than the former.
The diploid number of chromosomes in the male
germ cells of L. indica is twenty-three. The chromosomes

-128-
oan be olassified into three long, six medium and three
short ohromosomes. The sex ohromosome is the longest.
The general oourse of meiosis is of the classical type.
Mean ohiasma frequency per nucleus at diplotene, diakinesis
and metaphase I is 13.6 ! 1.49, 12.JO ! 1.12 and 9.71 ! 1.11
No seoond arms are visible at any stage. During mitosis,
the prophases of female somatio tissue exhibit an indis
tinot heteropyonotio mass. The second arms are not visible.
A oomparison of the lengths of germinal- and somatio
metaphase ohromosomes indioates that the ooiling is much
more loose in the latter.
The diploid number of ohromosomes in the m:~le
germ oells of A. crenul:;;ta is nineteen. The chromosomes
oan be olassified into three long, six medium and one
short ohromosomes. The sox ohromosome is the longest.
Meiosis has been reported elsewhere. During mitosis, the
sex ohromosome remains isopycnotic in the m81e somatio
tis3ue. The length of the somatic chromosomes at metapha.se
is almost double that of the spermatogon:i.al metaphase
ohromosomes.
The diploid number of chromosomes in the mole
germ gells of P. pictus is nineteen. The chromosomes can
be classified into three long, six medium ruid one short
chromosomes. Meiosis hss bean reported elsewhere. Mitosis
reveals the occurrence of second arms in the smallest
and one of the medium autoSOrlLes. The length of the second
arm varies from 0.9 to 1 micron. Different views are

-129-
available re)~aI'ding the po si tion of the centromere. There
is appreciable difference in the lengths of metaphase
chromosomes from the germinc:;l ;:;!1d somCltic tissues.
B. Abnormalities in the Natural ~~E~ations :
The abnormalities are scored inthe natural
populations of T. annUlate. t L. indlca, C. trachypterus
and A. crenulata a,mong one hundred - aud - six, fifty-two
eighty - six I:-illd thirty - five individuals, respectively.
T. armulate. cor.taiua extra fragment polyploid
cells, anaphase bridge [md breakage in its population.
~. indica contai!ls extra fragment, polyploid cells,
abnormal spindle and breakege. c. trachypterus contains
supernumerary chromosomes, polyploid oells, non-dif3junction,
breakage and extra fragment. Lastly, A. eranulata contains
breake:-lge and chromatid separation.
The percentage frequency of extra fragment is
nQ,t very significant in the species. It h&s not been found
to lag behind and interfere in spindle elongation.
Polyploidy is at the level of tetraploidy and originates
most probably due to failure of cleavage during mitosis.
No multivalent confi.gurations bre found and the sex
chromosomes do not show any tendency to synapse in the
tetraploid cells. The occurrence of brakage is significant
/'~ in L. indica only ( 29.41 'to ) • Here the breaks are Observed {
in the gonial metaphases affectinG both the chromatids of

-130-
the sex chromosome and the largest 8utosO,IJe. The
I oompleteness' of the break,:ige is doubtful in all the
instances. The dioentrio bridge has been observed in a
single oell in T. annulata.. A fragment of considers.hle
size aocompanies the bridge. Abnormal spindles with one
bifurcated pole hs.ve been found in L. indioa, the
peroentage frequenoy being 33.33 % • The origin of this
abnormatity is uncertain. The supernwnerary ohromosomes
occur only in C. trachlEter~s. Their number varies from
one to six in the oomplement. The behaviour is compared
and oontrasted with that of the supernumeraries of the
Ottu ( Haryana state ) - popul::'tion. Non-disjunotion
introduoes variable oonfigurations in the meiotio oells
of c. trachyPterus. The chromatids of the smallest
autosomal univalent divide preoociously in A. orenulata.
c. Banding Pattern of P. pi~ :
The nature and behc"Jviour of the ce tromere bi.:lS
been traoed in all the stages of spermatogenes~s with the
he~p of Giemsa staining. The o'bservations not only differ
in many respeots from the routine analysis of caryot;lpe,
but also oontradiot a few pOints. All the ohromosomes
possess well - marked C-bands, comprising the centric
cnromomeres and proximal band of heterochromatin of the
chromosoale arms" The sex chromosome possesses telomeric
bands 8S well. The smallest autosome has two bcll1ds in tbe

-131-
distal part. The C- band consists of four sub-units. The
centrio ohromomeres appear like 'snail - horns' at
paohytene. The sex chromosome, though showing hetaropyono
sia after general staining, stains as intensely as the
auto8omes with Giemsa. The prooedure is, therefore,
reliable for the looation of constitutive heterochromatin.
D. Effects of Dru~s :
The investigation includes the q,uantitative
analysis of the effects of caffeine and qualitative analyses
of the effeots of oaffeine, enteromycetin, formaline and
niootine on the germ oells of C. trachlE!~. The division
oyole has been consulted for caloulating and adjusting the
treatment - time for the analyses.
Ten conoentrations of oaffeine have been tested
quantitatively, keeping the treatment - time fixed.
Mortality ocours in the higher dosage range ( 0.5 % to 1.0 %)
The frequenoy of three types of abnormality has been soored
- fragmentation, clumping and polyploidy. Data are oollected
from diplotene, diakinesis and metaphase I stages, and
pooled and averaged acoording to the number of survived
speoimens, for each concentration of caffeine. The
frequency of olumping and polyploidy show a definite rela
tionship with eaoh other end with the inorease in oonoen
tration. No simple relationship oan be drawn, however, from
the frequency - study of fragmentation.

-132-
For qualitative analyses of the four drugs,
following ooncentrations have been selected - J.4 % oaffeine, 0.25 ~ enteromyoetin, 3.0 % formaline and 0.5 % niootine. The testes were fixed at the intervals of 4, 18,
48 and 72 hours and 6, 7, 8, 14 and 21 days post - injeotion.
The abnormalities have been oategorised into (i) mitotio
ohanges , (ii) changes in the meiotio prophase stages,
(iii) ohanges at metaphase I and II, and (iv) ohanges at
first and seoond post - metaphase stages.
Niootine has been found to be the most potent
drug of all. Probable modes of aotion of the various drugs
have been disoussed.
=*=* =* =: *= iI::: *= -lI::*= *=*=

VIII
BIBLIOGRAPHY

-133-
BIBLIOGRAPHY
As ana , J.J. 1934. "studies on the chromosomes of
Indian Orthoptera VI. The idiochromosomes of
Hierodul~ sp." CURR. SCI. 2 : 244 - 245.
2. Asana, J.J. and S. Makino. 1934. JOUR. UNIV. BOMBAY,
BlOt. SC1. 2 : 50.
3. As ana , J.J., S. Makino and H. Niyama. 1938. " A
chromosomal survey of some Indian inseots, I n.
J. FAC. HOKKAlDO UNIV. 6 : 211 - 234.
4. Asana, J.J., S. Makino and H. Niyama. 1939. tt A
chromosomal survey of some Indian inseots, II.
A comparative study of the chromosomes in six
species of Aorididae". JAP. J. GENET. 15 : 251-260
'5. Atoh1ey, W.R. 1974 ... Morphometrio differentaition in
chromosoma11y oharaoterised parapatrio races
of morabine grasshoppers ( Orthoptera :
Eumastaoidae ).11 AUST. J. ZOOL. 22(1) : 25 - 37. ".-"
/"6. Atkins, L., P.D. Taft and K.F. Da1a1. 1962.
ff Asynohronous DNA synthesis of sex ohromatin
in human interphase nuo1ei ". J. CELL. BIOL.
15 : 390 - 393.
7. Axe1rod, Ju1ius and Jules Reiohentha1. 1952. " The
fate of caffeine in man and a method for its
estimation in bio1ogioa1 material n. J. PHARM.
EXP. THERAPY. 1Q7 : 519.

-134-
8. Barkar, John F. 1966. " Climato10gical distribution
of a grasshopper ( Ml!:!!eleotettix maoulatus )
supernumerary ohromosome." EVOLUTION.
20(4) s 665 - 667.
9. Bhattaoharya, A.K. 1913. "The effects of semioarbazide
on spermatooyte ohromosomes of a grasshopper".
PROOf OF 60TH SESSION OF ISOA.
10. Bhattaoharya, A.K. and G. Dab. 1974. " Effects of arginine
on the spermatooyte ohromosomes of a grassho
pper". PROOf OF 61 ST SESSION OF ISCA.
11. Bhunya, S.P., B.B. Parida and S.N. Ghosh. 1974.
" Erythromyoin induoed spermatooyte
chromosome aberration in short - horned
grasshoppers ". PROOf OF 61 BT SESSION OF
ISCA.
12. Bhunya, S.P. and D.N. Aoharya. 1973. " Spermatooyte
chromosome aberrations induced by calcium
ohloride in a sandhopper qhrotogonus
traohypterus." PROC. OF 60TH SESSION OF ISCA.
13. Bhunya, S.P. and G.K. Manna. 1911. " Meiotio chromosomal
aberrations in a short - horned grasshopper
induoed by an antibiotio Griseofulvin ".
PROOf OF 58TH INDIAN SCIENOE CONGRESS.
14. Bhunya, S.P. and Nirupama Das. 1973. " Atropine induced
spermatocyte chromosome aberrations in a
grasshopper Poeci1ocera Eicta. "
PROOf OF 60TH INDIAN SCIENCE CONGRESS.

f'
-135-
15. Blakeslee, A.F. and A.G. Aver"!. 1937. If Methods of
induoing doubling of ohromosomes in plants".
J. HERED. 28 I 393 - 411.
16. Bre gman , A.A. 1973. n Lateral aub-units in C-bands of
unrep1ioated ohromosomes of Dissosteira ,.
oarolina ( L. ) ( Orthoptera : Acrididae ) ".
CAN. J. GENET. CYTOL. 15 (4) : 757 - 761.
Brown, !ndrea and P.J. Wilmore. 1974. n Looation .~ .. /
/ \,. of repeti tious DNA in the ohromosomes of the
181 Cardoso.
desert looust ( Sohistooeroa gregaria ) ".
CHROMOSOMA ( BERL. ) 47 (4) : 379 - 383.
H., F.A. Saez and N. Brumzorri11a. 1974.
" Looation, structure and behaviour of
C-heterochromatin during meiosis in
Diohroplus silveira~idoi ( Aorididae :
Orthoptera ) n. CHROMOSOMA ( BERL. )
48 (1) , 51 - 64.
i/ 19. Carlson, J.G. and M.E. Gaulden. " Gra.sshopper Neuro
blast Techniques ". In I " Methods in cell
Physiology tI, edited by D.M. Pressoott.
Vol. I. Aoademio Press. 1964.
20. Carothers, E.E. 1911. " The segregation and recombination
21. Oarroll,
of homologous ohromosomes as found in two
genera of Acrididae ( Orthoptera )n.
J. MORPH. 28 : 445 - 520.
M. 1920. If An extra dyad and an extra tetrad
in the spermatogenesis of Camnula pelluoida

-136-
( Orthoptera ) : Numerical variation in the
ohromosone oomplex within the individual ".
J. MORPH. 34 : 375 - 455.
Caspersson, T. and L. Zeoh. 1973 " Chromosome Identifi
oation : Teohnique and Application in
Biology and Medicine." ~
Caspersson, T., S. Farber, G.E. Po1ey, J. Kudynowski,
E. J. Modest, E. Simonsson, U. Wagh and
L. Zeoh. 1968. EXPTL. CELL RES. 49 : 219.
,~- 24. Cattanach, B.M. 1962. " Genetic effects of caffeine in
mice ". ZEITSCHRIFT FUR VEREBUNGS-LEHRE.
'./
25.
93 I 215 - 219.
Chatterjee, A.X. 1971. " Some oytological information
about two specie. of grasshoppers ( Orthoptera
: Aorididae ).n PROe. OF 58TH ,INDIAN SCIENCE
CONGRESS.
/'·'26. Chatterjee, K., A. Majh1 and S.K. Barik. 1971. " A study
27. Clark,
28. Coleman,
of ohromosomes during meiosis in four
species of Indian grasshoppers. " CARYOLOGIA.
24 (4) : 447 - 455.
A. M. and E. G. Clerk 1968. It The genetio effeots
of oaffeine in Drosophila melanogaster "
MUTATION RESEARCH. 6 : 227 - 234.
L.C. 1943. If Chromosome struoture in the
Aorididae with speoial referenoe to the X
ohromosome ". GENETICS 28 : 2 - 8.
'* 1',(,0 ceeding s

-137-
29. Crozier, R.H. 1968. " An acetic acid dissociation,
air - drying technique for il1sect
chromosomes with ace to - lactic orcein
, 30. Crozier,
staining. " STAH; TECH1~OL : 43 (3) : 171 - 173
R.H. 1970. " Karyotype of twenty-one ant
species ( Hymenoptera: Formicidae ), with
reviews of the known ant karyotypes. It
CANAD. JOUR. GENET. CYTOL. Vol. XII (1) :
109 - 128.
31. Dar1ington, C.D. 1936. " Crossing over and its mechanioal
relationship in Chorthippus and Stauroderus. "
J. GEN£T. 33 z 465 - 500
Dar1ington, C.D. 1956. It Chromosome Botany .. tt AlIen
and Unwin, London ..
., .. , 33. Darlington, C. D. and J. MoLeish. 1951. It Action of
maleic hydrazide on the cell .. tI NATURE.
167 : 407 - 408.
34. Dar1ington, C.D. and L.F. La Cour. 1950. It Hybridity
selection in Campanula.. tI HEREDITY.
"-'~ -'
\//'35.
36.
4 I 211 - 248.
Darlington, C.D. and L.F. La Cour. 1960. n The Handling
of Chromosomes. " 3rd Ed. George, AlIen and
Unwin Ltd.
Dar1ington, C.D. and M.B. Upoott. 1941. J. GENET.
41 : 291.
31. Darlington. C.D. and P.C. Kol1er 1947. If The chemical
breaknge of chromosomes. 11 HEREDITY. 1 : 187-221

-138-
// /<' 38. Dave, Iv:.J. 1965. n On unusual sex chromosomes found in
{
two species of the Looustidae. " OYTOLOGIA
30 (2) : 194 - 200.
/39. Desa!, V.B., M. Sirsi, M. Shankarappa and A.R. Kasturibai.
1966. " Studies on the tOxicity of
Abrus precatorius Linn. I. Effect of aqueous
ext~aots of seeds on mitosis and meiosis in
grasshopper, Poeci10oera Eicta. " INDIAN
JOUR. EIP. BlOL. 4 (3) : 164 - 166.
40. Dutt, M.IC. 1948 a • " A oytologica1 investigation on the
genus Phloeba ( Aorididae )." PROe. NAT.
INST. SOl. INDIA. 14 s 13 - 14.
41. Dutt, M.K. 1948b. It Meiosis and chiasma formation in
Tristia. pulvinata Uvarov. 11 PHOO. ZOOL. SOO.
BE.NG. 1 : 49 - 57.
42. Dutt, M.K. 1950. n studies on ohromosome behaviour and
sex ohromosome in Ohrotogonus sp. f1
PROe. ZOOL. SOO. BENG. 8 : III - 129.
43. Dutt, M.K. 1955. PROO. ZOOL. SOC. BENGAL. 8 : 111.
44. Dutta Gupta, A.K. and V.K. D1vakaran. 1959. " Response
of some grasshoppers ( Aorididae ) to
pyrethrum. n SCI. & CULT. , CALCUTTA.
25 : 273 - 274.
45. .1'Vans, W.L. 1954. AMER. NATURALIST. 88 : 21.
46. Fox, D.F. 1969. 11 DNA values in somatic tissues of
Dermestes ( Dermestidae : Coleoptera ) I.

-139-
Abdominal fat body and testis wall of the adult.,
CHROMOSOMA ( DERL ). 28 : 445 - 456.
47. Fox, V.P. 1970. " DNA values in somatic tissues of
Dermestes ( Dermestidae : Coleoptera ) 11.
Malpighian tubules of the adult male. n
CHROMOSOMA ( BERL. ) : 31 : 321 -330.
Gall, J.G., E.H. Cohen and M.L. Polan. 1971. n Repetitive
DNA sequences in Drosophila. It CHROMOSOMA
( BERL. ) 33 : 319 - 344.
'49. Gallaghar, A., G. Hewi tt and 1. Gibson. 1973. 11
» Differential Giemaa staining of hetero
chromatic B- chromosomes in Myrmeleotettix
maculatus ( Thunb. ) (Orthoptera : Acrididae ).
CHROMOSOY~ ( BERL. ) 40 : 161 - 112.
50. Gaulden, M.E:. and J.G. Carlson. 1951. 11 Cytological
effects of colchioine on grasahopper neurobla
sts in vitro with special reference to the
origin of spindle." EXP. CELL RES.
2 : 416 - 433.
51. Goldstein. A. and R. Warren. 1961. n Passage of caffeine
into human gonadal and fetal tissue. "
SHORT COr~dUNIC.ATIONS. 166 - 168.
52. Grouchy, J. de, C Turleau, M. Roubin and F. Chavin
Colin.. It Chrodlosoillal evolution of man and
primates ( Pan tro~lodytes, Gorilla !Orilla,
Pango pygmaeus ) " NOBEL. 23 ( 1913 ).
124 - 131. CHHOfJJOSOlv:E IDENTIFICATION.

-140-
/' 53. Gururaj, M.E. and M.R. Raje,sekarasetty. 1911.
,
It Oocurrenoe of B- ohromosomes, behaviour
and their influenoe on the ohiasma. frequency
in !Qt.:0tl1us.~berti~ ( Acridoidea I
Orthoptera )." PROC. OF 58TH INDIAN
SCIENCE CONGRESS.
54. Ha1dane, J.B.S. 1956. " The deteotion of autosomal
letha1s in mice induced by mutagenio agents."
J. GENET. 54 : 327 - 342.
55. Hareyama, S. 1941. " Studies on the chromosomes in
Locustidae." J. SOl. HIROSHIMA m~IV. B.
9 : 1 - 157.
1/ 56. Hennig, W., I. Hennig and H. Stein. 1970. " Repeated
sequences in the DNA of Droso~h~ and
their localisation in giant chromosomes."
CHROMOSOMA ( BERL. ) : 32 : 31 - 63.
Hewi tt, G.M. 1964. " Popule.tion oytology of British
grasshoppers I. Chiasma variation in
Chorthippus brunneus, C. parallelus and
Omocestus viridu1us. " ClrnOMOSOMA.
15 (2) I 212 - 230.
58. Hewltt, G.M. 1965 ... Population cytology of British
grasshoppers 11. Annual variation in
ohiasma frequeuoy. I, CHROMOSOMA.
16 (5) : 579 - 600.
Hewi tt, G.fyl. and B. John. 1967. It The B- chromosome
system of Mlrmeleotettix maou1atus (Thunb.) Ill.

-141-
The Statistics." CIlROMOSOMA. 21 (2) : 140 - 162 • .,.
i 60. Hewitt, G.M. and B. John. 1968. " Parallel polymorphism
for supernumerary segments in Chorthippus
parallelus ( Zetterstedt ) : I. British
populations." CHROMOSOMA. 25(3) : 319 - 342.
61. Hewitt, G.M. and B.John. 1972. » Inter - population sex
chromosome polymorphism in the grasshopper
Podisma pedestris. " CHROMOSOMA.
. /'
./ 62.
37 (I) : 23 - 42 •
Hau t T.C. 1972. Teohnioal Notes: " The Bandwagon."
In I MAMMALIAN CfmOMOSOME NEWS LETTER.
Vol. 13. No. 1. January.
Hsu, T.C. and K. Benirsohke. 1967. " An Atlas of
Mammalian ohromosomes." Vol. 11 SPRINGER -
VERLAG. N.Y. Ino.
(./ 64. Hau, T.C. and K. Benirsohke. 1968. " An Atl&s of
Mammalian Chromosomes. It Vol. 11 SPRINGER -
VERLAG. N.!. Inc. \
v 65. Hughes - Sohrader, S. 194"7. " The t premetaphase stretoh'
and kinetochore orientation in phasmids. 1t
/' CHROMOSOMA ( BERL. ) : 3 : 1 - 21. ,,/
/'
y' 66. Hughes - Sohrader, s. 1948. " AdVances in Genetios."
2 s 127 - 203. A.P.N.Y.
//67. Hug-hes - Schrader, S. 1950. n The ohromosomes of
mantids ( Orthoptera : Manteidae ) in
relation to taxonomy. " CHROMOSOMA.
4 : 1 - 55.

-142-
68. Htlghes - Sohrader, S. 1953. If Stlpplementary notes on
the oytotaxonomy of mantids ( Orthoptera :
Manteidae ). It CHRm~OSOMA. 6 : 79 - 90.
1~'69. Izawa, M., V.G. Alfrey and A.E. Mirsky. 1963 •
.. Composition of the nuoleus and the
ohromosomes in the lamp brush stage of the
newt oooyte." PROC. NAT. ACAD. SCI. 50 : 811 •
. / 70. Jaln, H.K. and Umrao Singh. 1967. It Aotinomyoin - D
71.
induoed ohromosome breakage and suppression
of meiosis in the looust SohistoQer~
gregaria .. Division of Genetios I.A.R.I. - .. -New Delhi. Dec. 12, 1966/ Feb. 27, 1967.
John, B. 1973. " The oytogenetio systems of grasshoppers
and loousts. 11. The origin and evolution of
supernumerary segments. It CHROMOSOMA ( BERL )
44 (2) : 123 - 146.
72. John, B. and G.w.. Hewltt. 1963 ... A spontaneous
interohange in Chorthippus brunneus with
extensive ohiasma formation in an interstitial
segment. It ~rlROMOSOMA ( BERL. )
14 : 638 - 650.
// 73. John, B. and G.M. Hewitt. 1965a. It The B- ohromosome
system of !lsmeleotettix maoulatus ( Thunb. )
I. The mechanios. .. CHRO~OSOMA.
16 (5) : 548 - 578.
74. John, B. and G.M. Hewitt 1965b. It The B-ohromosome
system of Myrmelcotettlx maoulatus (Thunb.) 11.
The statistios. If CHROMOSOMA.
17 (2) : 121 - 138.

-143- ,
75. John, B. and G.M. Hewitt. 1966. It Karyotype stability
and DNA variability in the Aorididae. tt
CHROMOSOMA ( BEEL.) 20: 155 - 172.
76. John, B. and G.M. Hew1tt. 1968 • ., Patterns and pathways
of ohromosome evolution within the
Orthoptera. It CHROMOSOM.A. 25 (1) z 4J -74.
77. John, B. and G.M. Hewitt. 1970 ... Inter - population
sex ohromosome polymorphism in the
grasshopper Podisma pedestris I. Fundamental
faots. " CHROMOSOMA. 31 (3) : 291 - 308.
John, B. and K.H. Lawis. 1965. ., Genetio speoiation in
the grasshopper Eu~re~oon~~is.plorans • .,
CHROMOSOMA 16 (3) : 308 - 344. >,,-
.,,' 79. John, B., K. R. Lewis and S. A. Henderson. 1960.
CHROMOSOMA. 11 : 1.
80. John, B. and S.A. Henderson. 1962. If Asynapsis and
polyploidy in Sohistooeroa p6ranensis. .,
CHROMOSOMA ( BERL. ) 13 I III - 147.
Jones, K. and C. Colden. 1968. .. The telooentrio
oomplement of !:r.odescantia micr~ntha.. "
CHROMOSOMA ( BERL. ) 24 : 135 - 157.
/'82. Kayano, H. and K. Nakamura. 1960. It Chiasma studies in
structural hybrids V. Heterozygotes for a
centric fusion and for translocl;;l,tion in
Acrida lata. " CYTOLOGIA ( TOK~O ).
25 : 476 - 480.
83. Kihlman, B. and A. Levan 1948. It The oytological

i
-144-
effeots of caffeine. « ABSTRACTS KURZE
MITTEILUNGEN. 109 - 111.
King, R.L. 1950. " Neo - Y chromosomes in HlPoohlora
~ and Mermiria intertexta ( Orthoptera :
Aoridoidea ). ft J. MORPH. 8'1 I 227 - 238.
85. King, R.L. and H.W. Beams. 1938. " The multiple
ohromosomes of ParatylotroEidia brunneri
Soudder ( Orthoptera : Acrididae ). "
J. MORPH. 63 : 289 - 299.
86. Klassen, W., T.R. Chang and P.E. Eide. 1969. tf Effects
of aphola,te on ohromosomes of germ cella in
the grasshopper testes. 11 CANAD. JOUR.
GENET. CYTOL. 11 : 829 - 833.
87. Klesterskat It A.T. Natarajan and C. Ramel. 1974.
" Heterochromatin distribution and ohiasma
looalisation in the grasshopper Brlodema
tuberoulata ( Fabr. ) (Aorididae ) ...
CHRO)10S0MA ( BERL. ) 44 : 393 - 4:)4.
d8. Kuhlman, W., li.G. Fromme, E.M. Heege and W. Ostertag.
1968. " The mutagenio action of caffeine in
higher orga:rlisms. ,. CANCER RESEARCH
28 : 2375 - 2389.
89. La Oourt L.F. and A. Rutishauser. 1954 ... X- ray
breakage experiments with endosperm. I.
sub-chromatid breakHge. n CHROMOSOMA.
6 : 696 -709.
90. Levant A., K. Fredga and A.V. Sandberg. 1964.

-145-
n Nomeno1ature for oentromerio position on
ohromosomes. .. HEREDITAS ( LUND )
52 I 201 - 220.
//<~"91. Ltawis, K.R. and B. John. 1959. n Breakdown and
restoration of ohromosome stability
following inbreeding in a looust. tI
CHROMOSOMA ( BERL. ) 10 : 589 - 618.
Lewis, K.R. and B. John. 1963 ... Chromosome Marker. ft
Churchill. London.
93. Lewis, K.R. and B. John. 1966. " The meiotic oonsequen-
ces of spontaneous ohromosome breakage. "
CHROMOSOMA 18 (2) : 287 - 304. I' V 94. Lewitsky, G.A. 1931. " The morphology of the ohromosomes"
/'
BULL. APPL. BOT. 27 I 19 - 173.
95. Lewontin, R.C. and M.J.D. White. 1960 ... Interaction
between inversion polymorphisms of two
ohromosome pairs in~e grasshopper Morabe
aourI's. " EVOLUTION ( Lanoaster, Pa. )
14 : 116 - 129.
96. Lima - de - Faria. A. 1956. " The role of the kinetoohore
in chromosome organization. If HEREDITAS
( LUND ) • 42 : 35 - 160.
~ 97. Lyon, M.F. 1961. " Gene action in the X- ohromosome of
the mouse ( Mus musculus L. ). " NATURE
/'/
/98.
190 : 372 - 373.
Lyon, M.F. 1962. " Sex ohromatin and gene aotion 1n the
mammalian X - chromosome. " AM. J. HUM .•

-146-
GE~ET. 14 : 135 - 148.
Makino, s. 1956. It A Review of ohromosome Numbers in
Animals. " Revised Edition, Hokuryu Kan.
Tokyo.
100. Makino, S. and E. Momma. 1950. " Observations on the
struoture of grasshopper ohromosomes
subjeoted to a new C!oetooarmine treatment. ,.
J. MORPH. 86 : 229 - 252.
'/' 101. Mangalangi, N.K. and !vl.K. Subramaniam. 1963. " A
haematoxylin squash technique for the study
of grasshopper ohromosomes. Jt CURi~.K~T
SCIEI\CE. 32 : 364 - 366. August.
102. Manna, G.K. 1954. " A study of ohromosomee during
meioLls in fifteen speoies of Indian
/' grasshoppers. " PROC. ZOOL. SOC. 7 : 39 - 58. //
V lQ3. Manna, G.K. 1967a. " Cytologioal analysis of the sex -----
ohromosomes from the testes oells of
grasshoppers: A review .ft NUCLEUS INST. J.
/' CYTOL. ALLIED TOP. 10 (l) : 64 - 80.
\ / ",/ 104. Manna, G.K. 1969. " Some Aspeots of ohromo:<ome
Cytology. " Presidential Address I 56 th
Indian Soienoe Congress, Bombay ( Powsi ).
105. Manns, G.K. and B.E. Parida. 1965a. " Differen~_lly
administered oolohioine effects upon the
testes oells of the grasshopper, Sp3.thosternum
/' prasiniferum.. 11 CYTOLOGIA. 30 : 392 .:. 401.
/'
106. Manna, G.K. and B.E. Parida. 1965b. tI Aluminium chloride

-141-
induoed meiotio ohromosome aberrations in the
grasshopper Ph10eba antennata • It
NATURWISJEN. 52 ( 23 ) : 641 - 648. ~ . .-' .-'"
.. 101. Manna, G. K. and B. B. Parida. 1967. tt Forma1ine induoed
sex ohromosome breakage in the spermatooyte
oe11s of the grasshopper Tristie pulvinata. 11
JOUR. CYTOL. & GENET. 2(1) : 86 - 91.
108. Manna, G.K. and B.B. Parida. 1968~ " Effeots of RUDR
( 5 bromodeoxyuridine ) on the spermatooyte
ohromosomes of the grasshopper Phloeba
antennata. " I\UCLEUS ( CALCUT'l'A )
11 (1) : 50 - 52.
>/' 109. Manna, G.K. and B.B. Parida. 1910. It Differentil:.il ma1eio
hydrazide ( MH ) sensitivity shown by the
spermatocyte chromosome in five speoies of
grasshoppers. It CYTOLOGIA. 35.
110. Manna, G.K. and B.B. Parida. 1911 ... Grasshopper spermato
oyte ohromo~ome aberrations induced by
phenols. I' PROC. I ST ALL INDIA CONGR. CY"TOL.
&: GENET. SYMPOSlur,f. CHANDIGARH.
111. Manna, G.K. and D. Roy. 1964. rr Grasshopper spermatooyte
ohromosomal ohanees induced by pH -
a1 trations. " DIB NATUHWIS;iEh SCHAFTEN.
51 (20) : 493 - 494.
112. Manna, G.K. and K. Chatterjee. 1963. " Polymorphio sex
ohromosomes in Euprepoonemis sp. I. The
meiosis in the XO type male and in the neo - X

-148-
and neo - Y type male • It THE NUCLEUS.
6 (2) : 121 - 1341
113. Manna, G.K. and M. Lahiri. 1966. 11 Effects of cupferron
on the spermatocyte or~omosomes of three s
speoies of grasshoppers. " JOUR. CYTOL. :'e
GE~ET. 1 (l) : 15 - 36.
114. Manna, G.K. and P.K. Mukherjee. 1966. a Spermatooyte
ohromosome aberrations in two species of
grasshoppers at two different ionic activities. It
~UCLEUS 9 (2) : 119 - 131 •
.,/ 115. Manna, G.K. and S.C. M.azumder. 1964. It Ethyl alcohol
induoed sex chromosome breakage in the
grasshopper Phloeba antennata. 11
~,.-, <
\ /'" NATURWISSLN. 51 (24) : 646.
~ 116. Manna, G.K. and S.C. Mazumder. 1961a. 11 Evolution of ! i
"', karyotype in an interesting species of
grasshopper, Tristia pulvinate Uvarov."
CYTOLOGIA. 32 (2) : 236 - 241.
\....-/ 111. Manna, G.K. and S. P. Bhunya. 1911. It Tetracycline induced
/ /
/
spermatocyte chromosomal aberrations in the
grasshopper S12athosternwp. p~asiniferu.m.fI
/ J. SOC. CYTOL. t GENET.
/ 118. Manna, G.K. and S.P. Bhunya. 19'72. " Insulin induced
grasshopper s~rmatocyte chromosome
aberration. " PROC. r~AT. ACAD. SeI. 42 (E). ,/'
/ / 119. Marks, G.E. 1957. " The cytology of Oxalis dispa!:
( Brown). " CHROlVlOSOMA ( BERL ) 8 : 650-610.

-149-
v,." 120. McClung, C.E. 1902. " The accessory ohromosome - sex - ~.----.--
determinant ? " BIOL. BULL. 3 : 43 - 84.
yf'"' 121. MoClung, C.E. 1902 ... The spermatooyte divisions of the
Looustidae. " KANSAS. U~IV. SC1. BULL.
1 : 125 - 131. ,.,...-" ... .,. .
. ,/ 122. MoC1ung, C.E. 1905. Jt The ohromosome oomplex of
Orthopteran spermatooytes. " BIOL. BULL.
WOODS HOLE. 9 I 304 - 340.
123. McC1ung, C.E. 1914. 11 A comparative study of the
ohromosomes in Orthopteran spermatooyte. et
J. MORPH. 25 I 65 - 749. "'''/'
//'124. Mesa, A. and R. S. de Mesa. 1967. 11 complex sex -
determining mechanism in three species of
Sout}! American grasshoppers ( Orthoptera I
ACl'idoidea ). " CHROMOSOMA.
21 (2) I 163 - 180. '\ ./
'/ 125. Mi ttwoch, U. 1964. n Sex - ohromatin." J. MED. GENET.
1 : 50 - 76.
126. Muel1er, G.A. , M.E. Gau1aen and W. TIrane. 1911 •
.. The effects of varying ooncentrations of
cOlohicine on the progression of grasshopper
neuroblasts into metaphase. " THE JOUR.NAL OF
CELL BIOLOGY. 40 : 253 - 265 •
.// 121. Mukherjee, B.E. 1965. " Cytological aspect of the
X - chromosome differentiation in normal and
abnormal oells. " CAN. GENET. CYTOL.
1 : 189 - 201.

-15J-
128. Mukherjee, B.B. and A.K. Sinha. 1963. " Further studies
on the pattern of chromosome duplioation in
cultured mammalian leucocytes." CAN. J.
.. ~ . GENET. CYTOL. 5: 490 - 495 •
129. Muller, H.J. 1927. It Artificial transmutation of genes."
SCIENCE. 66 : 84 - 87. , /'"
/' 130. Muller, H.J. 1940. " Bearing of the 'Drosophila.' work
on systematics. 1t p.p. 185 - 268 in I THE
NEW SYSTEMATICS OXFORD : CLAREDON PRESS. /' V/ 131. Nankivell, R.N. 1967. " A terminal assooiation of two
pericentric inversions in first metaphase
cells of the Australian grasshopper
Austriocetes interioris ( Acrididae )."
CHROllf.OSOMA ( BEHL. ) 22 : 42 - 68.
/132. Navashin, S.G. 1916. " On some sign.s of the internal /
crga~isation of chromosomes. " ( Russian ).
SBORN. K.A. TIMIRIAZEV. pp 185 - 214.
~/ 133. Nirmala, N. A. and M. R. Rajasekarasetty. 1971. n Chromosome
abnormalities in wild populations of
Pyrgomorpha bispinosa ( Acrididae : Orthoptera ~
PROe. OF 58TH INDIAN SCIENCE CONGRESS.
134. Nirmala, N.A. and M.R. Rajasekarasetty. 1973. .. Effeot
of p - benzoquinone on the male germ cells
of Poecilocerus pictus ( Aorididae :
Orthoptera )." PROC. OF THE 60TH INDIAN
SCIE~CE CONGRESS.

-151-
135. Nur, Uzi. 1961. " Meiotio behaviour of an unequal
biYa1ent in the grasshopper Ca11iptamus
pa1aestincnsis BDHR." CHROMOSOMA ( BERL. )
12 : 272 - 279.
136. Nur, Uzi. 1963. " A mitotioally unstable supernumerary
ohromosome with an aocumulation meohanism in
a grasshopper. fI CHROMOSOMA. 14 : 407 - 422. /
\ / 137. Nur, Uzi. 1968. " Synapsis and orossing over within a './
paracentrio inversion in the grasshopper.
Camnula pellucida. " CHROMOSOlV!A
25 (2) J 198 - 214.
138. Nur, Uzi. 1969. " Mitotio instability leading to an
aocumu1ation of B - ohromosomes in
grasshopper tt • CHROMOSOMA. 27 (1) , 1 - 19. ",<-
~~~ 139. Nur, Uzi. 1973~ n Random arrangement of ohromosomes in
,/
a radial metaphase configuration."
CHROMOSOMA ( BERL.) 40 I 263 - 267 • .• ~I".
140. Ohno, S. and T.S. Hausohka. 1960. " A11ocyc1y of the
x - ohromosome in tumors and normal tissues."
CANCER RES. 20 : 541 - 545.
141. Ostergren, G. 1947. n Proximal heteroohromatin,
struoture of centromere, and the meohanism of
its misdivision. tt BOT. NOT ( LUND ). 176 - 177. /'
V' 142. Ostergren, G., J. Mole - Bajer and A. Bajer. 1960. n An
interpretation of transport phenomena at
mitosis." ANN. N. Y. ACAD. SC1.
90 I 381 - 408.

-152-
\ 143. Ostertag, W., E. Duisberg and M. Sturmann. 1965. n The
mutagenio aotivity of oaffeine in man."
MUTAT. RES. 2 I 293 - 296.
144. Pati, P.C. and S.P. Bhunya. 1973 ... Effeot of a
tranquilliser drug, Largaotil, on the
spermatooytio ohromosomes of a grasshopper -
Poeoilooera piota." PROC. OF 60TH INDIAN
SCIENCE CONGRESS.
V'i45. Patterson, J.T. and W.S.stone. 1952 ... Evolution in the
genus Drosophila." Mao Mil1an Co. N.Y.
146. Pearson, P. It The limits of reoognition of human
ohromosomes in hybrid oells." NOBEL (23).
1973. 179 - 188. CHROMOSOME IDEI-:TIFICATION.
V'/' 147. Perry, P.E. and G.H. Jones. 1974. " Male and female
meiosis in grasshoppers I. Stethophyma
grossum." CHROMOSOMA. 47 (3) : 227 - 236.
148. Rahiman, M.A. and M.R. Rajasekarasetty. 1967a. " Nuolear
phenotype of Loousta migratoria .. CELLULAE.
66 (3) : 301 - 310.
149. Rao, S.R.V. 1960. " The aotion of urethene on ohromoso
mes of the grasshopper Poeoilooera piota
( Fabr.) Ion MEM. INDIAN BOT. SOC.
3 J 99 - 102.
150~ Rao, T.R. 1937. " A oomparative study of the ohromosomes
of eight genera of Indian Pyrgomorphini
( Acrididae ) lo" J. MORPH. 61 : 223 - 235.

-153-
151. Ray - Ohaudhuri, S.P. 1961 • " Induotion of ohromosome
aberrations in the spermatooytes of
grasshoppers." THE NUCLEUS. 4(1) I 47 - 66.
v/// 152. Ray - Chaudhuri, S.P.and A Gulla. 1952 ... X and neo - Y
mechanism of sex - determination in the
grasshopper Thisiocetrus ~ulcher." NATURE.
169 : 78 - 79.
/'
153. Ray - Chaudhuri, S.P. and A. Gulla. 1955. " Supernumerary
ohromosomes in two populations of the
grasshopper Aio1opus sp. and their behaviour
during spermatogenesis." J. GE~ET.
53 I 363 - 378.
154. Ray - Chaudhuri, S.P. and G.K. Manna. 1950. n On the
frequency of association between two
nonhomologoua chromosomes in the grasshopper
Gesonia punctifrons." J. EXP. ZOOL.
114 s 421 - 434. \ \ 155. Ray - Ohaudhuri, S. P. and G.K. Manna. 1951. I' Chromosome
evolution in the wild populations of
Aorid1dae. Part 1. On the Ooourrenoe of a
supernumerary ohromosome in Aio102US ap. ".
PROC. IND. ACAD. SC1. (B) 34 • 55 - 61.
156. Ray - Ohaudhuri, S.-P. and 1. Bose. 1948. " Meiosis in the
diploid and tetraploid spermatooytes of
Attractomorpha sp_ 11. PROG. ZOOL. SOO. BENG.
1 : 1 - 12.

-154-
157. Ray - Chaudhuri, S.P. and M.K. DlItt. 1947. " A oomparative
study of the chromosome struoture and
beha.viour in three genera of Indian grasshoppersll
PROC. ROY. SOC. EDIN. 62 : 292 - 298.
158. Ray - Chaudhuri, S.P. and T. Sharma. 1962. " Dicentric
bridge and cleavage in grasshopper
spermatocyte." PROe. ZOOL. SOC.
15 (2) I 83 - 88.
159. Reee, H. 1953. HEREDITY. 6 z 235.
160. Rees, H. and A. Jamieson. 1954. NATURE - 173 : 43.
; ./ ·161. Rhoadee, M.M. 1936. " A oytogenetio study Of a /'
ohromosome fragment in maize. tt GENETIOS.
21 : 491 - 502.
162. Rhoades, M.M. 1940. » Studies of a te1ooentrio ohromosome
in maize with referenoe to the stability of
its oentromere." GENETICS 25 I 483 - 520.
163. Robertson, W.R.B. 1915. J. MORPH. 26. 109.
164. Robertson, W.R.B. 1917. KANS UNIV. SCI. BULL. 10 • 275.
165. Rothfe1s, K.H. 1950. J. MORPH. 87 : 287.
166. Russell. L.B. 1961. " Genetios of mammalian sex -
ohromosomes." SCIENCE 133 : 1795 - 1803.
167. Saba, A.K. 1973. " Further observations on Uhe genesiS
of versene - induoed dioentric bridges in
grasshopper ohromosomes." PROO. OF 60TH
INDIAN SCIENCE CONGRESS.
168. Saba, A.K. 1974a. " Aotion of 2' - 4' - dihydroxyoha1oone
on grasshopper spermatooyte chromosomes."
CYTOLOGIA ( TOKYO ). 39(2) : 357 - 366.

-155-
/169. Baba, A.K. and A.R. Khudabaksh. 1974b. " Chromosome
aberrations induoed by melhy1 aloohol in
germinal oells of grasshopper." PROC OF
61ST INDIAN SCIENCE CONGRESS.
l70. Saha. A.K. and S. Chatterjee. 1973. " Observations on
the effeot of t - butyl aloohol ~on
grasshopper ohromosomes in the male germinal
line." PROe. OF 60TH INDIAN SCIENOE
CONGRESS.
171. Saba., A.K. and B. Ohatterjee. 1974. " Atropine - induced
chromosome anomalies in spermatocyte
chromosomes of grasshopper. " PROO. OF 61ST
INDIAN SOIENOE CONGRESS.
172. Sannom1ya, M. 1973. " Cytogenetio stUdies on natural
populations of grasshoppers with special
referenoe to B-ohromosomes : 11.
Attraotomorphas bedeli." OHROMOSOMA. ( BERL.)
44 (1) I 99 - 106.
1. 73. Sohrader, F. 1939. I' The struoture of the kinetochore
at meiosis." CHROMOSOMA (BEfiL.)
1 : 230 - 239.
174. Sohrader, F. 1944. " Mitosis: The movement of the
ohromosomes in oell division." N.Y. Columbia
University Press.
175. Sohroeter, G.L. and G.M. Godfrey. 1974. " The effeot of
supernumerary ohromatin in three speoies of
grasshopper." OAN. J. GENET. OYTOL. 16(2) :
285 - 296.

-156-
176. Sharma, A.X. and A. Sharma. 1960. " Spontaneous and
ohemioa11y induoed ohromosome breaks."
INTERN. REV. CITOL. 10 : 101 - 136.
)/"/- 177. Sharma. A.K. and A. Sharma. " Chromosome Teohniques -
Theory and Praotioe." Seoond Edition. 1912.
Butterworth and Co. Ltd.
/
178. Sharma, G.P., M.L. Gupta and G.S. Randhawa. 1967.
" Polysomy in Chroto~Onu8 traohypterus
( B1anohard ) ( Orthoptera : Aoridoidea :
Pyrgomorphidae ) from three more populations
and its possible role in animal speoiation."
RES. BULL. PUNJAB UNIY. 18(2) : 157 - 163.
179. Sharma, G.P., R. Parshad and H.M. Aggarwal. 1964.
"
" Cytoohemioal analysis of the lampbrush
fibres in the chromosomes of ChrotoS2nus
traohlEterus ( B1anchard ) ( Orthoptera :
Aoridodea I pyrgomorphidae ).It RES. BULL.
PUNJAB UNIV. SC1. : 15 (1) I 47 - 49.
180. Sharma, G.P., R. Parshad and M.L. Gupta. 1963e. " On
the seasonal variation in the mean ohiasma
frequency per nucleus in the wild population
of Chrotogonus traahY2t~ru~." RES. BULL.
PUNJAB UNIY. 14 : 313.
181. Sharma, G.P., R. Parshad and M.L. Gupta. 1965.
et Ohromosomal variation in the male germ oells
of Qhrotosonus traohypterus ( B1anohard )
( Orthoptera : Aoridoidea I Pyrgomorphidae )

-157-
from Ottu ( Punjab )." CELLULAE. 65 (3) :
,...,."., 295 - 314. ... /" '/
'// 182. Sharma, G.P., R. Parehad and 1'.5. Bed1. 1962.
" Breekdown of meiotio stability in
Chroto62nus traohy~terus ( Blanohard )
( Orthoptera : Aoridoidea : Pyrgomorphidae )."
RES. BULL. PUNJAB UNIV. 13 ( III - IV ) :
281 - 308.
183. Sharma, G.P., R. Parehad and T.S. Bedi. 1963. " A
translooation heterozygote in ChrotogonuB
traohypterus." JOUR. HEREDITY 54(2) : 62 - 65 ..
184. Shrivastav, Pushpa. 1975. " Karyologios1 studies in
three speoies of Aoridoidea." PROC. OF 62ND
INDIAr:i SCIENCE CONGRESS. pp 135 - 136.
i' 185. Slizynski, B.M. 1911. " Immediate effeots of oaffeine
mouse ohromosomes." CURRKNTS IR MODERN
BIOLOGY. 3 : 285 - 290. North Holland
Pllb1ishing Company.
186. SOllthern, D.I. 1961a. " Pseudo - multiple formation as
a oonsequenoe of prolonged nonhomo1ogous
assooiation in Metrioptera braohy;etera."
CHROMOSOMA ( BERL.) 21 : 272 - 284 •
. 181. Sri vas tavB, M.D.L. 1954. " studies on the stru,oture of
the chromosome of Chroto~onus inoertus
Bo1ivar ( Aorididae )." J. GEl'IET.
52 : 480 - 493.

-158-
\, ./'iS8. Srivastava. M. D.L. 1956. ft Studies on the struoture .".
and behaviour of the ohromosome of
Di880steira oarolina L." LA CELLULAE.
57 : 271 - 279.
189. Srivastava, M.D.L. 1959. " Meiotio aberrations in the
spermatogenesis of Poeoi1ooerus piotus. tt
LA CELLULAE. 61, 61 - 77.
,'190. Steffensen, D. 1955. " Breakage of ohromosomes at
meiosis by a oaloium defioienoy." NAT. ACAD.
SCI ( U.S. ) PROC. 41 : 155 - 160.
191. strid. A. 1968. " stable telooentrio ohromosomes
formed by spontanaeus misdivision in
Ni~ella d0f?rf1eri. It BOT. NOT .. ( LUND ).
121 : 153 - 164.
192. Subramanium, S. 1971. Personal oommunioation at
Bango1ore.
193. Sumner, A.T. 1972. " A simple teohnique for demonstrating
oentromerio heteroohromatin." EXPT. CELL
RES. 75 : 304 - 306.
,/194. Swansont C.P. 1958. " Cy1to1ogy and Cytogenetios." /
// Mo Mi110n Co., London.
195. Takizawa, S. and T. Narasawa. 1971. " Supernumerary
ohromosomes in two spe 01es OI' grasshoppers. ft
KROMOSOMA ( TOKYO ). 84 : 2679 - 2683 •
. " 196. Tjio, J.H. and A. Levan. 1954. n Chromosome analysis of '"
three hyperdiploid asoites tumours of the
mouse." K. FYSIOG SALLSK. HANDL. N.F. 65:51.

.,'
\
J""~'
-159-
191. Upoott, M.B. 1937. " The external meohanios of the
ohromosomes, VI. The behaviour of the oentro-
mere at meiosis." PROC. ROY. SOC. BENGAL.
124 : 336 - 361.
198. WaIters, M.S. 1954. ft A study of pseudo - bivalents in
meiosis of two interspeoifio hybrids of
Bromus." AMER. J. BOT. 41 : 160 - 171.
/'199. White, M.J.D. 1935. " The effeots of X - rays on mitosis /'
'" //
in the spermatogonial division of ~u~
migratoria." L. PROC. ROY. SOC. B
119 : 61 - 84.
200. White, M.J.D. 1940. " The origin and evolution of
multiple sex ohromosome mechanisms." J. GENET.
40 : 303 - 336.
201. White, M.J.D. 1941b. " The evolution of the sex
ohromosomes 11. The sex chromosome in the
Te11igoniidae and Acrididae and the principle
of evolutionary isolation of the X." "
J. GENET. 42 : 173 - 190.
202. White, M. J. D. 1949. GEl\ETICS. 34 • 537. •
203. White, M.J. D. 1951. " Cytogenetics of Orthopteroid
inseots." ADVANC. GEI\ET. 4 : 267 - 330.
204. White, M.J.D. 1953. " Multiple sex ohromosome mechani~ms
in the grasshopper genus Paratylotropidia."
AMER. NAT. 87 : 237 - 244. \ /
v' 205. White, M.J.D. 1954a. " An extreme form of ohiasma
looalisation in a speoies of Bryodema

-160-
( Orthoptera : Acrldidae ).It EVOLUTION
(Lancaster, Pa. ) 8: 350 - 358.
206. White, M.J.D. 1954b. " Animal Qytology and EvolutioIl."
3rd Edition. The University Press. Oambridge.
207. White, M.J.D. 1956. " Adaptive ohromosomal polymorphism
in an Australian €;Tasshopper." EVOLUTION.
10 : 298 - 313. , ("" .. '
'" 203. white, M.J.D. 1957a. " Cytogenetios of the grasshopper
Moraba scurra I. 'Meiosis of inter-racial and
inter-population hybrids." AU3T. J. ZOOL.
5 I 285 - 304.
209. white, M.J.D. 1957b. n Cytogenetios of the grasshopper
!,oraba scurra II. Heterotic systems anti their
interaction." AUST. J. ZOOL. 5 : 305 - 331.
210. White, M.J.D. 19570. n Cytogenetios of the grasshopper
Moraba sc~ra IV. Heterozygosity for -elastic
oonstrictions·." AUST. J. ZOOL. 5 , 348-354.
211. White, M.J.D. 1959. AUST. J. SOl. 22 : 32.
",," 212. White, M.J.D. 1961. tt Cytogenetics of the grasshopper ,,' Moraba sourra VI. A spontaneous pericentric
inversion." AUST. J. ZOOL. 9 : 784 - 790.
213. White, M.J.D. 1963 ... Cytogenetios of the grassho11per
Moraba sourra VIII. A oomp1ex spontaneous
translocation." CHROMOSOMA ( BERL.)
14 I 140 - 145.
214. White, M.J.D. 1965. It Chiasmatio and achiasmatio
meiosis in Afrioan Eumastaoid grasshoppers./f

-161-
CHROW.CSOMA (BERL.) 16(3): 271 - 301.
215. White, M.J.D. 1966. " A case of spontaneot:l.s chromosome
breakage at a specific locus oocurring at
meiosis ( undesoribed species of grasshopper
PW 21 - Orthopera : Eumestacidae :
Morabine.e ). 'I .A U8T. J. ZOOL.
/~ 14 (6) : 1027 - 1034.
~6. white, M.J.D. 1968. " Karyotypes and nuolear size in the
spermatogenesis of grasshoppers belonging to
the subfa:nilies Gompholllastacinde, Chininae
and Biroellinee ( Orthoptera : Eumastaoidae )."
// CARYOLOGIA. 21 (2) I 167 - 179.
j/" 217. White, M.J.D. 1970. " Ka,ryotypes and meiotic mechanisms
of some eumastacid grasshoppers from E. Africa,
rb"adagasoar, India arid South ftmerioa. "
CEROMOSO~~A. 30(1) : 62 - 97.
218. White, M.J.:9., A. lVJesa and R. Mesa .• 1967. " Neo - Xi
sex chromosome meohanisms in two speoies of
Tettigonioidea ( Orthoptera )." CYTOLOGIA.
32 (2) : 190 - 199.
/219. white, M.J.D. t H.L. Carson and J. Cheyney. 1964.
It Chromosomal races! ill the Aus·tralic.m
grasshopper Mora-ba viatica in 8, zOlle of
geogra.phio overlap." EVOLUTIOI~ ( LAWRENCE,
,/ KANSAS ). 18 : 417 429. //
,/ 220. White, M.J.D. and J. Cheyney. 1966. "Cytogenetics of the
cultrata group of morabine gra.sshopper-EI I.

-162-
A group of speoies with XY and X1X2Y sex
chromosome meohanism." AUST. J. ZOOL.
14 (5) : 821 - 834.
221. White, M.J.D., J. Cheyney and K.H.L. Key. 1963. MA
Parthenogenetio speoies of grasshopper with
complex structural heterozygosity
( Orthoptera : Acridoidea )." ADST. J. ZOOL.
11 I 1 - 19.
'/' 222. White, M. J. D. and K. H. L. Key. 1957d. 'f A cytotaxonomic l
study of the Pusi11a group of species in the
genus Austriocetes Uv. ( Orthoptera I
Acrididae ). n AUST. J. ZOOL. 5 : 56 - 87.
223. White, M.J.D. and L.E. Andrew. 1960. "Cytogenetics of
the grasshopper Moraba scurra V. Biometrio
effect of chromosomal inversions."
EVOLUTION ( LANCASTER, PA.) 14 : 284 - 292.
V/ 224. White, M.J.D. and L.E. Andrew. 1962. "Effeots of
chromosomal inversions on size and viability
in the grasshopper Moraba sourra. " In : The
Evolution of Living Organ~sms. ed. G.W.
Leeper. p. 94 - 101. Melbourne. Melbourne
University Press. oF'
/'
~ 225. White, M.J.D •• R.C. Lewontin and L.E. Andrew. 1963.
" Cytogenetios of the grasshopper Moraba
sourra VII. Geographio variation of adaptive
properties of inversions." EVOLUTION
( LAWRENCE, KANSAS ). 17(2) : 147 - 162.

\
-163-
226. White, M.J.D. t R.E. Blakith, R.M. Blakith and J. Cheyney
1967. "Cytogenetics of the viatica group of
morabine grasshoppers I. The 'coastal'
species." AUST. J. ZOOL. 15 : 263 - 302.
227. Wilson, E.B. 1906. "studies on ohromosomes Ill. The
sexual differenoes of the chromosomes in
Hemiptera, with some ,consideration on the
determination and inheritance of sex."
J. EXP. ZOOL. 3 I 1 -40.
228. Wilson. E.B. 1925. "The Cell in Development and
Heredity." Maomillan Company Limited. London.
229. Wilson, L. and M. Friedkin. 1967. " The biochemical
events of mitosis II. The in vivo and _. . in vitro binding of oolchicine in grasshopper
embryos and its possible relation to inhibition
of mitosis. " BIOCHBMISTRY.
6 (10) : 3126 - 3135.
=*=*=*I:=f= ~ *="*=:: :11: V =*=

IX
EXPLANATION OF FIGURES

Fig. 1
Fig. 2
Fig. 3
Fig. 4
Fig. 5
Fig. 6
Fig. 1
Fig. 8
Fig. 9
Fig. 10
Fig. 11
Fig. 12
-li4-
EXPLANATION OF FIGURES
Photograph of male speoimen of T. annulata.
Photograph of male specimen of
c. trachypterus.
Photograph of male specimen of L. indica.
Photograph of male specimen of A. crenulata.
Photograph of female specimen of P. pictus.
Photograph of cage used for rearing smaller
grasshoppers ( sliding glasS-door removed
from the front ).
Photograph of oage used for rearing P. piotus.
Photograph of diagrammatic representation of
approximate schedule of spermatogenesis in
c. trachlpterus.
Photograph of diagrammatio representation of
the method of counting metaphases ( or any
desired divisiona.l stage ) in the squash of a
testis - follicle.
X 800. Photomiorograph of spermatogonial
metaphase of T. ann~lata.
T. annulata. Photograph of spermatogonial
metaphase chromosomes arranged in order of
length. Sex chromosome is at the right hand
end.
X 800. T. annulata. Photomicrograph of
unsplit. heteropyonotio 'X' ohDomosome at

Fig. 13
.Fig. 14
Fig. 15
Fig. 16
Fig. 11
Fig. 18
Fig. 19
-165-
spermatogonial metaphase. A pachytene nucleus
at upper right - hand corner.
X 320. T. annulata. Photomicrograph of
pachytene nuclei with distinct positively
heteropyonotic sex - chromatin.
X 800 T. annulata. A pachytene nucleus with
clear split between the synapsed homologous
chromosomes.
X 800. T. ar~ulata. Pachytene nuclei with
centromerio ohromomeres which appear like
seoond arms t and interstitial blocks of
heterochromatin in the bivalents.
X 320. T. annulata. Photomicrograph of polar
view of metaphase I.
X 800. T. annuls!!. Photomicrograph of
bivalents at diplotene showing various confi
gurations aoquired due to varying number of
ohiasmata, the maximum number being three
( marked with arrow).
X 320. T. annulate. Photomicrograph of profiles
of metaphase I. The sex ohromosome lies
either 'above' or 'below' the equatorial
plate.
X 800. T. annulata. PhotomiorQgraph of
formation of • pseudo-chiasmata' by the distal '
ends of the separating chromosomes. at
anaphase I.

Fig. 20
Fig. 21
Fig. 22
Fig. 23
Fig. 24
Fig. 25
Fig. 26
Fig. 27
-166-
x 800. T. annu1ata. Photomicrograph of
beginning of movement of chromosomes towards
the opposite poles. A slightly heteropyonotic,
split 'X' chromosome can seen at the 'upper'
:.Dole.
X 800. T. annulata. Photomiorograph of late
anaphase I with 'X' - chromosome in the
split oondition at the right - hand side pole.
X 800. T. annulata. Photomicrograph of early
anaphase I with 'X'- ohromosome in unsplit
condition.
X 320. T. annulata. Photomiorograph of polar
view of metaphase 11 ( left oe11 ) and side
view of anaphase 11 ( right cell ) within the
marked area.
X 800. T. annulata. Photomiorograph of
magnified view of the area marked in Fig. 23,
showing metaphase 11 ( Right ) and anaphase
11 ( left ).
X 320. T. annulata. Photomiorograph of
telophase I with darkly stained spindle -
fibres stretched between the two poles.
X 320. T. annulata. Photomiorograph of late
telophase 11 ( three oells in the middle ).
X 320. T. annulata. Photomicrograph of
metaphase I ( left cell) with an extra
fragment at the equator.

Fig. 28
Fig. 29
Fig. 30
Pig. 31
Fig. 32
Fig. 33
Fig. 34
Fig. 35
Fig. 36
-161-
X 320. T. anntt1ata. Photomiorograph of two
metaphase I oel1s ( left and middle ) and one
anaphase I.,oe11 ( right ), eaoh wi th s.n extra
fragment.
X 320. T. annulat~. Photomiorograph of a
tetraploid cell ( middle ) at diakinesis.
X 320. T. annulata. Photomiorograph of two
tetraploid cells at diakinesis.
X 320. T. annulate. PhotomicrogTeph of one
tetraploid ( centre left ) and one normal
cell ( oentre fight ) at diakinesis.
T. annulate. Photograph of anaphase I with a
dioentric bridge aocompanied by a fragment.
X 800. T. annulata. Photomicrograph of polar
view of early anaphase I showing ohromatid -
breakage in the negatively heteropycnotic
sex chromosome.
x 800. T. annttlata. Photomicrograph of thirteen
chromosomes of a metaphase plate from the
male somatic tisstte ( gastric caeca ). Arrow
displays the indication of occttrrenoe of
second arms.
x 800. T. annula.ta. Photomicrograph of the
remaining ten ohromosomes of the same
metaphase plate shown in Fig. 34.
T. annulate. Photograph of diagrammatic
representation of the lengths of spermatogonial

Fig. 37
Fig. 38
Fig. 39
Fig. 40
Fig. 41
Fig. 42
-168-
metaphase chromosomes, based on the data
given in Table 1.
T. annulata. Photograph of diagrammatic
representa.tion of the lengths of' aomatio
metaphase chromosomes of male.
o. traohYRterus. Photograph of spermatogonial
metaphase chromosomes arranged in order of
size. Sex ohromosome at the right - hand end.
X 800. c. trachypterus. Photomicrograph of
paohytene nuoleus showing positively
heteropycnotic sex chromosome along with
a.nother similar, smaller mass of chromatin.
Destinct centromeric chromomeres can be seen.
X 800. o. trachyPterus. Photomicrograph of
side-view of early anaphase I, showing
'diamond' - shapes ( psuedochiasmata ) formed
by the bivalents. The cell at right - hand
end shows metaphase I.
X 800. c. trachypterus. Photomicrograph of
polar view of two cells at early anaphase I.
Pseudo chiasmata oan be seen in all the
bivalents. Sex chromosome shows a clear split
in the midcUe.
X 800. c. trachypterus. Photomicrograph of
eelrly anaphase one in the marked area, showing
the configuRations of the bivalents • Another
oell at metaphase can be seen at rl8ht-ha_d end.

Fig. 43
Fig. 44
Fig. 45
Fig. 46
Fig. 47
-169-
Photomicrograph of early anaphase I, shown in
Fig. 42, enlarged photographioally to reveal
the ns,ture of asoocia.tion of separating
chromosomes. Sex ohromosome shows a split
between its chromatids.
X 320. c. traohypterus. Photomiorograph of
cells at telophase 11. The single supernume
rary element can be seen lagging at equator
in top central oell.
X 800. c. tra.ohypterus. Photomiorograph showing
reduotion division of supernumerary bivalent
( right oell ) with 9dA + s at each pole;
and ( left oell ) with 9dA + aa at one pole
and 9dA + s at the other. Both the cells are
at anaphase I!.
X 320. o. trachlpterua. Photomiorograph
showing different behaviours of the super
numerary elements. In the n + 4s oondition
( Central cell ) 9dA + 1 es pass to eaoh pole.
In the n + 5s oondition ( upper right oell )
9dA + 1 ss pass to one pole and 9dA + 1 ss + 1 a
pass to the other. In the n+6s oondition,
ladA + 2ss + 2s oan be seen in the polar
view of anaphase 11 ( lower right oell ).
X 320. c. traohypterus. Photomiorograph of
two- tetraploid cells at diakinesis. The
chromosomal configJJ.ration in both oases is

Fig. 48
Fig. 49
Fig. 50
Fig. 51
Fig. 52
-110-
l8AA + 2x. The two sex ohromosomes lie away
from each other and show negative heteropycno
sis. Two normal oells at diakinesis can be
,seen ( extreme left and upper centrol
regions ).
X 800. c. traohypterus. Photomicrograph of a
tetraploid cell at metaphase 11. The
chromosomal configuration is 36dA + 2dx.
X 800. c. trachypterus. Photomiorograph of a
tetraploid oell at anaphase 11. Derived from
the binucleate oel1, the chromosomal
configurations of the two sete are
9dA + dx - 9dA + dx ( upper, right side ) and
9dA - 9dA ( lower side ), respectively.
X 320. c. traohYEterus. Photomiorograph of an
anaphase II oell with non-disjunction. The
ohromosomal oonfiguration is 8dA ( upper
pole) and lOdA ( lower pole ).
X 320. c. traohypterus. Photomiorograph of
ooourrenoe of non-cisjunotion at anaphase 11.
The ohromosomal oonfiguration is 8dA
( left pole ) and 10 dA ( right pole ). Other
oells at anaphase 11 show normal disjunotion
of the doughter ohromosomes.
X 800. c. traohlPterus. Photomicrograph of
oocurrenoe of non-disjunction at anaphase 11,
a oase where non- disjunotion had already

Fig. 53
Fig. 54
Fig. 55
Fig. 56
Fig. 57
-1'71-
occurred at anaphase I also. The latter oase
might have had 7A + 2,AA + x at one pole and
7A at the other. The 7A configuration
intead of. splitting into TdA - 7dA, splits
into 8dA - 6dA due to IJ.on-disjunction. A
normal cell at a:naphasf; 11 ( 9Wi + dx - 9dA
+ dx ) is seen on the left-hand side.
X 800. c. traohlEterus. Photom.iorograph of
anaphase I showing u..r..der - stained regions
in the ohromatids of' autosoma~ univa.lenta.
The latter are homologous and contain the
under - stained region in one chromatid only,
but at the same locus.
X 800. o. trachll?terus. Photomicrograph of'
anaphase 11 showing under - stained region
in one daughter univalent.
X 800. c. trachlpterus. Photomicrograph of
anaphase 11 showing under - stained region in
the daughteI' sex chromosome ( lower pole ).
X aOO. C. tr·aoh;YEtel'us. Photomicrograph of a
cell at anaphase 11 with a breakage in the
proximal region of a daughter autosome.
X 320. o. traohypterus. Photomicrograph
showing the presence of extra. fra§]Iler!t in two
oells at metaphase 11. Surrollnding oells
show different s-tages of' spermatogenesis.

Fig. 58
Fig. 59
Fig. 60
Fig. 61
Fig. 62
Fig. 63
Fig. 64
Fig. 65
-172-
x 800. c. traohlEterus. Photomiorograph of
metaphase from the gastrio oaeca of male
grasshopper. The sex ohromosome , being the
longest in the oomplement, oan be identified
only beoause of its length. Interphase oells
possess no positively heteropyonotio body.
c. traohlpterus. Photograph of diagramm&tio
representation of lengths of spermatogonial
metaphase chromosomes, based o~ the data
given in Table 4.
o. traohy?teras. Photograph of diagrammatio
representation of lengths of ohromosomes of
the somatio metaphase ( gas trio oaeoa ) of
male grasshopper.
X 800. Photomiorograph of spermatogonial
metaphase of Leva indioa.
L. indioa. Photograph of spermatogonial
metaphase ohromosomes arranged in order of
size. Sex ohromosome at the right - hand end.
L. lndioa. Photograph of diakinesis stage
showing an extra fragment.
~. indioa Photograph of polyploidy at anaphase
II. The total ohromosomal oonfiguration is
44dA, that is, lldA at Gaoh of the four poles.
L. indioa. Photograph of a oell at auaphase I,
posaessing an abnormal spindle. Ohromosomal
oonfiguration of the normal and bifid pole

Fig. 66
Fig. 67
Fig. 68
Fig. 69
Fig. 70
Fig. 71
Fig. 72
-173-
are lldA and 8dA - 3d., respectively.
L. indica. Photograph of spermatogonial
metaphase with breaka.ge in both the chromatids
of the long autosome.
L. ind10a Photograph of spermatogonial
metaphase with breakage in three long auto
aomes.
X 3~O. L. indica. Photomicrograph of
metephe.se from the gastric caeca of female
grasshopper. The two sex chromosomes oan be
identified only beceuse of their length,
being the lonBest in the oomplement. An
interphase cell at the bottom, shows an
indistinct heteropycnotic mass.
L. 1nd1ca. Photograph of diagrammatio
representation of lengths of spermatogonial
metaph~:!.se chrOG.lC'sC'IDE'!S, be.sed on the data.
given in Table 12.
L. 1ndica. Photograph of diGgrammatio
represente,t1on of lengths of chromosomes of
the SOI!l&tio metaphase ( gastrjc oaeca ) of
male grasshopper.
X 000. Photomicrograph of early spermatogonial
metaphase of Attr~tomorEha.~re~ulata.
A. cr~pu.lat~. Photograph of spermatogonial
metaphase chromosomes arranged in order of
length. Sex chromosome at the right - htomd end.

Fig. 13
Fig. 14
Fig. 15
Fig. 17
Fig. 18
Fig. 19
Fig. 80
Fig. 81
-174-
A. crenulata. Photograph of a cell at
anaphase I with a breaka.ge in one chromatid
of the sex ohromosome.
A. crenulata. Photograph of a oell at
anaphase I in the polar view. The chromatids
(dA) of the smallest autoBomes have
disju.ncted preoociously.
Photograph of early metaphase stage from the
gastric oaeoa of male nymph of A. orenulata.
Photograph of late metaphase stage from the
gastrio caeca of female nymph of A. crenu.lata.
A. orenulata. Photograph of diagrammatio
representation of lengths of spermatogonial
metaphase ohromosomes, based on the data
given in Table 15.
A. crenulata. Photograph of diagrammatic
representation of lengths of ohromosomes of
the somatic metaphase ohromosomes ( gastric
caeca ) of male grasshopper.
Photograph of somatic metaphase from the
gastrio caeoa of female nymph of P. pictus.
P. Eictus. Photograph of diagrammatic
r~presentation of the lengths of early
( black bars ) and lete ( dotted bars )
sperm:::1to::sonial metaphase ohromosomes.
P. pictus. Photograph of diagrammatically
representation of the lengths of the somatio

Fig. 82
Fig. 33
Fig. 84
Fig. 36
-175-
metaph~se chromosomes ( gastric caeca ) of
male grasshopper.
X 800. P. pictus. Sperma.togonial metaphase
with distinct bands of centromeric
heterochsomatin revealed by Giemsa staining.
The four subunits of the band can be clearly
seen in X - chromosome and M4 autosome.
X 800. P. pictus. Leptotene stage with the
oh~r~cteristic 'bouquet' arrangement,
Centromeres and telomeres centralize to form
the 'dark pole'. The positively heteropycno
tic sex ohr'omCltin lies at the oppoai te pole.
X 800. P. pictus. Pachytene stage showing
nine autosomal bivalents with distinct
oer:tromeric chromomerea and a jumb-bell
shaped sex - ohromatin body. Smallest
autosoID8J.. bivalenl; (SI) with clear telomeric
band.
x 800. P. pictus. Pachytene stage showing
distinot oentromeric chromomeres, oonstricted
off i'rom the main body of the chromosomes.
Another cell .at interphase ( 3xtrame left )
possesses a peculd:a.r, geometrio£;,lly sha.ped
heteroPycLotic body.
X jQ,). P. pictus. .Pa.cflytene nucleus showing
the presence of' a narrow, dark band at the
1;lroximal region t)f the chromosome (&.rrow marks).

Fig. 87
Fig. 88
Fig. 89
Fig. 90
Fig. 91
Fig. 92
-176-
X 80). P. pictus. Pachytene nucleus with the
centromeric chromomeres and proximal band of
heteroohromatin. Smallest bivalent ( SI )
looks uniformly dark because of the approxi
mation of diatal ~ proximal bands.
X 800. P. Eictus. Pachytene nucleus with very
distinct blobs of proxime.l heteroohromatin in
all the bivalents. Two dark bands are visible
in the diatel region of Sl* and 1 light band
in the sub-termim:tl distal pert of M6-
X 800. P. pictua. Dialiinesis stage showing
ole~rly the centromeric bands only. SI shows
almost uniform dark staining_ ·x· lies coiled
upon itself.
X 800. P. pictus. I'olar view of metaphase I.
The negctively heteropycnotic 'x· shows a dis
tinct oentromerio band. The bivalents also
show the oentromerio bands, which are not
very distinct are direoted towc~rds the oentre of
the spindle.
X 800. P. pictus. Side view of metaphase I.
Bands oannot be located in the much condensed
and over-stained biv':llents but the t X' does
possess dark regions at both the terminals.
X 800. P. pictus. Anaphase I with the sex
chromosome at the 'lower' pole. The bands of
·x· comprise one lighter band at the telomeric
end and one darker band at the proxima,l end.
The latter type of band is composed of 4.

Fig. 93
Fig. 94
Fig. 95
Fig. 96
Fig. 97
Fig. 98 I
-111-
subunits. Long and medium univalents have
olear oentromerio bands, while Sl-univalents
have both oentromerio and telomerio bands.
X 800. P. p1otus. Polar view of anaphase 11.
The ohromatids ( daughter ohromosomes ) show
oontraoted oentromerio bands at the proximal
ends, direoted towards the oentre of the
spindle. The two subunits of o-bands oan be
olearly seen in M2 and M5•
X 800. Effeot of oaffeine : High oondensation
of bivalents at diakinesis with oomplete
loss of 'lamp-brush' appearanoe. X -
ohromosome not distinguishable.
X 320. Effeot of oaffeine : Lesions in the
bivalents at diakinesis. Deohromatization
has resulted in the wavy outline of some of
the chromosomes.
x 800. Effeot of oaffeine : Polyploidy at
metaphase 11. The ohromosomal oonfiguration
of the tetraploid oell is 36A + 2X. The sex
ohromosomes oannot be distinguished on the f
basis of heteropyonosis as their staining
intensity is equal to that of the autosomes.
X 320. Effeot of oaffeine : Early stage of
olumping involving three or four ohromosomes
of eaoh oomplement at metaphase 11.
X 320. Effeot of oaffeine : Later stage of

Fig. 99
Fig. 100
Fig. 101
Fig. 102
Fig. 103
Fig. 104
-178-
clumping, involving almost all the chromo
somes of metaphase 11 cells. The identifica
tion of the divisional - stage is, however,
possible. Cell walls are not visible.
X 320. Effect of caffeine : Intense clumping
at metaphase 11 obtained after increasing the
treatment - time. The individual identity of
chromosomes is lost completely and the stage
can be identified with muoh difficulty. The
cell walls are not visible.
X 800. Effect of caffeine : Irregularity in
the arrangement of metaphase 11 chromosomes
because of the non-alignment of spindle -
fibres.
X 800. Effect of caffeine : Spindle dissolu
tion at metaphase 11, causing a scattering
of daughter chromosomes beyond the normal
dimensions.
X 800. Effect of caffeine : Breakage in the
smallest autosome~ Both the fragment and the
heteromorphic univalent pass to the same pole.
X 800. Effect of enteromycetin : Lesion in
the sex chromosome at spermatogonial
metaphase.
X 800. Effect of enteromycetin : Lesions in
both chromatids in the sex chromosome and one
of the longest autosomes, at spermatogonial

Fig. 105
Fig. 106
Fig. 107
Fig. 108
Fig. 109
Fig. 110
-179-
metaphase.
X 800. Effeot of enteromyoetin : Chromosome
-breakage at diplotene.
X 800. Effeot of enteromyoetin : One tetra
ploid oell at diakinesis ( right - hand side )
with the ohromosomal oonfiguration IBAA+2X.
The sex ohromosomes do not show any tendenoy
to synapse.
X 800. Effeot of enteromyoetin : Fragmentation
at metaphase 11. The "pseudo - satellites"
are due to the notohes whioh are the first
signs of the beginning of fragmentation.
X 800. Effeot of enteromyoetin : A tetraploid
oell at anaphase I showing stiokiness in the
telomerio ends of ohromosomes. All the
ohromosomes are highly condensed and no
separation between the ohromatids is visible.
X 800. Effeot of enteromyoetin : Two oells
at anaphase I with abnormal spindles. The
latter have one normal pole with the
ohromosomal configuration 9A+X. The other
pole is bifid and oontains 3A in the smaller
group and 6A in the larger group. The
univalents at the bifid pole enter telophase
earlier than the univalents at the normal
pole ( right cell ).
X 320. Effect of formalirle : Soattered

Fig. III
Fig. 112
Fig. 113
Fig. 114
Fig. 115
Fig. 116
-180-
bivalents at diplotene show dechromatization
and breakGge at chromosome level. There is an
indication of the destruction of the cell
wall as the bivalents lie scattered.
X 800. Effect of formaline : Diakinesis stage
showing the occurrence of interbivalent
connection. The latter comprises a thin
thread of chromatin connecting two of the
bivalents.
X 800. Effect of formaline : Late diplotene
stage with break2ge in one chromatid of a
medium bivalent.
X 800. Effect of forma1ine : A cell at
diakinesis showing clumpillg. The clumps are
formed by the grouping of 3 - 4 bivalents,
the identity of which is not completely lost.
The sex chromosome stains as intensely as
the Butosomes.
X 800. Effect of formaline : A cell at
prometaphase showing the linking of five
bivalents and the sex chromosome. Erosion of
chromatin material is also visible.
X 300. Effect of formaline : Metaphase I with
a chain of three bivalents and breakclge in
the clump ( left - hand side )
X 800. Effect of formaline : Destruction of
morphology of metaphase II chromosomes. A

Fig. 117
Fig. 118
Fig. 119
Fig. 120
Fig. 121
-181-
'pseudo-tetranucleate' appearance is found in
the cell ( upper right ) due to the grouping
of chromosomes. A pachytene nucleus in the s&me
photograph shows the 'snail-horn'-like
appearance.
X 800. Effect of formaline : Occurrence of
lesion in the sex chromosome at metaph&se 11.
The Butosomes hGve slightly affected morpho
logy.
X 800. Effect of formaline : Occurrenoe of
lesion in the sex ohromosome at metaphase 11
( right oell ). Another cell at metaph&se 11
( left-hand side ) shows a high degree of
dechromatization.
X 800. Effeot of formaline : Metaphase 11
showing a number of abnormalities. A promi
nent under-stained gap is present ill an
autosome. The sex chromosome shows a
stretohing of its oentromere. All the
chromosomes possess under-stained spots due
to dechromatiz~tion.
X 320. Effect of formaline : cells at meta
phase 11. The centromere of the sex ohromosome
is stretched to a great extent. Almost all
the chromosomes appear under-stained due to
extreme deohromatization.
X 300. Effect of formaline : 'star' configura-

Fig. 122
Fig. 123
Fig. 1~4
Fig. 125
Fig. 126
-182-
tion at metaphase 11 ( upper right corner ).
The stage is identifiable because some of the
univa1ents maintain their morphology by not
joining the tstar'. Another metaph&se 11
( lower left cor-ner) shows aberrant morphology
of the chromosomes.
X 800. Effects of nicotine : Fragmentation at
gonial metaphase. The sex chromosome is untou
ched and m~intains its negative heteropycnosis.
Almost all the autosomes have fragmented at
the chromatid-level.
X 800. Effect of nicotine : Diplotene stage
showing breaks in two bivalents. The fragments
lie away from the heteromorphic bivalents.
X 8JJ. Effect of nicotine : clumping at
diakinesis. The bivalents become thickened and
ag;~rega_te to form 'ball'-like structures.
X 800. Effect of nicotine : Clumping at
metaph<:.se I involving 2-3 bivalents.
X 800. Effect of nicotine : A diploid cell at
metaphase 11 with lesions in all the
chromosomes except the smaller ones. Some
chromosomes possess wavy outline.

X
ABBREVIATIONS

A
AA
asso. ch
bd
bif. pI
br
br. p.
c.c.
c.c.w.
oent. het.
chd. br
chr. br
cl
oon. bv
dA
dis. bI
dis. b2 dk. pI
dX
e.f
fr. ch
frg
g
h.b
into bv. con
-183-
ABBREVIATIONS
=
= = =
= =
= =
=
= = = =
= = = = = = = = = = = =
autosomal univalent.
autosomal bivalent.
assooiated ohromatids.
band.
bifid pole.
breakage.
broken piece.
oentrio ohromomeres.
oommon oell wall.
oentromerio heteroohromDtin.
ohromatid breakage.
chromosome breakage.
clump.
oonneoted biva.lents.
daughter autosome.
first distal band.
seoond distal band.
dark pole.
de,ughter sex ohromosome.
extra fragment.
free ohromatid.
fragment.
grass.
heteropyonotic body.
inter-bivalent connection.

into oh
into nu
Ll
L2
l.b.p.
le
lps
MI
m.l
m.s.
n
non. dis
p.c.
pro. b. het
prox. het
=
= =
::
= = ::
::
::
=
=
=
=
::
::
=
=
=
::
=
-184-
inters,ti tial ohiasma.
interpha.se nucleus.
first long autosome of P. piotus.
second long autosome of
P. piotus.
lagging broken pieoe.
lesion.
loops.
first medium autosome of
P. pictus.
seoond medium autosome of
P. pictus.
third medium autosome of
P. piotus.
fourth medium autosome of
P. :eictus.
fifth medium autosome of
P. Eiotus.
sixth medium autosome of
P. pictus.
movable lid.
moist sand.
haploid oomplement.
non-disjunction,
pseudo-chiasma.
proximal band of heteroohromatin.
proximal heterochromatin.

r. s.
S
SI
s
S. D.
sep. chd
sep. univ
sI. g. d.
sp. f
ss
st. confg
str. cen
tel. het
ter. ch
t.n.c.
t.p.c.
u.s.r.
w
X
-185-
=
= =
=
= = = = = =
=
=
=
=
= =
= =
reductional split.
standard diploid complement.
short autosome of P. piotus.
supernumerary univalent.
standard deviation.
separated ohromatids.
separating univalents.
sliding glass door.
spindle fibres.
supernumerary bivalent.
star oonfiguration.
stretohed centromere.
telomerio heterochromatin.
terminal chiasma.
texra-nucleate oell.
tetraploid oell.
understained region.
water.
sex chromosome.

Copyright © 2022 FDOKUMEN

