
The role of the left temporal region under the cognitive motor demands of shooting in skilled...
15
Biological Psychology 58 (2001) 263 – 277 The role of the left temporal region under the cognitive motor demands of shooting in skilled marksmen Scott E. Kerick a,1 , Kaleb McDowell a,b,2 , Tsung-Min Hung c , D. Laine Santa Maria a , Thomas W. Spalding d , Bradley D. Hatfield a,b, * a Department of Kinesiology, Uniersity of Maryland, College Park, MD 20742, USA b Program in Neuroscience and Cognitie Science, Uniersity of Maryland, College Park, MD 20742, USA c Graduate Institute of Sport and Exercise Science, Taipei Physical Education College, No 5, Tun -Hua N Rd, Taipei, Taiwan, ROC d Department of Public and Community Health, Uniersity of Maryland, College Park, MD 20742, USA Received 30 January 2001; received in revised form 30 April 2001; accepted 31 May 2001 Abstract A number of investigators have reported elevated left temporal alpha power in marksmen during response preparation. This finding has been interpreted to indicate the suppression of irrelevant cognitive processes. However, lower-order motor processes have not been excluded as a possible explanation. Event-related alpha power (11 – 13 Hz) was examined at sites T3, T4, C3, and C4 in eight skilled marksmen during shooting and two control tasks varying in perceptual-motor complexity. Over an 8-s period preceding the trigger pull, the marksmen exhibited higher power and slope at T3 than at all other sites during shooting compared with the control conditions. No such difference between conditions was detected at C3 and C4. The relative synchrony of left temporal alpha power during shooting, in conjunction with the lack of change at central sites, is inconsistent with the explanation that the effect is accounted www.elsevier.com/locate/biopsycho Partial support provided by the US Army Research Institute for the Behavioral and Social Sciences via contract MDA903-93-C-0054. * Corresponding author. Tel.: +1-301-405-2485; fax: +1-301-405-5578. E-mail address: [email protected] (B.D. Hatfield). 1 Co-first author. 2 Co-first author. 0301-0511/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved. PII:S0301-0511(01)00116-8
Transcript of The role of the left temporal region under the cognitive motor demands of shooting in skilled...

Biological Psychology 58 (2001) 263–277
The role of the left temporal region under thecognitive motor demands of shooting in skilled
marksmen�
Scott E. Kerick a,1, Kaleb McDowell a,b,2, Tsung-Min Hung c,D. Laine Santa Maria a, Thomas W. Spalding d,
Bradley D. Hatfield a,b,*a Department of Kinesiology, Uni�ersity of Maryland, College Park, MD 20742, USA
b Program in Neuroscience and Cogniti�e Science, Uni�ersity of Maryland, College Park,MD 20742, USA
c Graduate Institute of Sport and Exercise Science, Taipei Physical Education College, No 5,Tun-Hua N Rd, Taipei, Taiwan, ROC
d Department of Public and Community Health, Uni�ersity of Maryland, College Park,MD 20742, USA
Received 30 January 2001; received in revised form 30 April 2001; accepted 31 May 2001
Abstract
A number of investigators have reported elevated left temporal alpha power in marksmenduring response preparation. This finding has been interpreted to indicate the suppression ofirrelevant cognitive processes. However, lower-order motor processes have not been excludedas a possible explanation. Event-related alpha power (11–13 Hz) was examined at sites T3,T4, C3, and C4 in eight skilled marksmen during shooting and two control tasks varying inperceptual-motor complexity. Over an 8-s period preceding the trigger pull, the marksmenexhibited higher power and slope at T3 than at all other sites during shooting compared withthe control conditions. No such difference between conditions was detected at C3 and C4.The relative synchrony of left temporal alpha power during shooting, in conjunction with thelack of change at central sites, is inconsistent with the explanation that the effect is accounted
www.elsevier.com/locate/biopsycho
� Partial support provided by the US Army Research Institute for the Behavioral and Social Sciencesvia contract MDA903-93-C-0054.
* Corresponding author. Tel.: +1-301-405-2485; fax: +1-301-405-5578.E-mail address: [email protected] (B.D. Hatfield).1 Co-first author.2 Co-first author.
0301-0511/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved.
PII: S0301 -0511 (01 )00116 -8

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277264
for by ‘lower-order’ motor processes exclusively involving the central region. © 2001 ElsevierScience B.V. All rights reserved.
Keywords: EEG; Event-related alpha power; Marksmanship
1. Introduction
The investigation of physiological response patterns can provide valuable insightinto the real-time psychological processes associated with skilled motor perfor-mance. Self-paced motor tasks like target shooting have frequently been employedto examine such processes because of a number of methodological advantages overother tasks. Target shooting is an attentionally engaging task characterized by arelatively motionless state that enables psychophysiological recording. A majorfinding from research on participants of target shooting is that elevated lefttemporal electroencephalographic (EEG) alpha power is associated with skilledmarksmanship. Hatfield et al. (1984) originally observed that broadband alphapower (8–12 Hz) increased over the left temporal region of skilled marksmenduring the 7.5 s preceding the shot, while right temporal alpha power remainedrelatively constant over the same period. In a comparison between expert marks-men and novice shooters, Haufler et al. (2000) observed higher levels of lefthemispheric alpha II power (10–11 Hz) during a 6-s period preceding the triggerpull in the experts compared with novices, with the greatest differences occurring inthe temporal region. Further evidence of the relevance of left temporal alphaactivity to target shooting skill was provided in a longitudinal study of archerystudents (Landers et al., 1994). The main findings were those of an increase in left,but no change in right, temporal alpha power (10–12 Hz) during the aiming periodfollowing 14 weeks of instruction and practice compared with a pre-test assessment.It should be noted, however, that skilled shooters’ best trials are associated with lessthan maximal levels of alpha power implying a point of optimal activity and thepossibility of an inverted-U relationship between alpha power and performance(Hillman et al., 2000; Salazar et al., 1990). In summary, the existing evidencesuggests that appropriate synchronization of EEG alpha power over the lefttemporal region serves a significant and adaptive role in the orchestration ofvisual-spatial and motor processes during target shooting.
A number of investigators have inferred that heightened left temporal alphaactivity is related to the attenuation of covert verbal-analytical behavior (e.g.self-talk) and attentional focus on the perceptual-motor demands of shooting(Hatfield et al., 1984; Haufler et al., 2000; Landers et al., 1994; Lawton et al., 1998).This interpretation is derived primarily from two lines of research. First, cognitiveneuroscience studies have provided evidence through ablation and neuroimagingmethods that the left temporal region is associated with verbal-analytical processes(i.e. language processing and working memory, see Cohen, 1993). Second, themotor behavior literature suggests a reduction in such covert verbal processes (Fittsand Posner, 1967) and self-evaluative thoughts during highly skilled performances

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277 265
(Jackson, 1996; Ravizza, 1977; Williams and Krane, 1993). This reasoning isconsistent with the idea that the verbal-analytical demands of target shooting, andthe related verbal processing functions of the left temporal region, are likely to beminimal during skilled shooting in light of the achievement of behavioral auto-maticity. Several investigators have suggested that a reduction in regional corticalprocessing during task-specific engagement is related to inhibitory mechanisms(Kinsbourne, 1982; Pfurtscheller, 1992; Pfurtscheller et al., 1996; Smith et al., 1999).Thus, a reduction in verbal-analytical processes, at the neuronal level, may beaccounted for by inhibitory mechanisms in the left temporal region. Considerationof the aforementioned studies implies that those cognitive processes that are notessential to the task of shooting are inhibited in skilled performers so as not toconflict with response preparation (Hatfield et al., 1984; Haufler et al., 2000;Landers et al., 1994; Lawton et al., 1998). More specifically, engagement inverbal-analytical processes during actual performance may interfere with, or com-pete for, the neural resources required to optimally execute the shot. The inhibitionof such processes may function to facilitate the achievement of a stable responsepattern and thereby decrease the variability of the performance. The hypothesisthat covert verbal-analytical processes become inhibited as attention shifts to moretask-relevant processes during shooting is hereafter referred to as verbal-suppression.
Although the verbal-suppression account is consistent with observations ofelevated left temporal alpha power during skilled shooting (Hatfield et al., 1984;Haufler et al., 2000; Landers et al., 1994; Lawton et al., 1998), this response couldalso reflect processes associated with the motor demands. For example, the re-sponse could be indicative of volume conduction from established primary and/orsecondary motor areas or from low-frequency electromyographic activity (or har-monics of high-frequency activity) from the neck or facial muscles associated withthe shooting posture. Although few studies have attempted to test such competingexplanations, Salazar et al. (1990) did examine whether elevation of left temporalalpha power during target shooting was related to motor processes associated withthe muscular demands in a group of highly skilled archers. They found that lefttemporal alpha power (10–12 Hz) was significantly higher during actual targetshooting compared with a control condition in which the archers were required todraw a bow of equal draw weight, but not to aim or shoot. They concluded that theincreased alpha power during shooting was related to cognitive as opposed tomotor processes. However, the main comparative condition did not entirely excludethe potential influence of motor processes. In this regard, the comparison conditiondescribed above required the participants to draw the bow, but did not requirearrow release, which requires a relatively small yet distinct motor act. The actualshot execution would seem to require greater motor precision and integrative motorprocessing compared with the control condition. Additionally, volume conductionfrom adjacent cortical areas associated with motor control (i.e. somatomotorcortex) may have influenced the recordings at the temporal sites and Salazar et al.did not monitor activity at the central sites to assess the similarity of the EEGrecordings. Konttinen and Lyytinen (1993) also investigated motor and aiming

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277266
component processes in a group of sharpshooters by analyzing preparatory slowpotentials (SPs) in frontal, central, and occipital regions. However, because of thesite topography examined (Fz, C3, C4, and Oz) and the EEG measure analyzed(SP), their work is limited in terms of furthering our understanding of thefunctional basis of left temporal alpha synchronization during shooting. In light ofthese limitations, there is a need to further examine the potential influence of motorprocesses on left temporal alpha power during skilled shooting.
Observations of elevated left temporal alpha power may be related to (1)‘lower-order’ motor control (i.e. processes such as those occurring in the primarymotor area that are associated with the activation of muscles); (2) verbal-suppres-sion; and/or (3) ‘higher-order’ sensorimotor processes during the preparatoryperiod of shooting in skilled marksmen (i.e. processes that are relevant to theintegration of visual, somatosensory, vestibular, and motor aspects; see Mulhol-land, 1995). In an attempt to reduce the number of possible explanations, thepresent study sought to determine whether the earlier observed increase in lefttemporal alpha power is related to lower-order motor control. To address this issue,EEG alpha power during shooting and two control conditions was examined. Inthe control conditions, the postural and movement demands were similar to thoseduring shooting (i.e., required a trigger pull in one condition) but aiming demandswere minimized (see Section 2). Furthermore, several design and methodologicalimprovements were incorporated in an attempt to achieve a more accurate repre-sentation of the underlying processes in the left temporal region. First, activity overthe somatomotor region (C3, C4) was recorded and the spatial resolution in thatarea was improved by applying an estimation of the Laplacian operator tosupplement Cz-referenced data (Hjorth, 1975). Second, alpha band power wasexamined with greater specificity by independently examining lower (alpha I, 8–10Hz) and upper (alpha II, 11–13 Hz) alpha. An increase in alpha I power has beenshown to be associated with a generalized, or non-specific, increase in attentionalprocessing during task engagement, whereas alpha II is associated with moretask-specific attentional processing (Haufler et al., 2000; Klimesch, 1999;Pfurtscheller and Lopes da Silva, 1999; Smith et al., 1999). Third, the temporalresolution of alpha power estimates was improved to potentially reveal earlierobscured patterns in the time preceding the trigger pull. Finally, because of ourspecific interest in alpha II power over the left temporal region, absolute valueswere reported for each site rather than relative asymmetry indices between ho-mologous left and right hemisphere sites. Attempting to segregate the componentprocesses involved in shooting is difficult due to the increased motor precisiondemands required during actual target shooting relative to the comparative controlconditions. However, inclusion of the triggering action in one of the controlconditions, in addition to increased temporal and spatial resolution of the EEGmeasures, may help to provide valuable insight into the processes related to lefttemporal EEG activity during skilled target shooting.
Two lines of evidence would be needed to support the idea that left temporalalpha activity is related to either verbal-suppression or higher-order sensorimotorprocesses, and not to lower-order motor control processes. First, alpha II power

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277 267
should be greater in the left temporal region (T3) during the shooting conditionrelative to the motor control conditions. Second, the pattern of differences in lefttemporal alpha II power across conditions should be different from that observedin the somatomotor region (C3, C4) in terms of both mean levels and temporaldynamics, particularly when analyzed with increased spatial resolution (see Section2). Alpha II power was also examined in the right temporal region (T4) but was notexpected to differ across conditions. Additionally, because lower alpha power hasbeen shown to be less sensitive to task-specific demands, alpha I power was alsoexamined but not expected to differ between conditions.
2. Methods
2.1. Participants
Eight right-handed and right-eye-dominant skilled marksmen volunteered toparticipate. The age range of the participants was 20–32 year and formal shootingexperience ranged from 5 to 12 years. All participants were screened for neurolog-ical disorders and were asked not to ingest nicotine or caffeine at least 4 h prior todata collection.
2.2. Procedures
Informed consent was obtained from all participants in accord with the institu-tional review board guidelines. Testing was conducted at a regulation indoorshooting range and the participants were equipped with a .22-caliber competitionrifle. A small microphone was attached to the rifle barrel, which served as anevent-marker for the trigger pull (see Haufler et al., 2000 for details). A standardsmall-bore target (A-17) was placed 50 ft downrange at eye-level. After beingprepared for EEG acquisition, the participants were allowed 5–10 sighting trialsand then completed postural control (PC), movement control (MC), and shooting(SH) conditions in a randomized order. For all three conditions, the participantswere asked to assume a standard shooting posture from the standing position (i.e.with the feet approximately shoulder-width apart and perpendicular to the shootinglane). From this position, the rifle was raised in the direction of the target and wassupported by the left hand. The elbow of the left arm was braced against the leftside of the body to help stabilize the gun hold. The butt of the rifle was held firmlyin the anterior right shoulder region with the right hand and the trigger wasoperated with the right index finger (in the MC and SH conditions).
In the PC condition, the participants were instructed to raise the rifle to theshooting position and hold it stable continuously for 90 s. They were asked to lookstraight ahead, rather than focus on aligning the sights with the target, and not topull the trigger at any time. Since the PC condition did not require an event-relatedresponse, a relatively longer hold period was implemented to provide a morestationary, non-anticipatory comparison condition. The instructions for the MC

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277268
condition were similar to those for the PC condition except an event-relatedresponse was implemented by instructing the participants to pull the trigger(dry-fire) after a self-estimated 8-s period in the MC condition. In an attempt tocontrol for covert verbal processing during the MC condition, the participants wereinstructed not to internally count down the 8 s, but to approximate the duration ofthe hold based on their typical hold duration during normal shooting3. Forty trialswere repeated in the MC condition. In the SH condition, the participants wereasked to shoot at the target as they normally would in practice or competition andan 8 s period preceding each trigger pull was extracted for subsequent analyses (i.e.a fully integrated natural response). Forty trials were repeated in the SH condition.Thus, the PC condition closely simulated the gross motor demands of shooting (interms of maintaining a stable posture and gun hold) but it did not require preciseaiming processes. The MC condition incorporated the fine motor act of squeezingthe trigger but it did not require coincident timing at a precise optimal moment (interms of gun stabilization and alignment with aiming processes). The SH conditionrequired the integration of these general motor components with aiming processesand increased motor precision. Table 1 describes relative differences between thethree tasks with respect to accuracy, stability, and timing of the motor demands.
2.3. EEG acquisition
A stretchable lycra electrode cap (Electro-Cap Inc, Eaton, OH) was fitted to theparticipants for scalp recordings of EEG in accord with the standard international10–20 system (Jasper, 1958). The montage consisted of left and right central (C3,C4) and temporal (T3, T4) sites with Cz as the common reference. Eight additionalsites were included to allow for offline re-referencing of C3 and C4 by Laplacianderivation (Hjorth, 1975). The Hjorth Laplacian interpolation uses four nearest-neighbor electrodes situated equidistantly (2.5 cm) from the site of interest and atright angles to each other in an attempt to isolate electrocortical activity to a localregion under the site of interest. One limitation of the Laplacian derivation is thatperipheral sites (T3, T4) cannot be accurately referenced (Pfurtscheller, 1988).
Table 1Relative motor demands of shooting tasks
Condition Motor demand
Accuracy Stability Timing
NoneLow LowPCLow LowLowMC
High HighSH High
3 Variation in the intervals of time between raising the rifle and triggering during this conditionsupported that the participants adhered to the instructions and did not count down the 8 s. If theparticipant pulled the trigger before 8 s had elapsed the trial was repeated.

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277 269
Therefore, Laplacian-referenced data were not derived for the temporal regions.Eye-movement (EOG) artifact was monitored with a bipolar electrode montageusing two 10-mm diameter electrodes (Grass E5GH, gold plated) attached superiorto and on the external canthus of the right eye. Amplifiers were calibrated for eachparticipant separately through source input of a 50-�V 10-Hz sinusoidal wave at allchannels simultaneously. EEG and EOG signals were sampled at 256 Hz andamplified 50,000 (5000 for EOG) times using a 12-bit Grass model 12A5 NeurodataAcquisition system with band pass filter settings from 0.1 to 100 Hz. Electrodeimpedance values were between 5 and 10 K� for all recordings. Continuous EEGwas acquired using Neuroscan software (Ver. 4.0) and the recorded data were savedto optical disk.
2.4. Data reduction
Due to the absence of an event-related response in the PC condition, thecontinuous EEG time series was assumed to be stationary and, subsequently, wasepoched offline for comparable analyses with the epoched data from the SH andMC conditions. For the PC condition, the continuous 90-s time series was epochedinto 17 10-s windows with successive 50% overlapping sections. Epochs containingeye-movement artifact or amplitudes exceeding 75 �V were excluded from furtheranalyses. Artifact-free epochs were then filtered forward and backward using anarrow 2-Hz band pass filter (96 dB roll-off) centered at lower (8–10 Hz) and upper(11–13 Hz) alpha frequencies (Pfurtscheller and Lopes da Silva, 1999). The filtered10-s epochs were then squared and averaged over 125-ms segments (32 samples) tosmooth the data. One-second segments were cut from the beginning and end ofeach 10-s epoch to remove the edge effects of filtering. The remaining 8-s epochswere then ensemble-averaged, which reduced each channel of data to an averaged8-s power-time curve. This methodology allows for a trend analysis of the temporalchanges but, because a linear trend appeared to best represent the data, first-orderregression was used to model a line-of-fit by least-squares criteria. Finally, theslopes and intercepts of the line were derived for each site and then subjected tostatistical analyses. The intercept represents an interpolation of the mean level ofalpha power at the time of the trigger pull (hereafter referred to as mean alphapower), whereas the slope represents the average rate of change in alpha power overthe 8 s prior to the trigger pull. For the MC and SH conditions, the continuous rawdata were epoched into 10-s windows (−9 to 1 s) around the trigger pull for eachof the 40 trials and then inspected for artifact. Epochs containing eye-movementartifact or amplitudes exceeding 75 �V were rejected from further analyses. Arti-fact-free epochs were then processed using the same procedures as applied to thedata in the PC condition.
Traditionally, the changes associated with the preparatory period of shootinghave been assessed with temporal resolutions on the order of 1 s by Fast-Fouriertransform. By employing signal processing techniques that use digital filters toisolate narrow frequency bands (Hari et al., 1997; Pfurtscheller and Aranibar,1977), the temporal resolution of spectral estimates can be improved to as fine as 1

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277270
ms. The method outlined herein provides a power-time curve comprised of 64estimates per trial (eight samples per s×8 s) with 125 ms resolution. The percent-age of accepted epochs was 88, 78, and 81% for the PC, MC, and SH conditions,respectively.
2.5. Design and analyses
Mean alpha I and II power and slope estimates were analyzed in separate 3×4(condition×site) ANOVAs with repeated-measures on both factors for the Cz-ref-erenced data. Significant omnibus tests were followed by paired t-tests to examinedifferences at each site across conditions and between T3 and each of the other siteswithin conditions. Two separate one-factor ANOVAs, with three levels for condi-tion, were applied to the Laplacian-referenced data for C3 and C4 because the lackof a common reference does not allow these sites to be directly compared.
3. Results
3.1. Alpha II
Fig. 1 is a plot of the means and S.D. of the alpha II power-time curves over the8-s periods. Table 2 provides the means and standard deviations of alpha I and IIpower and slope values (for both Cz- and Laplacian-referenced data).
3.1.1. PowerANOVA applied to the Cz-referenced alpha II mean power revealed a significant
condition×site interaction, F(2.26, 15.81)=5.45, P�0.02. Post hoc comparisonsbetween conditions revealed that alpha II power at sites T3 and T4 was significantlyhigher in the SH condition than in the PC and MC conditions which did not differ,while no differences in power were observed across conditions for sites C3 and C4.Furthermore, within all conditions, T3 power was significantly higher than ob-served at each of the other sites (see Table 3 for effect sizes). The separateone-factor ANOVAs applied to the Laplacian-referenced data also revealed thatneither C3 nor C4 differed across conditions.
3.1.2. SlopesThe analyses of slopes for the Cz-referenced data also revealed a condition×site
interaction, F(2.10, 14.72)=4.37, P�0.03. Post hoc analyses between conditionsrevealed that the slopes were significantly higher at T3 in the SH conditioncompared with the PC and MC conditions, but did not differ between the PC andMC conditions. The slope for T4 was higher in the SH versus the MC condition,but not versus the PC condition. Within conditions, T3 slope was significantlyhigher than all three of the other sites in the SH condition only. The one-factorANOVAs applied to the Laplacian-referenced data for C3 and C4 were alsononsignificant.
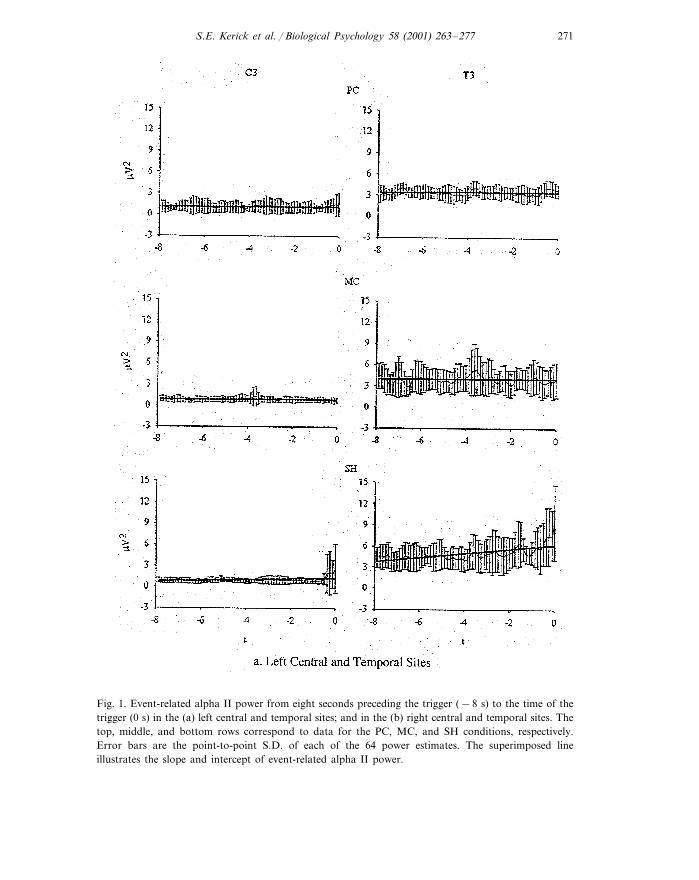
S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277 271
Fig. 1. Event-related alpha II power from eight seconds preceding the trigger (−8 s) to the time of thetrigger (0 s) in the (a) left central and temporal sites; and in the (b) right central and temporal sites. Thetop, middle, and bottom rows correspond to data for the PC, MC, and SH conditions, respectively.Error bars are the point-to-point S.D. of each of the 64 power estimates. The superimposed lineillustrates the slope and intercept of event-related alpha II power.
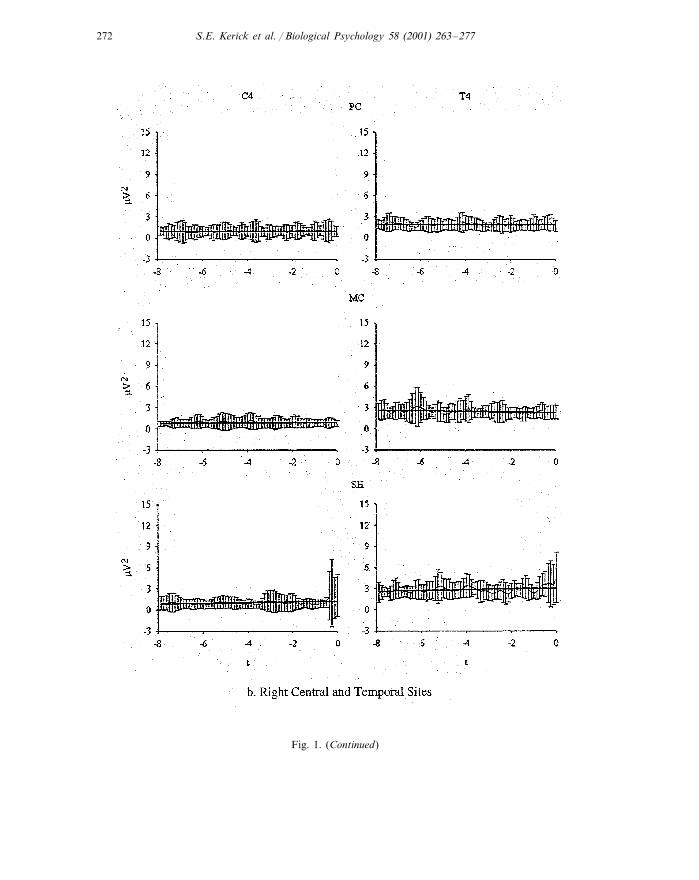
S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277272
Fig. 1. (Continued)
S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277 273
3.2. Alpha I
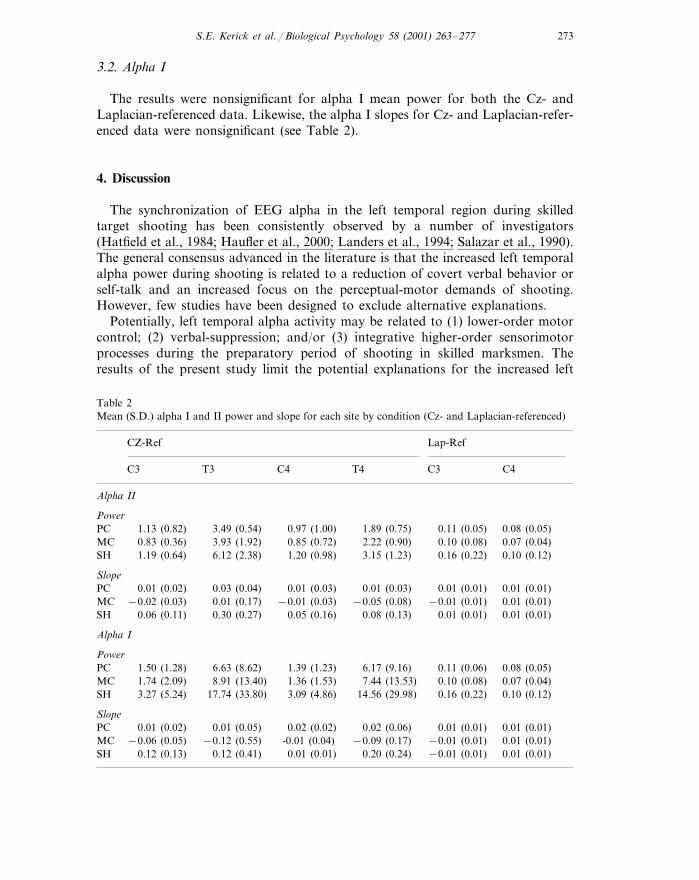
The results were nonsignificant for alpha I mean power for both the Cz- andLaplacian-referenced data. Likewise, the alpha I slopes for Cz- and Laplacian-refer-enced data were nonsignificant (see Table 2).
4. Discussion
The synchronization of EEG alpha in the left temporal region during skilledtarget shooting has been consistently observed by a number of investigators(Hatfield et al., 1984; Haufler et al., 2000; Landers et al., 1994; Salazar et al., 1990).The general consensus advanced in the literature is that the increased left temporalalpha power during shooting is related to a reduction of covert verbal behavior orself-talk and an increased focus on the perceptual-motor demands of shooting.However, few studies have been designed to exclude alternative explanations.
Potentially, left temporal alpha activity may be related to (1) lower-order motorcontrol; (2) verbal-suppression; and/or (3) integrative higher-order sensorimotorprocesses during the preparatory period of shooting in skilled marksmen. Theresults of the present study limit the potential explanations for the increased left
Table 2Mean (S.D.) alpha I and II power and slope for each site by condition (Cz- and Laplacian-referenced)
CZ-Ref Lap-Ref
T4 C3C3 C4T3 C4
Alpha II
Power1.89 (0.75)0.97 (1.00)3.49 (0.54) 0.08 (0.05)1.13 (0.82)PC 0.11 (0.05)2.22 (0.90) 0.10 (0.08)MC 0.07 (0.04)0.83 (0.36) 3.93 (1.92) 0.85 (0.72)
1.19 (0.64) 6.12 (2.38) 1.20 (0.98) 3.15 (1.23)SH 0.16 (0.22) 0.10 (0.12)
Slope0.01 (0.03)0.03 (0.04) 0.01 (0.01)0.01 (0.02)PC 0.01 (0.01)0.01 (0.03)
0.01 (0.01)−0.01 (0.01)−0.05 (0.08)−0.01 (0.03)MC 0.01 (0.17)−0.02 (0.03)0.08 (0.13) 0.01 (0.01)SH 0.01 (0.01)0.06 (0.11) 0.30 (0.27) 0.05 (0.16)
Alpha I
Power1.50 (1.28)PC 6.63 (8.62) 1.39 (1.23) 6.17 (9.16) 0.11 (0.06) 0.08 (0.05)1.74 (2.09) 0.10 (0.08) 0.07 (0.04)7.44 (13.53)MC 1.36 (1.53)8.91 (13.40)
3.09 (4.86)17.74 (33.80) 0.10 (0.12)3.27 (5.24) 0.16 (0.22)SH 14.56 (29.98)
Slope0.02 (0.06) 0.01 (0.01)PC 0.01 (0.01)0.01 (0.02) 0.01 (0.05) 0.02 (0.02)
−0.06 (0.05) -0.01 (0.04) −0.09 (0.17) −0.01 (0.01) 0.01 (0.01)MC −0.12 (0.55)0.12 (0.41) 0.01 (0.01) 0.20 (0.24) −0.01 (0.01) 0.01 (0.01)SH 0.12 (0.13)
S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277274
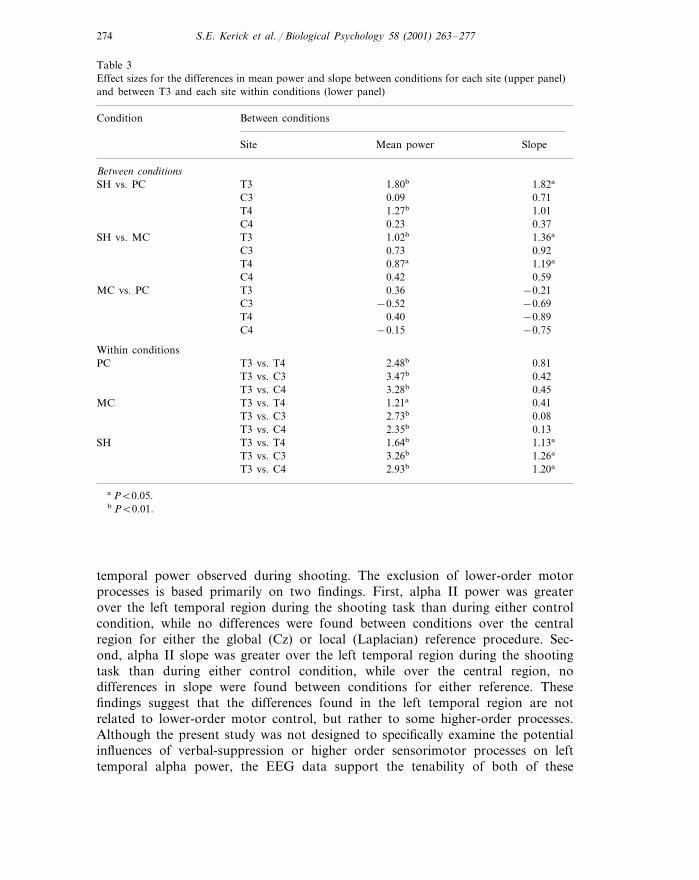
Table 3Effect sizes for the differences in mean power and slope between conditions for each site (upper panel)and between T3 and each site within conditions (lower panel)
Between conditionsCondition
Mean power SlopeSite
Between conditionsSH vs. PC 1.82aT3 1.80b
C3 0.09 0.711.01T4 1.27b
0.370.23C41.36aSH vs. MC T3 1.02b
0.920.73C31.19aT4 0.87a
0.590.42C4−0.21MC vs. PC T3 0.36−0.69−0.52C3
0.40T4 −0.89−0.75−0.15C4
Within conditionsPC 0.81T3 vs. T4 2.48b
0.423.47bT3 vs. C30.45T3 vs. C4 3.28b
0.411.21aT3 vs. T4MC0.08T3 vs. C3 2.73b
0.132.35bT3 vs. C4T3 vs. T4 1.64b 1.13aSH
1.26a3.26bT3 vs. C32.93bT3 vs. C4 1.20a
a P�0.05.b P�0.01.
temporal power observed during shooting. The exclusion of lower-order motorprocesses is based primarily on two findings. First, alpha II power was greaterover the left temporal region during the shooting task than during either controlcondition, while no differences were found between conditions over the centralregion for either the global (Cz) or local (Laplacian) reference procedure. Sec-ond, alpha II slope was greater over the left temporal region during the shootingtask than during either control condition, while over the central region, nodifferences in slope were found between conditions for either reference. Thesefindings suggest that the differences found in the left temporal region are notrelated to lower-order motor control, but rather to some higher-order processes.Although the present study was not designed to specifically examine the potentialinfluences of verbal-suppression or higher order sensorimotor processes on lefttemporal alpha power, the EEG data support the tenability of both of these

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277 275
accounts of cortical processing, which are not mutually exclusive. The currentfindings suggest that the increased synchronization of the left temporal regionduring skilled shooting is more strongly related to verbal-suppression and/orsensorimotor processing and less related to lower-order motor demands such asthe stance, gun hold, and trigger pull.
Several methodological improvements aided in the interpretation of the elec-trocortical activity associated with shooting. First, the improved temporal resolu-tion was instrumental in verifying the linear changes in alpha power across the 8s prior to triggering that had earlier been reported (see Fig. 1 Hatfield et al.,1984; Salazar et al., 1990). Second, the observed differences in alpha II but notalpha I bandwidths provided validation that shooting requires task-specific atten-tional processing above that required by the motor control conditions, however,general attentional processing was undifferentiated across conditions (Haufler etal., 2000; Klimesch, 1999; Pfurtscheller and Lopes da Silva, 1999; Smith et al.,1999). Third, the increased alpha power and slope at T3 relative to T4 duringshooting is consistent with earlier findings of asymmetry between the right andleft temporal regions (Hatfield et al., 1984; Haufler et al., 2000; Landers et al.,1994; Salazar et al., 1990). Fourth, the improved spatial resolution over thesomatomotor region allowed for greater localization of the underlying processesand aided in the interpretation that the activity observed over the left temporalregions was different from that over the central regions. Altogether, these find-ings provide further evidence that volume conduction from the central regionsdid not account for the observed differences in the left temporal region (Salazaret al., 1990). The greater temporal and spatial resolution of power estimates inspecific frequency bands provided valuable information that aided in the inter-pretation of the data. Future research may benefit from the employment of finertemporal-spatial resolution of the EEG metrics, when it is feasible to do so, interms of clarifying the assessment of such dynamic processes.
The present findings, along with those of others, support a robust effect ofsynchronous neural activity in the left temporal region associated with skilledmarksmanship. Neuroimaging studies on motor performance suggest that skillimprovement is associated with increased blood flow to the primary and supple-mentary motor cortex, basal ganglia, and cerebellum, and decreased neural activ-ity in less relevant cortical areas including the medial temporal region (seeGrafton et al., 2000; Ingvar and Petersson, 2000 for recent reviews). The syn-chronization observed at both of the temporal sites during the precision aimingperiod of shooting, relative to the control conditions, implies decreased activityin these areas. Several hypotheses have been proposed as to the basis for syn-chronization in temporal regions. According to Ingvar and Petersson (2000),activity decreases in the medial temporal area as performance on a motor taskreaches the level of automaticity and may be associated decreased working mem-ory and feedback demands. This perspective is consistent with the hypothesisthat the cortex can potentially be engaged in task-relevant and task-irrelevantprocessing, whereby task-irrelevant processes are inhibited or shut down during

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277276
the preparatory period of shooting through extended practice. The inhibition oftask-irrelevant processes may function to reduce interference with task-relevantsensory-motor processing (Pfurtscheller, 1992) and provides the basis for theverbal-suppression hypothesis (Hatfield et al., 1984). An alternative account ofthe synchronized activity during motor performance is that there are no task-ir-relevant processes, but that all cortical resources function in a complementarymanner to form a unitary pattern of behavior (Kinsbourne, 1982). According tothis view, activity within a specific cortical region, whether inhibited or excited,influences processing of other cortical regions (e.g., reciprocal inhibition betweencortical regions may provide a mechanism by which selective attention and be-havior are implemented). A difference between these two perspectives is whetherthe inhibited cortical area is actively or passively involved in the generation ofbehavioral responses. Of interest to note is the unexpected finding of increasedalpha II synchronization at T4 during shooting relative to the control conditions.However, the level of synchronization and the rate of alpha II power increaseacross the aiming period at T4 was significantly less than that observed at T3.These observations imply relative engagement of the right temporal area, a find-ing that is consistent with those of others who have examined EEG alpha duringthe performance of tasks that demand visuo-spatial processing (Smith et al.,1999). In light of the symbiotic relationship of the feature-detection and integra-tive processes of the left and right temporo–parietal areas, respectively, outlinedby Kinsbourne (1982) it may be that the synchronization of the left temporalarea during a visuo-spatial task like marksmanship is functionally related to thecortical activity observed in the right. Further examination of these processeswill ultimately lead to a better understanding of the general principles of corticalorganization associated with skilled human performance.
5. Conclusions
Synchronized alpha activity over the left temporal region has been associatedwith marksmanship skill and the time to trigger pull suggesting the importanceof the underlying processes during shooting. In other areas of research, the leftparietal– temporal–occipital pathway (PTO) has been associated with attention,working memory, visual-associative operations, auditory processing, language,memory consolidation, emotional-motivational processing, feature extraction ofspatial details, and the timing and sequencing of information (see Cohen, 1993).These findings indicate many possible functions of the left temporal region. Thisstudy attempted to eliminate one explanation for the synchronization of lefttemporal alpha activity by incorporating design and methodological refinements.The present results suggest that, while the pattern of activity in the left temporalregion was inconsistent with lower-order motor processes, it may be related toverbal-suppression processes and/or higher-order motor planning and controlduring the precision aiming period of target shooting.

S.E. Kerick et al. / Biological Psychology 58 (2001) 263–277 277
References
Cohen, R.A., 1993. The Neuropsychology of Attention. Plenum Press, New York.Grafton, S.T., Hari, R., Salenius, S., 2000. The human motor system. In: Toga, A.W., Mazziotta, J.C.
(Eds.), Brain Mapping: the Systems. Academic Press, San Diego, pp. 331–363.Fitts, P.M., Posner, M.I., 1967. Human Performance. Brookes/Cole Publishing Company, Belmont, CA.Hari, R., Salmelin, R., Makela, J.P., Salenius, S., Helle, M., 1997. Magnetoencephalographic cortical
rhythms. Int. J. Psychophysiol. 26, 51–62.Hatfield, B.D., Landers, D.M., Ray, W.J., 1984. Cognitive processes during self-paced motor perfor-
mance: an electroencephalographic profile of skilled marksmen. J. Sport Psychol. 6, 42–59.Haufler, A.J., Spalding, T.S., Santa Maria, D.L., Hatfield, B.D., 2000. Neuro-cognitive activity during
a self-paced visual-spatial task: comparative EEG profiles in marksmen and novice shooters. Biol.Psychol. 53, 131–160.
Hillman, C.H., Apparies, R.J., Janelle, C.M., Hatfield, B.D., 2000. An electrocortical comparison ofexecuted and rejected shots in skilled marksmanship. Biol. Psychol. 52, 71–83.
Hjorth, B., 1975. An on-line transformation of EEG scalp potentials into orthogonal source derivations.Electroencephalogr. Clin. Neurophysiol. 39, 526–530.
Ingvar, M., Petersson, K.M., 2000. Functional maps and brain networks. In: Toga, A.W., Mazziotta,J.C. (Eds.), Brain Mapping: the Systems. Academic Press, San Diego, pp. 111–139.
Jackson, S.A., 1996. Toward a conceptual understanding of flow experience in elite athletes. Res. Q.Exer. Sport 67, 76–90.
Jasper, H.H., 1958. The ten-twenty system of the International Federation. Electroencephalogr. Clin.Neurophysiol. 10, 371–375.
Kinsbourne, M., 1982. Hemispheric specialization and the growth of human understanding. Am.Psychol. 37, 411–420.
Klimesch, W., 1999. EEG alpha and theta oscillations reflect cognitive and memory performance: areview and analysis. Brain Res. Rev. 29, 169–195.
Konttinen, N., Lyytinen, H., 1993. Brain slow waves preceding time-locked visuo-motor performance. J.Sports Sci. 11, 257–266.
Landers, D.M., Han, M., Salazar, W., Petruzzello, S.J., Kubitz, K.A., Gannon, T.L., 1994. Effects oflearning on electroencephalographic and electrocardiographic patterns in novice archers. Int. J. SportPsychol. 25, 313–330.
Lawton, G.W., Hung, T.M., Saarela, P., Hatfield, B.D., 1998. Electroencephalography and mental statesassociated with elite performance. J. Sport Exer. Psychol. 20, 35–53.
Mulholland, T., 1995. Human EEG, behavioral stillness and biofeedback. Int. J. Psychophysiol. 19,263–279.
Pfurtscheller, G., 1988. Mapping of event-related desynchronization and type of derivation. Electroen-cephalogr. Clin. Neurophysiol. 70, 190–193.
Pfurtscheller, G., 1992. Event-related synchronization (ERS): an electrophysiological correlate of corticalareas at rest. Electroencephalogr. Clin. Neurophysiol. 83, 62–69.
Pfurtscheller, G., Aranibar, A., 1977. Event-related cortical desynchronization detected by powermeasurements of scalp EEG. Electroencephalogr. Clin. Neurophysiol. 42, 817–826.
Pfurtscheller, G., Lopes da Silva, F.H., 1999. Event-related EEG/MEG synchronization and desynchro-nization: basic principles. Clin. Neurophysiol. 110, 1842–1857.
Pfurtscheller, G., Stancak, A. Jr, Neuper, C., 1996. Event-related synchronization (ERS) in the alphaband-An electrophysiological correlate of cortical idling: a review. Int. J. Psychophysiol. 24, 39–46.
Ravizza, K., 1977. Peak experiences in sport. J. Human. Psychol. 17, 35–40.Salazar, W., Landers, D.M., Petruzzello, S.J., Crews, D.J., Kubitz, K.A., 1990. The effects of
physical/cognitive load on electrocortical responses preceding response execution in archery. Psycho-physiology 25, 478–479.
Smith, M.E., McEvoy, L.K., Gevins, A., 1999. Neurophysiological indices of strategy development andskill acquisition. Cogn. Brain Res. 7, 389–404.
Williams, J.M., Krane, V., 1993. Psychological characteristics of peak performance. In: Williams, J.M.(Ed.), Applied Sport Psychology: Personal Growth to Peak Performance. Mayfield, Mountain View,CA, pp. 137–147.




















