
Vegetation complexity—The influence of plant species diversity and plant structures on plant...
13
Basic and Applied Ecology 11 (2010) 383–395 REVIEW Vegetation complexity—The influence of plant species diversity and plant structures on plant chemical complexity and arthropods Barbara Randlkofer a , Elisabeth Obermaier b , Monika Hilker a , Torsten Meiners a,∗ a Freie Universität Berlin, Institute of Biology, Applied Zoology/Animal Ecology, Haderslebener Str. 9, 12163 Berlin, Germany b University of Würzburg, Department of Animal Ecology and Tropical Biology, Germany Received 5 April 2009; accepted 8 March 2010 Abstract Vegetation complexity is characterized by two major traits, i.e., plant chemical and plant structural complexity. Plant species diversity strongly determines these traits. Furthermore, plant structures affect microclimatic conditions, which in turn influence the emission and dispersion of plant volatiles (e.g., chemical complexity). Plant volatile chemical complexity may significantly affect orientation of herbivorous and carnivorous arthropods. Therefore, the way in which plant chemical and plant structural complexity act “in concert” may influence foraging and mating success of arthropods, and thus, finally, community composition. This review emphasizes an integrative view on the relationship between plant species diversity, plant structural complexity, plant volatiles (chemical complexity) and their effects on arthropods. Three new hypotheses are raised, which predict possible relations between plant volatile complexity and plant species diversity: (1) saturation-, (2) step-by-step, (3) incoherence-hypothesis. We conclude that arthropod orientation in natural environments is strongly determined by the relationship between plant volatile diversity and plant species diversity. Furthermore, we emphasize that structural complexity of the vegetation affects plant volatile diversity and thus, arthropod orientation. We review available information on how insects actually respond to complexity during olfactory and visual search and ask for both laboratory and field studies to further unravel the mechanisms of interactions between vegetation traits and their impact on arthropod orientation. Zusammenfassung Die Vegetationskomplexität ist hauptsächlich durch zwei Komponenten charakterisiert: chemische und strukturelle Kom- plexität der Pflanzen. Beide Komponenten werden im Besonderen durch die Pflanzenartendiversität bestimmt. Darüber hinaus beeinflussen Pflanzenstrukturen die mikroklimatischen Bedingungen, welche auf die Freisetzung und Verteilung von Pflanzendüften und damit auf die pflanzliche Duftkomplexität (=Chemische Komplexität) einwirken. Die pflanzliche Duftkom- plexität wiederum kann die Orientierung von herbivoren und karnivoren Insekten beeinflussen. Die Art und Weise, wie die chemische und strukturelle Komplexität der Pflanzen zusammen wirken, kann den Erfolg von Arthropoden bei der Nahrungs-, Eiablageplatz-oder Partnersuche und damit die Zusammensetzung der gesamten Arthropodengemeinschaft bee- influssen. Dieser Review beleuchtet integrativ die Beziehung zwischen Pflanzenartendiversität, struktureller Komplexität und chemischer Diversität der Pflanzen sowie ihrer Effekte auf Arthropoden. Dabei werden drei neue Hypothesen aufgestellt, die mögliche Beziehungen zwischen Pflanzenartendiversität, pflanzlicher struktureller Komplexität und pflanzlicher chemischer ∗ Corresponding author. Tel.: +49 3083855910; fax: +49 3083853897. E-mail address: [email protected] (T. Meiners). 1439-1791/$ – see front matter © 2010 Gesellschaft für Ökologie. Published by Elsevier GmbH. All rights reserved. doi:10.1016/j.baae.2010.03.003
-
Upload
uni-bayreuth -
Category
Documents
-
view
0 -
download
0
Transcript of Vegetation complexity—The influence of plant species diversity and plant structures on plant...

R
Vs
B
a
b
R
A
dtacTvbcdd
fo
Z
phPpdNicm
1d
Basic and Applied Ecology 11 (2010) 383–395
EVIEW
egetation complexity—The influence of plant species diversity and planttructures on plant chemical complexity and arthropods
arbara Randlkofera, Elisabeth Obermaierb, Monika Hilkera, Torsten Meinersa,∗
Freie Universität Berlin, Institute of Biology, Applied Zoology/Animal Ecology, Haderslebener Str. 9, 12163 Berlin, GermanyUniversity of Würzburg, Department of Animal Ecology and Tropical Biology, Germany
eceived 5 April 2009; accepted 8 March 2010
bstract
Vegetation complexity is characterized by two major traits, i.e., plant chemical and plant structural complexity. Plant speciesiversity strongly determines these traits. Furthermore, plant structures affect microclimatic conditions, which in turn influencehe emission and dispersion of plant volatiles (e.g., chemical complexity). Plant volatile chemical complexity may significantlyffect orientation of herbivorous and carnivorous arthropods. Therefore, the way in which plant chemical and plant structuralomplexity act “in concert” may influence foraging and mating success of arthropods, and thus, finally, community composition.his review emphasizes an integrative view on the relationship between plant species diversity, plant structural complexity, plantolatiles (chemical complexity) and their effects on arthropods. Three new hypotheses are raised, which predict possible relationsetween plant volatile complexity and plant species diversity: (1) saturation-, (2) step-by-step, (3) incoherence-hypothesis. Weonclude that arthropod orientation in natural environments is strongly determined by the relationship between plant volatileiversity and plant species diversity. Furthermore, we emphasize that structural complexity of the vegetation affects plant volatileiversity and thus, arthropod orientation.
We review available information on how insects actually respond to complexity during olfactory and visual search and askor both laboratory and field studies to further unravel the mechanisms of interactions between vegetation traits and their impactn arthropod orientation.
usammenfassung
Die Vegetationskomplexität ist hauptsächlich durch zwei Komponenten charakterisiert: chemische und strukturelle Kom-lexität der Pflanzen. Beide Komponenten werden im Besonderen durch die Pflanzenartendiversität bestimmt. Darüberinaus beeinflussen Pflanzenstrukturen die mikroklimatischen Bedingungen, welche auf die Freisetzung und Verteilung vonflanzendüften und damit auf die pflanzliche Duftkomplexität (=Chemische Komplexität) einwirken. Die pflanzliche Duftkom-lexität wiederum kann die Orientierung von herbivoren und karnivoren Insekten beeinflussen. Die Art und Weise, wieie chemische und strukturelle Komplexität der Pflanzen zusammen wirken, kann den Erfolg von Arthropoden bei der
ahrungs-, Eiablageplatz-oder Partnersuche und damit die Zusammensetzung der gesamten Arthropodengemeinschaft bee-nflussen. Dieser Review beleuchtet integrativ die Beziehung zwischen Pflanzenartendiversität, struktureller Komplexität undhemischer Diversität der Pflanzen sowie ihrer Effekte auf Arthropoden. Dabei werden drei neue Hypothesen aufgestellt, dieögliche Beziehungen zwischen Pflanzenartendiversität, pflanzlicher struktureller Komplexität und pflanzlicher chemischer
∗Corresponding author. Tel.: +49 3083855910; fax: +49 3083853897.E-mail address: [email protected] (T. Meiners).
439-1791/$ – see front matter © 2010 Gesellschaft für Ökologie. Published by Elsevier GmbH. All rights reserved.oi:10.1016/j.baae.2010.03.003

384 B. Randlkofer et al. / Basic and Applied Ecology 11 (2010) 383–395
Diversität vorhersagen: (1) Sättigungs-, (2) Stufenmodell-, (3) Inkohärenz-Hypothese. Wir kommen dabei zum Schluss, dass dieOrientierung von Arthropoden in natürlichen Umwelten durch den Zusammenhang von Pflanzenartendiversität und pflanzlicherDuftdiversität der Vegetation bestimmt wird. Es bedarf weiterer Untersuchungen in Labor und Freiland um die Mechanismen derInteraktionen zwischen diesen Merkmalen der Vegetation und ihrer Wirkung auf die Orientierung von Arthropoden aufzuklären.
© 2010 Gesellschaft für Ökologie. Published by Elsevier GmbH. All rights reserved.K ; Odou
I
haattaht&Hpat(
ecrtpticdTtcp(d(crdrhtoAScso&
&2
sheabwi(
may influence diversity of plant structures, e.g., plant archi-tecture or connectivity of plant parts, which may affectmobility of arthropods, and thus, colonization of habitats byarthropods (Kareiva 1983; Dennis, Aspinall, & Gordon 2002;
eywords: Habitat complexity; Plant structures; Chemical diversity
ntroduction
An important question in current ecological research isow plant species composition of vegetation influencesrthropod individuals, populations and communities. Thenswer to this question can provide insight in the mechanismshat shape interactions in complex food webs. Understandinghe effects of vegetation complexity on plant–arthropod inter-ctions is fundamental to manipulate environmental spatialeterogeneity and to modify managed habitats to main-ain ecosystem functioning and stability (Butler, Vickery,
Norris 2007; Tylianakis, Tscharntke, & Lewis 2007;ambäck, Björkman, Rämert, & Hopkins 2009). However,redictions of the influence of vegetation complexity onrthropods are difficult, because of the scarce knowledge ofhe interacting mechanisms in diverse habitats in the fieldPotting, Perry, & Powell 2005).
Several hypotheses have been raised to generalize orxplain the relationship between vegetation complexity andolonization of a habitat by arthropods. The associational-esistance hypothesis (Tahvanainen & Root 1972) andhe resource-concentration hypothesis (Root 1973) bothredict that plants in highly diverse plant communi-ies experience lower herbivore infestations than plantsn less diverse communities. According to the resource-oncentration hypothesis the host plant patch size and theistribution of host plants affect the degree of infestation.he composition of non-host plant vegetation surrounding
he host plants of an herbivorous arthropod has not beenonsidered in this hypothesis. The natural enemy hypothesisredicts a greater abundance and diversity of natural enemiespredators, parasitoids) in more diverse plant communitiesue to the greater availability of prey and microhabitatsRoot 1973). The disruptive crop hypothesis and the traprop hypothesis (Vandermeer 1989) were proposed to explainesults found in agricultural fields where interplanting ofisruptive (or trap) crops that repel (or attract) herbivo-ous arthropods may reduce damage of the target crops byerbivorous pest insects. A wide range of studies inves-igated the impact of heterogeneity of plant communitiesn arthropod orientation and performance (for reviews seegrawal, Lau, & Hambäck 2006; Kagata & Ohgushi 2006).ubstantial knowledge has been provided by studies of agroe-
osystems, where crop diversification is used as a tool touppress herbivorous pest insects or to enhance effectivenessf their antagonists (for reviews see Andow 1991; StampsLinit 1998; Landis, Wratten, & Gurr 2000; Sunderland
Fdaa
r plumes; Arthropod orientation; Herbivore–carnivore interactions
Samu 2000; Hooks & Johnson 2003; Norris & Kogan005).
In the above-mentioned hypotheses and agroecologicaltudies, the underlying proximate mechanisms acting uponerbivores and their natural enemies are little known. How-ver, this knowledge is required when trying to understandnd to predict outcomes of interactions between plants, her-ivorous arthropods, and their natural enemies. In this review,e refer to “vegetation complexity” when addressing chem-
cal and structural complexity of the vegetation in a habitatFig. 1).
Several studies have shown how plant species diversity
ig. 1. Outline of aspects addressed in this review. Plant speciesiversity affects vegetation complexity (=plant chemical complexitynd plant structural complexity), which in turn affects herbivorousnd carnivorous arthropods.

Applie
LRIdtaDh(O2
ic&t(soIopdMiEtaa
nmwttsr
tsnurtoL&csv2
bise
ptvippdfiiavta
Ec
aes2hvu(csaTadpffAitM
o&vccm(2uoi
B. Randlkofer et al. / Basic and
angellotto & Denno 2004; Meiners & Obermaier 2004;andlkofer, Jordan, Mitesser, Meiners, & Obermaier 2009).
n contrast, studies addressing the influence of plant speciesiversity on plant volatiles within a habitat are scarce, evenhough effects of plant metabolites on arthropod behaviournd performance have been studied intensively (Després,avid, & Gallet 2007). Especially plant volatiles of bothost and non-host plants may guide arthropod orientatione.g., Dethier 1982; Visser 1986; Vet & Dicke 1992; Gohole,verholt, Khan, Pickett, & Vet 2003; Steidle & van Loon003; Bruce, Wadhams, & Woodcock 2005).
Within a habitat, the composition and chemical complex-ty of the volatile bouquet might change with plant speciesomposition (Hadacek 2002; Dudareva, Negre, Nagegowda,
Orlova 2006), but also the soil and microorganisms inhe soil might be important modulators of volatile bouquetse.g., Karlovsky 2008). Possible interactions between plantpecies diversity, volatile chemical complexity, and arthropodrientation have not been addressed so far in this context.n addition to plant species diversity, the plant structuresf the vegetation might influence the patterns of emittedlant volatiles, since they may change microclimatic con-itions within habitats, and thus, evaporation of volatiles.oreover, plant structures can alter odour plume character-
stics like shape, size, and linearity (for reviews see Murlis,lkinton, & Cardé 1992; Vickers 2006). Thus, plant struc-
ures may interfere with the ability of an arthropod to locaten odour source or a plant with prey in case of a carnivorousrthropod.
The volatiles that are emitted by a given vegetation needot necessarily be the same an arthropod experiences whenoving through the vegetation. Due to its limited equipmentith sensory receptors and due to sensory filter mechanisms
he active odour space might be much more restricted. Thushe volatile complexity in a habitat also depends on the sen-ory capabilities and filter mechanisms of the animal at theeceiving end.
In highly diverse habitats, host plant and prey loca-ion might require higher sensory power (advanced sensoryystems) or better sensory integrative abilities (advancedeurological processing) due to the presence of variousnspecific plant cues (Bernays 2001; Hadacek, 2002). Theesponse of arthropods to volatile cues might be modified byhe overall odour bouquet composed of host and non-hostdours (Visser 1986; Hadacek, 2002; Meiners, Wäckers, &ewis 2003; Zhang & Schlyter 2004; Randlkofer, Obermaier,
Meiners 2007; Schröder & Hilker 2008). Physiologi-al studies revealed that generalist and specialist herbivoreshow sensitivity to both ubiquitous and taxon-specific plantolatiles (Barata, Mustaparta, Pickett, Wadhams, & Araujo002; Mustaparta 2002).
The influence of volatile complexity on arthropods has
een tested mainly in the laboratory. Almost no field stud-es are available so far. Moreover, interactions between planttructural and plant volatile complexity have not been consid-red yet. This review presents such an “integrative view” onGpg2
d Ecology 11 (2010) 383–395 385
lant structures and plant volatiles and investigates how struc-ures change the patterns of volatile emission (and thus plantolatile complexity) and how this in turn might affect forag-ng arthropods. We put forward three hypotheses that predictossible relationships between plant volatile complexity andlant species diversity. First we outline how plant volatileiversity is generated and how it could be measured in theeld. Then we provide information on how this complexity
nfluences insect behaviour by reviewing studies of the for-ging behaviour of arthropods in the field. We briefly refer toisual attributes of plants, since plant diversity and vegeta-ion complexity can affect not only olfactory but also visualrthropod orientation.
ffects of plant species diversity on volatilehemical complexity
The biochemistry of plant components is characterised byn enormous diversity and variability of compounds. Differ-nt plant species vary largely in quantity and quality of theirecondary metabolites (for reviews see Hadacek 2002; Ober005). Approximately 200,000 secondary plant compoundsave been identified to date (Ober 2005). Among these,olatile compounds are characterized by their low molec-lar weight, lipophilic character and high vapour pressurePichersky, Noel, & Dudareva 2006). About 1700 volatileonstituents from more than 90 plant families were detectedo far by headspace analyses (for reviews see Dudareva etl. 2006; Knudsen, Eriksson, Gershenzon, & Stahl 2006).hree major groups of plant volatiles can be differenti-ted: terpenoids, phenylpropanoids/benzenoids and fatty aciderivatives (Dudareva et al. 2006). Some of the secondarylant compounds appear to be ubiquitous to all plant species,or example, the green leaf volatiles (GLVs), which areormed in high amounts after damage of plant tissue (Visser &vé 1978; Matsui 2006). However, the quantitative and qual-
tative pattern of emitted GLVs and the response of insectso these compounds are species specific (e.g., Dickens 2000;
üller & Hilker 2000; Reinecke, Ruther, & Hilker 2002).The composition of odour bouquets can differ largely from
ne plant species to another (e.g., Cornu et al. 2001; SartorattoAugusto 2003; Bukovinszky, Gols, Posthumus, Vet, &
an Lenteren 2005). Hence, the composition and chemi-al complexity of the volatile bouquet of a habitat mighthange with species composition. Further, specific secondaryetabolites are found predominantly in only a few plant taxa
Schoonhoven, van Loon, & Dicke 2005; Pichersky et al.006). Glucosinolates, for example, whose hydrolysis prod-cts have volatile character, occur almost exclusively in therder Capparales, which includes the well-studied plant fam-ly Brassicaceae (for reviews see Renwick 2002; Halkier &
ershenzon 2006). Glucosinolates are further a good exam-le of a class of compounds, which is highly diverse within aroup of closely related species (Fahey, Zalcmann, & Talalay001; Benderoth et al. 2006).
3 Applie
cso(Ibvu(sRctwocT2
bsdd(2sdd2ps
ssopiIoascet
c(aSiWAa
t
owe(rr2
Pp
dfTcloptiTcif(
hoefcspavtTnwrtcravcs
p
86 B. Randlkofer et al. / Basic and
In addition to interspecific variation of volatile plantompounds, intraspecific variation between cultivars of apecies, between individual plants or even between devel-pmental stages may enhance the diversity of odour blendsGouinguené, Degen, & Turlings 2001; Lou et al. 2006).ntraspecific variation of plant odours may also be reflectedy plant chemotypes which differ in their amount and type ofolatile compounds. For instance, chemotypes occur in pop-lations of thyme (Thymus vulgaris, Lamiaceae) or yarrowAchillea millefolium, Asteraceae) and Scots pine trees (Pinusylvestris, Pinaceae) (Theis & Lerdau 2003; Orav, Arak, &aal 2006; Thoss, O’Reilly-Wapstra, & Iason 2007). Plantsan show a high degree of phenotypic plasticity with respecto the development and activation of certain chemical path-ays. Furthermore, environmental conditions like soil typer nutrient and water availability might contribute to intraspe-ific variation in plant volatile emission (e.g., Gouinguené &urlings 2003; Schmelz, Alborn, Engelberth, & Tumlinson003).
The variability of synthesis and emission of volatilesetween different plant parts may add further to the diver-ity of habitat odour bouquets. On the intraplant level,ifferent parts may emit qualitatively and quantitativelyifferent odour bouquets: leaves, buds, flowers or seedsKnudsen, Tollsten, & Bergström 1993; Irwin & Dorsett002; Pichersky & Gershenzon 2002). Abiotic or biotic stressuch as pathogen infection, herbivore feeding, insect eggeposition or plant-to-plant danger signaling (e.g., Dicke,e Boer, Höfte, & Rocha-Granandos 2003; Dicke & Hilker003; Heil & Ton 2008; Hilker & Meiners 2009) can tem-orarily alter the volatile blend by the induction of chemicalignals in the plants concerned.
Analysis of chemical complexity of plant volatiles istrongly determined by the sampling technique. Headspaceampling techniques are integrating chemical informationver time and do not represent adequately the chemical com-ounds that an arthropod experiences from the environmentn a certain moment (Riffell, Abrell, & Hildebrand 2008).n the field, arthropods most likely encounter only a fractionf the components collected. Real time measurements withdvanced techniques such as proton transfer reaction-masspectrometry (PTR-MS) or the Z-nose (Tholl et al. 2006)ould help to provide a more realistic assessment of the odournvironment which herbivores and carnivores experience inhe field.
It is possible to adopt accepted diversity indices to describehemical complexity (see Box 1, Appendix A): The ShannonShannon 1948; Krebs 1989) index, also after modificationpplied as chemodiversity index (Iason et al. 2005), and theimpson index of diversity (Simpson 1949). With the Horn
ndex (also known as Morisita–Horn index, cf. Cramer &illig 2005) the similarity of two bouquets can be compared.
ll these indices include the number of compounds detectednd their quantities.In addition to the indices, multivariate methods can be used
o analyse similarities and dissimilarities of complex patterns
wtmr
d Ecology 11 (2010) 383–395
f volatiles: for example, hierarchic clustering combinedith multidimensional scaling methods (e.g., Bukovinszky
t al. 2005), principal component and discriminant analysise.g., Mumm, Tiemann, Schulz, & Hilker 2004), or split-plotepeated measures analysis of variance and multivariate log-atio analyses (e.g., Pareja, Moraes, Clark, Birkett, & Powell007).
lant structures, odour blends, and odourlumes
In the field, the dynamics of airflows are a product ofirect interaction between the structure of the ground sur-ace and the physical characteristics of the air (Moen 1974).urbulences are either the result of thermal or mechanicalonditions prevailing in the environment. Thermal turbu-ences are caused by convective currents as a consequencef variations in air temperature. Convective currents abovelant canopies can develop when solar radiation hits onlyhe outer areas of a plant canopy, but does not reach thenner areas or the ground (Sun 1999; Zhang et al. 2007).his can also have profound implications for microclimaticonditions above and within plant canopies in terms of lightntensity, relative humidity, and soil moisture, which mighturther influence the spreading and perception of volatilesPlaza et al. 2005; Llusia, Penuelas, Alessio, & Estiarte 2006).
Vegetation characteristics such as height, density and shapeave a major impact on variations in wind velocity and thus,n turbulence intensity (Moen 1974; Whicker et al. 2002; Lit al. 2005). Homogeneous canopies form rather smooth sur-aces with low turbulence intensity, whereas heterogeneousanopies form rough surfaces with high turbulence inten-ity above or within. High turbulence intensity means thatlant volatiles are mixed up more intensively, carried away,nd diluted quickly, while under low turbulence intensity theolatiles stay longer near the source, and the source of attrac-ive odours can be detected easier by searching arthropods.he wind speed in dense grass meadows slows down expo-entially with decreasing height above ground, and the almostindless zone a few centimeters above the ground is inter-
upted frequently by gusts (Aylor, Wang, & Miller 1993). Ashe long and flexible leaves and stems of the grasses usuallyause an upward distraction of the airflow, odorant particleseleased by grass canopies may be quickly dispersed into thetmosphere (Conover 2007). In contrast, in forb-dominatedegetation with a high leaf density at the top level of theanopy, the airflow is slowed down already at the canopyurface.
An odour plume is formed when volatile chemical com-ounds are emitted by a source and are carried away by the
ind. An odour may be emitted continuously or intermit-ently. On a larger scale (>1 m) the shape of the plume as aeandering cloud may affect the odour-induced anemotactic
esponses exhibited by walking or flying arthropods orienting

Applie
iapia&&2atsdmbome(IatoacmtbK
wfbHdnmtS&
Pdc
tpisJIi1po
oi
(
(
B. Randlkofer et al. / Basic and
n upwind direction (reviewed in Cardé & Willis 2008). Onsmaller scale, the fluctuating odour concentrations withinlume stream filaments may influence the neural process-ng of the odour signal and the behavioural orientation ofrthropods to the source (Mafra-Neto & Cardé 1994; Vickers
Baker 1994; Mafra-Neto & Cardé 1998; Geier, Bosch,Boeckh 1999; Vickers, Christensen, Baker, & Hildebrand
001). Wind velocity and turbulence intensity are the mainbiotic factors influencing the shape of an odour plume andhe concentrations of compounds within the plume. Windpeed and airflow dynamics within the vegetation in turnepend on plant structures. Hence, plant structures deter-ine whether volatile plant compounds disperse in the air and
ecome thoroughly mixed within and above the plant canopyr concentrate below the canopy. Moreover, plant structuresay determine whether plant volatiles drift along a more lin-
ar guided band or spread out as a rather homogeneous cloudcf. Conover 2007; Riffell et al. 2008; and literature therein).n general, the further away from the source, the lower is usu-lly the concentration of the volatiles within the plume dueo dilution of volatiles (Murlis et al. 1992). However, studiesn the distribution of pheromone plumes in the field revealedlso that the surrounding vegetation absorbs volatile chemi-als and releases them later. Thus, releasing points of an odouray vary in the course of time, which in turn may influence
he quantitative composition of an odour that is searched fory an arthropod (Karg, Suckling, & Bradley 1994; Suckling,arg, & Bradley 1996; Perry, Wall, & Clark 1988).Flying insects can easily find their way to an odour source
hen the movement direction of plumes is linear by per-orming typical anemotactic upwind and crosswind flightehaviour (Willis & Cardé 1990; Vickers & Baker 1996).owever, in closed habitats with divergent aerodynamic con-itions (e.g., forests) individual odour segments show highlyon-linear trajectories (spreading patterns) even within a feweters. These non-linear trajectories might severely affect
he ability of arthropods to locate an odour source (Elkinton,chal, Ono, & Cardé 1987; Perry et al. 1988; Brady, Gibson,Packer 1989).
roposed relationship between plant speciesiversity, plant structural complexity, and planthemical complexity: three hypotheses
To understand how plant species diversity might be relatedo the complexity of odour blends in arthropod habitats, weropose three hypotheses that are based on accepted ecolog-cal hypotheses explaining the observed relations betweenpecies diversity and ecosystem function (see Lawton 1994;ohnson, Vogt, Clark, Schmitz, & Vogt 1996; Naeem 1998).n these theories adding new species can have different
mpacts on ecosystem function (e.g., Ehrlich & Ehrlich981). Similarly, in the vegetation complexity hypothesesut forward here adding a plant species with a certain setf chemical compounds and plant structures may increased Ecology 11 (2010) 383–395 387
verall vegetation complexity and, hence, the possibility ofnteractions with arthropods.
1) Saturation hypothesis (Fig. 2A). Chemical complexity ofan odour blend will rapidly increase in early successionof vegetation when plant species diversity and diversityof plant structures within the habitat increase. A prereq-uisite of this relationship is that each new plant speciesadds different chemical compounds to the overall habi-tat odour blend. This hypothesis also assumes that plantspecies diversity correlates positively with diversity ofplant structures. Plant species diversity enhances chemi-cal complexity by adding new compounds when adding anew species, and plant structures contribute to an increaseof chemical complexity by augmenting turbulence inten-sity and providing various microclimatic conditions. Forexample, studies investigating successional processesrevealed that structural heterogeneity increases duringsuccession (Cook, Yao, Foster, Holt, & Patrick 2005;Martinko, Hagen, & Griffith 2006). As plants produceonly a finite number of volatile compounds the increasein volatile chemical complexity will slow down whenthe number of plant species is high. Finally the chemicalcomplexity will approximate a plateau. This is in accor-dance with the species redundancy hypothesis (Walker1992).
2) Step-by-step hypothesis (Fig. 2B). Plant chemical com-plexity and plant structural complexity in habitatsincrease stepwise as the number of plant speciesincreases. A step is represented by the addition of anew plant taxon with a divergent type of volatile (sec-ondary) plant metabolites or plant architecture to theplant species pool of the habitat (Fig. 2B). This hypoth-esis takes into account that volatile chemical complexityand structural complexity may not change gradually withplant species diversity, but rather with the diversity ofgroups of plant taxa. Particular volatile chemical com-pounds are restricted to specific plant groups and closelyrelated plant species might release quite similar odourblends (Schoonhoven et al. 2005; Pichersky et al. 2006).Plant groups which release specific compounds mighttherefore represent distinct chemical functional groupsor “chemical guilds”.
A prerequisite for the step-by-step hypothesis might bea phylogenetic clustering of plant species showing chem-ical similarity when a community builts up. This might bemediated by habitat filtering and phylogenetic attractionof plants whose shared evolutionary history predisposesthem to share habitat tolerances and preferences (Webb,Ackerly, McPeek, & Donoghue 2002).
In addition to their chemical composition, plant groupsare distinguishable according to their spatial growth
form, for example, plant height and branching order.Hence, certain plant species may further constitute func-tional groups with respect to plant architecture (seeHooper et al. 2005). As in the keystone species hypothesis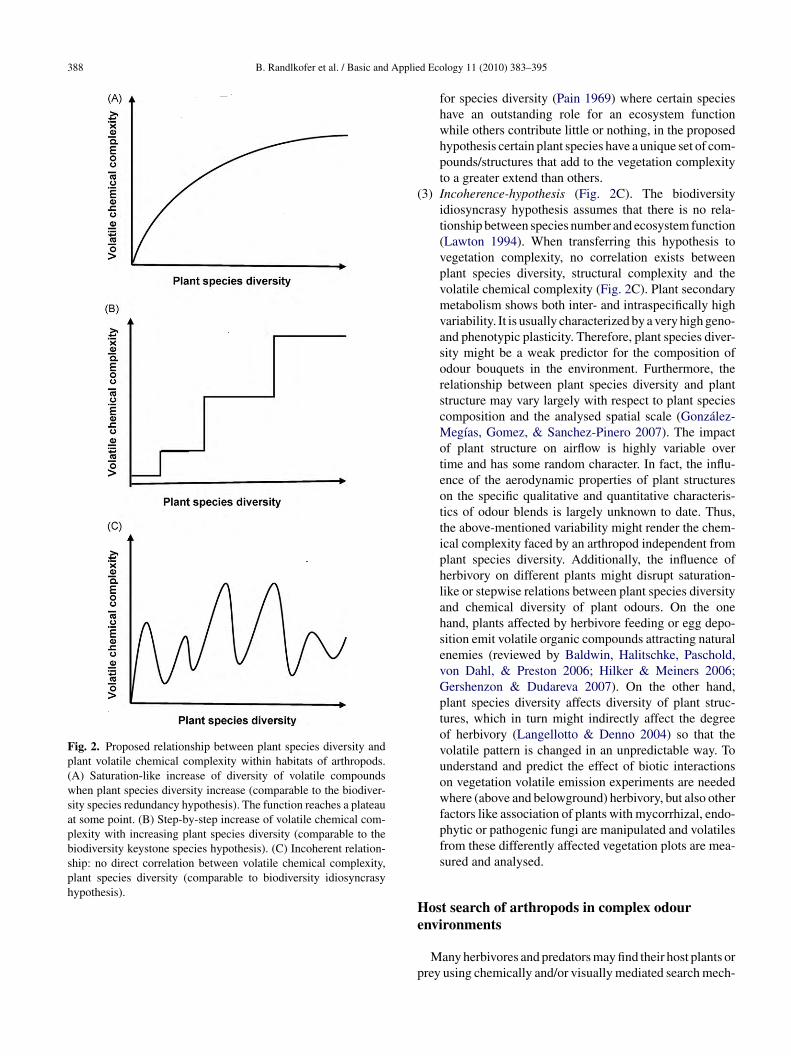
388 B. Randlkofer et al. / Basic and Applie
Fig. 2. Proposed relationship between plant species diversity andplant volatile chemical complexity within habitats of arthropods.(A) Saturation-like increase of diversity of volatile compoundswhen plant species diversity increase (comparable to the biodiver-sity species redundancy hypothesis). The function reaches a plateauat some point. (B) Step-by-step increase of volatile chemical com-plexity with increasing plant species diversity (comparable to thebiodiversity keystone species hypothesis). (C) Incoherent relation-ship: no direct correlation between volatile chemical complexity,plant species diversity (comparable to biodiversity idiosyncrasyhypothesis).
(
He
p
d Ecology 11 (2010) 383–395
for species diversity (Pain 1969) where certain specieshave an outstanding role for an ecosystem functionwhile others contribute little or nothing, in the proposedhypothesis certain plant species have a unique set of com-pounds/structures that add to the vegetation complexityto a greater extend than others.
3) Incoherence-hypothesis (Fig. 2C). The biodiversityidiosyncrasy hypothesis assumes that there is no rela-tionship between species number and ecosystem function(Lawton 1994). When transferring this hypothesis tovegetation complexity, no correlation exists betweenplant species diversity, structural complexity and thevolatile chemical complexity (Fig. 2C). Plant secondarymetabolism shows both inter- and intraspecifically highvariability. It is usually characterized by a very high geno-and phenotypic plasticity. Therefore, plant species diver-sity might be a weak predictor for the composition ofodour bouquets in the environment. Furthermore, therelationship between plant species diversity and plantstructure may vary largely with respect to plant speciescomposition and the analysed spatial scale (González-Megías, Gomez, & Sanchez-Pinero 2007). The impactof plant structure on airflow is highly variable overtime and has some random character. In fact, the influ-ence of the aerodynamic properties of plant structureson the specific qualitative and quantitative characteris-tics of odour blends is largely unknown to date. Thus,the above-mentioned variability might render the chem-ical complexity faced by an arthropod independent fromplant species diversity. Additionally, the influence ofherbivory on different plants might disrupt saturation-like or stepwise relations between plant species diversityand chemical diversity of plant odours. On the onehand, plants affected by herbivore feeding or egg depo-sition emit volatile organic compounds attracting naturalenemies (reviewed by Baldwin, Halitschke, Paschold,von Dahl, & Preston 2006; Hilker & Meiners 2006;Gershenzon & Dudareva 2007). On the other hand,plant species diversity affects diversity of plant struc-tures, which in turn might indirectly affect the degreeof herbivory (Langellotto & Denno 2004) so that thevolatile pattern is changed in an unpredictable way. Tounderstand and predict the effect of biotic interactionson vegetation volatile emission experiments are neededwhere (above and belowground) herbivory, but also otherfactors like association of plants with mycorrhizal, endo-phytic or pathogenic fungi are manipulated and volatilesfrom these differently affected vegetation plots are mea-sured and analysed.
ost search of arthropods in complex odour
nvironmentsMany herbivores and predators may find their host plants orrey using chemically and/or visually mediated search mech-
B.R
andlkoferetal./B
asicand
Applied
Ecology
11(2010)
383–395389
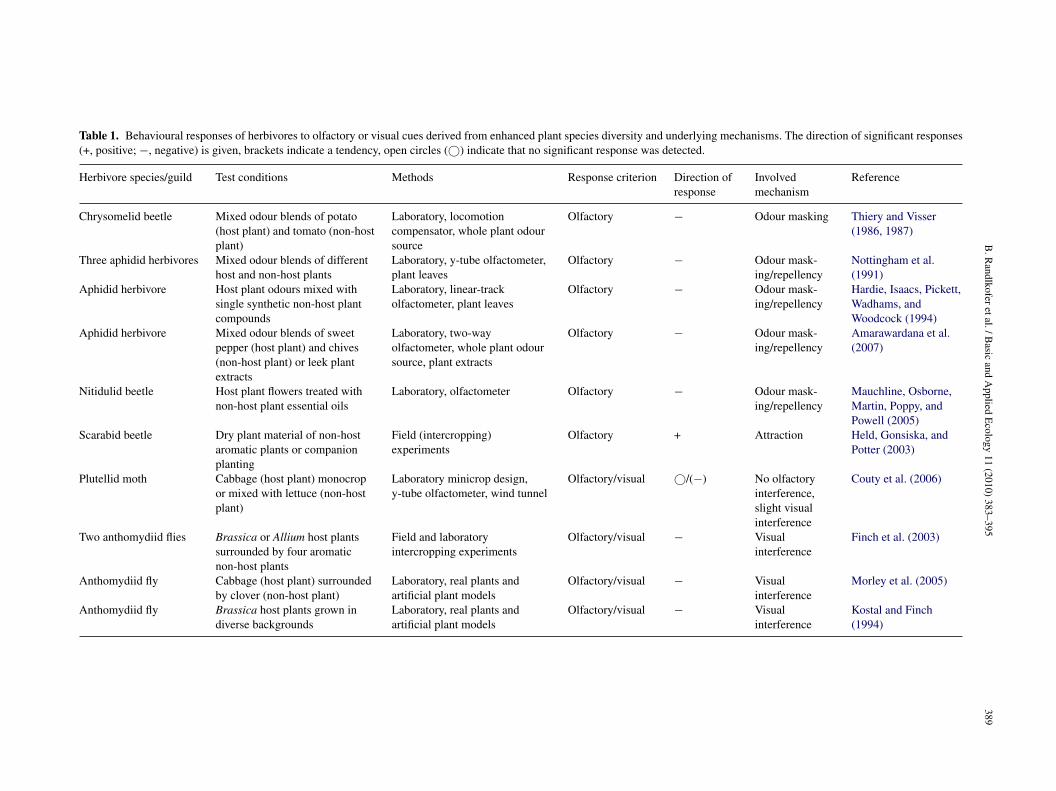
Table 1. Behavioural responses of herbivores to olfactory or visual cues derived from enhanced plant species diversity and underlying mechanisms. The direction of significant responses(+, positive; −, negative) is given, brackets indicate a tendency, open circles (©) indicate that no significant response was detected.
Herbivore species/guild Test conditions Methods Response criterion Direction ofresponse
Involvedmechanism
Reference
Chrysomelid beetle Mixed odour blends of potato(host plant) and tomato (non-hostplant)
Laboratory, locomotioncompensator, whole plant odoursource
Olfactory − Odour masking Thiery and Visser(1986, 1987)
Three aphidid herbivores Mixed odour blends of differenthost and non-host plants
Laboratory, y-tube olfactometer,plant leaves
Olfactory − Odour mask-ing/repellency
Nottingham et al.(1991)
Aphidid herbivore Host plant odours mixed withsingle synthetic non-host plantcompounds
Laboratory, linear-trackolfactometer, plant leaves
Olfactory − Odour mask-ing/repellency
Hardie, Isaacs, Pickett,Wadhams, andWoodcock (1994)
Aphidid herbivore Mixed odour blends of sweetpepper (host plant) and chives(non-host plant) or leek plantextracts
Laboratory, two-wayolfactometer, whole plant odoursource, plant extracts
Olfactory − Odour mask-ing/repellency
Amarawardana et al.(2007)
Nitidulid beetle Host plant flowers treated withnon-host plant essential oils
Laboratory, olfactometer Olfactory − Odour mask-ing/repellency
Mauchline, Osborne,Martin, Poppy, andPowell (2005)
Scarabid beetle Dry plant material of non-hostaromatic plants or companionplanting
Field (intercropping)experiments
Olfactory + Attraction Held, Gonsiska, andPotter (2003)
Plutellid moth Cabbage (host plant) monocropor mixed with lettuce (non-hostplant)
Laboratory minicrop design,y-tube olfactometer, wind tunnel
Olfactory/visual ©/(−) No olfactoryinterference,slight visualinterference
Couty et al. (2006)
Two anthomydiid flies Brassica or Allium host plantssurrounded by four aromaticnon-host plants
Field and laboratoryintercropping experiments
Olfactory/visual − Visualinterference
Finch et al. (2003)
Anthomydiid fly Cabbage (host plant) surroundedby clover (non-host plant)
Laboratory, real plants andartificial plant models
Olfactory/visual − Visualinterference
Morley et al. (2005)
Anthomydiid fly Brassica host plants grown indiverse backgrounds
Laboratory, real plants andartificial plant models
Olfactory/visual − Visualinterference
Kostal and Finch(1994)
390B
.Randlkofer
etal./Basic
andA
ppliedE
cology11
(2010)383–395
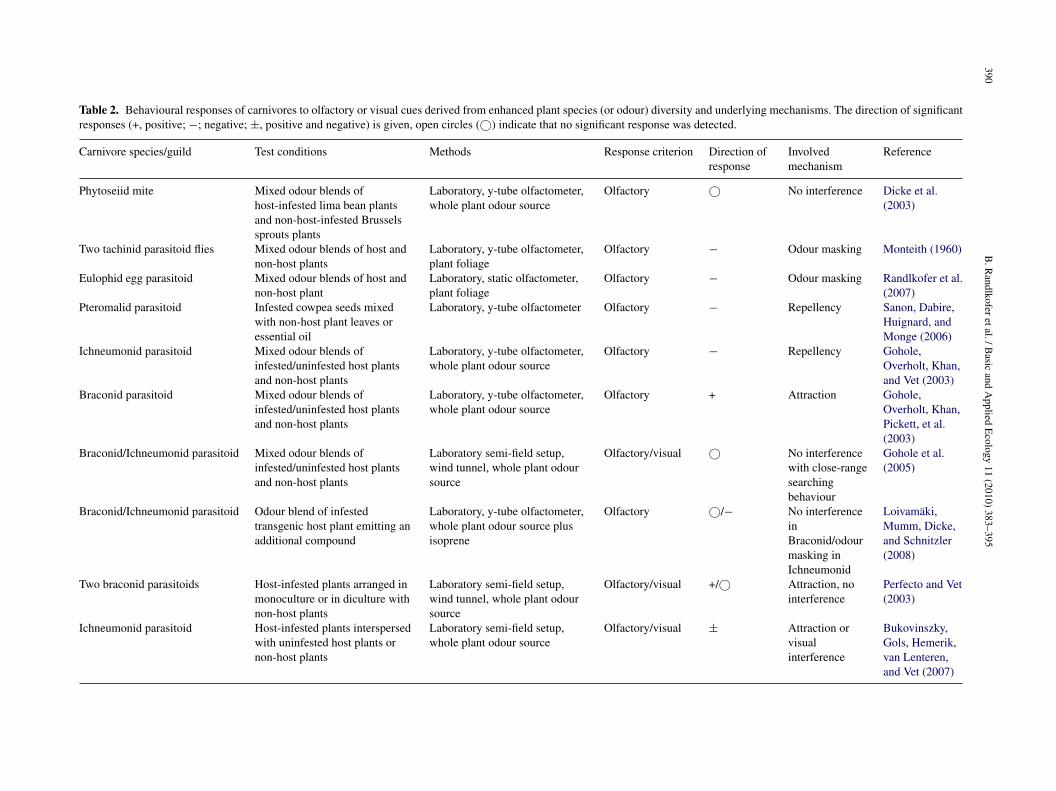
Table 2. Behavioural responses of carnivores to olfactory or visual cues derived from enhanced plant species (or odour) diversity and underlying mechanisms. The direction of significantresponses (+, positive; −; negative; ±, positive and negative) is given, open circles (©) indicate that no significant response was detected.
Carnivore species/guild Test conditions Methods Response criterion Direction ofresponse
Involvedmechanism
Reference
Phytoseiid mite Mixed odour blends ofhost-infested lima bean plantsand non-host-infested Brusselssprouts plants
Laboratory, y-tube olfactometer,whole plant odour source
Olfactory © No interference Dicke et al.(2003)
Two tachinid parasitoid flies Mixed odour blends of host andnon-host plants
Laboratory, y-tube olfactometer,plant foliage
Olfactory − Odour masking Monteith (1960)
Eulophid egg parasitoid Mixed odour blends of host andnon-host plant
Laboratory, static olfactometer,plant foliage
Olfactory − Odour masking Randlkofer et al.(2007)
Pteromalid parasitoid Infested cowpea seeds mixedwith non-host plant leaves oressential oil
Laboratory, y-tube olfactometer Olfactory − Repellency Sanon, Dabire,Huignard, andMonge (2006)
Ichneumonid parasitoid Mixed odour blends ofinfested/uninfested host plantsand non-host plants
Laboratory, y-tube olfactometer,whole plant odour source
Olfactory − Repellency Gohole,Overholt, Khan,and Vet (2003)
Braconid parasitoid Mixed odour blends ofinfested/uninfested host plantsand non-host plants
Laboratory, y-tube olfactometer,whole plant odour source
Olfactory + Attraction Gohole,Overholt, Khan,Pickett, et al.(2003)
Braconid/Ichneumonid parasitoid Mixed odour blends ofinfested/uninfested host plantsand non-host plants
Laboratory semi-field setup,wind tunnel, whole plant odoursource
Olfactory/visual © No interferencewith close-rangesearchingbehaviour
Gohole et al.(2005)
Braconid/Ichneumonid parasitoid Odour blend of infestedtransgenic host plant emitting anadditional compound
Laboratory, y-tube olfactometer,whole plant odour source plusisoprene
Olfactory ©/− No interferenceinBraconid/odourmasking inIchneumonid
Loivamäki,Mumm, Dicke,and Schnitzler(2008)
Two braconid parasitoids Host-infested plants arranged inmonoculture or in diculture withnon-host plants
Laboratory semi-field setup,wind tunnel, whole plant odoursource
Olfactory/visual +/© Attraction, nointerference
Perfecto and Vet(2003)
Ichneumonid parasitoid Host-infested plants interspersedwith uninfested host plants ornon-host plants
Laboratory semi-field setup,whole plant odour source
Olfactory/visual ± Attraction orvisualinterference
Bukovinszky,Gols, Hemerik,van Lenteren,and Vet (2007)

Applie
aBk(nSbma
ootm(neiabHpbtnwt
oTmtdOaahphrnbpC
tHhpoe
ssct
np
C
atfihgdcwFdttrectraifeh
A
tgFC1as
A
cd
R
B. Randlkofer et al. / Basic and
nisms instead of searching randomly (e.g., Saint-Germain,uddle, & Drapeau 2007). Arthropods searching for food arenown to pass through different phases of resource approach:host) habitat finding, (host) plant finding, (host) plant recog-ition, (host) plant or host acceptance (e.g., Godfray 1994;choonhoven et al. 2005). While visual cues are assumed toe of larger importance at longer distances, olfactory cuesight play a larger role at medium or short distances (withinfew decimetres to centimetres) to the host.How do insects actually respond to complexity during
lfactory search? Non-host plant odours can mask the odoursf the host plant in a way that a generally positive responseo the host plant is neutralized. The phenomenon of odour-
asking, which was described several times for herbivoresTable 1), has rarely been investigated and identified in car-ivorous arthropods (Table 2). Non-host plant odours mayven repel arthropods and thus impede host plant or host find-ng. The ratios of the offered amounts of plant material play
considerable role and already single compounds emittedy non-host plants may cause masking or repellent effects.owever, once landed on a host plant the presence of non-hostlants nearby might no longer change an arthropods foragingehaviour (e.g., Gohole, Overholt, Khan, & Vet 2005). Con-rary to masking or repellent effects, volatiles derived fromon-host plants can also attract herbivores and carnivoreshen they are also present in the host plant (Tables 1 and 2,
wo studies each).Plant diversity and vegetation complexity can affect not
nly olfactory but also visual arthropod orientation (see alsoables 1 and 2). Visual attributes of plants such as colouray affect herbivores and carnivores when orienting towards
he host plants. Similar to olfactory cues, visual signals canisrupt the host location process of insects. Prokopy andwens (1983) suggested that non-host vegetation might be
ttractive or disruptive through spectral characteristics suchs colour and could interfere with the visual apparency ofost plants. Finch and Collier (2000) developed the ‘appro-riate/inappropriate landings’ theory, which states that mosterbivorous insects are eager to land on any green object,ather than on bare soil. In more diverse vegetation a greaterumber of landings are supposed to be ‘inappropriate’,ecause they occur on non-host plants and thus, non-hostlants might disturb host plant finding (Finch, Billiald, &ollier 2003; Morley, Finch, & Collier 2005).Different studies provide evidence that visual characteris-
ics of plants have the potential to disturb host plant finding.owever, whether the ‘appropriate/inappropriate landings’ypothesis is applicable and host plant location is guidedredominantly by vision or olfaction seems also to dependn the biology of the investigated herbivore species (Coutyt al. 2006).
The examples shown in Tables 1 and 2 demonstrate that, in
pite of long evolved physiological adaptations of insects touccessfully locate their hosts by specific olfactory or visualues, these host location strategies can sometimes fail dueo high vegetation complexity. Specifically, the presence ofA
d Ecology 11 (2010) 383–395 391
on-host plant odours is able to mask the odour of the hostlant or even to repel herbivores or their natural enemies.
onclusions
We have only scarce knowledge on the interactive mech-nisms between plant structural and chemical traits and howhey affect arthropod behaviour in diverse habitats in theeld. Especially field studies, but also semi-field or green-ouse experiments as well as laboratory tests are needed toain deeper insight into the relationship between plant speciesiversity, plant structural complexity, and volatile chemicalomplexity. Well-designed experiments and field analysesill be necessary to test the three hypotheses raised here.ield experiments and analyses with varying plant speciesiversity, volatile chemical and structural complexity needo be accompanied by laboratory studies on arthropod orien-ation to a host plant in diverse plant settings. Such studiesequire the competence of experts in plant systematics, plantcology, plant chemistry, and arthropod behaviour. Closeooperation of specialists in these fields could provide bet-er insights into food web mechanisms (explaining trophicelationship) and functions (explaining biological diversitynd productivity). These studies can help to explore thenfluence of land use intensity in natural and agricultural ororest landscapes on vegetation complexity and to predict itsffect on arthropod biodiversity and the outcome of specificerbivore–natural enemy interactions.
cknowledgements
The authors thank the Berlin senate (stipends for inhe NaFöG scholarship programme and the ‘Berliner Pro-ramm zur Förderung der Chancengleichheit für Frauen inorschung und Lehre’ to B.R.) and the German Researchouncil (SPP 1374, Biodiversity Exploratories, DFG ME810/5 and DFG OB 185/2) for financial support. Threenonymous reviewers gave valuable criticism on earlier ver-ions of the manuscript.
ppendix A. Supplementary data
Supplementary data associated with this arti-le can be found, in the online version, atoi:10.1016/j.baae.2010.03.003.
eferences
grawal, A. A., Lau, J. A., & Hambäck, P. A. (2006). Commu-nity heterogeneity and the evolution of interactions betweenplants and insect herbivores. Quarterly Review of Biology, 81,349–376.

3 Applie
A
A
A
B
B
B
B
B
B
B
B
B
C
C
C
C
C
C
D
D
D
D
D
D
D
E
E
F
F
F
G
G
G
G
G
92 B. Randlkofer et al. / Basic and
marawardana, L., Bandara, P., Kumar, V., Pettersson, J., Ninkovic,V., & Glinwood, R. (2007). Olfactory response of Myzus per-sicae (Homoptera: Aphididae) to volatiles from leek and chive:Potential for intercropping with sweet pepper. Acta AgriculturaeScandinavica Section B – Soil and Plant Science, 57, 87–91.
ndow, D. A. (1991). Vegetational diversity and arthropod popula-tion response. Annual Review of Entomology, 36, 561–586.
ylor, D. E., Wang, Y. S., & Miller, D. R. (1993). Intermittentwind close to the ground within a grass canopy. Boundary-LayerMeteorology, 66, 427–448.
aldwin, I. T., Halitschke, R., Paschold, A., von Dahl, C. C., & Pre-ston, C. A. (2006). Volatile signaling in plant–plant interactions:“Talking trees” in the genomics era. Science, 311, 812–815.
arata, E. N., Mustaparta, H., Pickett, J. A., Wadhams, L. J., &Araujo, J. (2002). Encoding of host and non-host plant odoursby receptor neurones in the eucalyptus woodborer, Phoracanthasemipunctata (Coleoptera: Cerambycidae). Journal of Compar-ative Physiology A, 188, 121–133.
enderoth, M., Textor, S., Windsor, A. J., Mitchell-Olds, T., Ger-shenzon, J., & Kroymann, J. (2006). Positive selection drivingdiversification in plant secondary metabolism. Proceedings ofthe National Academy of Sciences of the United States of Amer-ica, 103, 9118–9123.
ernays, E. A. (2001). Neural limitations in phytophagous insects:Implications for diet breadth and evolution of host affiliation.Annual Review of Entomology, 46, 703–727.
rady, J., Gibson, G., & Packer, M. J. (1989). Odor movement,wind direction, and the problem of host-finding by tsetse flies.Physiological Entomology, 14, 369–380.
ruce, T. J. A., Wadhams, L. J., & Woodcock, C. M. (2005). Insecthost location: A volatile situation. Trends in Plant Science, 10,269–274.
ukovinszky, T., Gols, R., Hemerik, L., van Lenteren, J. C., &Vet, L. E. M. (2007). Time allocation of a parasitoid forag-ing in heterogeneous vegetation: Implications for host-parasitoidinteractions. Journal of Animal Ecology, 76, 845–853.
ukovinszky, T., Gols, R., Posthumus, M. A., Vet, L. E. M., & vanLenteren, J. C. (2005). Variation in plant volatiles and attractionof the parasitoid Diadegma semiclausum (Hellen). Journal ofChemical Ecology, 31, 461–480.
utler, S. J., Vickery, J. A., & Norris, K. (2007). Farmland biodi-versity and the footprint of agriculture. Science, 315, 381–384.
ardé, R. T., & Willis, M. A. (2008). Navigational strategies usedby insects to find distant, wind-borne sources of odour. Journalof Chemical Ecology, 34, 854–866.
onover, M. R. (2007). Predator-prey dynamics: The role of olfac-tion. Boca Raton: CRC Press.
ook, W. M., Yao, J., Foster, B. L., Holt, R. D., & Patrick, L. B.(2005). Secondary succession in an experimentally fragmentedlandscape: Community patterns across space and time. Ecology,86, 1267–1279.
ornu, A., Carnat, A.-P., Martin, B., Coulon, J.-B., Lamaison, J.L., & Berdagué, J.-L. (2001). Solid-phase microextraction ofvolatile components from natural grassland plants. Journal ofAgricultural and Food Chemistry, 49, 203–209.
outy, A., van Emden, H., Perry, J. N., Hardie, J., Pickett, J. A.,
& Wadhams, L. J. (2006). The roles of olfaction and vision inhost-plant finding by the diamondback moth, Plutella xylostella.Physiological Entomology, 31, 134–145.ramer, M. J., & Willig, M. R. (2005). Habitat heterogeneity, speciesdiversity and null models. Oikos, 108, 209–218.
G
d Ecology 11 (2010) 383–395
ennis, P., Aspinall, R. J., & Gordon, I. J. (2002). Spatial distributionof upland beetles in relation to landform, vegetation and grazingmanagement. Basic and Applied Ecology, 3, 183–193.
esprés, L., David, J.-P., & Gallet, C. (2007). The evolutionaryecology of insect resistance to plant chemicals. Trends in Ecologyand Evolution, 22, 298–307.
ethier, V. G. (1982). Mechanism of host plant recognition. Ento-mologia Experimentalis et Applicata, 31, 49–56.
icke, M., de Boer, J. G., Höfte, M., & Rocha-Granandos, M.C. (2003). Mixed blends of herbivore-induced plant volatilesand foraging success of carnivorous arthropods. Oikos, 101,38–48.
icke, M., & Hilker, M. (2003). Induced plant responses: Frommolecular to evolutionary biology. Basic & Applied Ecology, 4,3–14.
ickens, J. C. (2000). Orientation of Colorado potato beetle to nat-ural and synthetic blends of volatiles emitted by potato plants.Agricultural and Forest Entomology, 2, 167–172.
udareva, N., Negre, F., Nagegowda, D. A., & Orlova, I. (2006).Plant volatiles: Recent advances and future perspectives. CriticalReviews in Plant Sciences, 25, 417–440.
lkinton, J. S., Schal, C., Ono, T., & Cardé, R. T. (1987). Pheromonepuff trajectory and upwind flight of male gypsy moths in a forest.Physiological Entomology, 12, 399–406.
hrlich, P. R., & Ehrlich, A. H. (1981). Extinction: The causesand consequences of the disappearance of species. New York:Random House.
ahey, J. W., Zalcmann, A. T., & Talalay, P. (2001). The chemicaldiversity and distribution of glucosinolates and isothiocyanatesamong plants. Phytochemistry, 56, 5–51.
inch, S., Billiald, H., & Collier, R. H. (2003). Companion planting –do aromatic plants disrupt host-plant finding by the cabbage rootfly and the onion fly more effectively than non-aromatic plants?Entomologia Experimentalis et Applicata, 109, 183–195.
inch, S., & Collier, R. H. (2000). Host-plant selection by insects– a theory based on “appropriate/inappropriate landings” bypest insects of cruciferous plants. Entomologia Experimentaliset Applicata, 96, 91–102.
eier, M., Bosch, O. J., & Boeckh, J. (1999). Influence of odourplume structure on upwind flight of mosquitoes towards hosts.Journal of Experimental Biology, 202, 1639–1648.
ershenzon, J., & Dudareva, N. (2007). The function of terpenenatural products in the natural world. Nature Chemical Biology,3, 408–414.
odfray, H. C. J. (1994). Parasitoids – behavioral and evolutionaryecology. Princeton, New Jersey: Princeton University Press.
ohole, L. S., Overholt, W. A., Khan, Z. R., Pickett, J. A., & Vet,L. E. M. (2003). Effects of molasses grass, Melinis minutifloravolatiles on the foraging behavior of the cereal stemborer par-asitoid, Cotesia sesamiae. Journal of Chemical Ecology, 29,731–745.
ohole, L. S., Overholt, W. A., Khan, Z. R., & Vet, L. E. M. (2003).Role of volatiles emitted by host and non-host plants in theforaging behaviour of Dentichasmias busseolae, a pupal par-asitoid of the spotted stemborer Chilo partellus. EntomologiaExperimentalis et Applicata, 107, 1–9.
ohole, L. S., Overholt, W. A., Khan, Z. R., & Vet, L. E. M. (2005).Close-range host searching behavior of the stemborer parasitoidsCotesia sesamiae and Dentichasmias busseolae: Influence of anon-host plant Melinis minutiflora. Journal of Insect Behavior,18, 149–169.

Applie
G
G
G
H
H
H
H
H
H
H
H
H
H
I
I
J
K
K
K
K
K
K
K
K
L
L
L
L
L
L
L
M
M
M
B. Randlkofer et al. / Basic and
onzález-Megías, A., Gomez, J. M., & Sanchez-Pinero, F. (2007).Diversity-habitat heterogeneity relationship at different spatialand temporal scales. Ecography, 30, 31–41.
ouinguené, S., Degen, T., & Turlings, T. C. J. (2001). Variability inherbivore-induced odour emissions among maize cultivars andtheir wild ancestors (teosinte). Chemoecology, 11, 9–16.
ouinguené, S., & Turlings, T. C. J. (2003). The effects of abi-otic factors on induced volatile emissions in corn plants. PlantPhysiology, 129, 1296–1307.
adacek, F. (2002). Secondary metabolites as plant traits: Currentassessment and future perspectives. Critical Reviews in PlantSciences, 21, 273–322.
alkier, B. A., & Gershenzon, J. (2006). Biology and biochemistryof glucosinolates. Annual Review of Plant Biology, 57, 303–333.
ambäck, P. A., Björkman, M., Rämert, B., & Hopkins, R. J. (2009).Scale-dependent responses in cabbage herbivores affect attackrates in spatially heterogeneous systems. Basic and AppliedEcology, 10, 228–236.
ardie, J., Isaacs, R., Pickett, J. A., Wadhams, L. J., & Wood-cock, C. M. (1994). Methyl salicylate and (−)-(1R,5S)-myrtenalare plant-derived repellents for black bean aphid, Aphis fabaeScop. (Homoptera, Aphididae). Journal of Chemical Ecology,20, 2847–2855.
eil, M., & Ton, J. (2008). Long-distance signalling in plantdefence. Trends in Plant Science, 13, 264–272.
eld, D. W., Gonsiska, P., & Potter, D. A. (2003). Evaluating com-panion planting and non-host masking odors for protecting rosesfrom the Japanese beetle (Coleoptera: Scarabaeidae). Journal ofEconomic Entomology, 96, 81–87.
ilker, M., & Meiners, T. (2006). Early herbivore alert: Insecteggs induce plant defense. Journal of Chemical Ecology, 32,1379–1397.
ilker, M., & Meiners, T. (2009). How do plants “notice”attack by herbivorous arthropods? Biological Reviews,doi:10.1111/j.1469-185X.2009.00100.x
ooks, C. R. R., & Johnson, M. W. (2003). Impact of agriculturaldiversification on the insect community of cruciferous crops.Crop Protection, 22, 223–238.
ooper, D. U., Chapin, F. S., Ewel, J. J., Hector, A., Inchausti,P., Lavorel, S., et al. (2005). Effects of biodiversity on ecosys-tem functioning: A consensus of current knowledge. EcologicalMonographs, 75, 3–35.
ason, G. R., Lennon, J. J., Pakeman, R. J., Thoss, V., Beaton, J.K., Sim, D. A., et al. (2005). Does chemical composition ofindividual Scots pine trees determine the biodiversity of theirassociated ground vegetation? Ecology Letters, 8, 364–369.
rwin, R. E., & Dorsett, B. (2002). Volatile production by budsand corollas of two sympatric, confamilial plants, Ipomopsisaggregata and Polemonium foliosissimum. Journal of ChemicalEcology, 28, 565–578.
ohnson, K. H., Vogt, K. A., Clark, H. J., Schmitz, O. J., & Vogt,D. J. (1996). Biodiversity and the productivity and stability ofecosystems. Trends in Ecology & Evolution, 11, 372–377.
agata, H., & Ohgushi, T. (2006). Bottom-up trophic cascades andmaterial transfer in terrestrial food webs. Ecological Research,21, 26–34.
areiva, P. (1983). Influence of vegetation texture on herbivore pop-ulations: Resource concentration and herbivore movement. In R.F. Denno, & M. S. McClure (Eds.), Variable plants and herbi-vores in natural and managed systems (pp. 259–289). New York:Academic Press.
M
M
d Ecology 11 (2010) 383–395 393
arg, G., Suckling, D. M., & Bradley, S. J. (1994). Absorptionand release of pheromone of Epiphyas postvittana (Lepidoptera:Tortricidae) by apple leaves. Journal of Chemical Ecology, 20,1825–1841.
arlovsky, P. (2008). Secondary metabolites in soil ecology.Springer Berlin Heidelberg.
nudsen, J. T., Eriksson, R., Gershenzon, J., & Stahl, B. (2006).Diversity and distribution of floral scent. Botanical Review, 72,1–120.
nudsen, J. T., Tollsten, L., & Bergström, L. G. (1993). Floralscents – a checklist of volatile compounds isolated by headspacetechniques. Phytochemistry, 33, 253–280.
ostal, V., & Finch, S. (1994). Influence of background on host-plant selection and subsequent oviposition by the cabbage rootfly (Delia radicum). Entomologia Experimentalis et Applicata,70, 153–163.
rebs, C. (1989). Ecological methodology. New York: Harper-Collins.
andis, D. A., Wratten, S. D., & Gurr, G. M. (2000). Habitatmanagement to conserve natural enemies of arthropod pests inagriculture. Annual Review of Entomology, 45, 175–201.
angellotto, G. A., & Denno, R. F. (2004). Responses of inver-tebrate natural enemies to complex-structured habitats: Ameta-analytical synthesis. Oecologia, 139, 1–10.
awton, J. H. (1994). What do species do in ecosystems? Oikos, 71,367–374.
i, F. R., Kang, L. F., Zhang, H., Zhao, L. Y., Shirato, Y., &Taniyama, I. (2005). Changes in intensity of wind erosion at dif-ferent stages of degradation development in grasslands of InnerMongolia, China. Journal of Arid Environments, 62, 567–585.
lusia, J., Penuelas, J., Alessio, G. A., & Estiarte, M. (2006).Seasonal contrasting changes of foliar concentrations of ter-penes and other volatile organic compound in four domiantspecies of a Mediterranean shrubland submitted to a field exper-imental drought and warming. Physiologia Plantarum, 127,632–649.
oivamäki, M., Mumm, R., Dicke, M., & Schnitzler, J.-P. (2008).Isoprene interferes with the attraction of bodyguards by herba-ceous plants. Proceedings of the National Academy of Sciencesof the United States of America, 105, 17430–17435.
ou, Y. G., Hua, X. Y., Turlings, T. C. J., Cheng, J. A., Chen, X.X., & Ye, G. Y. (2006). Differences in induced volatile emis-sions among rice varieties result in differential attraction andparasitism of Nilaparvata lugens eggs by the parasitoid Ana-grus nilaparvatae in the field. Journal of Chemical Ecology, 32,2375–2387.
afra-Neto, A., & Cardé, R. T. (1994). Fine-scale structure ofpheromone plumes modulates upwind orientation of flyingmoths. Nature, 369, 142–144.
afra-Neto, A., & Cardé, R. T. (1998). Rate of realized interceptionof pheromone pulses in different wind speeds modulates almondmoth orientation. Journal of Comparative Physiology A, 182,563–572.
artinko, E. A., Hagen, R. H., & Griffith, J. A. (2006). Successionalchange in the insect community of a fragmented landscape.Landscape Ecology, 21, 711–721.
atsui, K. (2006). Green leaf volatiles: Hydroperoxide lyase path-way of oxylipin metabolism. Current Opinion in Plant Biology,9, 274–280.
auchline, A. L., Osborne, J. L., Martin, A. P., Poppy, G. M., &Powell, W. (2005). The effects of non-host plant essential oil

3 Applie
M
M
M
M
M
M
M
M
M
N
N
N
O
O
P
P
P
P
P
P
P
P
P
R
R
R
R
R
R
S
S
S
S
94 B. Randlkofer et al. / Basic and
volatiles on the behaviour of the pollen beetle Meligethes aeneus.Entomologia Experimentalis et Applicata, 114, 181–188.
einers, T., & Obermaier, E. (2004). Hide and seek on two spatialscales – vegetation structure effects herbivore oviposition andegg parasitism. Basic and Applied Ecology, 5, 87–94.
einers, T., Wäckers, F., & Lewis, W. J. (2003). Associative learn-ing of complex odours in parasitoid host location. ChemicalSenses, 28, 231–236.
oen, A. N. (1974). Turbulence and visualization of wind flow.Ecology, 55, 1420–1424.
onteith, L. G. (1960). Influence of plants other than food plantsof their host on host-finding by tachinid parasites. CanadianEntomologist, 92, 641–652.
orley, K., Finch, S., & Collier, R. H. (2005). Companion plant-ing – behaviour of the cabbage root fly on host plants andnon-host plants. Entomologia Experimentalis et Applicata, 117,15–25.
üller, C., & Hilker, M. (2000). The effect of a green leaf volatileon host plant finding by larvae of a herbivorous insect. Natur-wissenschaften, 87, 216–219.
umm, R., Tiemann, T., Schulz, S., & Hilker, M. (2004). Analysis ofvolatiles from black pine (Pinus nigra): Significance of wound-ing and egg deposition by a herbivorous sawfly. Phytochemistry,65, 3221–3230.
urlis, J., Elkinton, J. S., & Cardé, R. T. (1992). Odor plumesand how insects use them. Annual Review of Entomology, 37,505–532.
ustaparta, H. (2002). Encoding of plant odour information ininsects: Peripheral and central mechanisms. Entomologia Exper-imentalis et Applicata, 104, 1–13.
aeem, S. (1998). Species redundancy and ecosystem reliability.Conservation Biology, 12, 39–45.
orris, R. F., & Kogan, M. (2005). Ecology of interactions betweenweeds and arthropods. Annual Review of Entomology, 50,479–503.
ottingham, S. F., Hardie, J., Dawson, G. W., Hick, A. J., Pickett, J.A., Wadhams, L. J., et al. (1991). Behavioural and electrophysi-ological responses of aphids to host and plant volatiles. Journalof Chemical Ecology, 17, 1231–1242.
ber, D. (2005). Seeing double: Gene duplication and diversifica-tion in plant secondary metabolism. Trends in Plant Science, 10,444–449.
rav, A., Arak, E., & Raal, A. (2006). Phytochemical anal-ysis of the essential oil of Achillea millefolium L. fromvarious European Countries. Natural Product Research, 20,1082–1088.
ain, R. T. (1969). A note on tropic complexity and communitystability. American Naturalist, 103, 91–93.
areja, M., Moraes, M. C. B., Clark, S. J., Birkett, M. A., & Powell,W. (2007). Response of the aphid parasitoid Aphidius funebris tovolatiles from undamaged and aphid-infested Centaurea nigra.Journal of Chemical Ecology, 33, 695–710.
erfecto, I., & Vet, L. E. M. (2003). Effect of a nonhost planton the location behavior of two parasitoids: The tritrophic sys-tem of Cotesia spp. (Hymenoptera: Braconidae), Pieris rapae(Lepidoptera: Pieridae), and Brassica oleraceae. Environmental
Entomology, 32, 163–174.erry, J. N., Wall, C., & Clark, S. J. (1988). Close-range behavioor ofmale pea moths, Cydia nigircana, responding to sex-pheromonerereleased via the substrate. Entomologia Experimentalis etApplicata, 49, 37–42.
S
d Ecology 11 (2010) 383–395
ichersky, E., & Gershenzon, J. (2002). The formation and functionof plant volatiles: Perfumes for pollinator attraction and defense.Current Opinion in Plant Biology, 5, 237–243.
ichersky, E., Noel, J. P., & Dudareva, N. (2006). Biosynthesis ofplant volatiles: Nature’s diversity and ingenuity. Science, 311,808–811.
laza, J., Nunez, L., Pujadas, M., Perrez-Pastor, R., Bermejo, V.,Garcia-Alonso, S., et al. (2005). Field monoterpene emissionof Mediterranean oak (Quercus ilex). in the central IberianPeninsula measured by enclosure and micrometeorologicaltechniques: Observation of drought stress effect. Journal of Geo-physical Research-Atmospheres, 110, 1–12.
otting, R. P. J., Perry, J. N., & Powell, W. (2005). Insect behaviouralecology and other factors affecting the control efficacy of agro-ecosystem diversification strategies. Ecological Modelling, 182,199–216.
rokopy, R. J., & Owens, E. D. (1983). Visual detection of plantsby herbivorous insects. Annual Review of Entomology, 28,337–364.
andlkofer, B., Jordan, F., Mitesser, O., Meiners, T., & Obermaier,E. (2009). Effects of vegetation density, height and connectiv-ity on oviposition patterns of the leaf beetle Galeruca tanacetiL. (Coleoptera: Chrysomelidae). Entomologia Experimentalis etApplicata, 132, 134–146.
andlkofer, B., Obermaier, E., & Meiners, T. (2007). Mother’schoice of the oviposition site: Balancing risk of egg parasitismand need of food supply for the progeny with an infochemicalshelter? Chemoecology, 17, 177–186.
einecke, A., Ruther, J., & Hilker, M. (2002). The scent of food anddefence: Green leaf volatiles and toluquinone as sex attractantmediate mate finding in the European cockchafer Melolonthamelolontha. Ecology Letters, 5, 257–263.
enwick, J. A. A. (2002). The chemical world of crucivores: Lures,treats and traps. Entomologia Experimentalis et Applicata, 104,35–42.
iffell, J. A., Abrell, L., & Hildebrand, J. D. (2008). Phys-ical processes and real-time chemical measurement of theinsect olfactory environment. Journal of Chemical Ecology, 34,837–852.
oot, R. B. (1973). Organization of a plant-arthropod associationin simple and diverse habitats: The fauna of collards (Brassicaoleracea). Ecological Monographs, 43, 95–120.
aint-Germain, M., Buddle, C., & Drapeau, P. (2007). Primaryattraction and random landing in host-selection by wood-feedinginsects: A matter of scale? Agricultural and Forest Entomology,9, 227–235.
anon, A., Dabire, C., Huignard, J., & Monge, J. P. (2006). Influenceof Hyptis suaveolens (Lamiaceae) on the host location behaviorof the parasitoid Dinarmus basalis (Hymenoptera: Pteromali-dae). Environmental Entomology, 35, 718–724.
artoratto, A., & Augusto, F. (2003). Application of headspace solidphase microextraction and gas chromatography to the screen-ing of volatile compounds from some Brazilian aromatic plants.Chromatographia, 57, 351–356.
chmelz, E. A., Alborn, H. T., Engelberth, J., & Tumlinson, J. H.(2003). Nitrogen deficiency increases volicitin-induced volatile
emission, jasmonic acid accumulation, and ethylene sensitivityin Maize. Plant Physiology, 133, 295–306.choonhoven, L. M., van Loon, J. J. A., & Dicke, M. (2005).Insect-plant biology (2nd ed.). Oxford: Oxford UniversityPress.

Applie
S
S
SS
S
S
S
S
T
T
T
T
T
T
T
V
V
V
V
V
V
V
V
W
W
W
W
Z
B. Randlkofer et al. / Basic and
chröder, R., & Hilker, M. (2008). The relevance of backgroundodor in resource location by insects: A behavioural approach.Bioscience, 58, 308–316.
hannon, C. E. (1948). A mathematical theory of communication.Bell System Technical Journal, 27, 379–423, 623–656
impson, E. H. (1949). Measurement of diversity. Nature, 163, 688.tamps, W. T., & Linit, M. J. (1998). Plant diversity and arthropod
communities: Implications for temperate agroforestry. Agro-forestry Systems, 39, 73–89.
teidle, J. L. M., & van Loon, J. J. A. (2003). Dietary specializa-tion and infochemical use in carnivorous arthropods: Testinga concept. Entomologia Experimentalis et Applicata, 108,133–148.
uckling, D. M., Karg, G., & Bradley, S. J. (1996). Apple foliageenhances mating disruption of light-brown apple moth. Journalof Chemical Ecology, 22, 325–341.
un, J. L. (1999). Diurnal variations of thermal roughnessheight over a grassland. Boundary-Layer Meteorology, 92,407–427.
underland, K., & Samu, F. (2000). Effects of agricultural diversifi-cation on the abundance, distribution, and pest control potentialof spiders: A review. Entomologia Experimentalis et Applicata,95, 1–13.
ahvanainen, J. O., & Root, R. B. (1972). The influence of veg-etational diversity on the population ecology of a specializedherbivore, Phyllotreta cruciferae (Coleoptera: Chrysomelidae).Oecologia, 10, 321–346.
heis, N., & Lerdau, M. (2003). The evolution of function in plantsecondary metabolites. International Journal of Plant Sciences,164, 93–102.
hiery, D., & Visser, J. H. (1986). Masking of host plant odour in theolfactory orientation of the Colorado potato beetle. EntomologiaExperimentalis et Applicata, 41, 165–172.
hiery, D., & Visser, J. H. (1987). Misleading the Colorado potatobeetle with an odor blend. Journal of Chemical Ecology, 13,1139–1146.
holl, D., Boland, W., Hansel, A., Loreto, F., Roese, U. S. R., &Schnitzler, J.-P. (2006). Practical approaches to plant volatileanalysis. The Plant Journal, 45, 540–560.
hoss, V., O’Reilly-Wapstra, J., & Iason, G. R. (2007). Assessmentand implications of intraspecific and phenological variability in
monoterpenes of Scots pine (Pinus sylvestris) foliage. Journalof Chemical Ecology, 33, 477–491.ylianakis, J. M., Tscharntke, T., & Lewis, O. T. (2007). Habitatmodification alters the structure of tropical host-parasitoid foodwebs. Nature, 445, 202–205.
Z
d Ecology 11 (2010) 383–395 395
andermeer, J. (1989). The ecology of intercropping. Cambridge,UK: Cambridge University Press.
et, L. E. M., & Dicke, M. (1992). Ecology of infochemical useby natural enemies in a tritrophic context. Annual Review ofEntomology, 37, 141–172.
ickers, N. J. (2006). Winging it: Moth flight behavior and responsesof olfactory neurons are shaped by pheromone plume dynamics.Chemical Senses, 31, 155–166.
ickers, N. J., & Baker, T. C. (1994). Reiterative responses to sin-gle strands of odor promote sustained upwind flight and odorsource location by moths. Proceedings of the National Academyof Sciences of the United States of America, 91, 5756–5760.
ickers, N. J., & Baker, T. C. (1996). Latencies of behavioralresponse to interception of filaments of sex pheromone and cleanair influence flight track shape in Heliothis virescens (F.) males.Journal of Comparative Physiology A, 178, 831–847.
ickers, N. J., Christensen, T. A., Baker, T. C., & Hildebrand, J. G.(2001). Odour-plume dynamics influence the brain’s olfactorycode. Nature, 410, 466–470.
isser, J. H. (1986). Host odor perception in phytophagous insects.Annual Review of Entomology, 31, 121–144.
isser, J. H., & Avé, D. A. (1978). General green leaf volatilesin the olfactory orientation of the Colorado beetle, Leptino-tarsa decemlineata. Entomologia Experimentalis et Applicata,24, 738–749.
alker, B. H. (1992). Biodiversity and ecological redundancy. Con-servation Biology, 6, 18–23.
ebb, C. O., Ackerly, D. D., McPeek, M. A., & Donoghue, M. J.(2002). Phylogenies and community ecology. Annual Review ofEcology & Systematics, 33, 475–505.
hicker, J. J., Breshears, D. D., Wasiolek, P. T., Kirchner, T. B.,Tavani, R. A., Schoep, D. A., et al. (2002). Temporal and spatialvariation of episodic wind erosion in unburned and burned semi-arid shrubland. Journal of Environmental Quality, 31, 599–612.
illis, M. A., & Cardé, R. T. (1990). Pheromone-modulated opto-motor response in male gypsy moths, Lymantria dispar L.:Upwind flight in a pheromone plume in different wind velocities.Journal of Comparative Physiology A, 167, 699–706.
hang, G., Thomas, C., Leclerc, M. Y., Karipot, A., Gholz, H. L.,Binford, M., et al. (2007). On the effect of clearcuts on turbu-lence structure above a forest canopy. Theoretical and AppliedClimatology, 88, 133–137.
hang, Q. H., & Schlyter, F. (2004). Olfactory recognition andbehavioural avoidance of angiosperm nonhost volatiles byconifer-inhabiting bark beetles. Agricultural and Forest Ento-mology, 6, 1–19.




















