
Universidade Federal do Rio Grande
33
UNIVERSIDADE FEDERAL RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS DEPARTAMENTO DE OCEANOGRAFIA E LIMNOLOGIA VARIAÇÃO ESPAÇO-TEMPORAL DA MEGAFAUNA EPIBENTÔNICA DA PRAIA DE BAÍA FORMOSA, RIO GRANDE DO NORTE. ANA CARLA GREGOLIN MENDES Natal 2014
Transcript of Universidade Federal do Rio Grande

UNIVERSIDADE FEDERAL RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
DEPARTAMENTO DE OCEANOGRAFIA E LIMNOLOGIA
VARIAÇÃO ESPAÇO-TEMPORAL DA MEGAFAUNA EPIBENTÔNICA DA
PRAIA DE BAÍA FORMOSA, RIO GRANDE DO NORTE.
ANA CARLA GREGOLIN MENDES
Natal
2014

Monografia apresentada como
requisito parcial à obtenção do grau
de Bacharel em Ciências Biológicas da
Universidade Federal do Rio Grande
do Norte.
Orientação: Profª. Dra. Tatiana Silva
Leite
Co-orientação: Prof. Dr. Fúlvio Freire.
ANA CARLA GREGOLIN MENDES
VARIAÇÃO ESPAÇO-TEMPORAL DA MEGAFAUNA EPIBENTÔNICA DA
PRAIA DE BAÍA FORMOSA, RIO GRANDE DO NORTE.
Natal
2014

SUMÁRIO
LISTA DE FIGURAS.....................................................................................................ii
LISTA DE TABELAS....................................................................................................iii
1.INTRODUÇÃO............................................................................................................6
2.OBJETIVOS.................................................................................................................8
2.1.Objetivo Geral.........................................................................................................8
2.2.Objetivos Especificos ..............................................................................................8
3.METODOLOGIA.........................................................................................................9
3.1.Área de Estudo........................................................................................................9
3.2.Coletas......................................................................................................................9
3.3.Laboratório............................................................................................................10
3.4.Analises Estatísticas..............................................................................................10
4.RESULTADOS ...........................................................................................................12
5.DISCUSSÃO ..............................................................................................................23
6.CONSIDERAÇÕES FINAIS.....................................................................................27
REFERENCIAS.............................................................................................................28

LISTA DE FIGURAS
Figura 1: Localização da área de estudo enfatizando as estações de coleta e a influencia
do rio Cunhaú na porção mais ao norte da costa. Fonte: ALENCAR et al, 2014.
Figura 2: Precipitação acumulada por mês, medida no posto: Baía Formosa
(prefeitura). Gráfico baseado em dados da EMPARN.
Figura 3: Representação gráfica do percentual de indivíduos coletados e biomassa
relativa dos filos.
Figura 4: Número total de indivíduos dos táxons e/ou morfotipos mais abundantes
(acima de 10 indivíduos) nas amostragens de abril/2013 a março 2014.
Figura 5: Biomassa total dos táxons e morfotipos mais representativos (acima de 50
gramas) nas amostragens de abril/2013 a março/2014.
Figura 6: Variação temporal da abundância e biomassa totais durante o período do
estudo.
Figura 7: Dispersão da abundância e biomassa totais em relação à precipitação durante
os meses do estudo.
Figura 8: Comparação das médias de biomassa total para as estações chuvosa e seca
durante o período de estudo.
Figura 9: Variação sazonal dos índices de diversidade (Shannon), riqueza (Margalef) e
dominância ao longo do período de amostragem.
Figura 10: Variação mensal na abundância dos táxons dominantes.
Figura 11: Variação temporal na abundância relativa dos táxons dominantes.
Figura 12: Variação espacial do número total de indivíduos e biomassa conforme a
proximidade do estuário do Rio Cunhaú.
Figura 13: Variação espacial índices de diversidade (Shannon), riqueza (Margalef) e
dominância ao longo do período de amostragem.
Figura 14: Variação espacial na abundância relativa dos táxons dominantes

LISTA DE TABELAS
Tabela 1: Táxons de invertebrados bentônicos registrados em Baía Formosa e suas
abundancias (número total) e biomassas (gramas).

1.INTRODUÇÃO
Os organismos bênticos de regiões costeiras, principalmente nas áreas mais rasas
da plataforma, apresentam grande importância econômica e ecológica. Isto se dá tanto
pela sua participação na reciclagem de nutrientes e matéria orgânica depositada, quanto
pela sua utilização como itens alimentares pelo homem ou por peixes demersais de
importância econômica (Caddy, 1989). Crustáceos, equinodermas, corais, esponjas e
moluscos são os táxons de invertebrados que incluem o maior numero de espécies
encontradas nesses ambientes, e possuem como características gerais a baixa ou
nenhuma motilidade e vida relativamente longa (Soares-Gomes et al., 2009).
A comunidade faunística habitante dos fundos inconsolidados é estruturada pela
interação de fatores abióticos, biológicos e influências antrópicas (Amaral, 2011). A
hidrodinâmica, tipo de sedimento, salinidade, temperatura e poluição influenciam a
distribuição das assembleias bentônicas. Da mesma forma, o ciclo de vida dos
organismos e as interações biológicas são componentes que explicam flutuações de
espécies no espaço e no tempo (Peroni & Hernández, 2011).
Nos ambientes estuarinos ocorre uma ligação entre as águas interiores e o mar,
de forma que os mesmos sofrem influencia das marés e apresentam fortes gradientes
ambientais, desde águas doces próximos a sua cabeceira, águas salobras, e águas
marinhas próximas a sua desembocadura. Tais variações de salinidade acarretam na
variação da composição de espécies e riqueza diminuída. Organismos que vivem em
estuários estão sujeitos a estes gradientes e apresentam diversas estratégias para
sobreviver. Por exemplo, os crustáceos decápodes são capazes de osmorregular e estão
distribuídos por toda a região, já os equinodermos ocupam a zona mais marinha do
estuário e os gastrópodes fecham as conchas mediante a diminuição da salinidade
(Little, 2009).
A estrutura da comunidade animal refere-se aos atributos da sua composição,
abundância e diversidade de espécies. Assim, parâmetros usados na descrição da
estrutura da comunidade bentônica incluem: abundância ou densidade de algumas
espécies em relação às outras, frequência de ocorrência de uma espécie que é expressa
como uma porcentagem e a diversidade ou número de espécies encontradas. A possível
exclusão de determinadas espécies por atividades antrópicas, como a pesca de arrasto,
reflete no decréscimo da riqueza e diversidade específica, e pode alterar a distribuição e
a estrutura da comunidade faunística epibentônica (Reiss e Kroncke, 2004; Rábago-
Quiroz, 2011; DFO, 2012).
Teorias indicam que tanto a densidade como a biomassa de invertebrados
bênticos de plataformas tropicais e subtropicais tendem a ser reduzidas, com exceção
das áreas de ressurgência, onde a produção primária é mais elevada (Soares-Gomes et
al., 2009). Um estudo realizado por Vanin (1993) constatou que ocorre um aumento na
biomassa e abundância de organismos bentônicos no litoral de São Paulo nos períodos

de primavera – verão, devido a processos físicos como a entrada de frentes frias e
ressurgências. Outros estudos realizados no sul e sudeste do Brasil também
evidenciaram o registro de valores mais elevados de abundância e biomassa em meses
com temperaturas mais altas (Branco & Fracasso, 2004; Santos et al., 2000, Kotas,
1998). Sugere-se que para grandes invertebrados epibênticos ocorre um aumento na
diversidade em latitudes tropicais. Entretanto, segundo Lana (1996) em fundos arenosos
do nordeste do Brasil a biodiversidade é baixa com uma predominância de moluscos,
equinodermos e crustáceos.
Entre os aspectos antrópicos que afetam a fauna bentônica, as pescarias de
arrasto estão entre os mais frequentes. As redes de arrasto são pouco seletivas, causam
perturbações no fundo marinho, o deslocamento dos animais e impactos sobre o
recrutamento de juvenis, exercendo alterações na estrutura da comunidade dos
organismos bênticos (Duarte,2012). Analises dos dados relativos à epifauna capturada
acidentalmente na América do Norte - Canadá, Califórnia e Carolina do Norte - indicam
que as comunidades bentônicas em áreas intensamente exploradas pela pesca passam a
ser caracterizadas por grande abundância de organismos de pequeno porte ao invés
de espécies de elevada biomassa (Reiss e Kroncke, 2004; Rábago-Quiroz, 2011; DFO,
2012). Também se pode constatar que as atividades e o aumento da pressão de
exploração dos recursos sobre os ecossistemas costeiros ocasionam alterações na
dinâmica dos animais, afetando a qualidade do ambiente e a própria manutenção da
pesca (McConnaughey et al. 2000).
No estado do Rio Grande do Norte, com a faixa litorânea de 400 km divididos
em 25 municípios, a pesca de camarão ocorre do litoral norte ao sul, realizada em lagoas
com canoas ou em praias com o uso de barco a motor. Dentre as praias do litoral sul,
Baía Formosa destaca-se pela atividade que tem como características o uso de
embarcações de madeira, com casaria localizada na popa do convés, geralmente
tripuladas por dois pescadores e um motorista. As embarcações são menores que 12
metros e operam com apenas uma rede de arrasto, realizando viagens diárias com dois
arrastos com duração de 4 horas cada.
Sendo este tipo de modalidade pesqueira recorrente na localidade de Baia
Formosa, e considerando os possíveis impactos desta atividade sobre a comunidade
bentônica, mostra-se de grande importância caracterizar a variação espaço-temporal da
megafauna epibêntica de fundos inconsolidados nesta região.

2.OBJETIVOS
2.1.Objetivo geral:
Caracterizar a distribuição espaço-temporal da epifauna bentônica de fundo
inconsolidado na praia de Baía Formosa/RN.
2.2.Objetivos específicos:
Identificar os principais grupos de invertebrados da epifauna bentônica da região
Analisar a estrutura da comunidade bentônica em relação às estações seca e
chuvosa.
Verificar a distribuição dos diferentes grupos identificados em relação à
proximidade do estuário.

3.METODOLOGIA
3.1.Área de estudo
A praia de Baía Formosa encontra-se a 90 km da capital, no extremo sul do
litoral oriental do estado, a 6° 22’ 10” de latitude sul e 35° 00’ 28 ” de longitude
oeste . Segundo o IDEMA (2014) o clima da área é do tipo tropical úmido,
caracterizado por ocorrência de chuvas entre janeiro e agosto. A temperatura média
anual é de 27,3 °C com máxima de 29,6 °C e mínima de 23,9 °C. A maré apresenta
características semi-diurnas com a variação média das marés de sizígias de cerca de
2.30m e das marés de quadratura, de cerca de 0,85 m. As correntes de maré são mais
fortes nas vazantes de inverno, período este caracterizado por chuvas intensas. Na divisa
com o município de Canguaretama deságua o rio Cunhaú que devido à proximidade da
praia de Baía Formosa pode ter influencia na dinâmica oceanográfica desta localidade.
A economia da região baseia-se na agricultura e pesca (Freire, 2012).
3.2.Coletas
Para este estudo foi utilizado o material coletado, durante o Projeto “ESTUDO
DA DINÂMICA POPULACIONAL DOS CAMARÕES MARINHOS DE INTERESSE
ECONÔMICO NO LITORAL DO ESTADO DO RIO GRANDE DO NORTE COM
ÊNFASE NA ESPÉCIE XIPHOPENAEUS KROYERI (HELLER, 1862)
(CRUSTACEA, DECAPODA, PENAEIDAE).”, desenvolvido pelo Grupo de Estudos
em Ecologia e Fisiologia de Animais Aquáticos – GEEFAA/UFRN, sob a coordenação
do Prof. Dr. Fúlvio Aurélio de Morais Freire.
A amostragem da epifauna de invertebrados constou de 12 campanhas mensais,
realizadas durante o período de abril de 2013 a março de 2014, no litoral de Baia
Formosa. As operações de lançamento e recolhimento foram feitas manualmente pela
equipe do barco, composta por cinco integrantes do laboratório e os pescadores.
Os arrastos foram realizados entre 5 e 20m de profundidade da plataforma
continental em 6 transectos paralelos a costa, percorridos por 20 minutos, a uma
velocidade média de 1,08 nós. Os pontos de coleta foram predefinidos com o auxilio de
um GPS, visando duas áreas, uma próxima e outra distante da influencia do estuário do
rio Cunhaú. A arte de pesca utilizada consistiu na rede de arrasto com portas, empregada
na pesca comercial de camarão, a qual possui a malha do saco em torno de 25 mm.
Ainda a bordo, ocorreu uma triagem prévia dos táxons logo após o recolhimento
da rede. As amostras foram colocadas em sacos plásticos, etiquetadas - conforme o
trasecto amostrado - e armazenadas em recipiente de isopor resfriado com gelo.
Dados pluviométricos foram obtidos no site da Empresa de Pesquisa
Agropecuária do Rio Grande do Norte (EMPARN), em que foram utilizados os dados
de precipitação acumulada por mês nos anos de 2013 e 2014 para Baía Formosa.

Figura 1: Localização da área de estudo enfatizando as estações de coleta e a influencia
do rio Cunhaú na porção mais ao norte da costa. Fonte: ALENCAR et al, 2014.
3.3.Laboratório
Em laboratório, ocorreu a triagem dos exemplares por grandes grupos – e
morfotipos, que foram contados, pesados e fixados em álcool 70%. A identificação dos
espécimes foi realizada mediante a consulta de bibliografias especializadas (Rios, 2009;
Thomé et al, 2004; Clark & Downey, 1992; Matthews-Cascon, 2006; Absalão &
Pimenta, 2005) e com o apoio de especialistas de outros laboratórios. Todos esses dados
foram anotados e planilhados simultaneamente às analises.
3.4.Analises estatísticas
A partir dos registros, foram calculados os índices de riqueza de Margalef, de
diversidade de Shannon e dominância de Simpson utilizando-se o programa PAST.
Visando-se observar as variações mensais, para o cálculo dos índices foram somados
todos os valores de abundância das espécies dos 6 transectos amostrados em cada mês.
Para comparar os dados das estações do ano (chuva e seca), os meses de abril,
maio, junho, julho, agosto e setembro foram considerados réplicas para a estação de
maior pluviosidade e os meses de outubro, novembro, dezembro, janeiro, fevereiro e
março para a estação de estiagem. Com a finalidade de avaliar as variações espaciais, os
índices foram calculados considerando a abundância de todas as espécies das 12 coletas
mensais de cada localidade (transecto). Para verificar a influência do estuário, as

réplicas para a área distante do estuário foram os transectos 1, 2 e 3 e como réplicas para
a área mais próxima do estuário, os transectos 4, 5 e 6.
Tais dados foram comparados entre os meses e as regiões, para saber se existem
diferenças significativas entre as estações seca e chuvosa e entre a área mais próxima e
mais distante do estuário. Em função da não normalidade dos dados, fez-se o teste de
Kruscal-Walis no programa Statistica v7.
Para relacionar a quantidade e o peso dos organismos à pluviosidade foi feita
uma regressão linear simples, considerando a biomassa e a abundância como variável
dependente e a pluviosidade acumulada por mês como variável independente.

4.RESULTADOS
Em relação à pluviosidade observou-se que ocorrem variações na precipitação
em Baía Formosa, constando de estações: inverno e verão. O gráfico apresenta a
precipitação acumulada por mês durante o estudo. Pode-se observar o período chuvoso
inicia no mês de março e se estende até novembro, com destaque para o mês de julho
como o de maior pluviosidade. O período seco é composto pelos meses de outubro,
novembro e dezembro de 2013- classificados como muito secos- e janeiro e fevereiro de
2014, com baixa quantidade de chuvas.
Figura 2: Precipitação acumulada por mês, medida no posto: Baía Formosa
(prefeitura). Gráfico baseado em dados da Emparn.
Em relação às amostras biológicas, durante o período estudado foram capturadas
63 espécies/morfotipos, totalizando 1830 exemplares e 15,44 kg de invertebrados
bentônicos. Tais organismos pertenciam aos filos Mollusca (12 ssp.), Arthropoda-
Crustacea (44 ssp.), Echinodermata (5 ssp.) e Cnidaria (2 ssp.).
Tabela 1: Táxons de invertebrados bentônicos registrados em Baía Formosa e suas
abundâncias (número total) e biomassas (gramas).

Táxon N Biomassa (g)
CNIDARIA 10
Octocorallia
Renilla muelleri 2 0,53
Renilla sp. 8 11,95
MOLLUSCA 395
Bivalvia
Anadara sp. 4 16,57
Ventricolaria sp. 1 2
Cephalopoda
Loliginidae spp. 337 1422,57
Gastropoda
Anaspidea 1 4
Aplysia sp. 7 20
Cassis tuberosa 1 48
Natica canariensis 13 35
Sinum maculatum 1 0,72
Distorsio clathrata 22 225,4
Stramonita haemastoma floridana 1 5
Olivella sp. 1 0,05
Turbinella laevigata 6 971,52
CRUSTACEA 1146
Brachyura
Hepatus gronovii 3 31,13
Calappa ocellata 1 4,15
Calappa angusta 1 0,98
Stratiolibinia bellicosa 7 366,21
Acanthonyx dissimulatus 3 0,06
Epialtus bituberculatus 3 0,03
Pitho lherminieri 1 0,01
Persephona lichtensteinii 6 21,22
Persephona punctata 1 16,53
Microphrys sp. 1 0,36
Menippe nodifrons 1 47,92
Hexapanopeus caribbaeus 2 0,59
Pilumnus dasypodus 2 0,02
Pilumnus diomedeae 1 0,13
Pilumnus reticulatus 2 0,12
Dissodactylus crinitichelis 2 0,07
Xanthodius denticulatus 1 0,47
Callinectes ornatus 112 1906,22
Charybdis helleri 4 24,35

Portunus spp. 2 0,7
Arenaeus cribrarius 11 516,19
Cronius tumidulus 1 0,12
Callinectes sapidus 2 132,08
Callinectes danae 10 333,76
Callinectes spp. 55 87,59
Callinectes sp. 56 197,87
Anomura
Pagurus leptonyx 1 0,02
Dardanus venosus 19 343,25
Isocheles sawayai 1 3,02
Petrochirus diogenes 54 3945,21
Porcellana sayana 2 0,12
Achelata
Panulirus echinatus 1 61,13
Panulirus laevicauda 5 291,37
Parribacus antarcticus 1 3,23
Caridea
Alpheidae 3 0,98
Alpheus intrinsecus 5 1,78
Alpheus sp. 1 0,12
Exhippolysmata oplophoroides 350 351,74
Podochela cf. brasiliensis 1 0,01
Nematopalaemon schmitti 327 468,29
Caridea spp. 6 1,5
Dendrobranchiata
Rimapenaeus similis 2 1,94
Acetes americanus 62 3,19
Sicyonia dorsalis 14 7,06
ECHINODERMATA 279
Asteroidea
Astropecten sp. 5 25,53
Luidia senegalensis 202 2004,94
Echinoidea
Mellita sexisperforata 68 1475,17
Tripneustes ventricosus 1 3
Ophiuroidea
Ophiuroidea sp. 3 0,09
No total, os crustáceos foram os mais representativos, tanto em número de
indivíduos como em peso. Os moluscos contribuíram com 22% dos indivíduos e os
equinodermos com 15% da abundância total. No entanto, a biomassa percentual de

equinodermos superou a de moluscos. Os cnidários apresentaram baixa
representatividade com apenas 10 indivíduos e 20g de peso.
Figura 3: Representação gráfica do percentual de indivíduos coletados e biomassa
relativa dos filos
Dos 63 táxons e morfotipos capturados, 16 apresentaram 10 indivíduos ou mais.
Destacaram-se 2 espécies de camarão - Nematopolemon schimitti e Exhippolysmata
oplophoroides pertencentes à família Caridea, com importância econômica reduzida no
Brasil, as lulas da família Loliginidae, e a estrela-do-mar Luidia senegalensis com mais
de 200 indivíduos (figura 4).
Figura 4: Número total de indivíduos dos táxons e/ou morfotipos mais abundantes
(acima de 10 indivíduos) nas amostragens de abril/2013 a março 2014.
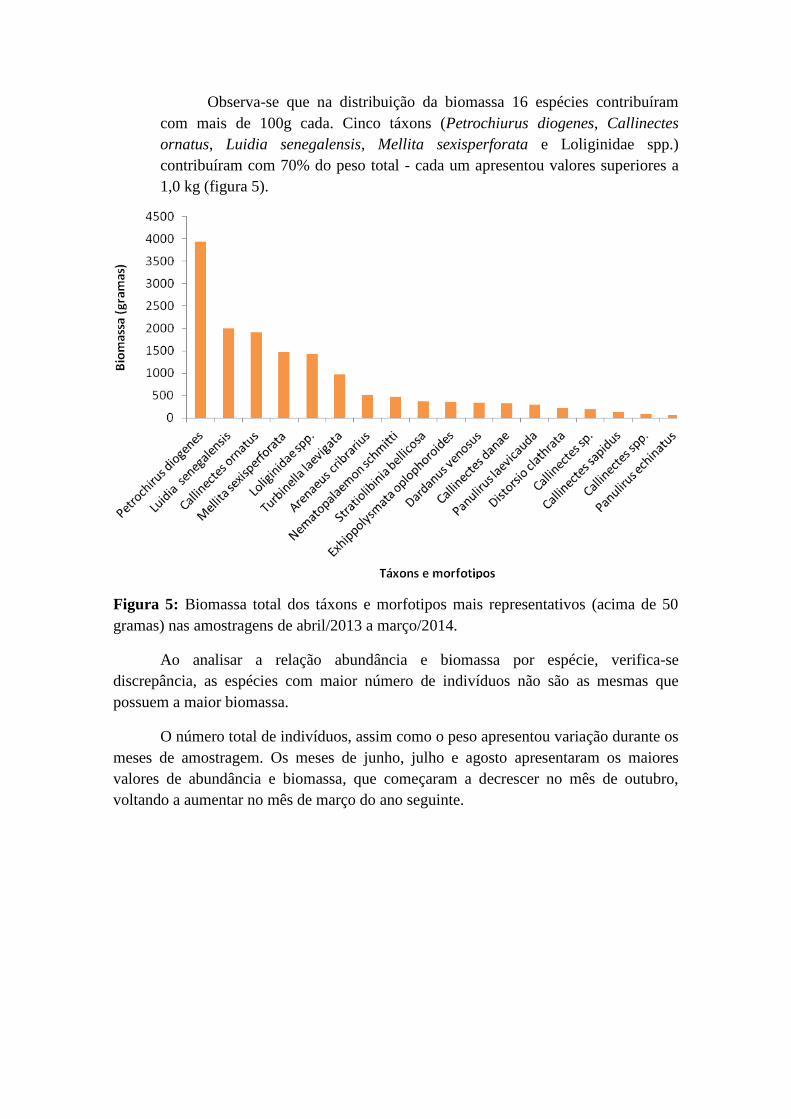
Observa-se que na distribuição da biomassa 16 espécies contribuíram
com mais de 100g cada. Cinco táxons (Petrochiurus diogenes, Callinectes
ornatus, Luidia senegalensis, Mellita sexisperforata e Loliginidae spp.)
contribuíram com 70% do peso total - cada um apresentou valores superiores a
1,0 kg (figura 5).
Figura 5: Biomassa total dos táxons e morfotipos mais representativos (acima de 50
gramas) nas amostragens de abril/2013 a março/2014.
Ao analisar a relação abundância e biomassa por espécie, verifica-se
discrepância, as espécies com maior número de indivíduos não são as mesmas que
possuem a maior biomassa.
O número total de indivíduos, assim como o peso apresentou variação durante os
meses de amostragem. Os meses de junho, julho e agosto apresentaram os maiores
valores de abundância e biomassa, que começaram a decrescer no mês de outubro,
voltando a aumentar no mês de março do ano seguinte.

Figura 6: Variação temporal da abundância e biomassa totais durante o período do
estudo.
Os valores de R² nas regressões lineares mostram que os valores de abundância
e biomassa são diretamente proporcionais à precipitação representada na figura abaixo.
Os valores são mais elevados no período chuvoso, e diminuem no período seco (R²=
0,6). Em dezembro foram capturados somente 17 espécimes, distribuídos em 9 táxons.
No entanto, a diferença sazonal entre as médias foi significativa apenas para a biomassa
(K-W, p<0,05) (figura 8).

Figura 7: Dispersão da abundância e biomassa totais em relação à precipitação durante
o período de estudo.

Figura 8: Comparação das médias de biomassa total para as estações chuvosa e seca
durante o período de estudo.
Os resultados obtidos para a estrutura da comunidade (figura 9) mostram uma
baixa variabilidade da diversidade entre os meses avaliados (1,8-2,3), sendo que os
menores valores foram encontrados no mês de agosto. A riqueza apresentou uma maior
variação quando comparada a diversidade (2,4-4,8), com maiores valores nos meses de
julho, setembro e março e menores nos meses de maio, agosto, outubro e novembro.
Entretanto, a dominância explica as alterações nos outros índices, pois se comporta de
forma inversa à riqueza e seus picos indicam os meses de menor equitabilidade. Ao
comparar as médias dos índices entre as estações do ano, verificou-se que não ocorreu
diferença significativa (K-W, p> 0,05) entre o período chuvoso e seco.
Figura 9: Variação sazonal dos índices de diversidade (Shannon), riqueza (Margalef) e
dominância ao longo do período de amostragem.

Durante o período de coleta, cinco táxons dominaram as amostras em todos os
meses estudados, a família Loliginidae, e as espécies Luidia senegalensis,
Exhippolysmata oplophoroides, Nematopalaemon schmitti e Callinectes ornatus. Os
táxons menos representativos foram classificados como “outros” e obtiveram maior
abundância apenas no mês de maio.
De uma forma geral os táxons dos Loliginidae e Callinectes foram os mais
frequentes durante o ano, ocorrendo em quase todos os meses, com exceção do mês de
maio. Dentre os 64 espécimes coletados neste mês, foram identificadas 13 espécies de
crustáceos. As lulas apresentaram uma evidente dominância em outubro e janeiro. Já a
estrela Luidia senegalensis não ocorreu nos meses de menor abundância, porém em
abril e junho compôs 38% e 25% do total, respectivamente. O siri, Callinectes ornatus,
mostrou um pico de dominância em dezembro e não esteve entre os mais numerosos de
julho. Os camarões carídeos contribuíram com o maior número de indivíduos na
maioria dos meses de estudo, exceto em abril, outubro, dezembro, janeiro.
Figura 11: Variação temporal na abundância relativa dos táxons dominantes.
Em relação à distribuição espacial observou-se que o número de indivíduos e a
biomassa total apresentaram diferentes valores para os transectos amostrados. A maior
abundância e peso ocorreu no transecto 1, local de menor profundidade e mais abrigado.
O transecto 6, longe da praia, onde a profundidade foi maior apresentou apenas 62
indivíduos e 750g coletados em 1 ano.
Quando comparado os transectos em conjunto para averiguação da influência do
estuário na estrutura das comunidades, verificou-se que, apesar da área de estudo mais
distante do estuário ter maiores valores para a biomassa e abundância esta diferença não

foi significativa (K-W, p>0,05) (figura 12).
Figura 12: Variação espacial do número total de indivíduos e biomassa conforme a
proximidade do estuário do Rio Cunhaú.
Em relação aos índices de diversidade, a diversidade mostrou uma tendência
geral a aumentar do T1 para o T6, enquanto que a dominância apresentou tendência
oposta (figura 13). Já a riqueza apresentou um pico no transecto 3, o mais distante da
costa e do rio. Apesar das tendências, as médias não apresentaram diferenças
significativas quando comparadas as áreas próxima e distante do estuário (K-W,
p>0,05).
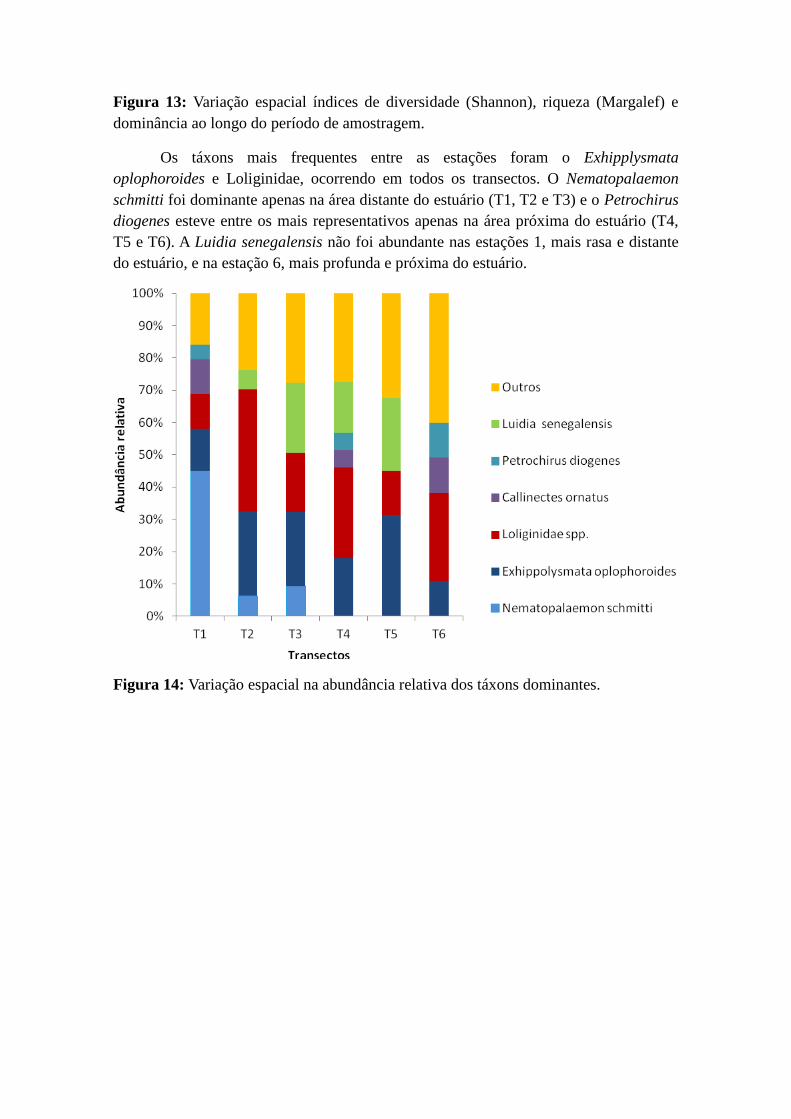
Figura 13: Variação espacial índices de diversidade (Shannon), riqueza (Margalef) e
dominância ao longo do período de amostragem.
Os táxons mais frequentes entre as estações foram o Exhipplysmata
oplophoroides e Loliginidae, ocorrendo em todos os transectos. O Nematopalaemon
schmitti foi dominante apenas na área distante do estuário (T1, T2 e T3) e o Petrochirus
diogenes esteve entre os mais representativos apenas na área próxima do estuário (T4,
T5 e T6). A Luidia senegalensis não foi abundante nas estações 1, mais rasa e distante
do estuário, e na estação 6, mais profunda e próxima do estuário.
Figura 14: Variação espacial na abundância relativa dos táxons dominantes.

5. DISCUSSÃO
Os resultados do presente estudo indicaram a presença de grupos de organismos
comuns na região Nordeste do Brasil, como crustáceos, equinodermos e moluscos que
habitam os fundos inconsolidados (Mota-Alves & Rodrigues 1977, Coelho & Santos
1993). Esses grandes grupos são principais representantes da epifauna bentônica em
diversas áreas tropicais e também em outras regiões do país (Branco & Fracasso, 2004;
Kotas, 1998; Braga, 2000; Branco & Verani, 2006; Pires-Vanin et al, 1995; Costa &
Medeira Di Beneditto, 2009).
De acordo com Carranza Frazer & Grande (1982) e Ruffino & Castelo (1992), a
abundância relativa dos grupos da megafauna epibêntica pode sofrer mudanças em
função da época do ano ou área de pesca. O presente estudo verificou um aumento do
peso e número de indivíduos capturados nos meses chuvosos o que evidenciou uma
variação sazonal em relação à abundância e biomassa de invertebrados bentônicos na
praia de Baía Formosa. O enriquecimento do substrato da costa pelo transporte de
matéria orgânica e inorgânica pelos rios (Soares-Gomes et al., 2009) poderia explicar a
biomassa significativamente maior no inverno (estação chuvosa). À medida que esses
nutrientes podem ser metabolizados pelos organismos, favorecem a entrada de energia
na teia alimentar, pois aumentam a proliferação de algas e a produção primária, além de
serem diretamente consumidos por outros níveis tróficos (Begon, et al. 2007).
No entanto, outros fatores abióticos de maior significância devem ser
considerados para explicar a variação espaço-temporal, como por exemplo, as correntes.
A velocidade da corrente influencia o tamanho das partículas do substrato, afetando a
distribuição de alimento e remoção de nutrientes (Allan, apud Almeida, 2003). O
revolvimento do sedimento por diferentes correntes, maior no inverno, também tem
efeitos na abundância dos organismos detritívoros e suspensívoros, à medida que torna
disponível a matéria orgânica. Os diferentes habitats que se formam devido o tipo de
sedimento, presença de algas ou folhiço, também oferecem esconderijos e podem
abrigar mais espécies/organismos (Soares-Gomes, 2009). Desta forma, análises futuras
de fatores ambientais suplementares, poderão elucidar aspectos mais particulares como
as ocorrências restrita de algumas espécies a determinadas áreas de coleta.
Na análise espacial os resultados de aumento de riqueza e diminuição da
dominância e abundância na área mais próxima ao rio Cunhaú foram opostos ao
esperado. Maiores abundâncias são observadas em direção ao estuário, em função do
enriquecimento da água pelos nutrientes, assim como uma diminuição da diversidade
devido a dominância das espécies resistentes, que se ajustam as situações de estresse
ambiental (Soares-Gomes et al., 2009; Coull, 1988). A diminuição do número de
organismos e biomassa na área de influência do deságue do rio, diferente do que consta
na literatura, pode ter uma explicação baseada nas correntes locais, até o momento
desconhecidas. A presença de uma corrente que leve a água oriunda do continente para

outra região explicaria porque a abundância, biomassa e dominância foram menores na
área mais próxima do estuário.
Através de observações pessoais, em mapas e em campo, pode-se perceber que a
área distante do estuário é abrigada, ou seja, mais protegida de correntes pela costa.
Além disso, esta possui sedimento de granulometria mais fina e presença de restos de
conchas em sua composição, o que pode gerar um acúmulo de nutrientes. Estudos
citados por Knox (2001) revelaram que em áreas onde o teor de matéria orgânica do
sedimento era alta, espécies com necessidades tróficas semelhantes coexistiram,
enquanto que em áreas com baixo teor de matéria orgânica, apenas espécies de
categorias tróficas diferentes conseguem sobreviver. Dessa forma, as espécies
dominantes de crustáceos (camarão-espinho, o camarão barriga-branca e o siri
Callinectes ornatus), ambos organismos detritivoros, teriam condições de coexistir na
área abrigada. Já na área próxima ao estuário, a estrela-do-mar foi encontrada em maior
abundância junto aos crustáceos, lulas e ermitões por serem animais que não têm
necessidades nutricionais tão semelhantes, e assim, concorrerem menos.
Conforme o que foi citado anteriormente, infere-se que a área abrigada e distante
do estuário tenha maior disponibilidade de alimentos e que comida está limitando
nessas comunidades de fundos moles, além de que, a concorrência é maior entre as
espécies com necessidades semelhantes. Dessa forma, outra explicação para o aumento
de indivíduos coletados no inverno é que a elevada pluviosidade pode ter favorecido a
deposição de matéria orgânica e, consequentemente, a oferta de alimentos para a
megafauna epibentônica. As correntes mais fortes no inverno também podem ser
relacionadas ao revolvimento do susbtrato e remoção da infauna do sedimento, trazendo
mais alimento para os consumidores que habitam a superfície do fundo marinho (Knox,
2001).
Neste estudo, o filo Crustacea foi o principal representante dos invertebrados
amostrados em número de espécies, abundância e biomassa. Fato que é realidade na
maioria das pescarias de arrasto de fundo, tanto em regiões temperadas quanto tropicais
(Graça Lopes et al, 2002; Vera et al; Cattani et al, 2012; Robert et al, 2007). Algumas
espécies de camarões da família Caridea, como Nematopolemon schimitti e
Exhippolysmata oplophoroides apresentam reduzida importância econômica no Brasil, e
dominaram o by catch da pesca artesanal de X. kroyeri em Baía Formosa.
Comparando a abundância dos táxons encontrados nesse estudo, verifica-se uma
semelhança em relação a grande abundância dos tá xons, por exemplo, do crustáceo
carídeo Exhippolysmata oplophoroides com o Paraná (Robert et al, 2007). Já na pesca
do camarão-rosa, realizada em Santa Catarina, a família Loliginidae foi o táxon
dominante nesta região em termos de abundancia e biomassa entre os moluscos (Kotas,
1998), o que também ocorreu no presente estudo.
Segundo Herrera (2013), a distribuição espaço-temporal de N. schmitti revelou
uma relação com os fatores ambientais analisados, principalmente com a granulometria

do sedimento e com a temperatura da água. Isto permitiu propor que tal espécie prefere
se distribuir em locais com predominância de areia fina e em períodos com temperatura
variando entre 20ºC e 22ºC. A citada espécie também teve maior representatividade em
locais com acumulo de folhas, que conferiram abrigos para este camarão (Almeida,
2007). Sabe-se, por observação durante as coletas em Baía Formosa, que os transectos
distantes do estuário localizam-se numa área mais abrigada e de sedimento de aspecto
lamoso, com parcela da região composta por sedimento bioclástico. Pode-se, assim,
inferir que tais características determinaram a dominância do camarão barriga-branca
apenas nessa porção da área analisada.
Apesar de todos os grupos seguirem o mesmo padrão de abundância
sazonalmente, o inverno, apesar de não rigoroso, pode ter favorecido a maior biomassa
de camarões que preferem menores temperaturas. Além das caracteristicas ideais para
cada espécie, a maior disponibilidade de nutrientes e favorecimento de fases do ciclo de
vida, contribuiram para a relevante diferença temporal na distribuição dos invertebrados
epibênticos da localidade estudada.
A estrela-do-mar Luidia senegalensis foi a mais representativa entre os
equinodermos de Baía Formosa. Esses animais são predadores generalistas que se
alimentam de pelicipodos, gastrópodes, asteroides e equinoides irregulares, braquiuros,
e outros pequenos crustáceos decápodos, copépodos, além de poliquetas (Moura, 1999).
A maior ocorrência dessa estrela nos meses de maior abundância é justificada pelo
maior número de presas disponíveis no período do inverno.
Já a bolacha-de-praia (ex. Mellita) é conhecida como agente modificador das
propriedades texturais do sedimento, principalmente na atividade de bioturbação da
meiofauna. Devido a sua capacidade de escavar o sedimento, a espécie influencia a
atividade microbiana pela alteração do tamanho dos detritos, pela homogeneização e
ressuspensão dos sedimentos, pela regeneração de nutrientes minerais, pelo aumento da
camada de atividade oxi-redução e pela redução da zona anaeróbica do sedimento.
Como consequência, eleva-se a biomassa microbiana e altera-se a estrutura das
comunidades de eucariontes (Dias, 2008). Esta espécie não esteve entre as dominantes,
mas seu elevado número de indivíduos nos meses chuvosos pode ter contríbuído para a
maior abundância total de invertebrados no inverno, disponibilizando maior quantidade
de matéria organica para os demais níveis tróficos da teia alimentar.
As águas costeiras são habitat da maioria dos loliginídeos (ex. Loligo) como
observada a dominância dessa família no presente estudo (Pires-Vanin, 1993). As lulas
são predadoras de crustáceos (ex. camarões e siris), de diversas espécies de peixes e de
moluscos, como outros cefalópodes, bivalves e gastrópodes (Ruppert, 1996). Algumas
capturadas durante a pesca de camarão em Baía Formosa possuíam pequenos ofiuros
entre seus braços, que indica que estão se alimentando próximo ao fundo. Assim, a
sobre-exploração de camarões e moluscos pode contribuir para a diminuição das
populações da família Loliginidae.

Embora não tenham aparecido dentre os mais representativos em Baía Formosa,
do ponto de vista ecológico, os bivalves desempenham um papel importante na
condução de energia e nutrientes da coluna de água, para o bentos no controle da
ciclagem de matéria entre esses compartimentos em águas rasas, podendo inclusive, em
caso de populações densas, controlar a eutrofização. Este grupo, ao lado dos
cefalópodes são os moluscos mais explorados economicamente (Pires-Vanin, 1993). A
baixa quantidade de bivalves encontrada no presente estudo pode estar relacionada às
correntes que aumentam a turbidez local ou à presença a presença abundante
depositivoros e turbadores, como a bolacha-de-praia e os camarões que causam um
amensalismo trófico, turbando o sedimento e dificultanto a captação de nutrientes por
essa classe de moluscos. O hábito de enterrar-se, típico do grupo, também dificulta a
captura por arrastão.
O filo Cnidaria, apesar de ter sido o menos representativo, em número de
indivíduos e espécies, tratou-se do primeiro registro para a costa sul do Rio Grande do
Norte, o que torna importante a presença de octocorais nas capturas em Baía Formosa.

6.CONSIDERAÇÕES FINAIS
De uma forma geral o presente estudo evidenciou que uma fauna diversificada
de organismos epibentônicos na praia de Baía Formosa, os quais podem ser capturados
de forma constante ao longo do ano. As quantidades de organismos coletados, peso e
número, sofreram visível variação sazonal. Entretanto, os resultados não apresentaram
grandes alterações de diversidade e dominância de espécies quando comparados os
meses de coleta, nem quando comparadas a área mais próxima e a mais distante do
estuário do rio Cunhaú. Tal fato pode estar relacionado à morfologia da área de estudo,
uma baía, com as duas áreas analisadas sem indícios diferenças aparentes entre si. O
formato de arco quase fechado desse tipo de praia, delimitada abruptamente por falésias
cobertas pelo campo de dunas e mais protegida de frentes de ondas incidentes,
garantiria condições mais estáveis a localidade, justificando a estabilidade dos índices
avaliados (Diniz, 1998; Amaral, 2000). Outras váriáveis, como o tipo de sedimento,
podem ser determinantes na distribuição da megaepifauna bêntica da região.

REFERÊNCIAS
ABSALÃO, R. S.; PIMENTA, A. D. Moluscos Marinhos da APA do Arquipélago de
Santana, Macaé, RJ. Rio de Janeiro: Editora Ciência Ltda., 2005.
ALENCAR, C. E. R. D; MORAES, A. B.; MORAES, S. A. S. N.; ARAÚJO, P. V. N.;
FREIRE, F. A. M. First record of the association between the porcellanid crab
Porcellana sayana and the brachyuran crab Stratiolibinia bellicosa. Marine
Biodiversity Records, v. 7, p. 1-5, 2014.
ALMEIDA, A. C.; FRANSOZO, V.; FURLAN, M.; GIOVANNETTI, N. Distribuição
ecológica do camarão Nematopalaemon schimitt (HOLTHUIS, 1950)(CRUSTACEA,
DECAPODA, CARIDEA) no litoral norte do estado de São Paulo, Brasil. Anais do VIII
Congresso de Ecologia do Brasil, Caxambu, 2007.
ALMEIDA, Reinaldo Pavan de. A fauna acompanhante do camarão sete barbas na pesca
artesanal de arrasto de portas na região costeira adjacente a Praia Mole e Carapebus-
Espirito Santo, Brasil. 53 pg. Monografia. Universidade Federal do Espírito Santo.
2004.
AMARAL, A.C.Z. (Org.); NALLIN, S.A.H. (Org.). Biodiversidade e Ecossistemas
Bentônicos Marinhos do Litoral Norte de São Paulo, Sudeste do Brasil. 1. ed.
Campinas (SP): UNICAMP/IB, 2011.
AMARAL, R. F. Contribuição ao estudo da evolução morfodinâmica do litoral
oriental sul do Rio Grande do Norte, entre Ponta de Búzios e Baía Formosa. Tese
de doutorado. UFRS. Porto Alegre, 2000.
BARROS, F.; COSTA, P.C.; CRUZ, I.; MARIANO, D.L.S.; MIRANDA R.J. Habitats
Bentônicos na Baía de Todos os Santos. Revista Virtual de Química, 4(5): 551-565,
2012.
BEGON, Michael; HARPER, John L.; TOWNSEND, Colin R. Ecologia - de Indivíduos a
Ecossistemas. 4° ed. Artmed: 2007.
BRAGA, M. S. C. A pesca de arrasto de camarões na zona costeira do município de
Fortaleza, estado do Ceará. 152 pg. Dissertação. Universidade Federal do Ceará. 2000.
BRANCO, J.O.; FRACASSO, H.A.A. Biologia populacional de Callinectes
ornatus Ordway, 1863 na Armação do Itapocoroy, Penha, Santa Catarina,
Brasil. Revista Brasileira de Zoologia, v. 21(1), p. 91-96, 2004.
BRANCO, J.O.; FRACASSO, H.A.A. Ocorrência e abundância da carcinofauna
acompanhante na pesca do camarão sete-barbas Xiphopenaeus kroyeri Heller

(Crustacea, Decapoda), na Armação do Itapocoroy, Penha, Santa Catarina,
Brasil. Revista Brasileira de Zoologia, v. 21(2), p. 295-301, 2004.
BRANCO, J.O.; VERANI, J.R. Pesca do camarão sete-barbas e sua fauna
acompanhante, na Armação do Itapocoroy, Penha, SC. In: BRANCO, Joaquim Olinto;
MARENZI, Adriano W. C. (Org.). Bases ecológicas para um desenvolvimento
sustentável: estudos de caso em Penha, SC. 291. Editora da UNIVALI, Itajaí, SC. p.
153-170.
BUSCHMANN, A.; ASTUDILLO, C.. Pesca de Arrastre, arrasando la vida marina.
Oceana. Santiago de Chile, 2004. pp 16.
CADDY, J. Recent developments in research and management for wild stocks of
bivalves and gastropods. p. 665-699 In: J. Caddy (Ed), Marine Invertebrate Fishery:
their assessment and management. John Wiley Sons, London. 699 p., 1989.
CATTANI, A. P. et al. Avaliação da ictiofauna da fauna acompanhante da pesca do
camarão sete-barbas do município de Pontal do Paraná, litoral do Paraná, Brasil.
Boletim do Instituto de Pesca, São Paulo, 37(2): 247 - 260, 2011.
CARRANZA-FRASER, J.; GRANDE, J.M. Experiencia de Mexico en el
aprovechamienro de la fauna de acompañamiento del camarón. Process of Gulf
Caribbean Fisheries Institute, Miami, 39: 109-111, 1982.
CLARK, A M. & DOWNEY, M. E. Starfishes of the Atlantic. London Chapman & Hall,
1992.
COELHO, P.A.; SANTOS, M.C.F. Época da reprodução do camarão sete-barbas,
(Xiphopenaeus kroyeri) (Heller, 1862) (Crustacea,Decapoda, Penaeidae) na região de
Tamandaré, PE. Técnico Científico CEPENE, Tamandaré, 1 (1): 171-186. 1993.
COELHO, J.A.P.; PUZZI, A.; GRAÇA-LOPEZ, R.; RODRIGUES, E.S.; PRETO JR, O.
Análise da rejeição de peixes na pesca artesanal dirigida ao camarão sete-barbas
(Xiphopenaeus kroyeri) no Litoral de São Paulo.Boletim do Instituto da Pesca, São
Paulo, 13 (2): 51-61. 1986.
COSTA, Igor David da; MADEIRA DI BENEDITTO, Ana Paula. Caracterización
preliminar de los invertebrados bentónicos capturados accidentalmente en la pesca de
camarones en el norte del estado de Río de Janeiro, sudeste de Brasil. Latin American
Journal of Aquatic Research, v. 37, n. 2, p. 259-264, 2009.
COULL, B. C. Ecology of marine meiofauna. In R. P. Higgins &H. Thiel, eds.
Introdution to the study of meiofauna. Washington, D. C., Smithsonian Institution Press,
p. 18-38, 1988.

DFO. Assessment of the impact of northern shrimp trawling on benthic habitats
communities in the Estuary and northern Gulf of St. Lawrence. DFO Can. Sci. Advis.
Sec. Sci. Advis. Rep. 2012/054, 2012.
DINIZ, R.F. Erosão costeira e o desenvolvimento de costa com baías em forma de zeta
no litoral oriental do Estado do Rio Grande do Norte. Resumos. 50a. Reunião Anual da
SBPC. Natal-RN, p.1041, 1998.
DUARTE, D. L. V. Caracterização da fauna acompanhante na pescaria de arrasto de
tangote dirigida a camarões no litoral sul do Brasil. 57 pg. Dissertação. Universidade
Federal do Rio Grande. 2012.
DUPLISEA, Daniel E. et al. A size-based model of the impacts of bottom trawling on
benthic community structure. Canadian Journal of Fisheries and Aquatic Sciences, v.
59, n. 11, p. 1785-1795, 2002.
Empresa de Pesquisa Agropecuária – EMPARN. Disponível em:
<http://www.emparn.rn.gov.br/>. Acesso em: setembro de 2014.
FREIRE, F. A. M. ESTUDO DA DINÂMICA POPULACIONAL DOS CAMARÕES
MARINHOS DE INTERESSE ECONÔMICO NO LITORAL DO ESTADO DO RIO
GRANDE DO NORTE COM ÊNFASE NA ESPÉCIE XIPHOPENAEUS KROYERI
(HELLER, 1862) (CRUSTACEA, DECAPODA, PENAEIDAE). Projeto de Pesquisa.
Universidade Federal do Rio Grande do Norte. 2012.
GRAÇA-LOPES, Roberto da. A pesca do camarão-sete-barbas Xiphopenaeus kroyeri
HELLER (1862) e sua fauna acompanhante no litoral do estado de São Paulo.106 pg.
Tese. Universidade Estadual Paulista. 1996.
GRAÇA-LOPES, Roberto da; et al. Fauna acompanhante da pesca camaroeira no litoral
do estado de São Paulo, Brasil. Boletim do Instituto de Pesca, São Paulo, 28(2): 173 -
188, 2002.
HERRERA, Daphine Ramiro. Distribuição ecológica e dinâmica populacional do
camarão barriga branca Nematopalaemon schmitti (Holthuis, 1950) (Caridea,
Palaemonidae) na região de Macaé/RJ, Brasil. Dissertação de mestrado. Unesp,
Botucatu: 2013.
IBRAHIM, Sakri et al. Seasonal abundance of benthic communities in coral areas of
Karah Island, Terengganu, Malaysia. Turkish Journal of Fisheries and Aquatic
Sciences, v. 6, p. 129-136, 2006.
Instituto de Desenvolvimento Sustentável e Meio Ambiente do Rio Grande do Norte –
IDEMA. Disponível em: <http://www.idema.rn.gov.br/>. Acesso em: março de 2014.

JOHNSON, K.A.A. Review of National and International Literature on the Effects of
fishing on Benthic Habitats. NOAA Technical Memorandun. NMFS-F/SPO-57,
August, 2002.
KNOX, G.A. The ecology of seashores. CRC Press, Londres: 2001.
KOTAS, J. E. Fauna Acompanhante nas Pescarias de Camarão em Santa Catarina.
IBAMA. Coleção Meio-Ambiente, Série Estudos Pesca, Brasília, 1998. v. 24, 76p.
LITTLE, Colin. The Biology of Soft Shores and Estuaries. New York: Oxford, 2009.
LONGHURST, Alan R.; PAULY, Daniel. Ecologia dos oceanos tropicais. São Paulo:
Edusp, 2007.
MADRID VERA, J. et al. Análisis de la fauna de acompañamiento capturado con redes
de arrastre prototipo en el alto golfo de California, como estrategia para reducir la
mortalidad de la vaquita marina. INAPESCA, 2010.
MATTHEWS-CASCON, H.; LOTUFO, T.M.C. Biota Marinha da Costa Oeste do
Ceará. MMA, Brasília. Série Biodiversidade, v. 24, 248p, 2006.
MATTHEWS-CASCON, H.; ROCHA-BARREIRA, C. A; MEIRELLES, C. A. O.
Desovas de Alguns Moluscos Brasileiros. Fortaleza: Expressão Grafica e Editora,
2011. 119 p.
McCONNAUGHEY, R. A.; MIER, K. L. & DEW, C. B. An Examination of chronic
trawling effects on soft-bottom benthos of the eastern Bering Sea. ICES Journal of
Marine Science, v. 57, p. 1377-1388, 2000.
MOTA-ALVES, M.I., RODRIGUES, M.M.. Aspectos da reprodução do camarão sete-
barbas Xiphopenaeuskroyeri (Heller,1862) (Decapoda, Macrura), na costa do estado do
Ceará. Arquivo de Ciência Marinha, Ceará,17 (1): 29-35. 1977.
PERONI, Nivaldo; HERNÁNDEZ, Malva Isabel Medina. Ecologia de populações e
comunidades. 1 ed. Florianópolis : CCB/EAD/UFSC, 2011.v.1.125 p.
PIRES, A. M. S. Structure and dynamics of benthic megafauna on the continental shelf
offshore of Ubatuba, southeastern Brazil. Marine Ecology Progress Series. v. 86, p.
63-76, 1992.
PIRES-VANIN, A. M. S. A Macrofauna Bentica da Plataforma Continental ao Largo de
Ubatuba, Sao Paulo, Brasil. Publicação Especial do Instituto Oceanográfico, v. 10, p.
137-158, 1993.
RÁBAGO-QUIROZ, Carlos Hiram et al. Distribución latitudinal y batimétrica de las
especies más abundantes y frecuentes en la fauna acompañante del camarón del Golfo
de California, México. Revista de Biología Tropical, v. 59, n. 1, p. 255-267, 2011.

REISS, Henning; KRÖNCKE, Ingrid. Seasonal variability of benthic indices: an
approach to test the applicability of different indices for ecosystem quality
assessment. Marine Pollution Bulletin, v. 50, n. 12, p. 1490-1499, 2005.
RIOS, Eliézer de Carvalho. Compendium of Brazilian Sea Shells. Rio Grande. Editora
Evangraf, 2009.
ROBERT, R.; BORZONE, C. A.; NATIVIDADE, C. D.. Os camarões da fauna
acompanhante na pesca dirigida ao camarão sete-barbas (Xiphopenaeus kroyeri) no
litoral do Paraná. Boletim do Instituto de Pesca, São Paulo, 33(2): 237 - 246, 2007.
RUEDA, Jose L.; SALAS, Carmen. Seasonal variation of a molluscan assemblage
living in a Caulerpa prolifera meadow within the inner Bay of Cádiz (SW
Spain). Estuarine, Coastal and Shelf Science, v. 57, n. 5, p. 909-918, 2003.
RUFFINO, M.L ; CASTELLO, J.P.. Alterações na ictiofauna acompanhante da pesca do
camarão-barba-ruça (Artemesia longinaris) nas imediações da Barra do Rio Grande.
Neurítica, Curitiba, 7(1-2): 43-55. 1992.
RUPPERT, E. E. Zoologia dos Invertebrados. 6.ed. São Paulo: Roca, 1996.
SANTOS, S.; RIEGER, P. S.; VIEIRA, R.R.R.; BARUTOT, R.A. Composição e
distribuição dos Crustacea (Decapoda) na Lagoa do Peixe, Rio Grande do Sul, Brasil.
Revista Brasileira Zoologia, v. 17(1), p. 213-223, 2000.
SANTOS, M.C.F.; PEREIRA, J. A.; IVO, C. T. C.. A pesca do camarão branco
Litopenaeus schmitti (Burkenroad, 1936) (Crustacea, Decapoda, Penaeidade) no
nordeste do Brasil. Bol. Téc. Cient. CEPENE, v.14, n.1, p. 33-58, 2006.
SOARES-GOMES, A.; PITOMBO, F.B.; PAIVA; P.C. Bentos de Sedimentos não
Consolidados. In: PEREIRA, R.C.; SOARES-GOMES, A.(Org.). Biologia Marinha. 2
ed. Rio de Janeiro: Interciência, 2009. p. 319-337.
TATARANNI, Mariella; LARDICCI, Claudio. Performance of some biotic indices in
the real variable world: A case study at different spatial scales in North-Western
Mediterranean Sea. Environmental pollution, v. 2010, 2010.
THOMÉ, J. W.; AYDOS BERGONCI, P. E.; GIL, G. M. As conchas das nossas praias:
guia ilustrado. Pelotas, RS: Editora USEB, 2004. 96 p.
TURRA, Alexander; BRANCO, Joaquim Olinto; SOUTO, Flávio Xavier Souto.
Population biology of the hermit crab Petrochirus diogenes (Linnaeus, 1758) in
Southern Brazil. Revta bras. Zool. 19(4):1043-1051, 2002

VELOSO, V. G.; NEVES, G. Praias Arenosas. In: PEREIRA, R.C.; SOARES-GOMES,
A.(Org.). Biologia Marinha. 2 ed. Rio de Janeiro: Interciência, 2009. p. 339-360.
ZOLDAN, Liliana Vala. Analise do conteúdo estomacal de duas espécies de estrela-
do-mar: Astropecten marginatus (Gray 1840) e Luidia senegalensis (Lamarck, 1816)
na Enseada de Caraguatatuba, SP. Sudeste do Brasil. Monografia. Centro
Universitário da Fundação de Ensino Octávio Bastos, São João da Boa Vista, SP: 2005














