
Evaluation of Local Banana Cultivars under Coconut Shade in ...

www.elsevier.com/locate/agee
Available online at www.sciencedirect.com
Agriculture, Ecosystems and Environment 124 (2008) 160–172
Tree species diversity and vegetation structure in shade
coffee farms in Veracruz, Mexico
Ana M. Lopez-Gomez, Guadalupe Williams-Linera *, Robert H. Manson
Instituto de Ecologıa, A.C., km 2.5 carretera Antigua a Coatepec 351, Xalapa, Veracruz 91070, Mexico
Received 15 February 2007; received in revised form 21 August 2007; accepted 17 September 2007
Available online 26 October 2007
Abstract
Using 15 shade coffee farms and two forest reserves located in central Veracruz, Mexico, we evaluated how tree vegetation structure and
richness changed as a function of management type and compared to that remnant forest fragments. Coffee farms were classified as shade
monocultures (SMs), simple polycultures (SPs), or diverse polycultures (DPs). A total of 124 sampling units representing 15.19 ha were
randomly located in farms with the number of sampling units varying as a function of farm area. Twenty additional plots were sampled in two
nearby cloud forest fragments. Forest vegetation structure was higher than in farms, except for mean canopy height that was similar to that in
SP and DP farms. Within farms, tree density was generally higher in SM, whereas basal area and both mean and maximum height were higher
in SP and DP sites. We recorded 2833 individuals representing 107 tree species �5 cm dbh in coffee farms, including 24 non-native species,
and 83 native species (33 primary, and 50 secondary tree species). Patterns of richness followed the expected pattern with 11 � 1.4 (S.E.) tree
species in SM farms, 14 � 1.8 in SP farms, 29 � 2.3 in DP farms, and 38 � 16 in forest control sites; however, rarefaction strongly suggests
that DP sites are actually richer in species than the forests studied. The proportion of native tree species in each coffee management type was
consistently elevated (78%). Since the majority of species (71%) were rare and found only in one or two farms, complementarity among types
of coffee management (68–77%), and coffee managements and forest (90–92%) was very high. SP and DP farms had the highest proportions
of animal-dispersed species and were similar to forest. Species richness was positively correlated with tree density for DP, and to basal area for
SM farms. Our results suggest that shade diversity is actively managed by coffee farmers and that all three types of coffee management studied
may have an important role to play in the conservation of regional biodiversity. Considering factors such as complementarity, landscape
heterogeneity, functional diversity, and the rigor of vegetation surveys may also help improve the validity, and thus the impact, of coffee
certification programs designed with the goal of conserving tropical montane biodiversity.
# 2007 Elsevier B.V. All rights reserved.
Keywords: Coffee agroecosystems; Tree diversity; Vegetation structure; Veracruz
1. Introduction
A number of studies have argued that the similarity of the
vegetation structure in traditional shade coffee plantations to
that in native forests remnants makes these agroecosystems
an important component of strategies for conserving tropical
montane biodiversity (Perfecto et al., 1996; Moguel and
Toledo, 1999). However, a recent debate regarding this
* Corresponding author. Tel.: +52 228 8421838/8421800x4206;
fax: +52 228 8421800x4222.
E-mail address: [email protected]
(G. Williams-Linera).
0167-8809/$ – see front matter # 2007 Elsevier B.V. All rights reserved.
doi:10.1016/j.agee.2007.09.008
assertion (Philpott and Dietsch, 2003; Rappole et al.,
2003a,b; O’Brien and Kinnaird, 2003, 2004; Dietsch et al.,
2004) suggests that more research is need to quantify the net
value of such farms in conserving forest structure
(biodiversity) and function (ecosystem services) and how
their promotion through mechanisms such as certification
may affect remnants of forest in the same region (Perfecto
et al., 2003; Lambin et al., 2003; Steffan-Dewenter et al.,
2007). While a lack of studies on important ecological
processes (Beer et al., 1998; Komar, 2006) and the economic
viability of different coffee management strategies (Gobbi,
2000; Gordon et al., 2007) are undoubtedly central in this
debate, the strong bias in the taxa studied in these

A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172 161
agroecosystems is also an important contributing factor
(Moguel and Toledo, 1999). In particular, there has been
much more focus on animal versus plant biodiversity and
within animal taxa birds and insects appear over-represented
in the literature. A variety of studies have established the
value of shaded coffee plantations as habitat for monkeys
(Williams-Guillen et al., 2006), birds (Greenberg et al.,
1997; Tejeda-Cruz and Sutherland, 2004; Komar, 2006),
bats, frogs and dung beetles (Pineda et al., 2005), and
various other groups of insects (Ricketts et al., 2001;
Armbrecht et al., 2004; Arellano et al., 2005). In contrast,
while some groups of plants have begun to be studied in
detail such as epiphytes (Solis-Montero et al., 2005), herbs,
and shrubs (Mayfield et al., 2005; Potvin et al., 2005),
relatively few studies have specifically focused on vegeta-
tion structure and tree assemblages in shade-grown coffee
farms (but see Bandeira et al., 2005; Soto-Pinto et al., 2001,
2007; Mendez et al., 2007).
This bias is important given the fact that trees are the
fundamental unit of shade coffee and other agroforestry
systems, providing habitat for multiple taxa, as well as
maintaining critical ecosystem functions such as nutrient
cycling, soil conservation, and productivity (Beer et al.,
1998; Soto-Pinto et al., 2000; Schroth et al., 2004). Besides
lessening the direct effects of habitat fragmentation and
habitat loss, the vegetation structure of shade coffee
plantations probably also serves as an effective buffer for
minimizing the indirect effects of such fragmentation (edge
effects) generated in landscapes with high structural
contrast such as those between forest reserves and
surrounding agricultural landscapes (Beer et al., 1998;
Perfecto and Vandermeer, 2002; Williams-Linera et al.,
2002). Additionally, the management and therefore
structure of coffee plantations is often extremely hetero-
geneous within the same region, thus providing a diverse
range of habitat types that facilitate the conservation of
native tree biodiversity (Bandeira et al., 2005; Soto-Pinto
et al., 2007), either by acting as reservoirs for species no
longer found in forest fragments (Williams-Linera et al.,
2005) or by contributing to a high beta diversity or species
turnover among patches that in turn results in high gamma
or landscape-scale diversity (Gordon et al., 2004; Williams-
Linera et al., 2005; Pineda et al., 2005; Arellano et al.,
2005).
The diversity and structure of shade in traditionally
managed coffee farms in Latin America is maintained by
farmers who use both planted and naturally established
species to provide a suite of important ecosystem goods and
services that lower production costs and increase sources of
revenue (Beer et al., 1998; Albertin and Nair, 2004; Soto-
Pinto et al., 2007; Mendez et al., 2007). These benefits are
typically obtained through a delicate balance of shade
intensity and composition by selecting introduced and native
tree species to minimize competition with coffee plants
while helping to ameliorate climate extremes, maintain soil
fertility, and provide important goods such as lumber,
fuelwood, and fruit (Beer et al., 1998; Albertin and Nair,
2004; Bandeira et al., 2005; Soto-Pinto et al., 2007).
Coffee is grown under diverse environmental conditions
in Mexico but its production is concentrated at altitudes
where quality is highest (between 600 and 1200 m) and
where high diversity forest ecosystems are found (Moguel
and Toledo, 1999). The structure and diversity of tree
canopies in coffee farms in Mexico may vary considerably
along a gradient of intensification and agrochemical use
(Nestel, 1995; Moguel and Toledo, 1999). However, such
shifts in intensification were particularly prominent in
eastern coastal states like Veracruz, where activities of the
governmental agency (INMECAFE) in charge of promoting
coffee ‘‘technification’’ were centered until the early 90s
(Potvin et al., 2005). Given their enhanced vulnerability to
the current coffee crises (Eakin et al., 2006), studies of
shade coffee canopies in this biodiversity rich state
(CONABIO, 1998) may be particularly relevant to regional
and national conservation efforts. This study is part of a
long-term multidisciplinary research project (BIOCAFE)
focused on quantifying changes in biodiversity (12 groups
of plants and animals), environmental services, and socio-
economic conditions in coffee farms under different
management strategies in central Veracruz. In this
particular study, we sought to compare patterns of tree
vegetation structure, richness, and diversity between shade
coffee agroecosystems and forest fragments remnants
typical for the region, and how this varied as a function
of general changes in management. We explored these
patterns using all species grouped together, or classified as
native or non-native, primary or secondary, and by seed
dispersal syndrome.
2. Methods
2.1. Study area
The study area is located in central Veracruz, Mexico
(198090–198270N and 968280–968580W) at altitudes ranging
from 1010 to 1340 m. This elevation corresponds with the
lower distributional limit of tropical montane cloud forest.
This coffee growing region is located largely between the
towns of Coatepec and Huatusco and includes over 20,000
farmers and almost 60,000 ha under cultivation, making it
the second most important coffee growing region in Mexico
(SAGARPA, 2007). Soils are mostly Andic Acrisols and
Cambisols. Mean annual temperature is 18–19 8C, and total
annual precipitation ranges between 1517 and 2200 mm.
Analysis of the landscape in a 2 km radius around all farms
using IKONOS satellite images revealed that annual crops
and sugar cane (42%), shade coffee farms (26%), and
pastures (19%) were the main elements of the matrix
surrounding highly fragmented and rapidly shrinking forest
remnants (currently 14%; Williams-Linera et al., 2002;
Lopez-Barrera and Landgrave, unpublished data).
A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172162
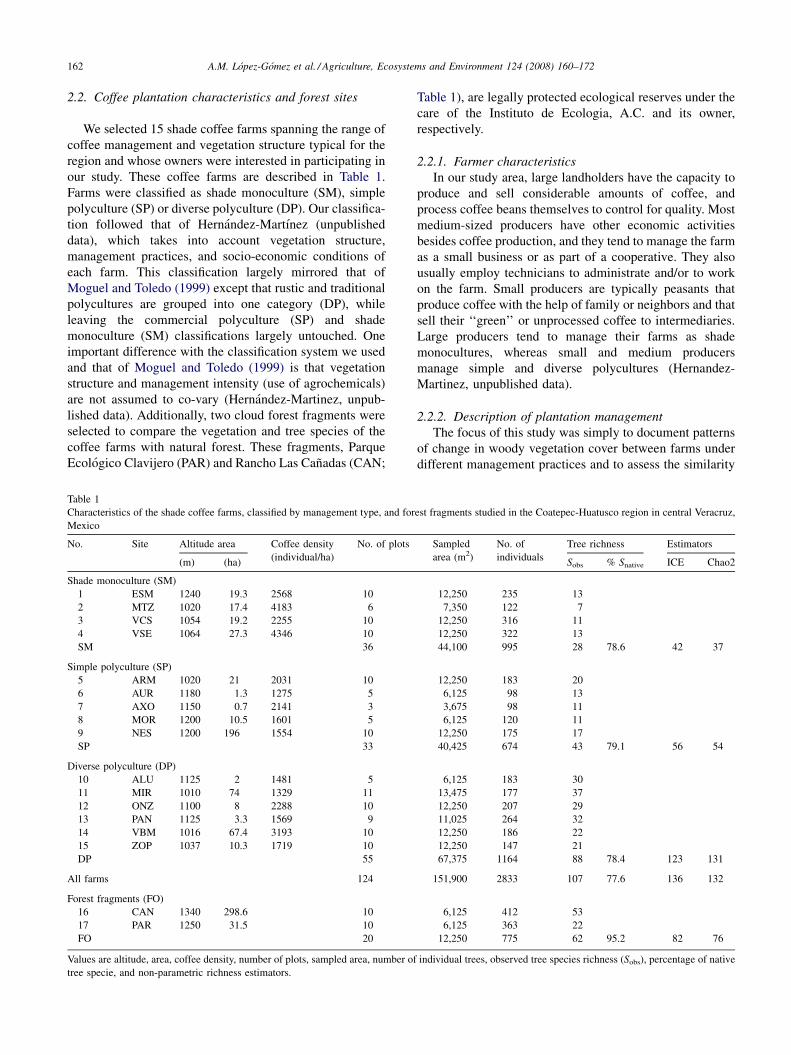
2.2. Coffee plantation characteristics and forest sites
We selected 15 shade coffee farms spanning the range of
coffee management and vegetation structure typical for the
region and whose owners were interested in participating in
our study. These coffee farms are described in Table 1.
Farms were classified as shade monoculture (SM), simple
polyculture (SP) or diverse polyculture (DP). Our classifica-
tion followed that of Hernandez-Martınez (unpublished
data), which takes into account vegetation structure,
management practices, and socio-economic conditions of
each farm. This classification largely mirrored that of
Moguel and Toledo (1999) except that rustic and traditional
polycultures are grouped into one category (DP), while
leaving the commercial polyculture (SP) and shade
monoculture (SM) classifications largely untouched. One
important difference with the classification system we used
and that of Moguel and Toledo (1999) is that vegetation
structure and management intensity (use of agrochemicals)
are not assumed to co-vary (Hernandez-Martinez, unpub-
lished data). Additionally, two cloud forest fragments were
selected to compare the vegetation and tree species of the
coffee farms with natural forest. These fragments, Parque
Ecologico Clavijero (PAR) and Rancho Las Canadas (CAN;
Table 1
Characteristics of the shade coffee farms, classified by management type, and for
Mexico
No. Site Altitude area Coffee density
(individual/ha)
No. of plots
(m) (ha)
Shade monoculture (SM)
1 ESM 1240 19.3 2568 10
2 MTZ 1020 17.4 4183 6
3 VCS 1054 19.2 2255 10
4 VSE 1064 27.3 4346 10
SM 36
Simple polyculture (SP)
5 ARM 1020 21 2031 10
6 AUR 1180 1.3 1275 5
7 AXO 1150 0.7 2141 3
8 MOR 1200 10.5 1601 5
9 NES 1200 196 1554 10
SP 33
Diverse polyculture (DP)
10 ALU 1125 2 1481 5
11 MIR 1010 74 1329 11
12 ONZ 1100 8 2288 10
13 PAN 1125 3.3 1569 9
14 VBM 1016 67.4 3193 10
15 ZOP 1037 10.3 1719 10
DP 55
All farms 124
Forest fragments (FO)
16 CAN 1340 298.6 10
17 PAR 1250 31.5 10
FO 20
Values are altitude, area, coffee density, number of plots, sampled area, number of
tree specie, and non-parametric richness estimators.
Table 1), are legally protected ecological reserves under the
care of the Instituto de Ecologia, A.C. and its owner,
respectively.
2.2.1. Farmer characteristics
In our study area, large landholders have the capacity to
produce and sell considerable amounts of coffee, and
process coffee beans themselves to control for quality. Most
medium-sized producers have other economic activities
besides coffee production, and they tend to manage the farm
as a small business or as part of a cooperative. They also
usually employ technicians to administrate and/or to work
on the farm. Small producers are typically peasants that
produce coffee with the help of family or neighbors and that
sell their ‘‘green’’ or unprocessed coffee to intermediaries.
Large producers tend to manage their farms as shade
monocultures, whereas small and medium producers
manage simple and diverse polycultures (Hernandez-
Martinez, unpublished data).
2.2.2. Description of plantation management
The focus of this study was simply to document patterns
of change in woody vegetation cover between farms under
different management practices and to assess the similarity
est fragments studied in the Coatepec-Huatusco region in central Veracruz,
Sampled
area (m2)
No. of
individuals
Tree richness Estimators
Sobs % Snative ICE Chao2
12,250 235 13
7,350 122 7
12,250 316 11
12,250 322 13
44,100 995 28 78.6 42 37
12,250 183 20
6,125 98 13
3,675 98 11
6,125 120 11
12,250 175 17
40,425 674 43 79.1 56 54
6,125 183 30
13,475 177 37
12,250 207 29
11,025 264 32
12,250 186 22
12,250 147 21
67,375 1164 88 78.4 123 131
151,900 2833 107 77.6 136 132
6,125 412 53
6,125 363 22
12,250 775 62 95.2 82 76
individual trees, observed tree species richness (Sobs), percentage of native

A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172 163
of this cover to that in nearby forest remnants and studies
performed in other regions. We did not seek to collect
extensive data on farmer management practices and other
factors that could be regulating the variation observed in
shade cover as this will be the subject of a future study.
Limited data collected to date suggests that shade
monocultures typically use few tree species, planted at
low densities, to create a minimal shade which alleviates
environmental stress for coffee plants while also insuring
enhanced coffee production. These farms usually employ a
more complete suite of conventional management practices
including commercial fertilizers, pest and weed control via
agrochemicals, systematic coffee pruning and shade
regulation by branch and epiphyte removal. Simple
polycultures are characterized by the removal of a large
proportion of the original forest canopy tree species and the
introduction of a set of some commercial species providing
appropriate shade for coffee cultivation. Management
practices are similar to those of shade monocultures
including sometimes pronounced use of agrochemicals for
fertilizer and the control of weeds and other pest species, as
well as coffee pruning. In diverse polycultures, coffee is
grown alongside numerous useful woody and herbaceous
plant species, forming a sophisticated system of managed
native and introduced species with the goal of increasing
revenues and lowering production costs. Main management
practices include manual weed control, coffee pruning and
chemical or organic fertilization. The characteristics of DPs
are intermixed in this classification, with some DPs being
more structurally complex and exhibiting greater diversity
than rustic farms. Thus, we did not separate a priori these
two management types in our classification (Hernandez-
Martınez, unpublished data).
2.3. Sampling design
We sampled a total of 124 35 m � 35 m (1225 m2) plots
distributed unevenly among the selected study farms due to
pronounced differences in their size (range 3–11 randomly-
located plots per site; Table 1). In each forest fragment, 10
plots were similarly sampled (Table 1). In each plot, dbh
(diameter at 1.3 m) and the height of all trees�5 cm dbh were
measured; this size class represents the great majority of trees
providing shade in coffee plantations. These data were then
used to calculate structural variables such as density, basal
area, and maximum and mean height per plot, study site, and
type of coffee management or forest. All woody plants in each
plot were identified to species or morphospecies following
Flora of Veracruz (Sosa and Gomez-Pompa, 1994) with
representative herbarium specimens collected and deposited
in the XAL herbarium in the Instituto de Ecologıa, A.C.
Tree species were classified as native or non-native.
Native species occur naturally in either lower montane or
premontane tropical forest, whereas non-native species are
exotic species introduced from any other type of vegetation
or region. Native species were further classified as either
primary or secondary tree species following Sosa and
Gomez-Pompa (1994) or the opinion of additional regional
experts. Finally, tree species were grouped according to seed
dispersal syndrome as either barochorous (seeds dispersed
by gravity), zoochorous (seeds dispersed by animals), or
anemochorous (seeds dispersed by wind).
2.4. Data analysis
We used a nested analysis of variance (ANOVA) with
sites (coffee plantations and forest fragments) nested in
categories. Prior to ANOVA, the normality of the structural
data was assessed using the Shapiro–Wilk’s test statistic and
logarithmic transformations were performed when neces-
sary. Relationship between species richness and structural
variables was analyzed using Pearson correlation coeffi-
cients. Analyses were performed using the statistical
package JMP version 3.2.2 (SAS, 1997). Tukey’s honest
statistical difference (HSD) was used for post-hoc compar-
isons. The proportions of the number of species or
individuals classified by dispersal syndromes as a function
of categories were compared using G-tests. Values reported
are mean � S.E. unless otherwise indicated.
Species–area curves for each management type were
constructed using EstimateS version 7.5.0 (Colwell, 2005),
and the non-parametric incidence-based estimators ICE and
Chao 2 were selected to calculate the completeness of the
inventories. To compare tree species richness between coffee
farms and forest, we computed rarefaction curves with the
expected richness function Mao Tau. The standardized
minimal number of individuals was used to estimate the
overall richness of each category, and tree richness for native
species classified as primary or secondary.
The species complementarity between coffee farms was
estimated as the proportion of total diversity in two sites that
occurred exclusively in only one site (Colwell and
Coddington, 1994). Also, species composition complemen-
tarities between coffee management type and forest were
estimated. Values of complementarity vary from zero
(identical species lists) to unity (completely distinct species
lists).
3. Results
3.1. Vegetation structure
The density, basal area and maximum height of
trees � 5 cm dbh were higher in forest than in coffee farms
(Fig. 1; Table 2), whereas mean height was statistically
similar between forest and SP and DP. Among farms,
structural variables were statistically similar between
polycultures (SPs and DPs), whereas tree density was
significantly higher in SMs and basal area, maximum and
mean height of trees � 5 cm dbh were higher in polycultures
(Fig. 1; Table 2).

A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172164
Fig. 1. Density, basal area, maximum and mean height for trees�5 cm dbh
in coffee farms and forests remnants in the Coatepec-Huatusco region of
central Veracruz, Mexico. Study sites are shade monocultures (SM), simple
polycultures (SP), diverse polycultures (DP), and forests (FO). Values are
means and error bars represent one S.E. Different letters indicate significant
differences (at alpha = 0.05).
Table 2
Results of ANOVA (nested design) performed using vegetation structure
variables for 15 shade coffee farms and two forest remnants in the Coatepec-
Huatusco region of central Veracruz, Mexico
Source DF SS F
Density
Site 13 1.41 6.63***
Category 3 4.98 101.49***
Basal area
Site 13 3.87 11.93***
Category 3 3.16 42.22***
Maximum height
Site 13 1935.32 20.34***
Category 3 1469.93 66.94***
Mean height
Site 13 1.70 23.97***
Category 3 0.84 51.40***
Study sites were nested within management type or forest (category).
3.2. Diversity analysis
3.2.1. Richness and abundance
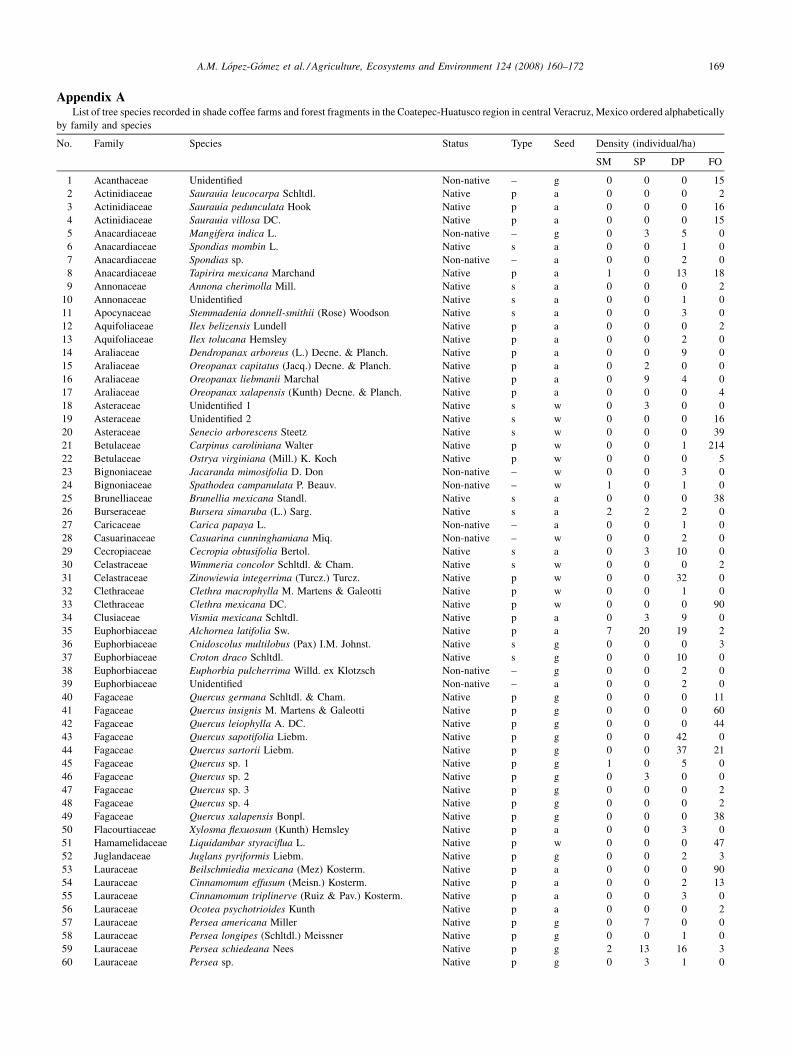
We identified 107 tree species � 5 cm dbh belonging to
78 genera and 41 families in the coffee farms included in this
study. Of these, 83 were native and 24 were non-native tree
species. Native trees comprised 50 primary and 33
secondary species. In total, including forest sites, we
recorded 153 tree species � 5 cm dbh, in 95 genera and 55
families (Appendix 1). Richness in SM farms averaged
11 � 1.4 species, compared to 14 � 1.8 in SPs, 29 � 2.3 in
DPs, and 38 � 16 in forest control sites (Table 1). Overall,
sampling revealed 28 tree species (22 natives) in SMs, 43
species (34 natives) in SPs, and 88 species (69 natives) in
DPs. In contrast, forests had only 62 tree species (59
natives). Interestingly, while total tree diversity changed
considerably with coffee management type the proportion of
native trees found in each class management was highly
consistent at around 79% (Table 1, Appendix 1).
The species–accumulation curves did not reach an
asymptote in any case indicating that the sampling effort
was not sufficient to complete the tree species inventories.
When all farm plots were pooled together, of the 107 species
of trees, 44 were rare (26 and 18, registered in just one or two
sites, respectively), and the non-parametric estimators ICE
and Chao2 indicated that 25–29 more species were needed to
complete the tree species inventories in the study region.
Some rare species recorded in only one farm and not in
forest were Ficus cotinifolia, F. obtusifolia, F. yoponensis,
Hamelia patens, Magnolia sp., Persea longipes, Pimenta
dioica and Prunus rhamnoides. Richness was analyzed
separately according to management type and species
classification as native and non-native. Observed richness
and estimated richness using ICE and Chao2 for each type of
management are presented in Table 1.
Rarefaction using common abundance levels for compar-
ing overall tree species richness (674 individuals), and
natives species (533 individuals) among coffee management
types and forest, revealed that richness was the highest in
DPs, followed by forest remnants, SPs, and SMs (Fig. 2A
and B). Likewise, tree richness was consistently highest for
primary species, followed by secondary species, and non-
native species.
Changes in the proportion of tree species with different
seed dispersal syndromes were observed both in terms of the
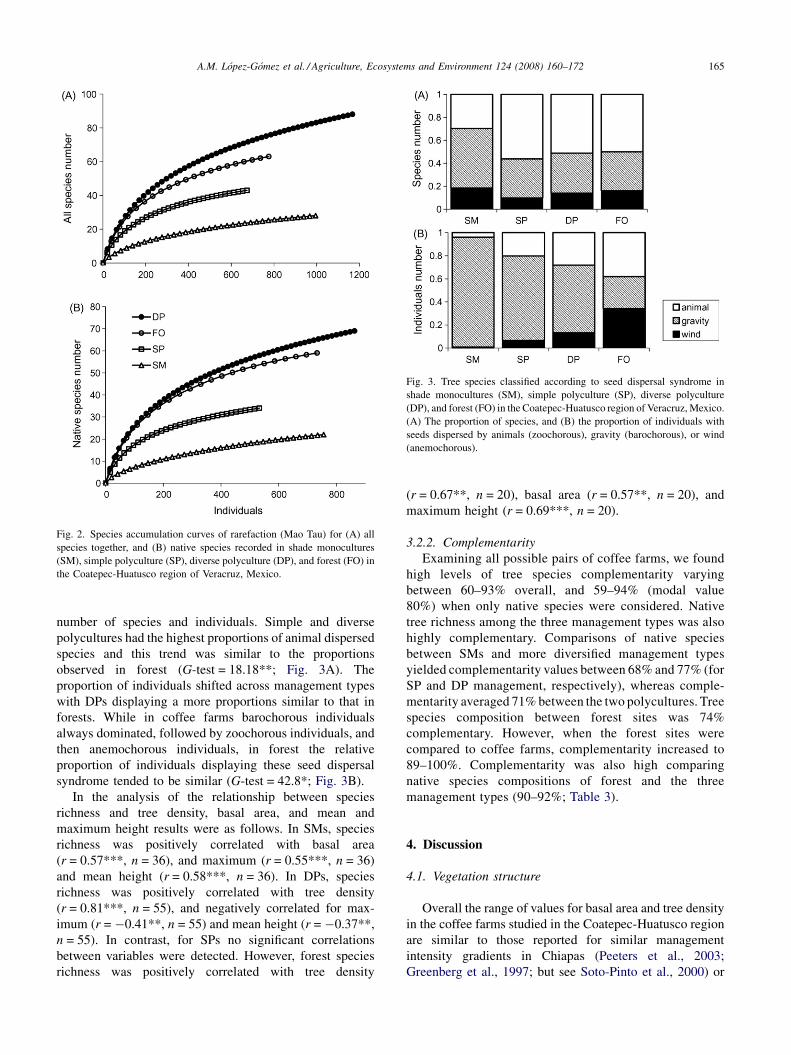
A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172 165
Fig. 2. Species accumulation curves of rarefaction (Mao Tau) for (A) all
species together, and (B) native species recorded in shade monocultures
(SM), simple polyculture (SP), diverse polyculture (DP), and forest (FO) in
the Coatepec-Huatusco region of Veracruz, Mexico.
Fig. 3. Tree species classified according to seed dispersal syndrome in
shade monocultures (SM), simple polyculture (SP), diverse polyculture
(DP), and forest (FO) in the Coatepec-Huatusco region of Veracruz, Mexico.
(A) The proportion of species, and (B) the proportion of individuals with
seeds dispersed by animals (zoochorous), gravity (barochorous), or wind
(anemochorous).
number of species and individuals. Simple and diverse
polycultures had the highest proportions of animal dispersed
species and this trend was similar to the proportions
observed in forest (G-test = 18.18**; Fig. 3A). The
proportion of individuals shifted across management types
with DPs displaying a more proportions similar to that in
forests. While in coffee farms barochorous individuals
always dominated, followed by zoochorous individuals, and
then anemochorous individuals, in forest the relative
proportion of individuals displaying these seed dispersal
syndrome tended to be similar (G-test = 42.8*; Fig. 3B).
In the analysis of the relationship between species
richness and tree density, basal area, and mean and
maximum height results were as follows. In SMs, species
richness was positively correlated with basal area
(r = 0.57***, n = 36), and maximum (r = 0.55***, n = 36)
and mean height (r = 0.58***, n = 36). In DPs, species
richness was positively correlated with tree density
(r = 0.81***, n = 55), and negatively correlated for max-
imum (r = �0.41**, n = 55) and mean height (r = �0.37**,
n = 55). In contrast, for SPs no significant correlations
between variables were detected. However, forest species
richness was positively correlated with tree density
(r = 0.67**, n = 20), basal area (r = 0.57**, n = 20), and
maximum height (r = 0.69***, n = 20).
3.2.2. Complementarity
Examining all possible pairs of coffee farms, we found
high levels of tree species complementarity varying
between 60–93% overall, and 59–94% (modal value
80%) when only native species were considered. Native
tree richness among the three management types was also
highly complementary. Comparisons of native species
between SMs and more diversified management types
yielded complementarity values between 68% and 77% (for
SP and DP management, respectively), whereas comple-
mentarity averaged 71% between the two polycultures. Tree
species composition between forest sites was 74%
complementary. However, when the forest sites were
compared to coffee farms, complementarity increased to
89–100%. Complementarity was also high comparing
native species compositions of forest and the three
management types (90–92%; Table 3).
4. Discussion
4.1. Vegetation structure
Overall the range of values for basal area and tree density
in the coffee farms studied in the Coatepec-Huatusco region
are similar to those reported for similar management
intensity gradients in Chiapas (Peeters et al., 2003;
Greenberg et al., 1997; but see Soto-Pinto et al., 2000) or

A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172166
Table 3
Native tree species complementarity (%) between coffee farms under different management: SM (1–4), SP (5–9), DP (10–15), and forest fragments FO (16–17)
in the Coatepec-Huatusco region of central Veracruz, Mexico
rustic farms located in the Chinantla region, Oaxaca
(Bandeira et al., 2005). Nevertheless, vegetation structure
appears more simplified than that observed in forest
fragments in the same region. Basal area, density and
height of the coffee farms included in our study were far
lower than the values reported for natural vegetation
(Williams-Linera, 2002; Williams-Linera et al., 2005).
Values of basal area obtained from the polyculture and shade
monoculture coffee farms in this study were actually similar
to values recorded in old-fields following 15–17 and 8–9
years of secondary succession, respectively (Muniz-Castro,
unpublished data).
Trees are smaller in SMs, mainly because of the species
composition, but also possibly because of more intensive
pruning of shade trees. The most frequent species in SMs
(Inga vera and Mimosa scabrella) do not reach great
heights or diameters, which could also help explain the
poor structure observed. Conversely, some abundant tree
species registered in DPs have large basal areas, as is the
case for Enterolobium cyclocarpum, Quercus sapotaefolia,
and Zinowewia integerrima. In SM farms, owners carry out
regular pruning practices to avoid interference with less
shade-tolerant coffee variants that are not as common in
shade polycultures (Hernandez-Martınez, personal com-
munication). Alternatively, this pattern is consistent with
the fact that SM farms are fairly young, most having been
established in the last few decades (BIOCAFE, unpub-
lished data). Canopy height, which differed markedly
among the management types we studied, is a factor
deemed very important by coffee growers in this (Cruz-
Angon and Greenberg, 2005) and other regions (Albertin
and Nair, 2004). As the diversity of several important taxa
such as birds and monkeys appear to be strongly positively
correlated with this variable (Greenberg et al., 1997;
Williams-Guillen et al., 2006; Gordon et al., 2007), the
overall net effect of the observed changes in vegetation
structure on regional faunistic diversity remains somewhat
unclear and warrants further study. The absence of
structural differences between SPs and DPs suggest that
in polycultures species composition may be a more
important focus for management by farmers than the tree
density, basal area, or height. The renewal of tree shade in
some polycultures (e. g. farms 8 and 11) and the partial
abandonment of some others due to low coffee prices
(farms 6 or 13), is not reflected in shade structure
differences between SPs and DPs, although an explicit
exploration of farmers management of shade structure
(e.g., Mendez et al., 2007; Soto-Pinto et al., 2007) in these
types of farms is missing for central Veracruz and is
urgently needed.

A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172 167
An important factor to consider in assessing the general
impact of changes in woody vegetation structure in coffee
farms on the conservation of regional biodiversity is the
landscape context (Schroth et al., 2004; Vandermeer and
Perfecto, 2007). Regional trends in the mountains of
central Veracruz suggest an overall simplification of the
landscape with forest cover being replaced by cattle
ranching, row crops and housing developments (Williams-
Linera et al., 2002; Lopez-Barrera and Landgrave,
unpublished data). In this type of setting, landscape
elements which enhance structural heterogeneity and
connectivity may promote biodiversity conservation
(Ricketts et al., 2001; Perfecto and Vandermeer, 2002;
Bandeira et al., 2005; Williams-Linera et al., 2005) and
ecosystem function (Klein et al., 2003). The strong site-
by-site variation in vegetation structure we observed in our
study sites (Fig. 1), suggests that coffee farms in central
Veracruz may indeed be helping to conserve habitat and
biological diversity in the region. That variation also
highlights the need for future studies to understand the
particular decisions made by farmers in their manage-
ment of shade trees (Kindt et al., 2004; Mendez et al.,
2007).
4.2. Species richness
The tree species richness reported in this study (153 tree
species � 5 cm dbh, 107 in coffee farms and 62 species in
forests) is high and similar to values reported for natural
vegetation in the same region. In central Veracruz, tree
species richness pooled over 10 forest fragments was 86
species, and when other landscapes elements such as active
(54 species) and abandoned (62 species) coffee plantations
and old fields (53 species) were incorporated in the
biodiversity analysis, the overall regional richness went
up to 156 tree species (Williams-Linera et al., 2005). Our
values are similar or slightly higher than those reported from
coffee plantations in Chiapas and Guerrero (74–98 species;
Rendon and Turrubiarte, 1985; Nunez, 1987; Soto-Pinto
et al., 2001, 2007), but are substantially higher than values
reported for Oaxaca (45 species, Bandeira et al., 2005) and
may reflect strong differences in the management strategies
employed within these regions (Nestel, 1995; Potvin et al.,
2005). Species accumulation curves generated for coffee
farms in central Veracruz imply that 25–29 more additional
species would be detected with additional sampling, which
might result in even greater disparities in tree diversity
between these coffee-growing regions. This result is
somewhat surprising since several authors have highlighted
the fact that the intensification and simplification of coffee
farms was more elevated in Veracruz than other coffee
growing regions in Mexico (Nestel, 1995; Potvin et al.,
2005)
Tree species richness was lower in SMs than in SPs and
DPs as has been observed in other studies (e.g., Peeters et al.,
2003). The fact that DPs displayed higher species richness
than forest sites may be related to the great beta diversity of
the regional montane cloud forest due to shift of tree species
composition from one site to the next one (Williams-Linera,
2002), and the fact that we only included two forests
fragments.
All three types of coffee management exhibited a
surprisingly high and consistent proportion of native
species (79%), which is similar to that found in coffee
farms located elsewhere in Mexico (Bandeira et al., 2005)
and Latin America (Mendez et al., 2007), but higher than
values reported in some African coffee farms (Degrande
et al., 2006). Such high proportions of native species in
coffee plantations is undoubtedly a reflection of farmer’s
intimate knowledge of their characteristics and potential
uses (Soto-Pinto et al., 2007), as well as their enhanced
availability due to seed dispersion and natural establish-
ment (Bandeira et al., 2005; Mendez et al., 2007), and the
fact that they are better adapted to local climatic conditions
(Albertin and Nair, 2004). However, while native tree
species typically dominate in these agroecosystems (Soto-
Pinto et al., 2001; Bandeira et al., 2005) they may not
necessarily be species with global conservation importance
(Gordon et al., 2003a; Mendez et al., 2007). In our study,
only three native species were identified that are of
international conservation concern: Juglans pyriformis,
Swietenia macrophylla and Cedrela odorata. The high
proportion of native species observed in this study suggests
that even with intensive management, shade coffee
plantations can make important contributions to the
preservation of the regional forest diversity (Soto-Pinto
et al., 2001; Bandeira et al., 2005).
This conclusion is further strengthened by the observa-
tion that the majority of species (71%) we surveyed were
rare and found only in one or two coffee farms, and that
comparisons between all coffee farms (range 60–93%) and
between groups of farms organized by management type
(68–77%) exhibited high species complementarity. Thus,
while the structure and composition of SMs differ
substantially from that in polycultures, farms under each
type of management appear to play a unique and important
role in conserving the regional pool of native tree species.
Previous studies have documented 27 species of trees no
longer found in sampled regional forests that were surviving
in coffee farms (Williams-Linera et al., 2005; unpublished
data). These findings support, at least in the region of central
Veracruz, the idea of coffee plantations as diversity refuges
(sensu Perfecto et al., 1996) and argue against the practice of
considering shade coffee farms as largely depauperate of
native tree species (Rappole et al., 2003a). Nevertheless,
more multi-taxon studies are needed to understand the net
importance of coffee agroecosystems for the conservation of
regional biodiversity pools (Perfecto et al., 2003; Steffan-
Dewenter et al., 2007).
Apart from the tree species richness, the functional
diversity of the species present is another important
consideration in evaluating the conservation value of

A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172168
shade coffee farms (Mayfield et al., 2005). Species of the
genus Inga are the most abundant in all three types of
coffee managements in central Veracruz, as well as in
polycultures and rustic systems in Chiapas and Oaxaca
(e.g., Soto-Pinto et al., 2001; Peeters et al., 2003;
Bandeira et al., 2005). These legume species have been
traditionally promoted by coffee growers as they are
thought to enhance nitrogen fixation, provide good shade,
abundant litterfall to enhance fertility and minimize the
erosion of soil, and as producers of edible fruits
(Villalvazo-Lopez, 2002). However, recent evidence
suggests the presence of these species does not improve
the quality of the coffee crops (Romero-Alvarado et al.,
2002). The abundance of other species registered in the
shade polycultures of our study, such as Citrus spp.,
Mangifera indica, Psidium guajava, and Persea schie-
deana, is also undoubtedly linked to their promotion by
farmers due to their value as fruit for human consumption,
or for fuelwood such as in the case of Cedrela odorata or
Trema micrantha (Beer et al., 1998; Albertin and Nair,
2004; Mendez et al., 2007). Peeters et al. (2003) have
shown that even when coffee production, and the density
of coffee and trees does not vary between management
types, timber and firewood production may be signifi-
cantly higher in DP versus SM. Considering these and
other ecosystem services, there appears little justification
for replacing traditional diverse plantations with those
dominated by a few species such as Inga spp. More
studies are needed in central Veracruz exploring manage-
ment practices of farms and how the selection of tree
species affects biodiversity and helps lower production
costs and increase sources of revenue (Albertin and Nair,
2004; Soto-Pinto et al., 2007). Such studies are
particularly relevant in areas hit hard by the current
crises of overproduction and low coffee prices (Eakin
et al., 2006).
High tree species richness and functional diversity in
coffee plantations is directly linked to the conservation of
many other plant and animal taxa (Perfecto et al., 1996;
Moguel and Toledo, 1999). In coffee plantations in the
Dominican Republic and elsewhere Inga vera was found
to strongly influence the foraging behavior and presence
of birds as did the overall abundance, variety, and
consistency of food resources provided by trees in these
agroecosystems (Wunderle and Latta, 1998). The struc-
tural and biological diversity of shade species in coffee
farms have also been shown to be important in
maintaining the diversity of twig-nesting ants in Colombia
(Armbrecht et al., 2004) and the epiphyte community in
central Veracruz (Solis-Montero et al., 2005). In the
current study, animal and gravity-dispersed species made
up a much large proportion of total tree species than wind-
dispersed species (Fig. 3). Both the richness and number
of animal seed dispersed species in particular increased
within more traditionally managed coffee farms suggest-
ing an important food resource for frugivorous birds, bats,
and intermediate-size mammals (e.g., Gallina et al., 1996;
Galindo-Gonzalez et al., 2000; Carlo et al., 2004). This
finding could also suggest that farmers use more naturally
establishing species in these types of coffee management
(unplanned diversity sensu Mendez et al., 2007; Bandeira
et al., 2005).
4.3. Management implications
In this study, we have sought to highlight a number of
factors that may be important in influencing tree species
richness and structural diversity in shade coffee planta-
tions in central Veracruz. Species richness was positively
related to tree density only in DPs and forest, suggesting
that DPs are more important for conservation; however,
since the highest number of individual trees was recorded
in SMs, overall species richness is not a function of higher
number of individuals, contrary to the finding of Mendez
et al. (2007). Exploring these factors in greater detail in
future studies should help clarify the debate concerning
the role of coffee agroecosystems in the conservation of
regional biodiversity pools (Philpott and Dietsch, 2003;
Rappole et al., 2003a,b; O’Brien and Kinnaird, 2003,
2004; Dietsch et al., 2004). In this context, data from this
and other studies suggest that it is imperative that other
land uses, such as shade coffee agroecosystems be
considered and explicitly included in regional conserva-
tion strategies (Gordon et al., 2003b; Schroth et al., 2004;
Williams-Linera et al., 2005; Vandermeer and Perfecto,
2007). Explicit consideration of factors such as com-
plementarity, landscape heterogeneity, and functional
diversity may also help improve the impact of coffee
certification programs designed to promote biodiversity
conservation. Results from this and other studies suggest
that coffee management systems need to be evaluated
using rigorous vegetation surveys to determine if they
meet certification criteria and to test the hypothesis that
such certified farms actual support higher levels of
biodiversity than those that are not certified (Mas and
Dietsch, 2004).
Acknowledgments
We thank the owners of the coffee farms included in this
study for their permission to work in their farms. We are
grateful to many people who helped with fieldwork
(Eduardo Martınez, Ines Morato, Anıbal Niembro), to
identify specimens at the Herbarium XAL (Claudia
Gallardo, Carlos Duran), and with useful discussion
(Gerardo Hernandez-Martınez). Two anonymous reviewers
and Dr. A. de Rouw provided many helpful comments on the
manuscript. This study is part of the project BIOCAFE
funded by SEMARNAT-CONACyT, 2002-C01-0194, and
was supported in part by the Department of Functional
Ecology, Instituto de Ecologıa, A.C. (IE 902-11).
A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172 169
Appendix AList of tree species recorded in shade coffee farms and forest fragments in the Coatepec-Huatusco region in central Veracruz, Mexico ordered alphabetically
by family and species
No. Family Species Status Type Seed Density (individual/ha)
SM SP DP FO
1 Acanthaceae Unidentified Non-native – g 0 0 0 15
2 Actinidiaceae Saurauia leucocarpa Schltdl. Native p a 0 0 0 2
3 Actinidiaceae Saurauia pedunculata Hook Native p a 0 0 0 16
4 Actinidiaceae Saurauia villosa DC. Native p a 0 0 0 15
5 Anacardiaceae Mangifera indica L. Non-native – g 0 3 5 0
6 Anacardiaceae Spondias mombin L. Native s a 0 0 1 0
7 Anacardiaceae Spondias sp. Non-native – a 0 0 2 0
8 Anacardiaceae Tapirira mexicana Marchand Native p a 1 0 13 18
9 Annonaceae Annona cherimolla Mill. Native s a 0 0 0 2
10 Annonaceae Unidentified Native s a 0 0 1 0
11 Apocynaceae Stemmadenia donnell-smithii (Rose) Woodson Native s a 0 0 3 0
12 Aquifoliaceae Ilex belizensis Lundell Native p a 0 0 0 2
13 Aquifoliaceae Ilex tolucana Hemsley Native p a 0 0 2 0
14 Araliaceae Dendropanax arboreus (L.) Decne. & Planch. Native p a 0 0 9 0
15 Araliaceae Oreopanax capitatus (Jacq.) Decne. & Planch. Native p a 0 2 0 0
16 Araliaceae Oreopanax liebmanii Marchal Native p a 0 9 4 0
17 Araliaceae Oreopanax xalapensis (Kunth) Decne. & Planch. Native p a 0 0 0 4
18 Asteraceae Unidentified 1 Native s w 0 3 0 0
19 Asteraceae Unidentified 2 Native s w 0 0 0 16
20 Asteraceae Senecio arborescens Steetz Native s w 0 0 0 39
21 Betulaceae Carpinus caroliniana Walter Native p w 0 0 1 214
22 Betulaceae Ostrya virginiana (Mill.) K. Koch Native p w 0 0 0 5
23 Bignoniaceae Jacaranda mimosifolia D. Don Non-native – w 0 0 3 0
24 Bignoniaceae Spathodea campanulata P. Beauv. Non-native – w 1 0 1 0
25 Brunelliaceae Brunellia mexicana Standl. Native s a 0 0 0 38
26 Burseraceae Bursera simaruba (L.) Sarg. Native s a 2 2 2 0
27 Caricaceae Carica papaya L. Non-native – a 0 0 1 0
28 Casuarinaceae Casuarina cunninghamiana Miq. Non-native – w 0 0 2 0
29 Cecropiaceae Cecropia obtusifolia Bertol. Native s a 0 3 10 0
30 Celastraceae Wimmeria concolor Schltdl. & Cham. Native s w 0 0 0 2
31 Celastraceae Zinowiewia integerrima (Turcz.) Turcz. Native p w 0 0 32 0
32 Clethraceae Clethra macrophylla M. Martens & Galeotti Native p w 0 0 1 0
33 Clethraceae Clethra mexicana DC. Native p w 0 0 0 90
34 Clusiaceae Vismia mexicana Schltdl. Native p a 0 3 9 0
35 Euphorbiaceae Alchornea latifolia Sw. Native p a 7 20 19 2
36 Euphorbiaceae Cnidoscolus multilobus (Pax) I.M. Johnst. Native s g 0 0 0 3
37 Euphorbiaceae Croton draco Schltdl. Native s g 0 0 10 0
38 Euphorbiaceae Euphorbia pulcherrima Willd. ex Klotzsch Non-native – g 0 0 2 0
39 Euphorbiaceae Unidentified Non-native – a 0 0 2 0
40 Fagaceae Quercus germana Schltdl. & Cham. Native p g 0 0 0 11
41 Fagaceae Quercus insignis M. Martens & Galeotti Native p g 0 0 0 60
42 Fagaceae Quercus leiophylla A. DC. Native p g 0 0 0 44
43 Fagaceae Quercus sapotifolia Liebm. Native p g 0 0 42 0
44 Fagaceae Quercus sartorii Liebm. Native p g 0 0 37 21
45 Fagaceae Quercus sp. 1 Native p g 1 0 5 0
46 Fagaceae Quercus sp. 2 Native p g 0 3 0 0
47 Fagaceae Quercus sp. 3 Native p g 0 0 0 2
48 Fagaceae Quercus sp. 4 Native p g 0 0 0 2
49 Fagaceae Quercus xalapensis Bonpl. Native p g 0 0 0 38
50 Flacourtiaceae Xylosma flexuosum (Kunth) Hemsley Native p a 0 0 3 0
51 Hamamelidaceae Liquidambar styraciflua L. Native p w 0 0 0 47
52 Juglandaceae Juglans pyriformis Liebm. Native p g 0 0 2 3
53 Lauraceae Beilschmiedia mexicana (Mez) Kosterm. Native p a 0 0 0 90
54 Lauraceae Cinnamomum effusum (Meisn.) Kosterm. Native p a 0 0 2 13
55 Lauraceae Cinnamomum triplinerve (Ruiz & Pav.) Kosterm. Native p a 0 0 3 0
56 Lauraceae Ocotea psychotrioides Kunth Native p a 0 0 0 2
57 Lauraceae Persea americana Miller Native p g 0 7 0 0
58 Lauraceae Persea longipes (Schltdl.) Meissner Native p g 0 0 1 0
59 Lauraceae Persea schiedeana Nees Native p g 2 13 16 3
60 Lauraceae Persea sp. Native p g 0 3 1 0

A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172170
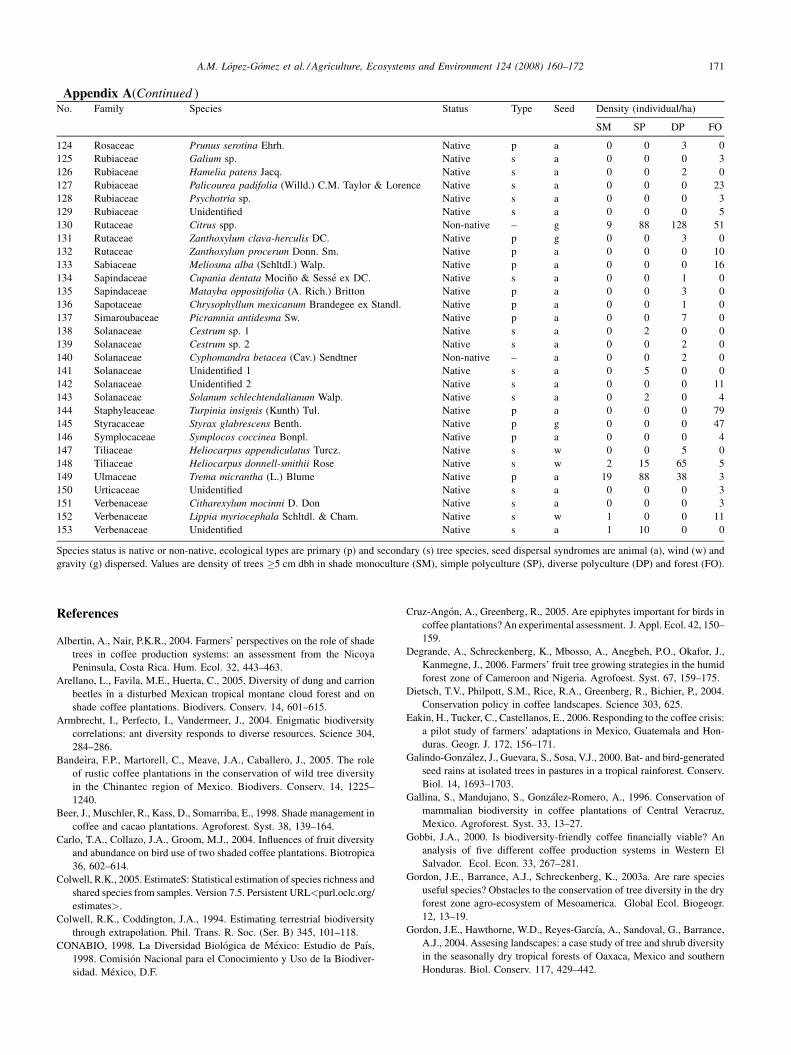
Appendix A(Continued )No. Family Species Status Type Seed Density (individual/ha)
SM SP DP FO
61 Leguminosae Acacia cornigera (L.) Willd. Native s g 2 0 0 0
62 Leguminosae Acacia pennatula (Schltdl. & Cham.) Benth. Native s a 0 1 1 0
63 Leguminosae Acrocarpus fraxinifolius Wright & Arn. Non-native – g 0 47 0 0
64 Leguminosae Bauhinia divaricata L. Non-native – g 0 0 24 0
65 Leguminosae Cojoba arborea (L.) Britton & Rose Native p g 1 0 4 4
66 Leguminosae Enterolobium cyclocarpum (Jacq.) Griseb. Native p g 1 40 39 0
67 Leguminosae Erythrina americana Miller Native s g 0 2 11 5
68 Leguminosae Erythrina poeppigiana (Walp.) Skeels. Non-native – g 47 0 0 0
69 Leguminosae Gliricidia sepium (Jacq.) Kunth ex Walp. Native s g 4 0 0 0
70 Leguminosae Inga jinicuil Schltr. Native s g 18 60 84 0
71 Leguminosae Inga latibracteata Harms Native s g 5 143 15 0
72 Leguminosae Inga punctata Willd. Native s g 7 12 23 0
73 Leguminosae Inga sp. 1 Non-native – g 0 10 0 0
74 Leguminosae Inga sp. 2 Non-native – g 0 0 13 0
75 Leguminosae Inga sp. 3 Native s g 0 0 0 10
76 Leguminosae Inga sp. 4 Native s g 0 0 0 12
77 Leguminosae Inga spuria Humb. & Bonpl. Ex Willd. Native s g 0 0 9 0
78 Leguminosae Inga vera Willd. Native s g 602 228 153 0
79 Leguminosae Unidentified 1 Non-native – g 0 5 0 0
80 Leguminosae Unidentified 2 Native s g 0 0 0 3
81 Leguminosae Leucaena leucocephala (Lam.) de Wit Non-native – g 3 0 1 2
82 Leguminosae Lonchocarpus guatemalensis Benth. Native p w 0 24 1 0
83 Leguminosae Mimosa scabrella Benth. Non-native – g 136 0 8 0
84 Leguminosae Piscidia piscipula (L.) Sarg. Native p g 0 0 1 0
85 Magnoliaceae Magnolia schiedeana Schltdl. Native p g 0 0 1 0
86 Malpighiaceae Bunchosia lindeniana A. Juss. Native s a 0 0 3 0
87 Malpighiaceae Byrsonima crassifolia (L.) Kunth Native s a 2 0 1 0
88 Malvaceae Hampea integerrima Schltdl. Native s a 0 0 0 51
89 Melastomataceae Unidentified 1 Native p a 0 2 1 0
90 Melastomataceae Unidentified 2 Native p a 0 0 0 7
91 Melastomataceae Miconia sp. 1 Native p a 0 0 1 0
92 Melastomataceae Miconia sp. 2 Native p a 0 0 0 2
93 Melastomataceae Tibouchina sp. Native s a 0 0 4 0
94 Meliaceae Cedrela odorata L. Native p w 2 18 17 0
95 Meliaceae Melia azedarach L. Non-native – a 0 14 6 0
96 Meliaceae Swietenia macrophylla King Native p w 0 0 14 0
97 Meliaceae Trichilia havanensis Jacq. Native p a 0 5 12 0
98 Monimiaceae Siparuna andina (Tul.) A. DC. Native s a 0 0 0 7
99 Moraceae Ficus calyculata P. Miller Native p a 2 11 8 0
100 Moraceae Ficus cotinifolia Kunth Native p a 0 0 1 0
101 Moraceae Ficus obtusifolia Kunth Native p a 0 1 0 0
102 Moraceae Ficus pertusa L. Native p a 0 2 4 0
103 Moraceae Ficus sp. 1 Non-native – a 0 2 0 0
104 Moraceae Ficus sp. 2 Native p a 0 0 1 0
105 Moraceae Ficus sp. 3 Native p a 0 0 1 0
106 Moraceae Ficus yoponensis Desv. Native p a 0 0 1 0
107 Moraceae Trophis mexicana (Liebm.) Bureau Native p a 0 0 0 2
108 Moraceae Trophis racemosa (L.) Urb. Native s g 0 0 3 0
109 Myrsinaceae Ardisia compressa Kunth Native s a 0 0 4 0
110 Myrsinaceae Myrsine coriacea (Sw.) R. Br. ex Roem. & Schult. Native p a 0 2 6 46
111 Myrtaceae Eugenia sp. Native p a 0 0 0 11
112 Myrtaceae Pimenta dioica (L.) Merr. Native p a 0 2 0 0
113 Myrtaceae Psidium guajava L. Native s a 0 1 22 0
114 Myrtaceae Psidium sartorianum (O. Berg) Nied. Native p a 1 0 3 0
115 Myrtaceae Syzygium jambos (L.) Alston Non-native – a 0 2 17 0
116 Piperaceae Piper nudum C. DC. Native p a 0 0 1 0
117 Platanaceae Platanus mexicana Moric. Native p w 0 0 0 8
118 Polygonaceae Coccoloba barbadensis Jacq. Native s a 1 0 0 0
119 Proteaceae Grevillea robusta A. Cunn. ex R. Br. Non-native – w 2 0 7 0
120 Proteaceae Macadamia tetraphylla L.A.S. Johnson Non-native – g 0 0 11 0
121 Rhamnaceae Rhamus capreifolia Schltdl. Native p a 0 0 0 3
122 Rosaceae Eriobotrya japonica Lindley Non-native – a 0 1 77 0
123 Rosaceae Prunus rhamnoides Koehne Native p a 0 0 2 0
A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172 171
Appendix A(Continued )No. Family Species Status Type Seed Density (individual/ha)
SM SP DP FO
124 Rosaceae Prunus serotina Ehrh. Native p a 0 0 3 0
125 Rubiaceae Galium sp. Native s a 0 0 0 3
126 Rubiaceae Hamelia patens Jacq. Native s a 0 0 2 0
127 Rubiaceae Palicourea padifolia (Willd.) C.M. Taylor & Lorence Native s a 0 0 0 23
128 Rubiaceae Psychotria sp. Native s a 0 0 0 3
129 Rubiaceae Unidentified Native s a 0 0 0 5
130 Rutaceae Citrus spp. Non-native – g 9 88 128 51
131 Rutaceae Zanthoxylum clava-herculis DC. Native p g 0 0 3 0
132 Rutaceae Zanthoxylum procerum Donn. Sm. Native p a 0 0 0 10
133 Sabiaceae Meliosma alba (Schltdl.) Walp. Native p a 0 0 0 16
134 Sapindaceae Cupania dentata Mocino & Sesse ex DC. Native s a 0 0 1 0
135 Sapindaceae Matayba oppositifolia (A. Rich.) Britton Native p a 0 0 3 0
136 Sapotaceae Chrysophyllum mexicanum Brandegee ex Standl. Native p a 0 0 1 0
137 Simaroubaceae Picramnia antidesma Sw. Native p a 0 0 7 0
138 Solanaceae Cestrum sp. 1 Native s a 0 2 0 0
139 Solanaceae Cestrum sp. 2 Native s a 0 0 2 0
140 Solanaceae Cyphomandra betacea (Cav.) Sendtner Non-native – a 0 0 2 0
141 Solanaceae Unidentified 1 Native s a 0 5 0 0
142 Solanaceae Unidentified 2 Native s a 0 0 0 11
143 Solanaceae Solanum schlechtendalianum Walp. Native s a 0 2 0 4
144 Staphyleaceae Turpinia insignis (Kunth) Tul. Native p a 0 0 0 79
145 Styracaceae Styrax glabrescens Benth. Native p g 0 0 0 47
146 Symplocaceae Symplocos coccinea Bonpl. Native p a 0 0 0 4
147 Tiliaceae Heliocarpus appendiculatus Turcz. Native s w 0 0 5 0
148 Tiliaceae Heliocarpus donnell-smithii Rose Native s w 2 15 65 5
149 Ulmaceae Trema micrantha (L.) Blume Native p a 19 88 38 3
150 Urticaceae Unidentified Native s a 0 0 0 3
151 Verbenaceae Citharexylum mocinni D. Don Native s a 0 0 0 3
152 Verbenaceae Lippia myriocephala Schltdl. & Cham. Native s w 1 0 0 11
153 Verbenaceae Unidentified Native s a 1 10 0 0
Species status is native or non-native, ecological types are primary (p) and secondary (s) tree species, seed dispersal syndromes are animal (a), wind (w) and
gravity (g) dispersed. Values are density of trees �5 cm dbh in shade monoculture (SM), simple polyculture (SP), diverse polyculture (DP) and forest (FO).
References
Albertin, A., Nair, P.K.R., 2004. Farmers’ perspectives on the role of shade
trees in coffee production systems: an assessment from the Nicoya
Peninsula, Costa Rica. Hum. Ecol. 32, 443–463.
Arellano, L., Favila, M.E., Huerta, C., 2005. Diversity of dung and carrion
beetles in a disturbed Mexican tropical montane cloud forest and on
shade coffee plantations. Biodivers. Conserv. 14, 601–615.
Armbrecht, I., Perfecto, I., Vandermeer, J., 2004. Enigmatic biodiversity
correlations: ant diversity responds to diverse resources. Science 304,
284–286.
Bandeira, F.P., Martorell, C., Meave, J.A., Caballero, J., 2005. The role
of rustic coffee plantations in the conservation of wild tree diversity
in the Chinantec region of Mexico. Biodivers. Conserv. 14, 1225–
1240.
Beer, J., Muschler, R., Kass, D., Somarriba, E., 1998. Shade management in
coffee and cacao plantations. Agroforest. Syst. 38, 139–164.
Carlo, T.A., Collazo, J.A., Groom, M.J., 2004. Influences of fruit diversity
and abundance on bird use of two shaded coffee plantations. Biotropica
36, 602–614.
Colwell, R.K., 2005. EstimateS: Statistical estimation of species richness and
shared species from samples. Version 7.5. Persistent URL<purl.oclc.org/
estimates>.
Colwell, R.K., Coddington, J.A., 1994. Estimating terrestrial biodiversity
through extrapolation. Phil. Trans. R. Soc. (Ser. B) 345, 101–118.
CONABIO, 1998. La Diversidad Biologica de Mexico: Estudio de Paıs,
1998. Comision Nacional para el Conocimiento y Uso de la Biodiver-
sidad. Mexico, D.F.
Cruz-Angon, A., Greenberg, R., 2005. Are epiphytes important for birds in
coffee plantations? An experimental assessment. J. Appl. Ecol. 42, 150–
159.
Degrande, A., Schreckenberg, K., Mbosso, A., Anegbeh, P.O., Okafor, J.,
Kanmegne, J., 2006. Farmers’ fruit tree growing strategies in the humid
forest zone of Cameroon and Nigeria. Agrofoest. Syst. 67, 159–175.
Dietsch, T.V., Philpott, S.M., Rice, R.A., Greenberg, R., Bichier, P., 2004.
Conservation policy in coffee landscapes. Science 303, 625.
Eakin, H., Tucker, C., Castellanos, E., 2006. Responding to the coffee crisis:
a pilot study of farmers’ adaptations in Mexico, Guatemala and Hon-
duras. Geogr. J. 172, 156–171.
Galindo-Gonzalez, J., Guevara, S., Sosa, V.J., 2000. Bat- and bird-generated
seed rains at isolated trees in pastures in a tropical rainforest. Conserv.
Biol. 14, 1693–1703.
Gallina, S., Mandujano, S., Gonzalez-Romero, A., 1996. Conservation of
mammalian biodiversity in coffee plantations of Central Veracruz,
Mexico. Agroforest. Syst. 33, 13–27.
Gobbi, J.A., 2000. Is biodiversity-friendly coffee financially viable? An
analysis of five different coffee production systems in Western El
Salvador. Ecol. Econ. 33, 267–281.
Gordon, J.E., Barrance, A.J., Schreckenberg, K., 2003a. Are rare species
useful species? Obstacles to the conservation of tree diversity in the dry
forest zone agro-ecosystem of Mesoamerica. Global Ecol. Biogeogr.
12, 13–19.
Gordon, J.E., Hawthorne, W.D., Reyes-Garcıa, A., Sandoval, G., Barrance,
A.J., 2004. Assesing landscapes: a case study of tree and shrub diversity
in the seasonally dry tropical forests of Oaxaca, Mexico and southern
Honduras. Biol. Conserv. 117, 429–442.

A.M. Lopez-Gomez et al. / Agriculture, Ecosystems and Environment 124 (2008) 160–172172
Gordon, J.E., Hawthorne, W.D., Sandoval, G., Barrance, A.J., 2003b. Trees
and farming in the dry zone of southern Honduras II: the potential for
tree diversity conservation. Agroforest. Syst. 59, 107–117.
Gordon, C., Manson, R.H., Sundberg, J., Cruz Angon, A., 2007. Biodi-
versity, profitability, and vegetation structure in coffee agroecosystems
of central Veracruz, Mexico. Agric. Ecosyst. Environ. 118, 256–266.
Greenberg, R., Bichier, P., Sterling, J., 1997. Bird populations in rustic and
planted shade coffee plantations of Eastern Chiapas, Mexico. Biotropica
29, 501–514.
Kindt, R., Simons, A.J., vanDamme, P., 2004. Do farm characteristics
explain differences in tree species diversity among Western kenyan
farms? Agroforest. Syst. 63, 63–74.
Klein, A.M., Steffan-Dewenter, I., Tscharntke, T., 2003. Fruit set of high-
land coffee increases with the diversity of pollinating bees. Proc. R. Soc.
Lond. (Ser. B) 270, 955–961.
Komar, O., 2006. Ecology and conservation of birds in coffee plantations: a
critical review. Bird Conserv. Int. 16, 1–23.
Lambin, E.F., Geist, H.J., Lepers, E., 2003. Dynamics of land-use and land-
cover change in tropical regions. Ann. Rev. Environ. Res. 28, 205–241.
Mas, A.H., Dietsch, T.V., 2004. Linking shade coffee certification to
biodiversity conservation: butterflies and birds in Chiapas, Mexico.
Ecol. Appl. 14, 642–654.
Mayfield, M.M., Boni, M.F., Daily, G.C., Ackerley, D., 2005. Species and
functional diversity of native and human-dominated plant communities.
Ecology 86, 2365–2372.
Mendez, V.E., Gliessman, S.R., Gilbert, G.S., 2007. Tree biodiversity in
farmer cooperatives of a shade coffee landscape western El Salvador.
Agric. Ecosyst. Environ. 119, 145–159.
Moguel, P., Toledo, V.M., 1999. Biodiversity conservation in traditional
coffee systems of Mexico. Conserv. Biol. 13, 11–21.
Nestel, D., 1995. Coffee in Mexico: internacional market, agricultural
landscape and ecology. Ecol. Econ. 15, 165–178.
Nunez, 1987. El agroecosistema cafetalero en tres ejidos de la Costa Grande
de Guerrero. Undergraduate thesis (Biology). Universidad Nacional
Autonoma de Mexico. Mexico, D.F. p. 61.
O’Brien, T.G., Kinnaird, M.F., 2003. Caffeine and conservation. Science
300, 587.
O’Brien, T.G., Kinnaird, M.F., 2004. Conservation policy in coffee land-
scapes—response. Science 303, 625–626.
Peeters, L.Y.K., Soto-Pinto, L., Perales, H., Montoya, G., Ishiki, M., 2003.
Coffee production, timber, and firewood in traditional and Inga-shaded
plantations in Southern Mexico. Agric. Ecosyst. Environ. 95, 481–493.
Perfecto, I., Mas, A.H., Dietsch, T., Vandermeer, J., 2003. Species richness
along an agricultural intensification gradient: A tri-taxa comparison in
shade coffee in southern Mexico. Biodivers. Conserv. 12, 1239–1252.
Perfecto, I., Rice, R., Greenberg, R., Van der Voort, M.E., 1996. Shade
coffee: a disappearing refuge for biodiversity. Bioscience 46, 598–608.
Perfecto, I., Vandermeer, J., 2002. Quality of agroecological matrix in a
tropical montane landscape: ants in coffee plantations in Southern
Mexico. Conserv. Biol. 16, 174–182.
Philpott, S.M., Dietsch, T., 2003. Coffee and conservation: a global context
and the value of farmer involvement. Conserv. Biol. 17, 1844–1846.
Pineda, E., Moreno, C., Escobar, F., Halffter, G., 2005. Frog, bat, and dung
beetle diversity in the cloud forest and coffee agroecosystems of
Veracruz, Mexico. Conserv. Biol. 19, 400–410.
Potvin, C., Owen, C.T., Melzi, S., Beaucage, P., 2005. Biodiversity
and modernization in four coffee-producing villages of Mexico.
Ecol. Society 10, 18 [online] URL:http://www.ecologyandsociety.org/
vol10/iss1/art18/.
Rappole, J.H., King, D.I., Vega, J.H., 2003a. Coffee and conservation.
Conserv. Biol. 17, 334–336.
Rappole, J.H., King, D.I., Vega, J.H., 2003b. Coffee and conservation III:
reply to Philpott and Dietsch. Conserv. Biol. 17, 1847–1849.
Rendon, A., Turrubiarte B.N., 1985. El cultivo del cafe: caracterizacion del
manejo y estructura de cuatro huertos en el ejido ‘‘El Quemado’’
Municipio de Atoyac de Alvarez, Guerrero. Undergraduate thesis (Biol-
ogy). Universidad Nacional Autonoma de Mexico. Mexico, D.F. p. 183.
Ricketts, T.H., Daily, G.C., Ehrlich, P.R., Fay, J.P., 2001. Countryside
biogeography of moths in a fragmented landscape: biodiversity in
native and agricultural habitats. Conserv. Biol. 15, 378–388.
Romero-Alvarado, Y., Soto-Pinto, L., Garcıa-Barrios, L., Barrera-Gaytan,
J.F., 2002. Coffee yields and soil nutrients Ander the shades of Inga sp.
vs. multiple species in Chiapas, Mexico. Agroforest. Syst. 54, 215–224.
SAGARPA 2007. Sistema de Informacion Agroalimentaria de Consulta,
Secretariat of Agriculture, Cattle, Rural Development, Fish and Food
(http://www.siap.sagarpa.gob.mx/ar_comanuar.html).
SAS, 1997. JMP User’s Guide. SAS Institute, Cary, North Carolina.
Schroth, G., Fonseca, G.A.B., Harvey, C.A., Gascon, C., Vasconcelos,
H.L., Izac, A.-M.N. (Eds.), 2004. Agroforestry and Biodiversity
Conservation in Tropical Landscapes. Island Press, Washington, D.C,
523 p.
Solis-Montero, L., Flores-Palacios, A., Cruz-Angon, A., 2005. Shade-coffee
plantations as refuges for tropical wild orchids in central Veracruz,
Mexico. Conserv. Biol. 19, 908–916.
Sosa, V., Gomez-Pompa, A., 1994. Flora de Veracruz. Lista Florıstica.
Fascıculo 82. Instituto de Ecologıa, A.C., Xalapa, Veracruz/University
of California, Riverside, California.
Soto-Pinto, L., Perfecto, I., Castillo, J., Caballero-Nieto, J., 2000. Shade
effect on coffee production at the Northern Tzeltal zone of the state of
Chiapas, Mexico. Agric. Ecosyst. Environ. 80, 61–69.
Soto-Pinto, L., Romero-Alvarado, Y., Caballero-Nieto, J., Segura, G., 2001.
Woody plant diversity and structure of shade-grown-coffee plantations
in Northern Chiapas, Mexico. Rev. Biol. Trop. 49, 977–987.
Soto-Pinto, L., Villalvazo-Lopez, V., Jimenez-Ferrer, G., Ramırez-Marcial,
N., Montoya, G., Sinclair, F.L., 2007. The role of local knowledge in
determining shade composition of multistrata coffee systems in Chia-
pas, Mexico. Biodivers. Conserv. 16, 419–436.
Steffan-Dewenter, I., Kessler, M., Barkmann, J., Bos, M.M., Buchori, D.,
Erasmi, S., Faust, H., Gerold, G., Glenk, K., Gradstein, S.R., Guhardja,
E., Harteveld, M., Herteld, D., Hohn, P., Kappas, M., Kohler, S.,
Leuschner, C., Maertens, M., Marggraf, R., Migge-Kleian, S., Mogea,
J., Pitopang, R., Schaefer, M., Schwarze, S., Sporn, S.G., Steingrebe, A.,
Tjitrosoedirdjo, S.S., Tjitrosoemito, S., Twele, A., Weber, R., Wolt-
mann, L., Zeller, M., Tscharntke, T., 2007. Tradeoffs between income,
biodiversity, and ecosystem functioning during tropical rainforest con-
version and agroforestry intensification. Proc. Natl. Acad. Sc. U.S.A.
104, 4973–4978.
Tejeda-Cruz, C., Sutherland, W.J., 2004. Bird responses to shade coffee
production. Anim. Conserv. 7, 169–179.
Vandermeer, J., Perfecto, I., 2007. The agricultural matrix and a future
paradigm for conservation. Conserv. Biol. 21, 274–277.
Villalvazo-Lopez, V.M., 2002. Especies de sombra y criterios locales de
seleccion en un sistema agroforestal de cafe en Chiapas, Mexico. M.Sc.
thesis, El Colegio de la Frontera Sur. San Cristobal de las Casas,
Chiapas. p. 23.
Williams-Guillen, K., McCann, C., Martınez-Sanchez, J.C., Koontz, F.,
2006. Resource availability and habitat use by mantled howling mon-
keys in a Nicaraguan coffee plantation: can agroforests serve as core
habitat for a forest mammal? Anim. Conserv. 9, 331–338.
Williams-Linera, G., 2002. Tree species richness complementarity, distur-
bance and fragmentation in a Mexican tropical montane cloud forest.
Biodivers. Conserv. 11, 1825–1843.
Williams-Linera, G., Lopez-Gomez, A.M., Muniz-Castro, M.A., 2005.
Complementariedad y patrones de anidamiento de especies de arboles
en el paisaje de bosque de niebla del centro de Veracruz (Mexico). In:
Halffter, G., Soberon, J., Koleff P., Melic, A., (Eds.), Sobre diversidad
biologica: el significado de las diversidades alfa, beta y gamma. pp.
153–164. m3m-Monografıas Tercer Milenio, Vol 4. SEA, CONABIO,
Grupo DIVERSITAS,CONACYT. Zaragoza, Spain.
Williams-Linera, G., Manson, R.H., Isunza Vera, E., 2002. La fragmenta-
cion del bosque mesofilo de montana y patrones de uso del suelo en la
region oeste del Xalapa, Veracruz, Mexico. Madera y Bosques 8, 69–85.
Wunderle, J.M., Latta, S.C., 1998. Avian resource use in Dominican shade
coffee plantations. Wilson Bull. 110, 271–281.
Copyright © 2022 FDOKUMEN




















