
Unveiling below-ground species abundance in a biodiversity experiment: a test of vertical niche...
11
Unveiling below-ground species abundance in a biodiversity experiment: a test of vertical niche differentiation among grassland species Liesje Mommer 1,2 *, Jasper van Ruijven 2 , Hannie de Caluwe 1 , Annemiek E. Smit-Tiekstra 1 , Cornelis A.M. Wagemaker 3 , N. Joop Ouborg 3 , Gerard M. Bo¨ gemann 1 , Gerard M. van der Weerden 4 , Frank Berendse 2 and Hans de Kroon 1 1 Institute for Water and Wetland Research, Experimental Plant Ecology, Radboud University Nijmegen, PO Box 9010, 6500 GL Nijmegen, The Netherlands; 2 Nature Conservation and Plant Ecology, Wageningen University, PO Box 47, 6700 AA Wageningen, The Netherlands; 3 Institute for Water and Wetland Research, Molecular Ecology, Radboud University Nijmegen, PO Box 9010, 6500 GL Nijmegen, The Netherlands; and 4 Institute for Water and Wetland Research, Botanical garden, Radboud University Nijmegen, PO Box 9010, 6500 GL Nijmegen, The Netherlands Summary 1. Plant diversity has profound effects on primary production. Plant diversity has been shown to correlate with increased primary production in nutrient-limited grassland ecosystems. This overyielding has been attributed to vertical niche differentiation among species below-ground, allowing for complementarity in resource capture. However, a rigorous test of this longstanding hypothesis is lacking because roots of different species could not be distinguished in diverse communities. 2. Here, we present the first application of a DNA-based technique that quantifies species abundances in multispecies root samples. We were thus able to compare root distributions in monocultures of two grasses and two forbs with root distributions in four-species mixtures. In order to investigate if vertical niche differentiation is driven by soil nutrient depletion, the topsoil layer of the communities were either nutrient-rich or -poor. 3. Immediately in the first year, 40% more root biomass was produced in mixtures than expected from the monocultures, together with significant below-ground complementarity effects, probably preceding above-ground overyielding. This below-ground overyielding appeared not to be the result of vertical niche differentiation, as rooting depth of the community tended to decrease, rather than increase in mixtures compared to monocultures. Roots thus tended to clump in the very dense topsoil layer rather than segregate over the whole profile in mixtures. The below-ground overyielding was mainly driven by enhanced root investments of one species, Anthoxanthum odoratum, in the densely rooted topsoil layer without retarding the growth of the other species. 4. Synthesis. Conventional ecological mechanisms, such as competition for nutrients, do not seem to be able to explain the increased root investments of A. odoratum in mixtures compared to monocultures, with apparently little effect on the root growth of the other species. Instead, the observed root responses are consistent with species-specific root recognition responses. From a community perspective, the observed early below-ground overyielding may initiate the recently reported increased soil organic matter, mineralization and N availability and thus may ultimately be responsible for the higher productivity at high plant species diversity. Key-words: biodiversity, determinants of plant community diversity and structure, ecosystem functioning, niche differentiation, nutrient depletion, root ecology, root interactions, species interactions *Correspondence author. E-mail: [email protected] Ó 2010 The Authors. Journal compilation Ó 2010 British Ecological Society Journal of Ecology 2010, 98, 1117–1127 doi: 10.1111/j.1365-2745.2010.01702.x
Transcript of Unveiling below-ground species abundance in a biodiversity experiment: a test of vertical niche...

Unveiling below-ground species abundance in a
biodiversity experiment: a test of vertical niche
differentiation among grassland species
Liesje Mommer1,2*, Jasper van Ruijven2, Hannie de Caluwe1, Annemiek E. Smit-Tiekstra1,
Cornelis A.M. Wagemaker3, N. Joop Ouborg3, Gerard M. Bogemann1,
Gerard M. van der Weerden4, Frank Berendse2 and Hans de Kroon1
1Institute for Water and Wetland Research, Experimental Plant Ecology, Radboud University Nijmegen, PO Box 9010,
6500 GL Nijmegen, The Netherlands; 2Nature Conservation and Plant Ecology, Wageningen University, PO Box 47,
6700 AA Wageningen, The Netherlands; 3Institute for Water and Wetland Research, Molecular Ecology, Radboud
University Nijmegen, PO Box 9010, 6500 GL Nijmegen, The Netherlands; and 4Institute for Water and Wetland
Research, Botanical garden, Radboud University Nijmegen, PO Box 9010, 6500 GL Nijmegen, The Netherlands
Summary
1. Plant diversity has profound effects on primary production. Plant diversity has been shown to
correlate with increased primary production in nutrient-limited grassland ecosystems. This
overyielding has been attributed to vertical niche differentiation among species below-ground,
allowing for complementarity in resource capture. However, a rigorous test of this longstanding
hypothesis is lacking because roots of different species could not be distinguished in diverse
communities.
2. Here, we present the first application of a DNA-based technique that quantifies species
abundances in multispecies root samples. We were thus able to compare root distributions in
monocultures of two grasses and two forbs with root distributions in four-species mixtures. In order
to investigate if vertical niche differentiation is driven by soil nutrient depletion, the topsoil layer of
the communities were either nutrient-rich or -poor.
3. Immediately in the first year, 40% more root biomass was produced in mixtures than
expected from the monocultures, together with significant below-ground complementarity
effects, probably preceding above-ground overyielding. This below-ground overyielding
appeared not to be the result of vertical niche differentiation, as rooting depth of the community
tended to decrease, rather than increase in mixtures compared to monocultures. Roots thus
tended to clump in the very dense topsoil layer rather than segregate over the whole profile in
mixtures. The below-ground overyielding was mainly driven by enhanced root investments of
one species, Anthoxanthum odoratum, in the densely rooted topsoil layer without retarding the
growth of the other species.
4. Synthesis. Conventional ecological mechanisms, such as competition for nutrients, do not seem
to be able to explain the increased root investments of A. odoratum in mixtures compared to
monocultures, with apparently little effect on the root growth of the other species. Instead, the
observed root responses are consistent with species-specific root recognition responses. From a
community perspective, the observed early below-ground overyielding may initiate the recently
reported increased soil organic matter, mineralization and N availability and thus may ultimately
be responsible for the higher productivity at high plant species diversity.
Key-words: biodiversity, determinants of plant community diversity and structure, ecosystem
functioning, niche differentiation, nutrient depletion, root ecology, root interactions, species
interactions
*Correspondence author. E-mail: [email protected]
� 2010 The Authors. Journal compilation � 2010 British Ecological Society
Journal of Ecology 2010, 98, 1117–1127 doi: 10.1111/j.1365-2745.2010.01702.x

Introduction
Anthropogenic exploitation of the natural environment
causes dramatic changes in the composition of ecological
communities (Chapin et al. 2000; Hooper et al. 2005). The
decline of plant diversity is one of the major issues in this
context, as plants are the primary producers in terrestrial
ecosystems. After two decades of conducting biodiversity
experiments, it is well established that plants produce more
above-ground biomass when they grow in mixtures than
would be expected from their monocultures (Cardinale et al.
2007). Enhanced biomass production in diverse communities
has been explained by selection and ⁄or complementarity
effects (Loreau & Hector 2001). The selection effect assumes
that the most productive species dominates the biomass of
the species mixtures as a result of chance. Complementarity
or facilitation occur when species exhibit niche differentia-
tion allowing for resources capture that is complementary in
space or time. Cardinale et al. (2007) statistically showed
that above-ground overyielding is largely due to species com-
plementarity.
To date it is unclear whether above-ground overyielding and
complementarity are mirrored underground, as most experi-
ments have focused on shoot biomass only. Only a few biodi-
versity studies quantified total root biomass and report mixed
results, ranging from 50% below-ground overyielding (Tilman
et al. 2001; Dimitrakopoulos & Schmid 2004; Reich et al.
2004) to no overyielding (Spehn et al. 2000; Gastine, Scherer-
Lorenzen & Leadley 2003), or even reduced biomass partition-
ing to roots in mixtures compared to monocultures (Bessler
et al. 2009). Until now, below-ground complementarity has
never been studied in diverse communities, as relative abun-
dance at the species level was unknown for root mixtures. This
omission was largely due to a lack of technical possibilities to
explore root distribution in diverse communities. We recently
developed a molecular method to quantify the proportional
abundance of different species in mixed-species root samples
(Mommer et al. 2008), allowing for tests of below-ground
complementarity.
One commonly proposed mechanism to reach functional
complementarity is vertical niche differentiation below-
ground. This, for example, is assumed to occur between deep-
rooting dicots and shallower-rooting grasses (Parrish&Bazzaz
1976; Berendse 1981, 1983; Dimitrakopoulos & Schmid 2004;
von Felten & Schmid 2008; Levine & HilleRisLambers 2009).
Complementarity would increase if roots of a given species
grew away from zones of intense nutrient competition with
neighbours (Gersani, Abramsky & Falik 1998; Semchenko,
John&Hutchings 2007b), leading to a further vertical segrega-
tion of roots of the different species, reminiscent of the two
barnacle species in the classic example of realized niche differ-
entiation (Connell 1961). Nutrient depletion would then be a
driving force behind niche differentiation (Casper, Schenk &
Jackson 2003). Dicots are thus expected to root even deeper
and grasses even shallower in species-rich mixtures than in
monocultures. Such functional complementarity in vertical
root distribution of different species should in theory lead to a
deeper and more even distribution of roots for the community
as a whole, and may lead to below-ground overyielding. This
overproduction allows amore complete exploitation of the soil
column and its available nutrients and in the end a higher
above-ground biomass production.
Here, we report about a biodiversity experiment with four
grassland species – two grasses and two dicots – with a below-
ground focus. We use the molecular technique of Mommer
et al. (2008) to unravel species abundances in root mixtures to
test the following specific hypotheses: (i) vertical root distribu-
tion will be different among the four species, and these differ-
ences will be more pronounced in mixtures than in
monocultures, leading to vertical segregation of root systems
of the different species. As it takes time for root systems to
establish, the vertical segregation will increase over time. (ii)
Vertical segregation of root systems results in a deeper and
more even distribution of roots at community level, and leads
to below-ground overyielding.
As net nitrogen mineralization typically decreases with soil
depth, most of the roots are present in the topsoil layer. There-
fore, competition for nutrients, and thus nutrient depletion, is
expected to be most intense in the topsoil. To investigate
whether rooting patterns were driven by soil nutrient deple-
tion, we included a treatment with a nutrient-poor topsoil
layer, to simulate intense nutrient competition and experimen-
tally speed up vertical niche differentiation. The third hypothe-
sis is, therefore, as follows: (iii) If nutrient depletion is
intensifying root competition and driving vertical niche differ-
entiation, segregation of rooting patterns of the species should
be enhanced in the poor topsoil treatment compared to the
nutrient-rich control.
Materials and methods
EXPERIMENTAL SETUP AND MEASUREMENTS
Two grasses, Anthoxanthum odoratum L. and Festuca rubra L., and
two forbs, Leucanthemum vulgare Lamk. and Plantago lanceolata L.,
which were also used in another biodiversity experiment (van Ruijven
& Berendse 2005), were grown in replicatedmonocultures and 1:1:1:1
mixtures in a substitutive design (Fig. S1) in plots (50 · 50 cm; 70 cm
depth; grouped by three in polyethylene containers) in the Nijmegen
Phytotron (http://www.ru.nl/phytotron). Soil depth was 64 cm,
divided in a 52-cm thick layer of a rich soil mixture (2:1:1 (v:v:v)
sand : loamy sand : potting soil) and a 12-cm layer consisting of
nutrient-poor riverine sand. The order of the layers depended on the
treatment: in the nutrient-poor topsoil treatment the sand layer was
on top of the nutrient-rich layer, whereas in the nutrient-rich topsoil
treatment the order was reversed. To determine the difference in
nutrient status in the two topsoil types, extractable nutrient content
of the freshlymixed soil was determined at the start of the experiment.
Soil samples (20 g, 20 mm diameter, 0–10 cm deep) were diluted in
50-mL demineralized water, shaken for 1 h gently to dissolve the
nutrients in the water and analysed with an Auto Analyser 3 system
(Bran+Luebbe, Norderstedt, Germany) for nitrate and phosphate.
Values of total extractable nitrogen (N) and phosphorus (P) (mg kg)1
dry soil) were 12.5±1.3 and 0.6±0.1, respectively, for the nutrient-
poor topsoil, whereas these values were 910.4±63.1 and 5.8±0.3,
respectively, for the nutrient-rich topsoil (which was also the bulk
1118 L. Mommer et al.
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127

soil; N = 5). At the bottom of all containers was a 4-cm layer with
pebbles, allowing drainage via the outlet at the bottom.
Assignment of the species (mono vs. mixture) · nutrient treat-
ments occurred randomly to a total of 36 plots (12 containers of three
plots each); resulting in monocultures N = 3–4; mixtures N = 4.
Different replicates of species · nutrients were always in different
containers. Five-week-old seedlings, originating from seeds of natural
populations occurring in the river forelands of the river Rhine, near
Nijmegen, the Netherlands, were planted in June 2006. In each plot
36 seedlings were planted (plant density 144 m2), but only the area of
the inner 4 · 4 plants (32 · 32 cm) was used for measurements to
avoid edge effects. In the growing season of 2006, plants were watered
2 L per plot three times a week with an automatic irrigation system
(PRIVA, de Lier, the Netherlands), giving constant moisture levels.
In 2007 this amount was increased to 4 L per plot, three times a week.
In winters, soil moisture levels weremaintained by wateringmanually
once a week.
In late September in 2006 and 2007, standing shoot biomass at the
end of the growing season (living and dead biomass) was harvested
by clipping 1–2 cm above soil surface, a standard practice for hay
meadows in which these species occur. In order to investigate stand-
ing root mass at the end of the growing season, soil cores (20 mm
diameter, two in monocultures, four in mixtures) were taken in seven
soil layers (0–6, 6–12, 12–18, 18–24, 24–40, 40–55, 55–66 cmdepth) at
the beginning of October 2006 and 2007. Distances to surrounding
individual plants were equal (see Fig. S1 in Supporting Information).
Roots (<2 mm) per soil increment were collected after carefully rins-
ing them with tap water. Fresh weight was determined with a micro-
balance (Sartorius, Nieuwegein, the Netherlands). Up to 100 mg of
fresh root material was stored at )80 �C for molecular analyses.
From a second subsample, root length was determined from root
scans (600 dpi, Epson expression 10000 XL; Regent Instruments,
Quebec, Canada) with WinrhizoTM software (‘very pale roots’-
option; Regent Instruments). Shoot and root dry weights were deter-
mined after drying at 70 �C for 48 h.
Quantitative real time polymerase chain reactions (RT-PCR) with
primers for non-coding species-specific markers were performed on
genomic DNA extracts from every mixed root sample separately
(Mommer et al. 2008). Using this technique, species abundance was
quantified in soil layers 1, 2, 4 and 6, comprising 60% (2006) and
80% (2007) of all root mass. Analyses were performed on the basis of
100 mg fresh rootmass. Datawere recalculated in terms of dry weight
as the ratio between dry weight and fresh weight of roots was constant
(mean±SE: 0.18±0.01,N = 50).
For the analysis ofN uptake, all shoot material of individual plants
(two per species per plot) was dried and pulverized. Of this shoot
material, 2–2.5 mg was used in a nitrogen analyser (EA 1110; Carlo
Erba – Thermo Electron, Milan, Italy) in combination with a mass
spectrometer (DEltaPlus; ThermoFinnigan, Bremen, Germany.
CALCULATIONS AND STATIST ICAL ANALYSES
All calculations and statistical analyses on roots were performed on
average plot values: i.e. root data of two cores were pooled in mono-
cultures, four cores in mixtures.
Mean rooting depth was calculated as the sum of the amount of
roots in soil layeri multiplied by the mean depth of layeri divided by
the total amount of roots in all layers. Analyses at the species level are
based on root mass density as determined from the four layers using
the RT-PCR technique. Coefficients of variation (CV) were deter-
mined as the standard deviation of root mass across the soil profile
divided by the mean. In order to compare the species results of mean
rooting depth and CV to the responses of the community as a whole,
analyses at the community scale were also based on these four soil
layers. However, analyses on the seven soil layers resulted in qualita-
tively similar results.
Net biodiversity effects, complementarity effects, selection effects
of shoot and root biomass were calculated according to Loreau &
Hector (2001). In this so-called additive partitioning method, the net
difference in yield for a mixture (DY, the net effect) is equal to the
observed yieldYOminus the expected yieldYE (i.e. the averagemono-
culture yield of the component species):
DY ¼ YO � YE
The net effect is assumed to be the sum of the complementarity
effect and the selection effect. The complementarity effect is calcu-
lated as:
N�mean ðDRYiÞ �mean ðMiÞ
in which N is the number of species, DRYi is the difference
between the observed and expected relative yield for species I and
Mi is the biomass in the monoculture of species i.
The selection effect is calculated as:
N� covarianceðDRYi;MiÞ
For roots, these biodiversity effects were calculated from the roots
in all seven soil layers. Species abundances of roots in soil layer 3 were
estimated from ratios in soil layer 4, roots in soil layers 5 and 7 from
ratios in soil layer 6. To compare observed and expected values,
deviation from expected values (D) were calculated as ln(observed))l-n(expected) for each species and layer. The expected value was always
based on the average of the monoculture(s); i.e. ¼ of the mono-
culture(s). Root : shoot ratios were calculated on the basis of above-
and below-ground biomass values perm2.
Statistical analyses were performed using full-factorial univariate
anovas (General Linear Model, SPSS 15.0), with diversity (monocul-
ture vs. mixture) and treatment (rich vs. poor topsoil layer) as fixed
factors. When analyses were performed at the species level, species
was also included as a fixed factor. Analyses were always run sepa-
rately for the two different years. In cases where D values were used,
significant levels of the intercepts of the predicted model were used to
determine whether these values deviated significantly from zero. This
was also done to evaluate net, complementarity and selection effects.
Further differences in performance (of species) between monoculture
and mixture were determined using t-tests (Compare Means, SPSS
15.0). If needed, variables were transformed in order to meet assump-
tions of anova.
Results
COMMUNITY EFFECTS: BELOW-GROUND
OVERYIELDING AND COMPLEMENTARITY
We observed a marked diversity-induced overyielding
below-ground in the rich topsoil treatment (Fig. 1; Table 1).
Immediately after the first growing season mixtures had
produced up to 35% more root biomass than expected on
the basis of the monocultures (mean relative yield total
RYT2006RichTop = 1.35). This below-ground overyielding,
also indicated by a significant net biodiversity effect (Table 1),
was mainly caused by a significant positive complementarity
effect. Selection effects were small and negative, indicating that
Below-ground species distributions in a biodiversity experiment 1119
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127

the subordinate species contributed more to the root growth
stimulation than the highly productive, dominant ones. The
positive effect of diversity on root biomass in the rich topsoil
treatment increased over the second growing season (mean
RYT2007RichTop = 1.48; Fig. 1, Table 1).
Biomass accumulation both below- and above-ground was
only slightly reduced in the poor topsoil compared to the rich
topsoil treatment (Fig. 1). However, overyielding and comple-
mentarity effects were retarded in the poor topsoil compared
to the rich topsoil as they became only significant in the second
year (meanRYT2007PoorTop = 1.62; Fig. 1, Table 1).
Above-ground, no increase of biomass production was yet
apparent in the mixtures in the first 2 years (Fig. 1, Table 1),
which appears to be common in the first years of biodiversity
experiments, particularly in the absence of nitrogen-fixing
legumes (van Ruijven & Berendse 2005). For the rich topsoil
treatment, net biodiversity effects for the shootwere even nega-
tive in the second year (Table 1). The biodiversity effects for
the community as a whole were not apparent, as clear signifi-
cant effects in roots were counterbalanced by negative effects
in the shoot.
NO EVIDENCE FOR ENHANCED VERTICAL NICHE
DIFFERENTIAT ION IN MIXTURES
Six weeks after planting, all species had grown roots already to
the deepest soil layers (minirhizotron observations, data not
shown), but most roots were allocated to the upper layers.
Over both treatments, 30% and 40% of the dicot and grass
roots, respectively, were present in the first 12 cm of soil after
Roo
t bio
mas
s (g
m–2
)
–2000
–1500
–1000
–500
0
A. odoratum F. rubraL. vulgareP. lanceolataExpectedObserved
Sho
ot b
iom
ass
(g m
–2)
0
500
1000
1500
2000
*
Rich topsoil
*
*
20072006Poor topsoil
20072006
*
Fig. 1. Above- and below-ground biomass (g dry weight m)2) for the four monocultures (simple bars), expected and observed values for the
mixtures (hatched and open stacked bars, respectively) in the rich and poor topsoil treatment at the end of the growing seasons of 2006 and 2007.
Anthoxanthum odoratum and Festuca rubra are grasses, Leucanthemum vulgare and Plantago lanceolata are dicots. The expected values are based
on the average of the monocultures. Data are mean±SE, N = 3–4. Significant differences (P < 0.05) between expected and observed values
are indicated by an asterisk, indicating significant over- or underyielding.
Table 1. Biodiversity effects for root, shoot and total biomass (g m)2), as well as shoot nitrogen (N) accumulation (g m)2), in both treatments
(rich topsoil and poor topsoil) in 2006 and 2007. Significant deviations (P < 0.05) from the intercept of the predicted model on square-root
transformed values are given in bold. Data are mean±SE,N = 3–4 plots
Rich topsoil treatment Root Shoot Total N uptakeshoot
2006 Overall net effect 112.0±27.7 71.3±124.3 183.3±112.2 )3.61±2.59
Complementarity effect 112. 5±24.8 94.1±118.2 179.9±112.2 )3.45±2.55
Selection effect )0.5±0.02 )22.8±7.2 3.4±5.0 )0.15±0.05
2007 Overall net effect 232.3±94.7 )114.0±29.9 118.3±102.5 )1.28±0.31
Complementarity effect 315.0±104.4 )142.6±32.6 26.1±100.8 )1.53±0.29
Selection effect )82.7±9.7 28.6±12.6 92.2±24.2 0.25±0.06
Poor topsoil treatment
2006 Overall net effect 5.5±46.6 162.8±69.3 168.4±73.1 )0.21±1.24
Complementarity effect 7.6±46.9 138.0±64.6 171.5±77.3 )0.61±0.96
Selection effect )2.1±0.3 24.8±6.07 )3.2±10.2 0.41±0.31
2007 Overall net effect 342.8±158.6 )13.0±71.6 329.8±150.6 )0.35±0.55
Complementarity effect 390.8±166.2 )16.4±68.6 281.6±148.8 )0.48±0.59
Selection effect )48.0±7.6 3.4±4.5 48.2±25.8 0.13±0.03
1120 L. Mommer et al.
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127
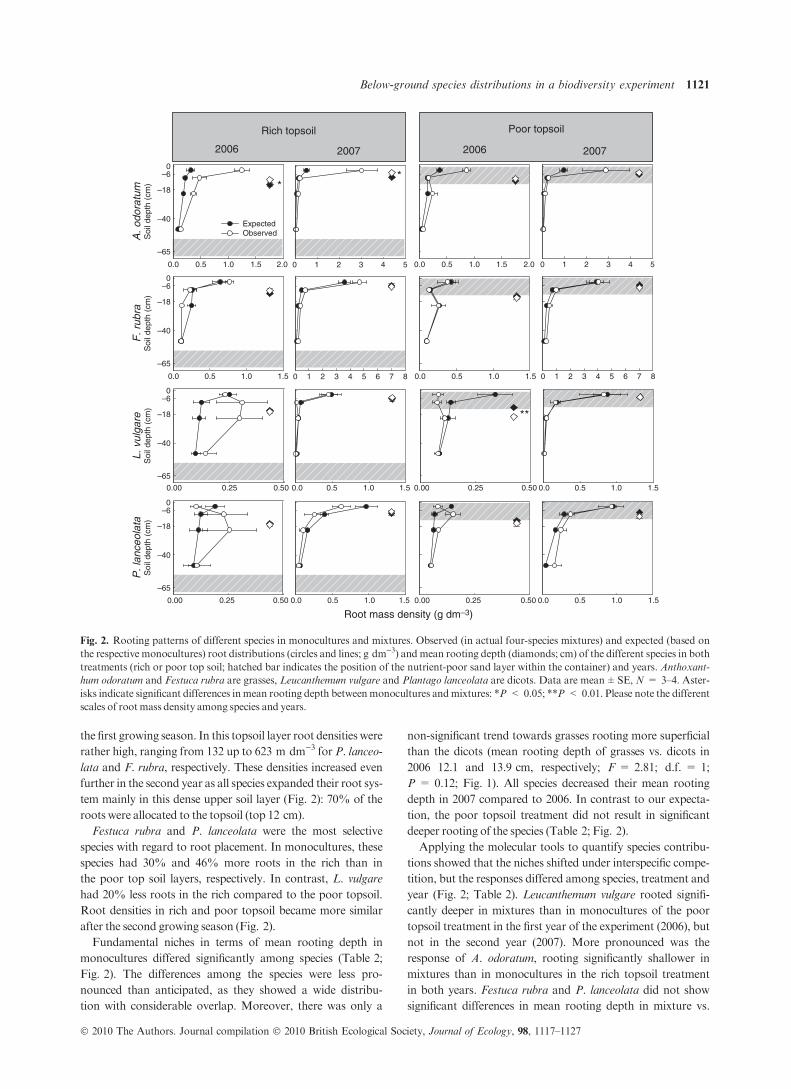
the first growing season. In this topsoil layer root densities were
rather high, ranging from 132 up to 623 m dm)3 for P. lanceo-
lata and F. rubra, respectively. These densities increased even
further in the second year as all species expanded their root sys-
tem mainly in this dense upper soil layer (Fig. 2): 70% of the
roots were allocated to the topsoil (top 12 cm).
Festuca rubra and P. lanceolata were the most selective
species with regard to root placement. In monocultures, these
species had 30% and 46% more roots in the rich than in
the poor top soil layers, respectively. In contrast, L. vulgare
had 20% less roots in the rich compared to the poor topsoil.
Root densities in rich and poor topsoil became more similar
after the second growing season (Fig. 2).
Fundamental niches in terms of mean rooting depth in
monocultures differed significantly among species (Table 2;
Fig. 2). The differences among the species were less pro-
nounced than anticipated, as they showed a wide distribu-
tion with considerable overlap. Moreover, there was only a
non-significant trend towards grasses rooting more superficial
than the dicots (mean rooting depth of grasses vs. dicots in
2006 12.1 and 13.9 cm, respectively; F = 2.81; d.f. = 1;
P = 0.12; Fig. 1). All species decreased their mean rooting
depth in 2007 compared to 2006. In contrast to our expecta-
tion, the poor topsoil treatment did not result in significant
deeper rooting of the species (Table 2; Fig. 2).
Applying the molecular tools to quantify species contribu-
tions showed that the niches shifted under interspecific compe-
tition, but the responses differed among species, treatment and
year (Fig. 2; Table 2). Leucanthemum vulgare rooted signifi-
cantly deeper in mixtures than in monocultures of the poor
topsoil treatment in the first year of the experiment (2006), but
not in the second year (2007). More pronounced was the
response of A. odoratum, rooting significantly shallower in
mixtures than in monocultures in the rich topsoil treatment
in both years. Festuca rubra and P. lanceolata did not show
significant differences in mean rooting depth in mixture vs.
0.00 0.25 0.50
0.0 0.5 1.0 1.5
0.0 0.5 1.0 1.5 2.0
ExpectedObserved
Root mass density (g dm–3)0.00 0.25 0.50
**
0.0 0.5 1.0 1.5
0 1 2 3 4 5 6 7 8
0 1 2 3 4 5
0.0 0.5 1.0 1.5
0 1 2 3 4 5 6 7 8
0 1 2 3 4 5
0.0 0.5 1.0 1.5
0.0 0.5 1.0 1.5
*
A. o
dora
tum
Soi
l dep
th (
cm)
F. r
ubra
Soi
l dep
th (
cm)
L. v
ulga
reS
oil d
epth
(cm
)P
. lan
ceol
ata
Soi
l dep
th (
cm)
0.0 0.5 1.0 1.5
0–6
–18
–40
–65
0.0 0.5 1.0 1.5 2.0
0–6
–18
–40
–65
0.00 0.25 0.50
0–6
–18
–40
–65
*
0.00 0.25 0.50
0–6
–18
–40
–65
Rich topsoil
20072006
Poor topsoil
20072006
Fig. 2. Rooting patterns of different species in monocultures and mixtures. Observed (in actual four-species mixtures) and expected (based on
the respective monocultures) root distributions (circles and lines; g dm)3) and mean rooting depth (diamonds; cm) of the different species in both
treatments (rich or poor top soil; hatched bar indicates the position of the nutrient-poor sand layer within the container) and years. Anthoxant-
hum odoratum and Festuca rubra are grasses, Leucanthemum vulgare and Plantago lanceolata are dicots. Data are mean±SE, N = 3–4. Aster-
isks indicate significant differences inmean rooting depth betweenmonocultures andmixtures: *P < 0.05; **P < 0.01. Please note the different
scales of rootmass density among species and years.
Below-ground species distributions in a biodiversity experiment 1121
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127

monoculture. While for two of the four species – A. odoratum
and (partly) L. vulgare – differences in mean rooting depth
were more pronounced in mixtures than in monocultures,
vertical segregation of root systems seemed to occur.
At the community level, the below-ground vertical segre-
gation of the two species did not result in a deeper and more
even distribution of roots. Mean rooting depth of the total
community was not increased in mixtures compared to
monocultures (Table 3). Different topsoil layers did not
affect the mean rooting depth of the community either
(Table 3). Moreover, there was a non-significant trend for
larger rather than smaller coefficients of variation of root
mass over soil layers in mixtures compared to monocultures
(Table 3), suggesting that root distributions tended to clump
rather than be more even in the diverse community.
ENHANCED ROOT GROWTH OF A. ODORATUM IN
MIXTURES
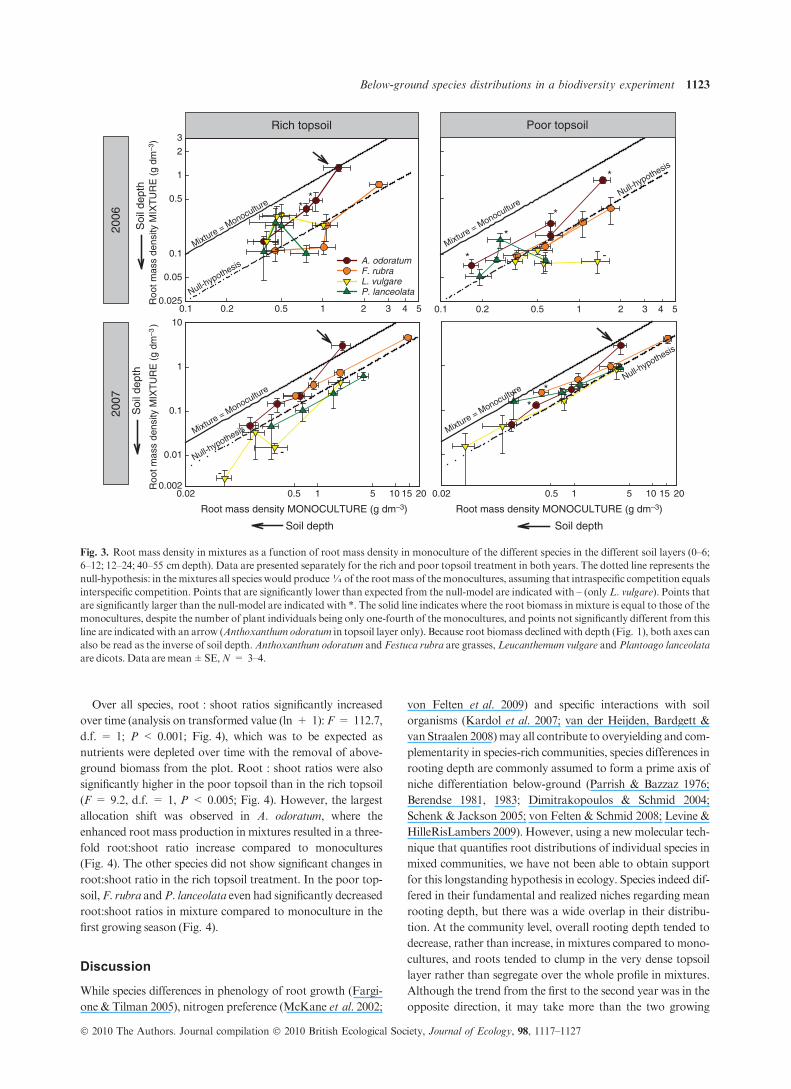
An appropriate null-hypothesis based on interspecific competi-
tive equivalence is that all species would produce¼ of the root
mass in the mixtures compared to the monocultures (dotted
line, Fig. 3). If species differ in competitive ability, some spe-
cies would do better (above the dotted line of Fig. 3), and some
worse (below the dotted line). Using the molecular tools to
unravel the species abundance in the mixtures, we found that
most root masses in mixtures were as high or higher than
expected from their respective monocultures, as most points
were above the dotted line, but not significantly different from
the 1 : 4 expectation (Fig. 3). Only a few root masses of L.
vulgare in mixtures were significantly lower than expected in
the null-hypothesis (indicated with ‘–’ in Fig. 3). In contrast,
most root masses of A. odoratum in mixtures, but also some
root masses of other species, were significantly larger than the
null hypothesis predicted (indicated with ‘*’ in Fig. 3). More-
over, the A. odoratum root masses of the topsoil were not sig-
nificantly different from the 1 : 1 line (indicated with an arrow
in Fig. 3), indicating that root mass in mixtures equals root
mass in monocultures in the top soil layer, while there were
only one-fourth of the number of individual plants in the mix-
tures than themonocultures.
In contrast to A. odoratum, the other grass species in the
experiment, F. rubra, was strikingly unresponsive to growth in
monoculture vs. mixture. Festuca rubra had by far the largest
root system (Fig. 1), but did not take advantage of the lower
root length density of the other species when growing in the
mixture (Fig. 3).
The described root patterns of A. odoratum and the other
species occurred in the rich top treatment immediately in the
first growing season, whereas in the poor topsoil treatment this
took more time. After two growing seasons, however, the pat-
terns were largely similar in the two treatments.
Table 3. Mean rooting depth (cm,MRD) and coefficient of variation of root mass over soil layers (CV) and anova results for the community as a
whole, calculated from root mass distribution over four soil layers (0–6; 6–12; 12–24; 40–55 cm depth). Treat. indicates the top soil treatment
(nutrient-rich or poor); Div. indicates the diversity level of the community (monoculture vs. four-species mixture). Mean rooting depths were
ln-transformed in order tomeet anova assumptions. *P < 0.05; †P < 0.10;N = 3–4 plots
Year Treat. Div.
MRD CV
Source D.f.
MRD CV
Mean±SE Mean±SE F-value F-value
2006 Rich top Mono 13.2±0.7 0.57±0.06 Div. 1 1.7 3.1
Mix 11.7±0.8 0.69±0.07 Treat. 1 0.3 3.9†
Poor top Mono 12.3±0.1 0.71±0.05 Div. · Tr. 1 0.6 0.1
Mix 11.9±0.7 0.80±0.04 Error (MS) 9 1.6 118.6
2007 Rich top Mono 6.4±0.02 1.36±0.02 Div. 1 0.3 0.9
Mix 6.1±0.2 1.47±0.01 Treat. 1 2.3 8.7*
Poor top Mono 6.5±0.7 1.32±0.07 Div. · Tr. 1 1.7 3.3
Mix 7.2±0.3 1.29±0.04 Error (MS) 9 0.4 46.7
Table 2. anova results, split per year, for mean rooting depth with
species, diversity (mono vs. mixtures) and treatment (i.e. rich vs. poor
topsoil) as fixed factors. Overall rooting depths were significantly
different among species. General diversity and treatment effects were
not apparent. To explore the significant interaction between species
and diversity, another anova, split per year and species, showed that
diversity effects were significant for Anthoxanthum odoratum in both
years (shallower rooting) and for Leucanthemum vulgare in 2006
(deeper rooting; analysis not shown). *P < 0.05; **P < 0.01;
***P < 0.001
Year Source d.f. F-value
2006 Species 3 17.5***
Diversity 1 0.77
Treatment 1 0.41
Species · Div 3 5.14**
Species · Treat 3 4.10*
Div · Treat 1 6.69*
Sp · Div · Treat 3 0.79
Error (d.f., MS) 38, 4.95
2007 Species 3 9.30***
Diversity 1 0.23
Treatment 1 0.14
Species · Div 3 3.95*
Species · Treat 3 1.40
Div · Treat 1 6.19*
Sp · Div · Treat 3 0.57
Error (d.f., MS) 38, 2.31
1122 L. Mommer et al.
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127
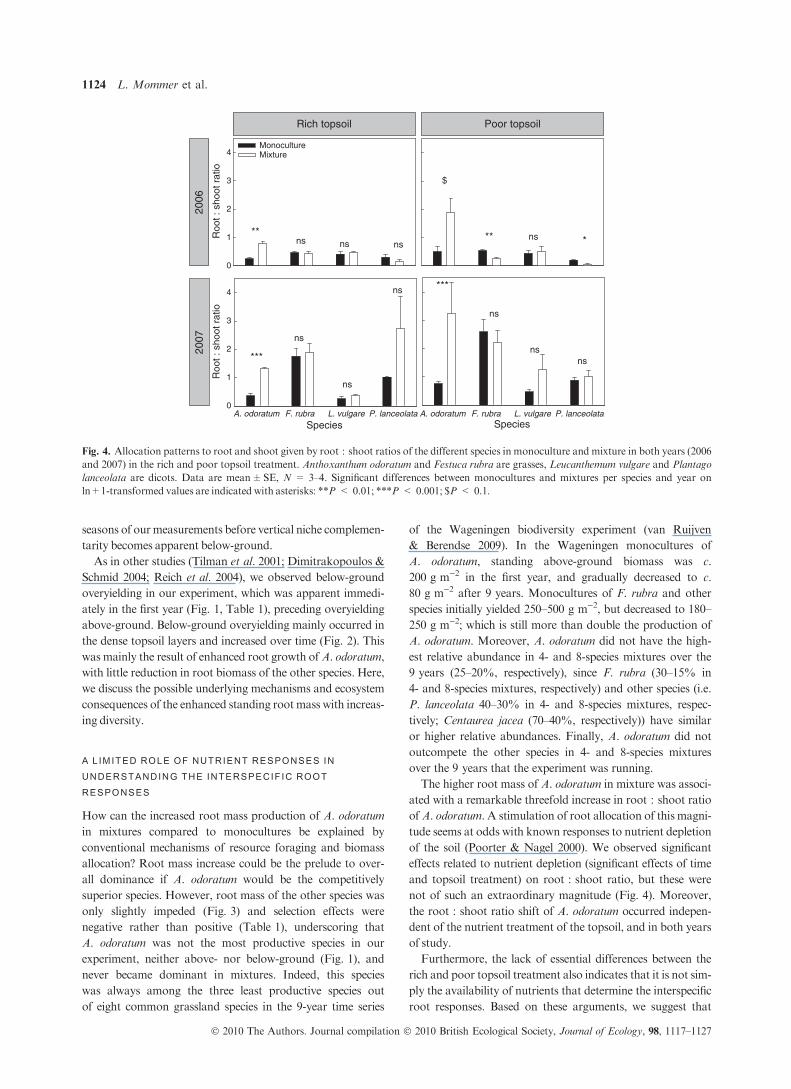
Over all species, root : shoot ratios significantly increased
over time (analysis on transformed value (ln + 1): F = 112.7,
d.f. = 1; P < 0.001; Fig. 4), which was to be expected as
nutrients were depleted over time with the removal of above-
ground biomass from the plot. Root : shoot ratios were also
significantly higher in the poor topsoil than in the rich topsoil
(F = 9.2, d.f. = 1, P < 0.005; Fig. 4). However, the largest
allocation shift was observed in A. odoratum, where the
enhanced root mass production in mixtures resulted in a three-
fold root:shoot ratio increase compared to monocultures
(Fig. 4). The other species did not show significant changes in
root:shoot ratio in the rich topsoil treatment. In the poor top-
soil, F. rubra andP. lanceolata even had significantly decreased
root:shoot ratios in mixture compared to monoculture in the
first growing season (Fig. 4).
Discussion
While species differences in phenology of root growth (Fargi-
one & Tilman 2005), nitrogen preference (McKane et al. 2002;
von Felten et al. 2009) and specific interactions with soil
organisms (Kardol et al. 2007; van der Heijden, Bardgett &
van Straalen 2008)may all contribute to overyielding and com-
plementarity in species-rich communities, species differences in
rooting depth are commonly assumed to form a prime axis of
niche differentiation below-ground (Parrish & Bazzaz 1976;
Berendse 1981, 1983; Dimitrakopoulos & Schmid 2004;
Schenk & Jackson 2005; von Felten & Schmid 2008; Levine &
HilleRisLambers 2009). However, using a newmolecular tech-
nique that quantifies root distributions of individual species in
mixed communities, we have not been able to obtain support
for this longstanding hypothesis in ecology. Species indeed dif-
fered in their fundamental and realized niches regarding mean
rooting depth, but there was a wide overlap in their distribu-
tion. At the community level, overall rooting depth tended to
decrease, rather than increase, in mixtures compared to mono-
cultures, and roots tended to clump in the very dense topsoil
layer rather than segregate over the whole profile in mixtures.
Although the trend from the first to the second year was in the
opposite direction, it may take more than the two growing
0.02 0.5 1 5 10 15 20
0.1 0.2 0.5 1 2 3 4 5
Null-hypothesis
Soil depth
-
*
*
**
Mixture = Monoculture
Mixture = Monoculture
Null-hypothesis
*
*
0.1 0.2 0.5 1 2 3 4 5
Roo
t mas
s de
nsity
MIX
TU
RE
(g
dm–3
)
0.025
0.05
0.1
0.5
1
2
3
A. odoratumF. rubraL. vulgareP. lanceolata
Mixture = Monoculture
Null-hypothesis
Root mass density MONOCULTURE (g dm–3) Root mass density MONOCULTURE (g dm–3)
0.02 0.5 1 5 10 15 20
Roo
t mas
s de
nsity
MIX
TU
RE
(g
dm–3
)
0.002
0.01
0.1
1
10
Soil depth
Soi
l dep
thS
oil d
epth
*
*
-
-Null-h
ypothesisMixture = Monoculture
*
2007
Poor topsoil20
06Rich topsoil
Fig. 3. Root mass density in mixtures as a function of root mass density in monoculture of the different species in the different soil layers (0–6;
6–12; 12–24; 40–55 cm depth). Data are presented separately for the rich and poor topsoil treatment in both years. The dotted line represents the
null-hypothesis: in themixtures all species would produce¼of the rootmass of themonocultures, assuming that intraspecific competition equals
interspecific competition. Points that are significantly lower than expected from the null-model are indicated with – (only L. vulgare). Points that
are significantly larger than the null-model are indicated with *. The solid line indicates where the root biomass in mixture is equal to those of the
monocultures, despite the number of plant individuals being only one-fourth of the monocultures, and points not significantly different from this
line are indicated with an arrow (Anthoxanthum odoratum in topsoil layer only). Because root biomass declined with depth (Fig. 1), both axes can
also be read as the inverse of soil depth.Anthoxanthum odoratum and Festuca rubra are grasses, Leucanthemum vulgare and Plantoago lanceolata
are dicots. Data are mean±SE,N = 3–4.
Below-ground species distributions in a biodiversity experiment 1123
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127
seasons of ourmeasurements before vertical niche complemen-
tarity becomes apparent below-ground.
As in other studies (Tilman et al. 2001; Dimitrakopoulos &
Schmid 2004; Reich et al. 2004), we observed below-ground
overyielding in our experiment, which was apparent immedi-
ately in the first year (Fig. 1, Table 1), preceding overyielding
above-ground. Below-ground overyielding mainly occurred in
the dense topsoil layers and increased over time (Fig. 2). This
was mainly the result of enhanced root growth ofA. odoratum,
with little reduction in root biomass of the other species. Here,
we discuss the possible underlying mechanisms and ecosystem
consequences of the enhanced standing rootmass with increas-
ing diversity.
A LIMITED ROLE OF NUTRIENT RESPONSES IN
UNDERSTANDING THE INTERSPECIF IC ROOT
RESPONSES
How can the increased root mass production of A. odoratum
in mixtures compared to monocultures be explained by
conventional mechanisms of resource foraging and biomass
allocation? Root mass increase could be the prelude to over-
all dominance if A. odoratum would be the competitively
superior species. However, root mass of the other species was
only slightly impeded (Fig. 3) and selection effects were
negative rather than positive (Table 1), underscoring that
A. odoratum was not the most productive species in our
experiment, neither above- nor below-ground (Fig. 1), and
never became dominant in mixtures. Indeed, this species
was always among the three least productive species out
of eight common grassland species in the 9-year time series
of the Wageningen biodiversity experiment (van Ruijven
& Berendse 2009). In the Wageningen monocultures of
A. odoratum, standing above-ground biomass was c.
200 g m)2 in the first year, and gradually decreased to c.
80 g m)2 after 9 years. Monocultures of F. rubra and other
species initially yielded 250–500 g m)2, but decreased to 180–
250 g m)2; which is still more than double the production of
A. odoratum. Moreover, A. odoratum did not have the high-
est relative abundance in 4- and 8-species mixtures over the
9 years (25–20%, respectively), since F. rubra (30–15% in
4- and 8-species mixtures, respectively) and other species (i.e.
P. lanceolata 40–30% in 4- and 8-species mixtures, respec-
tively; Centaurea jacea (70–40%, respectively)) have similar
or higher relative abundances. Finally, A. odoratum did not
outcompete the other species in 4- and 8-species mixtures
over the 9 years that the experiment was running.
The higher root mass of A. odoratum in mixture was associ-
ated with a remarkable threefold increase in root : shoot ratio
ofA. odoratum. A stimulation of root allocation of this magni-
tude seems at odds with known responses to nutrient depletion
of the soil (Poorter & Nagel 2000). We observed significant
effects related to nutrient depletion (significant effects of time
and topsoil treatment) on root : shoot ratio, but these were
not of such an extraordinary magnitude (Fig. 4). Moreover,
the root : shoot ratio shift of A. odoratum occurred indepen-
dent of the nutrient treatment of the topsoil, and in both years
of study.
Furthermore, the lack of essential differences between the
rich and poor topsoil treatment also indicates that it is not sim-
ply the availability of nutrients that determine the interspecific
root responses. Based on these arguments, we suggest that
SpeciesA. odoratum F. rubra L. vulgare P. lanceolata
nsns
ns
***
$
ns** ***
nsns ns
SpeciesA. odoratum F. rubra L. vulgare P. lanceolata
Roo
t : s
hoot
rat
io
0
1
2
3
4 ns
ns
ns
***
Roo
t : s
hoot
rat
io
0
1
2
3
4Monoculture Mixture
Rich topsoil Poor topsoil
2006
2007
Fig. 4. Allocation patterns to root and shoot given by root : shoot ratios of the different species in monoculture and mixture in both years (2006
and 2007) in the rich and poor topsoil treatment. Anthoxanthum odoratum and Festuca rubra are grasses, Leucanthemum vulgare and Plantago
lanceolata are dicots. Data are mean±SE, N = 3–4. Significant differences between monocultures and mixtures per species and year on
ln+1-transformed values are indicated with asterisks: **P < 0.01; ***P < 0.001; $P < 0.1.
1124 L. Mommer et al.
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127

competitive and allocation responses to soil nutrients alone are
unlikely to drive the elevated root mass in mixtures vs. mono-
cultures.
ALTERNATIVE EXPLANATIONS FOR THE SPECIES-
SPECIF IC ROOT INTERACTIONS
Recent advances in root ecology suggest that root growth and
distribution can be tuned to the identity of neighbouring roots
(Schenk 2006; de Kroon 2007) by distinguishing roots of the
same physiological individual from roots of a different geno-
type of the same species (Maina, Brown & Gersani 2002;
Gruntman&Novoplansky 2004; Dudley &File 2007) or roots
of a different species (Huber-Sannwald et al. 1998; Bartelhei-
mer, Steinlein & Beyschlag 2006; Li et al. 2007; Semchenko,
Hutchings & John 2007a). Responses to neighbours range
from inhibition to stimulation of root growth (Mahall & Call-
away 1991; Li et al. 2006; Semchenko, John & Hutchings
2007b). The mechanisms behind root recognition responses
are yet obscure, but non-nutritious cues, such as root exudates
(Bais et al. 2006) and ⁄or root rhizosphere biota (Kardol et al.
2007; van der Heijden, Bardgett & van Straalen 2008) may be
involved since the root responses appear very species-specific
and density-dependent.
We would argue that the root biomass responses of
A. odoratum are consistent with the operation of such non-
nutritious cues: growth enhancement at low densities (i.e. in
mixtures) and ⁄or inhibition of root growth at high densities
(monocultures), without affecting the root growth of the
other species. Density-dependent regulation of root growth
in monocultures and mixtures may have occurred via accu-
mulation of autotoxic root exudates (Bais et al. 2006) and ⁄orhost-plant specific pathogens (Van der Putten & Peters 1997;
Westover & Bever 2001). Alternatively, root growth of
A. odoratum in mixtures may have been stimulated by the
actual presence of roots of other species, directly, due to
growth-stimulating root exudates from these species and ⁄orindirectly, due to root growth-promoting rhizobacteria (Bais
et al. 2006; Berg & Smalla 2009). For our study species,
A. odoratum, it has been shown that seedlings grow better
on natural root growth-promoting isolates of Bacillus of
other species than on its own isolates (Westover & Bever
2001), suggesting that root growth promoting bacteria may
play a role in grassland systems. A more indirect explanation
for the enhanced root growth of A. odoratum in diverse com-
munities would be that high plant diversities affect soil
microbial communities (Kowalchuk et al. 2002; Bartelt-
Ryser et al. 2005), leading to suppression of (root) pathogens
(i.e. soil suppressiveness (Weller et al. 2002); cf. studies in
leaves (Mitchell, Tilman & Groth 2002)), although it usually
takes several years for the microbial communities to build
up (Steinbeiss et al. 2008). Such pathogens should then be
more deleterious for A. odoratum than for the other species
of our system in order to explain our results.
We conclude that our results are consistent with non-
nutritional, density-dependent feedback mechanisms as
reported for the spread of invasive species (Bais et al. 2003;
Levine et al. 2006) and plant succession (Bever 1994; Kardol
et al. 2007). However, although the interactions below-
ground may be essentially non-nutritious and depending on
species identity, the effects may improve the nutritional sta-
tus in mixtures vs. monocultures (cf. Li et al. 2007) or
increase competitiveness for soil nutrients. Future research is
needed to demonstrate what interactions are really involved
and to pinpoint the exact mechanisms in this apparent spe-
cies-specific root recognition.
ECOSYSTEM IMPLICATIONS
The ecosystem implications of the below-ground overyielding
reported here and in other studies (Tilman et al. 2001;
Dimitrakopoulos & Schmid 2004; Reich et al. 2004) may be
considerable, as the higher root mass in these grassland
mixtures appears to persist with possibly important conse-
quences for the soil C andN budget. In theWageningen biodi-
versity experiment (van Ruijven & Berendse 2005, 2009), root
masses were 40% higher in four-species mixtures than in the
average monoculture after 9 years (L. Mommer & J. van
Ruijven, unpubl. data). Similarly, in the Cedar Creek biodiver-
sity experiment, total root mass was 50–200% higher in the
eight-species mixtures than in the monocultures, 12 years after
the start of the experiment (Fornara, Tilman & Hobbie 2009).
Given the high turnover rates of roots (Van der Krift &
Berendse 2002), a higher root mass production will lead to
increased organic matter input into the soil. Indeed, after
4 years in the Jena Biodiversity Experiment, carbon stocks
were significantly higher in plots with higher species diversity
(Steinbeiss et al. 2008). In the long-term, increased organic
matter input can increase mineralization and N-availability
(Fornara, Tilman & Hobbie 2009), which in turn is held
responsible for the higher productivity in more diverse plant
communities.
Conclusions
Our experimental grassland communities revealed below-
ground species complementarity as expected, but found no
evidence for the associated vertical niche differentiation that
is generally assumed. While the exact mechanisms for the
observed root responses remain obscure, the results are
consistent with species-specific non-nutritional root interac-
tions. The root growth stimulation in species mixtures may
be the trigger to enhanced carbon sequestration and N
availability in the soil, finally leading to enhanced above-
ground production in diverse plant communities. If these
novel interactions indeed play a role in below-ground over-
yielding, they may be the drivers of fundamental ecosystem
processes.
Acknowledgements
We thank Jelle Eygensteyn for N measurements; Jan van Walsem and Frans
Moller for help during harvests and Jean Knops, Wim van der Putten, Ronald
Pierik and Eric Visser for comments on an earlier version of the manuscript.
Below-ground species distributions in a biodiversity experiment 1125
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127

L.M. is supported byNetherlands Organization for Scientific Research (NWO)
VENI grant 016091116.
References
Bais, H.P., Vepachedu, R., Gilroy, S., Callaway, R.M. & Vivanco, J.M. (2003)
Allelopathy and exotic plant invasion: from molecules and genes to species
interactions. Science, 301, 1377–1380.
Bais, H.P., Weir, T.L., Perry, L.G., Gilroy, S. & Vivanco, J.M. (2006) The role
of root exudates in rhizosphere interactions with plants and other organisms.
Annual Review of Plant Biology, 57, 233–266.
Bartelheimer, M., Steinlein, T. & Beyschlag, W. (2006) Aggregative root place-
ment: a feature during interspecific competition in inland sand-dune habi-
tats.Plant and Soil, 280, 101–114.
Bartelt-Ryser, J., Joshi, J., Schmid, B., Brandl, H. & Balser, T. (2005) Soil
feedbacks of plant diversity on soil microbial communities and subse-
quent plant growth. Perspectives in Plant Ecology Evolution and System-
atics, 7, 27–49.
Berendse, F. (1981) Competition between plant-populations with different
rooting depths 2. Pot experiments.Oecologia, 48, 334–341.
Berendse, F. (1983) Interspecific competition and niche differentiation between
Plantago lanceolata andAnthoxanthum odoratum in a natural hayfield. Jour-
nal of Ecology, 71, 379–390.
Berg, G. & Smalla, K. (2009) Plant species and soil type cooperatively shape the
structure and function of microbial communities in the rhizosphere. Fems
Microbiology Ecology, 68, 1–13.
Bessler, H., Temperton, V.M., Roscher, C., Buchmann, N., Schmid, B., Schu-
lze, E.D., Weisser, W.W. & Engels, C. (2009) Aboveground overyielding in
grassland mixtures is associated with reduced biomass partitioning to below-
ground organs.Ecology, 90, 1520–1530.
Bever, J.D. (1994) Feedback between plants and their soil communities in an
old field community.Ecology, 75, 1965–1977.
Cardinale, B.J., Wrigh, J.P., Cadotte, M.W., Carroll, I.T., Hector, A., Srivast-
ava, D.S., Loreau, M. & Weis, J.J. (2007) Impacts of plant diversity on bio-
mass production increase through time because of species complementarity.
Proceedings of the National Academy of Sciences of the United States of
America, 104, 18123–18128.
Casper, B.B., Schenk, H.J. & Jackson, R.B. (2003) Defining a plant’s below-
ground zone of influence.Ecology, 84, 2313–2321.
Chapin, F.S., Zavaleta, E.S., Eviner, V.T., Naylor, R.L., Vitousek, P.M., Rey-
nolds, H.L., Hooper, D.U., Lavorel, S., Sala, O.E., Hobbie, S.E., Mack,
M.C. & Diaz, S. (2000) Consequences of changing biodiversity.Nature, 405,
234–242.
Connell, J.H. (1961) Influence of interspecific competition and other factors on
distribution of barnacle Chthamalus stellatus.Ecology, 42, 710–717.
Dimitrakopoulos, P.G. & Schmid, B. (2004) Biodiversity effects increase line-
arly with biotope space.Ecology Letters, 7, 574–583.
Dudley, S.A. & File, A.L. (2007) Kin recognition in an annual plant. Biology
Letters, 3, 435–438.
Fargione, J. & Tilman, D. (2005) Niche differences in phenology and rooting
depth promote coexistence with a dominant C-4 bunchgrass.Oecologia, 143,
598–606.
von Felten, S. & Schmid, B. (2008) Complementarity among species in horizon-
tal versus vertical rooting space. Journal of Plant Ecology, 1, 33–41.
von Felten, S., Hector, A., Buchmann, N., Niklaus, P.A., Schmid, B. &
Scherer-Lorenzen, M. (2009) Belowground nitrogen partitioning in experi-
mental grassland plant communities of varying species richness. Ecology, 90,
1389–1399.
Fornara,D.A., Tilman,D.&Hobbie, S.E. (2009) Linkages between plant func-
tional composition, fine root processes and potential soil N mineralization
rates. Journal of Ecology, 97, 48–56.
Gastine, A., Scherer-Lorenzen, M. & Leadley, P.W. (2003) No consistent
effects of plant diversity on root biomass, soil biota and soil abiotic condi-
tions in temperate grassland communities. Applied Soil Ecology, 24,
101–111.
Gersani,M., Abramsky, Z. & Falik, O. (1998) Density-dependent habitat selec-
tion in plants.Evolutionary Ecology, 12, 223–234.
Gruntman, M. & Novoplansky, A. (2004) Physiologically mediated self ⁄ non-self discrimination in roots. Proceedings of the National Academy of Sciences
of theUnited States of America, 101, 3863–3867.
van der Heijden, M.G.A., Bardgett, R.D. & van Straalen, N.M. (2008) The
unseen majority: soil microbes as drivers of plant diversity and productivity
in terrestrial ecosystems.Ecology Letters, 11, 296–310.
Hooper, D.U., Chapin, F.S., Ewel, J.J., Hector, A., Inchausti, P., Lavorel, S.
et al. (2005) Effects of biodiversity on ecosystem functioning: a consensus of
current knowledge.Ecological monographs, 75, 3–35.
Huber-Sannwald, E., Pyke, D.A., Caldwell,M.M.&Durham, S. (1998) Effects
of nutrient patches and root systems on the clonal plasticity of a rhizomatous
grass.Ecology, 79, 2267–2280.
Kardol, P., Cornips, N.J., van Kempen, M.M.L., Bakx-Schotman, J.M.T. &
Van der Putten, W.H. (2007) Microbe-mediated plant-soil feedback causes
historical contingency effects in plant community assembly. Ecological
monographs, 77, 147–162.
Kowalchuk, G.A., Buma, D.S., de Boer, W., Klinkhamer, P.G.L. & van
Veen, J.A. (2002) Effects of above-ground plant species composition
and diversity on the diversity of soil-borne microorganisms. Antonie
Van Leeuwenhoek International Journal of General and Molecular
Microbiology, 81, 509–520.
de Kroon, H. (2007) Ecology – How do roots interact? Science, 318, 1562–
1563.
Levine, J.M. & HilleRisLambers, J. (2009) The importance of niches for the
maintenance of species diversity.Nature, 461, 254–257.
Levine, J.M., Pachepsky, E., Kendall, B.E., Yelenik, S.G. & HilleRisLambers,
J. (2006) Plant-soil feedbacks and invasive spread. Ecology Letters, 9, 1005–
1014.
Li, L., Sun, J.H., Zhang, F.S., Guo, T.W., Bao, X.G., Smith, F.A. & Smith,
S.E. (2006) Root distribution and interactions between intercropped species.
Oecologia, 147, 280–290.
Li, L., Li, S.M., Sun, J.H., Zhou, L.L., Bao, X.G., Zhang, H.G. & Zhang,
F.S. (2007) Diversity enhances agricultural productivity via rhizosphere
phosphorus facilitation on phosphorus-deficient soils. Proceedings of the
National Academy of Sciences of the United States of America, 104, 11192–
11196.
Loreau, M. & Hector, A. (2001) Partitioning selection and complementarity in
biodiversity experiments.Nature, 412, 72–76.
Mahall, B.E. & Callaway, R.M. (1991) Root communication among desert
shrubs. Proceedings of the National Academy of Sciences of the United States
of America, 88, 874–876.
Maina, G.G., Brown, J.S. & Gersani, M. (2002) Intra-plant versus inter-plant
root competition in beans: avoidance, resource matching or tragedy of the
commons.Plant Ecology, 160, 235–247.
McKane, R.B., Johnson, L.C., Shaver, G.R., Nadelhoffer, K.J., Rastetter,
E.B., Fry, B., Giblin, A.E., Kielland, K., Kwiatkowski, B.L., Laundre, J.A.
&Murray, G. (2002) Resource-based niches provide a basis for plant species
diversity and dominance in arctic tundra.Nature, 415, 68–71.
Mitchell, C.E., Tilman, D. & Groth, J.V. (2002) Effects of grassland plant spe-
cies diversity, abundance, and composition on foliar fungal disease. Ecology,
83, 1713–1726.
Mommer, L., Wagemaker, N., de Kroon, H. & Ouborg, N.J. (2008) Unravel-
ling belowground plant distributions: a real time PCR method for quantify-
ing species proportions in mixed root samples. Molecular Ecology Notes, 8,
947–953.
Parrish, J.A.D. &Bazzaz, F.A. (1976) Underground niche separation in succes-
sional plants.Ecology, 57, 1281–1288.
Poorter, H. & Nagel, O. (2000) The role of biomass allocation in the
growth response of plants to different levels of light CO2, nutrients and
water: a quantitative review. Australian Journal of Plant Physiology, 27,
595–607.
Reich, P.B., Tilman, D., Naeem, S., Ellsworth, D.S., Knops, J., Craine, J.,
Wedin, D. & Trost, J. (2004) Species and functional group diversity indepen-
dently influence biomass accumulation and its response to CO2 and N. Pro-
ceedings of the National Academy of Sciences of the United States of America,
101, 10101–10106.
van Ruijven, J. & Berendse, F. (2005) Diversity-productivity relationships:
Initial effects, long-term patterns, and underlying mechanisms. Proceedings
of the National Academy of Sciences of the United States of America, 102,
695–700.
van Ruijven, J. & Berendse, F. (2009) Long-term persistence of a positive plant
diversity-productivity relationship in the absence of legumes. Oikos, 118,
101–106.
Schenk, H.J. (2006) Root competition: beyond resource depletion. Journal of
Ecology, 94, 725–739.
Schenk, H.J. & Jackson, R.B. (2005) Mapping the global distribution of deep
roots in relation to climate and soil characteristics.Geoderma, 126, 129–140.
Semchenko,M., Hutchings,M.J. & John, E.A. (2007a) Challenging the tragedy
of the commons in root competition: confounding effects of neighbour pres-
ence and substrate volume. Journal of Ecology, 95, 252–260.
1126 L. Mommer et al.
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127

Semchenko, M., John, E.A. & Hutchings, M.J. (2007b) Effects of physical
connection and genetic identity of neighbouring ramets on root-placement
patterns in two clonal species.New Phytologist, 176, 644–654.
Spehn, E.M., Joshi, J., Schmid, B., Alphei, J. & Korner, C. (2000) Plant
diversity effects on soil heterotrophic activity in experimental grassland
ecosystems.Plant and Soil, 224, 217–230.
Steinbeiss, S., Bessler, H., Engels, C., Temperton, V.M., Buchmann, N., Ro-
scher, C., Kreutziger, Y., Baade, J., Habekost, M. & Gleixner, G. (2008)
Plant diversity positively affects short-term soil carbon storage in experimen-
tal grasslands.Global Change Biology, 14, 2937–2949.
Tilman, D., Reich, P.B., Knops, J., Wedin, D., Mielke, T. & Lehman, C. (2001)
Diversity and productivity in a long-term grassland experiment. Science,
294, 843–845.
Van der Krift, T.A.J. & Berendse, F. (2002) Root life spans of four grass species
from habitats differing in nutrient availability. Functional Ecology, 16, 198–
203.
Van der Putten,W.H. & Peters, B.A.M. (1997) How soil-borne pathogens may
affect plan competition.Ecology, 78, 1785–1795.
Weller, D.M., Raaijmakers, J.M., Gardener, B.B.M. & Thomashow, L.S.
(2002) Microbial populations responsible for specific soil suppressiveness to
plant pathogens.Annual Review of Phytopathology, 40, 309–348.
Westover, K.M. & Bever, J.D. (2001) Mechanisms of plant species coexistence:
roles of rhizosphere bacteria and root fungal pathogens. Ecology, 82, 3285–
3294.
Received 08 February 2010; accepted 18 June 2010
Handling Editor:Marina Semchenko
Supporting Information
Additional Supporting Information may be found in the online ver-
sion of this article:
Figure S1. Experimental setup of amixture plot.
Figure S2. Nutrient uptake at the community scale.
As a service to our authors and readers, this journal provides support-
ing information supplied by the authors. Such materials may be
re-organized for online delivery, but are not copy-edited or typeset.
Technical support issues arising from supporting information (other
thanmissing files) should be addressed to the authors.
Below-ground species distributions in a biodiversity experiment 1127
� 2010 The Authors. Journal compilation � 2010 British Ecological Society, Journal of Ecology, 98, 1117–1127




















