







Bahasa
Halaman
Hukum


Bioresource Technology 96 (2005) 1897–1906
Aerobic nitrification–denitrification by heterotrophic Bacillus strains
Joong Kyun Kim a,*, Kyoung Joo Park a, Kyoung Sook Cho a, Soo-Wan Nam b,Tae-Joo Park c, Rakesh Bajpai d
a Division of Food Science and Biotechnology, Pukyong National University, Pusan 608-737, Republic of Koreab Department of Biotechnology and Bioengineering, Dong-Eui University, Pusan 614-714, Republic of Koreac Department of Environmental Engineering, Pusan National University, Pusan 609-735, Republic of Korea
d Department of Chemical Engineering, University of Missouri-Columbia, Columbia, MO, USA
Received in revised form 3 January 2005; accepted 28 January 2005
Available online 29 March 2005
Abstract
Twenty-four Bacillus strains predominantly outgrown in a night soil treatment system were isolated and characterized. Under
various culture conditions, cell interactions took place among them and cell population changed. Maximum removal of NHþ4 -N
and cell production by the isolates occurred under the conditions of 30% DO and C/N ratio of 8. Five dominant isolates were iden-
tified to be species of Bacillus cereus, Bacillus subtilis and Bacillus licheniformis with similarities of 78–94%. Additions of 0.8% pep-
tone and 0.3% yeast extract to a basal medium influenced the growth of isolates and the removal of NHþ4 -N in flask culture. Metal
ions such as Ca2+, Fe2+ and Mg2+ had a similar effect. The specific growth rates of the five isolates were found to be in a range of
0.43–0.55 h�1. During the flask experiment of nitrogen removal under aerobic growth conditions, active nitrification by the isolates
occurred largely in 1 h with a decrease of COD and alkalinity reduced to only 74.6% of theoretical value. From the nitrogen balance,
the percentage of nitrogen lost in the flask culture was estimated to be 33.0%, which was presumed to convert to N2 gas. This con-
version of ammonia to N2 without formation of nitrous oxide under aerobic growth conditions was confirmed by GC analysis.
From all the results, it has been found that the Bacillus strains were able to occur simultaneously aerobic nitrification/denitrification
and the B3 process using the Bacillus strains seemed to possess some economic advantages.
� 2005 Elsevier Ltd. All rights reserved.
Keywords: Aerobic nitrification–denitrification; Bacillus strains; Nitrogen removal; Cell-population balance
1. Introduction
The presence of nitrogenous substances in wastewater
discharges has attracted attention because of the role of
nitrogen in eutrophication of receiving waters. Nitrogenremoval is an important aspect of present day wastewater
treatment processes, and biological nitrification–denitri-
fication is one of the most economical processes for nitro-
gen removal from municipal wastewaters (Gupta and
Gupta, 2001). The nitrogenous substances in municipal
0960-8524/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.biortech.2005.01.040
* Corresponding author. Tel.: +82 51 620 6186; fax: +82 51 620 6180.
E-mail address: [email protected] (J.K. Kim).
wastewater is mostly in the form of ammonium ion
requiring a treatment process involving the biological
oxidation of NHþ4 into NO
�3 (nitrification) followed by
the biological reduction of NO�3 into N2 (denitrification).
It has commonly been accepted that nitrification anddenitrification require aerobic and anoxic conditions,
respectively. However, there have been periodic reports
of anaerobic ammonium oxidation (anammox) (Fux
et al., 2002; Schmidt et al., 2003) and aerobic denitrifica-
tion (Meiberg et al., 1980; Robertson and Kuenen, 1983;
Su et al., 2001b; van Niel et al., 1992). The nitrification–
denitrification process has been also challenged by a one-
step process in which ammonium is oxidized directly to

1898 J.K. Kim et al. / Bioresource Technology 96 (2005) 1897–1906
N2 (Astrid et al., 1995; Mulder et al., 1995). Recently, re-
searches focus on nitrite nitrification, which might be a
short cut process for savings in oxygen for nitrification
and carbon requirements for denitrification (Eum and
Choi, 2002; Rahman et al., 1995).
In Korea, �B3 (Bio Best Bacillus) process� (KoreanPatent No. 151928) has been known as an advanced
wastewater treatment system in which Bacillus strains
have predominated. It has been reported that the process
is able to remove nitrogen and phosphorus as well as or-
ganic matter efficiently (Choi et al., 2000). Various
microorganisms participate in the nitrogen cycle in the
natural ecosystem. Among them, the nitrification process
has been thought to be carried out mainly by ammonia-and nitrite-oxidizing bacteria that are obligately aerobic
and chemoautotrophic. However, a number of hetero-
trophic microorganisms have been reported to nitrify
many types of nitrogen compounds (Focht and Verstra-
ete, 1977). Despite the diversity of the reactions, their
activities have known to be generally lower than those
of autotrophs and to be not essential for their growth.
Hence, their contribution to the nitrification processhas remained unclear in spite of the studies on heterotro-
phic nitrification (Papen et al., 1989; Rho, 1986; Robert-
son and Kuenen, 1990; Robertson et al., 1988).
Bacillus strains have known to be involved in hetero-
trophic nitrification (Mevel and Prieur, 2000), but other
characteristics of the Bacillus strains have not been stud-
ied in detail so far. Understanding the characteristics of
microorganisms used in the biological nitrification–deni-trification process is very important in order to remain
high efficiency of treatment at all times, and the possible
microbiological nitrogen conversions should be pro-
vided. The purpose of this study was to understand
Bacillus strains predominantly occurring in a night soil
treatment system by isolating pure cultures and charac-
terizing these isolates.
2. Methods
2.1. Isolation of Bacillus strains
The Bacillus sludge and cell suspension were obtained
from five different sites of night soil treatment systems in
which Bacillus strains were predominantly occurringand stable growth was maintained. The samples were
first agitated to obtain homogeneous suspensions in
sterile 0.2% NaCl. One milliliter of the suspended liquid
was pipetted into a 10 mL tube that contained a 0.8%
nutrient broth medium. After two days of incubation
at 30 �C and 150 rpm, cells were spread with a platinum
loop on 1.5% nutrient agar plates. Purified isolates of
Bacillus cells were obtained by repeated streaking onfresh agar plates. Each isolate was maintained on a
1.5% nutrient agar plate.
2.2. Characterization of isolates and their taxonomic
identification
To observe the change of cell population among
Bacillus strains under various culture conditions, a 5-L
continuous stirred reactor was used with the workingvolume of 3.5 L. The pH and temperature in the reactor
were controlled at 7 and 30 �C, respectively. The agita-tion speed was 300 rpm, and the concentration of dis-
solved oxygen (DO) in the reactor was adjusted by
both air and argon gas at the same time. The same
amount of isolates was cultivated in flasks, and 10% of
those mixed cells were used as an inoculum. The compo-
sition of a medium used in this experiment was (g L�1):glucose, 1.05; NH4Cl, 0.382; KH2PO4, 0.131; peptone,
0.05; yeast extract, 0.05; and 1 mL-mineral solution.
The mineral solution contained (g L�1): FeSO4 Æ7H2O,
3; H3BO3, 0.01; Na2MoO4 Æ2H2O, 0.01; MnSO4 ÆH2O,
0.02; CuSO4 Æ5H2O, 0.01; ZnSO4, 0.01; and ethylenedia-
mine tetraacetic acid (EDTA), 0.5. Glucose or NH4Cl
was used to provide for various C (total organic carbon,
TOC)/N (NHþ4 -N) ratios. All media used were sterilized
by autoclaving at 121 �C for 15 min. The experiment
was conducted as a repeated batch. After environmental
conditions (DO and C/N ratio) were changed, first two
batches were wasted not to obtain transient data. Sam-
ples were taken and analyzed from the third and fourth
batches. The experiment was initiated again when more
than 30% of spores were found in the mixed culture after
changing culture conditions. Each batch was executedfor 8 h and measurements were performed in triplicates.
In flask experiments to study effects of growth factors
on growth of isolates, a basal synthetic medium was
used, which contained (g L�1): KNO3, 1; K2HPO4, 1;
EDTA, 3.5 · 10�3; ZnSO4 Æ7H2O, 2 · 10�3; FeSO4 Æ7H2O, 10 · 10�3; MnSO4 Æ7H2O, 2 · 10�3; CuSO4Æ5H2O, 1 · 10�3; Co(NO3)2 Æ6H2O, 0.2 · 10�3; and
H3BO3, 1 · 10�3. Vitamin solution contained (g L�1):nicotinic acid, 0.2; thiamine-HCl, 0.4; nicotinamide,
0.2; and biotin, 0.008. The vitamin solution was added
to the flask after autoclaving. After growth factors were
determined, a synthetic medium containing additional
0.8% peptone and 0.3% yeast extract in the basal med-
ium was used for cultures of pure and mixed cells in later
experiments. A bacterial suspension (10% inoculum) of
the isolates was inoculated into duplicate flasks. Themedium pH was maintained at 7, and flask cultures were
incubated at 150 rpm and 30 �C for 14 h.
Capacity of isolates for aerobic denitrification was
tested in a tightly sealed 1 L-branched flask in duplicate.
The flask was evacuated and pure oxygen was fully pres-
surized into the flask before autoclaving (initial DO was
approximately 70% saturated). A septum was equipped
on the mouth of the branched flask for gas analysis.The night soil was used as a main substrate, and the sub-
strate composition was adjusted by glucose, NH4Cl and

J.K. Kim et al. / Bioresource Technology 96 (2005) 1897–1906 1899
KH2PO4 to simulate a typical composition of municipal
wastewater in Korea. The final composition of the sub-
strate was (mg L�1): chemical oxygen demand (COD),
250; NHþ4 -N, 35; NO
�2 -N, 0; NO
�3 -N, 0; total nitrogen
(TN), 50; and total phosphorus (TP), 7. The isolates
were cultivated very actively under the environmentalcondition of 30% DO, and 10% of those cells were
inoculated into the flask. The flask was incubated at
150 rpm and 30 �C. Samples were taken from the flasks
periodically for measurements of optical density (OD),
dry cell weight (DCW), NHþ4 -N, NO
�2 -N, NO
�3 -N,
TN, TP, COD, alkalinity, oxygen and nitrogen gas
concentrations.
To obtain microscopic features of isolates, the Gramstaining and agar-stab culture were made. Cell size,
motility and morphology of isolates were also deter-
mined microscopically (Axiostar plus microscope, Carl
Zeiss, German) (1000·). For taxonomic identification,API 50CH identification system (Version 6.0) was used.
2.3. Analyses
Growth of isolates was determined spectrophotomet-
rically at a 510-nm wavelength (OPRON-3000, Hanson
Technology Co., Ltd., Korea). The DCW was deter-
mined by weighing the cell pellet after being dried in
an oven at 100 �C for 12 h. The cell pellet was prepared
by centrifuging a 5 mL sample of broth culture at
5000 rpm for 10 min and then by decanting the superna-
tant after washing twice with distilled water.The concentrations of NHþ
4 -N, COD of medium were
determined by procedures given in the Standard Meth-
ods (APHA, 1992). Estimation of NHþ4 -N was made
by a phenate method that monitored the absorbency
at 635 nm. COD was measured by a closed reflux color-
imetric method at 600 nm. Nitrite and nitrate were esti-
mated by ion chromatography (Metrohm 792 Basic IC,
Switzerland). TN and TP were determined by ultravioletadsorption method and ascorbic acid reduction method,
respectively (APHA, 1992). Alkalinity was determined
by the titration method.
For oxygen and nitrogen determination, 20 lL sam-
ples (injection volume) were taken by a press-lock syr-
inge for GC/TCD (Perkin Elmer Instruments, USA)
analysis. The carrier gas was helium at a flow rate of
20 mL min�1. The column used was a �molecular sieve5A� (stainless steel, mesh 80/100, 6 ft · 1/8 in). The col-umn and detector temperatures were 70 and 120 �C,respectively. The amounts of both oxygen and nitrogen
were calculated by applying the ideal gas law.
2.4. Statistical analyses
Statistical analyses were done with measurements ob-tained from this study. Since the sample observations
were not arranged in a frequency distribution, the stan-
dard deviations were calculated by the following proce-
dures: each deviation was squared, the sum of the
squares was divided by (n � 1), one less than the sample
size (n), (this resulted in the sample variance) and finally
extraction of the square root recovered the original scale
of measurement. Comparisons of means were performedby the Tukey method (Neter et al., 1985) using the SAS
program, since all sample sizes were equal. Differences
were considered significant at P < 0.05.
3. Results and discussion
3.1. Isolation of Bacillus strains
Bacillus strains were isolated by streaking on agar
plates. Various distinct colonies developed on the agar
plates after two days incubation. The microscopic fea-
tures of the 24 isolates are tabulated in Table 1.
Although some isolates exhibited similar characteristics,
they had different colony types in elevation and surface
on agar. Only few cells were isolated identically fromfive different sites of night soil treatment systems. This
fact suggested that dominant Bacillus strains in a main
process of wastewater treatment system could be chan-
ged significantly by different environmental conditions
in which cell interactions took place among microbial
population (Purtschert and Gujer, 1999). Only one
(PK22) out of 24 colonies was found to be Gram nega-
tive cell. It is, thus, not a Bacillus strain and it might bePseudomonas or Zoogloea that is generally present in
activated sludge.
Hence, a detailed experiment was performed in a 5-L
bioreactor in order to investigate the change of cell pop-
ulation under different culture conditions.
3.2. Change of cell population among isolates
To investigate the change of cell population among
Bacillus strains, all the isolates were combined in a batch
reactor incubated for 8 h under various culture condi-
tions. The result is tabulated in Table 2. For determina-
tion of each cell�s population in the mixed culture,
samples were diluted such that 25–50 colonies were
formed on an agar plate, and the identification of each
species was based upon information in Table 1. The col-onies that were not clearly identifiable were classified as
remaining Bacillus strains. As mentioned in Section 3.1,
cell interactions among the isolates were observed
distinctly.
When the reactor was first operated under conditions
of 30% DO and a C/N ratio of 4 (400 ppm TOC/
100 ppm NHþ4 -N), the most dominating species was
found to be PK15, followed by PK11, PK8, PK5,PK16 and the other species. This reflected consider-
able microbial interactions among isolates during

Table 1
The microscopic features of 24 isolates
Isolates Gram strain Sizea (lm) Colony pigmentb Chain forming Motilityc Agar-stab cultured
PK1 + L: 2.5–4, W: 0.8 I None ++ F.A. (s.g.)
PK2 + L: 3–4, W: 0.7–1 L.B. Pair ++ F.A. (s.g.)
PK3 + L: 4.5–6, W: 1 I Pair ++ F.A. (d.g.)
PK4 + L: 4–5, W: 0.5–0.8 I Pair + F.A. (d.g.)
PK5 + L: 4–5, W: 1.2 I 2–3 +++ F.A. (d.g.)
PK6 + L: 2–3, W: 0.8 I Pair ++ F.A. (d.g.)
PK7 + L: 7–9, W: 1.2–1.5 I 2–3 + F.A. (s.g.)
PK8 + L: 3–4, W: 0.8–1 I None ++ Aerobic
PK9 + L: 4–5, W: 1 S.T. None + F.A. (s.g.)
PK10 + L: 3–4, W: 1–1.2 I Pair + F.A. (s.g.)
PK11 + L: 3–4, W: 1.2 I Pair ++ Aerobic
PK12 + L: 4–5, W: 1–1.2 I None ++ F.A. (s.g.)
PK13 + L: 4–5, W: 1 I 2–4 + F.A. (s.g.)
PK14 + L: 6–7, W: 1.5 I 2–4 + F.A. (s.g.)
PK15 + L: 2.5–3, W: 0.8 S.T. None +++ Aerobic
PK16 + L: 1.5–2, W: 0.7 I None +++ F.A. (d.g.)
PK17 + L: 4–5, W: 1–1.2 I None ++ F.A. (s.g.)
PK18 + L: 2.5–3, W: 0.8–1 L.Y. None ++ F.A. (s.g.)
PK19 + L: 1.5–2, W: 0.7 I None ++ F.A. (d.g.)
PK20 + L: 3–4, W: 1.2 I Several ++ F.A. (d.g.)
PK21 + L: 4–5, W: 1.2–1.5 I 2–4 ++ F.A. (d.g.)
PK22 � L: 1–1.2, W: 1 S.T. None ++ F.A. (d.g.)
PK23 + L: 3.5–5, W: 1 I 2–3 ++ F.A. (d.g.)
PK24 + L: 1–1.2, W: 0.8–1 L.Y. None ++ F.A. (d.g.)
a L means length and W means width.b Symbols mean as follows: I, ivory; L.B., light brown; S.T., semi-translucence; and L.Y., light yellow.c + means the degree of motility: +, weak; ++, moderate; and +++, strong.d F.A. means facultatively anaerobic; s.g. shallow growth and d.g. deep growth.
Table 2
Change of cell-population balance for 8 h under various culture conditionsa
Culture conditions Removal
of NHþ4 -N
(mg L�1)
Cell
production
(g L�1)
Population balance
30% DO and C/N = 4 (400 ppm TOC/100 ppm NHþ4 -N) 30 ± 2e 0.25 ± 0.01e 73 ± 7%ab PK15, 8 ± 4% PK11, 8 ± 4% PK8,
4 ± 2%b PK5, 4 ± 2%a PK16, 5 ± 4% others
30% DO and C/N = 8 (800 ppm TOC/100 ppm NHþ4 -N) 118 ± 4c 0.60 ± 0.03d 83 ± 6%ab PK15, 6 ± 4%b PK5, 6 ± 4%a PK16,
7 ± 3% others
30% DO and C/N = 4 (800 ppm TOC/200 ppm NHþ4 -N) 163 ± 4b 0.86 ± 0.02b 87 ± 6%a PK15, 4 ± 2%b PK5, 4 ± 2%a PK16,
4 ± 2% others
30% DO and C/N = 8 (1600 ppm TOC/200 ppm NHþ4 -N) 226 ± 5a 1.10 ± 0.02a 90 ± 6%a PK15, 4 ± 2%b PK5, 4 ± 2%a PK16,
2 ± 2% others
15% DO and C/N = 8 (800 ppm TOC/100 ppm NHþ4 -N) 116 ± 3c 0.68 ± 0.03cd 85 ± 5%ab PK15, 4 ± 2%b PK5, 7 ± 4%a PK16,
4 ± 3% others
5% DO and C/N = 8 (800 ppm TOC/100 ppm NHþ4 -N) 63 ± 3d 0.40 ± 0.04e 52 ± 8%b PK15, 28 ± 4%a PK5, 19 ± 4%a PK16,
2 ± 2% others
30% DO and C/N = 8 (800 ppm TOC/100 ppm NHþ4 -N) 120 ± 4c 0.76 ± 0.03bc 90 ± 4%a PK15, 5 ± 2%b PK5, 5 ± 2%a PK16,
2 ± 2% others
a Means in the same column with different superscript are significantly different (P < 0.05). Values represent mean ± S.D. of two replicates.
1900 J.K. Kim et al. / Bioresource Technology 96 (2005) 1897–1906
heterotrophic nitrification (Rho, 1986). The domination
of PK15 increased with the increase of C/N ratio, while
the populations of PK8 and PK11 reduced significantly.
Even at the same C/N ratio, the PK15 was predominant
in the medium containing high concentrations of TOC
and NHþ4 -N. The results showed that carbon and nitro-
gen components in the medium affected the nitrifying
activity of the isolates. When the DO concentration de-
creased from 30% to 15%, the removal amount of
NHþ4 -N reduced to half with minimal changes of cell-
population balance. At 5% DO and a C/N ratio of 8,
the populations of PK5 and PK16 increased markedly,
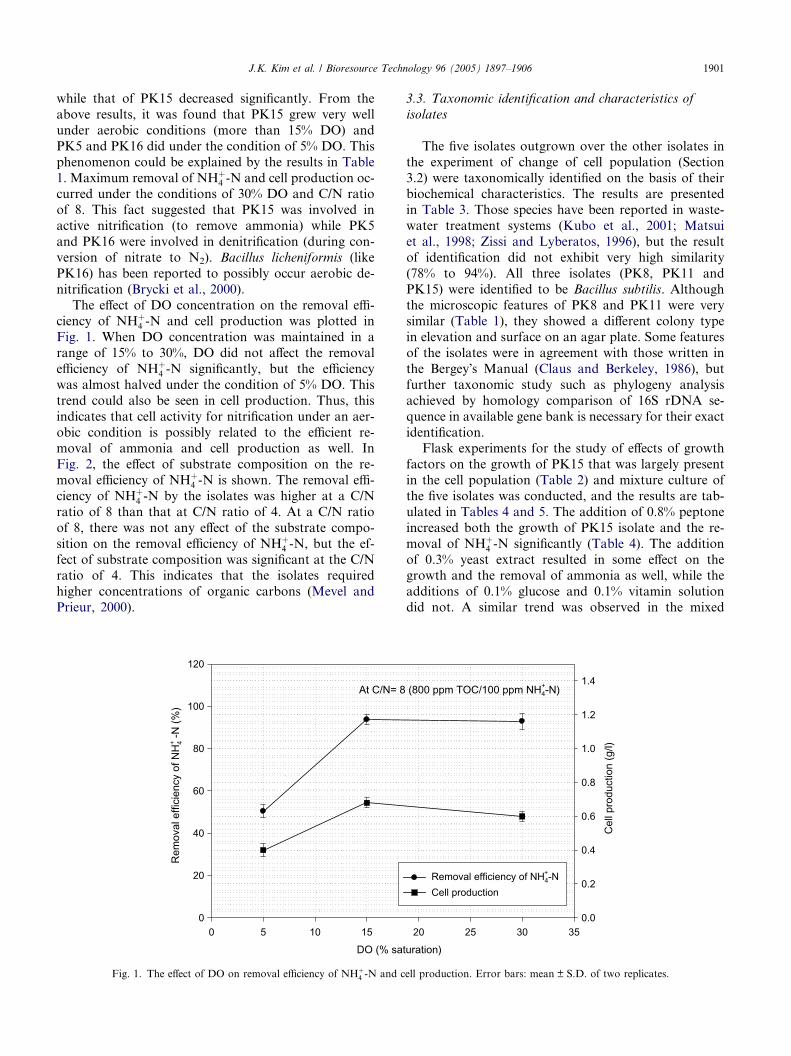
J.K. Kim et al. / Bioresource Technology 96 (2005) 1897–1906 1901
while that of PK15 decreased significantly. From the
above results, it was found that PK15 grew very well
under aerobic conditions (more than 15% DO) and
PK5 and PK16 did under the condition of 5% DO. This
phenomenon could be explained by the results in Table
1. Maximum removal of NHþ4 -N and cell production oc-
curred under the conditions of 30% DO and C/N ratio
of 8. This fact suggested that PK15 was involved in
active nitrification (to remove ammonia) while PK5
and PK16 were involved in denitrification (during con-
version of nitrate to N2). Bacillus licheniformis (like
PK16) has been reported to possibly occur aerobic de-
nitrification (Brycki et al., 2000).
The effect of DO concentration on the removal effi-ciency of NHþ
4 -N and cell production was plotted in
Fig. 1. When DO concentration was maintained in a
range of 15% to 30%, DO did not affect the removal
efficiency of NHþ4 -N significantly, but the efficiency
was almost halved under the condition of 5% DO. This
trend could also be seen in cell production. Thus, this
indicates that cell activity for nitrification under an aer-
obic condition is possibly related to the efficient re-moval of ammonia and cell production as well. In
Fig. 2, the effect of substrate composition on the re-
moval efficiency of NHþ4 -N is shown. The removal effi-
ciency of NHþ4 -N by the isolates was higher at a C/N
ratio of 8 than that at C/N ratio of 4. At a C/N ratio
of 8, there was not any effect of the substrate compo-
sition on the removal efficiency of NHþ4 -N, but the ef-
fect of substrate composition was significant at the C/Nratio of 4. This indicates that the isolates required
higher concentrations of organic carbons (Mevel and
Prieur, 2000).
DO (% sat0 5 10 15
Rem
oval
effi
cien
cy o
f NH
4 -N (%
)
0
20
40
60
80
100
120
At C/N= 8
Fig. 1. The effect of DO on removal efficiency of NHþ4 -N and c
3.3. Taxonomic identification and characteristics of
isolates
The five isolates outgrown over the other isolates in
the experiment of change of cell population (Section
3.2) were taxonomically identified on the basis of theirbiochemical characteristics. The results are presented
in Table 3. Those species have been reported in waste-
water treatment systems (Kubo et al., 2001; Matsui
et al., 1998; Zissi and Lyberatos, 1996), but the result
of identification did not exhibit very high similarity
(78% to 94%). All three isolates (PK8, PK11 and
PK15) were identified to be Bacillus subtilis. Although
the microscopic features of PK8 and PK11 were verysimilar (Table 1), they showed a different colony type
in elevation and surface on an agar plate. Some features
of the isolates were in agreement with those written in
the Bergey�s Manual (Claus and Berkeley, 1986), but
further taxonomic study such as phylogeny analysis
achieved by homology comparison of 16S rDNA se-
quence in available gene bank is necessary for their exact
identification.Flask experiments for the study of effects of growth
factors on the growth of PK15 that was largely present
in the cell population (Table 2) and mixture culture of
the five isolates was conducted, and the results are tab-
ulated in Tables 4 and 5. The addition of 0.8% peptone
increased both the growth of PK15 isolate and the re-
moval of NHþ4 -N significantly (Table 4). The addition
of 0.3% yeast extract resulted in some effect on thegrowth and the removal of ammonia as well, while the
additions of 0.1% glucose and 0.1% vitamin solution
did not. A similar trend was observed in the mixed
uration)20 25 30 35
Cel
l pro
duct
ion
(g/l)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Removal efficiency of NH4-NCell production
(800 ppm TOC/100 ppm NH4-N)
+
+
ell production. Error bars: mean ± S.D. of two replicates.

C/N Ratio
C/N = 4 C/N = 8
Rem
oval
effi
cien
cy o
f NH
4 -N (%
)
0
20
40
60
80
100
120
400 ppm TOC/100 ppm NH4 -N
800 ppm TOC/200 ppm NH4 -N
800 ppm TOC/100 ppm NH4 -N
1600 ppm TOC/200 ppm NH4 -N
Under the condition of 30% DO+
+
+
+
Fig. 2. The effect of substrate composition on removal efficiency of NHþ4 -N. Error bars: mean ± S.D. of two replicates.
Table 3
The result of identification for the dominant Bacillus species among
isolatesa
Dominant species Identification Similarity (%)
PK5 B. cereus 78 ± 2
PK8 B. subtilis 92 ± 0
PK11 B. subtilis 89 ± 1
PK15 B. subtilis 84 ± 2
PK16 B. licheniformis 94 ± 0
a Values represent mean ± S.D. of two replicates.
Table 4
The effects of growth factors on the growth of isolates cultivated at pH 7 an
Addition of growth factors PK15
DOD510 � initial OD510 % removal of NH
+0.1% glucose 0.47 ± 0.06c 9.3 ± 1.3c
+0.8% peptone 7.25 ± 0.19a 64.7 ± 1.6a
+0.3% yeast extract 2.56 ± 0.13b 29.9 ± 1.1b
+0.1% vitamin solution 0.27 ± 0.06c 3.2 ± 0.7c
a Means in the same column with different superscript are significantly diffb Mixture of PK5, PK8, PK11, PK15 and PK16 cultures.
Table 5
The effects of metal ions on the growth of PK15 and the removal of NHþ4 -N
Measurement Ca2+ Co2+ Cu2+ Fe2+
DOD510 � initial OD510 21.67 ± 0.44a 1.11 ± 0.11d 1.22 ± 0.11d 19.56
% removal of NHþ4 -N 87.9 ± 2.0a 4.8 ± 0.5d 5.3 ± 0.5d 81.2
a Means in the same row with different superscript are significantly differe
1902 J.K. Kim et al. / Bioresource Technology 96 (2005) 1897–1906
culture studies. Hence, in later experiments we used a
synthetic medium containing 0.8% peptone and 0.3%yeast extract additionally in the basal medium.
As shown in Table 5, 0.5-mM metal ions such as
Ca2+, Fe2+ and Mg2+ influenced the growth of PK15
and removal of NHþ4 -N as well. However, Co2+, Cu2+
and Zn2+ did not show their effects on either the growth
or the removal of ammonia, while Mn2+ and Mo2+ did
significantly only on the growth of PK15. These phe-
nomena can be explained in that the metal ions have
d 30 �Ca
Mixed cultureb
þ4 -N DOD510 � initial OD510 % removal of NHþ
4 -N
0.73 ± 0.09c 14.7 ± 1.3c
8.31 ± 0.31a 60.3 ± 2.0a
3.77 ± 0.15b 32.2 ± 1.7b
0.42 ± 0.08c 3.9 ± 0.6d
erent (P < 0.05). Values represent mean ± S.D. of two replicates.
a
Mg2+ Mn2+ Mo2+ Zn2+
± 0.44b 21.56 ± 0.33a 18.44 ± 0.23b 18.89 ± 0.33b 6.89 ± 0.22c
± 1.9a 85.5 ± 1.5a 28.5 ± 1.9c 40.1 ± 1.9b 9.2 ± 0.9d
nt (P < 0.05). Values represent mean ± S.D. of two replicates.

Culture time (h)0 2 4 6 8 10 12 14
OD
510
0.0
0.5
1.0
1.5
2.0
2.5
PK5PK8PK11PK15PK16
Fig. 3. Growth profiles of the five isolates obtained from each pure culture. Error bars: mean ± S.D. of two replicates.
J.K. Kim et al. / Bioresource Technology 96 (2005) 1897–1906 1903
been known to be involved in enzyme catalysis in a vari-
ety of ways and that trace metals Mo2+ and Fe2+ are
found in nitric-oxide reductase (Palmer, 1995).
To characterize the growth of each pure isolate, flask
cultures were incubated with the synthetic medium con-
taining 0.8% peptone and 0.3% yeast extract. Thegrowth profiles of five isolates were plotted in Fig. 3.
The data points represent average values for two repli-
cates. All isolates reached stationary phase in 8 h, and
the cultures of PK5 and PK16 exhibited longer lag times
than the cultures of PK8, PK11 and PK15. The values of
specific growth rates were calculated to be 0.51, 0.43,
0.47, 0.55 and 0.45 h�1 for PK5, PK8, PK11, PK15
and PK16, respectively. Among the three B. subtilis spe-cies that were found to be aerobes, the specific growth
rate of PK15 was highest. This fact might result in over-
growth of PK15 over the other two species, PK8 and
PK11 (described in the experiment in Section 3.2),
although cell interaction can be very complex in mixed
culture (Purtschert and Gujer, 1999).
3.4. Capacity of the isolates for aerobic denitrification
In Fig. 4, changes of various components in the flask
culture during the experiment of nitrogen removal under
an aerobic condition are shown. The main substrate for
this experiment was night soil because the isolates re-
quired high C/N ratio with a high concentration of or-
ganic carbon. The concentration of NHþ4 -N decreased
significantly in 1 h, and as did TN. The same trend couldbe seen in both the removal of COD and the decrease in
alkalinity. This indicates that active nitrification oc-
curred largely in 1 h with a decrease of COD. At this
time, nitrate began to accumulate by nitrification, and
remained as denitrification occurred simultaneously,
without any nitrite build-up. The similar result has been
seen in other studies using B. subtilis sp. (Sakai et al.,1996). It has been reported that during aerobic denitrifi-
cation by autotrophic Nitrosomonas europaea, dinitro-
gen was produced as an end product, whereby nitrous
oxide was observed as an intermediate (Shrestha et al.,
2002). Thus, the heterotrophic Bacillus strains possessed
less complex metabolic pathways for removal of ammo-
nia than did autotrophs. The initial DO was measured
to be 5.29 mg L�1, decreased to 1.93 mg L�1 in 1 hand then was reduced slowly to 0.44 mg L�1 (about
8% saturated) after 4 h. The amount of DO derived
from oxygen pressurized fully in the tightly sealed flask
did not match consumption of DO by Bacillus strains
due to the low solubility of oxygen (Nielsen and Villad-
sen, 1994). The concentrations of NHþ4 -N, TN, COD
and alkalinity did not decrease significantly after 2 h,
probably due to the decrease of DO in the flask. How-ever, the production of N2 increased steadily. This indi-
cated that a low partial oxygen pressure was favorable
for N2 production, and this result is in agreement with
that from study of autotrophic Nitrosomonas europaea
(Shrestha et al., 2002). From the results obtained in Sec-
tion 3.2, it can be inferred that PK15 was largely in-
volved in nitrification and PK5 and PK16 in
production of N2 during aerobic nitrification–denitrification.

Culture time (h)0 2 4
NH
4 -N
, NO
2 -N
, NO
3 -N
(mg/
l)
0
10
20
30
40
NH4+ -N
NO2- -N
NO3- -N
Culture time (h)0 2 3
TN (m
g/l)
0
10
20
30
40
50
60
Org
-N, N
2, D
O (m
g/l)
0
2
4
6
8
10
12
14
16
18
TNOrg-NN2
DO
Culture time (h)0 2 3
CO
D, A
lkal
inity
(mg/
l)
0
50
100
150
200
250
300
CODAlkalinity
Culture time (h)0 2 3
TP (m
g/l)
0
2
4
6
8
Dry
cel
l wei
ght (
mg/
l)
0.40
0.45
0.50
0.55
0.60
TPDCW
(a) (b)
(c) (d)
4 514 51
1 3 5 4 51
Fig. 4. Changes of NHþ4 -N, NO
�2 -N and NO�
3 -N (a), TN, Org-N and N2 (b), COD and alkalinity (c), TP and DCW (d), in the flask culture of mixed
isolates. Error bars: mean ± S.D. of two replicates.
1904 J.K. Kim et al. / Bioresource Technology 96 (2005) 1897–1906
The organic nitrogen (Org-N) was almost depleted by
the isolates after 4 h. The removal efficiencies of
NHþ4 -N, TN, COD and TP for 4 h were found to be
72.0, 57.6, 89.5 and 21.4%, respectively. The low re-
moval efficiency of TP could result from low production
of cells (only 100 mg L�1 produced for 4 h) (Su et al.,2001a). The alkalinity destructed in the culture was
5.28 mg L�1 (as CaCO3) per each gram of NHþ4 -N oxi-
dized. In conventional active sludge, substantial quan-
tity of alkalinity is lost during nitrification, but the
alkalinity decreased in this study was only 74.6% of
the theoretical value (Grady et al., 1999). When the con-
centration of organic nitrogen decreased, cell produc-
tion was largely retained and substantial removal ofCOD and NHþ
4 -N did not continue. The ratio of milli-
gram of COD removed per milligram of NHþ4 -N re-
moved was calculated to be 7.46.
Table 6
Nitrogen balance for the flask culturea (units: mg L�1)
Initial TN Final NHþ4 -N Final NO�
2 -N Final NO�3 -N
50.0 ± 1.5 9.8 ± 0.1 0 ± 0 11.3 ± 0.1
a Values represent mean ± S.D. of triplicates.b Calculated value.c Biomass composition was assumed to be C5H7O2N.d % N lost = 100 · {(initial TN) � (final NHþ
4 -N) � (final NO�2 -N) � (fina
From the nitrogen balance (Table 6), 14.5 mg L�1
(33% of initial TN) of nitrogen in the medium disap-
peared after 4 h. The amount of N2 gas measured was
12.0 mg L�1 (24% of initial TN). Hence, 9% of initial
TN was missing during nitrogen removal by the isolate.
The difference might result from measurement errors,but the similar result could be found in the previous re-
port, showing that on an average 8% of influent nitrogen
did not appear in the effluent in nitrogen balance between
the influent and the effluent (Gonenc and Harremoes,
1985). From these results, the mixed Bacillus cells were
found to lead to aerobic denitrification. Heterotrophs
that have been reported to occur aerobic denitrification
includeAlcaligenes faecalis (vanNiel et al., 1992),Hypho-
microbium X (Meiberg et al., 1980), Paracoccus denitrifi-
cans (previously named Thiosphaera pantotropha)
(Robertson et al., 1988) and Pseudomonas stutzeri
Final Org-Nb N in Biomassc % N lostd
0.5 ± 0.2 12.4 ± 0.3 33.0
l NO�3 -N) � (final Org-N) � (N in biomass)}/(initial TN).

J.K. Kim et al. / Bioresource Technology 96 (2005) 1897–1906 1905
(Su et al., 2001b). In Bacillus strains, the possibility of
aerobic denitrification by B. licheniformis has been re-
ported (Brycki et al., 2000). However, microbial interac-
tion between mixed Bacillus strains and their
characteristics in removal of nitrogen have not been
reported.
4. Conclusions
To understand characteristics of Bacillus strains used
in a biological nitrification–denitrification process, 24
species were isolated from the sludge in a night soil
treatment system. Under different environmental condi-
tions, cell interactions took place among them and cell
population changed. Additions of 0.8% peptone and
0.3% yeast extract to a basal medium influenced thegrowth of isolates and the removal of NHþ
4 -N, and as
did metal ions such as Ca2+, Fe2+ and Mg2+. It was
found that PK15 (identified to be B. subtilis) was in-
volved largely in nitrification and PK5 (identified to be
Bacillus cereus) and PK16 (identified to be B. lichenifor-
mis) were involved in production of N2 during aerobic
nitrification–denitrification. A low partial oxygen pres-
sure was favorable for the N2 production. From thenitrogen balance, the percentage of nitrogen lost in a
nitrification–denitrification process was estimated to be
33.0%, which was presumed to have been converted to
N2 gas. The Bacillus strains were found to convert
ammonia to N2 without formation of nitrous oxide
under aerobic conditions. Thus, in removal of ammonia,
the heterotrophic Bacillus strains had less complex met-
abolic pathways than autotrophs. The ability to both ni-trify and denitrify makes Bacillus strains attractive
candidates for application in the removal of ammonia
nitrogen from wastewater. Thus, the B3 process using
Bacillus strains seemed to have some economic advanta-
ges, and the development of this aerobic denitrification
process may serve as an alternative to the costly nitrifi-
cation–denitrification processes.
Acknowledgements
This research has been supported by theKorea Science
Engineering Foundation (KOSEF) (Grant No. R12-
1996-009203-0) through the Institute for Environmental
Technology and Industry at Pusan National University.
Technical assistance of Mr. Ill-Ho Jung (Daekyong En-
Tech International Co., Seoul) is also acknowledged.
The students, Kyoung Joo Park and Kyoung SookCho, were supported by the Brain Korea 21 Project.
References
APHA, 1992. Standard Methods for the Examination of Water and
Wastewater, 18th ed. American Public Health Association, Wash-
ington, DC.
Astrid, A., van de Graaf, A.A., Mulder, A., de Bruijn, P., Jetten, M.S.,
Robertson, L.A., Kuenen, J.G., 1995. Anaerobic oxidation of
ammonium is a biologically mediated process. Applied and
Environmental Microbiology 61, 1246–1251.
Brycki, B., Seifert, K., Szymanska, K., Domka, F., 2000. The effect of
oxidizing biocides on desulfurication and denitrification processes.
Polish Journal of Environmental Studies 9 (5), 363–367.
Choi, Y.S., Hong, S.W., Kim, S.J., Chung, I.H., 2000. Development of
a biological process for livestock wastewater treatment using a
technique for predominant outgrowth of Bacillus species. Water
Science and Technology 45, 71–78.
Claus, D., Berkeley, R.C.W., 1986. Endospore-forming Gram-positive
rods and cocci. In: Sneath, P., Mair, N., Sharpe, M.E., Hilt, J.
(Eds.), Bergey�s Manual of Systematic Bacteriology, vol. 2.
Williams and Wilkins, Baltimore, pp. 1105–1139.
Eum, Y., Choi, E., 2002. Optimization of nitrogen removal from
piggery waste by nitrite nitrification. Water Science and Techno-
logy 45, 89–96.
Focht, D.D., Verstraete, W., 1977. Biochemical ecology of nitrification
and denitrification. Advances in Microbial Ecology 1, 135–214.
Fux, C., Boehler, M., Huber, P., Brunner, I., Siegrist, H., 2002.
Biological treatment of ammonium-rich wastewater by partial
nitrification and subsequent anaerobic ammonium oxidation
(anammox) in a pilot plant. Journal of Biotechnology 99, 295–306.
Grady Jr., C.P.L., Daigger, G., Lim, H., 1999. Biological Wastewater
Treatment, second ed.MarcelDekker, Inc., NewYork, pp. 216–221.
Gonenc, I.E., Harremoes, P., 1985. Nitrification in rotating disc
system-I: criteria for transition from oxygen to ammonia rate
limitation. Water Research 19, 1119–1127.
Gupta, A.B., Gupta, S.K., 2001. Simultaneous carbon and nitrogen
removal from high strength domestic wastewater in an aerobic
RBC biofilm. Water Research 35, 1714–1722.
Kubo, M., Hiroe, J., Murakami, M., Fukami, H., Tachiki, T., 2001.
Treatment of hypersaline-containing wastewater with salt-tolerant
microorganisms. Journal of Bioscience and Bioengineering 91,
222–224.
Matsui, S., Yamamoto, H., Shimizu, Y., Harada, J., Einaga, D., 1998.
Humic substances affecting the limitation of the activated sludge
process for removal of micropollutants. Water Science and
Technology 38, 217–225.
Meiberg, J.B., Bruinenberg, M.P.M., Harder, W., 1980. Effect of
dissolved oxygen tension on the metabolism of methylated amines
in Hyphomicrobium X in the absence and presence of nitrate:
evidence for aerobic denitrification. Journal of General Micro-
biology 120, 453–463.
Mevel, G., Prieur, D., 2000. Heterotrophic nitrification by a thermo-
philic Bacillus species as influenced by different culture conditions.
Canadian Journal of Microbiology 46, 465–473.
Mulder, A., van de Graaf, A.A., Robertson, L.A., Kuenen, J.G., 1995.
Anaerobic ammonium oxidation discovered in a denitrifying
fluidized bed reactor. FEMS Microbiology Ecology 16, 177–184.
Nielsen, J., Villadsen, J., 1994. Bioreaction Engineering Principles.
Plenum Press, New York, pp. 296–300.
Neter, J., Wasserman, W., Kutner, M.H., 1985. Applied Linear
Statistical Models, second ed. IRWIN, Homewood, pp. 574–579.
Palmer, T., 1995. Understanding Enzymes, fourth ed. Prentice Hall,
London, pp. 202–206.
Papen, H., Berg, R., Hinkel, I., Thoene, B., Rennenberg, H., 1989.
Heterotrophic nitrification by Alcaligenes faecalis: NO�2 , NO
�3 ,
N2O, and NO production in exponentially growing cultures.
Applied and Environmental Microbiology 55, 2068–2072.
Purtschert, I., Gujer, W., 1999. Population dynamics by methanol
addition in denitrifying wastewater treatment plants. Water Science
and Technology 39, 43–50.
Rahman, H., Rols, J.L., Capdeville, B., Cornier, J.C., Deguin, A.,
1995. Nitrite removal by a fixed culture in a submerged granular
biofilter. Water Research 29, 1745–1753.

1906 J.K. Kim et al. / Bioresource Technology 96 (2005) 1897–1906
Rho, J., 1986. Microbial interactions in heterotrophic nitrification.
Canadian Journal of Microbiology 32, 243–247.
Robertson, L.A., Kuenen, J.G., 1983. Thiosphaera pamtotropha gen.
nov., sp. nov., a facultatively anaerobic, facultatively autotrophic
sulphur bacterium. Journal of General Microbiology 129,
2847–2855.
Robertson, L.A., Kuenen, J.G., 1990. Combined heterotrophic nitri-
fication and aerobic denitrification in Thiosphaera pantotropha and
other bacteria. Antonie van Leeuwenhoek 57, 139–152.
Robertson, L.A., van Niel, E.W.J., Torremans, R.A.M., Kuenen, J.G.,
1988. Simultaneous nitrification and denitrification in aerobic
chemostat of Thiosphaera pantotropha. Applied and Environmental
Microbiology 54, 2812–2818.
Sakai, K., Ikehata, Y., Ikenaka, Y., Wakayama, M., Moriguchi, M.,
1996. Nitrite oxidation by heterotrophic bacteria under various
nutritional and aerobic conditions. Journal of Fermentation and
Bioengineering 82, 613–617.
Schmidt, I., Sliekers, O., Schmid, M., Bork, E., Fuerst, J., Kuenen,
J.G., Jetten, M.S.M., Strous, M., 2003. New concept of microbial
treatment processes for the nitrogen removal in wastewater. FEMS
Microbiology Reviews 27, 449–557.
Shrestha, N.K., Hadano, S., Kamachi, T., Okura, I., 2002. Dinitrogen
production from ammonia by Nitrosomonas europaea. Applied
Catalysis A: General 237, 33–39.
Su, J.-J., Liu, B.-Y., Chang, Y.-C., 2001a. Identifying an interfering
factor on chemical oxygen demand (COD) determination in piggery
wastewater and eliminating the factor by an indigenous Pseudomo-
nas stutzeri strain. Letters in Applied Microbiology 33, 440–444.
Su, J.-J., Liu, B.-Y., Lin, J., Yang, C.-P., 2001b. Isolation of an aerobic
denitrifying bacterial strain NS-2 from the activated sludge of
piggery wastewater treatment systems in Taiwan possessing deni-
trification under 92% oxygen atmosphere. Journal of Applied
Microbiology 91, 853–860.
van Niel, E.W.J., Braber, K.J., Robertso, L.A., Kuenen, J.G., 1992.
Heterotrophic nitrification and aerobic denitrification in Alcalige-
nes faecalis strain TUD. Antonie van Leeuwenhoek 62, 231–237.
Zissi, U., Lyberatos, G., 1996. Axo-dye biodegradation under anoxic
conditions. Water Science and Technology 34, 495–500.
Top Related
Copyright © 2022 FDOKUMEN

