
ANTI-TUMOR ACTIVITY OF FRUIT EXTRACTS OF MOMORDICA DIOICA ROXB
Upload
independentCategory
view
3download
0
Cancer Letters 324 (2012) 66–74
Contents lists available at SciVerse ScienceDirect
Cancer Letters
journal homepage: www.elsevier .com/ locate/canlet
The MAP30 protein from bitter gourd (Momordica charantia) seeds promotesapoptosis in liver cancer cells in vitro and in vivo
Evandro Fei Fang a,1,2, Chris Zhi Yi Zhang b,c,1, Jack Ho Wong a, Jia Yun Shen d, Chuan Hao Li e,Tzi Bun Ng a,⇑a School of Biomedical Sciences, Faculty of Medicine, The Chinese University of Hong Kong, Shatin, Hong Kongb State Key Laboratory of Oncology in Southern China, Sun Yat-Sen University Cancer Center, Guangzhou, Chinac Department of Pathology, Sun Yat-Sen University Cancer Center, Guangzhou, Chinad Institute of Digestive Disease and Department of Medicine and Therapeutics, Li Ka Shing Institute of Health Sciences, The Chinese University of Hong Kong, Hong Konge Department of Chemistry, Faculty of Science, The Chinese University of Hong Kong, Hong Kong
a r t i c l e i n f o a b s t r a c t
Article history:Received 17 March 2012Received in revised form 28 April 2012Accepted 2 May 2012
Keywords:MAP30Bitter gourdMomordica charantiaHepatocellular carcinomaApoptosis
0304-3835/$ - see front matter � 2012 Elsevier Irelanhttp://dx.doi.org/10.1016/j.canlet.2012.05.005
⇑ Corresponding author.E-mail address: [email protected] (T
1 These authors contributed equally to this work.2 Present address: Laboratory of Molecular Geron
Aging, National Institutes of Health, Baltimore, MD, US
Human hepatocellular carcinoma Hep G2 cells and Hep G2-bearing mice were used as in vitro and in vivomodels to assess the efficacy and safety of MAP30, a natural component from Momordica charantia, as ananticancer agent against liver cancer. Molecular studies disclosed the contribution of both caspase-8 reg-ulated extrinsic and caspase-9 regulated intrinsic caspase cascades in MAP30-induced cell apoptosis. Theantitumor potential was also effective in Hep G2-bearing nude mice. Since bitter gourd is a staple inmany Asian countries, MAP30 would serve as a novel and relatively safe agent for prophylaxis and treat-ment of liver cancer.
� 2012 Elsevier Ireland Ltd. All rights reserved.
1. Introduction RNA which is necessary for the binding of elongation factors [7].
Bitter gourd/BG (binominal name Momordica charantia belong-ing to Family Cucurbitaceae) is widely distributed in Asia, Africa,and some places in South America [1,2]. Its bitter fruit is a commonand popular vegetable in Asian countries because of its rich tasteand nutritional value. Furthermore, BG is also applied in folkloremedicine to treat illnesses such as metabolic disorders and viralinfections [1]. In recent decades, a large number of nutritional/medicinal components in BG have been purified and characterized,such as cucurbitane-type triterpenoids, linolenic acids, potato- andsquash-type protease inhibitors, ribonucleases (RNase MC1 andRNase MC2), type 1 ribosome inactivating proteins/RIPs (napin-like RIP, momorcharins, and MAP30), and a type 2 RIP (M. charantialectin/MCL) [1,3,4]. The credited anti-diabetic, anti-HIV, and anti-tumor potentials of BG are at least partially attributed to the re-ported components [1,5,6].
RIPs are RNA glycosylases that cleave an adenine–riboseglycosidic bond within a conserved loop in the 28S ribosomal
d Ltd. All rights reserved.
.B. Ng).
tology, National Institute onA.
The representative RIP protein MAP30 was first purified by Lee-Huang and coworkers in 1990 [8] where in-depth mechanisticstudies of the anti-HIV activity of MAP30 have been carried out.MAP30 was active against infection and replication of both herpessimplex virus (HSV) [9] and human immunodeficiency virus (HIV)[10,11]. For example, MAP30 could inhibit the HIV-1 infection inT lymphocytes/monocytes and repress virus replication in cellsalready infected, both with mechanisms associated with impedi-ment of HIV viral DNA integration [10,11]. Furthermore, MAP30inhibited proliferation of AIDS-related lymphoma cells infectedwith Kaposi’s sarcoma-associated virus by modulation of differentviral and cellular genes necessary for viral and cell proliferationand apoptosis [12]. Intriguingly, recombinant MAP30 produced indifferent expression systems, such as E. coli and squash leaves, man-ifested comparable biological activities against HIV-1, HSV, andHHV8 viruses as its native counterpart [13,14]. The anti-HIV andanti-cancer properties may be attributed to its functions as a DNAglycosylase and/or DNA apurinic/apyrimidinic lyase [11], but pre-clude its RNA N-glycosidase (ribosome inactivating) activity [15].
Besides anti-HIV activity, initial screening studies disclosed thatMAP30 inhibited proliferation of tumor cells, such as brain glio-blastoma U87GM, breast carcinoma BT20, epidemoid carcinomaA431, melanoma Malme-3 M, myeloma U266, neuroblastomaSK-N-SH, prostate carcinoma DU145, and hepatoma Hep 3B cells

E.F. Fang et al. / Cancer Letters 324 (2012) 66–74 67
[14]. However, only few in-depth investigations reported themechanisms of action of the observed activities. Firstly, MAP30manifested antitumor activity toward estrogen-independent hu-man breast cancer MDA-MB-231 cells by down-regulating theexpression of HER2 in both in vitro and in vivo studies. Second,Fan and coworkers found that recombinant MAP30 inhibited cellproliferation and induced apoptosis in human colorectal carcinomaLoVo cells by up-regulation of Bax as well as down-regulation ofBcl-2 [16]. In this study, we have extended the application rangeof MAP30 to liver cancer. We observed that MAP30-induced apop-tosis of Hep G2 cells was mediated through activation of bothcaspase-8 and caspase-9 signaling pathways. The apoptosis-induc-ing activity of MAP30 was also significant in Hep G2-bearing nudemice.
2. Materials and methods
2.1. Reagents and antibodies
BG seeds were purchased in a local market and authenticated by Dr. Shiu-yingHu, Honorary Professor of Chinese Medicine at the Chinese University of HongKong. N-benzyloxycarbonyl-Val-Ala-Asp-fluoromethyl ketone (Z-VAD-FMK) waspurchased from Sigma, and the MAPK caspase inhibitors were provided byTocriset™ (1878). All abovementioned inhibitors were dissolved in dimethylsulfox-ide/DMSO (Sigma–Aldrich, USA). The final concentration of DMSO used (<0.01%)had no adverse effect on cell viability. The antibodies used in this study were as fol-lows: primary antibodies for Bid (sc-11423, polyclonal), p53 (sc-126, monoclonal),tubulin (sc-9104, polyclonal), PARP (sc-25780, polyclonal), cleaved PARP (sc-23461-R, polyclonal), and Bak (sc-832, polyclonal) were purchased from Santa Cruz Bio-technology (Santa Cruz, CA). Antibodies for caspase 8 (9746, monoclonal), caspase9 (9508, monoclonal), caspase 3 (9665, polyclonal), Bcl-2 (2870, monoclonal), p-p38 (9215, monoclonal), p38 (9212, monoclonal), p-ERK (4376, monoclonal), ERK(4695, monoclonal), p-JNK (4671, monoclonal) and JNK (9252, polyclonal) wereprovided by Cell Signaling (Danvers, MA).
2.2. Purification of MAP30
MAP30 was purified according to the methods reported by Lee-Huang et al.with modifications [8]. For routine preparation, 250 g of dried BG seeds was soakedin deionized water overnight, followed by homogenization and centrifugation(16,000 g, 30 min, 4 �C), and the aqueous supernatant was loaded on a Blue-Sepharose column in 0.02 M Tris–HCl. After washing off the unadsorbed fraction,the eluate obtained by elution with 1 M NaCl was dialyzed, and subjected to anSP-Sepharose column. After removing the unadsorbed and loosely-adsorbed impu-rities with 0.02 M NH4OAc containing 0.05 M NaCl, the SP- Sepharose column waseluted with 0.02 M NH4OAc containing 0.1 M NaCl. Homogeneous MAP30 waseluted, pooled, dialyzed, lyophilized, and stored at �20 �C for further studies.
2.3. Cell culture, cell viability/proliferation, anchorage-independent, and wound healingassays
Hep G2 cells were maintained in DMEM medium with 10% fetal bovine serumand antibiotics. Cell viability assay was carried out using MTT reagent (Sigma, USA)as previously described [4]. For cell proliferation assay, counting of total cell num-ber as well as number of dead cells was performed using a haemocytometer aftertrypan blue staining of the cells [4]. Anchorage-independent assay was performedand photographed using a light microscope [17]. In addition, wound healing assayunder mono layer conditions was done per the protocol as described elsewhere[18]. All experiments were performed in triplicate and at least two repetitive exper-iments were performed for each result.
2.4. Cell cycle analysis and apoptosis studies
Cell cycle analysis was performed using propidium iodide (PI) staining as previ-ously reported from our lab [3]. To detect MAP30-induced cell apoptosis, DNA frag-mentation using an In Situ Cell Death Detection Kit/TUNEL (Roche, Indianapolis, IN),quantitative analysis using Annexin V-PI double staining, and detection of typicalnuclear morphological changes using Hoechst 33342 staining (Sigma, USA) werecarried out as we reported in detail elsewhere [3,4]. All flow cytometry data wereanalyzed by FCS Express 4 software. Furthermore, for inhibitor interference studies,Z-VAD-FMK (final concentration 20 lM) was employed to inhibit the phosphoryla-tion of caspase cascades, and Tocriset™ kit was used to inhibit the phosphorylationof MAPKs. After pretreatment with the abovementioned inhibitors for 1 h, MAP30was added and incubated for 24 h. Apoptosis was measured with flow cytometry,followed by Annexin V and PI staining.
2.5. Western blotting
The effects of MAP30 on the regulation of caspase cascades were studied byWestern blotting as we reported before [3,19]. Hep G2 cells were cultured withMAP30 at 14.3 lM and 28.6 lM for 24, 36, and 48 h, and whole cell extracts wereprepared by lysing cells in ice-cold lysis buffer. Insoluble material was separatedby high speed centrifugation, and protein concentration of the supernatantwas determined using the BCA protein assay kit. Twenty lg of protein was size-fractionated using SDS–PAGE, following which proteins in the gels were transferredonto polyvinylidene difluoride membranes. Membranes were firstly blocked in 5%fresh milk for 1 h, followed by incubation with a specific primary antibody over-night at 4 �C. Finally, membranes were incubated with a horseradish peroxidase-conjugated anti-mouse or anti-rabbit secondary antibody (Cell Signaling, Danvers,MA) for 30 min, and bands were visualized by using ECL detection system (Amer-sham Life Science). On the other hand, in order to investigate the effect of MAP30on the phosphorylation levels of Akt and different MAPKs, whole cell extracts wereprepared using lysis buffer with phospho-stop solution (Roche). Other procedureswere the same. Quantification of protein bands was done using ImageJ software.
2.6. Assay of mitochondrial depolarization
Measurement of changes of mitochondrial transmembrane potential was per-formed as we reported before [3]. Briefly, after different treatments, cells were har-vested and stained with JC-1 dye at a concentration of 2.5 lg/ml for 15 min, andapplied to a FACSort flow cytometer (Becton Dickinson) for analysis. Data were ana-lyzed by FCS Express 4 software.
2.7. In vivo studies in nude mice
Ten BALB/c nude mice were purchased from the Laboratory Animal ServicesCentre at The Chinese University of Hong Kong, and all experimental procedureshad been approved and were performed under the direction of the University Ani-mal Research Ethics Committee. HepG2 cells (2 � 107) in 0.2 ml DMEM mediumwere injected subcutaneously into the right flank of each nude mouse using a 26-gauge needle. It was observed that by 17 days a solid tumor was apparent in allHep G2-injected nude mice. On the same day, mice were randomly divided intotwo groups (5 mice in each group), and there was no statistical difference of tumorvolumes between the groups. Mice in the MAP30 group received an intraperitonealinjection of 2.0 mg MAP30/kg body weight every other day (total 7 injections,13 days), and the control mice were treated with PBS instead. Tumor diameterswere serially measured using an electronic caliper, and tumor volumes were calcu-lated using the following formula as we used before [3]: tumor volume (cm3) =0.5 � tumor length (cm) � tumor width2 (cm2). After seven MAP30 injections, allmice were sacrificed. Tumors were excised, weighted, followed by fixation in 10%formalin, and embedding in paraffin wax for further immunohistochemicalanalysis.
2.8. Immunohistochemistry
Immunohistochemistry was performed as described elsewhere [3]. For histopa-thology analyses of tumor tissues, tissue slides were stained with Hematoxy-lin&Eosin (H&E). Apoptosis was examined by anti-cleaved caspase-3 (CellSignaling, MA), cleaved PARP (sc-23461-R, Santa Cruz Biotechnology, CA), andTUNEL detection kit (Roche, Germany). All sections were coverslipped and stainingwas scored by an experienced pathologist blinded to treatment group. The activatedcaspase-3, cleaved PARP, and TUNEL-positive cells were quantitated as percentageof cells positive from 5 high-powered fields in each section. Slides treated with Iso-type-matched IgG but without each primary antibody were served as negativecontrol.
2.9. Statistics
Data were analyzed using SPSS 11.0 (SPSS, Chicago) software. All data wereshown as mean ± standard deviation (S.D.) from at least two independent exper-iments. For both in vitro experiments and in vivo mouse experiments, a two-sided student t test was applied for the comparison of continuous variables be-tween two groups. Differences are considered significant when the p valueswere <0.05.
3. Results
3.1. MAP30 inhibits Hep G2 cell proliferation and motility
MAP30 was purified to homogeneity as judged by the presenceof a single band in SDS–PAGE (Fig. 1A). Using a HP 1000A Edmandegradation unit and a HP 1000 HPLC system, the sequence ofthe first 20 N-terminal amino acids was acquired which was in

Fig. 1. MAP30 exhibits cytotoxicity in Hep G2 cells. (A) Purification of MAP30 by liquid chromatography. Lane 1, protein ladder; Lane 2, crude extract of BG seeds; Lane 3,column fraction adsorbed on Blue-Sepharose; Lane 4, purified MAP30. (B) MAP30 reduced cell viability in Hep G2 cells. Cells were treated with different concentrations ofMAP30 (0.98–62.5 lM) for 24 h and 48 h, and then the residual cell viability was measured by MTT assay. (C and D) MAP30 manifested cytotoxic and cytostatic effects towardHep G2 cells. After MAP30 treatment, cells were counted with a haemocytometer using the trypan blue exclusion assay. (E) MAP30 reduced anchorage-independentproliferation in Hep G2 cells. Cells were seeded into soft agar containing PBS (control) or 1.95 lM MAP30. After 30 days incubation, colonies were photographed and countedunder a light microscope, and clonogenicity was determined. (F) MAP30 retarded cell motility. A sterile pipette tip was used to make a straight scratch, simulating a wound.After culture with 1.95 lM MAP30 for 24 h, bright-field images were taken on a NIKON TE2000 microscope. For Figs. B–E, results represent mean ± SD (n = 3).
68 E.F. Fang et al. / Cancer Letters 324 (2012) 66–74
accordance with previous reports (data not shown) [1,8]. We thenused the MTT assay to test the cytotoxic effect of MAP30 on Hep G2cells. Results showed that MAP30 inhibited cell viability in a time-and dose-dependent manner with an IC50 value of 28.6 lM for 24 hand 7.8 lM for 48 h (Fig. 1B). In view of the fact that the MTT assaydoes not allow a discrimination of cytostatic and cytotoxic effects,counting of total cell number as well as number of dead cells wasperformed using a haemocytometer and trypan blue staining of thecells. As shown in Figs. 1C and D, MAP30 exerted both cytostaticand cell death-inducing activities in a time and/or dose-dependentfashion. Furthermore, application of MAP30 at a very small concen-tration (1.95 lM, one quarter of the IC50 at 48 h) suppressed colonygrowth of Hep G2 cells in soft agar. On day 30, significant colonyformation appeared in the control dish, whereas there was almostno colony formation detected in the dish with addition of MAP30.The percentages of clonogenicity were statistically significant(p < 0.05) between the two groups (Fig. 1E). Moreover, MAP30 sup-pressed cell motility in Hep G2 cells (Fig. 1F).
3.2. MAP30 induces cell cycle arrest and apoptosis
PI staining revealed that MAP30 induced S-phase arrest in HepG2 cells. After culture with 28.6 lM MAP30 for 24 h, accumulation
of cells in S phase of the cell cycle increased from a baseline valueof 36.69–69.52% (Fig. 2A). The results were consistent with thecytostatic activity shown in Fig. 1D. We further assessed whetherMAP30 triggered apoptosis in Hep G2 cells. Firstly, Annexin V-PIdouble staining showed that the number of apoptotic cells wasdose-dependently elevated in Hep G2 cells treated with MAP30(2.24% ± 0.5% for control vs. 60.7% ± 5.6% for 28.6 lM MAP30,Fig. 2B). Secondly, typical morphological changes in MAP30-treated Hep G2 cells were detected using Hoechst 33342 staining.As Fig. 2C indicates, stereotypical apoptotic features, such askaryorrhexis, chromatin condensation, and DNA fragmentationwere noticed. Thirdly, the TUNEL results quantitatively confirmedthat MAP30 increased the number of cells undergoing DNAfragmentation (Fig. 2D).
3.3. Both extrinsic and intrinsic caspase pathways were involved inMAP30-induced apoptosis
We further investigated the molecular mechanisms involved inMAP30-induced apoptosis. Western blotting showed that MAP30reduced protein expression of p53 in a time- and dose-dependentmanner (Fig. 3A). To discover whether caspase-dependent path-ways were involved in apoptosis, activation levels of caspase-8,

Fig. 2. MAP30 arrests cell cycle progression at the S-phase and induces apoptosis in Hep G2 cells. (A) MAP30 induces S-phase cell cycle arrest. Cells were treated withdifferent concentrations of MAP30 for 24 h, following which cells were stained with PI and analyzed on a flow cytometer. (B) MAP30 treatment brought about apoptosis inHep G2 cells. After the same treatment shown in A, cells were stained with Annexin V-PI double staining and analyzed on a flow cytometer. Results represent mean ± SD(n = 3). (C) Stereotypical morphological changes of cell apoptosis was noticed in MAP30-treated Hep G2 cells. Cells were treated with MAP30 at specified doses for 24 h, and inturn stained with Hoechst 33342. Nuclear morphology was observed with a fluorescence microscope. (D) MAP30 induced DNA fragmentation in Hep G2 cells. Cells weretreated with MAP30 for 24 h and DNA fragmentation was detected with a TUNEL kit.
E.F. Fang et al. / Cancer Letters 324 (2012) 66–74 69
-9, -3, and PARP were assessed after treatment with MAP30 at twodoses (14.3 and 28.6 lM) for 24, 36, and 48 h. As shown in Fig. 3B,MAP30 displayed the ability to initiate the activation of all threecaspases. It subsequently induced the cleavage of PARP which fi-nally lead to apoptosis. In view of the possibility that some B-celllymphoma-2 (Bcl-2) family members are involved in caspase-in-volved apoptosis and cell cycle arrest [20,21], the effect ofMAP30 on the protein expression levels of Bcl2, Bak, and Bid wasdetermined. The results demonstrated that MAP30 reduced theexpression of the anti-apoptotic protein in a time- and dose-dependent manner, Nevertheless, there was no statistically signif-icant effect on the expression of either of the two pro-apoptoticproteins, Bak and Bid (Fig. 3C). It has been reported that apoptosis-inducing agents always trigger mitochondrial membrane depolar-ization which is a pre-requisite for caspase-9 translocation frommitochondria to the nucleus and in turn for its activation[3,22,23]. JC-1 staining revealed that MAP30 increased the percent-age of cells undergoing membrane depolarization (cytosol mono-meric JC-1 positive staining) from the baseline of 26.49%(control) to 56.91%. In addition, the pancaspase inhibitor z-VAD-FMK diminished MAP30-induced cell apoptosis which corrobo-rated the importance of caspase action in this process (Fig. 3E).
3.4. Regulation of Akt and MAPKs play a role in MAP30-inducedapoptosis
Emerging data indicated that regulation of phosphorylationof Akt and MAPKs was detected in different tumor cells afterexposure to chemotherapeutic agents [3,4,24]. We performed anon-time monitoring of the phosphorylation levels of Akt and threeMAPK members, p38 MAPK, extracellular signal-regulated kinase(ERK), and Jun kinase (JNK). We found that MAP30 increased thephosphorylation of Akt from 0.5 h to 8 h. Similarly, the phosphor-ylation levels of p38 MAPK as well as ERK were noticed, whichare shown in the Western blotting data as increased protein levelsof p-p38 MAPK and p-ERK, respectively (Fig. 4A). However, no sig-nificant phosphorylation changes of JNK were observed (data notshown). To further confirm that initiation of the phosphorylationof MAPKs was important to MAP30-induced apoptosis in Hep G2cells, SB203580 (a specific p38 MAPK inhibitor) and PD98059 (aspecific ERK inhibitor) were utilized in our studies. Consistently,we found that pretreatment of SB203580 or PD98059 for 1 himpaired induction of apoptosis by MAP30, suggesting thatMAP30-induced phosphorylation of p38 MAPK and ERK was atleast partially attributed to its cytotoxicity.
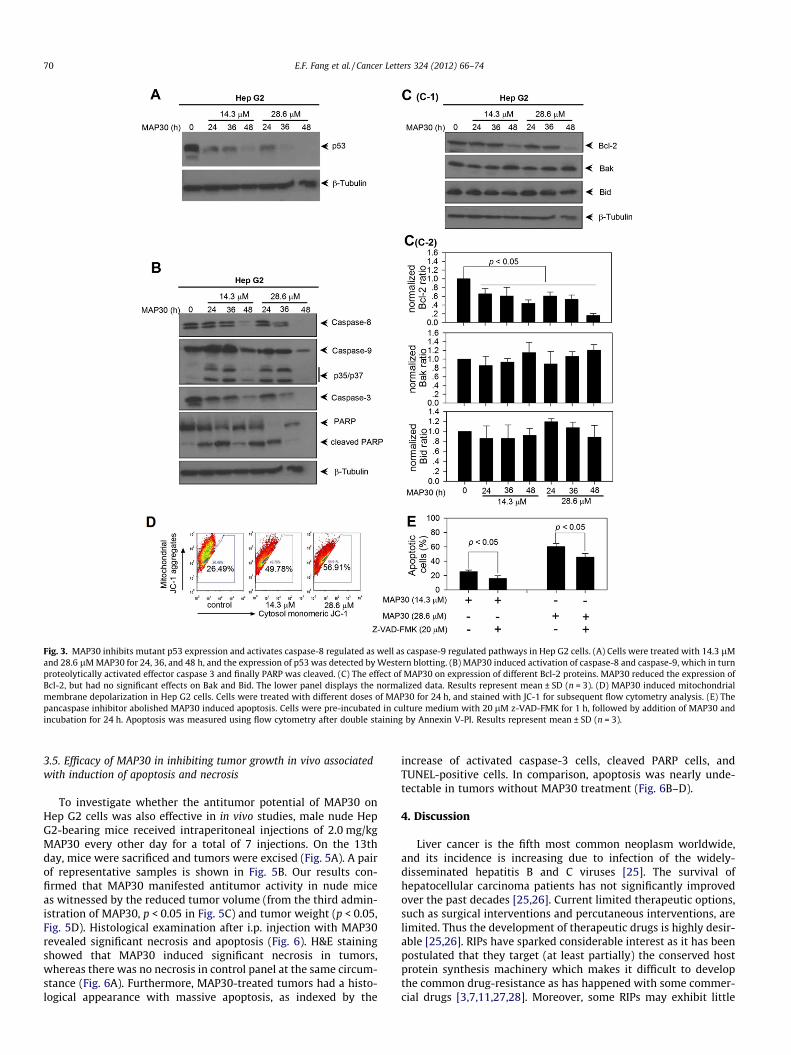
Fig. 3. MAP30 inhibits mutant p53 expression and activates caspase-8 regulated as well as caspase-9 regulated pathways in Hep G2 cells. (A) Cells were treated with 14.3 lMand 28.6 lM MAP30 for 24, 36, and 48 h, and the expression of p53 was detected by Western blotting. (B) MAP30 induced activation of caspase-8 and caspase-9, which in turnproteolytically activated effector caspase 3 and finally PARP was cleaved. (C) The effect of MAP30 on expression of different Bcl-2 proteins. MAP30 reduced the expression ofBcl-2, but had no significant effects on Bak and Bid. The lower panel displays the normalized data. Results represent mean ± SD (n = 3). (D) MAP30 induced mitochondrialmembrane depolarization in Hep G2 cells. Cells were treated with different doses of MAP30 for 24 h, and stained with JC-1 for subsequent flow cytometry analysis. (E) Thepancaspase inhibitor abolished MAP30 induced apoptosis. Cells were pre-incubated in culture medium with 20 lM z-VAD-FMK for 1 h, followed by addition of MAP30 andincubation for 24 h. Apoptosis was measured using flow cytometry after double staining by Annexin V-PI. Results represent mean ± SD (n = 3).
70 E.F. Fang et al. / Cancer Letters 324 (2012) 66–74
3.5. Efficacy of MAP30 in inhibiting tumor growth in vivo associatedwith induction of apoptosis and necrosis
To investigate whether the antitumor potential of MAP30 onHep G2 cells was also effective in in vivo studies, male nude HepG2-bearing mice received intraperitoneal injections of 2.0 mg/kgMAP30 every other day for a total of 7 injections. On the 13thday, mice were sacrificed and tumors were excised (Fig. 5A). A pairof representative samples is shown in Fig. 5B. Our results con-firmed that MAP30 manifested antitumor activity in nude miceas witnessed by the reduced tumor volume (from the third admin-istration of MAP30, p < 0.05 in Fig. 5C) and tumor weight (p < 0.05,Fig. 5D). Histological examination after i.p. injection with MAP30revealed significant necrosis and apoptosis (Fig. 6). H&E stainingshowed that MAP30 induced significant necrosis in tumors,whereas there was no necrosis in control panel at the same circum-stance (Fig. 6A). Furthermore, MAP30-treated tumors had a histo-logical appearance with massive apoptosis, as indexed by the
increase of activated caspase-3 cells, cleaved PARP cells, andTUNEL-positive cells. In comparison, apoptosis was nearly unde-tectable in tumors without MAP30 treatment (Fig. 6B–D).
4. Discussion
Liver cancer is the fifth most common neoplasm worldwide,and its incidence is increasing due to infection of the widely-disseminated hepatitis B and C viruses [25]. The survival ofhepatocellular carcinoma patients has not significantly improvedover the past decades [25,26]. Current limited therapeutic options,such as surgical interventions and percutaneous interventions, arelimited. Thus the development of therapeutic drugs is highly desir-able [25,26]. RIPs have sparked considerable interest as it has beenpostulated that they target (at least partially) the conserved hostprotein synthesis machinery which makes it difficult to developthe common drug-resistance as has happened with some commer-cial drugs [3,7,11,27,28]. Moreover, some RIPs may exhibit little

Fig. 5. MAP30 demonstrates antitumor efficacy in Hep G2 xenograft model. Hep G2 cells were implanted subcutaneously in the right flank of male nude mice. On the 17thday of tumor cell injection, treatment was initiated at a dose of 2.0 mg MAP30/kg body weight every other day for a total of seven injections. There was no detectable lethalityor weight changes in both groups. Mice were sacrificed on the 13th day of MAP30 treatment, and tumors were excised. (A) A pair of representative mice is shown, (B) MAP30administration produced complete tumor stasis as evidenced by the smaller tumor volumes, (C) as well as tumor weight, and (D) compared with the control panel. Resultsrepresent mean ± SD (n = 5).
Fig. 4. MAP30 induces phosphorylation of Akt and two MAPK members which are implicated in cell apoptosis. (A) MAP30 induced up-regulation of p-Akt, p-p38 MAPK, andp-ERK. Hep G2 cells were incubated with various concentrations of MAP30 for 0–12 h. Protein expression was determined by Western blotting. (B) Phosphorylation of p38MAPK and ERK were implicated in MAP30-induced cell apoptosis. Cells were pre-incubated with the p38 inhibitor SB203580 or the ERK inhibitor PD98059 for 1 h.Subsequently MAP30 was added at a final concentration of 14.3 lM followed by incubation for 24 h. Apoptosis was examined by flow cytometry using Annexin V-PI staining.Results represent mean ± SD (n = 3).
E.F. Fang et al. / Cancer Letters 324 (2012) 66–74 71
or no detectable adverse effect on normal cells for they may recog-nize features on membranes unique/dominant to tumor cells[3,11,27,28]. For example, MAP30 was not toxic to human sperma-tozoa at the doses it inhibited HIV-1 and herpes simplex virus[11,27]. Our interest in MAP30 stems from its potential therapeuticutility against both HIV and tumors, and we demonstrate hereinthat MAP30 is a potentially promising anti-liver cancer therapeuticagent.
Previous screening studies showed that MAP30 inhibitedgrowth of different tumor cells, including liver hepatoma Hep 3Bcells [14]. Here we further found that MAP30 manifested bothcytostatic and cytotoxic effects in Hep G2 cells. MAP30 also
inhibited the colony formation of Hep G2 cells in soft agar. Impor-tantly, Hep G2 cell motility was also retarded by MAP30 treatment.This is of more than trivial interest since tumor metastasis is still abig challenge for liver cancer patients in both chemotherapy andafter surgical interventions [25]. Furthermore, we found thatMAP30 induced cell cycle arrest in S-phase and this may explainits cytostatic activity. The cell cycle arrest ability of MAP30 wasin consistence with other reports of different BG components. Forexample, the crude extract of BG fruits induced G2-M phase accu-mulation in breast cancer MCF-7 cells [29], inhibited G1/S-specificcyclin D1, D2, and D3 expression in adrenocortical cancer cells[30], whereas the S-phase arrest was noticed in prostate cancer

Fig. 6. MAP30 induced necrosis and apoptosis in Hep G2 tumor tissues. Matched sections of control (left panel) and MAP30-treated tumors were stained with H&E (A),activated caspase-3 (B), cleaved PARP (C), and TUNEL kit (D) as fully described in Section 2. In Fig. A, significant necrosis was detected in MAP30-treated tumor tissues(indicated in asterisks). For B–D, histological figures in the right of each panel were quantitative data from 5 high-powered fields in each section. There were significantdifferences between control and MAP30-treated group (p < 0.05). Magnification, �40 (upper panel of Fig. A) and �200 (lower panel of Figs. A–D).
72 E.F. Fang et al. / Cancer Letters 324 (2012) 66–74
cells (PC3 and LNCaP) under the same treatment [31]. Wepreviously reported that MCL, a type II RIP existed in both BG fruitsand seeds, caused G1 cell-cycle arrest in nasopharyngeal carci-noma CNE-1 and CNE-2 cells [3].
It has been reported that p53 gene mutation is frequentlyoccurring in liver cancer, and p53 could markedly increaseinsulin-like growth factor II (IGF-II) expression [32]. IGF-II servesas a mitogenic growth factor in tumor development and also playsa role as a surviving factor which contributes to the selection oftransformed hepatocellular cells (by HBV infections) with an in-creased resistance to apoptosis [32]. It is now critical to developtherapeutic strategies to circumvent the possible anti-apoptoticfunction of mutant p53. Such strategies are currently the focus ofintense research [33]. The Hep G2 cell line used here has anN-ras mutation at codon 61 position 2 [34]. In this regard, ourresults found that MAP30 strongly inhibited mutant p53 expres-sion in Hep G2 cells. This inspiring result is reminiscent of Brennanand coworkers’ observation that BG extract inhibited cell prolifer-ation of adrenocortical cancer [30]. They found that BG extracttreatment significantly reduced the level of insulin-like growth fac-tor 1 receptor and its downstream signaling pathway, though therewas no p53 mutation in the used cells. Inhibition of mutant p53expression exists in other chemotherapeutic agents-treated tumorcells [3,35]. An example is MCL. It inhibited both mutant p53 andits downstream p21 expression in nasopharyngeal carcinomaCNE-1 and CNE-2 cells [3].
Growing evidence suggests that apoptosis is always involved inchemotherapeutic agents induced tumor cell death [3,4,23]. In thiscase, we found that MAP30 induced apoptosis in Hep G2 cells, asevidenced by the increase of Annexin V-positive cells, more cellsundergoing DNA fragmentation, and the appearance of stereotypi-cal nuclear morphological changes (Fig. 2). Furthermore, our re-sults disclosed that both caspase-8 regulated extrinsic as well ascaspase-9 regulated intrinsic pathways were involved in MAP30-induced cell apoptosis. In MAP30 treated cells, we found the phos-phorylation of initiator caspases-8 and -9, and executer caspse-3,and cleavage of the downstream PARP. Because PARP cleavage isclosely linked with DNA damage responses [36], these results cor-roborate our observation of the increased percentage of cellswith DNA fragmentation (Fig. 2D). In addition, inhibition of theactivation of caspase cascades by z-VAD-FMK could compromiseapoptosis-inducing function of MAP30, which suggest a role forcaspase cascades in the apoptosis process.
Results of cell cycle arrest, mitochondrial depolarization, andthe activation of caspases raise the possibility that the involvementof Bcl-2 protein family in MAP30-induced cell death. It has beenpostulated that apoptotic cell death is regulated by complicatednetworks between two subgroups of pro-apoptotic members (mul-tidomain proapoptotic proteins and BH3-only proapoptotic pro-teins) and antiapoptotic proteins of the Bcl-2 protein family [20].For instance, the proapoptotic proteins Bak and Bid can regulatethe mitochondrial membrane potential, inhibit cell-cycle progres-

Fig. 7. A diagram showing the pathways involved in MAP30-induced apoptosis in Hep G2 cells. MAP30 induced phosphorylation of ERK and p38, as well as the activation ofboth caspase-9- and caspase-8-mediated caspase cascades, which finally contributed to apoptosis. For more details, please refer to Section 4.
E.F. Fang et al. / Cancer Letters 324 (2012) 66–74 73
sion, inhibit cell survival, and increase apoptosis [20,21,37]. Thepro-survival protein Bcl-2 provides a distinct survival signal tothe cell and contributes to neoplasia [38]. In accordance with this,we found that MAP30 inhibited Bcl-2 expression in a dose- andtime-dependent way. The effect is commensurate with its functionin human colorectal carcinoma LoVo cells [16]. However, MAP30had no statistically significant effects on the protein levels of Bakand Bid. The propinquity effect of relaying an apoptotic signal fromdeath stimulus to the activation of caspase-9 may be substitutedby other Bcl-2 members [20].
Akt and MAPKs have been reported to be involved in theantitumor effects of some chemotherapeutics. For example, inthe process of chemotherapeutic-induced tumor cell death, thephosphorylation of Akt can be both up-regulated [4] and inhibited[39]. For the MAPK members, the phosphorylation of p38 MAPKand JNK is proapoptotic [3,4,40,41], whereas ERK phosphorylationhas been associated with both antitumor effect [4,41] and involvedin tumor progression and angiogenesis [24]. In this regard, we alsoinvestigated the effects of MAP30 on the phosphorylation of Aktand MAPK members. Treatment of Hep G2 cells with MAP30 re-sulted in phosphorylation of Akt, p38 MAPK and ERK at as earlyas 0.5 h and persisted to 8 or 12 h. This activation may contributeto MAP30-induced cytotoxicity, because inhibition of the p38MAPK by SB 203580 could partially reverse MAP30- mediatedapoptosis. This effect is amplified to the ERK pathway for inhibitionof the ERK with PD98059 also rescued cells from MAP30-inducedapoptosis. MAP30 had no significant effect on JNK activationthough another two BG components, including MCL and RNaseMC2, induced JNK activation in other tumor cells [3,4].
It has been reported that treatment of human breast cancerMDA-MB-231-bearing nude mice with MAP30 at 10 lg/injectionEOD for 10 times resulted in significant improvement of survival[42]. In the present study, MAP30 also inhibited Hep G2 xenograftgrowth in nude mice. We found that MAP30 administration for atotal of seven intraperitoneal injections at a pharmacologically safedose profoundly inhibited tumor volume as well as tumor weight.Side effects were not observed with the dose. Our further immuno-histochemical analysis indicated that MAP30 induced necrosis, andthe activation of apoptosis signaling pathways as evidenced bythe activation of caspase-3, PARP cleavage, and increase ofTUNEL-positive cells in tumor tissues. These findings suggest that
MAP30 can inhibit Hep G2 tumor progression through inductionof both necrosis and apoptosis. A diagram to summarize all the sig-naling pathways involved in MAP30-induced cell apoptosis in HepG2 cells is shown in Fig. 7.
It is known that chemotherapeutic agents exert their functionsby targeting on signaling proteins which are usually membranereceptors or kinases. The membrane protein(s) in tumor cells forMAP30 to bind on is unknown. However, the studies on trichosan-thin (TCS) may prove some clues. TCS is also a type I RIP protein,purified from the root tuber of Trichosanthes kirilowii, which sharesa 59% sequence homology with MAP30 [7]. The way for TCS to enterinto the tumor cells is associated with the binding on two receptorproteins (with molecular weights of 50 and 60 kDa, respectively),and/or depending on a2-macroglobulin receptor/low density lipo-protein receptor related protein (LRP)-mediated endocytosis [7].Future studies on the cell entry process of MAP30 may help to fur-ther clarify its anti-cancer molecular mechanism.
In conclusion, we have demonstrated that MAP30 could inhibitHep G2 cell growth in both cell culture settings and xenograft-bearing nude mice. The antitumor activity of MAP30 in Hep G2cells is attributed to (a) activation of both extrinsic caspase-8and intrinsic caspase-9 pathways, and (b) phosphorylation of bothAkt and some MAPK members. In view of that MAP30 was purifiedfrom the common edible vegetable bitter gourd, suitable ways ofMAP30 administration (such as intraperitoneal injection and intra-tumoral treatment) may be expected to be well tolerated. Thus,one can envision two possible scenarios in the medicinal applica-tions of MAP30: one is the application to fight against HIV andthe other is its tremendous therapeutic promise on tumors. Thelatter scenario merits clinical trials using MAP30 in the manage-ment of liver cancer.
References
[1] E.F. Fang, T.B. Ng, Bitter gourd (Momordica charantia) is a cornucopia of health:a review of its credited antidiabetic, anti-HIV, and antitumor properties, Curr.Mol. Med. 11 (2011) 417–436.
[2] H. Schaefer, S.S. Renner, A three-genome phylogeny of Momordica(Cucurbitaceae) suggests seven returns from dioecy to monoecy and recentlong-distance dispersal to Asia, Mol. Phylogenet. Evol. 54 (2010) 553–560.
[3] E.F. Fang, C.Z. Zhang, T.B. Ng, J.H. Wong, W.L. Pan, X.J. Ye, Y.S. Chan, W.P. Fong,Momordica Charantia Lectin, a Type II ribosome inactivating protein, exhibits

74 E.F. Fang et al. / Cancer Letters 324 (2012) 66–74
antitumor activity toward human nasopharyngeal carcinoma cells in vitro andin vivo, Cancer Prev. Res. (Phila) 5 (2011) 109–121.
[4] E.F. Fang, C.Z. Zhang, W.P. Fong, T.B. Ng, RNase MC2: a new Momordicacharantia ribonuclease that induces apoptosis in breast cancer cells associatedwith activation of MAPKs and induction of caspase pathways, Apoptosis 17(2011) 377–387.
[5] S.D. Xiong, K. Yu, X.H. Liu, L.H. Yin, A. Kirschenbaum, S. Yao, G. Narla, A. DiFeo,J.B. Wu, Y. Yuan, S.M. Ho, Y.W. Lam, A.C. Levine, Ribosome-inactivatingproteins isolated from dietary bitter melon induce apoptosis and inhibithistone deacetylase-1 selectively in premalignant and malignant prostatecancer cells, Int. J. Cancer 125 (2009) 774–782.
[6] E.F. Fang, T.B. Ng, Can bitter gourd (Momordica Charantia) be a novel therapyfor human cancers?, Medicinal Aromatic Plants 1 (2012) 1–2.
[7] E.F. Fang, T.B. Ng, P.C. Shaw, R.N.S. Wong, Recent progress in medicinalinvestigations on trichosanthin and other ribosome inactivating proteins fromthe plant genus trichosanthes, Curr. Med. Chem. 18 (2011) 4410–4417.
[8] S. Lee-Huang, P.L. Huang, P.L. Nara, H.C. Chen, H.F. Kung, P. Huang, H.I. Huang,MAP 30: a new inhibitor of HIV-1 infection and replication, FEBS Lett. 272(1990) 12–18.
[9] A.S. Bourinbaiar, S. Lee-Huang, The activity of plant-derived antiretroviralproteins MAP30 and GAP31 against herpes simplex virus in vitro, Biochem.Biophys. Res. Commun. 219 (1996) 923–929.
[10] S. Lee-Huang, P.L. Huang, A.S. Bourinbaiar, H.C. Chen, H.F. Kung, Inhibition ofthe integrase of human immunodeficiency virus (HIV) type 1 by anti-HIV plantproteins MAP30 and GAP31, Proc. Natl. Acad. Sci. USA 92 (1995) 8818–8822.
[11] Y.X. Wang, N. Neamati, J. Jacob, I. Palmer, S.J. Stahl, J.D. Kaufman, P.L. Huang,H.E. Winslow, Y. Pommier, P.T. Wingfield, S. Lee-Huang, A. Bax, D.A. Torchia,Solution structure of anti-HIV-1 and anti-tumor protein MAP30: structuralinsights into its multiple functions, Cell 99 (1999) 433–442.
[12] Y. Sun, P.L. Huang, J.J. Li, Y.Q. Huang, L. Zhang, S. Lee-Huang, Anti-HIV agentMAP30 modulates the expression profile of viral and cellular genes forproliferation and apoptosis in AIDS-related lymphoma cells infected withKaposi’s sarcoma-associated virus, Biochem. Biophys. Res. Commun. 287(2001) 983–994.
[13] T. Arazi, P. Lee Huang, P.L. Huang, L. Zhang, Y. Moshe Shiboleth, A. Gal-On, S.Lee-Huang, Production of antiviral and antitumor proteins MAP30 and GAP31in cucurbits using the plant virus vector ZYMV-AGII, Biochem. Biophys. Res.Commun. 292 (2002) 441–448.
[14] S. Lee-Huang, P.L. Huang, H.C. Chen, A. Bourinbaiar, H.I. Huang, H.F. Kung, Anti-HIV and anti-tumor activities of recombinant MAP30 from bitter melon, Gene161 (1995) 151–156.
[15] P.L. Huang, Y. Sun, H.C. Chen, H.F. Kung, S. Lee-Huang, Proteolytic fragments ofanti-HIV and anti-tumor proteins MAP30 and GAP31 are biologically active,Biochem. Biophys. Res. Commun. 262 (1999) 615–623.
[16] J.M. Fan, J. Luo, J. Xu, S. Zhu, Q. Zhang, D.F. Gao, Y.B. Xu, G.P. Zhang, Effects ofrecombinant MAP30 on cell proliferation and apoptosis of human colorectalcarcinoma LoVo cells, Mol. Biotechnol. 39 (2008) 79–86.
[17] W. Chen, J.D. Arroyo, J.C. Timmons, R. Possemato, W.C. Hahn, Cancer-associated PP2A Aalpha subunits induce functional haploinsufficiency andtumorigenicity, Cancer Res. 65 (2005) 8183–8192.
[18] K.L. Kong, D.L. Kwong, L. Fu, T.H. Chan, L. Chen, H. Liu, Y. Li, Y.H. Zhu, J. Bi, Y.R.Qin, S.Y. Law, X.Y. Guan, Characterization of a candidate tumor suppressorgene uroplakin 1A in esophageal squamous cell carcinoma, Cancer Res. 70(2010) 8832–8841.
[19] C.Z. Zhang, H.T. Zhang, G.G. Chen, P.B. Lai, Trichostatin A sensitizes HBx-expressing liver cancer cells to etoposide treatment, Apoptosis 16 (2011) 683–695.
[20] P.N. Kelly, A. Strasser, The role of Bcl-2 and its pro-survival relatives intumourigenesis and cancer therapy, Cell Death Differ. 18 (2011) 1414–1424.
[21] S. Maddika, S.R. Ande, S. Panigrahi, T. Paranjothy, K. Weglarczyk, A. Zuse, M.Eshraghi, K.D. Manda, E. Wiechec, M. Los, Cell survival, cell death and cell cyclepathways are interconnected: implications for cancer therapy, Drug Resist.Update 10 (2007) 13–29.
[22] S. Krajewski, M. Krajewska, L.M. Ellerby, K. Welsh, Z. Xie, Q.L. Deveraux, G.S.Salvesen, D.E. Bredesen, R.E. Rosenthal, G. Fiskum, J.C. Reed, Release ofcaspase-9 from mitochondria during neuronal apoptosis and cerebralischemia, Proc. Natl. Acad. Sci. USA 96 (1999) 5752–5757.
[23] S. Seoane, J.C. Montero, A. Ocana, A. Pandiella, Effect of multikinase inhibitorson caspase-independent cell death and DNA damage in HER2-overexpressingbreast cancer cells, J. Natl Cancer Inst. 102 (2010) 1432–1446.
[24] S.M. Wilhelm, C. Carter, L. Tang, D. Wilkie, A. McNabola, H. Rong, C. Chen, X.Zhang, P. Vincent, M. McHugh, Y. Cao, J. Shujath, S. Gawlak, D. Eveleigh, B.Rowley, L. Liu, L. Adnane, M. Lynch, D. Auclair, I. Taylor, R. Gedrich, A.Voznesensky, B. Riedl, L.E. Post, G. Bollag, P.A. Trail, BAY 43-9006 exhibitsbroad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathwayand receptor tyrosine kinases involved in tumor progression and angiogenesis,Cancer Res. 64 (2004) 7099–7109.
[25] J.M. Llovet, A. Burroughs, J. Bruix, Hepatocellular carcinoma, Lancet 362 (2003)1907–1917.
[26] H.B. El-Serag, Hepatocellular carcinoma, N. Engl. J. Med. 365 (2011) 1118–1127.
[27] C.A. Schreiber, L. Wan, Y. Sun, L. Lu, L.C. Krey, S. Lee-Huang, The antiviralagents, MAP30 and GAP31, are not toxic to human spermatozoa and may beuseful in preventing the sexual transmission of human immunodeficiencyvirus type 1, Fertil. Steril. 72 (1999) 686–690.
[28] C.D. Putnam, J.A. Tainer, The food of sweet and bitter fancy, Nat. Struct. Biol. 7(2000) 17–18.
[29] R.B. Ray, A. Raychoudhuri, R. Steele, P. Nerurkar, Bitter melon (Momordicacharantia) extract inhibits breast cancer cell proliferation by modulating cellcycle regulatory genes and promotes apoptosis, Cancer Res. 70 (2010) 1925–1931.
[30] V.C. Brennan, C.M. Wang, W.H. Yang, Bitter melon (Momordica charantia)extract suppresses adrenocortical cancer cell proliferation through modulationof the apoptotic pathway, steroidogenesis, and insulin-like growth factor Type1 receptor/RAC-alpha serine/threonine-protein kinase signaling, J. Med. Food15 (2012) 325–334.
[31] P. Ru, R. Steele, P.V. Nerurkar, N. Phillips, R.B. Ray, Bitter melon extract impairsprostate cancer cell-cycle progression and delays prostatic intraepithelialneoplasia in TRAMP model, Cancer Prev. Res. (Phila) 4 (2011) 2122–2130.
[32] Y.I. Lee, S. Lee, G.C. Das, U.S. Park, S.M. Park, Activation of the insulin-likegrowth factor II transcription by aflatoxin B1 induced p53 mutant 249 iscaused by activation of transcription complexes; implications for a gain-of-function during the formation of hepatocellular carcinoma, Oncogene 19(2000) 3717–3726.
[33] K. Ohnishi, I. Ota, A. Takahashi, K. Yane, H. Matsumoto, T. Ohnishi, Transfectionof mutant p53 gene depresses X-ray- or CDDP-induced apoptosis in a humansquamous cell carcinoma of the head and neck, Apoptosis 7 (2002) 367–372.
[34] I.C. Hsu, T. Tokiwa, W. Bennett, R.A. Metcalf, J.A. Welsh, T. Sun, C.C. Harris, P53gene mutation and integrated hepatitis B viral DNA sequences in human livercancer cell lines, Carcinogenesis 14 (1993) 987–992.
[35] C.A. Kanashiro, A.V. Schally, K. Groot, P. Armatis, A.L. Bernardino, J.L. Varga,Inhibition of mutant p53 expression and growth of DMS-153 small cell lungcarcinoma by antagonists of growth hormone-releasing hormone andbombesin, Proc. Natl. Acad. Sci. USA 100 (2003) 15836–15841.
[36] A.H. Boulares, A.G. Yakovlev, V. Ivanova, B.A. Stoica, G. Wang, S. Iyer, M.Smulson, Role of poly(ADP-ribose) polymerase (PARP) cleavage in apoptosis.Caspase 3-resistant PARP mutant increases rates of apoptosis in transfectedcells, J. Biol. Chem. 274 (1999) 22932–22940.
[37] S. Shimizu, M. Narita, Y. Tsujimoto, Bcl-2 family proteins regulate the releaseof apoptogenic cytochrome c by the mitochondrial channel VDAC, Nature 399(1999) 483–487.
[38] D.L. Vaux, S. Cory, J.M. Adams, Bcl-2 gene promotes haemopoietic cell survivaland cooperates with c-myc to immortalize pre-B cells, Nature 335 (1988) 440–442.
[39] S.S. Ng, M.S. Tsao, T. Nicklee, D.W. Hedley, Wortmannin inhibits pkb/aktphosphorylation and promotes gemcitabine antitumor activity in orthotopichuman pancreatic cancer xenografts in immunodeficient mice, Clin. CancerRes. 7 (2001) 3269–3275.
[40] A. Mansouri, L.D. Ridgway, A.L. Korapati, Q. Zhang, L. Tian, Y. Wang, Z.H. Siddik,G.B. Mills, F.X. Claret, Sustained activation of JNK/p38 MAPK pathways inresponse to cisplatin leads to Fas ligand induction and cell death in ovariancarcinoma cells, J. Biol. Chem. 278 (2003) 19245–19256.
[41] J. Chen, M. Rusnak, R.R. Luedtke, A. Sidhu, D1 dopamine receptor mediatesdopamine-induced cytotoxicity via the ERK signal cascade, J. Biol. Chem. 279(2004) 39317–39330.
[42] S. Lee-Huang, P.L. Huang, Y. Sun, H.C. Chen, H.F. Kung, W.J. Murphy, Inhibitionof MDA-MB-231 human breast tumor xenografts and HER2 expression by anti-tumor agents GAP31 and MAP30, Anticancer Res. 20 (2000) 653–659.
Copyright © 2022 FDOKUMEN




![IJCBS RESEARCH PAPER VOL. 1 [ISSUE 4] JUNE, 2014 ISSN:-2349–2724 Novel Formulation, Preparation and Quality Evaluation of Sweet Gourd ketchup](https://static.fdokumen.com/doc/165x107/6321140bb257cd26d003bb7a/ijcbs-research-paper-vol-1-issue-4-june-2014-issn-23492724-novel-formulation.jpg)







![Descriptors for Sponge Gourd [Luffa cylindrica (L.) Roem.]](https://static.fdokumen.com/doc/165x107/63187e763394f2252e02b92e/descriptors-for-sponge-gourd-luffa-cylindrica-l-roem.jpg)



