







Bahasa
Halaman
Hukum


0024-3
doi:10
* C
E-m
www.elsevier.com/locate/lifescie
Life Sciences 74 (2003) 663–673
Aniseed oil increases glucose absorption and reduces urine
output in the rat
Sawsan Ibrahim Kreydiyyeha,*, Julnar Ustab, Khuzama Knioa,Sarine Markossiana, Shawky Daghera
aDepartment of Biology, Faculty of Arts and Sciences, American University of Beirut, Beirut, LebanonbDepartment of Biochemistry, Faculty of Medicine, American University of Beirut, Beirut, Lebanon
Received 20 March 2003; accepted 18 July 2003
Abstract
Anise (Pimpinella anisum) has been used as a traditional aromatic herb in many drinks and baked foods because
of the presence of volatile oils in its fruits commonly known as seeds. Hot water extracts of the seeds have been
used also in folk medicine for their diuretic and laxative effect, expectorant and anti-spasmodic action, and their
ability to ease intestinal colic and flatulence. The aim of this work was to study the effect of aniseed oil on
transport processes through intestinal and renal epithelia and determine its mechanism of action. The essential oils
were extracted from the seeds by hydrodistillation and analyzed by gas chromatography. Aniseed oil enhanced
significantly glucose absorption from the rat jejunum and increased the Na+-K+ ATPase activity in a jejunal
homogenate in a dose dependent manner. The oil, however, exerted no effect on water absorption from the colon
and did not alter the activity of the colonic Na+-K+ ATPase. When added to drinking water, it reduced the volume
of urine produced in the rat and increased the activity of the renal Na+-K+ ATPase even at extremely low
concentrations. It was concluded that aniseed oil increases glucose absorption by increasing the activity of the Na+-
K+ ATPase and consequently the sodium gradient needed for the sugar transport. Its anti-diuretic effect is also
mediated through a similar mechanism in the kidney whereby a stimulation of the Na+-K+ pump increases tubular
sodium reabsorption and osmotic water movement. The colonic Na+-K+ ATPase was however, resistant to the oil.
D 2003 Elsevier Inc. All rights reserved.
Keywords: Aniseed; Glucose; Na+-K+ ATPase; Antidiuretics
205/$ - see front matter D 2003 Elsevier Inc. All rights reserved.
.1016/j.lfs.2003.07.013
orresponding author. Fax: +961-1744461.
ail address: [email protected] (S.I. Kreydiyyeh).

S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673664
Introduction
Traditional herbal medicine is gaining recently more popularity and phytotherapy has become a
more acceptable scientific approach to disease treatment and prevention. Plant extracts are
considered nowadays, as potential bioactive agents that can interfere and alter positively or
negatively different cellular processes. In most instances, volatile oils constitute the most common
active ingredients in medicinal herbs. Anise (Pimpinella anisum) is a plant that produces fruits
commonly referred to as seeds which are rich in such oils. The oils are used in perfumery,
medicine, soaps and beverages (Duke, 1985). Aniseed and aniseed oils are employed also in food
industry as flavoring agents (e.g., Greek Ouzo, Lebanese arak, anisette) and a concoction of seeds
in hot water is used as a carminative, antiseptic, diuretic, digestive, and a folk remedy to insomnia
and constipation (Bisset, 1994). Most of these reported biological effects have rarely been supported
by any scientific background and rely mainly on empirical information. Because aniseeds are highly
used in Lebanon in baked goods and beverages, a study of their physiological properties was
deemed necessary and informative.
This work aims at studying, in the rat, the effect of aniseed oil on the absorption of glucose from the
jejunum, and water from the colon and kidney tubules.
Materials and methods
Isolation of the essential oils
The fruits of the anise plant(Pimpinella anisum), commonly known and thereafter referred to as
aniseeds, were obtained from the local market and allowed to dry at room temperature for 24 hours. The
essential oils were extracted by hydrodistillation using a ratio of 1:6 (g/ml) of seed weight to volume of
distilled water.
The distillation process was carried out over a period of 6 hours using a round bottom container, and a
receiver for oils lighter than water to collect the essential oils. The collected oil was measured and dried
over anhydrous sodium sulfate.
Gas chromatography/mass spectroscopy
Essential oil composition was analyzed using a HP6890 gas chromatograph hooked to HP 5972 mass
selective detector. A fused silica column loaded with 5% diphenyl 95% dimethylpoly siloxane was used.
Column dimensions were: length 30 m, i.d. 0.25 mm. The operating conditions were as follows: linear
velocity 36 cm/sec; split ratio 1:100; ionization voltage 70 eV; ionization current 50 AA; scanning speed
30–500 amus/sec; volume of injection 1 AL of pure oil.
Components of the essential oil were screened using the libraries: Wiley 275 and NBS.
Animal treatments
Male Sprague-Dawley rats (Rattus norvegicus) weighing 150–200 g were used and handled all
through in accordance with the Guide for Laboratory Animal Facilities and Care, US Department of
Health, Education and Welfare.

S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673 665
Measurement of 14C glucose absorption by the upper jejunum
Rats were anesthetized by i.p. injection of pentobarbital (5 mg/100 g body weight) and laid dorsally
on a heating pad maintained at 37 jC throughout the experiment. The abdomen was opened to expose
the intestines and a 10 cm segment of the upper jejunum situated 15 cm from the pylorus was isolated
keeping the mesenteries and vasculature intact. An ‘‘L-shaped’’ inlet and outlet catheter were
introduced at each end of the segment and in the same plane of the segment, to allow for buffer
infusion and collection. The perfusion of the segment followed a single pass method and the perfusion
rate was maintained all through at 0.8 ml/min. The intestinal segment was perfused first for 15 min
with oxygenated (95% O2/5%CO2) Krebs improved Ringer buffer pH 7.5 (KIRB) (123.3 mM NaCl;
6.17 mM KCl; 3.29 mMCaCl2�7H2O; 0.78 mM MgSO4�7H2O; 32.14 mM NaHCO3; 1.54 mM
KH2PO4; 6.4 mM sodium pyruvate; 6.4 mM sodium glutamate; 7 mM sodium fumarate; 11.1 mM
glucose) to clear it from wastes. In a second step the jejunum was perfused for 30 min at the same
rate, with KIRB to which 180 AM D[U-14C]glucose (9.9 GBq/mmol; Sigma) was added.The effluent
buffer was collected at different time intervals and its volume measured and compared to the volume
of buffer infused during the same period. When the effect of aniseed oil was studied it was dissolved
in DMSO and added to the buffer during both perfusion steps at a concentration of 5 � 10� 4 Al oil/Al buffer. The oil was added to the first perfusion buffer to give it enough time to exert its effect, if
such an effect is present, before measurements of glucose absorption are started. The same volume of
the vehicle was added in the control treatment. At the end of the experiment the perfused intestinal
segment was isolated, cut longitudinally and opened to expose the mucosal surface. The width and
length of the obtained rectangle were measured, and its surface area calculated. Samples were
withdrawn from the effluent buffers collected at different time intervals during the second perfusion
period and assayed for radioactivity together with samples from the infused buffer. From the
radioactivity data obtained and the measured volumes of the effluent and infused buffers the total
radioactivity entering and leaving the intestine during each interval were computed. Absorption of
glucose was calculated as the difference between the total radioactivity infused and the total
radioactivity appearing in the collected buffer divided by the area of the segment perfused.
Measurement of luminal water movements in the rat colon
The effect of the oil on water movements in the colon was studied on another group of animals. The
abdomen of anesthetized rats was opened through a mid line incision to expose this time the colon. The
whole colon was perfused in a two-step process, and as described before for the jejunum, with KIRB.
The volume of the effluent buffer collected during the second perfusion step was measured and
compared to the volume of infused buffer. Anise oil when tested was added to the two perfusion steps at
the same concentration as for the jejunum (0.5 ml/L). All buffers used were oxygenated (95%O2,
5%CO2) at 37 jC and their pH adjusted to 7.4.
At the end of the second perfusion, the colon was excised, cut longitudinally, its width and length
measured, and its surface area determined. Water movements during the second 30-min perfusion step
were calculated as the difference in volume between the infused and collected buffers divided by the
surface area of the perfused colon.
Measurement of the volume of urine eliminated
Rats of similar weights were housed in metabolic cages and after an overnight fast were
allowed free access to water and food. The volume of water consumed and the volume of urine

S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673666
produced over 24 hours were measured. The animals were then offered water to which aniseed oil
dissolved in DMSO was added at a concentration of 0.5 ml oil/L water. Water intake and urine
production were measured as before. Thus every animal served as it own control. In the control
treatment a similar volume of DMSO was added to the drinking water. The ratio of urine
produced/water intake was calculated and considered as a standardized indicator of urine
elimination.
In vitro assay of the Na+-K+ ATPase activity In a homogenate of the jejunum and colon
The whole colon and an upper jejunal segment were excised from three rats that have not
received any treatment. The segments were opened longitudinally and washed well with KIRB. The
mucosal surface was then scraped with a glass slide and the scrapings were suspended in a buffer
with the following composition (200 mM NaCl; 5 mM MgCl2�6H2O; 2 mM EGTA; 5 mM KCL;
200 mM Tris-HCl, pH 7.4), and homogenized in a glass-Teflon homogenizer with 20 strokes at
2100 rpm (Arthur Thomas Scientific Apparatus, Philadelphia,PA). Samples from the homogenate
were diluted with the same buffer to a concentration of 2 mg protein/ml and used to assay for the
Na+-K+ ATPase activity. After a 30-min incubation period with saponin (0.02% final concentration)
at room temperature, samples (50 Al) were pre-incubated at 37 jC for 10 min in presence or
absence of ouabain (4 mM final concentration). The reaction was then initiated by addition of ATP
to a final concentration of 1.25 mM and terminated after a one hour incubation at 37 jC by
addition of trichloroacetic acid (200 Al, 11%). The samples were centrifuged at 3000 g for 5 min
and the amount of inorganic phosphate liberated in the supernatant was measured colorimetrically
according to the method of Taussky and Shorr (Taussky and Shorr, 1953). Since ouabain is a
specific inhibitor of the Na+-K+ ATPase, the activity of the enzyme was determined by measuring
the ouabain-inhibitable inorganic phosphate released.
Results are reported as percentage of the enzymatic activity in control, as follows:
PiðoilÞ � Piðoil þ ouabainÞPiðcontrolÞ � Piðcontrol þ ouabainÞ � 100
In vitro assay of the Na+-K+ ATPase in kidney homogenate
The abdomen of anesthetized rats was opened through a lateral incision and the intestine and
stomach were pushed aside to expose the left kidney. A heparinized catheter was then introduced
in the left renal vein to allow for drainage, and ligated to fix it in place. Another heparinized
catheter was introduced in the aorta just above the level of the left renal artery to allow for the
infusion of the buffer. The mesenteric artery and the right renal artery were both ligated. The left
kidney was perfused with gassed Krebs improved Ringer buffer (KIRB) for 10 min at a rate of 5
ml/min to clear it from blood and then excised, its capsule removed, cut into small pieces and
homogenized in KIRB buffer in a glass-Teflon homogenizer with 20 strokes at 2100 rpm (Arthur
Thomas Scientific Apparatus, Philadelphia, PA). Its protein content was determined using the Bio-
Rad protein assay (Bio-Rad Laoratories, 2000 Alfred Nobel Drive, Hercules, CA 94547, USA) and
samples were taken to assay for the Na+-K+ ATPase activity which was conducted as described
above.

S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673 667
Statistical analysis
Results are reported as means F SEM. Statistical significance between ratios of water intake/urine
volume was tested by a paired Student t test and by a one-way analysis of variance followed by a Tukey-
Kramer multiple comparisons test for all remaining values.
Results
Essential oil composition
The extraction procedure resulted in a yield of 1.4 ml oil/100 g dry sample. Relative percentage
composition of essential oils extracted from aniseeds appears in Table 1. From the compounds
identified in aniseed oil, trans-anethole (76.7%) was found to be the most abundant followed by
anisalacetone (7.1%), estragole (6.1%) and anisaldehyde (1.5%). Linalool, germacrene-d, ar-
curcumene and h-bisabolene were the least abundant and were all present in the same amount
(0.2%).
Glucose absorption
Aniseed oil added to the perfusion buffer at a final concentration of 0.05% increased significantly
(Fig. 1) jejunal glucose absorption during a 15 (P < 0.002), 25 (P < 0.002) and 30 (P < 0.003) min.
perfusion period.
Effect of aniseed oil on the volume of water reabsorbed from the colon
Aniseed oil at a 0.05% concentration did not exert any significant effect on the volume of water
absorbed from the colon (Fig. 2).
Table 1
Relative percentage composition of essential oils isolated from aniseeds
Name RT(min) CAS number %F SEM
Linalool 13.55 000078-70-6 0.2 F 0.049
Estragole 16.92 000140-67-0 6.1 F 2.001
Anisaldehyde 18.92 000123-11-5 1.5 F 0.601
Trans-anethole 19.87 004180-23-8 76.7 F 12.79
Anisalacetone 23.12 000122-84-9 7.1 F 6.59
Germacrene-d 26.00 023986-74-5 0.2 F 0.049
Ar-curcumene 26.15 000644-30-4 0.2 F 0.098
h-bisabolene 26.86 000495-61-4 0.2 F 0.049
Identified 92.1
Yield (ml oil/100 g dry sample) 1.4
Values are Means F SEM of 3 observations.
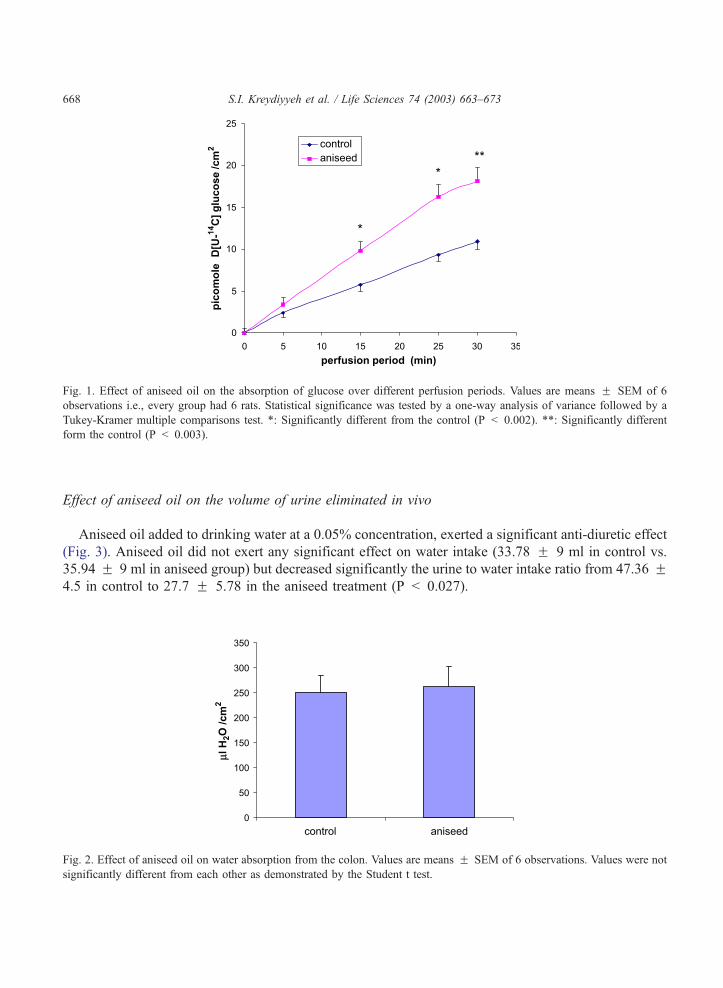
Fig. 1. Effect of aniseed oil on the absorption of glucose over different perfusion periods. Values are means F SEM of 6
observations i.e., every group had 6 rats. Statistical significance was tested by a one-way analysis of variance followed by a
Tukey-Kramer multiple comparisons test. *: Significantly different from the control (P < 0.002). **: Significantly different
form the control (P < 0.003).
S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673668
Effect of aniseed oil on the volume of urine eliminated in vivo
Aniseed oil added to drinking water at a 0.05% concentration, exerted a significant anti-diuretic effect
(Fig. 3). Aniseed oil did not exert any significant effect on water intake (33.78 F 9 ml in control vs.
35.94 F 9 ml in aniseed group) but decreased significantly the urine to water intake ratio from 47.36 F4.5 in control to 27.7 F 5.78 in the aniseed treatment (P < 0.027).
Fig. 2. Effect of aniseed oil on water absorption from the colon. Values are means F SEM of 6 observations. Values were not
significantly different from each other as demonstrated by the Student t test.

Fig. 5. Effect of aniseed oil on the Na+-K+ ATPase activity in colonic homogenate. Values are means F SEM of three
observations. No significant differences were detected between the different values as demonstrated by the Tukey-Kramer
multiple comparisons test.
Fig. 3. Urine/water intake in control rats and rats given aniseed oil. Values are means F SEM. Every group had 5 rats and every
animal served as its own control. *: Significantly different from the control (P < 0.027) as demonstrated by the Student t-test.
Fig. 4. Effect of different concentrations of aniseed oil on the Na+-K+ ATPase activity in jejunal homogenates. All samples
contained 0.8 mg protein. Statistical difference between the different values was tested by a one-way analysis of variance
followed by a Tukey-Kramer multiple comparisons test. *: Significantly different from the control (P < 0.01). **: Significantly
different from the control (P < 0.05).
S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673 669
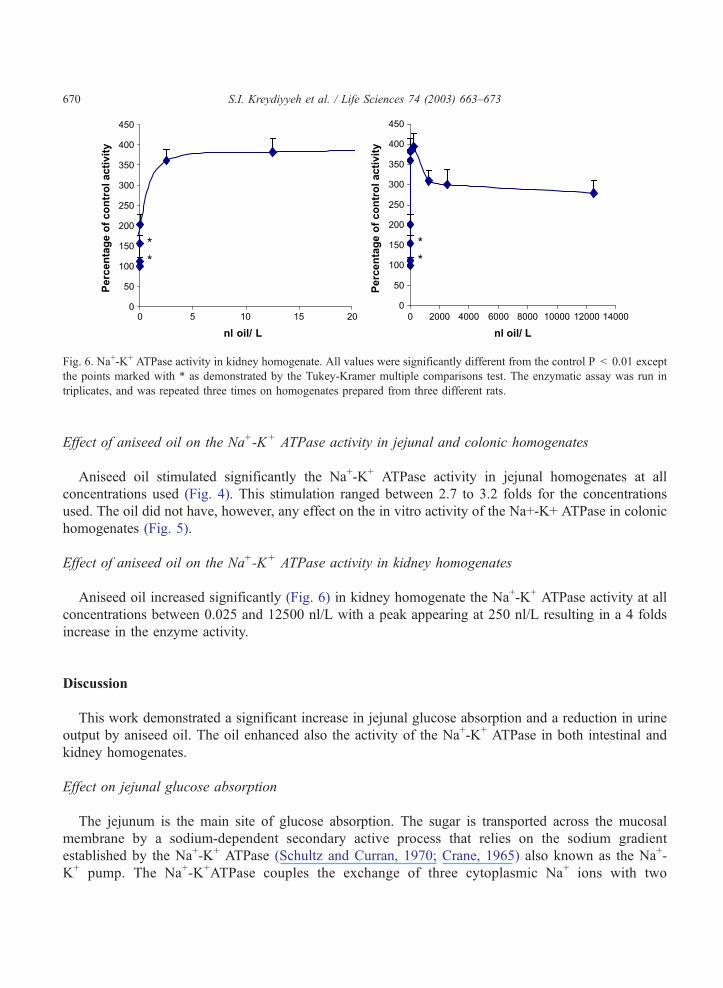
Fig. 6. Na+-K+ ATPase activity in kidney homogenate. All values were significantly different from the control P < 0.01 except
the points marked with * as demonstrated by the Tukey-Kramer multiple comparisons test. The enzymatic assay was run in
triplicates, and was repeated three times on homogenates prepared from three different rats.
S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673670
Effect of aniseed oil on the Na+-K+ ATPase activity in jejunal and colonic homogenates
Aniseed oil stimulated significantly the Na+-K+ ATPase activity in jejunal homogenates at all
concentrations used (Fig. 4). This stimulation ranged between 2.7 to 3.2 folds for the concentrations
used. The oil did not have, however, any effect on the in vitro activity of the Na+-K+ ATPase in colonic
homogenates (Fig. 5).
Effect of aniseed oil on the Na+-K+ ATPase activity in kidney homogenates
Aniseed oil increased significantly (Fig. 6) in kidney homogenate the Na+-K+ ATPase activity at all
concentrations between 0.025 and 12500 nl/L with a peak appearing at 250 nl/L resulting in a 4 folds
increase in the enzyme activity.
Discussion
This work demonstrated a significant increase in jejunal glucose absorption and a reduction in urine
output by aniseed oil. The oil enhanced also the activity of the Na+-K+ ATPase in both intestinal and
kidney homogenates.
Effect on jejunal glucose absorption
The jejunum is the main site of glucose absorption. The sugar is transported across the mucosal
membrane by a sodium-dependent secondary active process that relies on the sodium gradient
established by the Na+-K+ ATPase (Schultz and Curran, 1970; Crane, 1965) also known as the Na+-
K+ pump. The Na+-K+ATPase couples the exchange of three cytoplasmic Na+ ions with two

S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673 671
extracellular K+ ions, to the hydrolysis of one molecule of ATP (Berribi-Bertran et al., 2001) resulting in
the establishment of an electrochemical gradient across the plasma membrane. It has been estimated that
the pump consumes around 25% of the energy spent by a cell (Lingrel and Kuntzweiler, 1994). The
sodium gradient created by the pump provides the driving force for several secondary active transport
processes like the one responsible for the absorption of glucose from the lumen of the intestine and
kidney tubules.
Any change in the activity of the Na+-K+ ATPase is expected to increase or decrease the sodium
gradient and consequently alter glucose absorption.
Aniseed oil increased significantly glucose absorption in the rat jejunum. To see if this is a
consequence of an increase in the Na+-K+ ATPase activity, the direct in vitro effect of the oil on the
enzyme activity in jejunal homogenates was studied. Aniseed oil increased in a dose-dependent manner,
the ATPase activity, and the effect appeared at a concentration as low as 0.025%.
The data suggest that the observed increase in the intestinal absorption of glucose with aniseed oil is
mediated through a stimulation of the Na+-K+ ATPase which increases the sodium gradient that gears the
mucosal glucose transport.
Effect on water absorption from the colon
One of the aims of this work was to demonstrate the laxative effect of aniseed oil if present.
A laxative agent may decrease net water absorption from the colon by either decreasing electrolyte
absorption, or increasing electrolyte secretion, or both. These absorptive and secretory processes are
regulated by different carriers and result from the transport of different ions (Halm and Frizzel, 1991).
The mammalian colon absorbs sodium and chloride, and secretes potassium and bicarbonate (Binder and
Sandle, 1994). The neutral NaCl mucosal transport is the predominant sodium absorptive mechanism in
the rat (Binder et al., 1987; Zimmerman and Binder, 1984) and involves the dual ion exchangers Na/H
and Cl/HCO3 which are coupled by intracellular pH (Goldfarb et al., 1988) and energized by the sodium
gradient created by the Na+-K+ ATPase (Binder and Sandle, 1994). Active potassium secretion is geared
also by the Na+-K+ pump (Binder and Sandle, 1994). The Na+-K+ ATPase is thus the key player in the
regulation of electrolytes and consequently water movements.
Aniseed oil added at a 0.05% concentration did not have any significant effect on colonic water
absorption. This finding was supported further by the absence of any effect of the oil on the in vitro
activity of the Na+-K+ ATPase in homogenates from the colon.
It seems thus that the ATPase in the colon is resistant to the oil effect. The Na+-K+-ATPase is a highly-
conserved integral membrane protein that is expressed in virtually all cells of higher organisms. The
ATPase has two functional subunits: a and h. In mammals, four alpha-subunit and at least three beta-
subunit isoforms have been identified in addition to two gamma-subunits in the kidney. Different
combinations of subunit isoforms are present in different cells (Ewart and Klip, 1995). The alpha subunit
(f 113 kD) is the action hero of the pair. It binds ATP and both sodium and potassium ions, and contains
the phosphorylation site. The smaller beta subunit (f 35 kDa glycoprotein) is absolutely necessary for
the activity of the complex (Lingrel and Kuntzweiler, 1994), and appears to be critical in facilitating
plasma membrane localization and activation of the alpha subunit.
The resistance of the colonic enzyme to the oil effect may be due to the presence in the colon of
different isoform combinations than in the jejunum in which the binding site for the active component is
absent or not accessible.

S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673672
The use of aniseed concoctions as a traditional remedy for constipation is not in contradiction with our
findings since the concoctions may contain some active ingredients extracted with water but not
appearing in the oil.
Effect on urine volume
Aniseed oil added to drinking water caused a significant reduction in the volume of urine produced
and thus exerted an anti-diuretic effect. Water movements in the kidney tubules are usually secondary to
movements of electrolytes. An increase in electrolytes reabsorption would increase the osmotic gradient
that drives water movements in the same direction leading to a decrease in the volume of urine produced.
Sodium is the major electrolyte reabsorbed, and the process is geared by by the Na+-K+ pump. By
extruding sodium from the cytosol, the pump builds up a concentration gradient for sodium across the
luminal membrane that favors its entry to the inside of the cell. Intracellular sodium is then removed by
the Na+-K+ ATPase to the extracellular space and water follows.
Aniseed oil decreased significantly the volume of urine produced and increased the in vitro activity of
the Na+-K+ ATPase in kidney homogenates. It could be concluded that the anti-diuretic effect of the oil is
due to a stimulation of the Na+-K+ pump.
The renal Na+-K+ ATPase as compared to the jejunal one, seems however to be much more sensitive
to the oil. While a two fold stimulation was observed with a concentration of 0.1% in the jejunum, a
concentration of only 0.025 nl/L produced the same effect in the kidney. Here again this may be ascribed
to the presence of different isoforms in the two tissues.
This work has demonstrated a stimulation of the Na+-K+ ATPase activity in the jejunum and kidney
which resulted in an increase in glucose absorption and a decrease in the volume of urine produced. The
oil did not exert however any effect on water absorption from the colon nor on its ATPase activity. The
incorporation of aniseed oil in the diet will thus make a greater portion of ingested glucose available in
blood for use by the cells, and in a hot and dry weather, the oil would help conserve water in the body
and prevent dehydration.
The work did not determine, however, the nature of the active ingredient(s).Trans-anethole constituted
76.7% of the total oils extracted from aniseeds This value is consistent with the 70–90% range reported
in the literature (Bisset, 1994). Since the active ingredient may not necessary be the most abundant one
in the oil mixture, each of the identified components in addition to trans-anethole may be responsible for
the observed physiological effects. Identification of the active ingredient(s) will be conducted in a
follow-up project.
Acknowledgements
This work was supported by a grant from the University Research Board.
References
Berribi-Bertran, I., Robert, P., Camelin, J.C., Bril, A., Souchet, M., 2001. The gamma subunit of (Na+,K+)-ATPase: a
representative example of human single transmembrane protein with a key regulatory role. Cell Mol Biol. 47, 285–296.

S.I. Kreydiyyeh et al. / Life Sciences 74 (2003) 663–673 673
Binder, H.J., Sandle, G.J., 1994. Electrolyte Transport in the Mammalian Colon. In: Johnson, L.R. (Ed.), Physiology of the
Gastrointestinal Tract, 3rd ed., Raven Press, New York, pp. 2133–2171.
Binder, H.J., Foster, E., Budinger, M.E., Hayslett, J.P., 1987. Mechanism of Electroneutral Sodium-Chloride Absorption in
Distal Colon of the Rat. Gastroenterology 93, 449–455.
Bisset, N.G. (Ed.), 1994. Herbal drugs and phytopharmaceuticals, CRC Press, Medpharm, Stuttgart, pp. 73–75.
Crane, R.K., 1965. Na+-dependent transport in the intestine and other animal tissues. Proc Fedn Am. Soc. Exp. Biol. 24,
1000–1006.
Duke, J.A., 1985. Handbook of medicinal Herbs, CRC Press, Boca Raton, pp. 374–375.
Ewart, H.S., Klip, A., 1995. Hormonal regulation of the Na(+)-K(+)-ATPase: mechanisms underlying rapid and sustained
changes in pump activity. Am J Physiol. 269, C295–C311.
Goldfarb, D.S., Egnor, R.W., Charney, A.N., 1988. Effects of acid–base variables on ion transport in rat colon. J Clin Invest 81,
1903–1910.
Halm, D.R., Frizzel, R.A., 1991. Ion transport across the large intestine. Handbook of Physiology. Gastrointestinal System.
Intestinal Absorption and Secretion. Am. J. Physiol. Soc. vol. 4, 257–274 (Bethesda, MD).
Lingrel, J.B., Kuntzweiler, T., 1994. Na(+),K(+)-ATPase. J Biol Chem. 269, 19659–19662.
Schultz, S.G., Curran, P.F., 1970. Coupled transport of sodium and organic solutes. Physiol. Rev. 40, 637–719.
Taussky, H., Shorr, E., 1953. Microcolorimetric method for determination of inorganic phosphorous. J. Biol. Chem. 202,
675–685.
Zimmerman, T.W., Binder, H.J., 1984. Serotonin-induced alteration of colonic. Electrolyte transport in the rat. Gastroenterology
86, 310–317.
Top Related
Copyright © 2022 FDOKUMEN

