
HSP-1/2, a major protein of equine seminal plasma, exhibits chaperone-like activity
Upload
independentCategory
view
2download
0
The human oesophageal squamous epithelium exhibits a novel typeof heat shock protein response
Adam Yagui-Beltran1, Ashley L. Craig1, Laura Lawrie2, Devon Thompson3, Sarka Pospisilova1,4,David Johnston1, Neil Kernohan1, David Hopwood1, John F. Dillon1 and Ted R. Hupp1
1Cancer Research Campaign Laboratories, the Department of Molecular and Cellular Pathology, University of Dundee, Scotland;2University of Aberdeen, UK Protein Sequencing Facility, Scotland; 3Department of Surgery, Stanford University, CA, USA; 4Masaryk
Cancer Institute, Brno, Czech Republic;
The human oesophageal epithelium is subject to damage
from thermal stresses and low extracellular pH that can play
a role in the cancer progression sequence, thus identifying a
physiological model system that can be used to determine
how stress responses control carcinogenesis. The classic
heat shock protein HSP70 is not induced but rather is
down-regulated after thermal injury to squamous epithelium
ex vivo; this prompted a longer-term study to address
the nature of the heat shock response in this cell type. An
ex vivo epithelial culture system was subsequently used to
identify three major proteins of 78, 70, and 58 kDa, whose
steady-state levels are elevated after heat shock. Two of the
three heat shock proteins were identified by mass spectro-
metric sequencing to be the calcium-calmodulin homologue
transglutaminase-3 (78 kDa) and a recently cloned oeso-
phageal-specific gene called C1orf10, which encodes a
53-kDa putative calcium binding protein we have named
squamous epithelial heat shock protein 53 (SEP53). The
70-kDa heat shock protein (we have named SEP70) was not
identifiable by mass spectrometry, but it was purified and
studied immunochemically to demonstrate that it is distinct
from HSP70 protein. Monoclonal antibodies to SEP70
protein were developed to indicate that: (a) SEP70 is
induced by exposure of cultured cells to low pH or glucose
starvation, under conditions where HSP70 protein was
strikingly down-regulated; and (b) SEP70 protein exhibits
variable expression in preneoplastic Barrett’s epithelium
under conditions where HSP70 protein is not expressed.
These results indicate that human oesophageal squamous
epithelium exhibits an atypical heat shock protein response,
presumably due to the evolutionary adaptation of cells
within this organ to survive in an unusual microenvironment
exposed to chemical, thermal and acid reflux stresses.
Keywords: HSP70; SEP53; SEP70; p53; cancer.
The incidence and biology of oesophageal cancer in thedeveloped world has undergone dramatic changes in the lasttwo decades. The timescale of this change suggests adramatic alteration in the microenvironment of the oeso-phagus due to alterations in patient life-style and under-standing how this tissue responds to external stresses istherefore of fundamental interest. Cancer progression ispresumably is driven in part by gastro-oesophageal refluxdisease and progression to Barrett’s epithelium, which is apreneoplastic cell type representing an intermediate in thecancer progression sequence to adenocarcinoma. The oeso-phagus is exposed directly to thermal injury, acid or bilestresses, and oxidizing chemicals from the environment.These agents can damage DNA and proteins over the life-span of the cell, contribute to tissue damage, promote aging,and initiate diseases like cancer. Thus, it is of interest todefine the upstream signal transduction pathways that
orchestrate a cellular damage response in the oesophagealepithelium and to define how dysregulation of these stress-responsive networks contributes to tissue injury and cancerdevelopment.
Two major clinical models that have thus far givenmechanistic insight into the multistep nature of humanneoplasia include oesophageal and colorectal cancer [1,2].The development of neoplasia in both of these tissue typesinvolve mutations in tumour suppressor genes such as p53,but vary strikingly in the stage of the neoplastic sequencein which such mutations occur. In colorectal cancers,mutations in p53 often occur relatively late in the progres-sion sequence to carcinoma, while in the oesophagus p53mutations can often occur earlier in dysplastic or evenmetaplastic cells [3,4]. The earlier mutation pattern of p53in the adenocarcinoma progression sequence of the oeso-phagus suggests a unique requirement for inactivation of thep53-dependent damage-induced pathway and presumablyinvolves, in part, the type of microenvironmental damageimposed upon cells of the oesophagus. Consistent with this,it is important to point out that oesophageal cancers arisefrom two distinct natural histories with respect to p53 pathwayperturbation. Squamous cell carcinomas of the oesophaguscontain signature mutations in the p53 gene that suggest theinvolvement of DNA damaging agents or carcinogens,presumably due to life-style patterns such as cigarettesmoking. In contrast, in metaplasia of the oesophagus or
Correspondence to T. R. Hupp, Cancer Research Campaign
Laboratories, the Department of Molecular and Cellular Pathology,
University of Dundee, Dundee DD1 9SY, Scotland.
Tel.: 1 44 1382 496 430, E-mail: [email protected]
(Received 5 July 2001, accepted 21 August 2001)
Abbreviations: HSP, heat-shock protein; DMEM, Dulbecco[apos ]s
modified Eagle[apos ]s medium; SEP, squamous epithelial heat shock
protein
Eur. J. Biochem. 268, 5343–5355 (2001) q FEBS 2001

Barrett’s epithelium, within the adenocarcinoma lineage, thep53 mutations arise from spontaneous deamination [3],suggesting that a distinct life-style or environmental risk isinvolved in this type of cancer development. Further,morphologically indistinguishable adenocarcinoma canfurther be divided into two types that either do or do notoveramplify a novel region of chromosome 5p [5], thushighlighting that the study of this tissue is giving rise to newinsights into the molecular nature of cancer progression.
Types of naturally occurring environmental agents thatcan activate p53 protein and that play a role in acid-refluxdisease or progression to Barrett’s epithelium in this tissueinclude stresses such as lowered oxygen concentration, lowextracellular pH, and thermal injury. Such cellular damageor trauma appears to be an initiating event in the evolutionof ulceration, p53 mutation, metaplasia, dysplasia, andadenocarcinoma [6–12]. Cells of the normal oesophagealepithelium and Barrett’s epithelium have therefore presum-ably evolved specific mechanisms to tolerate or repair injuryinduced by exposure to these and other damaging agents thatare relatively unique to this tissue. It is of interest thereforeto develop an understanding of the cellular defense systemsthat operate in parallel to p53 in the normal and Barrett’sepithelium of the human oesophagus.
One of the most evolutionarily conserved and widelyrecruited cellular defense pathways comprise the stressprotein or heat shock protein family members. These poly-peptides termed molecular chaperones are classified basedon differences in molecular masses, which generally includeproteins of 25, 40, 60, 70, 90, and 110 kDa [13]. The bio-chemical function of molecular chaperones is thought torevolve around the regulation of protein folding, unfolding,intracellular transport, and protein degradation [14]. Thebiological consequences of stress protein induction in manycell types can involve, not only repair of damaged poly-peptides and cellular survival after injury, but acquisition ofthermotolerance or protection of cells from normally lethallevels of damage [15]. Although most studies on the stress-protein responses in mammals have been centered onin vitro studies using tissue culture cell lines, the effects ofin vivo hyperthermia, inflammation, or damage on HSPinduction are beginning to be unraveled using animal ororgan models [16]. One of the most striking results toemerge from animal studies is the remarkable discordance inthe expression of the HSP genes after physiological stress orheat shock in vivo [17,18]. Together these observationsindicate that environmentally or physiologically generatedsignals can activate the HSP pathway in vivo, but this iscell-specific, suggesting the existence of cell-type specificfactors that control the organ responses to stress.
Given the attraction of using the epithelium of the humanoesophagus as a model for heat, acid, and genotoxic-dependent signaling pathways that activate stress-proteinand p53 protein pathways, we had previously undertakenbasic research to determine whether the major heat shock-response pathway can be studied ex vivo with humanoesophageal biopsies and to determine the types of stress-protein responses that are activated in this tissue. The initialwork has defined the conditions for ex vivo organ cultureand has shown that this tissue has the unusual property ofsurprisingly down-regulating the normal molecular chaperoneHSP70 after thermal stress [19]. In this current study, wehave developed methods to identify, purify, sequence, and
characterize the major HSPs from normal squamous epi-thelium. Our data demonstrate that the squamous epitheliumexhibits a relatively novel type of heat shock response thushighlighting a group of gene products whose function mayregulate normal cellular integrity in squamous epithelialcells. The further study of this heat shock system may assistin shedding light on the regulatory changes that occur duringthe development of inflammation, dysplasia, and carcinomain this relatively unique microenvironment.
E X P E R I M E N T A L P R O C E D U R E S
Ex vivo treatment and radiolabeling of normal humanoesophageal epithelium
Oesophageal squamous epithelial biopsies were obtainedfrom patients undergoing upper gastro-intestinal endoscopy.Biopsy specimens were taken from 28 to 34 cm from thepatients teeth and the tissue was macroscopically normal,unless indicated to be Barrett’s epithelium. All patients gaveinformed consent and the study has been approved by theTayside Medical Ethical Committee. Samples werecollected into NaCl/Pi prechilled at 0 8C, the cells weretransferred to a polyethylene dish in 200 mL of Hams-F10(Gibco BRL) media preincubated at 37 8C in an incubatorwith 5% CO2 and, after 5 min, transferred to fresh mediacontaining sodium arsenite (250 mM or 500 mM) or ethanol(2 or 4%), as indicated. In order to induce the heat shockresponse, epithelial biopsies were transferred from NaCl/Pi
into Hams-F10 preincubated to 37 8C and either incubatedat 37 8C or subjected to a 15-min heat shock (at 42, 45, or55 8C) followed by an incubation for the indicated times at37 8C. Following the ex vivo stress, the cells were eitherfrozen in liquid nitrogen in order to examine the steady statepattern of protein synthesis using Coommassie stained SDSpolyacrylamide gels or incubated with radioactive methion-ine for radiolabeling of proteins. To label the protein syn-thesized after or during stress, cells that had been incubatingfor 2 or 4 h in either sodium arsenite (500 mM) or ethanol(4%) were incubated in labeling media at 37 8C for 30 minprior to harvesting. Labeling media contained methionine-and cysteine-free Dulbecco’s modified Eagle’s medium(DMEM; ICN) containing 10% dialyzed fetal bovine serum,2 mM glutamine, 100 mCi:mL21 of [35S]methionine, andeither sodium arsenite (500 mM) or ethanol (4%), as indi-cated in the figure legends. For labeling the polypeptides inheat shock cells, labeling media was also added andincubated at 37 8C for 30 min prior to harvesting, usingepithelial biopsies that had been incubating for 2 or 4 h aftera 15-min heat shock (55 8C). Relatively short radiolabelingtimes were employed, as prolonged exposure of cellsto radioisotopes can activate a p53-dependent radiation-responsive stress pathway in normal cells [20]. Cells werefrozen in liquid nitrogen and lysed as indicated below.
Lysis of squamous and Barrett’s epithelium
Following ex vivo treatment of epithelial samples in definedmedia, frozen samples were lysed by homogenizing in lysisbuffer (1% NP-40; 25 mM Hepes, pH 7.6; 5 mM dithio-threitol; 0.4 M KCl; 5 mM EDTA; 10 mM NaF; 2 mg:mL21
Pefabloc (Boehringer-Mannheim); 20 mg:mL21 leupeptin;1 mg:mL21 aprotinin; 2 mg:mL21 pepstatin; 10 mg:mL21
5344 A. Yagui-Beltran et al. (Eur. J. Biochem. 268) q FEBS 2001

trypsin inhibitor; and 1 mM benzamidine) and incubated at0 8C for 15 min. Soluble supernatant was obtained aftercentrifugation at 14 000 r.p.m. in a minicentrifuge at 2 8Cfor 10 min and protein concentrations were determinedby the method of Bradford [21]. One- or two-dimensionalgel electrophoresis was carried out as described previously[22].
Mass-spectrometric analysis of heat shock proteins innormal squamous and Barrett’s epithelium
Populations of normal squamous epithelium were lysedusing urea lysis buffer (8 M urea; 50 mM Hepes, pH 7.6;5 mM dithiothreitol; 0.4 M KCl; 1 mM benzamidine, 50 mM
(NaF) or lysis buffer (as above) to extract the polypeptides.Two-dimensional gel electrophoresis, where used, wascarried out by adding 100 mg of total protein into 100 mL ofsolubilization buffer (8 M urea, 1%, v/v, Triton X-100,40 mM Tris base, and 10 mM dithiothreitol), applied to aPharmacia Immobiline Dryplate (pH 4.0–7.0) equilibratedin denaturation buffer (8 M urea, 0.05%, v/v, Triton X-100;0.1 M dithiothreitol; and 0.5%, v/v, Pharmalyte, pH 3–10),and focused for 7 kVh with a circulating water cooling unitset at 15 8C. The second dimension electrophoresis wasperformed using standard SDS/PAGE and stained withCoomassie blue. Polypeptides whose expression wasdramatically distinct were processed for mass spectrometricsequencing by the UK Protein Sequencing Facility inAberdeen. In a typical manual processing, the protein bandwas excised from the gel and cut into 1-mm cubes witha clean scalpel blade on a clean glass plate. The gel piecewas washed in 3 band vol. of water for 15 min, then anequivalent volume of acetonitrile (HPLC grade) was addedfor a further 15 min The supernatant was removed and 3band vol. of 0.1 M NH4HCO3(BDH) was added for 15 minfollowed by an equivalent volume of acetonitrile for another15 min. Three volumes of acetonitrile were then added todehydrate the gel piece and the gel pieces were dried in aspeedy-vac. The polypeptides in the gel slices were alky-lated in situ by first adding NH4HCO3 and incubating at56 8C for 1 h followed by the addition of an equal volume of50 mM iodoacetamide in NH4HCO3. After incubation in thedark for 45 min at room temperature, the supernatant wasremoved and the gel slice was washed in 3 band volumes of0.1 M NH4HCO3. After removal of the wash solution, 3 vol.of acetonitrile were added to dehydrate the gel piece andthe gel pieces were dried in a speedy vac. Two gel volumesof 12.5 mg:mL21 modified trypsin (sequencing gradeBoehringer) in 20 mM NH4HCO3 was added to the driedgel slices and incubated at 30 8C for 30 min with shaking.After a further 18-h incubation without shaking, the aqueoussupernatant was removed for mass spectrometric analysis asdescribed below. The trypsinized peptide extract was passedthrough a GELloader tip which contained a small volume ofPOROS R2 solvent (PerSeptive Biosystems, Framingham,MA, USA). The absorbed peptides were washed and theneluted in 0.5 mL of a saturated solution of a-cyano-4-hydroxy-cinnamic acid (Aldrich Chemical, Poole, UK) in50% acetonitrile/5% formic acid. The mass spectra wereacquired on a PerSeptive Biosystems Voyager-DE STRMALDI-TOF mass spectrometer operated in the reflectron-delayed extraction mode. Spectra were calibrated internallyusing trypsin autodigestion products. A nonredundant
protein sequence database was used for searches. Trypticfragment masses (m/z ) were submitted to the Peptide MassFingerprint search form andanalysed using the MASCOT
search engine (Matrix Science, http://www.matrixscience.com) and the NCBI primary sequence databases withparameters including: cysteine as S-carbamidomethylderivative and maximum peptide mass error of 50 p.p.m.[47,48]. Protein identification was determined by thegeneration of a high Mowse score (P , 0.05).
Purification of the radiolabeled 70-kDa heat shockprotein
Human oesophageal epithelial cells from 60 biopsies derivedfrom 20 patients (approximately 2 mg of total protein) wereheat shocked, radiolabeled with [35S]methionine, and lysedas described above. Soluble lysates were pooled and diluted20-fold in buffer B (20% glycerol; 25 mM Hepes, pH 7.6;5 mM dithiothreitol; 0.1% Triton X-100; and 1 mM
benzamidine) to a conductivity equivalent to 20 mM KCland the diluent was applied to a Q50 ion-exchange column(1 mL) equilibrated in buffer B containing 20 mM KCl. Thebound protein was eluted with a 40-col.-vol. linear gradientfrom 20 mM to 500 mM KCl in buffer B at a 1-col.-vol. per hflow rate. Fractions (0.5 mL) were collected and localiz-ation of SEP70 protein during purification was performed asdescribed below by fluorography. Following electrophor-esis, polyacrylamide gels were fixed with 50% methanol/10% acetic acid and impregnated with Amplify (Amersham)for 30 min prior to drying. Gels from the purification wereexposed to X-ray film at 270 8C for 2 months to visualizethe radiolabeled stress protein.
Generation of monoclonal antibodies to the 70-kDaheat shock protein
The column fraction containing the bulk of the 70-kDaheat shock protein (named SEP70 protein) was used as anantigen to immunize mice for development of hybridomacell lines to the protein using standard procedures [23].Purified fractions of SEP70 protein were injected into twomice (1 mg per injection � three injections) and spleeno-cytes were fused to SP2 myeloma cells to generatehybridoma cells. Individual hybridoma supernatants werefirst screened for IgG and then tested for antibodies thatbind to purified SEP70 protein by denaturing immuno-blots. Following hybridoma cloning, the antibody insupernatants was used directly for immunoblotting or forcell staining as indicated in the figure legends. Theantibody used in this report to SEP70 protein is namedMB-H2 (Moravian Biotechnologies).
Immunochemical methods
Following electrophoresis, protein from polyacrylamidegels was transferred to nitrocellulose (Amersham; HybondC) and protein blots were blocked by incubating with NaCl/Pi/Tween/milk (NaCl/Pi/0.1% Tween-20/5% milk), incu-bated with purified monoclonal antibodies (2 mg:mL21 inNaCl/Pi/Tween/milk) overnight at 4 8C, and developed withanti-(mouse IgG) secondary Ig conjugated to peroxidaseusing ECL (Amersham). Wistar rats (adult male) weresubjected to whole body hyperthermia by first elevating the
q FEBS 2001 Stress protein induction in epithelial cells (Eur. J. Biochem. 268) 5345

internal body temperature from 38 to 42 8C over a period of15 min using a fan-assisted incubator, as indicatedpreviously [17,24]. Lysates were prepared using lysis bufferfrom the indicated tissues of animals, where body tem-perature had re-equilibrated to normal temperature 1 h or 2 hafter the hyperthermia regime, and were a gift from B.Vojtesek, Masaryk Cancer Institute, Czech Republic. Themonoclonal antibodies were obtained as indicated: mono-clonal antibody to HSP70 protein (SPA-710) was obtainedfrom Stressgen; the monoclonal antibody with broad cross-reactivity to HSP70; HSC70; and HSP70-2 (MB.H1) wasgenerated as described previously [19].
Cell staining
Using paraffin embedded human tissue blocks, 4- to 6-mmsections were cut, mounted on slides, de-waxed in xyleneand rehydrated in decreasing alcohol concentrations. Endo-genous peroxidase was blocked by treatment with 1%hydrogen peroxide in NaCl/Pi for 20 min After washing inNaCl/Pi, the sections were blocked by treatment with 5%nonfat dried milk in NaCl/Pi for 60 min The MB-H2monoclonal antibody in hybridoma supernatant was appliedat a dilution 1 : 40 for 1 h at room temperature. After threewashes in NaCl/Pi, the biotinylated secondary antibody/peroxidase–streptavidin complex was applied and incu-bated for 60 min at room temperature and washed inNaCl/Pi. The color reaction was developed using a diamino-benzidine peroxide solution (0.2% diaminobenzidine in0.2% hydrogen peroxide in NaCl/Pi for 5 min). Finally, afterwashing in water and 70% ethanol, the nuclei werecounterstained in Harris’ haemotoxylin, washed in water,dehydrated, cleared and mounted.
Development of a cultured cell model to study low pHinduction of SEP70 protein
A cancer cell line derived from squamous epithelium (HeLacells) were grown at 37 8C in DMEM medium containing10% serum. Cells were exposed to low pH as describedpreviously using methods to demonstrate p53 induction [25]and glucose starvation was performed using DMEM medialacking glucose (Gibco). Whole cell lysates from HeLacells were prepared by adding detergent in lysis Buffer[1% IGEPAL (Sigma); 50 mM Hepes (pH 7.6); 5 mM
dithiothreitol; 0.4 M KCl; 1 mM benzamidine, and including50 mM NaF, and 120 nM okadaic acid to preserve phospho-epitopes] to cell pellets and incubating on ice for 15 minFollowing centrifugation at 10 000 g for 10 min at 4 8C, theextracted protein was quantitated using the method ofBradford [21].
R E S U L T S
Heat shock stress in normal squamous epitheliumselectively decreases HSP70 protein levels
Following a 15-min heat shock at 55 8C, the ex vivoincubation of squamous epithelial biopsies at 37 8C for fourhours (Fig. 1A, lane 3) gave rise to a striking reduction inthe steady-state levels of HSP70 protein (compared toFig. 1A, lanes 1 and 2). The addition of the heat shock
mimetic, arsenite, did not alter the steady-state levels ofHSP70 (Fig. 1A, lanes 5 and 6 vs. lane 4), indicating that thedown-regulation of HSP70 protein is not a nonspecific toxiceffect of damage to the epithelial cells. In addition, theex vivo incubation of squamous epithelial biopsies with 4%(v/v) ethanol for 4 h (Fig. 1A, lane 8) also gave rise to areduction in the steady-state levels of HSP70 protein(compared to Fig. 1A, lanes 7 and 4), indicating that stressesother than thermal injury can result in the down-regulationof HSP70 protein in this cell type.
This down-regulation of HSP70 protein after heat shockor ethanol stress to a cell is relatively unique and a series ofcontrols was performed in order to further clarify thespecificity of the response. First, it may be possible that thesquamous epithelial cells are simply killed by the rathersevere heat shock of 55 8C resulting in degradation of
Fig. 1. HSP70 protein is down-regulated after heat shock and
ethanol treatment in squamous epithelium. (A) Examination of
HSP70 protein levels after stress. Oesophageal squamous epithelial
biopsies were incubated at 37 8C as indicated in the Experimental
procedures. In order to induce the stress response, epithelial biopsies
were either: left untreated and incubated at 37 8C (lane 1 and lanes 4),
subjected to a 15-min heat shock at 45 8C (lane 2) or 55 8C (lane 3);
transferred to fresh media containing 250 mM sodium arsenite (lane 5)
or 500 mM sodium arsenite (lane 6); or transferred to fresh media
containing 2% (v/v) ethanol (lane 7) or 4% (v/v) ethanol (lane 8). Four
hours after the heat shock, and 4 h after the after arsenite or ethanol
incubation, the cells were frozen in liquid nitrogen and subsequently
processed in order to examine HSP70 protein levels by immunoblotting
with the HSP70-specific monoclonal antibody (Stressgen), as indicated
in the Experimental procedures. A ‘1’ above the lanes indicate the type
of stress imposed on the cells and HSP70 protein is marked by an
asterisk. (B) Examination of HSC70 protein levels after stress.
Oesophageal squamous epithelial biopsies (lanes 2–5 and 7–10) were
incubated at 37 8C as indicated in the Experimental procedures.
Epithelial biopsies were either: left untreated and incubated at 37 8C
(lane 2 and lane 7), subjected to a 15-min heat shock at 42 8C (lane 3
and 8), 45 8C (lane 4 and 9) or 55 8C (lane 5 and 10), and transferred to
fresh media. Four hours after the heat shock, the cells were frozen in
liquid nitrogen and subsequently processed in order to examine HSP70
protein levels by immunoblotting with the HSP70-specific monoclonal
antibody (lanes 1–5) or immunoblotted with the antibody MB-H1 that
can recognize HSC70 (lanes 6–10), as indicated in the Experimental
procedures. The immunoreactivity of purified HSP70 (lane 1) and
HSC70 (lane 6) are displayed as markers.
5346 A. Yagui-Beltran et al. (Eur. J. Biochem. 268) q FEBS 2001

total protein. However, the human oesophageal epitheliumin vivo can be subjected to temperature stresses of up to65 8C and higher [46], suggesting that it may have a potentstress-response survival system. In addition, the drinking ofvery hot beverages, but not ethanol intake or smoking, is animportant risk factor for cancer progression in theoesophagus [26]. The cancer risk associated with heatshock of the oesophagus may be due to thermal injuryinduced on the cells and associated tissue regeneration thatoccurs in the presence of refluxed acid from the stomach.These data highlight the unusual heat stresses imposed uponthe human oesophagus. The use of ex vivo organ culturescontaining human oesophageal biopsies was pioneeredpreviously, where it was shown that the cells remain viableas defined by trypan-blue exclusion [19,45]. In addition, theheat-shocked or ethanol-treated cells can synthesize adistinct, but overlapping set of polypeptides after stress (seebelow), indicating that the cells are viable four hours afterinjury with respect to stress-induced protein synthesis. Theepithelial cells can survive in culture and continue toundergo protein synthesis for at least 24 h after heat shock[19].
A second concern with respect to the reduction in HSP70protein levels after heat stress to squamous epithelial cells isthat it may simply be a consequence of total proteinreduction. To address this, we also immunoblotted for therelated HSP70 family member, HSC70 (Fig. 1B). Althoughincreasing the temperatures from 37 8C up to 55 8C resultedin a gradual reduction in the steady-state levels of theclassically stress-induced HSP70 (Fig. 1B, lanes 2–5), thesteady-state levels of the constitutive HSC70 protein familymember remained unchanged (Fig. 1B, lanes 7–10).Additionally, the major 70-kDa squamous epithelial cell-specific heat stress protein we have named SEP70 proteincan be induced after ex vivo heat shock of squamousbiopsies (see below). Together, these data demonstrate that arelatively specific down-regulation of HSP70 protein canoccur in this cell type after heat stress.
A third concern with respect to the reduction in HSP70protein levels after heat stress to squamous epithelial cells isthat we have not demonstrated that the immunochemicalassay we have used can actually measure the classicincreases in HSP70 protein levels in normal tissue after heatshock. Previous studies had demonstrated the conditions forinducing a heat shock response in vivo in the Wistar ratstrain [17,24] and this assay was used to determine whetherHSP70 protein increases were in fact a consequence of heatshock in many different organs. Thus, whole body hyper-thermia at 42 8C in Wistar rats gave rise to an increase inHSP70 protein in the lung, liver, kidney, spleen, and adrenalgland 1 h or 2 h after a 15-min heat shock (Fig. 2, lanes 3and 2 vs. lane 1). The temperature gradient shows that at42 8C there is a reduction in the steady-state levels of HSP70in squamous epithelium ex vivo (Fig. 1B), which is withinthe range of the temperatures achieved during in vivohyperthermia. The small intestine, thymus, cardiac tissue,and skeletal muscle also exhibited an increase in HSP70protein levels after heat shock (data not shown). This alsoindicates that the squamous epithelium exhibits a relativelyunusual type of heat shock response involving the downregulation of HSP70 protein and prompted an analysis of thetypes of proteins that are in fact synthesized after heat shockin this cell type.
Three abundant polypeptides are selectively synthesizedin heat shocked squamous epithelium
An investigation was subsequently carried out to determinewhether squamous epithelium could in fact synthesizespecific sets of polypeptides after stress. The ex vivo treat-ment of squamous epithelial biopsies with heat shock,ethanol, or arsenite gave rise to an overlapping but distinctchange in the pattern of protein synthesis. In all three cases,the steady-state level of protein synthesis after stress wasmonitored by radiolabeling the cells with [35S]methionineand under these conditions protein synthesis was reduced bygreater than 95% (data not shown). A 30-min pulse-labelwith [35S]methionine in cells incubated for 4 h in 2% or 4%(v/v) ethanol resulted in the selective synthesis of a 70-kDaand a 78-kDa polypeptide, in comparison to untreated orarsenite treated cells (Fig. 3, lanes 3 and 4 vs. lanes 1 and 2).The 4% ethanol-exposed cells also resulted in the synthesisof a 33-kDa protein that was not observed in untreated,arsenite treated, or heat shocked cells (Fig. 3, lane 4 vs. allother lanes).
Focusing on heat shock, since it may be in part acontributing factor to cancer development [26], a set of threekey proteins exhibiting molecular masses of 58, 70, and78 kDa are reproducibly elevated when biopsies fromdifferent patients were analyzed after a 45 8C or a 55 8C heatshock by radiolabeling of cells with [35S]methionine (Fig. 3,lanes 5 and 6, vs. lanes 1–4). There is therefore somesimilarity between ethanol and heat shocked squamousepithelium: (a) HSP70 protein is down regulated (Fig. 1)
Fig. 2. HSP70 protein accumulates after heat shock in many tissue
types. Tissues were subjected to hyperthermia for 15 min as indicated
in the Experimental procedures and lysates were processed for
immunoblotting using the HSP70-specific monoclonal antibody
(Stressgen) in order to quantitate changes in HSP70 protein levels, as
reported in the legend for Fig. 1. From the top to bottom panels, lung,
liver, kidney, spleen, and adrenal gland were either untreated (lane 1) or
harvested 1 h (lane 2) or 2 h (lane 3) after the heat shock. A ‘ 1’above
the lane indicates whether stress was imposed on the cells and HSP70
protein is marked by an asterisk.
q FEBS 2001 Stress protein induction in epithelial cells (Eur. J. Biochem. 268) 5347
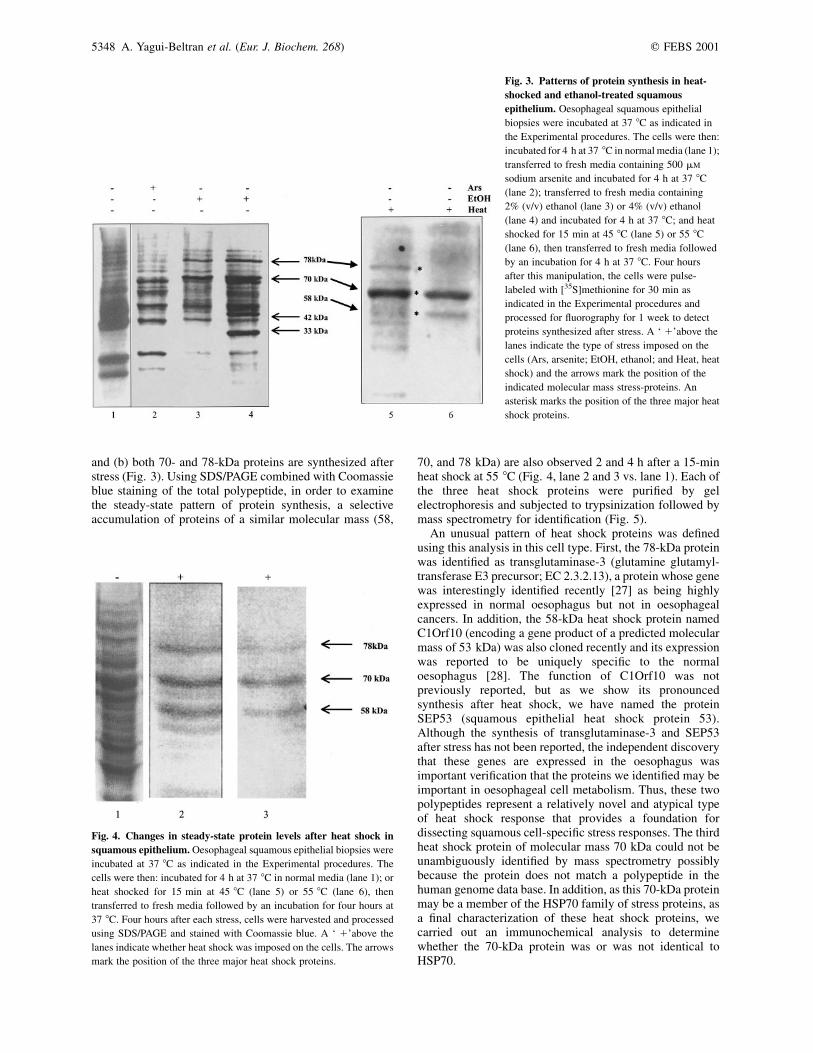
and (b) both 70- and 78-kDa proteins are synthesized afterstress (Fig. 3). Using SDS/PAGE combined with Coomassieblue staining of the total polypeptide, in order to examinethe steady-state pattern of protein synthesis, a selectiveaccumulation of proteins of a similar molecular mass (58,
70, and 78 kDa) are also observed 2 and 4 h after a 15-minheat shock at 55 8C (Fig. 4, lane 2 and 3 vs. lane 1). Each ofthe three heat shock proteins were purified by gelelectrophoresis and subjected to trypsinization followed bymass spectrometry for identification (Fig. 5).
An unusual pattern of heat shock proteins was definedusing this analysis in this cell type. First, the 78-kDa proteinwas identified as transglutaminase-3 (glutamine glutamyl-transferase E3 precursor; EC 2.3.2.13), a protein whose genewas interestingly identified recently [27] as being highlyexpressed in normal oesophagus but not in oesophagealcancers. In addition, the 58-kDa heat shock protein namedC1Orf10 (encoding a gene product of a predicted molecularmass of 53 kDa) was also cloned recently and its expressionwas reported to be uniquely specific to the normaloesophagus [28]. The function of C1Orf10 was notpreviously reported, but as we show its pronouncedsynthesis after heat shock, we have named the proteinSEP53 (squamous epithelial heat shock protein 53).Although the synthesis of transglutaminase-3 and SEP53after stress has not been reported, the independent discoverythat these genes are expressed in the oesophagus wasimportant verification that the proteins we identified may beimportant in oesophageal cell metabolism. Thus, these twopolypeptides represent a relatively novel and atypical typeof heat shock response that provides a foundation fordissecting squamous cell-specific stress responses. The thirdheat shock protein of molecular mass 70 kDa could not beunambiguously identified by mass spectrometry possiblybecause the protein does not match a polypeptide in thehuman genome data base. In addition, as this 70-kDa proteinmay be a member of the HSP70 family of stress proteins, asa final characterization of these heat shock proteins, wecarried out an immunochemical analysis to determinewhether the 70-kDa protein was or was not identical toHSP70.
Fig. 3. Patterns of protein synthesis in heat-
shocked and ethanol-treated squamous
epithelium. Oesophageal squamous epithelial
biopsies were incubated at 37 8C as indicated in
the Experimental procedures. The cells were then:
incubated for 4 h at 37 8C in normal media (lane 1);
transferred to fresh media containing 500 mM
sodium arsenite and incubated for 4 h at 37 8C
(lane 2); transferred to fresh media containing
2% (v/v) ethanol (lane 3) or 4% (v/v) ethanol
(lane 4) and incubated for 4 h at 37 8C; and heat
shocked for 15 min at 45 8C (lane 5) or 55 8C
(lane 6), then transferred to fresh media followed
by an incubation for 4 h at 37 8C. Four hours
after this manipulation, the cells were pulse-
labeled with [35S]methionine for 30 min as
indicated in the Experimental procedures and
processed for fluorography for 1 week to detect
proteins synthesized after stress. A ‘ 1’above the
lanes indicate the type of stress imposed on the
cells (Ars, arsenite; EtOH, ethanol; and Heat, heat
shock) and the arrows mark the position of the
indicated molecular mass stress-proteins. An
asterisk marks the position of the three major heat
shock proteins.
Fig. 4. Changes in steady-state protein levels after heat shock in
squamous epithelium. Oesophageal squamous epithelial biopsies were
incubated at 37 8C as indicated in the Experimental procedures. The
cells were then: incubated for 4 h at 37 8C in normal media (lane 1); or
heat shocked for 15 min at 45 8C (lane 5) or 55 8C (lane 6), then
transferred to fresh media followed by an incubation for four hours at
37 8C. Four hours after each stress, cells were harvested and processed
using SDS/PAGE and stained with Coomassie blue. A ‘ 1’above the
lanes indicate whether heat shock was imposed on the cells. The arrows
mark the position of the three major heat shock proteins.
5348 A. Yagui-Beltran et al. (Eur. J. Biochem. 268) q FEBS 2001

The squamous-cell specific 70-kDa heat shock protein andHSP70 exhibit distinct expression profiles
In order to generate monoclonal antibodies to the majorsquamous-cell specific 70 kDa stress protein (now namedSEP70 for squamous epithelial heat shock protein 70), apurification scheme was first developed to enrich for thispolypeptide from normal epithelial biopsies obtained frompatients undergoing endoscopic surveillance for gastro-reflux disease. Although purification of the 70-kDa SEP70protein based on its molecular mass is possible, given thatother proteins of a similar size may copurify with SEP70,like HSP70, we developed a radioactive assay to monitorits elution profile during chromatography. As HSP70 proteinis down-regulated by heat shock and presumably notsynthesized (Fig. 1), this tagging assay should enrich forSEP70 and reduce contamination by HSP70. Biopsies werefirst pulse-labeled for 4 h with [35S]methionine after a15-min heat shock in order to ‘tag’ the stress-induced SEP70protein. Approximately 60 biopsies were collected, heat-shocked, pulse-labeled, and frozen over a period of 18months, following which radioactively labeled SEP70 proteinwas then detected during the purification using SDS/PAGEand fluorography in order to localize its elution peak. The twomajor radiolabeled and heat induced polypeptides (SEP70and SEP53) eluted differentially at a conductivity equivalentto buffer B containing approximately 75 mM KCl, withthe bulk of C1Orf10 protein (SEP53) eluting prior to thatof SEP70 protein (Fig. 6A, lanes 3–5; upper arrow). ACoomassie blue stained SDS polyacrylamide gel shows therelative purity of the individual fractions in this region of thegradient (Fig. 6B, lanes 3–5). The yield of SEP70 proteinwas approximately 20% with 7 mg acquired from 2 mg oflysate. An immunoblot was used to confirm that SEP70protein (Fig. 1 A and 1B, lanes 3–5) is not identical to the
major stress-70 family isoforms, based on its lack of bindingto monoclonal antibody MB-H1 (Fig. 6C, lanes 1–7),an antibody that cross-reacts with HSP70, HSP70-2, andHSC70 proteins. Five stable hybridoma cell lines thatproduce monoclonal antibodies to SEP70 protein weregenerated and reported here is the characterization of one ofthe monoclonal antibodies that has been classified as IgG1(MB-H2). An immunochemical blot with MB-H2 demon-strates its specificity for SEP70 protein eluting from theQ-50 column (Fig. 6D, lanes 3–5) and its lack of binding toHSP70 protein (Fig. 7).
Further immunoblotting analysis with MB-H2 shows thatit detects from crude lysates derived from biopsies of humanoesophageal epithelium a single protein at a molecular massof 70 kDa (Fig. 7A, lane 1), and that it does not bind toHSC70 or HSP70 proteins (Fig. 7A, lanes 3 and 4), indicat-ing that MB-H2 has a high specificity for SEP70 protein. Inaddition, after ex vivo heat shock, the steady-state SEP70protein levels increase (Fig. 7A, lane 2 vs. lane 1 and lanes 6vs. 5), under conditions where HSP70 protein levels decreaseand HSC70 protein levels remain constant (Fig. 1B). Asimilar mutually exclusive expression of SEP70 and HSP70can be seen in cultured cells (see below). Immunohisto-chemical cell staining of paraffin-embedded blocks wasperformed to determine the cell types where SEP70 proteinwas expressed within the epithelial biopsy. A strikingstaining pattern was observed for SEP70 protein in thesuprabasal layer of the normal oesophageal epithelium andSEP70 protein was predominantly cytoplasmic with somenuclear staining (Fig. 7B).
In addition to heat shock, low extracellular pH may be animportant factor that effects the stress response in epithelialcells. However, we have not been able to demonstrate thatpH stress can induce SEP70 in ex vivo biopsies (data notshown), possibly because the cells have been exposed to
Fig. 5. Mass spectrometric identification of the
stress proteins in squamous epithelium. The
observed molecular mass fragments obtained using
a Perseptive Biosystems Voyager-DE STR
Biospectrometry Workstation were matched to the
indicated polypeptides using Matrix Science
Mascot Search, as indicated in the Experimental
procedures. The peptide sequence matching the
observed molecular mass and the corresponding
amino-acid positions are indicated in the Figure.
(A) 78-kDa protein (glutamine glutamyltransferase
E3 precursor); (B) 53-kDa protein
(C1Orf10¼SEP53).
q FEBS 2001 Stress protein induction in epithelial cells (Eur. J. Biochem. 268) 5349

maximal levels of acid reflux prior to endoscopy. As such,cultured cell lines were also screened to determine whetherlow extracellular pH affects SEP70 protein expression. Thecell line that was best able to respond was a HeLa cell,which is a tumour cell line derived originally from cervicalsquamous epithelium. Although the relationship betweenHeLa cells and human squamous epithelium is not clear, theuse of human biopsies as a routine model is very com-plicated and a proliferating cell model is required that atleast mimics the biochemical observations that HSP70 andSEP70 expression responds differently in squamousepithelium. Although reduction in the pH of the mediafrom 7.4 to 7.0 had little effect on SEP70 levels or HSP70protein levels in HeLa cells (data not shown), furtherreduction in extracellular pH led to a striking and relativelyrapid induction of SEP70 protein (Fig. 8A, lanes 1–4, toppanel) and down regulation of HSP70 protein (Fig. 8A,lanes 1–4, bottom panel). Such mutually exclusiveexpression of SEP70 and HSP70 is observed in stressed
human oesophageal biopsies (Fig. 7 vs. Fig. 1) and provideda unique model to dissect the acid-dependent signalingpathways that regulate SEP70 protein synthesis. Four hoursfollowing the acidification of the media, SEP70 proteinlevels declined back to basal levels and the HSP70 proteinlevels rose to preacidification levels (Fig. 8A, lane 4 vs.lane 3). This phenomenon is probably due to the changesin the extracellular pH that occurs in bicarbonated tissueculture media in a CO2-controlled incubator. When thetissue culture media is initially buffered to a lower pH, thereis a relatively rapid drift back to neutral pH when placed inthe incubator due to the bicarbonate buffering mechanism(Fig. 8C). It is presumably after the re-neutralization of themedia that the SEP70 protein declines back to its normallevels and HSP70 down-regulation is reversed.
The exposure of cells to a variety of DNA damagingagents did not induce SEP70 in cultured cells (data notshown), distinguishing SEP70 from other commonHSP70-responsive stress pathways. Given that the pleio-tropic effects of acid on cell viability may involve alterationsin energy metabolism we investigated whether metabolicstarvation also affected SEP70 expression. A slower, butprolonged induction of SEP70 protein was achieved byeither glucose starvation (Fig. 8B, lanes 1–4, top panel) orpyruvate starvation (data not shown). A similar down-regulation of HSP70 occurred when SEP70 was induced(Fig. 8B, lanes 1–4, bottom panel). These latter resultsindicate that SEP70 protein belongs to the glucoseregulated family of stress proteins including GRP78,HSP75, and the 70-kDa redox protein ERP72/CaBP2.Antibodies to these GRP family members, however, failedto cross-react with SEP70 and SEP70-specific monoclonalantibodies did not react with these isoforms (data notshown).
Variable expression of SEP70 protein in Barrett’sepithelial cells
Having established that SEP70 is a functional acid-respon-sive stress protein of the glucose-regulated family ofproteins that is immunochemically distinct from HSP70protein, we hoped to use our panel of anti-SEP70 mono-clonal antibodies to determine whether HSP70 and SEP70protein exhibit altered expression in Barrett’s epithelium, apreneoplastic lesion on the cancer progression sequenceleading to adenocarcinoma. Two distinct outcomes werepredicted from the onset regarding SEP70 protein and/orHSP70 protein expression in Barrett’s epithelium. First, asBarrett’s epithelium is a tissue where p53 mutations arebeginning to be observed, a down-regulation in the majoracid-responsive stress-protein SEP70 and/or HSP70 in thischemically harsh microenvironment is postulated to contri-bute in part to an accelerated rate of oxidative cell damageover the patients life. Alternatively, stresses imposed uponBarrett’s epithelial cells may up-regulate SEP70 protein orHSP70 protein in order to confer ‘survival’ advantage toproliferative cells growing under chronic exposure to acid/bile reflux and/or carcinogens from food intake. Either ofthese two possibilities are consistent with the known role ofstress proteins as anti-apoptotic factors (contributing tosurvival of damaged neoplastic cells) [16] or as factors thatcan maintain cell integrity (whose loss contributes toaccelerated cell damage) [18].
Fig. 6. Q-50 Sepharose chromatographic purification of SEP70
protein. SEP70 protein was purified from biopsied and heat-shocked
human squamous cell epithelium as indicated in the Experimental
procedures. Soluble lysates were diluted, applied to an ion-exchange
column, and the bound protein was eluted as indicated in the
Experimental procedures. Individual fractions were assayed by
electrophoresis as follows: (A) fluorographic exposure for 2 months
to detect radioactive SEP70 (top arrow) and SEP53 (bottom arrow)
proteins using an 8% polyacrylamide gel to enhance electrophoretic
separation of SEP53 from SEP70; (B) Coomassie blue stained gel of
column fractions using 12% SDS polyacrylamide gel to detect the bulk
of protein; (C) an immunochemical blot with antibodies to HSP70/
HSC70 (MB-H1) using the protein in the column fractions; and (D) an
immunochemical blot with a monoclonal antibody generated to SEP70
protein (MB-H2) using the protein in the column fractions. The samples
are as follows; lanes 1–7 (fractions 1–7 from 60 mM to 90 mM KCl
which eluted from the ion exchange column during the gradient and
which contain the peak of radioactive SEP70 protein). The ‘*’ in
fraction 4 (lane 4) marks the peak elution of SEP70 protein.
5350 A. Yagui-Beltran et al. (Eur. J. Biochem. 268) q FEBS 2001
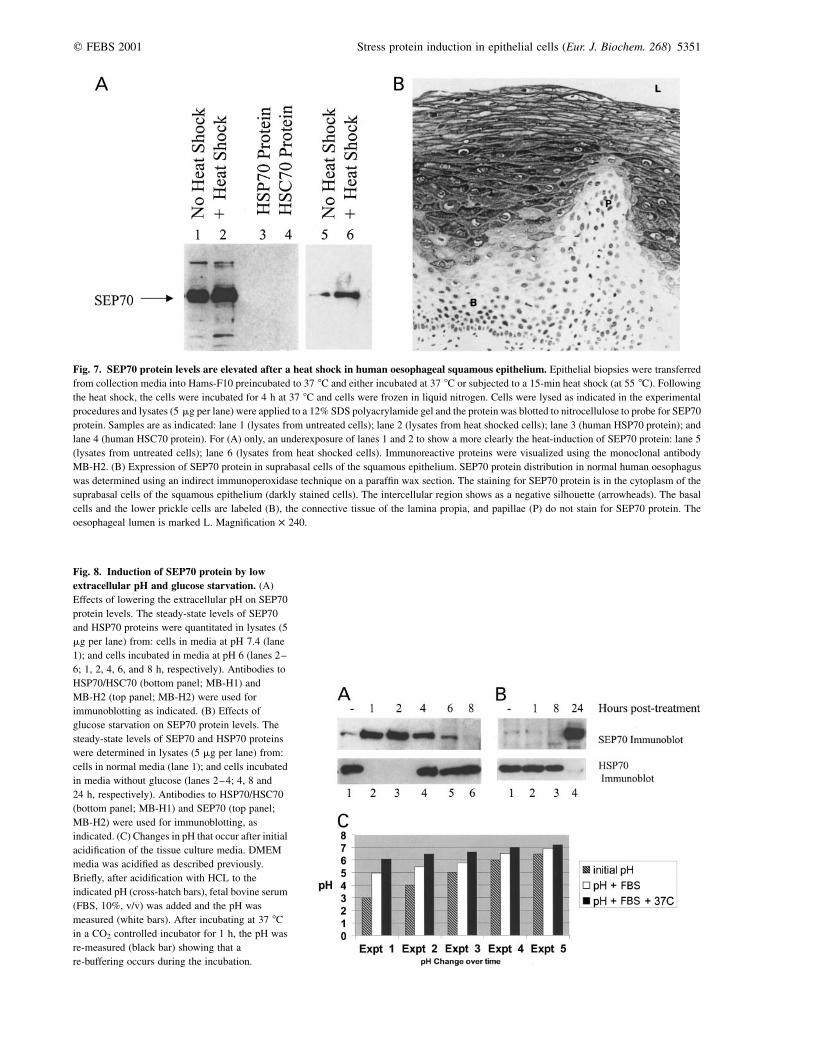
Fig. 7. SEP70 protein levels are elevated after a heat shock in human oesophageal squamous epithelium. Epithelial biopsies were transferred
from collection media into Hams-F10 preincubated to 37 8C and either incubated at 37 8C or subjected to a 15-min heat shock (at 55 8C). Following
the heat shock, the cells were incubated for 4 h at 37 8C and cells were frozen in liquid nitrogen. Cells were lysed as indicated in the experimental
procedures and lysates (5 mg per lane) were applied to a 12% SDS polyacrylamide gel and the protein was blotted to nitrocellulose to probe for SEP70
protein. Samples are as indicated: lane 1 (lysates from untreated cells); lane 2 (lysates from heat shocked cells); lane 3 (human HSP70 protein); and
lane 4 (human HSC70 protein). For (A) only, an underexposure of lanes 1 and 2 to show a more clearly the heat-induction of SEP70 protein: lane 5
(lysates from untreated cells); lane 6 (lysates from heat shocked cells). Immunoreactive proteins were visualized using the monoclonal antibody
MB-H2. (B) Expression of SEP70 protein in suprabasal cells of the squamous epithelium. SEP70 protein distribution in normal human oesophagus
was determined using an indirect immunoperoxidase technique on a paraffin wax section. The staining for SEP70 protein is in the cytoplasm of the
suprabasal cells of the squamous epithelium (darkly stained cells). The intercellular region shows as a negative silhouette (arrowheads). The basal
cells and the lower prickle cells are labeled (B), the connective tissue of the lamina propia, and papillae (P) do not stain for SEP70 protein. The
oesophageal lumen is marked L. Magnification � 240.
Fig. 8. Induction of SEP70 protein by low
extracellular pH and glucose starvation. (A)
Effects of lowering the extracellular pH on SEP70
protein levels. The steady-state levels of SEP70
and HSP70 proteins were quantitated in lysates (5
mg per lane) from: cells in media at pH 7.4 (lane
1); and cells incubated in media at pH 6 (lanes 2–
6; 1, 2, 4, 6, and 8 h, respectively). Antibodies to
HSP70/HSC70 (bottom panel; MB-H1) and
MB-H2 (top panel; MB-H2) were used for
immunoblotting as indicated. (B) Effects of
glucose starvation on SEP70 protein levels. The
steady-state levels of SEP70 and HSP70 proteins
were determined in lysates (5 mg per lane) from:
cells in normal media (lane 1); and cells incubated
in media without glucose (lanes 2–4; 4, 8 and
24 h, respectively). Antibodies to HSP70/HSC70
(bottom panel; MB-H1) and SEP70 (top panel;
MB-H2) were used for immunoblotting, as
indicated. (C) Changes in pH that occur after initial
acidification of the tissue culture media. DMEM
media was acidified as described previously.
Briefly, after acidification with HCL to the
indicated pH (cross-hatch bars), fetal bovine serum
(FBS, 10%, v/v) was added and the pH was
measured (white bars). After incubating at 37 8C
in a CO2 controlled incubator for 1 h, the pH was
re-measured (black bar) showing that a
re-buffering occurs during the incubation.
q FEBS 2001 Stress protein induction in epithelial cells (Eur. J. Biochem. 268) 5351

A collection of Barrett’s biopsies and normal squamousepithelium were first examined microscopically to confirmmorphological character. In addition, proteomic analysiswas performed in order to fingerprint these tissue typesmolecularly and to provide us with a better method fornormalization of normal and Barrett’s epithelial biopsiesbetween patients. Lysates from normal and Barrett’sepithelial cells were then separated using two-dimensionalgel electrophoresis to identify proteins that are differentially
expressed (data not shown). The identification by massspectrometry of one the major proteins specificallyexpressed in Barrett’s epithelial cells as anterior gradient-2protein [29] was confirmed by immunoblotting (Fig. 9A,lanes 4–6 vs. lanes 1–3). Thus, these molecular analysesprovided a fingerprint of candidate polypeptides of knownidentity whose differential expression provide us an internalprofile to normalize the integrity and identity of the differentBarrett’s epithelial cells and normal biopsies from differentpatients. Additionally, the mutually exclusive expression ofthese marker proteins makes it certain that normalepithelium does not contaminate the Barrett’s epithelialcells and vice versa.
The same biopsied material as that used in experimentsdescribed in Fig. 9A from normal and Barrett’s epitheliumwere used to analyze SEP70 and HSP70/HSC70 proteinlevels by denaturing immunoblotting. Interestingly, HSP70/HSC70 proteins are not expressed in the Barrett’s epithelialcells (Fig. 9B, lanes 4–6 vs. lanes 1–3), providing evidencethat stress-protein down-regulation can occur in preneo-plastic cells of the oesophagus in a microenvironment wherestress-protein protective pathways will presumably beimportant to maintain cell integrity and minimize injury.In contrast to hAG-2 and HSP70/HSC70 proteins that areuniquely expressed in one or the other epithelium, SEP70expression in Barrett’s epithelium was divided into threedistinct classes (Fig. 9C, lanes 4–6). One class (27%) didnot synthesize detectable levels of SEP70 protein at all(Fig. 9C, lane 6). A second class (27%) produced largeamounts of SEP70 protein (Fig. 9C, lane 5), which cannotbe explained by contamination of normal tissue in theBarrett’s biopsy, as this biopsy expresses high levels ofhAG-2 (Fig. 9A, lane 5) and no HSP70/HSC70 (Fig. 9B,lane 5). A third group (45%) expresses very small amountsof SEP70 protein along with a higher molecular mass cross-reacting species (Fig. 9C, lane 4). These data distinguishSEP70 protein from HSP70 protein in Barrett’s epithelialcell populations. Additionally, this indicates that one the ofthe major squamous epithelial heat shock proteins (SEP70)is variably expressed in Barrett’s epithelial cells and identifya stress pathway whose altered expression may play a role inregulating the extent of tissue injury and cancer progression.
D I S C U S S I O N
Although the link between genetic lesions and carcinogen-esis has clearly established the role that oncogene andtumour suppressor mutations play in growth control,growing evidence suggests that environmental damageleading to normal tissue trauma may be associated withearly stages of neoplasia [4,11,12]. The human oesophagealepithelium provides a unique model to study role of tissuecell repair pathways in modifying the oncogenic and tumoursuppressor pathways that regulate proliferation control. Asprimary stress responses are likely to play an important rolein minimizing oesophageal tissue damage and injury, wehave set up a system to study the basic damage responses inhuman epithelial biopsies. Previous results have indicatedthat stress-protein responses in human oesophagealepithelial cells can be studied ex vivo and parameters havebeen defined that permit analysis of the stress responses[19]. Critical to such analyses using tissue biopsies is therelatively narrow window permitting analysis of stress
Fig. 9. Proteomics provides a molecular fingerprint to normalize
the integrity of normal and Barrett’s epithelium: SEP70 and HSP70
protein exhibit distinct expression profiles in Barrett’s epithelial
cells. Three distinct class of Barrett’s epithelium were found based on
differences in SEP70 protein levels. Representative lysates from three
normal squamous epithelial biopsies (5 mg per lane, lanes 1–3 and three
Barrett’s epithelial biopsies, 5 mg per lane, lanes 4–6) were separated
using a one-dimensional polyacrylamide gel. (A) A novel marker
protein to ensure the purity of Barrett’s epithelium: hAG-2 protein is
selectively expressed in Barrett’s epithelial cells. A novel marker
protein specific for Barrett’s epithelium was identified by this lab using
mass spectrometric fingerprinting as anterior gradient protein-2 (hAG-2)
(data not shown). Epithelial lysates were applied to a 12% SDS poly-
acrylamide gel and the protein was blotted to nitrocellulose to probe for
hAG-2 protein levels using a polyclonal antibody that was generated
from the recombinant hAG-2 gene [29]. An asterisk marks the migration
of the 18-kDa protein hAG-2. (B) A marker protein to ensure the purity
of squamous epithelium:HSP70/HSC70 protein is selectively expressed
in normal squamous epithelial cells. Epithelial lysates (5 mg per lane)
from normal (lanes 1–3) and Barrett’s epithelial biopsies (lanes 4–8)
were applied to a 10% SDS polyacrylamide gel and the protein was
blotted to nitrocellulose to probe for HSP70/HSC70 protein levels using
the monoclonal antibody MB-H1. An asterisk marks the migration of
the HSP70/HSC70 protein. (C) SEP70 protein is variably expressed in
Barrett’s epithelial cells. Epithelial lysates (5 mg per lane) from normal
(lanes 1–3) and Barrett’s epithelial biopsies (lanes 4–8) were applied to
a 10% SDS polyacrylamide gel and the protein was blotted to nitro-
cellulose to probe for SEP70 protein levels using monoclonal antibody
MB-H2. The lower asterisk marks the migration of the 70-kDa SEP70
protein and the upper asterisk marks the migration of a larger species
that cross-reacts with the monoclonal antibody to SEP70 protein,
MB-H2.
5352 A. Yagui-Beltran et al. (Eur. J. Biochem. 268) q FEBS 2001

responses before the cells become metabolically damagedex vivo. In addition, the atypical decline in steady statelevels of HSP70 protein after heat shock or ethanol exposurealong with the predominant synthesis of a set of poly-peptides ranging from 58 to 78 kDa (Fig. 3), spurred furtherinvestigation into the basic stress-responses in this cell type.
We first report on the identification of two of the threemajor heat shock proteins in normal squamous epitheliumusing mass spectrometry and the characterization of thethird stress protein that was undefined by this sequencingmethod. Transglutaminase-3 is an enzyme that can catalyzethe cross-linking of peptide-bound glutamine residues toprimary amines [30]. Intriguingly, the cross-linking ofinterleukin-2 by transglutaminase can also activate celldeath by p53 [31], suggesting that this stress protein willbe linked to tumour suppression by p53. Consistent withthis, a recent study has shown that transglutaminase-3 is notexpressed in over 90% of oesophageal cancers, compared tonormal mucosa from the same patient, suggesting that lossof its function can be associated with cancer development[27]. In addition, a new calcium-binding protein of 146amino acids, named calmodulin-like skin protein (CLSP),was shown to be 100% identical to transglutaminase-3 [32].Thus, our identification of this enzyme as a stress proteinsuggests an involvement of calcium-dependent andp53-dependent signaling pathways in squamous stressresponses.
Interestingly, the 58-kDa stress protein was identified as anovel gene (C1Orf10) that also contains putative calciumbinding motifs. C1Orf10 (SEP53) is highly expressed inoesophageal tissue, but not 15 other glandular tissues sitesand it is not expressed in over 95% of oesophageal cancerexamined [28]. Additionally, in searching human and mouseexpressed sequence tag databases, C1Orf10 (SEP53) wasfound expressed in human cervical squamous epitheliumand in murine skin (data not shown). These data suggest thatC1orf10 (SEP53) is a squamous cell-specific polypeptide.The 70-kDa protein named SEP70 protein could not bedefined at the primary amino-acid level using mass spectro-metry presumably due to the possibility that the SEP70 geneis not yet accessible to translation using the currentdatabase. Because the atypical feature of the squamousepithelium was the down-regulation of HSP70 protein(Fig. 1), we therefore developed set of biochemical criteriato determine whether SEP70 is distinct from or similar toHSP70 protein.
The first key criteria was the mutually exclusiveexpression of SEP70 protein and HSP70 protein in heatshocked squamous epithelium and in acid-treated or glucosestarved cultured cell lines. The rapid synthesis of SEP70after low pH or glucose stress indicates that it is a member ofthe 70-kDa glucose-regulated stress protein family, whilethe striking down-regulation of steady-state levels of HSP70under these same conditions is unusual for this latter stress-protein family member. The classic mammalian stress-70protein (‘heat-shock’) family is composed of constitutiveand inducible homologues that can play distinct roles in themetabolic control of normal or damaged cells anddifferences in peptide-binding specificities amongst variousmembers of the stress-70 protein family suggest thatpeptide-binding determines, in part, the actual function ofthe protein. HSP70 protein is the classic heat inducedpolypeptide that is synthesized in most prokaryotic and
eukaryotic cell types in response to thermal stress [14].HSP70 protein synthesis in mammals is increased aftercellular injury due to activation of the heat shock tran-scription factors HSF1 or HSF2, which bind to and activatetranscription from the hsp70 gene [33]. p75 is a mito-chondrial member of the stress-70 family of proteins thatcan be expressed in response to glucose deprivation orischemia [34]. HSC70 protein is constitutively expressed ineukaryotic cells and, although it presumably participates in avariety of biochemical reactions, it can clearly play a role inthe dissociation of clathrin from coated vesicles permittingvesicle fusion with its target [35]. BiP (Grp78) is localized inthe ER and is required for translocation of newly syn-thesized polypeptides across the ER membrane [36].In addition to the induction of stress-70 homologues (mito-chondrial p75 and Grp78) by glucose starvation, a 70-kDastress protein named Erp72, with homology to proteindisulfide isomerases, is also induced by glucose starvation[37]. Available antibodies to the glucose regulated Erp72,Grp78, and p75 do not cross-react with SEP70 (data notshown) indicating that SEP70 is distinct from theseisoforms.
The second key criteria developed to determine whetherSEP70 protein was distinct from HSP70 involved examiningwhether SEP70 and HSP70 expression in Barrett’sepithelium was distinct. Either SEP70 and/or HSP70 proteinwould be up-regulated in Barrett’s epithelial cells to controlcell integrity and enhance preneoplastic cell survival andclonal expansion in response to chemical stress or SEP70and/or HSP70 protein would be down-regulated in Barrett’sepithelial cells thus possibly contributing to accelerated celldamage. These hypotheses are based on the behavior ofwell-studied heat shock proteins, as organ models in mam-mals have indicated a therapeutic effect of activation of theheat shock pathway on tissue integrity and repair. Wholebody hyperthermia [38,39] or in vivo transfection with theHsp70 gene can protect from myocardial damage in responseto ischemia and repurfusion [40]. Heat shock by perfusion inkidney can protect the tissue from damage induced by warmischemic injury [41,42]. Strikingly, whole body hyperther-mia 20 h prior to a normally lethal dose of radiation canprevent chromatin fragmentation as well as thymocyte andbone marrow cell death [43], indicating that hyperthermiacan be a potent radioprotector from p53-dependentapoptotic pathways in vivo. Our data showing that HSP70protein is not expressed in Barrett’s epithelial cells whilstSEP70 protein is variably expressed first suggests that thetwo pathways are indeed distinct. Second, that fluctuatingSEP70 protein expressions occurs in a microenvironmentwhere chronic chemical stress will presumably play a role indisease progression focuses attention on this signal trans-duction pathway as important for future studies. Our ana-lysis of heat shock protein expression in an oral cancerprogression model has also demonstrated that SEP70 proteinexpression is elevated in the transition for normal tolow-grade dysplasia and HSP70 protein changes itsintracellular localization under the same conditions (G.Ogden, N. Kernohan and T. Hupp, unpublished results). Ourcurrent interest is in determining whether transglutaminase-3,SEP53, and SEP70 stress protein pathways respond to thesame signaling lesion and whether their altered expressioncan further predispose a cell to acid and thermal injury andinactivation of p53 protein or, alternatively, whether forced
q FEBS 2001 Stress protein induction in epithelial cells (Eur. J. Biochem. 268) 5353

re-expression of transglutaminase-3, SEP53, and/or SEP70could reduce the rate of chronic oxidative injury in Barrett’sepithelial cells, which would provide evidence for theimportance of these protective pathway in regulating the rateof cancer progression, as has been observed with theantioxidant GST in animal models of chemical oxidant-induced skin cancer development [44]. Thus, chaperonerepair networks may prove to be important modifiers ofhuman tissue injury or repair and our identification oftransglutaminase-3, SEP53, and SEP70 protein as stressproteins highlights the importance of investigating molecu-lar chaperones as modifiers of disease progression.
A C K N O W L E D G E M E N T S
T. R. H. is supported by the Cancer Research Campaign, the UK
Medical Research Council, the Association for International Cancer
Research, and Moravian Biotechnologies. A. Y. B. was supported by the
Daniels Memorial Cancer Fellowship. S. P. was supported by EMBO
fellowship AST9443.
R E F E R E N C E S
1. Neshat, K., Sanchez, C.A., Galipeau, P.C., Cowan, D.S., Ramel, S.,
Levine, D.S. & Reid, B.J. (1994) Barrett’s esophagus: a model of
human neoplastic progression. Cold Spring Harb. Symp. Quant.
Biol. 59, 577–583.
2. Vogelstein, B. & Kinzler, K.W. (1994) Colorectal cancer and the
intersection between basic and clinical research. Cold Spring Harb.
Symp. Quant. Biol. 59, 517–521.
3. Campomenosi, P., Conio, M., Bogliolo, M., Urbini, S., Assereto, P.,
Aprile, A., Monti, P., Aste, H., Lapertosa, G., Inga, A.,
Abbondandolo, A. & Fronza, G. (1996) p53 is frequently mutated
in Barrett’s metaplasia of the intestinal type. Cancer Epidemiol.
Biomarkers Prev. 5, 559–565.
4. Prevo, L.J., Sanchez, C.A., Galipeau, P.C. & Reid, B.J. (1999)
p53-mutant clones and field effects in Barrett’s esophagus. Cancer
Res. 59, 4784–4787.
5. Stocks, S.C., Pratt, N., Sales, M., Johnston, D., Thompson, A.,
Carey, F.A. & Kernohan, N.M. (2001) Chromosomal imbalances in
gastric and esophageal adenocarcinoma: specific CGH detected
abnormalities segregate with junctional adenocarcinomas. Genes
Chromosomes Cancer 32, 50–58.
6. Tobey, N.A. & Orlando, R.C. (1991) Mechanisms of acid injury to
rabbit esophageal epithelium. Role of basolateral cell membrane
acidification. Gastroenterology 101, 1220–1228.
7. Seto, Y. & Kobori, O. (1993) Role of reflux oesophagitis and acid in
the development of columnar epithelium in the rat oesophagus. Br.
J. Surg. 80, 467–470.
8. Jaskiewicz, K. & De Groot, K.M. (1994) p53 gene mutants
expression, cellular proliferation and differentiation in oesophageal
carcinoma and non-cancerous epithelium. Anticancer Res. 14,
137–140.
9. Spechler, S.J. & Goyal, R.K. (1996) The columnar-lined
esophagus, intestinal metaplasia, and Norman Barrett. Gastro-
enterology 110, 614–621.
10. Reid, B.J., Barrett, M.T., Galipeau, P.C., Sanchez, C.A., Neshat, K.,
Cowan, D.S. & Levine, D.S. (1996) Barrett’s esophagus: ordering
the events that lead to cancer. Eur. J. Cancer Prev. 5 (Suppl. 2),
57–65.
11. Jankowski, J.A., Wright, N.A., Meltzer, S.J., Triadafilopoulos, G.,
Geboes, K., Casson, A.G., Kerr, D. & Young, L.S. (1999)
Molecular evolution of the metaplasia-dysplasia-adenocarcinoma
sequence in the esophagus. Am. J. Pathol. 154, 965–973.
12. Barrett, M.T., Sanchez, C.A., Prevo, L.J., Wong, D.J., Galipeau,
P.C., Paulson, T.G., Rabinovitch, P.S. & Reid, B.J. (1999) Evolution
of neoplastic cell lineages in Barrett oesophagus. Nat. Genet. 22,
106–109.
13. Welch, W.J. (1993) How cells respond to stress. Sci. Am. 268,
56–64.
14. Welch, W.J. & Brown, C.R. (1996) Influence of molecular and
chemical chaperones on protein folding. Cell Stress Chaperones 1,
109–115 (published erratum appears in Cell Stress Chaperones 1,
207).
15. Minowada, G. & Welch, W.J. (1995) Clinical implications of the
stress response. J. Clin. Invest. 95, 3–12.
16. Hupp, T.R. (2000) Development of physiological models to study
stress protein responses. In Stress Responses: Methods and
Protocols (Keyes, S., ed.). Humana Press, Totowa, NJ.
17. Blake, M.J., Gershon, D., Fargnoli, J. & Holbrook, N.J. (1990)
Discordant expression of heat shock protein mRNAs in tissues of
heat-stressed rats. J. Biol. Chem. 265, 15275–15279.
18. Hu, Y., Metzler, B. & Xu, Q. (1997) Discordant activation of stress-
activated protein kinases or c-Jun NH2- terminal protein kinases in
tissues of heat-stressed mice. J. Biol. Chem. 272, 9113–9119.
19. Hopwood, D., Moitra, S., Vojtesek, B., Johnston, D.A., Dillon, J.F.
& Hupp, T.R. (1997) Biochemical analysis of the stress protein
response in human oesophageal epithelium. Gut 41, 156–163.
20. Yeargin, J. & Haas, M. (1995) Elevated levels of wild-type p53
induced by radiolabeling of cells leads to apoptosis or sustained
growth arrest. Curr. Biol. 5, 423–431.
21. Loffler, B.M. & Kunze, H. (1989) Refinement of the Coomassie
brilliant blue G assay for quantitative protein determination. Anal.
Biochem. 177, 100–102.
22. Craig, A.L., Burch, L., Vojtesek, B., Mikutowska, J., Thompson, A.
& Hupp, T.R. (1999) Novel phosphorylation sites of human tumour
suppressor protein p53 at Ser20 and Thr18 that disrupt the binding
of mdm2 (mouse double minute 2) protein are modified in human
cancers. Biochem. J. 342, 133–141.
23. Harlow, E. & Lane, D. (1988) Antibodies: A laboratory Manual.
Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New
York.
24. Blake, M.J., Fargnoli, J., Gershon, D. & Holbrook, N.J. (1991)
Concomitant decline in heat-induced hyperthermia and HSP70
mRNA expression in aged rats. Am. J. Physiol. 260, R663–R667.
25. Williams, A.C., Collard, T.J. & Paraskeva, C. (1999) An acidic
environment leads to p53 dependent induction of apoptosis in
human adenoma and carcinoma cell lines: implications for clonal
selection during colorectal carcinogenesis. Oncogene 18,
3199–3204.
26. Casson, A.G., Tammemagi, M., Eskandarian, S., Redston, M.,
McLaughlin, J. & Ozcelik, H. (1998) p53 alterations in
oesophageal cancer: association with clinicopathological features,
risk factors, and survival. Mol. Pathol. 51, 71–79.
27. Chen, B.S., Wang, M.R., Xu, X., Cai, Y., Xu, Z.X., Han, Y.L. &
Wu, M. (2000) Transglutaminase-3, an esophageal cancer-related
gene. Int. J. Cancer 88, 862–865.
28. Xu, Z., Wang, M.R., Xu, X., Cai, Y., Han, Y.L., Wu, K.M., Wang,
J., Chen, B.S., Wang, X.Q. & Wu, M. (2000) Novel human
esophagus-specific gene c1orf10: cDNA cloning, gene structure,
and frequent loss of expression in esophageal cancer. Genomics 69,
322–330.
29. Thompson, D.A. & Weigel, R.J. (1998) hAG-2, the human homo-
logue of the Xenopus laevis cement gland gene XAG-2, is
coexpressed with estrogen receptor in breast cancer cell lines.
Biochem. Biophys. Res. Commun. 251, 111–116.
30. Hitomi, K., Ikura, K. & Maki, M. (2000) GTP, an inhibitor of
transglutaminases, is hydrolyzed by tissue-type transglutaminase
(TGase 2) but not by epidermal-type transglutaminase (TGase 3).
Biosci. Biotechnol. Biochem. 64, 657–659.
31. Eizenberg, O., Faber-Elman, A., Gottlieb, E., Oren, M., Rotter, V.
& Schwartz, M. (1995) Direct involvement of p53 in programmed
cell death of oligodendrocytes. EMBO J. 14, 1136–1144.
5354 A. Yagui-Beltran et al. (Eur. J. Biochem. 268) q FEBS 2001

32. Mehul, B., Bernard, D., Simonetti, L., Bernard, M.A. & Schmidt,
R. (2000) Identification and cloning of a new calmodulin-
like protein from human epidermis. J. Biol. Chem. 275,
12841–12847.
33. Cotto, J.J. & Morimoto, R.I. (1999) Stress-induced activation of the
heat-shock response: cell and molecular biology of heat-shock
factors. Biochem. Soc. Symp 64, 105–118.
34. Massa, S.M., Longo, F.M., Zuo, J., Wang, S., Chen, J. & Sharp, F.R.
(1995) Cloning of rat grp75, an hsp70-family member, and its
expression in normal and ischemic brain. J. Neurosci. Res. 40,
807–819.
35. Ungewickell, E., Ungewickell, H., Holstein, S.E., Lindner, R.,
Prasad, K., Barouch, W., Martin, B., Greene, L.E. & Eisenberg, E.
(1995) Role of auxilin in uncoating clathrin-coated vesicles. Nature
378, 632–635.
36. Sanders, S.L., Whitfield, K.M., Vogel, J.P., Rose, M.D. &
Schekman, R.W. (1992) Sec61p and BiP directly facilitate
polypeptide translocation into the ER. Cell 69, 353–365.
37. Mazzarella, R.A., Srinivasan, M., Haugejorden, S.M. & Green, M.
(1990) ERp72, an abundant luminal endoplasmic reticulum protein,
contains three copies of the active site sequences of protein
disulfide isomerase. J. Biol. Chem 265, 1094–1101.
38. Donnelly, T.J., Sievers, R.E., Vissern, F.L., Welch, W.J. & Wolfe,
C.L. (1992) Heat shock protein induction in rat hearts. A role for
improved myocardial salvage after ischemia and reperfusion?
Circulation 85, 769–778.
39. Currie, R.W., Tanguay, R.M. & Kingma, J.G. Jr (1993) Heat-shock
response and limitation of tissue necrosis during occlusion/
reperfusion in rabbit hearts. Circulation 87, 963–971.
40. Suzuki, K., Sawa, Y., Kaneda, Y., Ichikawa, H., Shirakura, R. &
Matsuda, H. (1997) In vivo gene transfection with heat shock
protein 70 enhances myocardial tolerance to ischemia-reperfusion
injury in rat. J. Clin. Invest. 99, 1645–1650.
41. Perdrizet, G.A., Heffron, T.G., Buckingham, F.C., Salciunas, P.J.,
Gaber, A.O., Stuart, F.P. & Thistlethwaite, J.R. (1989) Stress
conditioning: a novel approach to organ preservation. Curr. Surg.
46, 23–26.
42. Chatson, G., Perdrizet, G., Anderson, C., Pleau, M., Berman, M. &
Schweizer, R. (1990) Heat shock protects kidneys against warm
ischemic injury. Curr. Surg. 47, 420–423.
43. Shen, R.N., Crabtree, W.N., Wu, B., Young, P., Sandison, G.A.,
Hornback, N.B. & Shidnia, H. (1992) A reliable method for
quantitating chromatin fragments by flow cytometry to predict the
effect of total body irradiation and hyperthermia on mice. Int.
J. Radiat Oncol. Biol. Phys. 24, 139–143.
44. Henderson, C.J., Smith, A.G., Ure, J., Brown, K., Bacon, E.J. &
Wolf, C.R. (1998) Increased skin tumorigenesis in mice lacking pi
class glutathione S-transferases. Proc. Natl Acad. Sci. USA 95,
5275–5280.
45. Hopwood, D., Spiers, E.M., Ross, P.E., Anderson, J.T.,
McCullough, J.B. & Murray, F.E. (1995) Endocytosis of
fluorescent microspheres by human oesophageal epithelial cells:
comparison between normal and inflamed tissue. Gut 37, 598–602.
46. De Jong, U.W., Day, N.E., Mounier-Kuhn, P.L. & Haguenauer, J.P.
(1972) The relationship between the ingestion of hot coffee and
intraoesophageal temperature. Gut 13, 24–30.
47. Cash, P., Argo, E., Ford, L., Lawrie, L. & McKenzie, H. (1999) A
proteomic analysis of erythromycin resistance in Streptococcus
pneumoniae. Electrophoresis 20, 2259–2268.
48. Votyakova, T.V., Wallace, H.M., Dunbar, B. & Wilson, S.B. (1999)
The covalent attachment of polyamines to proteins in plant
mitochondria. Eur J. Biochem. 260, 250–257.
q FEBS 2001 Stress protein induction in epithelial cells (Eur. J. Biochem. 268) 5355
Copyright © 2022 FDOKUMEN




















