







Bahasa
Halaman
Hukum


1
Association analysis of functional variants of the FcgRIIa and FcgRIIIa genes with type
1 diabetes, celiac disease and rheumatoid arthritis
Behrooz Z. Alizadeh1,2,*, Gustavo Valdigem1, Marieke J. H. Coenen3, Alexandra
Zhernakova1, Barbara Franke3, Alienke Monsuur1, Piet L. C. M. van Riel4, Pilar Barrera4,
Timothy R. D. J. Radstake4, Bart. O. Roep2, Cisca Wijmenga1,5 and Bobby P.C. Koeleman1,2
1 Complex Genetics Section, Dept. of Medical Genetics, University Medical Center Utrecht,
Utrecht, the Netherlands.
2 Dept. of Immunohematology & Blood Transfusion, Leiden University Medical Center,
Leiden, the Netherlands.
3 Dept. of Anthropogenetics, Radboud University Nijmegen Medical Center, Nijmegen, the
Netherlands.
4 Dept. of Rheumatology and Experimental Rheumatology and Advanced Therapeutics,
Radboud University Nijmegen Medical Center, Nijmegen, the Netherlands.
5 Dept. of Genetics, University Medical Center Groningen, the Netherlands.
*Correspondence to:
Behrooz Z. Alizadeh, MD, PhD
Complex Genetics Section, Department of Medical Genetics, University Medical Center
Utrecht, PO Box 85060, 3508 AB Utrecht, the Netherlands. Tel: +31-30-2537925, Fax: +31-
30-2538479. E-mail: [email protected]
© The Author 2007. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: [email protected]
HMG Advance Access published July 25, 2007 by guest on February 21, 2016
http://hmg.oxfordjournals.org/
Dow
nloaded from

2
ABSTRACT
Background FcgRIIa and FcgRIIIa are potent modulators of the immune system which bind
(auto)antibodies and activate immune cells. The FcgRIIa*A519G and FcgRIIIa* A559C
functional variants have been associated with several immune-related diseases. We studied
FcgRIIa*A519G and FcgRIIIa*A559C SNPs in type 1 diabetes (T1D), celiac disease (CD),
and rheumatoid arthritis (RA) patients and controls, and included a meta-analysis of all recent
studies of FcgRIIIa*A559C and RA.
Method Our cohorts comprised 350 T1D, 519 CD, 639 RA patients and 1,359 controls, who
were genotyped for FcgRIIa*A519G and FcgRIIIa*A559C variants. Regression and EM-
algorithm-based haplotype analyses were used for the data analysis.
Results We found significant differences in genotype frequencies of FcgRIIa between
controls and patients with T1D (P=0.04), CD (P=0.000005) and RA (P=0.04). The
FcgRIIa*519GG genotype showed an increased risk for both T1D (OR=1.51; 95%CI=1.08-
2.12; P=0.015), and CD (OR=1.81, 95%CI=1.35-2.37; P=0.000004), but not for RA. There
was no difference in the frequency of FcgRIIIa*A559C genotypes or allelotypes between
controls with T1D, CD, and RA. We found that FcgRIIa and FcgRIIIa haplotypes frequencies
differed significantly between controls and patients with T1D (P=0.05), and with CD
(P=0.00038), but not with RA patients. Our meta-analysis showed a significant 1.37(1.14-
1.66) fold increased risk of RA for the FcgRIIIa*559CC (158VV) genotype (P=0.001).
Conclusion This is the first report that the FcgRIIa*519GG genotype predisposes to T1D and
CD. We confirmed the FcgRIIIa*559CC genotype is associated to RA. If replicated, our
findings would suggest the FcgRIIa*519G as a common risk factor for auto-immune diseases.
This may have clinical implications with regard to efficacy or safety of antibody-based
immuno-modulator therapies.
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

3
INTRODUCTION
Fc receptors I, II, and III (FcgRI, FcgRII, and FcgRIII) have evolved as crucial immune
response-modulating molecules that participate in reactivity to environmental antigens (1-3).
Eight genes clustered on chromosome 1q21-q24 encode three classes of FcgRs that are
expressed at the cell surface, namely the high-affinity receptor FcgRI (CD64) that binds
monomeric IgG, and FcgRII (CD32) and FcgRIII (CD16) that bind to multivalent IgG.
FcgRII and FcgRIII have different subclasses. FcgRIIa and FcgRIIIa associate with the
common FcRgamma-chain containing a stimulatory ITAM motif that is also present in the
intracellular tail of FcgRIIa, whereas FcgRIIb contains an inhibitory ITIM motif in the
cytoplasmic domain. FcgRIIa and FcgRIIIa stimulatory receptors are expressed by most
leukocytes, including monocytes, dendritic cells, macrophages, natural killer cells, platelets,
and endothelial cells, and a subpopulation of T-cells whereas FcgRIIb is expressed by B
lymphocytes, macrophages and dendritic cells (FcgRb2) (4). Upon binding of antibodies or
autoantibodies, FcgRIIa and FcgRIIIa activate immune cell functions, including phagocytosis,
and the release of inflammatory mediators, whereas FcgRIIb nullifies cell activation (3, 5).
Thus, FcgRs are part of an important regulatory system in intercepting and digesting
(auto)antibodies, which modulates antibody-mediated cellular cytotoxicity (1, 3, 6, 7).
FcgR isoforms were therefore linked to the pathogenic consequences triggered by
autoantibodies or immune complexes in autoimmune diseases such as rheumatoid arthritis
(RA) and systemic lupus erythematosus (SLE) (1, 3, 7).
FcgRIIa can have either histidine (H131) or arginine (R131) at amino acid position
131 located in the IgG binding site, which is encoded FcgRIIa*A519G SNP (8, 9). The
FcgRIIa*519G allele encodes the H131 high-binding isoform to IgG2, and the FcgRIIa*519A
encodes the low-binding R131 isoform (3, 9). The FcgRIIIa gene expresses two isoforms,
namely V158 (or V176) and F158 (or F176), which differ at amino acid 158 (or 176) in the
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

4
extracellular domain and are encoded by FcgRIIIa*A559C SNP. The FcgRIIIa*559C allele
encodes the high-binding isoform to IgG1 and IgG3 valine isoform (i.e. V158), and the
FcgRIIIa*559A encodes the 158 low-binding phenylalanine isoform (i.e. F158) (10, 11). It is
interesting that several independent genetic studies have linked chromosome 1q22-q24 to RA
and SLE (7, 12-14). However, extensive studies yielded different results on the relationship
between genetic polymorphisms in the FcgR genes, in particular the FcgRIIa and FcgRIIIa
high/low binding polymorphisms, to autoimmune diseases. On the one hand, meta-analyses
have confirmed association of the FcgRIIa*519G variant with susceptibility to SLE
worldwide (1, 15), and an association of FcgRIIIa*559C with SLE nephritis (16) and several
other inflammatory diseases (1, 4, 16-29). On the other hand, the association of these two
genes with RA remains interesting, but inconclusive for FcgRIIIa*559C, whereas no
association was found in multiple sclerosis (22, 25, 27, 30-32).
Like SLE and RA, type 1 diabetes (T1D) and celiac disease (CD) are common
autoimmune diseases, which are characterized clinically by the presence of autoantibodies:
against insulin-producing beta cells in T1D or intestinal mucosal cells in CD (33-37). The
experimental-clinical studies are of great interest and have implied a role for FcgRs in the
efficacy and side-effects of anti-CD3 immuno-modulatory therapy in T1D (30, 38-40).
However, the relevance of the FcgRIIa or FcgRIIIa genes to the pathogenesis of T1D and CD
has not been investigated so far. In this study, we investigated FcgRIIa*A519G and
FcgRIIIa*A559C and their joint effects on susceptibility to T1D, CD and RA, and performed
a meta-analysis on the association between FcgRIIIa*A559C with RA.
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

5
RESULTS
FcgRIIa
Overall, the frequencies of FcgRIIa*A519G genotypes differed significantly between
controls and T1D patients (P=0.04; Table 1). Individuals homozygous for the FcgRIIa*519G
variant (corresponding to the high-binding 131H isoform) were more frequent in T1D patients
(29.01%) than controls (22.65%), yielding a 1.51-fold increased risk for T1D in carriers
(95%CI 1.08–2.12; P=0.015, Table 1). Similarly, we found a significant difference in the
frequency of FcgRIIa*T519G genotypes in controls and CD patients (P=0.000005). CD
patients were also more frequently (32.61%) homozygous for the high-binding FcgRIIa*519G
than controls, leading to a 1.81-fold (1.35–2.37; P=0.000004) increase in risk for CD in
carriers (Table 1). Overall, the frequency of FcgRIIa*A519G genotypes differed significantly
between controls and RA patients (P=0.047; Table 1). RA patients were more frequently
heterozygous for this SNP than controls (54.08% vs. 48.50%), and were less often
homozygous for both A (27.20%) and G (18.72%) alleles than the controls (28.85%, and
22.65%, respectively). When the data was analyzed per chromosome, the high-binding
FcgRIIa*519G allele was significantly more frequent in patients with T1D (frequency 0.52;
P=0.01) and CD (0.55; P=0.000002) than in controls (0.47), yielding an increased risk of 1.24
(0.93-1.55) and 1.39 (1.04-1.74) for T1D and CD, respectively. We found no significant
difference in the frequencies of the FcgRIIa*519G allele in controls and RA patients.
FcgRIIIa
Overall, there was no significant difference of FcgRIIIa*A559C genotypes between controls
and T1D, CD, or RA (Table 1). Also, the frequency of the FcgRIIIa*559C allele did not differ
significantly among the controls and patients with T1D, CD or RA (Table 1).
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

6
Linkage disequilibrium and haplotype analysis
Since the FcgRIIa and FcgRIIIa genes reside close to each other in the same chromosomal
region, we tested whether there was an allelic association between these two genes in our
cohorts (Table 2). We found that FcgRIIa*A519G and FcgRIIIa*A559C were significantly
associated in the controls (P=5.97*10-19), T1D (7.06*10-8), CD (1.70*10-5) and RA patients
(5.30*10-6). To quantify the strength of LD, we calculated the D’ between the two loci (Table
2). The D’ as well as R2 between the FcgRIIa*A519G and FcgRIIIa*A559C variants were
low in our cohorts, indicating that there is a low LD between the studied variants.
Next, we tested whether there was a specific disease-associated FcgRIIa*A519G
_FcgRIII*A559C haplotype for T1D and CD. Overall, we found significant differences in the
frequencies of FcgRIIa*A519G_FcgRIII*A559C haplotypes between controls and T1D
(P=0.03), CD (P=0.00038) but not with RA patients (P=0.58; Table 2). The haplotype specific
risks indicate that it is mainly the FcgRIIa* 519G allele that explains the association of the
FcgRIIa-FcgRIIIa haplotypes to T1D and CD, while there were no differences in the
frequencies of these haplotypes between RA patients and controls (Table 2).
We further fitted the statistical model with interaction terms between FcgRIIa*A519G
and FcgRIIIa*A559C genotypes. We found no significant evidence of interaction between the
two loci in susceptibility to T1D or CD suggesting an FcgRIIIa-independent association of
FcgRIIa*519G variant with T1D and CD.
Meta-analysis of FcgRIIIa genotypes
To further clarify the inconclusive association between FcgRIIIa and RA we conducted a
meta-analysis of 11 studies together with our current data (Fig. 1). The heterogeneity test was
not statistically significant in the analysis of the FcgRIIIa*559CC genotype (P=0.35) whereas
it was significant for the analysis of the FcgRIIIa*559CA genotype (P=0.03). In Caucasians,
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

7
we found that the FcgRIIIa*559CC genotype was associated with a significant, 1.37 (95%CI
1.14 - 1.66; P=0.001) increased risk of RA in carriers (Fig. 1) while the FcgR*559CA
genotype was not associated to RA (P=0.33). We found no association of FcgRIIIa*559CC or
of FcgRIIIa*559CA to RA in Asians (Fig. 1).
DISCUSSION
We demonstrated that the FcgRIIa*519G variant is associated with both T1D and CD. We
found a relatively low LD between FcgRIIa*A519G and FcgRIIIa*A559C, and hence the
FcgRIIa–FcgRIIIa haplotypes showed different frequencies in the healthy controls and T1D or
CD patients that is mainly explained by FcgRIIa*519G, but not between RA patients and
controls. Our meta-analysis showed that FcgRIIIA*559CC genotype is significantly
associated with a mild increase in the risk of RA.
In our cohorts, homozygosity for the FcgRIIa*519G variant was consistently
associated with T1D and CD, which agrees with the studies that demonstrated homozygosity
for the FcgRIIa*519G variant is consistently associated with other autoimmune disorders (16,
20, 24-26). A meta-analysis of a large number of SLE patients and controls, confirmed the
FcgRIIa*519G variant as a genetic risk factor to SLE (16). Others have found an association
between this SNP and Guillain-Barre syndrome (GB), and RA (1, 4, 17-30).
Our findings also fit with the functional characteristics of this stimulatory variant in
determinations of immune hyper-reactivity, and thus suggest that the FcgRIIa gene is a
predisposing factor for several autoimmune diseases (1, 4, 16-29). This finding seems to be
similar to the associations of CTLA4 and PTPN22 with autoimmunity in general (41).
PTPN22 and CTLA4 are functionally completely different molecules and are involved in the
regulation of T cell function (41). There may be also a point of concern: the effect of FcgRIIa
varies with autoimmune diseases, in the sense that the FcgRIIa*519G variant is associated to
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

8
SLE and GB, and in our study, to CD and T1D, but it shows no consistent association with
multiple sclerosis (MS) or RA (22).Also in our study, the association of FcgRIIa*G to RA
was weaker and not convincing. The association was due to an excess of heterozygosity,
which lacks a plausible biological meaning. In contrast, we detected novel association of
FcgRIIa with CD and T1D with excess of homozygosity, which supports the hypothesis that
FcgRIIa is a crucial determinant of susceptibility to several autoimmune diseases. Thus,
FcgRIIa genotypes may serve as a marker for distinguishing the underlying basic pathologic
heterogeneity in autoimmune diseases.
We could not confirm an association of the FcgRIIIa*A559C genotypes to RA in our
study. Indeed, reports on this association are contradictory. Some have shown an association
of this marker with RA in the English, Indian and Pakistani populations (26, 31, 42), but
several other studies found no association between the FcgRIII*559C variant and RA in
Japanese, Taiwanese, Spanish, Norwegian, Dutch, or Spanish populations (22, 25, 43-45).
However, our meta-analysis confirmed an association of FcgRIIIa*559CC genotypes to RA in
Caucasians. This finding agrees with the results of an earlier pervious meta-analysis that
confirmed the association between FcgRIIIa*559CC and SLE (15). It should be noted that the
latter studies reported a positive association between FcgRIIIa and RA in subgroups of
patients who shared a particular clinical characteristic such as shared anti-GPI positivity (46),
severe in hands RA (25) nodular arthritis (26) or only in men (25), which are indicators of
disease severity. These observations fit with the finding that the FcgRIIIa*559C variant is
associated with lupus nephritis (1), a disease complication, implying that the FcgRIIIa gene
may be involved in the course of autoimmune diseases. This would partly explain the
contradictory findings in RA, including those in the present study. Altogether, and given the
findings of our meta-analysis we concluded that FcgRIIIa is associated to RA in Caucasians,
most likely in those with a severe form of RA.
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

9
Our study focused on the FcgRIIa and FcgRIIIa genes, two activating receptors for
immuno-effector cells. However, there are effective classes of inhibitory FcgRs, such as
FcgRIIb and FcgRIIIb, which were also clustered to chromosome 1q21-q24, often present as
pairs on the cell surface, and associated with several autoimmune diseases (3, 15, 47). The
inhibitory receptors nullify stimulation signals from FcgRIIa and FcgRIIIa. Interestingly,
association between copy number variation of FcgRIIIb and lupus nephritis, SLE and
Wegner’s granulomatosis has been reported (48, 49). The low LD FcgRIIa and FcgRIIIa and
the close proximity of FcgRIIIb to the latter indicates that FcgRIIIb may be an independent
risk locus in this region that warrants further investigation (48). Our risk estimates may
therefore be skewed, due to the modifying effects of “functionally” interacting variants in
neighboring inhibitory FcgRs.
We have tested FcgRIIa*A519G and FcgRIIIa*A559C alleles on the basis of prior
evidence of previous reported association in other autoimmune diseases. Thus, multiple
testing may not be applicable to our study. Nevertheless, significant association would still
remain for the CD study when a conservative six-fold Bonferroni correction for multiple-
hypothesis testing is used.
It has been shown that the efficacy and compliance to the immunomodulatory
monoclonal antibody against CD3 (anti-CD3 Ab) therapy differs according to imuunoglobolin
FcgRIIa, and FcgRIIIa isoforms due to variation in the encoding genes (39, 40, 50, 51), while
the non-FcR binding anti-CD3 is less immunogenic than FcR-binding forms (52-54).
Furthermore it has been shown that FcgRs variations influences the release of cytokines,
which may underlie the occurrence of side-effects after initiation of anti-CD3 mAb (54). On
the basis of these observations, several studies investigated the role variations in the FcgRIIa,
FcgRIIIa genes in the efficacy of anti-CD3 therapy in different immune diseases, and they
found conflicting and non-consistent results (55-57). Therefore, our findings may further
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

10
delineate that the disease risk genotype of FcgRIIa may modify the efficacy of anti-CD3 Ab
oriented therapies, a hypothesis which remains to be tested.
In conclusion, this is the first report of homozygosity for FcgRIIa*519G as a
predisposing factor to T1D and CD but not to RA. Our novel findings need to be replicated by
others. We also found that homozygosity for FcgRIIIa*559C was associated to RA in the
meta-analysis. Our findings provide basic insight into the possible mechanism of AIDs and
may well have clinical implications with respect to the efficacy, and side-effects, of immuno-
modulator interventions, such as anti-CD3 therapy.
MATERIALS AND METHODS
Study populations
Type 1 Diabetes. Patients were retrieved from the Kolibri T1D cohort that includes 350
Dutch patients with juvenile onset T1D (median 8.7 yrs, range 1-17 yrs). The cohort was
collected consecutively after diagnosis by pediatricians in the Southwestern part of the
Netherlands between 1995 and 1999. The diagnosis was made according to International
Society of Pediatric and Adolescent Diabetes (ISPAD) and WHO criteria.
Celiac Disease. Patients were included from cohorts of Dutch CD patients that included
children and adults. All the 519 CD patients have been diagnosed according to the revised
ESPGHAN criteria (58). More than 90% of the patients were HLA-DQ2 positive. The
patients’ initial biopsy specimens were retrieved and all showed a Marsh III lesion on re-
evaluation by experienced pathologists.
Rheumatoid Arthritis. The characteristics of patients with RA have been described
elsewhere (59). In brief, the RA patients included in our study were recruited from an ongoing
early-RA inception study that was started in 1985 at the Department of Rheumatology,
Radboud University Nijmegen Medical Centre (RUNMC) in the Netherlands. All the patients
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

11
were diagnosed according to the American College of Rheumatology criteria for RA (60), had
a disease duration of less than one year, and had no prior use of disease-modifying anti-
rheumatic drugs or biological agents before presentation. All patients in the early-RA
inception cohort are regularly monitored for disease phenotype, severity, and outcome. In
total, 639 Dutch patients with RA were included in our study.
The T1D, CD and RA patients were also born in the Netherlands and had at least three out of
four grandparents also born in the Netherlands.
Control subjects. A total of 1,359 unrelated Dutch individuals were selected for being born
in the Netherlands and had at least three out of four grandparents also born in the Netherlands.
All the patients and controls gave their informed consent and the medical ethical
committee of the University Medical Center Utrecht or the Radboud University Nijmegen
Medical Centre approved this study.
Genotyping
We genotyped our study cohorts for the FcgRIIa* A519G SNP (rs1801274), and the
FcgRIIIa*A559C SNP (rs396991). The genotyping of FcgRIIa*A519G SNP was successful
for 1,324 controls, 324 T1D patients, 509 CD patients, and 625 RA patients, while the
genotyping for FcgRIIIa* A559C succeeded in 1,326 controls, 319 T1D patients, 510 CD, and
601 RA patients. Participants’ genotypes for both the FcgRIIa*A519G and FcgRIIIa*A559C
variants were available for 1,290 controls, 314 T1D patients, 503 CD patients and 587 RA
patients. Genotype frequencies of FcgRIIa* A519G and FcgRIIIa* A559C variants were in
Hardy-Weinberg proportions in controls.
The Taqman(R) SNP genotyping assays for PCR were supplied by Applied Biosystems
(Nieuwerkerk a/d Ijssel, the Netherlands) for FcgRIIa*A519G (ABI assay identification
number C__9077561_20), or for FcgRIIIa*A559C (C_25815666_10).
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

12
Meta-analysis. We searched Medline for all publications relating to association studies
using the combinations of quarries of “FcgRIIa*A519G”, “FcgRIIIa*A559C”, “IIIA”, “IIA”,
“RA”, “Rheumatoid”, “Arthritis”, “FcgR”, “FcgammaRIIIA”, “FcgammaRIIA” and checked
the references from retrieved publications for additional studies. We identified 14 articles: two
performed analysis in 2 different ethnic populations i.e. Caucasians and Indians (26, 27), of
which each analysis was treated as a separate entity in the meta-analysis. One study provided
family-based association study(44); and one study presented data only for patients (61). These
studies were not included in our meta-analysis. All the studies used the same diagnostic
criteria for RA and patients were diagnosed according to the American College of
Rheumatology criteria (60). In total we included 11 studies in our meta-analysis, of which 8
were conducted in Caucasian populations (25-27, 42, 45, 62, 63) including two analysis in
Indians (26, 27), and three in Asians (22, 30, 43). In total the meta-analysis covered
chromosomes from 3341 patients and 4161 controls.
Data analyses
Genotype and allele frequencies were calculated by direct counting. Hardy-Weinberg
equilibrium was checked by using GenePOP software. First the data was analyzed overall by
genotypes. Chi-squared tests were used to compare frequencies. Regression analysis was used
to first estimate genotypic main-effect odds ratios (OR) and the corresponding 95%
confidence intervals (95% CI), and then the main effect of the risk allele of the two SNPs and
the interaction effect OR (95% CI) between variants of the FcgRIIa*A519G and
FcgRIIIa*A559C SNPs. Coefficient (D’) and corresponding 95% confidence bound of pair-
wise linkage disequilibrium as well as r-Squared (r2) between FcgRIIa*A519G and
FcgRIIIa*A559C using the LD plot module implemented in the Haploview software, version
3.2. We tested for allelic association between these SNPs independently in the controls, T1D,
CD, and RA patients using an expectation maximization (EM) algorithm as described
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

13
elsewhere (64). A two-tailed p-value < 0.05 was considered statistically significant. Data
analysis was performed using UNPHASED (65, 66), and STATA statistical software, version
8.0 for MS Windows.
Meta-analysis. For each study the frequency of FcgRIIIa genotypes were derived from
counting method in patients and controls. In all the studies, allele frequencies were consistent
with Hardy-Weinberg Equilibrium. In addition to the total group, we classified the studies into
Caucasians and Asian. The effect of the FcgRIIIa*A559C genotypes were assessed by
comparing the frequency of the FcgRIIIa*559AC and FcgRIIIa*559CC genotype versus the
FcgRIIIa*559AA genotype in patients and controls. We used funnel plots to examine
publication bias of reported associations. The study of Milicic et al.(2002) had a very skewed
frequency for the FcgRIIIa* 559CC genotype that led to this study being an oulier. We
excluded this study from the analysis. To accommodate the effect of different ethnic
backgrounds on the association between FcgRIIIa*559AC and RA, heterogeneity between
studies was tested using the Chi-squared test, and the confidence interval for the odds ratio
was estimated using a random effect model. We included only FcgRIIIa*A559C in the meta-
analysis; since there were not enough data on the association of FcgRIIa*A519G variant and
RA. The meta-analysis was conducted using the Cochrane Review Manager, version 4.1.2.
ACKNOWLEDGMENTS
This study was supported by grants from the Dutch Diabetes Research Foundation (97.137),
the Netherlands Organization for Health Research and Development (ZonMW), and Novo
Nordisk A/S. Collection and genotyping of controls was part of a celiac disease project
supported by grants from the Netherlands Organization for Scientific Research (NWO grants
902-22-094 and 912-02-028), the Dutch Digestive Diseases Foundation (grants 97-44 and 03-
06) and the Celiac Disease Consortium, an innovative cluster approved by the Netherlands
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

14
Genomics Initiative and partially funded by the Dutch government (grant BSIK03009). We
thank Eric Strengman for technical assistance, Harry van Someren as database manager and
Jackie Senior for critically reading the manuscript.
Conflict of Interests statement. None
REFERENCES
1. Karassa, F.B., Trikalinos, T.A. and Ioannidis, J.P. (2004) The role of FcgammaRIIA
and IIIA polymorphisms in autoimmune diseases. Biomed Pharmacother, 58, 286-
291.
2. Dijstelbloem, H.M., van de Winkel, J.G. and Kallenberg, C.G. (2001) Inflammation in
autoimmunity: receptors for IgG revisited. Trends Immunol, 22, 510-516.
3. Takai, T. (2005) Fc receptors and their role in immune regulation and autoimmunity. J
Clin Immunol, 25, 1-18.
4. Karassa, F.B., Bijl, M., Davies, K.A., Kallenberg, C.G., Khamashta, M.A., Manger,
K., Michel, M., Piette, J.C., Salmon, J.E., Song, Y.W. et al. (2003) Role of the
Fcgamma receptor IIA polymorphism in the antiphospholipid syndrome: an
international meta-analysis. Arthritis Rheum, 48, 1930-1938.
5. Nakamura, A., Akiyama, K. and Takai, T. (2005) Fc receptor targeting in the
treatment of allergy, autoimmune diseases and cancer. Expert Opin Ther Targets, 9,
169-190.
6. Bredius, R.G., Fijen, C.A., De Haas, M., Kuijper, E.J., Weening, R.S., Van de Winkel,
J.G. and Out, T.A. (1994) Role of neutrophil Fc gamma RIIa (CD32) and Fc gamma
RIIIb (CD16) polymorphic forms in phagocytosis of human IgG1- and IgG3-
opsonized bacteria and erythrocytes. Immunology, 83, 624-630.
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

15
7. Corper, A.L., Sohi, M.K., Bonagura, V.R., Steinitz, M., Jefferis, R., Feinstein, A.,
Beale, D., Taussig, M.J. and Sutton, B.J. (1997) Structure of human IgM rheumatoid
factor Fab bound to its autoantigen IgG Fc reveals a novel topology of antibody-
antigen interaction. Nat Struct Biol, 4, 374-381.
8. Salmon, J.E., Edberg, J.C., Brogle, N.L. and Kimberly, R.P. (1992) Allelic
polymorphisms of human Fc gamma receptor IIA and Fc gamma receptor IIIB.
Independent mechanisms for differences in human phagocyte function. J Clin Invest,
89, 1274-1281.
9. Bredius, R.G., Derkx, B.H., Fijen, C.A., de Wit, T.P., de Haas, M., Weening, R.S., van
de Winkel, J.G. and Out, T.A. (1994) Fc gamma receptor IIa (CD32) polymorphism in
fulminant meningococcal septic shock in children. J Infect Dis, 170, 848-853.
10. Salmon, J.E., Edberg, J.C. and Kimberly, R.P. (1990) Fc gamma receptor III on
human neutrophils. Allelic variants have functionally distinct capacities. J Clin Invest,
85, 1287-1295.
11. Wu, J., Edberg, J.C., Redecha, P.B., Bansal, V., Guyre, P.M., Coleman, K., Salmon,
J.E. and Kimberly, R.P. (1997) A novel polymorphism of FcgammaRIIIa (CD16)
alters receptor function and predisposes to autoimmune disease. J Clin Invest, 100,
1059-1070.
12. Lee, Y.H. and Nath, S.K. (2005) Systemic lupus erythematosus susceptibility loci
defined by genome scan meta-analysis. Hum Genet, 118, 434-443.
13. Forabosco, P., Gorman, J.D., Cleveland, C., Kelly, J.A., Fisher, S.A., Ortmann, W.A.,
Johansson, C., Johanneson, B., Moser, K.L., Gaffney, P.M. et al. (2006) Meta-analysis
of genome-wide linkage studies of systemic lupus erythematosus. Genes Immun, 7,
609-614.
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

16
14. Choi, S.J., Rho, Y.H., Ji, J.D., Song, G.G. and Lee, Y.H. (2006) Genome scan meta-
analysis of rheumatoid arthritis. Rheumatology, 45, 166-170.
15. Karassa, F.B., Trikalinos, T.A. and Ioannidis, J.P. (2002) Role of the Fcgamma
receptor IIa polymorphism in susceptibility to systemic lupus erythematosus and lupus
nephritis: a meta-analysis. Arthritis Rheum, 46, 1563-1571.
16. Karassa, F.B., Trikalinos, T.A. and Ioannidis, J.P. (2003) The Fc gamma RIIIA-F158
allele is a risk factor for the development of lupus nephritis: a meta-analysis. Kidney
Int, 63, 1475-1482.
17. Zuniga, R., Ng, S., Peterson, M.G., Reveille, J.D., Baethge, B.A., Alarcon, G.S. and
Salmon, J.E. (2001) Low-binding alleles of Fcgamma receptor types IIA and IIIA are
inherited independently and are associated with systemic lupus erythematosus in
Hispanic patients. Arthritis Rheum, 44, 361-367.
18. Dijstelbloem, H.M., Scheepers, R.H., Oost, W.W., Stegeman, C.A., van der Pol, W.L.,
Sluiter, W.J., Kallenberg, C.G., van de Winkel, J.G. and Tervaert, J.W. (1999)
Fcgamma receptor polymorphisms in Wegener's granulomatosis: risk factors for
disease relapse. Arthritis Rheum, 42, 1823-1827.
19. Salmon, J.E., Ng, S., Yoo, D.H., Kim, T.H., Kim, S.Y. and Song, G.G. (1999) Altered
distribution of Fcgamma receptor IIIA alleles in a cohort of Korean patients with lupus
nephritis. Arthritis Rheum, 42, 818-819.
20. Trikalinos, T.A., Karassa, F.B. and Ioannidis, J.P. (2001) Meta-analysis of the
association between low-affinity Fcgamma receptor gene polymorphisms and
hematologic and autoimmune disease. Blood, 98, 1634-1635.
21. Lehrnbecher, T., Foster, C.B., Zhu, S., Leitman, S.F., Goldin, L.R., Huppi, K. and
Chanock, S.J. (1999) Variant genotypes of the low-affinity Fcgamma receptors in two
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

17
control populations and a review of low-affinity Fcgamma receptor polymorphisms in
control and disease populations. Blood, 94, 4220-4232.
22. Kyogoku, C., Tsuchiya, N., Matsuta, K. and Tokunaga, K. (2002) Studies on the
association of Fc gamma receptor IIA, IIB, IIIA and IIIB polymorphisms with
rheumatoid arthritis in the Japanese: evidence for a genetic interaction between HLA-
DRB1 and FCGR3A. Genes Immun, 3, 488-493.
23. Kyogoku, C., Dijstelbloem, H.M., Tsuchiya, N., Hatta, Y., Kato, H., Yamaguchi, A.,
Fukazawa, T., Jansen, M.D., Hashimoto, H., van de Winkel, J.G. et al. (2002)
Fcgamma receptor gene polymorphisms in Japanese patients with systemic lupus
erythematosus: contribution of FCGR2B to genetic susceptibility. Arthritis Rheum, 46,
1242-1254.
24. van der Pol, W.L., van den Berg, L.H., Scheepers, R.H., van der Bom, J.G., van
Doorn, P.A., van Koningsveld, R., van den Broek, M.C., Wokke, J.H. and van de
Winkel, J.G. (2000) IgG receptor IIa alleles determine susceptibility and severity of
Guillain-Barre syndrome. Neurology, 54, 1661-1665.
25. Kastbom, A., Ahmadi, A., Soderkvist, P. and Skogh, T. (2005) The 158V
polymorphism of Fc gamma receptor type IIIA in early rheumatoid arthritis: increased
susceptibility and severity in male patients (the Swedish TIRA project). Rheumatology
(Oxford), 44, 1294-1298.
26. Morgan, A.W., Griffiths, B., Ponchel, F., Montague, B.M., Ali, M., Gardner, P.P.,
Gooi, H.C., Situnayake, R.D., Markham, A.F., Emery, P. et al. (2000) Fcgamma
receptor type IIIA is associated with rheumatoid arthritis in two distinct ethnic groups.
Arthritis Rheum, 43, 2328-2334.
27. Milicic, A., Misra, R., Agrawal, S., Aggarwal, A., Brown, M.A. and Wordsworth, B.P.
(2002) The F158V polymorphism in FcgammaRIIIA shows disparate associations
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

18
with rheumatoid arthritis in two genetically distinct populations. Ann Rheum Dis, 61,
1021-1023.
28. Chu, Z.T., Tsuchiya, N., Kyogoku, C., Ohashi, J., Qian, Y.P., Xu, S.B., Mao, C.Z.,
Chu, J.Y. and Tokunaga, K. (2004) Association of Fcgamma receptor IIb
polymorphism with susceptibility to systemic lupus erythematosus in Chinese: a
common susceptibility gene in the Asian populations. Tissue Antigens, 63, 21-27.
29. Siriboonrit, U., Tsuchiya, N., Sirikong, M., Kyogoku, C., Bejrachandra, S.,
Suthipinittharm, P., Luangtrakool, K., Srinak, D., Thongpradit, R., Fujiwara, K. et al.
(2003) Association of Fcgamma receptor IIb and IIIb polymorphisms with
susceptibility to systemic lupus erythematosus in Thais. Tissue Antigens, 61, 374-383.
30. Matsumoto, I., Zhang, H., Muraki, Y., Hayashi, T., Yasukochi, T., Kori, Y., Goto, D.,
Ito, S., Tsutsumi, A. and Sumida, T. (2005) A functional variant of Fcgamma receptor
IIIA is associated with rheumatoid arthritis in individuals who are positive for anti-
glucose-6-phosphate isomerase antibodies. Arthritis Res Ther, 7, 1183-1188.
31. Morgan, A.W., Barrett, J.H., Griffiths, B., Subramanian, D., Robinson, J.I., Keyte,
V.H., Ali, M., Jones, E.A., Old, R.W., Ponchel, F. et al. (2006) Analysis of Fcgamma
receptor haplotypes in rheumatoid arthritis: FCGR3A remains a major susceptibility
gene at this locus, with an additional contribution from FCGR3B. Arthritis Res Ther,
8, 5.
32. Breij, E.C., van der Pol, W.L., van Winsen, L., Jansen, M.D., Dijkstra, C.D., van de
Winkel, J.G. and Uitdehaag, B.M. (2003) No association of Fc gamma RIIa, Fc
gamma RIIIa and Fc gamma RIIIb polymorphisms with MS. J Neuroimmunol, 140,
210-215.
33. Eisenbarth, G.S. (1986) Type I diabetes mellitus. A chronic autoimmune disease. N
Engl J Med, 314, 1360-1368.
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

19
34. Atkinson, M.A. and Eisenbarth, G.S. (2001) Type 1 diabetes: new perspectives on
disease pathogenesis and treatment. Lancet, 358, 221-229.
35. Narendran, P., Williams, A.J., Elsegood, K., Leech, N.J. and Dayan, C.M. (2003)
Humoral and cellular immune responses to proinsulin in adults with newly diagnosed
type 1 diabetes. Diabetes Metab Res Rev, 19, 52-59.
36. Colman, P.G., McNair, P.D., Gellert, S., Kewming, K., Schmidli, R.S., Steele, C.E.
and Harrison, L.C. (2002) Development of autoantibodies to islet antigens during
childhood: implications for preclinical type 1 diabetes screening. Pediatr Diabetes, 3,
144-148.
37. Monsuur, A.J. and Wijmenga, C. (2006) Understanding the molecular basis of celiac
disease: What genetic studies reveal? Annals of Medicine, 38, 578-591.
38. Magnusson, V., Johanneson, B., Lima, G., Odeberg, J., Alarcon-Segovia, D. and
Alarcon-Riquelme, M.E. (2004) Both risk alleles for FcgammaRIIA and
FcgammaRIIIA are susceptibility factors for SLE: a unifying hypothesis. Genes
Immun, 5, 130-137.
39. Chatenoud, L. (2003) CD3 antibody treatment stimulates the functional capability of
regulatory T cells. Novartis Found Symp, 252, 279-290.
40. Keymeulen, B., Vandemeulebroucke, E., Ziegler, A.G., Mathieu, C., Kaufman, L.,
Hale, G., Gorus, F., Goldman, M., Walter, M., Candon, S. et al. (2005) Insulin needs
after CD3-antibody therapy in new-onset type 1 diabetes. N Engl J Med, 352, 2598-
2608.
41. Brand, O., Gough, S. and Heward, J. (2005) HLA , CTLA-4 and PTPN22 : the shared
genetic master-key to autoimmunity? Expert Rev Mol Med, 7, 1-15.
42. Morgan, A.W., Keyte, V.H., Babbage, S.J., Robinson, J.I., Ponchel, F., Barrett, J.H.,
Bhakta, B.B., Bingham, S.J., Buch, M.H., Conaghan, P.G. et al. (2003)
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

20
FcgammaRIIIA-158V and rheumatoid arthritis: a confirmation study. Rheumatology
(Oxford), 42, 528-533.
43. Chen, J.Y., Wang, C.M., Wu, J.M., Ho, H.H. and Luo, S.F. (2006) Association of
rheumatoid factor production with FcgammaRIIIa polymorphism in Taiwanese
rheumatoid arthritis. Clin Exp Immunol, 144, 10-16.
44. Radstake, T.R., Petit, E., Pierlot, C., van de Putte, L.B., Cornelis, F. and Barrera, P.
(2003) Role of Fcgamma receptors IIA, IIIA, and IIIB in susceptibility to rheumatoid
arthritis. J Rheumatol, 30, 926-933.
45. Nieto, A., Caliz, R., Pascual, M., Mataran, L., Garcia, S. and Martin, J. (2000)
Involvement of Fcgamma receptor IIIA genotypes in susceptibility to rheumatoid
arthritis. Arthritis Rheum, 43, 735-739.
46. Suzuki, T., Muraki, Y., Yasukochi, T., Zhang, H., Kori, Y., Wakamatsu, E., Hayashi,
T., Goto, D., Ito, S., Tsutsumi, A. et al. (2005) Immunoglobulin G from anti-glucose-
6-phosphate isomerase antibodies positive patient with rheumatoid arthritis induces
synovitis in cynomolgus monkeys. Autoimmun Rev, 4, 475-478.
47. Edberg, J.C., Langefeld, C.D., Wu, J., Moser, K.L., Kaufman, K.M., Kelly, J., Bansal,
V., Brown, W.M., Salmon, J.E., Rich, S.S. et al. (2002) Genetic linkage and
association of Fcgamma receptor IIIA (CD16A) on chromosome 1q23 with human
systemic lupus erythematosus. Arthritis Rheum, 46, 2132-2140.
48. Aitman, T.J., Dong, R., Vyse, T.J., Norsworthy, P.J., Johnson, M.D., Smith, J.,
Mangion, J., Roberton-Lowe, C., Marshall, A.J., Petretto, E. et al. (2006) Copy
number polymorphism in Fcgr3 predisposes to glomerulonephritis in rats and humans.
Nature, 439, 851-855.
49. Fanciulli, M., Norsworthy, P.J., Petretto, E., Dong, R., Harper, L., Kamesh, L.,
Heward, J.M., Gough, S.C., de Smith, A., Blakemore, A.I. et al. (2007) FCGR3B copy
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

21
number variation is associated with susceptibility to systemic, but not organ-specific,
autoimmunity. Nat Genet, 39, 721-723.
50. Herold, K.C., Hagopian, W., Auger, J.A., Poumian-Ruiz, E., Taylor, L., Donaldson,
D., Gitelman, S.E., Harlan, D.M., Xu, D., Zivin, R.A. et al. (2002) Anti-CD3
monoclonal antibody in new-onset type 1 diabetes mellitus. N Engl J Med, 346, 1692-
1698.
51. Herold, K.C., Gitelman, S.E., Masharani, U., Hagopian, W., Bisikirska, B.,
Donaldson, D., Rother, K., Diamond, B., Harlan, D.M. and Bluestone, J.A. (2005) A
single course of anti-CD3 monoclonal antibody hOKT3gamma1(Ala-Ala) results in
improvement in C-peptide responses and clinical parameters for at least 2 years after
onset of type 1 diabetes. Diabetes, 54, 1763-1769.
52. Routledge, E.G., Falconer, M.E., Pope, H., Lloyd, I.S. and Waldmann, H. (1995) The
effect of aglycosylation on the immunogenicity of a humanized therapeutic CD3
monoclonal antibody. Transplantation, 60, 847-853.
53. Alegre, M.L., Tso, J.Y., Sattar, H.A., Smith, J., Desalle, F., Cole, M. and Bluestone,
J.A. (1995) An anti-murine CD3 monoclonal antibody with a low affinity for Fc
gamma receptors suppresses transplantation responses while minimizing acute toxicity
and immunogenicity. J Immunol, 155, 1544-1555.
54. Tax, W.J., Frenken, L.A., Glaudemans, C.A., Tamboer, W.P. and Koene, R.A. (1995)
Polymorphism of Fc receptor (Fc gamma RII) is reflected in cytokine release and
adverse effects of mIgG1 anti-CD3/TCR antibody during rejection treatment after
renal transplantation. Transplant Proc, 27, 867-868.
55. Lin, T.S., Flinn, I.W., Modali, R., Lehman, T.A., Webb, J., Waymer, S., Moran, M.E.,
Lucas, M.S., Farag, S.S. and Byrd, J.C. (2005) FCGR3A and FCGR2A
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

22
polymorphisms may not correlate with response to alemtuzumab in chronic
lymphocytic leukemia. Blood, 105, 289-291.
56. Farag, S.S., Flinn, I.W., Modali, R., Lehman, T.A., Young, D. and Byrd, J.C. (2004)
Fc gamma RIIIa and Fc gamma RIIa polymorphisms do not predict response to
rituximab in B-cell chronic lymphocytic leukemia. Blood, 103, 1472-1474.
57. Anolik, J.H., Campbell, D., Felgar, R.E., Young, F., Sanz, I., Rosenblatt, J. and
Looney, R.J. (2003) The relationship of FcgammaRIIIa genotype to degree of B cell
depletion by rituximab in the treatment of systemic lupus erythematosus. Arthritis
Rheum, 48, 455-459.
58. The United European Gastroenterolog (2001) When is a coeliac a coeliac? Report of a
working group of the United European Gastroenterology Week in Amsterdam, 2001.
Eur J Gastroenterol Hepatol, 13, 1123-1128.
59. Welsing, P.M. and van Riel, P.L. (2004) The Nijmegen inception cohort of early
rheumatoid arthritis. J Rheumatol, 69, 14-21.
60. Arnett, F.C., Edworthy, S.M., Bloch, D.A., McShane, D.J., Fries, J.F., Cooper, N.S.,
Healey, L.A., Kaplan, S.R., Liang, M.H., Luthra, H.S. et al. (1988) The American
Rheumatism Association 1987 revised criteria for the classification of rheumatoid
arthritis. Arthritis Rheum, 31, 315-324.
61. Hughes, L.B., Criswell, L.A., Beasley, T.M., Edberg, J.C., Kimberly, R.P., Moreland,
L.W., Seldin, M.F. and Bridges, S.L. (2004) Genetic risk factors for infection in
patients with early rheumatoid arthritis. Genes Immun, 5, 641-647.
62. Brun, J.G., Madland, T.M. and Vedeler, C.A. (2002) Immunoglobulin G fc-receptor
(FcgammaR) IIA, IIIA, and IIIB polymorphisms related to disease severity in
rheumatoid arthritis. J Rheumatol, 29, 1135-1140.
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

23
63. Stewart-Akers, A.M., Cunningham, A., Wasko, M.C. and Morel, P.A. (2004) Fc
gamma R expression on NK cells influences disease severity in rheumatoid arthritis.
Genes Immun, 5, 521-529.
64. Chiano, M.N. and Clayton, D.G. (1998) Fine genetic mapping using haplotype
analysis and the missing data problem. Ann Hum Genet, 62, 55-60.
65. Dudbridge, F. (2003) Pedigree disequilibrium tests for multilocus haplotypes. Genet
Epidemiol, 25, 115-121.
66. Cordell, H.J. and Clayton, D.G. (2002) A unified stepwise regression procedure for
evaluating the relative effects of polymorphisms within a gene using case/control or
family data: application to HLA in type 1 diabetes. Am J Hum Genet, 70, 124-141.
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from

24
LEGENDS FOR TABLES
Table 1.
The association of the functional FcgammaRIIaA519G and FcgammaRIIIaT559G variants to
type 1 diabetes, celiac disease and rheumatoid arthritis
Table 2.
LD between FcgammaRIIa*A519G_FcgammaRIIIa*A559C and their joint association to type
1 diabetes (T1D), celiac disease (CD) and rheumatoid arthritis (RA)
Figure 1,
A meta-analysis of the frequency of FcgammaR*559CC and FcgammaR*559CA genotypes
compared to that of FcgammaR*559AA in RA patients versus controls. OR represents odds
ratio with 95 percent confidence interval estimated using random model (Random). n1
represents number of subjects with CC genotypes; N1, number of sum of subjects with CC
and AA genotypes, n2, number of subjects with CA genotype, and N2 represents sum of the
number of subjects CA and AA genotypes. Vertical grey line serves reference line for no
association i.e. OR=1.
by guest on February 21, 2016http://hm
g.oxfordjournals.org/D
ownloaded from
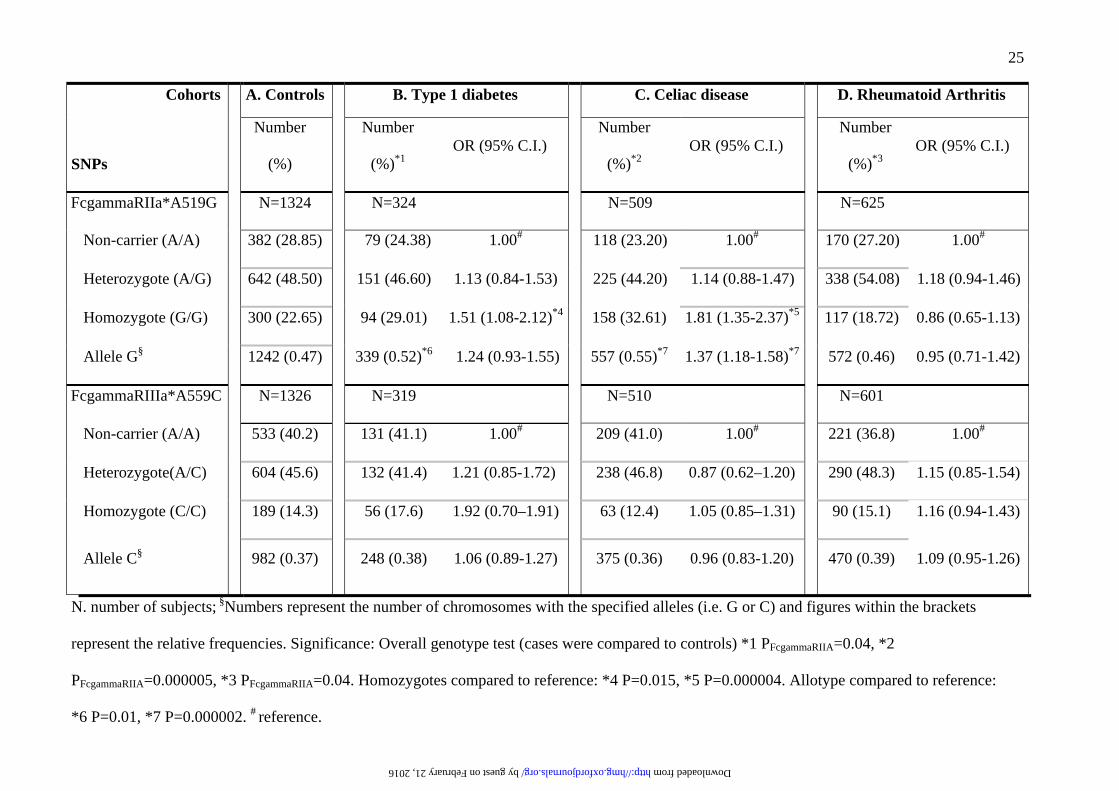
25
Cohorts A. Controls B. Type 1 diabetes C. Celiac disease D. Rheumatoid Arthritis
SNPs
Number
(%)
Number
(%)*1OR (95% C.I.)
Number
(%)*2OR (95% C.I.)
Number
(%)*3OR (95% C.I.)
FcgammaRIIa*A519G N=1324 N=324 N=509 N=625
Non-carrier (A/A) 382 (28.85) 079 (24.38) 1.00# 118 (23.20) 1.00# 170 (27.20) 1.00#
Heterozygote (A/G) 642 (48.50) 151 (46.60) 1.13 (0.84-1.53) 225 (44.20) 1.14 (0.88-1.47) 338 (54.08) 1.18 (0.94-1.46)
Homozygote (G/G) 300 (22.65) 94 (29.01) 1.51 (1.08-2.12)*4 158 (32.61) 1.81 (1.35-2.37)*5 117 (18.72) 0.86 (0.65-1.13)
Allele G§ 1242 (0.47) 339 (0.52)*6 1.24 (0.93-1.55) 557 (0.55)*7 1.37 (1.18-1.58)*7 572 (0.46) 0.95 (0.71-1.42)
FcgammaRIIIa*A559C N=1326 N=319 N=510 N=601
Non-carrier (A/A) 533 (40.2) 131 (41.1) 1.00# 209 (41.0) 1.00# 221 (36.8) 1.00#
Heterozygote(A/C) 604 (45.6) 132 (41.4) 1.21 (0.85-1.72). 238 (46.8) 0.87 (0.62–1.20) 290 (48.3) 1.15 (0.85-1.54)
Homozygote (C/C) 189 (14.3) 56 (17.6) 1.92 (0.70–1.91) 63 (12.4) 1.05 (0.85–1.31) 90 (15.1) 1.16 (0.94-1.43)
Allele C§ 982 (0.37) 248 (0.38) 1.06 (0.89-1.27) 375 (0.36) 0.96 (0.83-1.20) 470 (0.39) 1.09 (0.95-1.26)
N. number of subjects; §Numbers represent the number of chromosomes with the specified alleles (i.e. G or C) and figures within the brackets
represent the relative frequencies. Significance: Overall genotype test (cases were compared to controls) *1 PFcgammaRIIA=0.04, *2
PFcgammaRIIA=0.000005, *3 PFcgammaRIIA=0.04. Homozygotes compared to reference: *4 P=0.015, *5 P=0.000004. Allotype compared to reference:
*6 P=0.01, *7 P=0.000002. # reference.
by guest on February 21, 2016 http://hmg.oxfordjournals.org/ Downloaded from

26
Controls
(N=2396)
T1D
(N=628)
CD
(N=924)
RA
(N=1196)
FcgRIIa
*A519G
FcgIIIa
*A559C N (freq.) N (freq.) OR (95%CI) N (freq.) OR (95%CI) N (freq.) OR (95%CI)
A A 790 (0.31) 141 (0.22) 1.00# 227 (0.25) 1.00# 338 (0.28) 1.00#
A C 582 (0.23) 155 (0.25) 1.19 (0.89–1.49) 188 (0.20) 0.89 (0.67-1.33) 312 (0.26) 1.05 (0.75-1,25)
G A 836 (0.32) 248 (0.39) 1.36 (1.03-1.71) 368 (0.40) 1.22 (0.92-1.53) 391 (0.33) 0.92 (0.69-1.38)
G C 370 (0.14) 84 (0.13) 1.43 (1.07-1.79) 139 (0.15) 1.49 (1.12-1.87) 155 (0.13) 1.10 (0.83-1.38)
Overall statistics for disease association:
chi2 value (df); p value. 8.88 (3), P=0.03 19.94 (3); 0.00017 1.93 (3); 0.59
Analysis of LD between the variants
D’ (95% confidence
bound);
R-squared
0.33 (0.27-0.43);0.06 0.21 (0.12-0.30); 0.08 0.28 (0.17-0.37); 0.03
N: presents the number of chromosomes included in the analysis; D’ coefficient of pair-wise linkage disequilibrium; LR: likelihood ratio; df: degree of
freedom; LD: linkage disequilibrium.
0.28 (0.17-0.37); 0.04
by guest on February 21, 2016 http://hmg.oxfordjournals.org/ Downloaded from

27
FcγRIIIA*559CA vs. AA FcγRIIIA*559CC vs. AA
1817/3608
261/752
077/173
149/354
091/179
1201/2276
056/122
058/098
025/061
364/720
063/120
188/353
051/095
085/155
290/511
045/102
Controlsn2 / N2
317/70675/464080/471Total
Asians170/325033/121046/201Chen _Taiwan (43)
046/105010/050008/067Mprgan _India (26)
042/093023/089023/089Akres _USA (63)
1456/2856366/1442377/1784Total
132/277028/233026/171Kyogoku _Japan (23)
059/150014/110008/099Matsumoto _Japan (30)
596/1128090/311189/728This Study _Dutch
161/329026/09633/201Kastbom _Sweden (25)
032/075017/061014/057 Brun _Norway (62)
214/386048/213034/206Milicic _UK (27)
048/113021/078011/076Morgan _UK (26)
1518/2982441/1906457/2255Total
035/079002/038014/044Milicic _India (27)
241/505116/472056/320Morgan _UK (42)
076/122015/072020/066Nieto_Spanish (45)
Caucasians
Patientsn2 / N2
Patientsn1 / N1
Controlsn1 / N1Studies
FcγRIIIA*559CA vs. AA FcγRIIIA*559CC vs. AA
1817/3608
261/752
077/173
149/354
091/179
1201/2276
056/122
058/098
025/061
364/720
063/120
188/353
051/095
085/155
290/511
045/102
Controlsn2 / N2
317/70675/464080/471Total
Asians170/325033/121046/201Chen _Taiwan (43)
046/105010/050008/067Mprgan _India (26)
042/093023/089023/089Akres _USA (63)
1456/2856366/1442377/1784Total
132/277028/233026/171Kyogoku _Japan (23)
059/150014/110008/099Matsumoto _Japan (30)
596/1128090/311189/728This Study _Dutch
161/329026/09633/201Kastbom _Sweden (25)
032/075017/061014/057 Brun _Norway (62)
214/386048/213034/206Milicic _UK (27)
048/113021/078011/076Morgan _UK (26)
1518/2982441/1906457/2255Total
035/079002/038014/044Milicic _India (27)
241/505116/472056/320Morgan _UK (42)
076/122015/072020/066Nieto_Spanish (45)
Caucasians
Patientsn2 / N2
Patientsn1 / N1
Controlsn1 / N1Studies
*P=0.001
Not included
by guest on February 21, 2016 http://hmg.oxfordjournals.org/ Downloaded from
Top Related
Copyright © 2022 FDOKUMEN

