







Bahasa
Halaman
Hukum


www.elsevier.com/locate/brainres
2004) 164–173
Brain Research 998 (0006-8
doi:10.
* C
9918.
E-m
Research report
Age-related changes in growth hormone (GH) cells in the pituitary gland
of male mice are mediated by GH-releasing hormone but not by
somatostatin in the hypothalamus
Sachi Kuwaharaa, Dwi Kesuma Saria, Yasuhiro Tsukamotoa, Shin Tanakab, Fumihiko Sasakia,*
aLaboratory of Veterinary Anatomy, Graduate School of Agriculture and Biological Sciences, Osaka Prefecture University, 1-1 Gakuen-cho,
Sakai, Osaka 599-8531, JapanbLaboratory Animal Research Facilities, National Institute for Longevity Sciences, 36-3 Gengo, Morioka, Obu, 474-8522, Japan
Accepted 21 October 2003
Abstract
Using immunocytochemical and morphometric methods, we examine changes with age of growth hormone-releasing hormone (GHRH)
in the arcuate nucleus (ARC), changes of somatostatin (SS) in the periventricular nucleus (PeN) of the hypothalamus, and changes of growth
hormone (GH) cells in the anterior pituitary in male C57BL/6J mice at 2 months old (2 M), 4 M, 12 M and 24 M. The number of GHRH-ir
neurons decreased significantly with age. The number of SS-ir neurons did not differ significantly between these all age groups. The volume
of the anterior pituitary and the number of adenohypophysial parenchymal cells fell dramatically from 4 to 12 M. The proportion of GH-ir
cells decreased significantly with age, and in absolute number from 4 to 12 M and in size from 2 to 4 M and from 4 to 12 M. These results
suggest that the reduction in GH-ir cells in male mice is modulated by the reduction in GHRH-ir neurons, but not by SS-ir neurons.
D 2003 Elsevier B.V. All rights reserved.
Theme: Development and regeneration
Topic: Aging process
Keywords: GH-ir cell; GHRH-ir neuron; SS-ir neuron; Immunocytochemistry; Morphometry
1. Introduction
It is widely acknowledged that secretion of growth
hormone (GH) declines with age in animals [22] and in
humans [3]. Pulsatile release and the mean concentration of
GH in the anterior pituitary are attenuated in male rats with
age [32]. This decrease in GH may be responsible for a loss
of muscle mass, increase in adipose tissue mass, and a
deterioration in several tissue and organ functions [3].
Release of GH from the anterior pituitary is regulated by
at least two hypothalamic neuropeptides: growth hormone-
releasing hormone (GHRH), which stimulates GH synthesis
and release, and somatostatin (SS), which inhibits GH
synthesis and release [10]. GHRH neurons are located in
993/$ - see front matter D 2003 Elsevier B.V. All rights reserved.
1016/j.brainres.2003.10.060
orresponding author. Tel.: +81-72-254-9475; fax: +81-72-254-
ail address: [email protected] (F. Sasaki).
the arcuate nucleus (ARC) of humans [1,2], rats [6,9,24,31]
and mice [25,29,30] and SS neurons are located in the
periventricular nucleus (PeN) of rats [9,12,14,19] and mice
[25]. They directly secrete each neuropeptide to the median
eminence (ME) so as to modulate GH secretion. The age-
related decline in GH secretion may therefore be responsible
for neuropeptides from GHRH and SS neurons. It has been
suggested that the age-related decline of GH secretion is a
result of more reduced responsiveness of GH cells to GHRH
[3,32] than any defect in the mechanisms of GH release
[35]. However, the contribution of SS to the reduced
secretion of GH from GH cells remains controversial
[11,21,32,34]. We have already reported that pituitary GH
immunoreactive (-ir) cells [27,30] and ARC GHRH-ir
neurons and PeN SS-ir neurons [25] mature in differently
during postnatal development. It is important to study the
hypothalamo (GHRH/SS)-pituitary (GH)-somatic axis in
mature animals in order to understand the physiological
mechanism of aging; we believe that this has not been

S. Kuwahara et al. / Brain Research 998 (2004) 164–173 165
studied in detail. However, there is little information about
age-related changes in hypothalamic ARC GHRH neurons
and PeN SS neurons [21]. An immunocytochemical and
morphometric study was therefore conducted to examine
age-related changes in ARC GHRH neurons, PeN SS
neurons and pituitary GH cells.
2. Materials and methods
2.1. Animals
Adult male C57BL/6J mice were obtained from the
National Institutes on Aging (NIA, MD, USA). These mice
were in four age groups: 2 months old (2 M), 4 months old
(4 M), 12 months old (12 M) and 24 months old (24 M).
Five mice from each group were anaesthetized by ether
inhalation, and killed in the morning between 1000 and
1200 h. The body weight in 2, 4, 12 and 24 M mice was
30.31F 0.86 (meanF S.E.), 30.79F 1.22, 29.98F 1.12
and 33.41F 0.92 g, respectively [the body weight in 24
M mice was significantly heavier (F = 2.26, P < 0.05) than
that in 2 and 12 M mice].
After immersion in Bouin’s solution for 30 min, the
hypothalamus was taken from the brain as follows (Fig. 1):
their brains were cut frontally at the anterior part of the optic
chiasma (a) and at the anterior part of the cerebellum (b), cut
longitudinally at both lateral sides (c,d) of hypothalamus,
Fig. 1. Ventral view of hypothalamus with the frontal plane of the anterior
part of the optic chiasma. Their brains were cut frontally at the anterior part
of the optic chiasma (a) and at the anterior part of the cerebellum (b), cut
longitudinally at both lateral sides (c,d) of hypothalamus, and cut
horizontally at the part of the corpus callosum (e) at the plane of the
anterior part of the optic chiasma. cc: corpus callosum, LV: lateral ventricle,
on: optic nerve, oc: optic chiasma.
and cut horizontally at the part of the corpus callosum (e) at
the plane of the anterior part of the optic chiasma. It was
further placed in the same fixative for 24 h. The pituitary
glands from mice of each age were fixed in 10% neutral
formalin for 24 h. All specimens were dehydrated in a
graded series of ethanol solutions, cleared in xylene, and
embedded in Tissue Prep (Fisher Scientific, Pittsburgh, PA,
USA).
2.2. Experimental procedures
The parts of ARC and PeN in all hypothalami were
frontally cut in serial sections of 10 Am as described
previously [25] and were mounted on poly-L-lysine (50
Ag/ml)-coated slides. In all immunocytochemical proce-
dures, sections were processed by the avidin–biotin–per-
oxidase complex (ABC) method, using either anti-mouse
GHRH rabbit serum (1:5000) or anti-mouse SS rabbit serum
(1:1000), both of which were donated by Dr. Miki (Tokyo
Women’s Medical College) as described previously [25].
The absorption tests for the GHRH antiserum and SS
antiserum are described in Refs. [20,25], respectively.
Sections from the lateral, parasagittal, and midsagittal
parts of the pituitary gland were stained immunocytochem-
ically by ABC method, using anti-rat GH rabbit serum
(1:8000), which was donated by Dr. A. F. Parlow (NIDDK,
National Hormone and Pituitary Program) as described
previously [15]. The specificity of this antiserum was
verified by absorption testing using antigen. The absorp-
tion test for the GH antiserum was performed by adding
100 Ag/ml rGH (Biogenesis, UK) to the corresponding
antiserum.
Proliferating cell nuclear antigen (PCNA) and in situ
apoptosis tests were performed to observe proliferation of
cells and nuclear DNA fragmentation in ARC and PeN
neurons and adenohypophysial cells, respectively, as de-
scribed previously [25]. Furthermore, the in situ method
[antibody against single-stranded DNA (Dako, Tokyo, Ja-
pan)] by ABC method was used to observe apoptic cells of
ARC and PeN neurons and adenohypophysial cells.
2.3. Morphometry
2.3.1. Hypothalamic GHRH-ir and SS-ir neurons
Serial sections of the ARC [total number of sections per
mouse: 71.2F 1.0 (meanF S.E.)] were cut along the ante-
rior–posterior axis of the ARC and selected systematically
at intervals of four sections from all mice. The number of
sections for ARC GHRH-ir neurons selected from each
animal was 18.4F 0.5 for 2 M, 17.6F 0.2 for 4 M,
18.2F 0.6 for 12 M and 17.0F 0.3 for 24 M mice. From
serial sections of the PeN (total number of sections per
mouse: 78.9F 1.1), which were cut along the anterior–
posterior axis of the PeN, sections were selected from each
mouse at intervals of four sections in order to count the PeN
SS-ir neurons. The number of sections obtained per animal

Fig. 2. Number of GHRH-ir neurons in the ARC of male mice versus age.
The number falls significantly with age. Differences were significantly
tested with the previous younger group. *P< 0.05.
S. Kuwahara et al. / Brain Research 998 (2004) 164–173166
was 20.3F 0.9 for 2 M, 19.2F 0.6 for 4 M, 19.4F 0.6 for
12 M and 19.6F 0.7 for 24 M mice.
The total number (T) of immunoreactive cells in the ARC
or PeN was derived by counting the number of neurons
(SNı) in the number of the sections (ı) selected from the
A
VV
C
V
V
Fig. 3. GHRH-ir neurons in the ARC of male mice at 2 M (A), 4 M (B), 12 M (C) a
from 2 to 4 M and only a few GHRH-ir neurons were detected at 12 and 24 M
represents 50 Am (A–D).
number of total sections (n) in the nucleus in each mouse,
and then multiplying by SNı by n/ı according to the
formula: T= n/ı�SNı [25,26].
2.3.2. Adenohypophysial GH-ir cells
To estimate the volume of the anterior pituitary, pituitaries
from 20 mice (5 per group) were sagittally and serially cut 5
Am thick, and sections weremounted on poly-L-lysine (50 Ag/ml)-coated slides. The areas (A1, A2,. . ., An) of the anterior
pituitary were measured in every 10th section with an image
analyzer (Cosmozone, 1SB, Nikon, Tokyo, Japan). The
volume (V) of the anterior pituitary was calculated as
V= 5� 10�SAı Am3.
The proportion, number and size of GH-ir cells were
evaluated as follows. A square of side 40 Am was drawn
at a magnification of � 400 on the screen of a personal
computer. For each animal, about 2000 nuclei of the
adenohypophysial cells in these squares were counted
with a rule in the lateral, parasagittal and midsagittal
parts of the anterior lobe [28]. The nuclei of cells
immunoreactive with GH antiserum were also counted.
The numbers of cells immunoreactive with GH antiserum
B
D
nd 24 M (D). The number of GHRH-ir neurons in the ARC fell dramatically
. GHRH-ir neurons are indicated by arrows. V: third ventricle. Scale bar

Fig. 4. The ME stained for GHRH in male mice at 2 M (A), 4 M (B), 12 M
(C) and 24 M (D). The numbers of immunoreactive fibers were clearly
diminished. V: third ventricle. Scale bar represents 100 Am (A–D).
Fig. 5. Number of SS-ir neurons in PeN of male mice versus age. There is
no significant difference in number between all age groups.
S. Kuwahara et al. / Brain Research 998 (2004) 164–173 167
were expressed as percentages of a total of about 2000
nuclei.
For each animal, the mean number (N) of nuclei of
adenohypophysial cells in these squares (402 Am2) was
calculated, and was transformed into the mean number
(N3/2 Am3) of nuclei of adenohypophysial cells (T) using
the formula: T= N3/2�V/403 [29]. The numbers of GH-ir
cells were then calculated by multiplying this quantity by
the proportion of GH-ir cells.
About 100 cells per mouse with nuclei having entire cells
inside the squares were randomly chosen from each prepara-
tion of cells immunoreactive to GH antiserum, and their areas
were calculated using the image analyzer.
2.4. Statistical analysis
Results were expressed as meanF S.E. for five mice. The
data were analyzed by one-way ANOVA followed by Fish-
er’s least significant difference test, with significant differ-
ences, which were assigned at the 5% probability level.
3. Results
3.1. Number of GHRH-ir neurons in the ARC and
immunocytochemical detection of GHRH in the ME
The number of GHRH-ir neurons [2 M (1023F 50
neurons), 4 M (478F 52 neurons), 12 M (299F 52 neu-
rons) and 24 M (155F 12 neurons)] decreased significantly
(F = 98.71, P < 0.05) with age (Figs. 2 and 3).
The immunoreactive of fibers in the ME were similar in
immunoreactive intensity for 2 and 4 M mice. The intensi-
ties of immunoreactive fibers showed a slight reduction at
12 and 24 M compared with those at 2 and 4 M, but the
numbers of immunoreactive fibers were clearly diminished
(Fig. 4).
3.2. Number of SS-ir neurons in the PeN and immunocy-
tochemical detection of SS in the ME
The number of SS-ir neurons did not differ significantly
between 2 M (2163F 38 neurons), 4 M (2091F 69 neu-
rons), 12 M (2358F 152 neurons) and 24 M (2154F 68
neurons in 24 M) (Figs. 5 and 6).
No clear difference was observed between 2, 4, 12 and
24 M for SS reactivity in the ME (Fig. 7).
3.3. PCNA and apoptosis observation in the ARC and PeN
Proliferation cells or DNA fragmentation cells were not
found in the ARC or PeN at any age.
3.4. Volume of the anterior pituitary
The volume of the anterior lobe increased significantly
(F = 24.68, P < 0.05) from 2 M (0.55F 0.02 mm3) to 4 M
(0.71F 0.06 mm3), but 12 M (0.33F 0.02 mm3) decreased
significantly compared to 4 M. There was no significant
difference in the volume of the anterior lobe between 12
and 24 M (0.38F 0.01 mm3) (Fig. 8A).

A
V
B
V
C
V
D
V
Fig. 6. PeN SS-ir neurons of male mice at 2 M (A), 4 M (B), 12 M (C) and 24 M (D). SS-ir neurons are indicated by arrows. The number of SS-ir neurons do
not differ significantly between all age groups. V: third ventricle. Scale bar represents 50 Am (A–D).
S. Kuwahara et al. / Brain Research 998 (2004) 164–173168
3.5. Number of adenohypophysial cells
The number of adenohypophysial cells did not differ
significantly between 2 M [37.01F1.72 (� 105)] and 4 M
[38.75F 2.16 (� 105)], but it decreased significantly by 12
M [22.20F 0.82 (� 105)] (F = 32.41, P < 0.05) compared
to 4 M mice. The number of adenohypophysial cells did
not differ between 12 and 24 M [23.46F 0.47 (� 105)]
mice (Fig. 8B).
3.6. Proportion of GH-ir cells
The proportion of GH-ir cells fell significantly
(F = 29.30, P < 0.05) with age: 2 M (55.36F 0.70%), 4
M (51.42F 1.11%), 12 M (46.75F 0.86%) and 24 M
(43.36F 1.13%) (Figs. 9A and 10).
3.7. Number of GH-ir cells
The number of GH-ir cells did not differ significantly
between 2 M [20.47 F 0.87 ( � 105)] and 4 M
[19.89F 1.02 (� 105)], but the number of GH-ir cells
decreased significantly (F = 53.26, P < 0.05) between 4
and 12 M [10.39F 0.51 (� 105)]. There was no significant
difference in number between 12 and 24 M [10.19F 0.45
(� 105)] (Figs. 9B and 10).
3.8. Size of GH-ir cells
The size of GH-ir cells decreased significantly (F = 39.57,
P < 0.05) between 2 M (46.39F 0.47 Am2) and 4 M
(40.49F 1.03 Am2), and between 4 and 12 M (37.15F 0.51
Am2). GH-ir cells in 24 M (35.13F 1.01 Am2) mice did not
differ in size from those in the 12 M mice (Figs. 9C and 10).
3.9. PCNA and Apoptosis observation in the anterior
pituitary
Many proliferation cells were detected in the anterior
pituitary in 2 M mice. In 4 M mice, they were also detected,
but in smaller numbers than in 2 M mice. Few proliferation
cells were detected in the 12 and 24 M mice (Fig. 11).
DNA fragmentation cells found abundantly in 2 M mice,
but only a few were detected in the older groups (Fig. 12).
4. Discussion
We have found that the number of ARC GHRH-ir
neurons in male mice decreases with age from 2 to 24 M.
In particular, the number of ARC GHRH-ir neurons in 4 M
mice was approximately half that in 2 M mice, and GHRH
immunoreactivity in the ME was slightly reduced in 12 and
24 M, but the number of immunoreactive fibers is clearly
diminished in 24 M. In elderly (24 M) male rats treated with
colchicines, GHRH-ir neurons in the ARC show no change
in distribution or immunoreactive intensities from those in
young (3 M) rats [21], but GHRH-ir intensities in the ME
[7,21] and hypothalamic GHRH mRNA [7] are markedly
reduced in elderly rats. It is reported that colchicines is
necessary to detect perikaryal immunolabeling for GHRH in
the hypothalamus, but colchicine can itself induce the
expression of genes that are not active under physiological
conditions in a cell [4,23]. In our study, no apoptotic cells
were found in the ARC or PeN neurons at any age
examined, so we believe that GHRH neurons do not
themselves reduce in number even in old age; but only
GHRH neurons that played an active function were immu-
nologically stained. In postnatal development, the number

Fig. 8. The volume of the anterior pituitary (A) and the number of
adenohypophysial cells (B) during aging. The volume of the anterior
pituitary and the number of adenohypophysial cells in 12 M are
significantly smaller than those in 4 M. Significant difference (*P< 0.05)
compared with the previous younger group.
Fig. 7. The ME stained for SS in male mice at 2 M (A), 4 M (B), 12 M (C)
and 24 M (D). No clear difference was observed across the four age groups
in their SS—immunoreactivity in the ME. V: third ventricle. Scale bar
represents 100 Am (A–D).
S. Kuwahara et al. / Brain Research 998 (2004) 164–173 169
of ARC GHRH-ir neurons decreased dramatically from 5 to
20 days, gradually increased until day 40, then decreased
until day 60 [25]. From our results ([25], present data), it
appears that the number of ARC GHRH-ir neurons in mice
reaches its peak value just after onset of puberty, at day 40,
then decreases with age.
The number of PeN SS-ir neurons was not significantly
different, and the intensities and number of SS-ir fibers in
the ME were almost unchanged with age in our mice. In
elderly (24 M) male rats which were treated with colchicine,
SS-ir neurons in the PeN, and SS-ir intensities in the ME,
showed no change from those of young (3 M) rats [21].
However, Hoffman and Sladek [11] reported that the num-
ber of SS-ir fibers fell markedly in the ME with advancing
aging in male rats at 3, 12, 20 and 30 M. This conflicting
result remains unresolved. SS mRNA expression in the PeN
also showed an age-related decrease [33]. During postnatal
development, the number of SS-ir neurons increased signif-
icantly from days 5 to 30. The number of SS-ir neurons did
not increase significantly between days 30 and 40, and
increased between ages 40 and 60 of days [25]. These data
show that in mice, the number of PeN SS-ir neurons
changes during postnatal development, but does not change
after 2 M.
In our present study, the volume of the anterior
pituitary had decreased dramatically in 12 M mice from
its value in younger mice. In male rats, the weight of the
pituitary showed no significant differences between 6, 12,
and 18 M, but female pituitary glands at 12 and 18 M
were significantly heavier than in 6 M females [34]. In our
mice, many anterior pituitary cells that were stained by
PCNA, which has been used as a marker of cell prolifer-
ation [16,18], were detected at 2 and 4 M, but few PCNA-
ir cells were found in the adenohypophysis at 12 and 24
M. Furthermore, many apoptotic cells were found in 2 M
mice, but only a few apoptotic cells were detected
thereafter. The data presented here indicate that at 12
and 24 M, the proliferation of pituitary cells was extreme-
ly small and the volume and number of adenohypophysial
cells had decreased. Thus, the capability of regeneration of
the adenohypophysial cells may decrease with age. On the
other hand, GHRH is essential for the proliferation of the
somatotrope population during early development of the
pituitary [17]. With increasing age, GHRH synthesis and
secretion in GHRH neurons decreased, so that the anterior

Fig. 9. Proportion (A), number (B) and size (C) of GH-ir cells in the
anterior pituitary of male mice versus age. The proportion of GH-ir cells
(A) decreased with age. The number of GH-ir cells (B) in 4 M was
approximately half that in 12 M. The size of the GH-ir cells (C) fell from 2
to 4 M and from 4 to 12 M. Significant difference (*P < 0.05) compared
with the previous younger group.
S. Kuwahara et al. / Brain Research 998 (2004) 164–173170
pituitary cells decreased: this implies a marked decrease in
GH cells, because about 50% of anterior pituitary cells are
GH cell.
We observed that the proportion of GH-ir cells de-
creased significantly with age, that the number of GH-ir
cells reduced from 4 to 12 M, and the size of GH-ir cells
was smaller at 4 M than 2 M and at 12 M than 4 M. It has
previously been reported that no differences exist in basal
plasma levels of GH between 12 and 28 M in C57BL/6J
male mice [8], but Crew et al. [5] have reported that GH
mRNA levels fall with age and they suggested that
transcriptional activity is reduced in specific genes of
elderly male mice. In old male rats, the amplitude and
duration of GH pulse are significantly decreased, reducing
secretion of GH to about one third of that in young male
rats [32]. In male rats, GH-ir cells and GH cells, which
contain large secretory granules ultrastructurally (Type I
cells), decrease with age from 12 M [13,34], and pituitary
GH level at 18 M is significantly lower than at 6 M [34].
The age-related changes in GH secretion may be not due
to a change in the pituitary response to GHRH [36], but
due to the dysfunction in the synthesis, transport or release
of GHRH [21,35]. In our morphological study, however,
the reduction in the proportion, number and size of GH-ir
cells with age in male mice may therefore be caused by the
decrease in the number of GHRH-ir neurons, with no
involvement of SS-ir neurons.
In conclusion, the fall in the number of GHRH-ir neurons
reduces the number of GH-ir cells.
Acknowledgements
We thank Dr. A.F. Parlow (NIDDK) for the gift of rGH
antiserum and Dr. N. Miki (Tokyo Women’s Medical
College) for the gifts of mGHRH and mSS antisera. This
work was supported by a Grant-in-Aid for Scientific
Research from the Ministry of Education, Science, Sports,
and Culture of Japan to F.S. (No. 13660303).
References
[1] P. Ciofi, G. Tramu, B. Bloch, Colocalization of GHRF and NPY
immunoreactives in neurons of the infundicular area of the human
brain, Neuroendocrinology 47 (1988) 469–472.
[2] P. Ciofi, G. Tramu, B. Bloch, Comparative immunohistochemical
study of the distribution of neuropeptide Y, growth hormone-releasing
factor and carboxyterminus of precursor protein GHRF in human
hypothalamic infundibular area, Neuroendocrinology 51 (1990)
429–436.
[3] E. Corpas, S.M. Harman, S. Blackman, Human growth hormone and
human aging, Endocr. Rev. 14 (1993) 20–39.
[4] R. Cortes, S. Ceccatelli, M. Schalling, T. Hokfelt, Differential effects
of intracerebroventricular colchicine administration on the expression
of mRNAs for neuropeptides and neurotransmitter enzymes, with
special emphasis on galanin: an in situ hybridization study, Synapse
6 (1990) 369–391.
[5] M.D. Crew, S.R. Spindler, R.L. Walford, A. Koizumi, Age-related
decrease of growth hormone and prolactin gene expression in the
mouse pituitary, Endocrinology 121 (1987) 1251–1255.
[6] S. Daikoku, S. Hisano, H. Kawano, M. Chikamori-Aoyama, Y. Ka-
gotani, R. Zhang, K. Chihara, Ultrastructural evidence for neuronal
regulation of growth hormone secretion, Neuroendocrinology 47
(1988) 405–415.
[7] V. De Gennaro Colonna, M. Zoli, D. Cocchi, A. Maggi, P. Marrama,
L.F. Agnati, E.E. Muller, Reduced growth hormone releasing factor
(GHRF)-like immunoreactivity and GHRF gene expression in the
hypothalamus of aged rats, Peptides 10 (1989) 705–708.
[8] C.E. Finch, C. Jonec, J.R. Wisner Jr., Y.N. Sinha, J.S. de Vellis,
R.S. Swerdloff, Hormone production by the pituitary and testes of
male C57BL/6J mice during aging, Endocrinology 101 (1977)
1310–1317.
[9] M. Fodor, Z. Csaba, C. Kordon, J. Epelbaum, Growth hormone-
releasing hormone, somatostatin, galanin and h-endorphin afferent
to the hypothalamic periventricular nucleus, J. Chem. Neuroanat. 8
(1994) 61–73.
[10] L.A. Frohman, T.R. Doens, P. Chomocanski, Regulation of growth
hormone secretion, Front. Neuroendocrinol. 13 (1992) 344–405.

Fig. 10. GH-ir cells in the anterior pituitary of male mice at 2 M (A), 4 M (B), 12 M (C) and 24 M (D). At 2 M, GH-ir cells were larger and more intensively
stained than at other ages. The number and size of GH-ir cells fell from 4 to 12 M. Scale bar represents 20 Am (A–D).
S. Kuwahara et al. / Brain Research 998 (2004) 164–173 171
[11] G.E. Hoffman, J.R. Sladek Jr., Age-related changes in dopamine,
LHRH and somatostatin in the rat hypothalamus, Neurobiol. Aging
1 (1980) 27–37.
[12] K. Ishikawa, Y. Taniguchi, K. Kurosumi, M. Suzuki, M. Shinoda,
Immunohistochemical identification of somatostatin-containing neu-
rons projecting to median eminence of the rat, Endocrinology 121
(1987) 94–97.
[13] S. Jurado, G. Console, C.G. Dumm, Sexually dimorphic effects of
aging on rat somatotroph cells. An immunohistochemical and ultra-
structural study, J. Vet. Med. Sci. 60 (1998) 705–711.
[14] H. Kawano, S. Daikoku, Somatostatin-containing neurons systems in
the rat hypothalamus: retrograde tracing and immunohistochemical
studies, J. Comp. Neurol. 271 (1988) 293–299.
[15] S. Kuwahara, T. Mizukami, M. Omura, M. Hagihara, Y. Iinuma, Y.
Shimizu, H. Tamada, Y. Tsukamoto, T. Nishida, F. Sasaki, Seasonal
Fig. 11. PCNA-ir cells in the anterior pituitary of male mice at 2 M (A), 4 M (B),
mice, but few in 12 and 24 M mice. Scale bar represents 50 Am (A–D).
changes in the hypothalamo-pituitary-testes axis of the Japanese wood
mouse (Apodemus speciosus), Anat. Rec. 260 (2000) 366–372.
[16] G. Landberg, G. Roos, Antibodies to proliferating cell nuclear antigen
as S-phase probes in flow cytometric cell cycle analysis, Cancer Res.
51 (1991) 4570–4574.
[17] S.-C. Lin, C.R. Lin, I. Gukovsky, A.J. Lusis, P.E. Sawchenko,
M.G. Rosenfeld, Molecular basis of the little mouse phenotype
and implications for cell type-specific growth, Nature 364 (1993)
208–213.
[18] M.D. Linden, C.K. Ma, J. Kubus, R.D. Brown, R.J. Zarbo, Ki-67 and
proliferating cell nuclear antigen tumor proliferative indices in DNA
diploid colorectal adenocarcinomas. Correlation with histopathologic
characteristics and cell cycle analysis with two-color DNA flow
cytometry, Am. J. Clin. Pathol. 100 (1993) 206–212.
[19] I. Merchenthaler, G. Seralo, C. Csontos, P. Petrusz, B. Flerko, A.
12 M (C) and 24 M (D). Many PCNA-ir cells were detected in 2 and 4 M
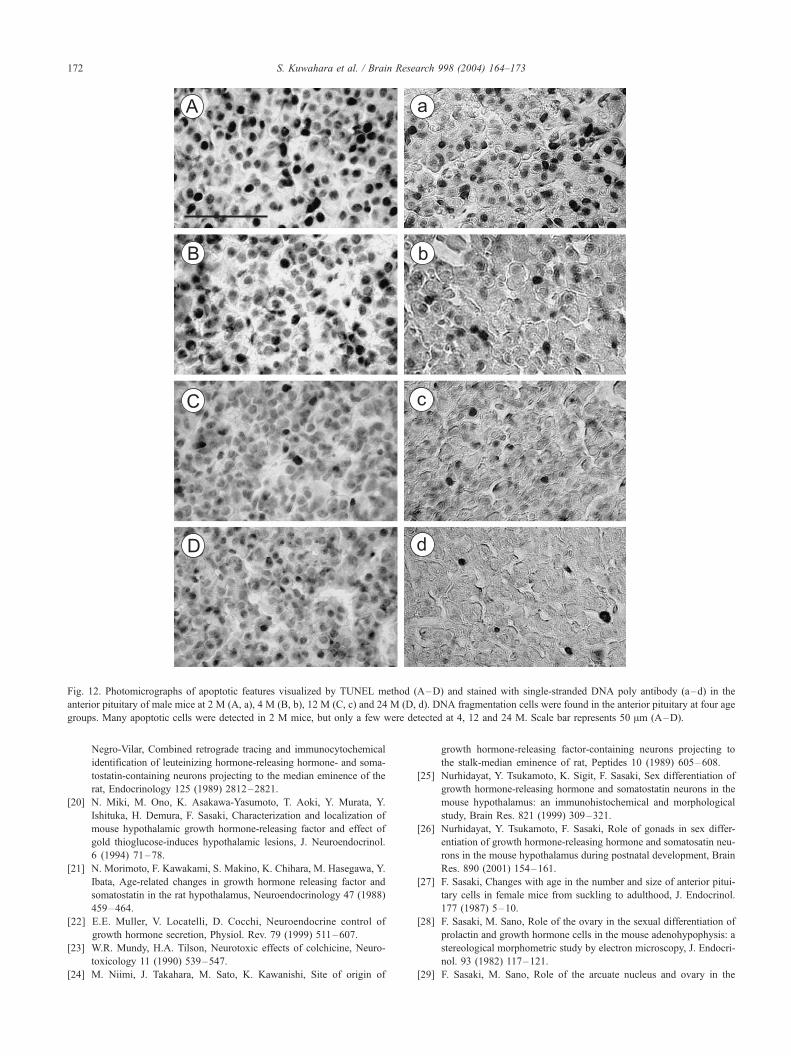
Fig. 12. Photomicrographs of apoptotic features visualized by TUNEL method (A–D) and stained with single-stranded DNA poly antibody (a–d) in the
anterior pituitary of male mice at 2 M (A, a), 4 M (B, b), 12 M (C, c) and 24 M (D, d). DNA fragmentation cells were found in the anterior pituitary at four age
groups. Many apoptotic cells were detected in 2 M mice, but only a few were detected at 4, 12 and 24 M. Scale bar represents 50 Am (A–D).
S. Kuwahara et al. / Brain Research 998 (2004) 164–173172
Negro-Vilar, Combined retrograde tracing and immunocytochemical
identification of leuteinizing hormone-releasing hormone- and soma-
tostatin-containing neurons projecting to the median eminence of the
rat, Endocrinology 125 (1989) 2812–2821.
[20] N. Miki, M. Ono, K. Asakawa-Yasumoto, T. Aoki, Y. Murata, Y.
Ishituka, H. Demura, F. Sasaki, Characterization and localization of
mouse hypothalamic growth hormone-releasing factor and effect of
gold thioglucose-induces hypothalamic lesions, J. Neuroendocrinol.
6 (1994) 71–78.
[21] N. Morimoto, F. Kawakami, S. Makino, K. Chihara, M. Hasegawa, Y.
Ibata, Age-related changes in growth hormone releasing factor and
somatostatin in the rat hypothalamus, Neuroendocrinology 47 (1988)
459–464.
[22] E.E. Muller, V. Locatelli, D. Cocchi, Neuroendocrine control of
growth hormone secretion, Physiol. Rev. 79 (1999) 511–607.
[23] W.R. Mundy, H.A. Tilson, Neurotoxic effects of colchicine, Neuro-
toxicology 11 (1990) 539–547.
[24] M. Niimi, J. Takahara, M. Sato, K. Kawanishi, Site of origin of
growth hormone-releasing factor-containing neurons projecting to
the stalk-median eminence of rat, Peptides 10 (1989) 605–608.
[25] Nurhidayat, Y. Tsukamoto, K. Sigit, F. Sasaki, Sex differentiation of
growth hormone-releasing hormone and somatostatin neurons in the
mouse hypothalamus: an immunohistochemical and morphological
study, Brain Res. 821 (1999) 309–321.
[26] Nurhidayat, Y. Tsukamoto, F. Sasaki, Role of gonads in sex differ-
entiation of growth hormone-releasing hormone and somatosatin neu-
rons in the mouse hypothalamus during postnatal development, Brain
Res. 890 (2001) 154–161.
[27] F. Sasaki, Changes with age in the number and size of anterior pitui-
tary cells in female mice from suckling to adulthood, J. Endocrinol.
177 (1987) 5–10.
[28] F. Sasaki, M. Sano, Role of the ovary in the sexual differentiation of
prolactin and growth hormone cells in the mouse adenohypophysis: a
stereological morphometric study by electron microscopy, J. Endocri-
nol. 93 (1982) 117–121.
[29] F. Sasaki, M. Sano, Role of the arcuate nucleus and ovary in the

S. Kuwahara et al. / Brain Research 998 (2004) 164–173 173
maturation of growth hormone, prolactin, and nongranulated cells in
the mouse adenohypophysis during postnatal development: a stereo-
logical morphometric study by electron microscopy, Endocrinology
119 (1986) 1682–1689.
[30] F. Sasaki, T. Kawai, M. Ohta, Immunohistochemical evidence of
neurons with GHRH or LHRH in the arcuate nucleus of male mice
and their possible role in the postnatal development of adenohypo-
physial cells, Anat. Rec. 240 (1994) 255–260.
[31] K. Shirasu, W.E. Stumpf, M. Sar, Evidence for direct action of estra-
diol on growth hormone-releasing factor (GRF) in rat hypothalamus:
localization of [3H]estradiol in GRF neurons, Endocrinology 127
(1990) 344–349.
[32] W.E. Sonntag, R.W. Steger, L.J. Forman, J. Meites, Decreased pulsa-
tile release of growth hormone in old male rat, Endocrinology 107
(1980) 1875–1879.
[33] W.E. Sonntag, R.L. Boyd, R.M. Booze, Somatostatin gene expression
in hypothalamus and cortex of aging male rats, Neurobiol. Aging 11
(1990) 409–416.
[34] S. Takahashi, Immunocytochemical and immuno-microscopical study
of growth hormone cells in male and female rats of various ages, Cell
Tissue Res. 266 (1991) 275–284.
[35] R.F. Walker, S.W. Yang, B.B. Bercu, Robust growth hormone se-
cretion in aged female rats co-administrated GH-releasing hexapep-
tide (GHRP-6) and growth hormone factor, Life Sci. 49 (1991)
459–464.
[36] W.B. Wehrenberg, N. Ling, The absence of an age-related change in
the pituitary response to growth hormone-releasing factor in rats,
Neuroendocrinology 37 (1983) 463–466.
Top Related
Copyright © 2022 FDOKUMEN

