
ULTRASTRUCTURE AND CHAETOTAXY OF SENSORY RECEPTORS IN THE CERCARIAE OF A SPECIES OF CREPIDOSTOMUM...
15
The American Society of Parasitologists is collaborating with JSTOR to digitize, preserve and extend access to The Journal of Parasitology. http://www.jstor.org Ultrastructure and Chaetotaxy of Sensory Receptors in the Cercariae of a Species of Crepidostomum Braun, 1900 and Bunodera Railliet, 1896 (Digenea: Allocreadiidae) Author(s): T. Bogéa and J. N. Caira Source: The Journal of Parasitology, Vol. 87, No. 2 (Apr., 2001), pp. 273-286 Published by: The American Society of Parasitologists Stable URL: http://www.jstor.org/stable/3285041 Accessed: 24-07-2015 03:09 UTC Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at http://www.jstor.org/page/ info/about/policies/terms.jsp JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTC All use subject to JSTOR Terms and Conditions
-
Upload
celsolisboa -
Category
Documents
-
view
4 -
download
0
Transcript of ULTRASTRUCTURE AND CHAETOTAXY OF SENSORY RECEPTORS IN THE CERCARIAE OF A SPECIES OF CREPIDOSTOMUM...

The American Society of Parasitologists is collaborating with JSTOR to digitize, preserve and extend access to The Journal ofParasitology.
http://www.jstor.org
Ultrastructure and Chaetotaxy of Sensory Receptors in the Cercariae of a Species of Crepidostomum Braun, 1900 and Bunodera Railliet, 1896 (Digenea: Allocreadiidae) Author(s): T. Bogéa and J. N. Caira Source: The Journal of Parasitology, Vol. 87, No. 2 (Apr., 2001), pp. 273-286Published by: The American Society of ParasitologistsStable URL: http://www.jstor.org/stable/3285041Accessed: 24-07-2015 03:09 UTC
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at http://www.jstor.org/page/ info/about/policies/terms.jsp
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected].
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions

J. Parasitol., 87(2), 2001. p. 273-286 ? American Society of Parasitologists 2001
ULTRASTRUCTURE AND CHAETOTAXY OF SENSORY RECEPTORS IN THE CERCARIAE OF A SPECIES OF CREPIDOSTOMUM BRAUN, 1900 AND BUNODERA RAILLIET, 1896 (DIGENEA: ALLOCREADIIDAE)
T. Bogea* and J. N. Caira Department of Ecology and Evolutionary Biology, University of Connecticut, Storrs, Connecticut, 06269-3043
ABSTRACT: Previous investigations of cercarial sensory systems have focused on chaetotaxy and ultrastructure of sensory recep- tors and have revealed chaetotaxic patterns within families, genera, and species as well as different types of sensory receptors. However, chaetotaxic and ultrastructural observations have rarely been combined. We investigated the ultrastructure of cercarial sensory receptors in conjunction with the chaetotaxy and neuromorphology in 2 allocreadiid species belonging to the genera Crepidostomum and Bunodera. Cercariae were treated with acetylthiocholine iodide and silver nitrate, and for scanning electron microscopy (SEM) and transmission electron microscopy (TEM). Similar cholinergic nerve networks were revealed. Chaetotaxy was consistent with that of other allocreadiids. Seven and 6 types of receptors were distinguished with SEM in Crepidostomum sp. and Bunodera sp., respectively. Types differed in number of cilia (1 or 2), cilium length (short, moderately long, or long), presence or absence of a tegumentary collar and a domelike base, and tegumentary collar length (low, moderately low, or high). TEM of some types revealed unsheathed cilia, basal body, and thickened nerve collars. Some receptor types were site specific. Thus, long uniciliated receptors were concentrated on the dorsal surface. Other types, such as short uniciliated receptors, were widespread throughout most regions. Ultrastructure and site-specificity observations suggest that most receptors are mechanore- ceptors.
The morphology of sensory receptors across the cercarial tegument has been under investigation for > 100 yr (Bettendorf, 1897). Recently, ultrastructural studies have revealed various types of cercarial sensory receptors within a single species (Short and Cartrett, 1973; Zdarska, 1992; Abdul-Salam and Sreelatha, 1998) and among different species (Matricon-Gon- dran, 1971; Koie, 1981; Krejci and Fried, 1994). These receptor types have been distinguished by the presence or absence of a ciliary sheath, a tegumentary collar surrounding the cilium, and villi on the tegumentary collar as well as number of cilia, cilium length, tegumentary collar length, and tegumentary collar width. Characters such as presence of a basal body, a basal plate, a ciliary rootlet, and membrane reinforcements within the nerve bulb also have been used to characterize the different types (Niewiadomska and Czubaj, 1996).
Other studies have emphasized the distribution of sensory re- ceptors across the larval tegument. Using the method of silver nitrate impregnation, Richard (1971) and Bayssade-Dufour (1979) among others have demonstrated a marked uniformity in elements of chaetotaxic patterns at the family, genus, and species levels. Recently, neuromorphological investigations have contrib- uted to the chaetotaxic nomenclatures (Bayssade-Dufour et al., 1989, 1993; Grabda-Kazubska and Kiseliene, 1989).
Richard (1971) recognized that chaetotaxic investigations would profit from combination with ultrastructural observa- tions, mainly because silver nitrate impregnation does not dis- tinguish among the various types of cercarial sensory receptors. Thus, a detailed study of the cercaria of a species combining silver nitrate impregnation and electron microscopy techniques would allow description of each sensory receptor type observed in the chaetotaxic pattern, providing a new perspective on the study of cercarial sensory systems (Bayssade-Dufour, 1979).
The development of a standard procedure that combines ultra-
structural, chaetotaxic, and neuromorphological observations is, therefore, fundamental. In this sense, standard protocols must be developed not only for combining various techniques but also for each technique individually to overcome the limitations of even the most widely used techniques (Sweeting, 1976; Blair, 1977; Bayssade-Dufour et al., 1989; Niewiadomska, 1994).
In the present study, we investigated the ultrastructure of cer- carial sensory receptors in conjunction with their distribution on the tegument and their position relative to the nerve network, aiming to develop a standard procedure for the examination of species using a combination of techniques. For this purpose, cercariae of 2 allocreadiid species belonging to Crepidostomum Braun, 1900 and Bunodera Railliet, 1896 were studied with the acetylthiocholine method for cholinesterase, silver nitrate im- pregnation, scanning electron microscopy (SEM), and trans- mission electron microscopy (TEM).
MATERIALS AND METHODS Specimen collection and identification
Bottom sediment was collected with dip nets and was sieved through 0.25-inch-mesh aluminum screening. A total of 121 individuals of the fingernail clams Musculium partumeium (Say, 1822) and Pisidium cas- ertanum (Poli, 1795) (Bivalvia: Sphaeriidae) was collected in Halls Pond, Windham, Connecticut, from April through September 1996. A total of 356 individuals of the fingernail clams Pisidium aequilaterale (Lea, 1852) (Bivalvia: Sphaeriidae) and P. casertanum was collected in Pachaug Pond, Griswold, Connecticut, from July through September 1998. The clams were transported to the laboratory in plastic vials (2 feet long x 1.5 feet wide) containing pond water.
In the laboratory, the infected clams were identified and individually isolated in glass vials with 5 ml of pond water. They were then exposed to artificial light (60 watts) and temperature (approximately 30 C for up to 1 hr) every 2 days for up to 2 wk to induce cercarial emergence. Cercariae were obtained from 7 infected clams collected in Halls Pond and from 6 infected clams collected in Pachaug Pond, respectively. The most actively emerging cercariae were collected with fine glass pipettes and processed according to the following protocols.
Twenty cercariae of each species were processed as whole mounts (Cable and Peters, 1986). They were fixed in hot 5% buffered formalin phosphate, transferred to 70% ethanol, hydrated in ethanol, stained with Semichon's acetocarmine, dehydrated in ethanol, cleared in methyl salic- ylate, and mounted in Canada balsam. Illustrations of each specimen showing general morphology were made with the aid of a drawing tube.
Received 31 March 2000; revised 22 August 2000; accepted 22 Au- gust 2000.
* Departamento de Medicina Tropical, Instituto Oswaldo Cruz, Funda-
qio Oswaldo Cruz, Av. Brasil 4365, Manguinhos, Rio de Janeiro 21045-900, Brazil.
273
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions

274 THE JOURNAL OF PARASITOLOGY, VOL. 87, NO. 2, APRIL 2001
Taxonomic identifications followed descriptions by Hopkins (1934), Can- non (1971), and Caira (1989). Ultrastructural characters such as presence of spherical ventral sucker protrusions and relative sizes of anterior and posterior ventral sucker protrusions were used to verify the identification of the cercariae to family and genus, respectively (Caira, 1989). Moreover, previous parasite surveys conducted at the collection sites were consid- ered (State Board of Fisheries and Game, 1959; Olson, 1985). The present taxa possibly belong in the species Crepidostomum cooperi Hopkins, 1931 and Bunodera sacculata Van Cleave and Mueller, 1932. However, they are referred to here as Crepidostomum sp. and Bunodera sp. because cercariae were not reared through the second intermediate and definitive hosts and thus specific identification could not be confirmed. Voucher specimens of the cercariae of each species have been deposited in the U.S. National Parasite Collection, Beltsville, Maryland, and in the Larry R. Penner Parasitology Collection, Department of Ecology and Evolu- tionary Biology, University of Connecticut, Storrs.
Specimen preparation and study Acetylthiocholine method for cholinesterase: The acetylthiocholine
method for cholinesterase of Thompson (1966) was adapted for the mapping of the nerve networks and for the identification of distinct nerve regions and sensory receptors associated with each region in both species. Twenty cercariae of each species were fixed in cold 4% para- formaldehyde in phosphate buffer (pH 7.4) for approximately 10 min, incubated in acetylthiocholine iodide substrate at 37 C for 40 min, rinsed in a saturated solution of sodium sulfate, washed thoroughly in distilled water, and developed with 3% silver nitrate for 1 min. Cercariae were then treated with Bodian's developer (Presnell and Schreibman, 1997), fixed in a solution of 1.5% glutaraldehyde/1.5% formaldehyde in 0.1 M cacodylate buffer (pH 7.35), dehydrated in ethanol, cleared in methyl salicylate, and mounted in Canada balsam. Controls consisted of preincubation of 20 specimens of each species in 1 x 10-5 M eserine sulfate in phosphate buffer to inactivate nonspecific cholinesterases, fol- lowed by treatment with the above protocol. Illustrations of each spec- imen recording the basic arrangement of the nerve network were made with the aid of a drawing tube. A single illustration summarizing the arrangements seen in all cercariae represented a map of the basic nerve network for the species. Nerve commissures and cords were named according to the lettering system proposed by Richard (1971) and Bays- sade-Dufour et al. (1993).
Silver nitrate impregnation: The standard procedure for silver nitrate impregnation as described by Combes et al. (1976) allowed the mapping of sensory receptors on the cercarial tegument of each species. Ap- proximately 100 cercariae of each species were fixed in cold 3% silver nitrate solution, exposed to an artificial source of ultraviolet light for 10 min, rinsed in cold distilled water, and mounted in Hoyer's medium. Illustrations recording the distribution of sensory receptors on the cer- carial tegument of each specimen were made with the aid of a drawing tube. The chaetotaxy of each species was subsequently determined us- ing the procedure described by Bog6a (1999). A grid schematically subdivided each specimen into chaetotaxic sites. The comparison of the illustrations obtained for each specimen allowed the determination of the range of receptors for each site. This range was divided into classes of 1-receptor intervals, and the frequency of each class was recorded. The modal class was selected to represent the chaetotaxy of each site. A single illustration summarizing the illustrations of each specimen showed the basic chaetotaxic pattern for each species.
SEM: Micrographs obtained with SEM were used to develop a de- scription of the external ultrastructure of sensory receptors and to iden- tify characters that might reflect their sensory modalities. Fifty cercariae of each species were fixed in a cold solution of 1.5% glutaraldehyde/ 1.5% formaldehyde in 0.1 M cacodylate buffer (pH 7.35) overnight, postfixed in cold aqueous solution of 1% osmium tetroxide overnight, transferred to 70% ethanol, and hydrated in ethanol. The specimens were rinsed in cold distilled water, dehydrated in ethanol, air dried in hexamethyldisilazane (Ted Pella, Redding, California) for 10 min, and mounted on aluminum stubs with double-sided adhesive carbon tape. Specimens were then sputter-coated with approximately 100 A of gold/ palladium and examined using a Leo/Zeiss DSM982 Gemini field emis- sion scanning electron microscope.
The external ultrastructure of each sensory receptor type was de- scribed based on characters previously established in the literature. The types of sensory receptors were distinguished in at least 10 different
specimens of each species based on number of cilia, cilium width (de- fined as the arithmetic mean of largest and smallest cilium widths), presence or absence of a tegumentary collar, tegumentary collar width (defined as the arithmetic mean of largest and smallest tegumentary collar widths), and presence or absence of a domelike base. They were also characterized by cilium length (defined as the visible portion of the cilium extending beyond the tegument) and tegumentary collar length (defined as the length of the extension from the point of its insertion in the tegument to the tip of its collar). Thus, the sensory receptors could exhibit very short (<1.0 p m long), short (approximately 1.0 Vpm long), moderately long (approximately 2.0-3.0 Vpm long), or long (>3.0 Vpm long) cilia and a low (<0.3 Vpm long), a moderately low (approximately 0.3-0.5 Vpm long), a high (approximately 0.5 Vpm long), or a very high (>0.5 Vpm long) tegumentary collar. All measurements are in micro- meters. Means are followed by standard deviations and number of struc- tures measured.
For identification purposes, each type of sensory receptor was des- ignated with a letter code. The first letter indicates the number of cilia, where U = uniciliated and B = biciliated. This letter is followed by a Roman numeral, which indicates the receptor type designation, and the letters C or B corresponding to Crepidostomum and Bunodera, respec- tively.
TEM: Micrographs obtained with TEM were used to identify addi- tional characters that might reflect the sensory modalities in a subset of types of receptors. Fifty cercariae of each species were fixed in a cold solution of 1.5% glutaraldehyde/1.5% formaldehyde/1.0% acrolein in 0.1 M cacodylate buffer (pH 7.35) for 2 hr, rinsed in cold 0.1 M cac- odylate buffer for a total of I hr, and postfixed in 1% osmium tetroxide in 0.1 M cacodylate buffer for 2 hr on ice. The specimens were washed in cold distilled water, dehydrated in ethanol, and embedded in 100% LRWhite (viscosity of 8 centipoises at 25 C) resin (Polysciences, War- rington, Pennsylvania). Gold serial sections from at least 10 specimens of each species were cut using a LKB Ultratome III microtome with a diamond knife and then mounted on uncoated copper grids. The sec- tions were stained with 2% uranyl acetate and 2.5% lead citrate and examined and photographed using a Phillips transmission electron mi- croscope model 300.
The internal ultrastructure of sensory receptors was characterized based on the following features defined by Lyons (1969): presence or absence of a sensory bulb, a basal plate, a basal body, a ciliary rootlet, a tegumentary membrane, nerve collars, and neuronal vesicles, diameter of neuronal vesicles, shape of junctional complexes, and microtubular arrangement of the cilium.
Combination of methods: For each species, the results obtained were combined as follows. First, chaetotaxic maps were superimposed on the nerve network maps to identify the position of each receptor relative to the transverse commissures of each nerve region using the nomenclature proposed by Richard (1971), Bayssade-Dufour (1979), Bayssade-Du- four et al. (1989, 1993), and Grabda-Kazubska and Kiseliene (1989). Chaetotaxic maps were then used in the SEM studies to investigate the staining of tegumentary structures other than sensory receptors with silver nitrate and to describe the external ultrastructure of each sensory receptor detected on the cercarial tegument. A final map combining ultrastructure and chaetotaxy of sensory receptors of each species al- lowed the investigation of site specificity among the types of receptors.
RESULTS
Nerve network
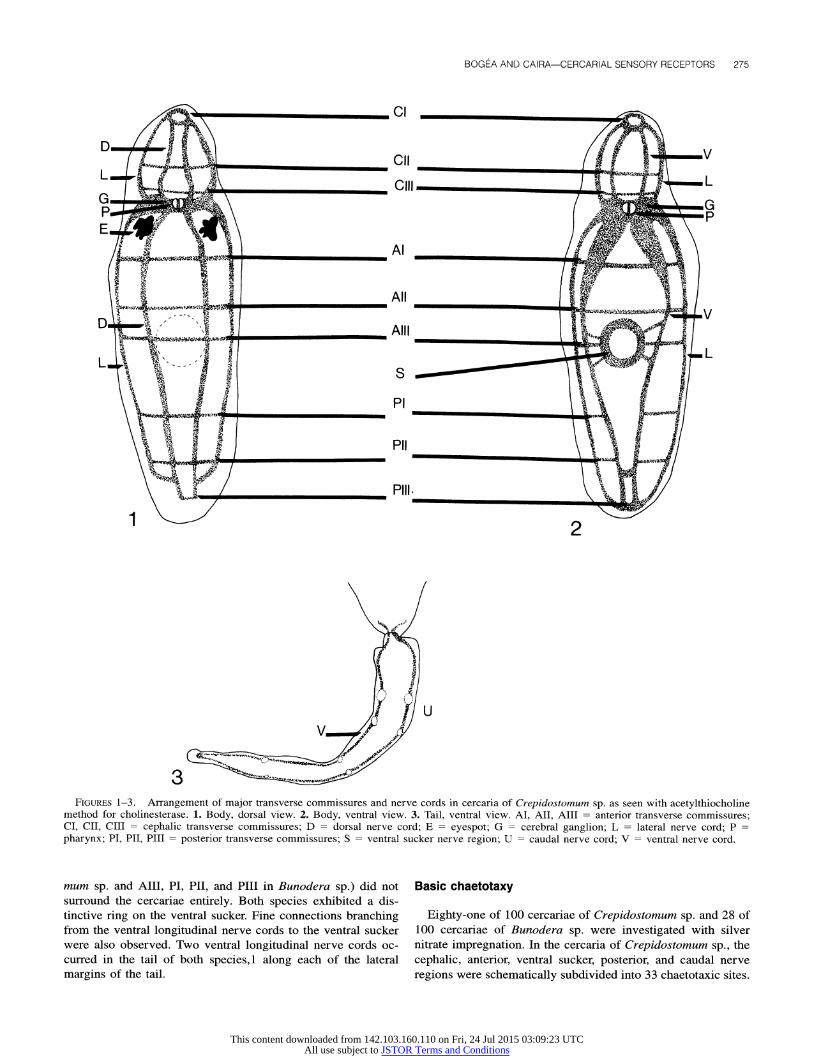
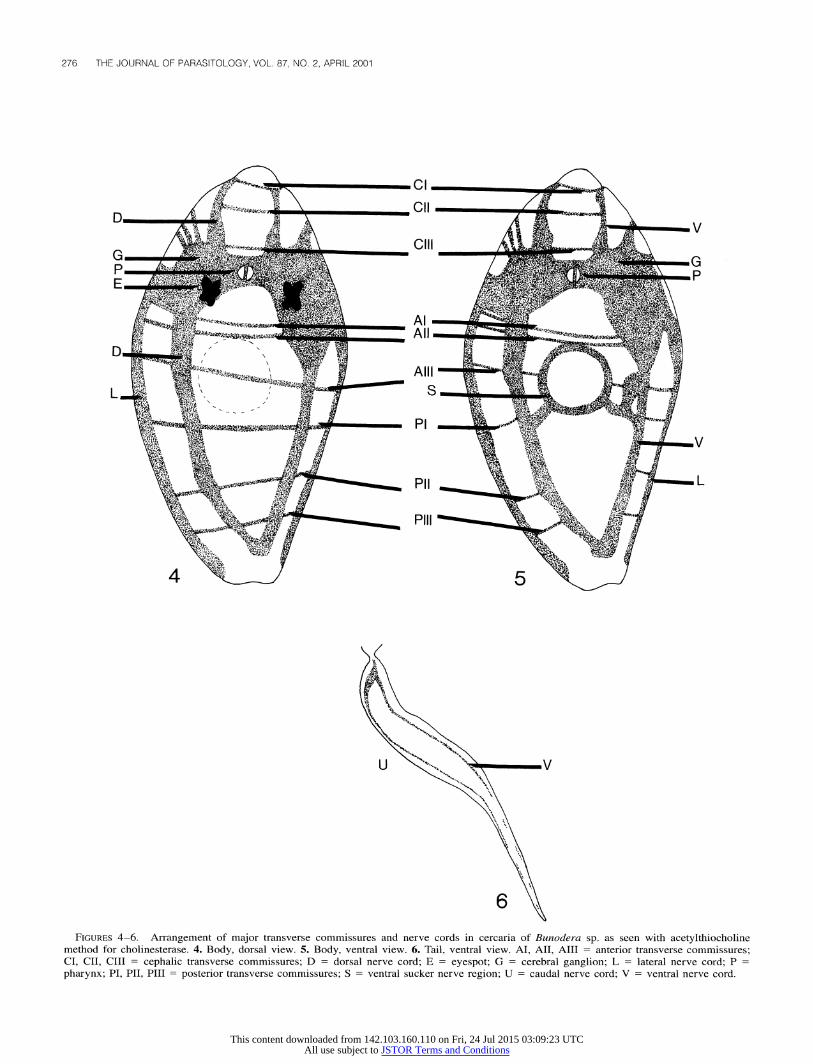
Ten of 20 cercariae of each species were studied with the acetylthiocholine method for cholinesterase. The basic arrange- ments of the nerve networks are presented in Figures 1-3 for Crepidostomum sp. and in Figures 4-6 for Bunodera sp. In both species, the nervous system consisted of a bilobed cerebral gan- glion at the level of the pharynx and 3 pairs of lateral, dorsal, and ventral longitudinal nerve cords, which extended anteriorly and posteriorly from the cerebral ganglion. Three cephalic (CI, CII, and CIII), 3 anterior (AI, AII, and AIII), and 3 posterior (PI, PII, and PIII) transverse commissures were identified. Some of these commissures (AIII, PI, and PII in Crepidosto-
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions
BOGEA AND CAIRA-CERCARIAL SENSORY RECEPTORS 275
Cl
Do
A ill G G
Al
SAll
Alll
i,.
P i
Pii PN
Pill -~
FIGURES 1-3. Arrangement of major transverse commissures and nerve cords in cercaria of Crepidostomum sp. as seen with acetylthiocholine method for cholinesterase. 1. Body, dorsal view. 2. Body, ventral view. 3. Tail, ventral view. AI, AII, AIII = anterior transverse commissures; CI, CII, CIII = cephalic transverse commissures; D = dorsal nerve cord; E = eyespot; G = cerebral ganglion; L = lateral nerve cord; P pharynx; PI, PII, PIII = posterior transverse commissures; S = ventral sucker nerve region; U = caudal nerve cord; V = ventral nerve cord.
mum sp. and AIII, PI, PII, and PIII in Bunodera sp.) did not surround the cercariae entirely. Both species exhibited a dis- tinctive ring on the ventral sucker. Fine connections branching from the ventral longitudinal nerve cords to the ventral sucker were also observed. Two ventral longitudinal nerve cords oc- curred in the tail of both species,1 along each of the lateral margins of the tail.
Basic chaetotaxy
Eighty-one of 100 cercariae of Crepidostomum sp. and 28 of 100 cercariae of Bunodera sp. were investigated with silver nitrate impregnation. In the cercaria of Crepidostomum sp., the cephalic, anterior, ventral sucker, posterior, and caudal nerve regions were schematically subdivided into 33 chaetotaxic sites.
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions
276 THE JOURNAL OF PARASITOLOGY, VOL. 87, NO. 2, APRIL 2001
iCI
.',• ..•.
,MO . ,ill
F~fv .-f
A. 5f• •
J'. :.-'
!' ,--. 'r ... -'t• ,
L._ ', N.,
NNR
G G PM:M4
Not W
-` "tt•"•:•""•~~-f~;
l~,~~
Wu 0-
~L
3~~F?;?;~t,~ ?S~;~t......U
\ . 'X I , :~"-~.. \
1;6z
Pilli
FIGURES 4-6. Arrangement of major transverse commissures and nerve cords in cercaria of Bunodera sp. as seen with acetylthiocholine method for cholinesterase. 4. Body, dorsal view. 5. Body, ventral view. 6. Tail, ventral view. AI, AII, AIII = anterior transverse commissures; CI, CII, CIII = cephalic transverse commissures; D = dorsal nerve cord; E = eyespot; G = cerebral ganglion; L = lateral nerve cord; P pharynx; PI, PII, PIII= posterior transverse commissures; S = ventral sucker nerve region; U = caudal nerve cord; V = ventral nerve cord.
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions
BOGEA AND CAIRA-CERCARIAL SENSORY RECEPTORS 277
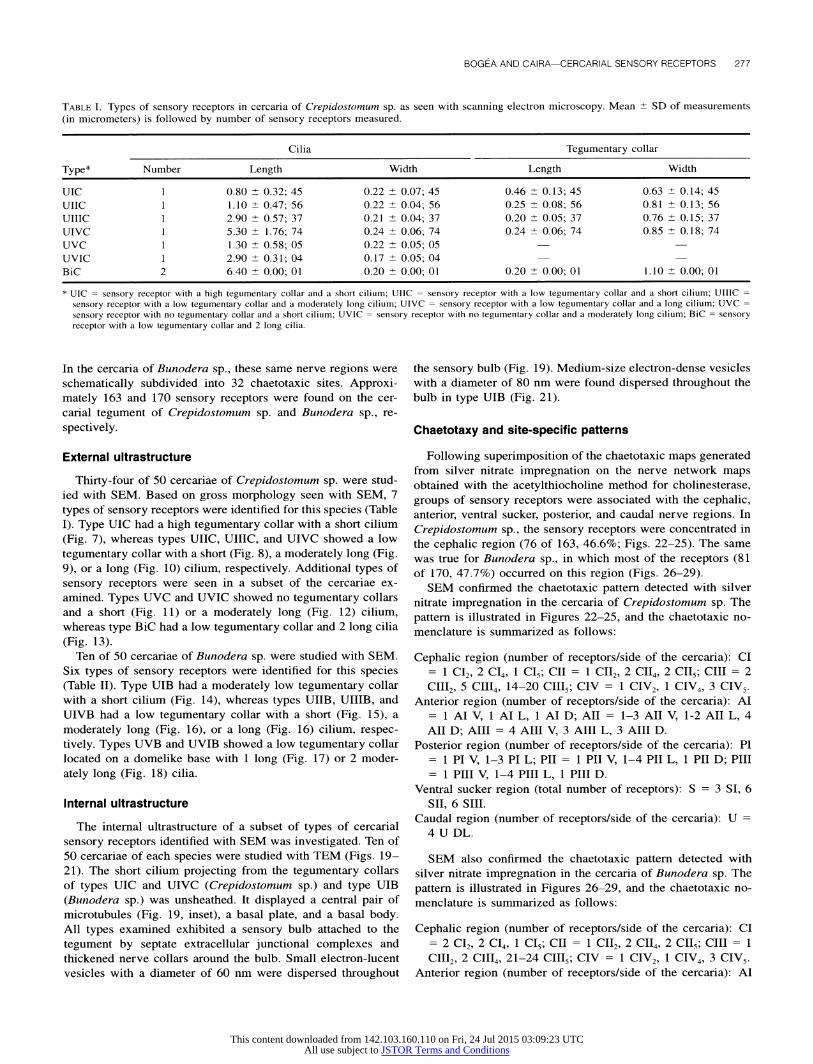
TABLE I. Types of sensory receptors in cercaria of Crepidostomum sp. as seen with scanning electron microscopy. Mean ?- SD of measurements (in micrometers) is followed by number of sensory receptors measured.
Cilia Tegumentary collar
Type* Number Length Width Length Width
UIC 1 0.80 1 0.32; 45 0.22 1 0.07; 45 0.46 + 0.13; 45 0.63 1 0.14; 45
UIIC 1 1.10 1 0.47; 56 0.22 1 0.04; 56 0.25 1 0.08; 56 0.81 1 0.13; 56
UIIIC 1 2.90 1 0.57; 37 0.21 0.04; 37 0.20 1 0.05; 37 0.76 1 0.15; 37
UIVC 1 5.30 1 1.76; 74 0.24 1 0.06; 74 0.24 1 0.06; 74 0.85 1 0.18; 74
UVC 1 1.30 1 0.58; 05 0.22 1 0.05; 05
UVIC 1 2.90 1 0.31; 04 0.17 1 0.05; 04
BiC 2 6.40 1 0.00; 01 0.20 1 0.00; 01 0.20 1 0.00; 01 1.10 1 0.00; 01
* UIC = sensory receptor with a high tegumentary collar and a short cilium; UIIC = sensory receptor with a low tegumentary collar and a short cilium; UIIIC = sensory receptor with a low tegumentary collar and a moderately long cilium; UIVC = sensory receptor with a low tegumentary collar and a long cilium; UVC = sensory receptor with no tegumentary collar and a short cilium; UVIC = sensory receptor with no tegumentary collar and a moderately long cilium; BiC = sensory receptor with a low tegumentary collar and 2 long cilia.
In the cercaria of Bunodera sp., these same nerve regions were
schematically subdivided into 32 chaetotaxic sites. Approxi- mately 163 and 170 sensory receptors were found on the cer- carial tegument of Crepidostomum sp. and Bunodera sp., re-
spectively.
External ultrastructure
Thirty-four of 50 cercariae of Crepidostomum sp. were stud- ied with SEM. Based on gross morphology seen with SEM, 7
types of sensory receptors were identified for this species (Table I). Type UIC had a high tegumentary collar with a short cilium (Fig. 7), whereas types UIIC, UIIIC, and UIVC showed a low
tegumentary collar with a short (Fig. 8), a moderately long (Fig. 9), or a long (Fig. 10) cilium, respectively. Additional types of
sensory receptors were seen in a subset of the cercariae ex- amined. Types UVC and UVIC showed no tegumentary collars and a short (Fig. 11) or a moderately long (Fig. 12) cilium, whereas type BiC had a low tegumentary collar and 2 long cilia
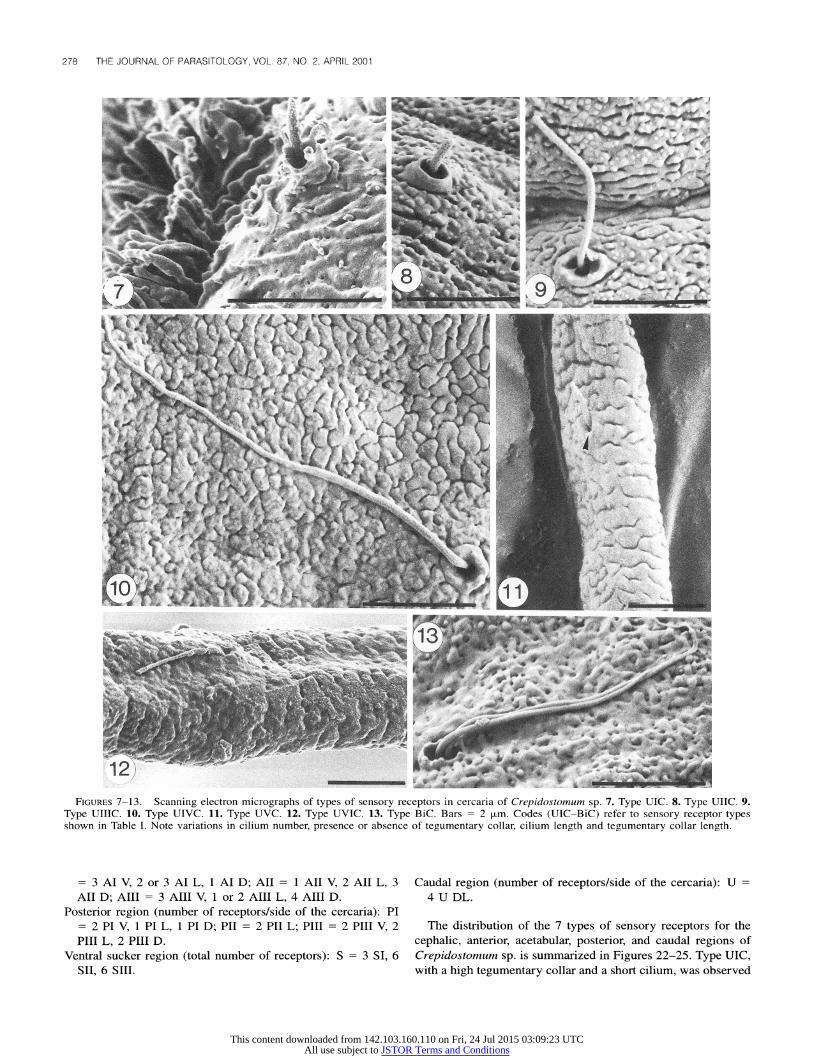
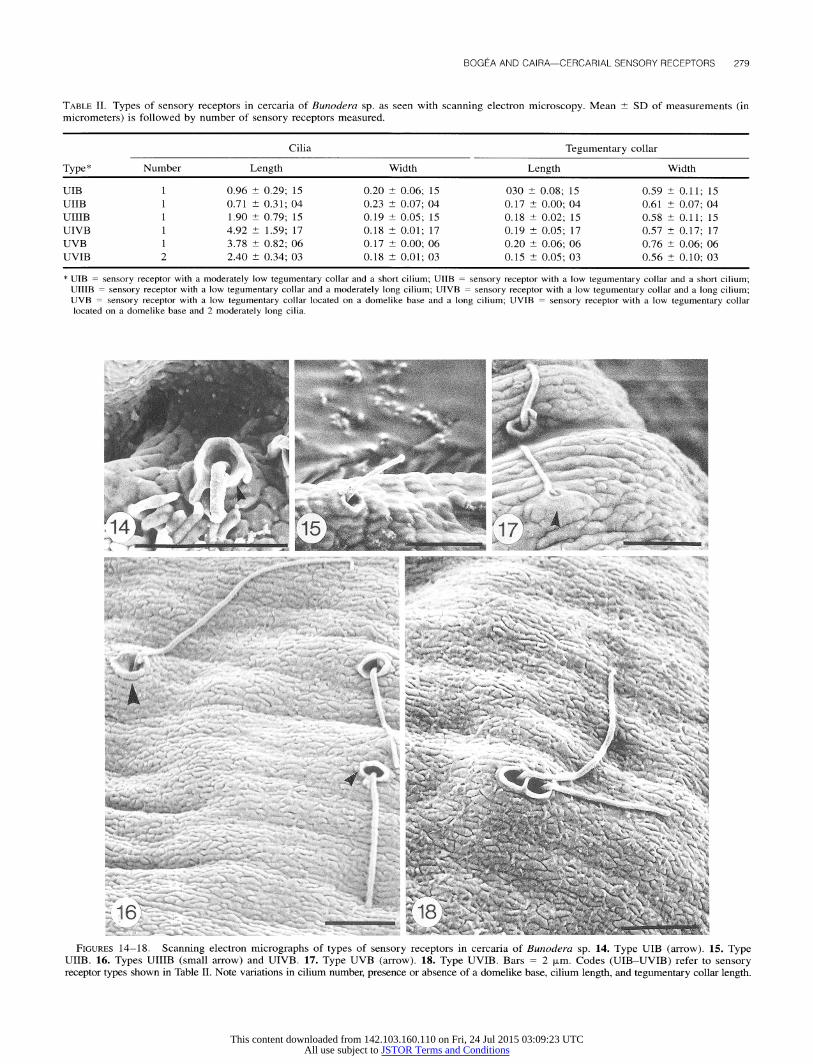
(Fig. 13). Ten of 50 cercariae of Bunodera sp. were studied with SEM.
Six types of sensory receptors were identified for this species (Table II). Type UIB had a moderately low tegumentary collar with a short cilium (Fig. 14), whereas types UIIB, UIIIB, and UIVB had a low tegumentary collar with a short (Fig. 15), a
moderately long (Fig. 16), or a long (Fig. 16) cilium, respec- tively. Types UVB and UVIB showed a low tegumentary collar located on a domelike base with 1 long (Fig. 17) or 2 moder-
ately long (Fig. 18) cilia.
Internal ultrastructure
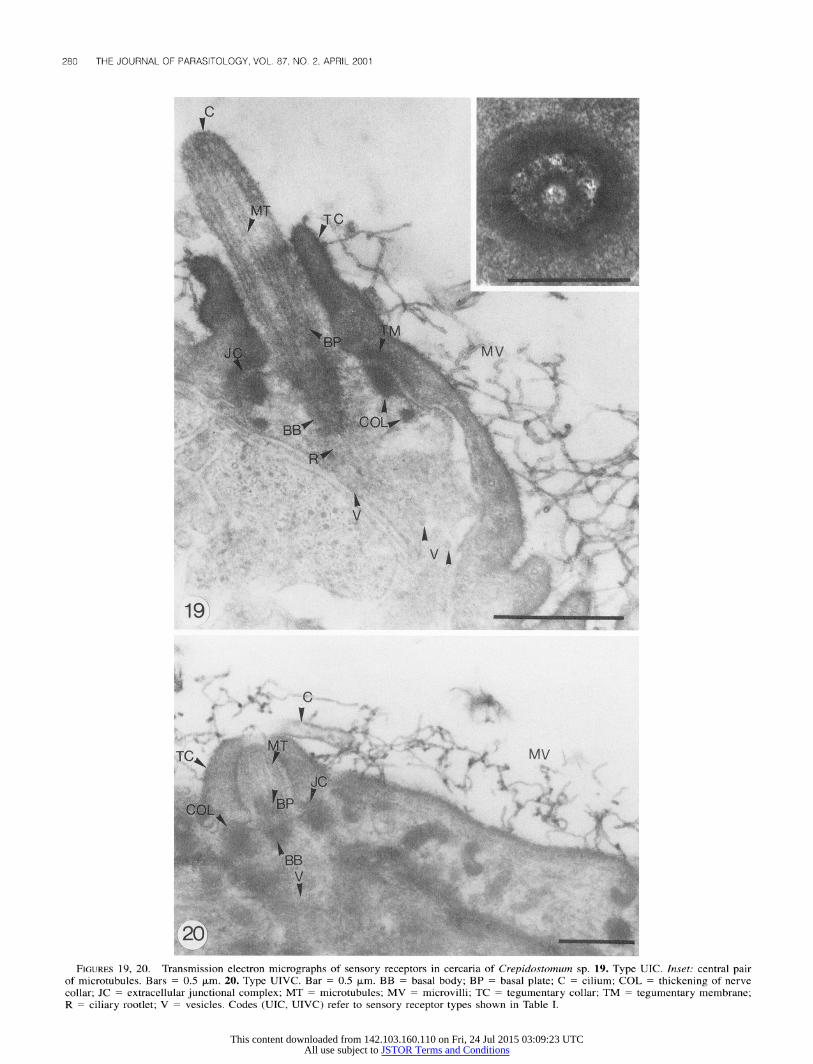
The internal ultrastructure of a subset of types of cercarial
sensory receptors identified with SEM was investigated. Ten of 50 cercariae of each species were studied with TEM (Figs. 19- 21). The short cilium projecting from the tegumentary collars of types UIC and UIVC (Crepidostomum sp.) and type UIB (Bunodera sp.) was unsheathed. It displayed a central pair of microtubules (Fig. 19, inset), a basal plate, and a basal body. All types examined exhibited a sensory bulb attached to the
tegument by septate extracellular junctional complexes and thickened nerve collars around the bulb. Small electron-lucent vesicles with a diameter of 60 nm were dispersed throughout
the sensory bulb (Fig. 19). Medium-size electron-dense vesicles with a diameter of 80 nm were found dispersed throughout the bulb in type UIB (Fig. 21).
Chaetotaxy and site-specific patterns
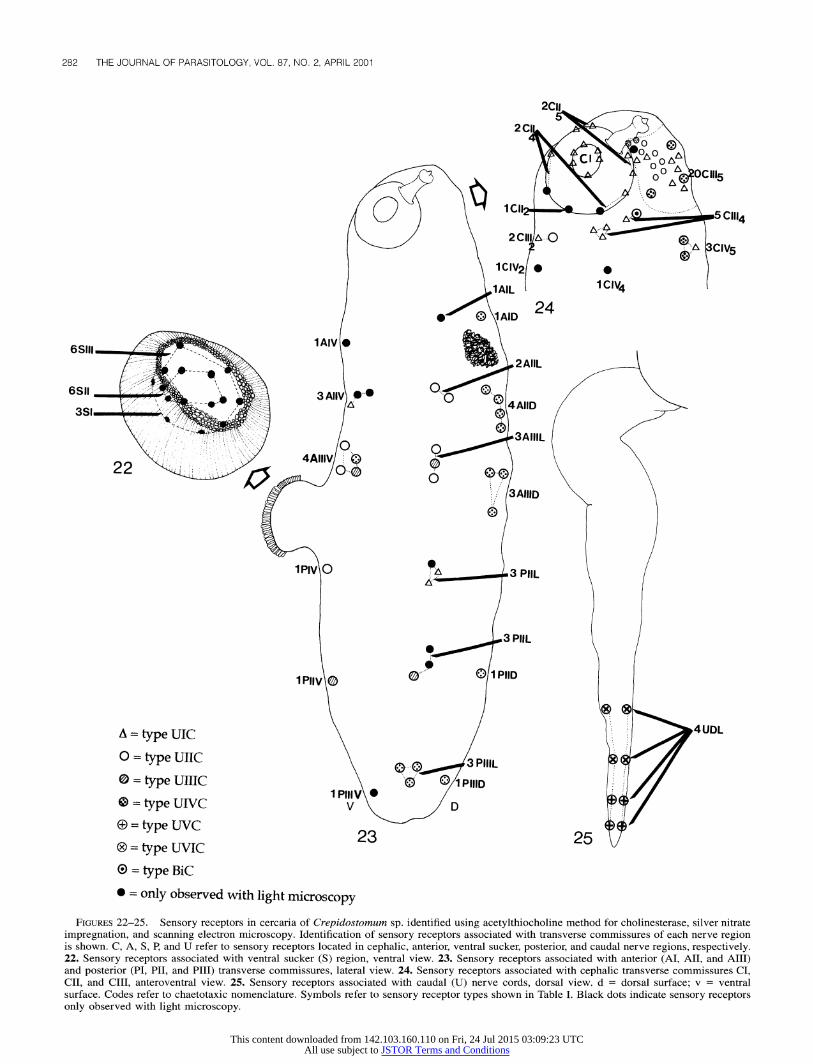
Following superimposition of the chaetotaxic maps generated from silver nitrate impregnation on the nerve network maps obtained with the acetylthiocholine method for cholinesterase, groups of sensory receptors were associated with the cephalic, anterior, ventral sucker, posterior, and caudal nerve regions. In
Crepidostomum sp., the sensory receptors were concentrated in the cephalic region (76 of 163, 46.6%; Figs. 22-25). The same was true for Bunodera sp., in which most of the receptors (81 of 170, 47.7%) occurred on this region (Figs. 26-29).
SEM confirmed the chaetotaxic pattern detected with silver nitrate impregnation in the cercaria of Crepidostomum sp. The
pattern is illustrated in Figures 22-25, and the chaetotaxic no- menclature is summarized as follows:
Cephalic region (number of receptors/side of the cercaria): CI = 1 CI2, 2 CI42, 2 CII4, 1 C2 CII; CII CIII = 1 C2, 2 CII, 2 Cu5; CII = 2
CIII2, 5 CIII4, 14-20 CIII5; CIV = 1 CIV2, 1 CIV4, 3 CIVs. Anterior region (number of receptors/side of the cercaria): AI
= 1 AI V, 1 AI L, 1 AI D; AII = 1-3 AII V, 1-2 AII L, 4 AII D; AIII = 4 AIII V, 3 AIII L, 3 AIII D.
Posterior region (number of receptors/side of the cercaria): PI = 1 PI V, 1-3 PI L; PII = 1 PII V, 1-4 PII L, 1 PII D; PIII = 1 PIII V, 1-4 PIII L, 1 PIII D.
Ventral sucker region (total number of receptors): S = 3 SI, 6 SII, 6 SIII.
Caudal region (number of receptors/side of the cercaria): U = 4 U DL.
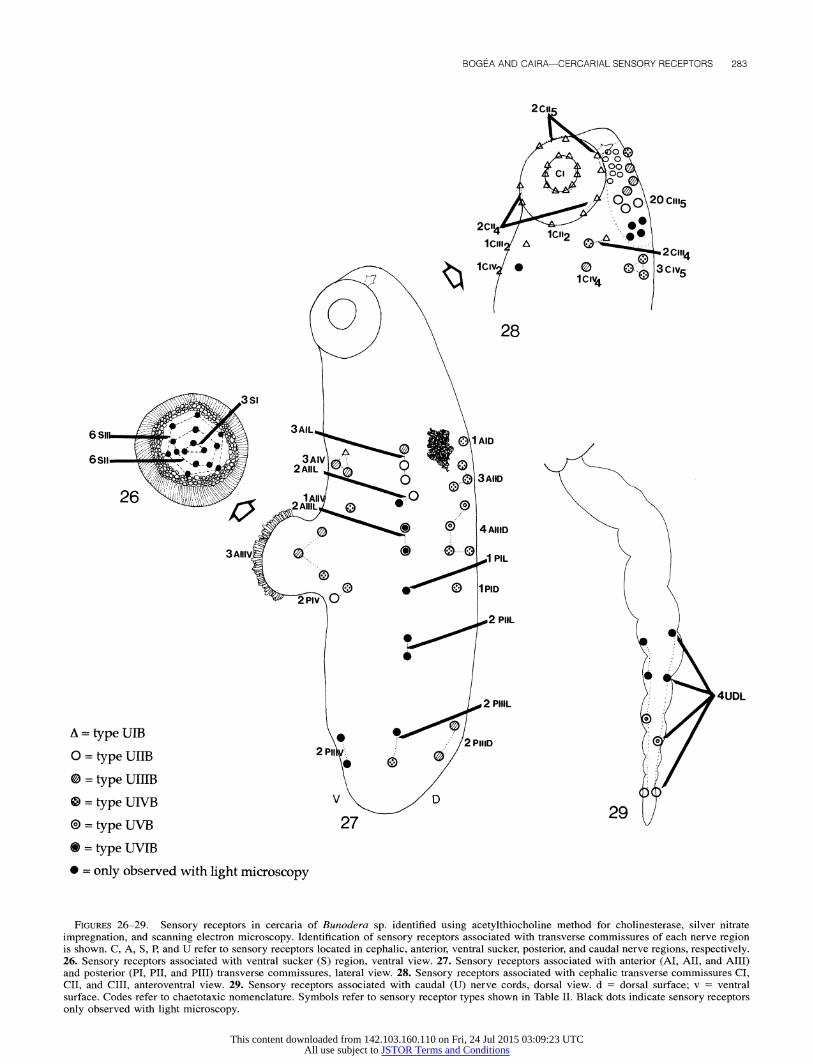
SEM also confirmed the chaetotaxic pattern detected with silver nitrate impregnation in the cercaria of Bunodera sp. The
pattern is illustrated in Figures 26-29, and the chaetotaxic no- menclature is summarized as follows:
Cephalic region (number of receptors/side of the cercaria): CI = 2 CI2, 2 CI4, 1 C5; CII = 1 CII2, 2 CII4, 2 CII5; CIII = 1
CIII2, 2 CIII4, 21-24 CIII5; CIV = 1 CIV2, 1 CIV4, 3 CIVs. Anterior region (number of receptors/side of the cercaria): AI
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions
278 THE JOURNAL OF PARASITOLOGY, VOL. 87, NO. 2, APRIL 2001
Ar rAPr AV 0 ~C1) zPc ra
AlJ APBc
WOW 00~ 3; 't~wow
Ira
Aa!,J "W ~I
%
c f. t 9 -i I++IN
~?t? ~dlc~00
;oar
SW" 40; 4;
FIGURES 7-13. Scanning electron micrographs of types of sensory receptors in cercaria of Crepidostomum sp. 7. Type UIC. 8. Type UIIC. 9. Type UIIIC. 10. Type UIVC. 11. Type UVC. 12. Type UVIC. 13. Type BiC. Bars = 2 p~m. Codes (UIC-BiC) refer to sensory receptor types shown in Table I. Note variations in cilium number, presence or absence of tegumentary collar, cilium length and tegumentary collar length.
= 3 AI V, 2 or 3 AI L, 1 AI D; AII = 1 AII V, 2 AII L, 3
All D; AIII = 3 AIII V, 1 or 2 AIII L, 4 AIII D. Posterior region (number of receptors/side of the cercaria): PI
= 2 PI V, 1 PI L, 1 PI D; PII = 2 PII L; PIII = 2 PIII V, 2 PIII L, 2 PIII D.
Ventral sucker region (total number of receptors): S = 3 SI, 6 SII, 6 SIII.
Caudal region (number of receptors/side of the cercaria): U = 4 U DL.
The distribution of the 7 types of sensory receptors for the cephalic, anterior, acetabular, posterior, and caudal regions of
Crepidostomum sp. is summarized in Figures 22-25. Type UIC, with a high tegumentary collar and a short cilium, was observed
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions
BOGEA AND CAIRA-CERCARIAL SENSORY RECEPTORS 279
TABLE II. Types of sensory receptors in cercaria of Bunodera sp. as seen with scanning electron microscopy. Mean ? SD of measurements (in micrometers) is followed by number of sensory receptors measured.
Cilia Tegumentary collar
Type* Number Length Width Length Width
UIB 1 0.96 + 0.29; 15 0.20 + 0.06; 15 030 + 0.08; 15 0.59 + 0.11; 15 UIIB 1 0.71 + 0.31; 04 0.23 + 0.07; 04 0.17 + 0.00; 04 0.61 + 0.07; 04
UIIIB 1 1.90 + 0.79; 15 0.19 ? 0.05; 15 0.18 + 0.02; 15 0.58 + 0.11; 15 UIVB 1 4.92 + 1.59; 17 0.18 + 0.01; 17 0.19 + 0.05; 17 0.57 + 0.17; 17 UVB 1 3.78 + 0.82; 06 0.17 + 0.00; 06 0.20 + 0.06; 06 0.76 + 0.06; 06 UVIB 2 2.40 + 0.34; 03 0.18 + 0.01; 03 0.15 + 0.05; 03 0.56 + 0.10; 03
* UIB = sensory receptor with a moderately low tegumentary collar and a short cilium; UIIB = sensory receptor with a low tegumentary collar and a short cilium; UIIIB = sensory receptor with a low tegumentary collar and a moderately long cilium; UIVB = sensory receptor with a low tegumentary collar and a long cilium; UVB = sensory receptor with a low tegumentary collar located on a domelike base and a long cilium; UVIB = sensory receptor with a low tegumentary collar located on a domelike base and 2 moderately long cilia.
*Awl or lk t"-j3
.14 1
A- Alla:
k4, ~L XCL*S' .~X IP
vor"
4i A?:
Ilrr doll. 4ow- 7~"'C" : "-;
,W.4; *at4?4;, .dPs ~ -?c *3""`'S l~h I~C~\i-f~~:Wf
~'::~ Y?MW
r, ; ? -~18'
FIGURES 14-18. Scanning electron micrographs of types of sensory receptors in cercaria of Bunodera sp. 14. Type UIB (arrow). 15. Type UIIB. 16. Types UIIIB (small arrow) and UIVB. 17. Type UVB (arrow). 18. Type UVIB. Bars = 2 Vpm. Codes (UIB-UVIB) refer to sensory receptor types shown in Table II. Note variations in cilium number, presence or absence of a domelike base, cilium length, and tegumentary collar length.
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions
280 THE JOURNAL OF PARASITOLOGY, VOL. 87, NO. 2, APRIL 2001
C
vc v
.... ?
i4-
Iv
19",: --Ice
4w.. ..
TC m vai- d?? - i
"100
FIGURES 19, 20. Transmission electron micrographs of sensory receptors in cercaria of Crepidostomum sp. 19. Type UIC. Inset: central pair of microtubules. Bars = 0.5 ?pm. 20. Type UIVC. Bar = 0.5 ?pm. BB = basal body; BP = basal plate; C = cilium; COL = thickening of nerve collar; JC = extracellular junctional complex; MT = microtubules; MV = microvilli; TC = tegumentary collar; TM = tegumentary membrane; R = ciliary rootlet; V = vesicles. Codes (UIC, UIVC) refer to sensory receptor types shown in Table I.
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions

BOGEA AND CAIRA-CERCARIAL SENSORY RECEPTORS 281
~41
_r.!
21
ANN'
FIGURE 21. Transmission electron micrograph of a sensory receptor (type UIB) in cercaria of Bunodera sp. Bar = 0.6 Rm. BB = basal body; BP = basal plate; C = cilium; COL = thickening of nerve collar; JC = extracellular junctional complex; MT = microtubules; TC = teg- umentary collar; TM = tegumentary membrane; V = vesicles. Code (UIB) refers to sensory receptor type shown in Table II.
on both ventral (around the oral opening) and dorsal surfaces of the cephalic region (Fig. 24). Types UIIC and UIIIC, with low tegumentary collars and either a short or a moderately long cilium, were widespread throughout the tegument and on the ventral and lateral surfaces of the cercarial body, respectively (Fig. 23). Type UIVC, with a low tegumentary collar and a
long cilium, occurred mainly on the dorsal surfaces of the ce-
phalic region and cercarial body (Figs. 23, 24). Type BiC, with a low tegumentary collar and 2 long cilia, was restricted to the ventral surface of the cephalic region (Fig. 24), whereas those with no tegumentary collars and a short or a moderately long cilium were found on the tail (Fig. 25).
The distribution of the 6 types of sensory receptors for the
cephalic, anterior, acetabular, posterior, and caudal regions of Bunodera sp. is summarized in Figures 26-29. Type UIB, with a moderately low tegumentary collar and a short cilium, was observed mainly around the oral opening (Fig. 28). Types UIIB and UIIIB, with a low tegumentary collar and either a short or a moderately long cilium, were widespread throughout the teg- ument (Figs. 27-29). Type UIVB, with a low tegumentary col- lar and a long cilium, occurred mainly on the dorsal surfaces of the cephalic region and cercarial body (Figs. 27, 28). Types UVB and UVIB, with a low tegumentary collar located on a domelike base with 1 long or 2 moderately long cilia, were found on the dorsal surface of the cercarial body (Fig. 27), on the tail (Fig. 29), and on the lateral surface of the cercarial body (Fig. 27), respectively.
DISCUSSION
SEM observations suggest that there are 7 and 6 basic types of sensory receptors in the cercaria of Crepidostomum sp. (Ta- ble I; Figs. 7-13) and Bunodera sp. (Table II; Figs. 14-18), respectively. These types are distinguished by the lack of a
tegumentary collar, possession of a domelike base, 1 or 2 cilia that are short, moderately long, or long, and a low, a moderately low, or a high tegumentary collar.
Some of the types of sensory receptors distinguished in the cercariae of Crepidostomum sp. and Bunodera sp. were site
specific. For example, receptors with a moderately low or high tegumentary collar and a short cilium were most commonly found on the cephalic region (Figs. 24, 28). Sensory receptors with a low tegumentary collar and a short cilium were wide-
spread throughout the tegument, whereas those with a low teg- umentary collar and a long cilium predominated on the dorsal surface (Figs. 23, 27). Receptors with a low tegumentary collar and a moderately long cilium were restricted to the ventral and lateral surfaces of the cercarial body of Crepidostomum sp. (Fig. 23) but were widespread throughout the tegument of Bun- odera sp. (Fig. 27). Sensory receptors with no tegumentary col- lar only occurred on the tail of Crepidostomum sp. (Fig. 25). Types with a low tegumentary collar located on a domelike base and 1 long cilium were observed on the tail and on the dorsal surface of the body of Bunodera sp. (Figs. 27, 29), whereas those with 2 moderately long cilia occurred on the lateral surface of the body in this species (Fig. 27). Site-specific patterns including receptors with no tegumentary collar or a domelike base restricted to the cercarial tail, long uniciliated
receptors concentrated on the dorsal surface, and short unici- liated receptors widespread throughout the tegument are ob- served in other free-swimming cercariae (Ruiz, 1989; Niewia- domska and Czubaj, 1996; Bog6a, 1998; Kock and Bockeler, 1998).
The internal ultrastructure of 3 types of sensory receptors was studied in the cercariae of Crepidostomum sp. and Bun- odera sp. The receptors shared features such as an unsheathed cilium, a basal plate, and a basal body. They also exhibited a sensory bulb attached to the tegument by septate junctional complexes and thickened nerve collars. These observations suggest that such types most likely respond to the same sensory modality. They also indicate that types UIC and UIB are similar. The internal ultrastructure of types UIC, UIVC, and UIB is comparable to that of sensory organs found in other cercariae,
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions
282 THE JOURNAL OF PARASITOLOGY, VOL. 87, NO. 2, APRIL 2001
2CII
2CI o o
1 CI
2 C0Il= p -A DCIV5
1CIV2 2 2 1AIL 1CIV4
=tpAID 24
6SIII. 1AIV AI
2AIIL
6SII 3 AI0l. ..
O S IL4
AIID
3AI11L
4AI11V
22 o .. o
,. 3AI11D
1PIV 0 3 PIIL
3 PIlL
1PIIV\ 1
...PIID
A = type UIC 4UDL
@ = type UIVC V D 0 = type
UVC235 G = type UVIC
( = type BiC * = only observed with light microscopy
FIGURES 22-25. Sensory receptors in cercaria of Crepidostomum sp. identified using acetylthiocholine method for cholinesterase, silver nitrate impregnation, and scanning electron microscopy. Identification of sensory receptors associated with transverse commissures of each nerve region is shown. C, A, S, P, and U refer to sensory receptors located in cephalic, anterior, ventral sucker, posterior, and caudal nerve regions, respectively. 22. Sensory receptors associated with ventral sucker (S) region, ventral view. 23. Sensory receptors associated with anterior (AI, AII, and AIII) and posterior (PI, PII, and PIII) transverse commissures, lateral view. 24. Sensory receptors associated with cephalic transverse commissures CI, CII, and CIII, anteroventral view. 25. Sensory receptors associated with caudal (U) nerve cords, dorsal view. d = dorsal surface; v = ventral surface. Codes refer to chaetotaxic nomenclature. Symbols refer to sensory receptor types shown in Table I. Black dots indicate sensory receptors only observed with light microscopy.
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions
BOGEA AND CAIRA-CERCARIAL SENSORY RECEPTORS 283
2 cuI
00
OO20 c'm5
1O1i 2 . 1CI A @0
28 28
V" 3si
6 sil 3 AL 1 AID
6sII 3AIV O
.O 3 AIID
26 1AIIO0 2AI1L
0 4 AllD
3AIIIV 0 0 PIL
1PID ?20 IiiD
2 PIIL
2 PHIL 4UDL
S= type UIB 2 PIIID o
O = type U11B
O = type UIIIB S = type UIVB V 2lD
@= type UVB 27 * = type UVIB
* = only observed with light microscopy
FIGURES 26-29. Sensory receptors in cercaria of Bunodera sp. identified using acetylthiocholine method for cholinesterase, silver nitrate impregnation, and scanning electron microscopy. Identification of sensory receptors associated with transverse commissures of each nerve region is shown. C, A, S, P, and U refer to sensory receptors located in cephalic, anterior, ventral sucker, posterior, and caudal nerve regions, respectively. 26. Sensory receptors associated with ventral sucker (S) region, ventral view. 27. Sensory receptors associated with anterior (AI, AII, and AIII) and posterior (PI, PII, and PIII) transverse commissures, lateral view. 28. Sensory receptors associated with cephalic transverse commissures CI, CII, and CIII, anteroventral view. 29. Sensory receptors associated with caudal (U) nerve cords, dorsal view. d = dorsal surface; v = ventral surface. Codes refer to chaetotaxic nomenclature. Symbols refer to sensory receptor types shown in Table II. Black dots indicate sensory receptors only observed with light microscopy.
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions

284 THE JOURNAL OF PARASITOLOGY, VOL. 87, NO. 2, APRIL 2001
including those of Echinostoma revolutum (Fr6lich, 1802) and Diplostomum pseudospathaceum Niewiadomska, 1984, as ob- served by Zdarska (1992) and Niewiadomska and Czubaj (1996), respectively. In both species, these sensory receptors were presumed to be mechanoreceptors, although the hypoth- esis that they may also serve the double function of mechano- chemoreception cannot be discarded. TEM investigations on the remaining types of sensory receptors detected in the cercariae of Crepidostomum sp. and Bunodera sp. are needed to test this hypothesis.
Comparative studies on sensory receptor ultrastructure are useful for further elucidating the sensory modalities with which the receptors are associated. The internal ultrastructure of types UIC, UIVC, and UIB indicates that the transducer mechanism of these types may be similar to those described in sensory organs of other invertebrates (Lyons, 1969) and vertebrates (Thurm, 1968). Transduction would be triggered by an input stimulus applied to the cilium. Such displacement would bend the basal body and would result in local changes in membrane potential, in neuronal synthesis, and in secretion of extracellular signaling molecules or chemical messengers responsible for co- ordinating and controlling the response of the appropriate ef- fector organ (Halton et al., 1997). These molecules would be sequestered in neuronal vesicles. The development of novel neurophysiological recording methods for cercarial sensory re- ceptors would be particularly relevant to verify this assumption.
Lyons (1969) suggested that the input stimulus for sensory receptors such as types UIC, UIVC, and UIB would probably be mechanical, because the cilium found in platyhelminth sen- sory receptors usually has a central pair of microtubules char- acteristic of motile cilia of invertebrates and vertebrates (Thurm, 1968). The presence of thickened nerve collars around the sensory bulb supports this idea for platyhelminths (Lyons, 1969). These collars might act as membrane reinforcements that maintain the sensory receptors in place in the tegument. These reinforcements, possibly keratinous, indicate the levels of me- chanical tension to which these receptors may be subjected. Consequently, types UIC, UIVC, and UIB, all of which possess nerve collars, probably belong in the class of mechanorecep- tors.
Ultrastructural characters concerning tegumentary collar and cilium lengths provide additional information on the mechan- osensitivity of cercarial sensory receptors. For example, the moderately low and high tegumentary collars of types UIB and UIC would seem to restrict cilium movement more than those of types UIIC, UIIIC, UIIB, and UIIIB. In addition, the low tegumentary collars of types UIVC, BiC, UIVB, and UVB may allow long cilia to respond more promptly to low mechanical stimuli such as those elicited by water currents. Long uniciliated sensory receptors that tended to occur on the dorsal surface of the cercarial body in both species suggest that they may play a role in detecting water currents. Short uniciliated sensory re- ceptors were widespread throughout the tegument, indicating that they may be associated with touch, which is crucial during host penetration.
The chaetotaxy of species of Crepidostomum and Bunodera has been previously studied. Richard (1971) and Bayssade-Du- four (1979) investigated the chaetotaxy of Crepidostomum me- toecus (Braun, 1900) Braun, 1900 and Crepidostomum farionis (Miiller, 1784) Nicoll, 1909 from Europe. Although the chae-
totaxic maps were different for each species, these authors rec- ognized the arrangements of CI (2 CI V, 2 CI L, 1 CI D) and CII (1 CII V1, 1 CII V2, 1 CII L1, 2 CII L2), of anterior circles AI D (1 AI D) and A V (1 AI V, 1 AII V, 1 AIII V), of midbody circle M V (4 M V), of ventral sucker circles S (3 SI, 6 SII, 6 SIII), and of caudal circle U (4 U DL) as diagnostic characters for Crepidostomum. The chaetotaxic map of North American Crepidostomum sp. studied here is similar, but not identical, to the maps of C. metoecus and C. farionis previously obtained (Figs. 22-25). The present cercaria had numbers of sensory receptors on circles CI and CII (1 CI2, 2 CI4, 1 CI5, and 1 CII2, 2 CII4, 2 CII5, respectively) similar to those observed for C. farionis and C. metoecus. However, Crepidostomum sp. had more receptors on the ventral surface of the anterior region (1 AI V, 1-3 All V, 4 AIII V) than has been reported in either species. Based on the work of Bayssade-Dufour et al. (1993), it is proposed that Crepidostomum be characterized by chae- totaxic arrangements 1-2 CI2 (previously identified as 2 CI V), 2 CI4 (previously identified as 2 CI L), 1 CI5 (previously iden- tified as 1 CI D), 1 CII2 (previously identified as 1 CII VI), 2
CII4 (previously identified as 1 CII V2 and 1 CII LI), 2 CII5
(previously identified as 2 CII L2), 1 AI D, 1 AI V, 1-3 AII V (previously identified as 1 All V and 1 AIII V), 4 AIII V (pre- viously identified as 4 M V), 3 SI, 6 SII, 6 SIII, and 4 U DL.
In Crepidostomum sp., the distribution of sensory receptors on the cercarial tegument was verified with SEM. However, it differed slightly from that of the chaetotaxic map previously obtained with silver nitrate impregnation. Specifically, 2 pairs of ventral sensory receptors (1 on the cephalic region and 1 on the posterior region) were detected with SEM but not with sil- ver nitrate. The distribution of sensory receptors as revealed by SEM, however, was consistent with the basic chaetotaxic map. The distribution of sensory receptors as revealed with silver nitrate also differed from that obtained with SEM in the cer- cariae of Opisthioglyphe ranae (Fr6lich, 1791), Trichobilharzia franki Muiller and Kimmig, 1994, and Trichobilharzia ocellata (La Valette, 1855) (Busta and Nasincova, 1988; Kock and B6ckeler, 1998).
Cannon (1971) established chaetotaxic maps for the cercariae of Bunodera sacculata Van Cleave and Mueller, 1932 and Bun- odera luciopercae (Muller, 1776) Luehe, 1909. An objective interpretation of the patterns obtained, however, remains ten- tative because the chaetotaxic nomenclature was not determined by Cannon. Chaetotaxic arrangements in cephalic (2 CI2, 2 CI4, 1 C5), anterior (3 AI V, 1 All V, 2-4 AIII V, 1 AI D), and ventral sucker (4 SI, 6 SIII) regions were identical in both spe- cies. The chaetotaxy of Bunodera sp. is similar to those of B. sacculata and B. luciopercae in the arrangements in cephalic (2 CI2, 2 CI4, 1 C5), anterior (3 AI V, 2-3 AI L, 1 AI D, 1 AII
V, 2 AII L, 3 AII D, 3 AIII V, 1-2 AIII L, 4 AIII D), posterior (2 PI V, 1 PI L, 1 PI D, 2 PII L, 2 PIII V, 2 PIII L, 2 PIII D), and ventral sucker (3 SI, 6 SII, 6 SIII) regions (Figs. 26-28). Additional studies are needed to determine the chaetotaxy of the genus Bunodera.
Phylogenetic characters have been demonstrated in the cer- carial chaetotaxy of Allocreadiidae. Within this family, Bays- sade-Dufour (1979) identified a pair of mediodorsal sensory receptors found at the eyespot level (1 AI D) as the diagnostic characters for the family. These receptors were reported in the cercariae of C. metoecus and C. farionis by Richard (1971), of
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions

BOGEA AND CAIRA-CERCARIAL SENSORY RECEPTORS 285
B. sacculata and B. luciopercae by Cannon (1971), and of Al- locreadium ictaluri Pearse, 1924 by Cable and Peters (1986). The presence of these receptors in Crepidostomum sp. and Bun- odera sp. further confirms their identity as allocreadiid cercar- iae.
The arrangements of cholinergic nerve networks are intro- duced here for the first time for the cercariae of Allocreadiidae. Cholinergic components were demonstrated in the cerebral gan- glion, nerve cords, and transverse commissures of Crepidosto- mum sp. and Bunodera sp. Fine connections branching from the ventral nerve cords to the ventral sucker suggest that such cords innervate the sensory receptors found in this region. Five nerve regions were clearly recognized. Shishov (1991) ob- served similar arrangements in the cholinergic nerve networks of the adult stage of Bunodera luciopercae. The present results indicate that cercarial nerve networks may closely resemble those of adult stages in this family.
The identification of cephalic and anterior nerve regions in Crepidostomum sp. and Bunodera sp. deserves attention, main- ly because the identification of these regions is related to the position of the cerebral ganglion, a feature that may vary among species. In the cercaria of the diplostomid Diplostomum pseu- dospathaceum, for instance, the cerebral ganglion is located be- low the pharynx, resulting in 4 cephalic rings (Niewiadomska, 1994). In the cercariae of Crepidostomum sp. and Bunodera sp., the cerebral ganglion was located at the level of the phar- ynx (Figs. 1, 2, 4, 5). Consequently, these allocreadiids exhib- ited 3 cephalic rings, whereas the number of transverse com- missures remained equal.
Recent investigators have demonstrated that cercarial ner- vous systems are far more complex than originally presumed. The vast array of neuronal substances detected in this system indicates that there are serotonergic, aminergic, and peptidergic components (Halton et al., 1997). Peptidergic and cholinergic elements seem to co-occur in the neurons, whereas serotonergic components are located in different sets of neurons. These re- sults prompted us to send cercariae to Dr. David Halton, who conducted a preliminary analysis in the nervous system of the cercaria of Bunodera sp. This analysis indicated that serotoner- gic and peptidergic components occur in this species. This find- ing supports the idea that cercarial nervous and sensory systems may be complex enough to enable these larvae to sense and respond to specific stimuli to complete their development in a timely manner (Bog6a et al., 1990, 1996).
The results of the present study indicate that the proposed standard procedure using a combination of techniques enhances the study of cercarial sensory systems. The use of neurocyto- chemical methods permits distinction among different nerve re- gions and the naming of cercarial sensory receptors after their actual position relative to the nerve network (Bayssade-Dufour et al., 1989; Niewiadomska, 1994). Increasing the number of cercariae of a species processed with silver nitrate and using the modal class to determine the chaetotaxic pattern helps to minimize the subjective identification of dorsal, ventral, and lateral sensory receptors and the effect of intraspecific vari- ability (Short and Cartrett, 1973; Bog6a, 1999). In addition, SEM and TEM allow the verification of staining of pores and tegumentary elevations, as suggested by Chapman and Wilson (1970). The proposed standard procedure, therefore, is a useful approach to further investigate cercarial sensory systems. It
may not only allow a better understanding of the functional roles of cercarial sensory receptors but may also reveal novel features of phylogenetic utility for the taxa involved.
ACKNOWLEDGMENTS
We thank S. Curran, T. Ferrara, S. Goshroy, C. Healy, S. Hains, K. Jensen, C. Latieri, M. Rush, R. Rodrigues, and S. Zahner for collecting fingernail clams. We thank H. Keller and M. Mercer for granting permission for the collections to be made in Pachaug Campground and Marina. G. Mackie kindly identified the clams. D. Scott provided us with the protocol for preparing whole mounts of cercariae. We appreciate the tech- nical assistance and advice of K. Tiekotter, J. Romanow, M. Cantino, C. Henry, E. Jokinen, and L. Khairallah. We are grate- ful to C. Schaefer for critically reviewing an earlier version of the manuscript. We thank D. Halton for conducting a prelimi- nary analysis of serotonergic and peptidergic systems in the cercaria of Bunodera sp. We thank 2 anonymous reviewers for helpful comments. All collections were conducted according to conditions set forth in collecting permit 203 issued to J.N.C. by the Fisheries Division of the Connecticut State Department of Environmental Protection. This work was supported by doc- toral scholarships from the Conselho Nacional de Desenvolvi- mento Cientifico e Tecnol6gico (200142/94-1), Brazil, and the University of Connecticut to T.B.
LITERATURE CITED ABDUL-SALAM, J., AND B. N. S. SREELATHA. 1998. Studies on cercariae
from Kuwait Bay. VIII. Description and surface topography of Cer- caria kuwaitae VIII sp. n. (Digenea: Opecoelidae). Parasitology International 47: 87-94.
BAYSSADE-DUFOUR, C. 1979. L'appareil s6nsoriel des cercaries et la sys- t6matique des tr6matodes digen6tiques. M6moires du Museum Na- tional d'Histoire Naturelle Paris S6rie A 113: 1-81.
, J.-L. ALBARET, B. GRABDA-KAZUBSKA, AND A. G. CHABAUD. 1989. Nomenclature propos6e pour la ch6totaxie des cercaires de Plagiorchiata. Annales de Parasitologie Humaine et Compar6e 64: 426-432.
, J.-P HUGOT, AND J.-L. ALBARET. 1993. Analyse ph6n6tique des Microphalloidea (Trematoda) d'apris la ch6totaxie des cercaires. Systematic Parasitology 25: 1-24.
BETTENDORF, H. 1897. Uber Musculatur und Sinneszellen der Trema- toden. Zoologische Jahrburcher Anatomie 10: 307-358.
BLAIR, D. 1977. A key to cercariae of British strigeoids (Digenea) for which the life-cycles are known, and notes on the characters used. Journal of Helminthology 51: 155-166.
BOGEA, T 1998. Distribution and morphology of sensory papillae in the cercaria of Allassogonoporus marginalis (Digenea: Lecithodendri- idae). Parasitology International 47: 120.
. 1999. Ultrastructure and chaetotaxy of cercarial sensory recep- tors: A comparative analysis of four species of Digenea (Platyhel- minthes). Ph.D. Dissertation. University of Connecticut, Storrs, Connecticut, 253 p.
, T C. FAVRE, L. ROTENBERG, H. S. SILVA, AND O. S. PIERI. 1990. Chronobiological aspects in the emission of Schistosoma mansoni cercariae from Biomphalaria spp. under outdoor conditions: Mono and plurimiracidial infections. Journal of Interdisciplinary Cycle Research 21: 171-173.
, - , , AND . 1996. Circadian pattern of cercarial emergence in Schistosoma mansoni (Platyhelminthes: Digenea) from isolated Biomphalaria glabrata. Chronobiology In- ternational 13: 93-101.
BUSTA, J., AND V. NASINCOVA. 1988. Scanning electron microscopy of the egg, sporocyst, cercaria, metacercaria, and adult of Opisthiog- lyphe ranae (Fr6lich, 1791) (Trematoda: Plagiorchiidae). Folia Par- asitologica 35: 97-103.
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions

286 THE JOURNAL OF PARASITOLOGY, VOL. 87, NO. 2, APRIL 2001
CABLE, R. M., AND L. E. PETERS. 1986. The cercariae of Allocreadium ictaluri Pearse (Digenea: Allocreadiidae). Journal of Parasitology 72: 369-371.
CAIRA, J. N. 1989. A revision of the North American papillose Allo- creadiidae with independent cladistic analyses of larval and adult forms. Bulletin of the University of Nebraska State Museum 11: 1-58.
CANNON, L. R. G. 1971. The life cycles of Bunodera sacculata and B. luciopercae (Trematoda: Allocreadiidae) from Algonquin Park. Ca- nadian Journal of Zoology 49: 1417-1429.
CHAPMAN, H. D., AND R. A. WILSON. 1970. The distribution and fine structure of the integumentary receptors of the cercariae of Hi- masthla secunda (Nicoll). Parasitology 61: 219-227.
COMBES, C., C. BAYSSADE-DUFOUR, AND J. CASSONE. 1976. Sur
l'impregnation et le montage des cercaires pour l'6tude ch6tota- xique. Annales de Parasitologie Humaine et Compared 51: 399- 400.
GRABDA-KAZUBSKA, B., AND V. KISELIENE. 1989. Chaetotaxy and excre-
tory system of the cercaria of Echinoparyphium recurvatum (Lin- stow, 1873) (Digenea, Echinostomatidae). Acta Parasitologica Po- lonica 34: 325-335.
HALTON, D. W., A. G. MAULE, AND C. SHAW. 1997. Trematode neuro-
biology. In Advances in trematode biology, B. Fried and T K. Graczyk (eds.). CRC Press, Boca Raton, Florida, p. 345-382.
HOPKINS, J. 1934. The papillose Allocreadiidae. Illinois Biological Monographs 13: 1-79.
KOCK, S., AND W. BOCKELER. 1998. Observations on cercarial chaeto-
taxy as a means for the identification of European species of Trichobilharzia Skrjabin and Zakharow, 1920 (Digenea: Schisto- somatidae). Systematic Parasitology 43: 159-166.
KOIE, M. 1981. On the morphology and life-history of Podocotyle re- flexa (Creplin, 1825) Odhner, 1905, and a comparison of its de- velopmental stages with those of P. atomon (Rudolphi, 1802) Odhner, 1905 (Trematoda, Opecoelidae). Ophelia 20: 17-43.
KREJCI, K. G., AND B. FRIED. 1994. Light and scanning electron micro-
scopic observations of the eggs, daughter rediae, cercariae, and encysted metacercariae of Echinostoma trivolvis and E. caproni. Parasitology Research 80: 42-47.
LYONS, K. M. 1969. Sense organs of monogean skin parasites ending in a typical cilium. Parasitology 59: 611-623.
MATRICON-GONDRAN, M. 1971. Etude ultrastructurale des rec6pteurs sensoriels t6gumentaires de quelques tr6matodes Digen6tiques lar- vaires. Zeitschrift ftir Parasitenkunde 35: 318-333.
NIEWIADOMSKA, K. 1994. Adjusting of the pattern of chaetotaxy in cer- cariae of Diplostomum Nordmann, 1832 (Digenea) to the structure of their nervous system. Acta Parasitologica 39: 187-191.
--, AND A. CZUBAJ. 1996. Sensory endings in Diplostomum pseu- dospathaceum Niewiadomska, 1984 cercariae (Digenea, Diplos- tomidae). Acta Parasitologica 41: 20-25.
OLSON, B. J. 1985. Metazoan parasites of yellow perch Percaflavescens (Mitchill) from Pachaug Pond, Connecticut. M.S. Thesis. Univer- sity of Connecticut, Storrs, Connecticut, 69 p.
PRESNELL, J. K., AND M. P. SCHREIBMAN. 1997. Humanson's animal tis- sue techniques, 5th ed. Johns Hopkins University Press, Baltimore, Maryland, 572 p.
RICHARD, J. 1971. La ch6totaxie des cercaires: valeur systematique et phyl6tique. M6moires du Museum National d'Histoire Naturelle Paris S6rie A 67: 1-179.
RuIz, L. J. R. 1989. Ultrastructura de la superficie del cuerpo de la cercaria de Paragonimus mexicanus. Anales del Instituto de Biol- ogia de la Universidade Nacional Aut6noma de Mexico Serie Zool- ogia 60: 299-310.
SHORT, R. B., AND M. L. CARTRETT. 1973. Argentophilic 'receptors' of Schistosoma mansoni cercariae. Journal of Parasitology 59: 1041- 1059.
SHISHOV, B. A. 1991. Aminergic elements in the nervous system of helminths. Studies in Neurosciences 13: 113-152.
STATE BOARD OF FISHERIES AND GAME. 1959. A fishery survey of the lakes and ponds of Connecticut. State Board of Fisheries and Game Publications, Hartford, Connecticut, 232 p.
SWEETING, R. 1976. An experimental demonstration of the life cycle of a Diplostomulum from Lampetra fluviatilis Linnaeus, 1758. Zeit- schrift fUir Parasitenkunde 49: 233-242.
THOMPSON, S. W. 1966. Selected histochemical and histopathological methods. Charles C Thomas, Springfield, Illinois, 1,639 p.
THURM, U. 1968. Steps in the transducer process of mechanoreceptors. Symposia of the Zoological Society of London 23: 199-216.
ZDARSKA, Z. 1992. Transmission electron microscopy of sensory recep- tors of Echinostoma revolutum (Froelich, 1802) cercaria (Digenea: Echinostomatidae). Parasitology Research 78: 598-606.
This content downloaded from 142.103.160.110 on Fri, 24 Jul 2015 03:09:23 UTCAll use subject to JSTOR Terms and Conditions




















