
The fossil hominoids of Lufeng, Yunnan Province, the People's Republic of China: A series of...
55
YEARBOOK OF PHYSICAL ANTHROPOLOGY 27~1-55 (1984) The Fossil Hominoids of Lufeng, Yunnan Province, The People’s Republic of China: A Series of Translations DENNIS A. ETLER Departinent ofAnthropology, University of Wisconsin, Milwaukee, Wisconsin 53201 KEY WORDS Lufeng, Miocene hominoids, Ramapithecus, Sivapithecus TRANSLATORS INTRODUCTION The following translations represent the major research papers published in Chinese journals that detail the discovery of hominoid fossils from late Miocene deposits of Lufeng county, Yunnan Province, in the Peoples’s Republic of China. This important body of descriptive and comparative work on the Lufeng hominoids has been previously available to the west only in short summary translations which do not do full justice to the contents of the articles. It is hoped that these translations will illuminate for a western audience the significant research into hominoid phylo- geny now going on in China. The hominoid fossils recovered from the Lufeng basin constitute one of the most extensive in situ collections of hominoid materials ever assembled. As of this writing there are nearly 1,000 specimens representing tens if not hundreds of individuals. This material includes five reasonably complete fossil ape crania, six fragmentary crania, nine mandibles, a dozen or so maxillary fragments, and innumerable man- dibular fragments, tooth rows, and isolated teeth. The description, analysis, and interpretation of this collection of fossil hominoids is essential for understanding the late Miocene radiation of the Hominoidea and emergence of man and the modern apes. The article by Qi (1979) gives a preliminary account of the geology and stratigra- phy of the Shihuiba fossil site at Lufeng. The article by Sun and Wu (1980) is a detailed palynological analysis of the hominoid-bearingdeposits at Shihuiba. The articles by Xu et al. (1978) and Xu and Lu (1979) are systematic descriptions of the holotypes of Ramapithecus lufengensis and Sivapithecus yunnanensis. The article by Wu et al. (1982) gives details of the 1981 excavation and briefly describes some new material recovered during that field season. The article by Lu et al. (1981) contains a preliminary account and comparative analysis of the Lufeng Sivapithecus cranium, while the article by Wu et al. (1983) gives further details on this cranium, suggesting a different interpretation for the Lufeng collection. Translator’s note: The article by Sun and Wu (1980) begins with a section on the geology and stratigraphy of the Shihuiba site which basically recounts that given in Qi (1979).This section (#1) has not been translated as it was felt to be redundant. The article by Xu and Lu (1979) repeats the description of the R. lufengensis mandibular dentition for C-M3 as given in Xu et al. (1978). This is noted in the translated text. The article by Wu et al. (1982) is accompanied by a full translation of the first section, which deals with stratigraphy and which was not worked on by the trans- lator. The first section is included in this collection for reference. INTRODUCTION by Wu Rukang (Woo Ju-Kand March 1984 in BelJing A fossil primate tooth was found by an officer of the local museum on the hill slope Miaoshanpo, 9 km north of the town Lufeng, northwest of Kunming in Yunnan 0 1984 Alan R. Liss, Inc.
Transcript of The fossil hominoids of Lufeng, Yunnan Province, the People's Republic of China: A series of...

YEARBOOK OF PHYSICAL ANTHROPOLOGY 27~1-55 (1984)
The Fossil Hominoids of Lufeng, Yunnan Province, The People’s Republic of China: A Series of Translations
DENNIS A. ETLER Departinent ofAnthropology, University of Wisconsin, Milwaukee, Wisconsin 53201
KEY WORDS Lufeng, Miocene hominoids, Ramapithecus, Sivapithecus
TRANSLATORS INTRODUCTION
The following translations represent the major research papers published in Chinese journals that detail the discovery of hominoid fossils from late Miocene deposits of Lufeng county, Yunnan Province, in the Peoples’s Republic of China. This important body of descriptive and comparative work on the Lufeng hominoids has been previously available to the west only in short summary translations which do not do full justice to the contents of the articles. It is hoped that these translations will illuminate for a western audience the significant research into hominoid phylo- geny now going on in China.
The hominoid fossils recovered from the Lufeng basin constitute one of the most extensive in situ collections of hominoid materials ever assembled. As of this writing there are nearly 1,000 specimens representing tens if not hundreds of individuals. This material includes five reasonably complete fossil ape crania, six fragmentary crania, nine mandibles, a dozen or so maxillary fragments, and innumerable man- dibular fragments, tooth rows, and isolated teeth. The description, analysis, and interpretation of this collection of fossil hominoids is essential for understanding the late Miocene radiation of the Hominoidea and emergence of man and the modern apes.
The article by Qi (1979) gives a preliminary account of the geology and stratigra- phy of the Shihuiba fossil site at Lufeng.
The article by Sun and Wu (1980) is a detailed palynological analysis of the hominoid-bearing deposits at Shihuiba.
The articles by Xu et al. (1978) and Xu and Lu (1979) are systematic descriptions of the holotypes of Ramapithecus lufengensis and Sivapithecus yunnanensis.
The article by Wu et al. (1982) gives details of the 1981 excavation and briefly describes some new material recovered during that field season.
The article by Lu et al. (1981) contains a preliminary account and comparative analysis of the Lufeng Sivapithecus cranium, while the article by Wu et al. (1983) gives further details on this cranium, suggesting a different interpretation for the Lufeng collection.
Translator’s note: The article by Sun and Wu (1980) begins with a section on the geology and stratigraphy of the Shihuiba site which basically recounts that given in Qi (1979). This section (#1) has not been translated as it was felt to be redundant.
The article by Xu and Lu (1979) repeats the description of the R. lufengensis mandibular dentition for C-M3 as given in Xu et al. (1978). This is noted in the translated text.
The article by Wu et al. (1982) is accompanied by a full translation of the first section, which deals with stratigraphy and which was not worked on by the trans- lator. The first section is included in this collection for reference.
INTRODUCTION by Wu Rukang (Woo Ju-Kand March 1984 in BelJing
A fossil primate tooth was found by an officer of the local museum on the hill slope Miaoshanpo, 9 km north of the town Lufeng, northwest of Kunming in Yunnan
0 1984 Alan R. Liss, Inc.

2 YEARBOOK OF PHYSZCAL ANTHROPOLOGY [Vol. 27, 1984
Province. It came from beds of lignite-brown coal which was collected by the local people for fuel. The locality finally became a fossil site that promises to considerably expand our knowledge of ramapithecids, an extinct group of hominoid primates. Since 1975, the site has been studied by a team of paleoanthropologists and verte- brate paleontologists of the Institute of Vertebrate Paleontology and Paleoanthropol- ogy of the Chinese Academy of Sciences, Beijing. Members of the Provincial Museum of Yunnan also took part in the excavation for several seasons.
The Lufeng site is a paleoanthropological gem. Up to the present, five fairly complete or partial skulls from Ramapithecus and Siuapithecus, more than 40 upper and lower jaws, and over 1,000 teeth both in situ or isolated have been found. Other primate specimens include gibbonoids and lorisoids.
The site is probably about eight million years old. It represents an instant of geological time, which holds as much detail as the terrestrial fossil record ever yields. It is a single outcrop of interbedded silt and lignite which contains vertebrate, invertebrate, and plant fossils. In addition to primates, other mammals that have been recovered include a diverse array of Insectivora (Anourosorex sp.), Rodentia (Brachyrhizomys sp.), Lagomorpha (Alilepus sp.), Carnivora (Siuaonyx bathygnathus, Zctitherium gaudryi, Epimachairodus fires), Perissodactyla (Hipparion sp., Chiloth- erium sp., Macrotherium salinum), Artiodactyla (Lophochoerus lufengensis, Dorcath- erium sp., Antilospira sp.), and so on.
Interpretations of the evolutionary relationships between ramapithecids and hom- inids differ among specialists, and this issue continues to be studied. Preliminary studies of different aspects of the site have been published. As most of the papers were published in Chinese with only an English summary or abstract, the language barrier prevents many paleoanthropologists and students of related subjects in the United States and the Western countries from gaining access to them. The English translations of these papers will surely strengthen the communication between scholars of China and the English-speaking world.
As the leader of the Lufeng research group, I am pleased to make this brief introduction.
XU QINGHUA, LU QINGWU, PAN YUERONG, &I GUOQIN, ZHANG XINGYONG, AND ZHENG LIANG (1978) THE MANDIBLE OF RAMAPITHECUS LUFENGENSZS. KEXUE TONGBAO
(SCIENCE BULLETIN) #9:554-556+546.
In November of 1976 during excavations at the Shihuiba colliery site in Lufeng county, Yunnan, members of a joint field team of the Chinese Science Academy IVPP and the Yunnan Provincial Museum discovered a lower jaw of Ramapithecus (see Fig. 1). This is a t present the most complete specimen of Ramapithecus known; moreover, it has morphological features that indicate that it is transitional between ape and man. Because of these facts the discovery and study of these remains has much to contribute to an understanding of the theory of human origins.
This fossil site has also yielded other hominoid material and many fossil mam- mals. Initial research has determined that the lignite beds of Shihuiba are of an early Pliocene age.
Below is a brief review of the preliminary research conclusions regarding the new mandible.
Description of the specimen Family: Hominidae Genus: Ramapithecus Ramapithecus lufengensis sp. nov.
Holotype-IVPP PA580. A relatively complete mandible with full dentition except for the central incisors
Diagnosis This new species of Ramapithecus is distinguished from others of the genus by the
following characteristics: the mandibular dental arcade assumes a regular arcuate
Etler] FOSSIL HOMINOIDS OF LUFENG 3
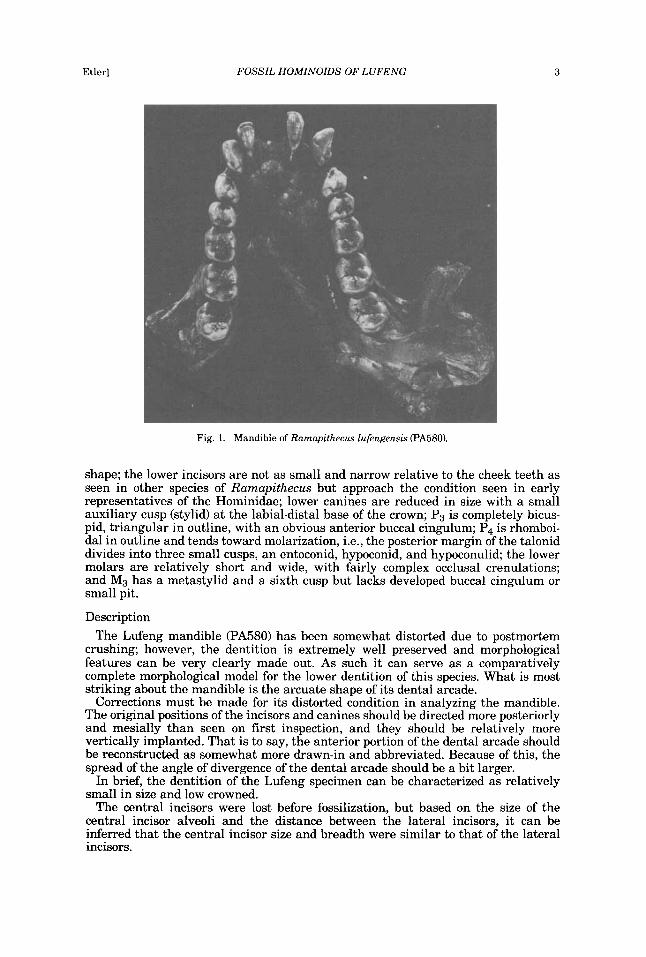
Fig. 1. Mandible of Ramapithecus lufengensis (PA580).
shape; the lower incisors are not as small and narrow relative to the cheek teeth as seen in other species of Ramapithecus but approach the condition seen in early representatives of the Hominidae; lower canines are reduced in size with a small auxiliary cusp (stylid) a t the labial-distal base of the crown; P3 is completely bicus- pid, triangular in outline, with an obvious anterior buccal cingulum; P4 is rhomboi- dal in outline and tends toward molarization, i.e., the posterior margin of the talonid divides into three small cusps, an entoconid, hypoconid, and hypoconulid; the lower molars are relatively short and wide, with fairly complex occlusal crenulations; and M3 has a metastylid and a sixth cusp but lacks developed buccal cingulum or small pit.
Description The Lufeng mandible (PA580) has been somewhat distorted due to postmortem
crushing; however, the dentition is extremely well preserved and morphological features can be very clearly made out. As such it can serve as a comparatively complete morphological model for the lower dentition of this species. What is most striking about the mandible is the arcuate shape of its dental arcade.
Corrections must be made for its distorted condition in analyzing the mandible. The original positions of the incisors and canines should be directed more posteriorly and mesially than seen on first inspection, and they should be relatively more vertically implanted. That is to say, the anterior portion of the dental arcade should be reconstructed as somewhat more drawn-in and abbreviated. Because of this, the spread of the angle of divergence of the dental arcade should be a bit larger.
In brief, the dentition of the Lufeng specimen can be characterized as relatively small in size and low crowned.
The central incisors were lost before fossilization, but based on the size of the central incisor alveoli and the distance between the lateral incisors, it can be inferred that the central incisor size and breadth were similar to that of the lateral incisors.

4 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
The cutting margin of the lateral incisors is level mesially and slopes downward distally. The cutting margin and the mesial surface intersect a t right angles; the distal intersection is obtuse. The lingual surface of the crown has a weak “V”-shaped basal marginal crest. The crown is much wider labiolingually than mesiodistally.
The lower canines are relatively small and low and project only slightly beyond the surface of the tooth row. Again the labiolingual breadth is very much wider than the mesiodistal length.
The upper half of the labial and lingual surfaces of the crown intersect a t the anteroexterior corner to form an anterior marginal ridge. Below this is a triangular mesial surface with a noticeable primary ridge constituting the posterior marginal ridge of the lingual surface. Between this and the posterior marginal ridge of the labial surface, there is a deep vertical groove. At the base of the lingual surface there is a minute “W”-shaped protruding cingulum which runs along to the inferior portion of the posterior marginal ridge of the labial surface forming a triangular stylid (auxiliary cusp). On the distal surface of the stylid there is a very small contact facet. Corresponding to this, on the basal portion of the mesiointerior surface of the P3 crown is a similar-sized contact facet, showing that the lower canine and P3 were originally contiguous and not separated, as it now appears. The canine root is somewhat longer than the lateral incisor root and its inferior portion turns distally outward.
P3 has a very transitional character between ape and man, and can serve as a prototype. It has already clearly differentiated into a bicuspid tooth. This is the first such example seen during the late Miocene-early Pliocene. In this bicuspid tooth, the buccal protoconid is relatively large and somewhat tall and the lingual meta- conid is relatively small and low; between the two cusps is an obvious vertical groove. (In the dryopithecines and Siuapithecus, it is often possible to see a deeply inclined groove between the buccal posterior marginal ridge and the primary lingual ridge that extends from the posterior forea to the apex of the protoconid.) The talonid broadens out and its distal surface becomes flat. The buccal surface is abbreviated and the mesial lingual surface is slanted and slightly raised, making the outline of the crown triangular. The basal cingulum of the mesiolingual surface turns ante- riorly as an obvious anterior buccal cingulum; posteriorly, it terminates at the metaconid and does not continue to the posterior margin. The root divides into anterior and posterior branches.
P4 crown is rhomboidal in outline. The protoconid and metaconid are approxi- mately equal in size. There is a longitudinal groove separating the two cusps. A transverse ridge develops opposite the apices of the two cusps and converges with the longitudinal groove. There is a slender transverse anterior fovea in front of this. Posteriorly there is a large, broad talonid basin. The sloping posterior surfaces of the two cusps each have an obvious inclined groove and several delicate crenula- tions. The posterior margin is rough and blunted and has a tendency to divide into three very small cusps, the entoconid, hypoconid, and hypoconulid. The dental root divides into anterior and posterior branches.
The lower molar crowns are relatively short and wide with low-lying cusps. The cusps on the two lateral sides are fairly wide apart. The talonid basins are relatively large and broad. The five main cusps are arranged in a Y-5 pattern as a general feature.
M1 has a relatively small crown; the entoconid projects a slight bit posteroin- teriorly. A trace of cingulum still exists on the buccal surface between the protoconid and hypoconid. Left M1 hypoconulid is damaged by a relatively large enamel win- dow. The root is divided into anterior and posterior branches.
Mz is a bit larger and wider than MI, but does not show the great disparity in size (heterodonty) seen in the dryopithecines or Siuapithecus. The buccal cingulum is completely lost and the posterior fovea is fairly well-marked.
M3 is a little smaller than Ma. It differs from the other two lower molars in having two auxiliary cusps (stylids). The posterior part of the crown narrows, and there are fairly complex crenulations on the occlusal surface. The first auxilary cusp is posi-

Etler] FOSSIL HOMINOIDS OF LUFENG 5
tioned posterointeriorly to the metaconid. Since it differentiated from the metaconid, it is recognized as metastylid. The second auxiliary cusp is positioned between the entoconid and the hypoconulid. It occupies the position of the posterior fcvea. Since it is hard to tell from which cusp it arose, it is called a sixth cusp. M3 does not have a buccal cingulum.
Since the mandibular corpus has been deformed and damaged, it is not described here.
Comparison and discussion From the above morphological characteristics it can be seen that the Lufeng
specimen is unlike either the dryopithecine apes or Sivapithecus, and is more like Ramapithecus. The dryopithecine apes and Sivapithecus have “U”-shaped dental arcades, relatively large tooth size, high crowns, tall, large, and sharply pointed canines, single-cusped, sectorial P3, and P4 set obliquely in the tooth row with an elongated talonid. MI is much smaller than Mz, and M3 and is relatively high cusped. Given these characteristics it can be seen that the Lufeng specimen cannot belong to either Dryopithecus or Siuapithecus.
Among the Ramapithecines, the late Miocene Kenyan right mandibular specimen KNM-FT45 assigned by Andrews (1971) to R. wickeri has an arcuate dental arcade. Its shape and that of the Lufeng specimen are, however, not the same. The Kenyan specimen shows an irregular arc which curves inward at Pq. This shape can also be seen among the more primitive Proconsul; it is, therefore, a primitive character. In like manner, the left mandible discovered in early Pliocene deposits of Pakistan in 1969 by D. Pilbeam and assigned to R. punjabicus also evidences an irregular arcuate shape of the dental arcade. Among all hominids the dental arcade is regu- larly arcuate. It can be seen that the shape of the Lufeng dental arcade closely approximates this progressive character.
The new mandibular material of Ramapithecus from the early Pliocene of Paki- stan displays incisors that are especially small, and do not match up with the cheek teeth. The lower incisors of the Lufeng specimen, however, are moderate in size, similar to early hominids such as H. habilis and H. erectus, showing that the Lufeng specimen is indeed close to Homo.
From the lower canine alveolus preserved in the Pakistani and Kenyan mandibles, it can be inferred that the lower canines were relatively small. Their roots also seem to be a bit shorter than would be the case in the Lufeng material.
The P3 of the Kenyan left mandible (KNM-FT45) already shows incipient bicuspi- dization. There is, however, no obvious vertical groove between the two cusps. The metaconid is very small; the lingual cingulum runs around the base of the metaconid and continues onto the posterior margin. The crown is semisectorial. These charac- teristics all express a rather primitive condition unlike the Lufeng specimen, which approaches the human condition. The Kenyan specimen P4 talonid is angled toward the posterointerior corner. This is, likewise, a primitive condition not seen in the Lufeng specimen.
The dental specimen of R. wickeri from Turkey shows a P4 talonid basin that is especially elongate, causing the coronal outline to be all the more long and narrow. The lower molars all clearly evidence buccal cingulum. This type of primitive condition is not seen in the Lufeng specimen.
The three mandibular fragments discovered in India (DGSI-118/119 and YPM138141, and the dental series discovered in Kaiyuan, China (PA75:1-5), show lower molars that are small sized with low crowns, cusps that hug the dental margins, and relatively broad talonid basins. The M3’s in particular all have a metastylid and sixth cusp. These characteristics are very similar to the Lufeng specimen. In contrast to the Lufeng specimen, the molars all have buccal cingulum and crowns are somewhat low, small, long, and narrow (cf. YPM13814), clearly displaying a more primitive condition.
Two mandibular fragments from lower Pliocene lignite deposits of Hungary (Rud- 1 and Rud-17) have been referred, by some workers, to Ramapithecus. From original

6 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
photographs it is apparent that the dental arcade is not arcuate; the canines are large, tall, and sharply pointed. P3 is unicusped, sectorial, and larger in size than Pq. These are all apelike characteristics, fundamentally different from the Lufeng specimen.
In summary the following points are offered: (1) The dental size and morphology of the Lufeng PA580 mandibles clearly show it
to belong to Ramapithecus, but it has certain "specific characteristics" that distin- guish it from other members of the genus. Because of this we deemed it proper to classify it as R. lufengensis sp. nov.
(2) R. lufengensis is morphologically closer to Homo in character than any previ- ously known specimens of Ramapithecus. This is especially evident in the arcuate shape of the dental arcade, the moderate size of the lower incisors, the completely bicuspid nature of P3, the short, broad crowns of the lower molars, and the lack of buccal cingulum. These are all traits reflecting a transition from ape to man. From materials presently available, it can be argued that R. lufengensis is a late Miocene/ early Pliocene member of the hominids which tends to most closely approximate Homo.
(3) The discovery of R. lufengensis goes to further demonstrate that southwest China is a most important region in human origins. It, moreover, attests to the vast potential that the Neogene lignite beds spread throughout Yunnan have in the search for hominid ancestors.
$1 GUOQIN (1979) PLIOCENE MAMMALIAN FAUNA O F LUFENG, YUNNAN. VERTEBRATA PALASIATICA. 1711): 14-22.
The Lufeng fossil site in China's Yunnan province has recently gained interna- tional attention for its hominoid-bearing deposits. In the past, the Lufeng locality was known for its prolific remains of Mesozoic dinosaurs and mammal-like reptiles. Cenozoic deposits yielded a near complete blank as far as vertebrate fossils were concerned.
In April 1975, numerous fossil teeth were unearthed at the Lufeng Shihuiba colliery site. These finds were reported to the Yunnan Provincial Museum by Wang Zhengju of the Lufeng Cultural Relics Bureau. Initial excavations of the coal beds recovered fossil hominoid teeth of the Tertiary period. In the winter of 1975, a joint work team from the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) Academia Sinica, and the Yunnan Provincial Museum conducted further excavations which continued into the spring and winter field seasons of 1976. Hominoid and mammalian remains recovered from the Lufeng lignite beds have been abundant and representative, giving a relatively complete picture of the Pli- ocene mammalian fauna of south China.
Geology and topography of Shihuiha and environs The Lufeng Shihuiba colliery site (IVPP 75033; 102"4'E, 25'3") is situated on
the southern slope of Miaoshanpo hill, about 9 km north of Lufeng city on the left bank of the Xihe river. In general, the area has been elevated and subject to erosion for a long period of time. Exposed bedrock near the site is mainly of the Presinian Kunyan group and the Lufeng series of the lower Jurassic.
The Kunyan group is located east of the Xihe river in the area surrounding locality 75033. It consists of greyish white, greyish blue, and in part deep-red, small-grained crystalline dolomitic limestone, intercalated with purplish mica-slate-laden second- ary belts of quartz. This stratum is identified as the upper member of the Luzhijiang formation. It is oriented to the southeast at an inclination of 15"-160" with an angle of inclination of 40"-45". The surface has developed a thick, red, aeolian crust, topographically expressed as well-rounded, low-lying hills and mountains to the east of the Xihe river, north of Lufeng.
Red Mesozoic deposits, consisting of dark purple sandy shale in part intercalated with a small amount of greenish-grey mudstone, are located primarily west of the Xihe river. They are identified as the lower Lufeng series of the early Jurassic, expressed topographically as mid- to high-ranging mountains. Looking southwest

Etler] FOSSIL HOMINOIDS OF LUFENG I
from the railroad bridge over the Xihe river along the railway, the lower Lufeng series can be seen to intermittently cover the exposed Luzhijiang formation.
Cenozoic deposits are primarily spread along the foothills of the hills and moun- tains formed by Kunyan group strata, generally as fluviaUlacustrine deposits of gravel, sand, sandy clay, clay, and lignite. Various Cenozoic sections deposited at different locations on Miaoshanpo hill that have been excavated are described below. Strata are recorded from top to bottom in turn (see Fig. 2, Diagram 1).
Section A Located about 100 m west of Shihuiba, trend 45 O NE, exposed thickness about 18 m. (1) Purplish-red weathered crust with large and small gravels and sand, about 1.5
m thick. (2) Yellowish, brownish-yellow calciferous sandy clay in horizontal beds with thick
overlying gravel deposits, about 4 m thick, clearly demarcated from lower deposits by an erosion surface.
(3) Greyish-yellow sandy clay, upper part containing a belt of brown carbonaceous clay about 2.5 m thick. (4) Greyish clay containing gravels about 3 m thick. (5) Purplish-red sandy clay mixed with gravel, gravel becoming coarser toward
bottom, about 7 m thick.
Section B Located about 50 m west of section A, trend 45" NE, exposed thickness about 15
m. These layers are basically similar to those of section A but are finer grained. (1) Purplish-red, reddish-yellow weathered crust containing assorted gravels about
1 m thick. (2) Brownish-yellow sand and gravel; gravel composed of quartz, sandstone, and
slate; average diameter 5 cm, maximum diameter 18 cm; moderately rounded about 1.5 m thick, over an erosion surface.
(3) Brownish-yellow sandy clay intercalated with brown carbonaceous clay, about 3.2 m thick.
(4) Greyish-yellow clay with calcareous mud balls and sand and gravels composed of bedrock, about 6 m thick.
Fig. 2. Site of the Lufeng hominoids

8 YEARBOOK OF PHYSICAL ANTHROPOLOGY vol. 27, 1984
(5) Purplish-red sandy clay containing a large amount of sand and gravel. Com- posed mostly of sandstone and slate. Maximum diameter about 20 cm, turning a deep purple when exposed to weathering; buried deposits are brown to brownish- grey, about 3 m thick.
Section C
exposed thickness about 5 m.
aceous gravels, about 1 m thick.
Located 150 m west of section B, and 50 m south of section D, trend NE 15",
(1) Brownish-yellow sandy clay, containing calcium and a small amount of carbon-
(2) Grey, greyish-yellow clay containing some sand, about 1 m thick. (3) Purplish-red sandy clay containing sand but comparatively thinner than in
section A or B, about 3 m thick.
Section D (Excavated worksite 75033) about 150 m west of section B, adjacent to the road
leading from Shihuiba to Miaoshanpo, oriented in a northlsouth direction, exposed thickness about 6 m.
(1) Greyish-yellow, yellowish-brown sand, sandy clay and clay, often with sand lenses, containing coal slack and well-rounded, worn quartzite and sandstone peb- bles. Fossils show abrasive scars from alluvial transport. There are a small number of mammalian fossils such as Hipparion, elephant, and rhinoceros. The color, lithog- raphy, and folding of the lower level of the stratum clearly show traces of undulating wave action, disconformably situated over the lower strata about 0.7-2.0 m thick. Palynological sequence (P.S.)I-2,1-3.
(2) Greyish-yellow, grey sandy clays, and lignite. The lignite bed is thin, 10-30 cm. There are three to four coal seams intercalated with sand lenses, containing abun- dant mammalian, soft-shelled turtle and other fossils. Fossil hominoids are very rare, 0.6-2.5 m thick (P.S.)II-2:7.
(3) Upper section of hominoid-fossil-bearing stratum. Lignite and greyish-white sands, characterized by well-formed chunks of lignite, 40-80 cm thick. Mammalian and hominoid fossils are very abundant. The Ramapithecus skull and important mandibular and maxillary materials are found throughout this section. This is the most important location for fossil hominoids in the Shihuiba formation, 1.2-1.5 m thick (P.S.)1112:6. (4) Lower section of hominoid-fossil-bearing stratum. Lignite and pure, thick grey-
ish-white, fine-coarse sand, evenly distributed, with many fossil hominoids, fish, soft-shelled turtle and molluscs common, 2-2.2 m thick (P.S.)IV-1.
(5) Lignite and greyish-black, greyish-white sandy clays. Upper section has lignite seam 20-30 cm thick. Lower section consists of greyish-black sandy clays and contains rhinoceros, elephants, and other large mammalian types. Hominoid fossil remains very scarce. 0.7 m thick (P.S.)V-1.
The layers of section D are deposited continuously and incline to SW 220°, angle of inclination 10"-12". All layers of section D correspond to layers 3 and 4 of section B.
Section E Situated about 20 m south of section D, trend 15" NE, exposed thickness 3 m. (1) Varigated sands and gravel intercalated with clay. Gravel is composed of well-
rounded sandstone, limestone, quartz, slate, etc., diameters of gravels gradually increase toward top of layer averaging about 3 cm, maximum diameter 15 cm. There are three thin strata of clay contained in level 1. Clay is brownish-red to brownish- yellow, increasingly washed out bottom to top, about 1 m thick.
(2) Purplish-red to yellowish-orange weathered crust, earth-formed accumulations containing angular gravels, about 2 m thick.
(3) Greyish-yellow , brownish-yellow sand and sandy clay containing irregular brown belts of carbonaceous clay similar to level 1 of section D, about 0.1 m thick.

Dia
gram
1. G
eolo
gic
cros
s-se
ctio
n, L
ufen
g Sh
ihui
ba fo
ssil
ape
site
' -
MQ
~
'Edi
tor'
s N
ote:
Oth
er l
ine
draw
ings
pre
sent
ed i
n th
is a
rtic
le h
ave nut
been
cil
ed in
the
text
. The
y ha
ve b
een
deri
ved
from
the
orig
inal
Chi
nese
art
icle
s an
d ar
c pr
esen
ted
here
for
the
read
er's
rmlig
hknm
ent.

10 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
The above sections and their correspondences are shown in Diagram 1. Section A and section B basically summarize the Cenozoic stratigraphy of the Shihuiba area as follows: the ancient bedrock to the erosion surface (i.e., sections A and B midstrata 3-5) represent Pliocene deposits; above the erosion surface there are possible post- Pliocene (?&) accumulations. The strata of section D correspond to strata 3 and 4 of sections A and B and strata 1 and 2 of section C. Sections C and D lack stratum 1 of sections A and B (i.e., weathered crust). The sand and gravel deposits of sections A and B, from their erosion surfaces to their respective weathered crusts, and the sand and gravel strata of section E overlying the weathered crust, were apparently laid down at different times. The sand and gravel deposits underlying the weathered crust of sections A and B are possibly early remnant terrace accumulations laid down by the Xihe river. The sand and gravel deposits overlying the weathered crust of section E, which are intercalated with clay, are most likely later terrace accumu- lations of the Xihe river. They are today widely distributed throughout the surround- ing area and can be seen from site 75033 west toward the banks of the Xihe river.
In summary, it can be seen that during the Pliocene, Shihuiba was near a small, low-lying piedmont depression centered close to present section D. In this depression there were initial deposits of purplish-red sandy clays intercalated with sand and gravel. Subsequent depositions were of grey and greyish-yellow sandy clays and clays. Toward the center of the depression were marshlands, later forming the present-day lignite beds.
The mammalian fauna of Lufeng and its biochronologic placement Of the above geologic sections, only section D has yielded mammalian fossils and
these have been primarily unearthed from strata 2-4. Initial analysis of the mam- malian fauna is as follows:
Primates Ramapithecus lufengensis sp. nov. Sivapithecus yunnanensis sp. nov. Lorisiformes gen. Hylobatinae gen. Macaca sp.
Rodentia Sciuridae Hystrix sp. Scaptochirus sp. Brachyrhizomys Castoridae
Lagomorpha Alilepus sp. nov. spp. (1,2,3)
Carnivora Zctitherium gaudryi Viverrids spp. (1,2,3) Mustelinae Sivaonyx bathygnathus Epimachairodus sp. nov. Eomellivora sp.
Proboscidea Serridentinus sp. Gomphotheridae gen. and sp. indet.
Perissodactyla Hipparion cf: nagriensis Chilotherium sp. nov. Aceratherium sp. nov. Macrotherium salinum Tapirus sp.
Artiodactyla Potamochoerus cf: salinus Hyotherium cf: palaeochoerus Dicorychoerus sp. Suidae gen. et sp. indet. Doratherium minus Palaeomeryx sp. Cervidae Dorcabune Nuntiacus sp. nov. Metacervulus cf: simplex Antilospira sp. Moschus cf: primaeus
A preliminary count of the above faunal specimens gives a total of 40 types, including five species of Primates, six species of Carnivora, five species of Rodentia, one species of Lagomorpha, two species of Proboscidea, five species of Perissodactyla, and 13 species of Artiodactyla. The fossils are not well preserved and consist mainly of isolated teeth with a few crania, mandibulae, and maxillae. Of these, higher primates and large-bodied ungulates are most common.

Etler] FOSSIL HOMINOIDS OF LUFENG 11
According to traditional geological thinking, the Old World Pliocene began with the arrival of Hipparion. It is generally recognized in Europe that Hipparion first appears during the Vallesian or the lower part of the middle Sarmatian. In Asia there has been a long history of research into the stratigraphy and mammalian faunas of the Neogene. The most detailed studies have been done in the Siwalik Mountain region of India and Pakistan. In the past, various investigators have recognized the earliest appearance of Hipparion to be the Chinji formation. In 1969, S.T. Hussian restudied the Hipparion of the Siwaliks and placed its earliest appear- ance in the Nagri formation, naming the species Hipparion nagriensis. Recently, D. Pilbeam et al. (1977a,b), in their articles concerning new Tertiary materials from the Siwaliks, stated that the earliest appearance of Hipparion is in the lower Nagri formation. At Lufeng, fossil remains of Hipparion are scarce. From the form and metrics of their teeth, they appear to be very similar to H. nagriensis (Hussian). Other members of the Lufeng fauna, i.e., Ictitherium gaudyi, Chilotherium, Siuaonyx bathygnathus, Hyotherium cf. palaeochoerus, and Epimachairodus, can also be seen in the faunal compilations for the Nagri as reported by E.H. Colbert (1935) and Pilbeam et al. (1977a,b), and in fauna from contemporary north China and Europe.
Among the Lufeng fauna there are also representative types from the Eurasian Miocene, such as Macrotherium, Aceratherium, and Potamochoerus. As can be seen, the Lufeng fauna show typical Hipparion and other Pliocene faunal representatives known from the Siwalik Nagri formation and Eurasia, and other typically Miocene species; accordingly the geologic age can be initially regarded as early Pliocene. According to Pilbeam et al. (1977a,b), the most important section for Nagri formation faunal collection is close to its upper member. Pilbeam et al. recognize the Nagri formation faunal age to be possibly correlated with the late Vallesian or early Turolian of Europe. Considering the similarity between the Nagri and Lufeng fauna, and the recently discovered Hipparion fauna from Tibet, which are possibl older than that from Lufeng, it can be suggested that the faunal age of the h ‘ e n g material is late Vallesian or early Turolian, i.e., late early Pliocene or early mid- Pliocene.
There has been over a half century of research into the Pliocene stratigraphy and mammalian fauna of China. Before liberation, however, most fossiliferous localities investigated were in the provinces of Shanxi, Shaanxi, Gansu, Henan, and Shan- dong. Besides the determinations made a t Baode (N;)’ and Jingle (Ng) in northwest Shanxi and those made at the Yushe formations (IN;, IINg, IIIN;) in southeast Shanxi, all other Hipparion deposits have been subsumed under the nomenclature of “Liandi period Hipparion red clay.” However, insofar as much of the fossil material from Baode and Yushe was purchased at Chinese drugstores or from the populace at large, there still exist many questions as to their exact geologic prove- nance. Since liberation, wide-ranging large-scale investigations have been conducted at Lantian, Shaanxi province. Based on faunal and geomorphic correlations estab- lished among Pliocene deposits along a stretch of the Bahe river, the Bahe formation has been established as (NhP2) and the Lantian formation as (NZ), largely similar to conditions seen within the Fenhe gorge in south central Shanxi.
At present only a small number of the Lufeng fauna which occupy broad ecological zones such as Ictitherium, Epimachairodus, Mustelidae, and Antilospira are found in the Hipparion fauna of north China. It is, therefore, still relatively difficult to establish concrete comparisons and correspondences between the age of the northern Hipparion fauna and that of Lufeng.
The composition of the Lufeng fauna has its own distinct color. It contains many primates, Viverridae, Doratherium, etc., of a southern type; yet, it also contains species frequently seen in more northern Hipparion fauna. Lufeng’s geographical position, however, aligns it with southern regions such as the Siwaliks of India and Pakistan. It has also been determined that certain types that should be expected in
‘Translator’s note: In Chinese nomenclature N refers to Neogene; subscript 1 refers to Miocene, subscript 2 to Pliocene. Superscripts 1,2, 3 refer to early, middle and late divisions of an epoch.

12 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
such fatma, i.e., Giraidae, are absent at Lufeng either because environmental conditions were inappropriate or because the excavations have not yet been thor- ough enough.
From the above, it can be seen that the Lufeng faunal composition is basically analogous to that of the Nagri formation, although there are a number of differences. As far as the northern Hipparion fauna are concerned, there are some similarities, but the differences are comparatively greater.
As with other mammalian fauna that contain Ramapithecus, the Lufeng fauna has various kinds of primates, Sciuridae, otter, forest rhinos, viverrids, as well as primitive munjak and deer which are mostly forest-dwelling species. Nevertheless, there are also a few genera, which are numerically well represented, of large herbivorous forms such as Antilospira (a caprine). From the nature of the fossils recovered from the lignite beds and palynological analysis still in progress, the area around Lufeng, Shihuiba, at the time in question was characterized by a warm, humid climate and a natural environment of marshland, spread along a piedmont basin in the vicinity of forest and grassland.
SUN XIANGJUN AND WU YUSHU (1980) PALEOENVIRONMENT DURING THE TIME OF RA MAPITHECUS LUFENGENSIS. VERTEBRATA PALASIATICA 18(3):247-255.
[Translater’s note: Translation begins with second section of the article.]
A palynological analysis of section D distinguishes three distinct developmental General outline of pollen-spore assemblages
stages of pollen-spore assemblages. Stage I. Fern lizard-tail floral stage
Pteridophyte spores and Saururaceae pollens predominate, referrable to the lowest part of the fifth stratum of the section. Fern (pteridophyte) spores constitute 43% of this assemblage. Pollen from herbaceous plants and small shrubs represents 35%, while woody plants account for nearly 22%. Within the spore sample there is a preponderance of genera and species of Polypodiacae, i.e., Polypodium argtum, f? niponicum, Neolepisorus ovatus, Lepisorus oligolepidus, and Drynaria. Also present are the Pteridaceae (brake ferns); Pteris latealata, f? brevifolia, l? baksaensis, Dav- alliaceae; Davallia and a large number of Psilate monulites. Among the herbaceous plants, Saururus (lizard-tail) pollens, constituting 20% of palynomorphs, have the highest percentage. Umbelliferae (carrot family) are also present. Woody plants and trees are well represented, especially chinquapin (Castanopsis) 7%, alder (Alnus nepalensis) 8%, along with the palmlike angiosperm Cycas 3%, oak (Quercus) 2%, and a small amount of Sumac (Rhus), olive (Oleacea), and birch (Betula) pollens, etc. Stage 11. Bayberry/alder floral stage
Bayberry (Myrica) and alder (A lnus nepalensis) pollens predominate, referrable to strata 4-2 of section D. There is a preponderance of tree pollens (46-71%), followed by spores (7-30%) and pollen from herbaceous plants and small shrubs (8-28%). Among trees bayberry and alder, and among vines, the southern snake vine (MoncF celastrurn) are very prevalent. There are three kinds of bayberry-Myrica esculenta, M. nana, and M. rubra-present. Among the alders Alnus nepalensis is most impor- tant. In addition chinquapin (Castanopsis), Tarenna, oak (Quercus), hornbean (Car- pinus), hazelnut (Corylus and Ostrya), black dye tree (Platycarya), yellow-basket “willow” (Engelhardtia), hickory (Carya), walnut (Juglans), wingnut (Pterocarya), sweetleaf (Symplocos), elm (Ulmus), Zelkova, chinaberry (Melia), Murraya ponicu- lata, sourgum (Nyssa), Scheflera, holly (Zlex), and Culamus are present. The palm- like Cycas, seen in stage I, is not evident. Spores, both in quantity and form, are not as common as in Stage I; most important are Polypodiaceae and Davalliaceae such as Goniophlebium subauriculatum and Davallia sinensis, etc. Among herbaceous plants the most significant pollens include Umbelliferae (1-14%) and among others, Fatoua, knotweed (Polygonum), crowroot (Ranunculacea), lizard-tail (Sausurea), and a small amount of freshwater algae (Spirogyra).

Etler] FOSSIL HOMINOIDS OF LUFENG 13
Stage III. Alder/pine floral stage Alnus and Pinus prevail. Two samples are referrable to the upper part of stratum
1 of section D. This stage is quite distinct from the previous two. In the previous two assemblages samples showed only a moderate quantity of disaccate pine pollens. This assemblage, in contrast, contains up to 9-13% with Pinus being most common. There are also small amounts of Chinese fir (Keteleeria evelyniana), cedar (Cedus), Chinese hemlock (Tsuga), and dragon spruce (Picea). Alders also increase dramati- cally, approaching 2443%. At the same time Myrica (bayberry) declines abruptly and is seldom seen. The chinquapin (Castanopsis) likewise falls off. Besides these differences, the pollen-spore types of this assemblage and that of stage 11 are largely the same.
Environmental conditions at the time of Ramapithecus The above three pollen-spore assemblages reflect the palaeobotanical and palaeo-
climatic conditions extant at and before the time of Ramapithecus in the Lufeng area over 10 million years B.P. and the division of this period into three distinct stages.
The first stage does not yet show evidence of fossil hominoids. This could perhaps represent a stage before the first appearance of Ramapithecus in the area. This stage is characterized by the predominance of pteridophyte spore and Saururaceae pollens. Saururus is a perennial herb widespread thorughout Japan and China, south to the Philippines. Yunnan has one species (Saururus chinensis) distributed over the Dianmian (YunnarBurmese) border region growing in dense forest, prefer- ring a hot, humid climate. Among the pteridophytes (ferns) Polypodium argtum and Pteris latealata are now found in primeval forests and thickets of southeast and southwest Yunnan. Pteris baksuensis is found on Hainan Island. Other species are today evenly distributed throughout central Yunnan.
Although tree pollens are not well represented in this assemblage there are an abundance of ferns. Ferns are forest-floor dwellers and today a large number of species such as Polypodium argtum, Pteris sp., and Neolepisorus sp. are found only in such an environment. Others, i.e., Davallia and Drynaria, for the most part grow on tree trunks or rocks and are associated with a forest ecosystem. Saururus is also a ground-growing forest species. It can be postulated, therefore, that the assemblage indicates a time when a forest environment prevailed.
At the time of deposition, a hot, humid, tropical or south Asian tropical climate is indicated. It was warmer and moister than in the present-day region around Lufeng, perhaps similar to the climate now seen in southern Yunnan.
The second assemblage represents a time when Ramapithecus and associated fauna were present at Lufeng. The pollen-spore assemblage reflects that the area near the depositional section was at the forest margin in a mixed woodlands zone composed of evergreen and deciduous trees and shrubs. Among the evergreens the Myricaceae (bayberry) Myrica esculenta, M. nana, and M. rubra were most signifi- cant. Among the deciduous trees, the alder (Alnus nepalensis) predominated. A diverse group of other deciduous families were also evident, i.e., Juglandaceae (walnuts and their allies), Betulaceae (hazels, etc.), Aquifollaceae (holly), and Sym- plocaceae (sweetleaf). Scattered within the forests and along the water margins were meadowlands as indicated by the presence of Umbelliferae, Polygonaceae, Moraceae (mulberry), and Saururacea among the herbs and Polypodiaceae among the ferns within the microflora.
The sampling of algae demonstrates that the area had pools and lakes, along which grew a great quantity of A l m s nepalensis.
From an analysis of the pollen-spore assemblage it can be seen that not far from the sampling area grew a montaine evergreen, Castanopsis, and oak forest. The forest floor consisted of low-lying shrubs, such as Myriaceae, Tarenna, etc. Climbing the tree trunks were vines, such as Monocelastrum, and ferns such as Davallia grew in the vicinity.

14 YEARBOOK OF PHYSICAL ANTHROPOLOGY 1Vol. 27, 1984
Based on present-day conditions, it can be inferred that the climate during the time of Ramapithecus was quite warm. Compared to the previous period it was somewhat drier. Today Myrica, Engelhardtia, etc., grow on dry hillsides. Other than Myrica and Davallia the flora represented is currently seen most often in central Yunnan, so it can be estimated that the temperature a t the time of deposition was somewhat higher than in present-day Lufeng, similar to conditions encountered now in central and southern Yunnan.
The third assemblage is from the uppermost part of the section from which, until now, no hominoid fossils have been recovered. This strata could perhaps represent a period when hominoids were no longer present in the area. The palynological character of this stage indicates the spread of coniferous forest. The mountain slopes, however, still had a Castanopsisloak forest. The bayberry (Myrica) forest on the gently rolling slopes characteristic of the former hominoid occupation is gone. Only the Alders remain as the predominant members of the mixed forest.
The appearance of pine pollens and the decline in bayberry and Castanopsis pollens indicate a fall in temperature.
Comparison with the environment of Ramapithecus i n the Siwaliks Ramapithecus fossils were first discovered in the Siwalik mountains of India and
Pakistan. The exposed thickness of the sands, gravels, and silts of the Siwalik group approaches 2,000 feet. Since there are abundant vertebrate fossils, the Siwaliks are very famous. Based on the vertebrate fossils present, the Siwalik group is divided into three periods: lower, middle, and upper, the time ranging from the mid-Miocene to the lower Pleistocene. Rarnapithecus is found in the middle Siwaliks.
The paleobotany of the Siwaliks has been extensively investigated and reported (Ghosh and Ghosh, 1958; Rawat, 1964; Verma, 1968; Patnak, 1969; Mathur, 1974). The presence of Dipterocarpoxylon, Anisopteroxylon, and Borraginaceae reflect a tropical environment at the time of deposition.
Bachejce (1968), Ghosh (19771, Lukos (1969), Nandi and Bandyopadhyay (1970), and Nandi (1972) have done palynological studies of the deposits. Ghosh (1977) based on materials from six sectional outcroppings, distinguishes four biostratigraphic zones with Ramapithecus assigned to the third zone which contains the most abun- dant pollen-spore fossils. Among these angiosperm pollens are highest in quantity, i.e., Longapertites, Betulaepollenites, Caryapollenites, Myrtaceidites, Symplocospol- lenites, Monoporopollenites, and Alnipollenites. Gymnosperm pollens are relatively abundant, but Pteridophyte spores are rather scarce. Based on the character of the pollen-spore assemblage, Ghosh (1977) discusses the paleoclimatic conditions at the time the Siwalik deposits were laid down. The evolutionary trend from the lower to mid-Siwalik was from a humid, tropical or subtropical climate to a comparatively drier and cooler one. In the upper Siwaliks, the diversity of pollen-spores declines appreciably. Ghosh (1977) speculates that this could represent the fourth Himalayan Uplift.
Lukos (1969) also reports on the pollen-spore assemblage from the middle Siwaliks composed of Polypodiaceae, Pteris, Schizea, Lygodium, Myricaceae, Juglandaceae, Magnoliaceae, Mimosaceae, Moraceae, Pinus, Tsuga, and Podocarpus. In these de- posits there are also abundant animal fossils, representing the tail end of the Miocene.
In summary, the paleobotanical material shows that the representative flora of the Siwaliks during the time of Ramapithecus included Dipterocarpoxylon, Mora- ceae, Palmae, Myrtaceae, Magnoliaceae, and Myrica, Juglans, Carya, etc. The moun- tains had a Pinus, Tsuga, Podocarpus, etc., coniferous forest.
The Lufeng Ramapithecus strata and the Siwalik group have many similarities. They contain a diverse spectrum of floral types, such as Polypodiaceae, Pteris, Betulaceae, Myriaceae, and Juglandaceae. This reflects the fact that the natural environments of Lufeng and the Siwaliks a t the time of Ramapithecus were compa- rable. Of interest is the evolution of the climate as reflected in the palynological

Etler] FOSSIL HOMINOIDS OF LUFENG 15
record. Both areas in general are alike, going from a very hot and humid climate to a warm and dryer one, and then to a cool climate.
According to the initial comparison of the palynological data, the hominoid fossil bearing fourth and higher strata of section D could be upper Miocene to lower Pliocene in age. The fifth and lower strata could be of the mid-Miocene.
XU QINGHUA AND LU QINGWU (1979) THE MANDIBLES OF RAMAPITHECUS AND SIVAPZTHECUS FROM LUFENG, YUNNAN. VERTEBRATA PALASIATICA 17(1);1-13.
In May 1975, Wang Zhengiu of the Lufeng County Cultural Bureau discovered the lower molar of a Tertiary ape and other mammalian fossils while conducting a survey of the Shihuiba colliery site. In the winter of that year the IVPP (Academia Sinica) and the Yunnan Provincial Museum organized a joint field team to investi- gate and excavate the fossiliferous locality, and then large-scale excavations were continued in the spring and winter of 1976. Two comparatively complete fossil hominoid mandibles and more than 100 associated fossil teeth were recovered. The two fossil jaws are morphologically distinct and represent two types, one seemingly transitional between ape and man, and the other close in form to the fossil orangutan.
Ramapithecus lufengensis (Xu et al., 1978) The holotype of this species is a comparatively complete mandible (PA850). This
article amplifies on the description, comparison, and discussion given by Xu et al. (1978).
Description The most important distinguishing characteristic of the mandible is its regularly
arcuate shape. The specimen was crushed and deformed during deposition and fossilization. The jaw is broken anteriorly between the right lateral incisor and canine, the crack passing through the symphysial region, producing a fractural deformity everting the right lateral tooth row and splaying the canine and incisors downward and out.
Based on the position of the lingual alveolar margins of the canines and incisors, and the observation that incisors, canines and P3’s have corresponding contact facets, it can be assumed that these teeth were arranged contiguously along the tooth row. During the reconstruction of the dental arcade, the anterior portion of the mandible was consequently foreshortened and directed a bit medially. The incisors and canines are implanted more vertically in the alveolus (see Fig. 1).
11 was lost before fossilization, but its size can be inferred to have been about the same as 12.12 is much smaller than the equivalent in the Lufeng Siuapithecus and approximates that seen in early representatives of Homo. It is much wider labiolin- gually than mesiodistally. The cutting edge is level mesially and slopes downward distally, intersecting the mesial surface at a right angle and the distal surface at an obtuse angle. The lingual surface has weak mesial and distal marginal ridges which converge at the base of the crown forming a “V”; there are, Lowever, no lingual tubercles [translator’s note: see Xu et al. 1978 for description of C, Ps, Pq, M1-31.
Comparison and discussion It can clearly be seen that the Lufeng mandible (PA580) is morphologically and
dimensionally distinct from Dryopithecus or Sivapithecus and more similar to Ramapithecus.
The genus Ramapithecus was first established by G.E. Lewis in 1934. A fragmen- tary right maxilla (YPM 13799) discovered in the Siwaliks region contiguous to India and Pakistan was assigned by Lewis to R. brevirostris. Later, Simons and Pilbeam (1965) revised the nomenclature, reducing this specimen and other morpho- logically similar specimens into a single species-R. punjabicus. In the 1970s fossil remains of Ramapithecus increased considerably so that today there are materials from Africa (Kenya), Asia (India, Pakistan, Turkey, China), and Europe (Hungary and Greece).

16 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
Among these discoveries many, which were initially assigned new genera, have later been included within Ramapithecus. It is well to take note, however, that morphologically many of these specimens retain a number of real differences. They are widely distributed both spatially and temporally throughout Eurasia from the middle Miocene to the middle-Pliocene. As many investigators have frequently created new species based purely on their own individual assessments, there is still today no general agreement as to how to classify these specimens on the species level. We will attempt a broad comparison and try to clarify the classificatory position and systematic relationships of R. lufengensis.
The Kenyan material was discovered in late Miocene deposits a t Fort Ternan and has an absolute date of approximately 14 million years B.P. The initial specimen was an upper jaw assigned by Leakey (1962) to Kenyapithecus wickeri. In 1971 Andrews, while researching a left mandibular fragment (KNM-FT45) recovered from the same locality, noticed that it occluded with the previously mentioned maxilla (KNM-FT46). He consequently referred both specimens to the same species and individual, and revised the nomenclature as R. wickeri. According to the recon- struction of the dental arcade proposed by Walker and Andrews (1973) the tooth rows are straight and diverge slightly posteriorly. In this respect the dental arcade has a pongid character unlike the arcuate arcade of the Lufeng PA580 mandible. The incisors of the Kenyan mandible would have been very small. On first glance, the P3 appears unicuspid but it actually has developed an incipient metaconid tending in this regard in the direction of the Hominidae. The P3 is low crowned and semisectorial, and has lingual cingulum that continues to the posterior margin. These characters are primitive Proconsul-like retentions. The talonid of P4 is elon- gate and its long axis is set obliquely in the tooth row. The Lufeng PA580 mandible has moderate-sized incisors. P3 is definitely bicuspid and triangular in outline. The lingual cingulum does not continue to the posterior margin. P4 has a relatively short talonid which is not turned inward. These characteristics are much more derived than those seen in the Kenyan mandible.
A lower jaw discovered in late Miocene deposits in Candir, Turkey, was initially classified by Tekkaya (1974) as Sivapithecus alpani. Subsequently, Tekkaya and Andrews (1976) referred it to R. wickeri. This mandible is very well preserved, holding left P3-M3 and right P4-M3. Its dental arcade is like that of the recon- structed mandible KNM-FT45; the long axis of P3 is obliquely set; the dental row from P3 to M3 is straight and diverges slightly posteriorly. The angle of divergence of the dental arc is very small. In contrast, the dental arcade of PA580 is regularly arcuate; the dental row gradually curves inward from M3 to 12, and P3 does not sit obliquely. Consequently the anterior portion of the dental arcade is rounded as in the Hominidae and the angle of divergence of the dental arcade is rather large. Based on the size of their alveoli, the incisors and canines of the Candir mandible appear to have been small. P3 is a low-crowned elongated tooth possessing a primary cusp (protoconid) and a small secondary cusp (metaconid), but it is not as obviously bicuspid as the P3 of the PA580 mandible. P4 is also obliquely set whereas the Lufeng P4 is not. Molar size and morphology are similar to R. lufengensis, with low- cusped even crowns and steep lateral sides. M3 has two auxiliary cusps. All the molars of the Candir mandible have traces of buccal cingulum; this is especially obvious on M3. Buccal cingulum is a primitive trait of Proconsul where it is highly developed. It is nonexistent on M3 of the Lufeng specimen.
Fossil teeth have been discovered from mid-Miocene deposits a t Pasalar in Turkey. These have been assigned to two types by Andrews and Tobein (1977): Sivapithecus darwini and R. wickeri. The tooth size and morphology of the latter are both analogous to the Candir material and somewhat more primitive than Lufeng.
From the above exposition it can be seen that R, wickeri is in a number of evolutionarily significant ways clearly distinct from R. lufengensis. R. wickeri is comparatively primitive, retaining quite a few characteristics of Proconsul; R. lufen- gensis is comparatively progressive, showing features that approximate Homo.
The lower jaws of Ramapithecus discovered in India were all initially assigned to different genera. For example, the two mandibular fragments discovered by Pilgrim

Etler] FOSSIL HOMINOIDS OF LUFENG 17
(1915) in the Chinji deposits of the Siwaliks (GSI D-118/119) were at first classified as D. punjabicus.
Another mandibular fragment (YPM 13814), likewise from the Chinji deposits, was described by Lewis (1934) as Bramapithecus thorpi. Later, Simons and Pilbeam (1965) subsumed both under R. punjabicus. These three mandible fragments all have small sized, low-crowned molars with M3’s having a metaconulid and sixth cusp similar to the condition seen in PA580. The Indian specimens’ molars are, however, somewhat smaller in size with a relatively long, narrower coronal outline. They have obvious buccal cingulum which surrounds a small fovea, suggesting that they are more primitive than the Lufeng specimen. Deserving special attention is the M3 of the GSI D-118 mandible, which in the arrangement of the five primary cusps and the form of the two auxiliary cusps, the structure of the buccal cingulum, and the squared contours of the dental crown, approximates the left M3 of P ufricanus (Clark and Leakey, 1951). The latter M3 is smaller, with more developed cingulum which has a blunted denticulate form. Their similarities, however, reflect the close phylogenetic relationship between the two genera.
In 1956, five lower teeth (PA75:l-5) were discovered in late Miocene lignite deposits at the Xiaolongtan colliery, Kaiyuan, Yunnan. These specimens were orig- inally ascribed by Wu (1957, 1958) to D. keiyuanensis and later referred to as ramapithecus sp. Comparing the Kaiyuan and Lufeng PA580 lower dentitions, the former is somewhat lower crowned and smaller in size, and P4 has an elongated talonid. All indicate a more primitive condition than in the latter and are very much akin to the Kenyan material. The Kaiyuan Mz and M3 are morphologically similar to the Indian specimens GSI D-118/119. Mid and late Miocene African and Asian ramapithecines can be seen, then, to have been morphologically quite comparable.
In the last several years Ramapithecus and Sivapithecus fossils have been discov- ered in early and middle-Pliocene beds of the Potwar Plateau of Pakistan. Pilbeam et al. (1977) have ascribed the ramapithecine specimens to R. punjabicus. There are two particularly important specimens in this material, a comparatively complete lower jaw containing left M1-M3 and right M3 along with sockets and alveoli of the missing teeth (GSP4622/4857), and a right mandibular fragment containing P4-M3 (GSP6153). The molars of these two specimens are in morphology and size very similar to the Lufeng PA580 molars with thick-enameled crowns and enamel win- dows on the occlusal surface. There are a number of important traits, however, which distinguish the Lufeng mandible from the Pakistani material: (1) The angle of divergence of the dental arcade of GSP4622/4857 is especially large. This large degree of divergence is never seen in early representatives of Homo but is seen in Australopithecus. In contrast, there is less of a degree of angular divergence of the PA580 mandible dental arcade, making it more comparable to the condition seen in early Homo. (2) The dental arcade of the Pakistani mandible is not regularly arcuate. P3 projects outward; P4 and M1 are deflected inward, causing this area of the tooth row to be bow-shaped. This is never seen in Homo. PA580, however, has a regularly arcuate dental arcade very close in shape to that seen in early man. (3) The incisor region of the Pakistani mandible (GSP4622/4857) is very narrow, indicating that its incisors were very small. Its molars are, in contrast, fairly large. This condition is comparable to that of Australopithecus. The incisor size of the PA580 specimen is moderate. The whole dental series approaches that of early man.
Among other Pakistani materials, the P3 from the GSP7619 mandibular fragment has a small but obvious metaconid, thereby resembling PA580 in being incipiently bicuspid. The basal mesiolingual cingulum is bow-shaped and continues to the posterior margin. The tooth lacks anterior buccal cingulum. These characters are unlike that seen in the Lufeng specimen and are more similar to those seen in the Kaiyuan specimen.
In 1975, Kretzoi made a brief description and comparison of ramapithecine mate- rial discovered in lower Pliocene deposits from Rudabaya, Hungary. It was named Rudapithecus hungricus, but many scholars have referred it to Ramapithecus. The material includes two mandibular fragments, Rud-l and Rud-17. The straight ap-

18 YEARBOOK OF PHYSZCAL ANTHROPOLOGY [Vol. 27, 1984
pearance of the tooth row from the premolars to the molars seems to indicate that the dental arcade could not have been arcuate as in the Lufeng specimen. In its dental features, the Rud-1 P4 is broader than MI, and M3 is especially long. The Rud-17 canine is relatively high crowned and sharply tipped. P3 is unicuspid and sectorial and much larger than Pq. It is pongid in character and clearly unlike the Lufeng specimen.
In Greece, a mandible was discovered early in World War 11, but it was not until 1972 that it was formally described by Von Koenigswald and designated Graecopi- thecus freyburgi. This specimen is from Miocene deposits. Unfortunately, its denti- tion has been extensively damaged. Not long afterward, Simons (1977) in a review of the ramapithecines, pointed out the close relationship between this mandible and those from Fort Ternan and Candir.
From the above systematic comparisons we come to the following conclusions: (1) Given the present evidence, R. lufengensis more closely approaches early Homo
than do other members of the genus. As such, it is possible that it is a direct hominid ancestor transitional between ape and man.
(2) From the mandibular evidence it can be seen that the ramapithecines of the middle and late Miocene generally have primitive characters close to Proconsul, indicating a close phylogenetic relationship. Early and middle-Pliocene specimens from Pakistan, Hungary, and China show a tendency toward morphological differ- entiation. This phenomenon is most likely related to worldwide orogenesis and concomitant changes in climate and habitat. Based on these considerations we suggest the following preliminary assessment of the origin and development of the ramapithecines: Ramapithecus probably arose from the early Miocene Proconsul of Africa. At first, most likely in the middle-Miocene, it became dispersed in Africa and neighboring regions, finally becoming widespread throughout southern Europe and Asia. In response to continued and varied climatic shifts and corresponding adjustments in lifeways and diet the ramapithecines of the early and middle-Pli- ocene began to differentiate regionally, possibly in diverse ways.
(3) In the past, China has recorded the discovery of the Kaiyuan Ramapithecus of the late Miocene, early Pleistocene teeth of A ustralopithecus from Jianshi, Hubei, and of H. erectus from Yuanmou, Yunnan; and fossils of H. erectus from Lantian and Beijing. The discovery of R. lufengensis not only fills a gap in the documentation of the evolutionary transition from ape to man during the Chinese Pliocene; it also proves that southern China was an important region in the origin of mankind and moreover that this area has a very high probability of producing middle and late Pliocene to early Pleistocene hominid fossils.
Sivapithecus yunnanensis
Family: Pongidae Genus: Sivapithecus Sivapithecus yunnanensis sp. nov. Holotype-A relatively complete mandible containing a full complement of ante-
rior teeth (I,C,P) and left MI and Mz. In addition isolated right MI and Mz belonging to the same specimen (PA548). Diagnosis
The mandibular symphysis is very deep and the mandibular corpus becomes gradually shallower posteriorly. The inferior surface of the symphysial region has a clear spina digastrica, and its two sides have a broad flat fossa digastrica. There is a posteriorly projecting, thick simian shelf. The incisors are especially high crowned; the canines are tall, sharply tipped, and pyramidal. The molars have a deep notch on the posterior lingual margin of the metaconid apex. The crenulations on the occlusal surface of the cheek teeth are very complex. Description
The PA548 mandible is robust and thick, and is of an adolescent male. The mandibular symphysis is very deep. The center of its exterior surface bulges slightly,
Etlerl FOSSIL HOMINOIDS OF LUFENG 19
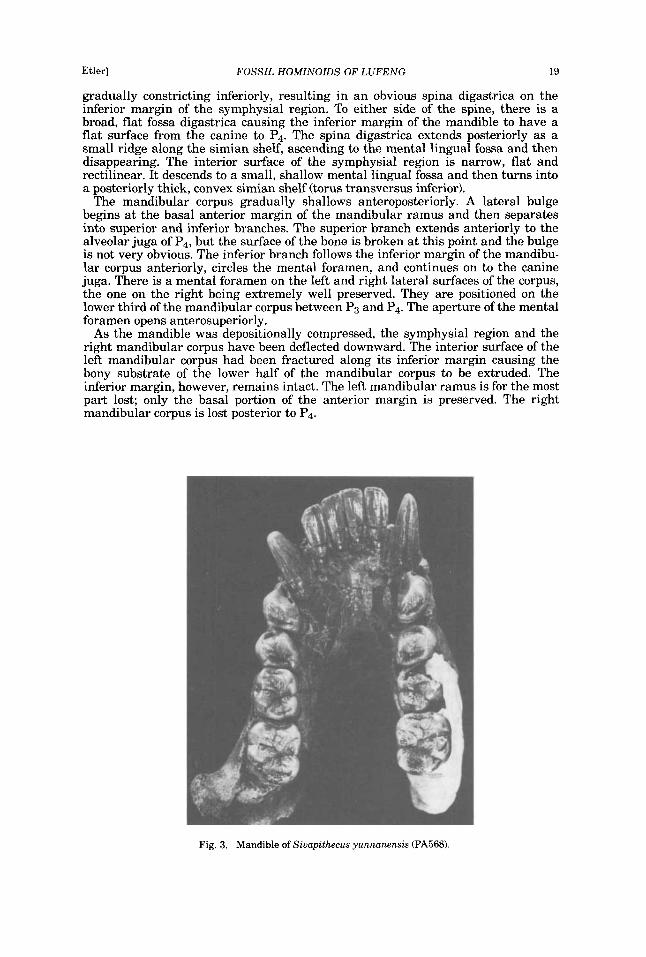
gradually constricting inferiorly, resulting in an obvious spina digastrica on the inferior margin of the symphysial region. To either side of the spine, there is a broad, flat fossa digastrica causing the inferior margin of the mandible to have a flat surface from the canine to P4. The spina digastrica extends posteriorly as a small ridge along the simian shelf, ascending to the mental lingual fossa and then disappearing. The interior surface of the symphysial region is narrow, flat and rectilinear. It descends to a small, shallow mental lingual fossa and then turns into a posteriorly thick, convex simian shelf (torus transversus inferior).
The mandibular corpus gradually shallows anteroposteriorly. A lateral bulge begins at the basal anterior margin of the mandibular ramus and then separates into superior and inferior branches. The superior branch extends anteriorly to the alveolar juga of P4, but the surface of the bone is broken at this point and the bulge is not very obvious. The inferior branch follows the inferior margin of the mandibu- lar corpus anteriorly, circles the mental foramen, and continues on to the canine juga. There is a mental foramen on the left and right lateral surfaces of the corpus, the one on the right being extremely well preserved. They are positioned on the lower third of the mandibular corpus between P3 and P4. The aperture of the mental foramen opens anterosuperiorly.
As the mandible was depositionally compressed, the symphysial region and the right mandibular corpus have been deflected downward. The interior surface of the left mandibular corpus had been fractured along its inferior margin causing the bony substrate of the lower half of the mandibular corpus to be extruded. The inferior margin, however, remains intact. The left mandibular ramus is for the most part lost; only the basal portion of the anterior margin is preserved. The right mandibular corpus is lost posterior to P4.
Fig. 3. Mandible of Siuupithecus yunnanensis (PA568).

20 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
Although the symphysial region and the right mandibular corpus have been shifted, the left tooth row from the canine to M2 is preserved in its original orienta- tion. Because of this, an estimate of the form of the dental arcade can be made. As the left cheek teeth are arranged in a straight line, the P3 and canine are only slightly directed medially, and the canine juga is obviously rotated, the dental arcade can be seen to be close to “U”-shaped with only a small angle of divergence.
The incisor crowns are especially high. The incisors are far wider labiolingually than anteroposteriorly. The central and lateral incisors are equally broad, and the cutting margin is thin. The cutting margin of the central incisor is level, intersecting the mesial and distal surface at right angles. The distal end of the cutting margin of the lateral incisors slants downward, the distal angle is obtuse, and the mesial angle is right. The lingual surfaces of the incisors all have obvious marginal ridges.
The canines are tall crowned, sharp, and pyramidal in shape. Their apices are well above the dental plane of the cheek teeth. The base of the crown is oval in cross section, wider anteroposteriorly than labiolingually. The anterior marginal ridge is sharper than the posterior marginal ridge. The posterior marginal ridge divides into labial and lingual branches. In between, there is a deep vertical groove. There are shallow vertical grooves on the labial surface of the crowns to the side of the anterior and posterior marginal ridges. Vertical crenulations are spread over the canines’ lingual surfaces. There is obvious cingulum at the base of the mesial lingual surface. The canine has weak buccal cingulum. The canine root is implanted at a slight incline in the alveolus.
P3 is sectorial, unicuspid, tall, and sharply tipped. A sharp primary ridge slopes from the apex of the crown toward the base of the posterointerior corner, stopping at the lingual cingulum. It divides the lingual surface into two sloping planes. The mesial lingual plane is relatively flat; superiorly it has several vertically directed crenulations. The distal lingual plane is concave similar to a talonid basin; medially it has many minute crenulations. The mesial marginal ridge and distal marginal ridge are relatively sharp. The whole crown is shaped like a three-sided pyramid, which is also characteristic of the Pleistocene fossil orangutan. Midway along the primary ridge is a slightly swelled cuspulid. The mesial lingual cingulum is very heavy. It curves anteriorly toward the anterior buccal cingulum and posteriorly toward the posterior margin. The base of the buccal surface has level growth lines.
The P4 crown is piriform in outline. Its posterointerior corner protrudes strongly. A transverse ridge which runs between the buccal protoconid and the lingual metaconid, which intersects the vertical groove. Anterior to the ridge there is a small transverse anterior fovea. Posteriorly there is a large, deep talonid basin. Within the two depressions there are minute crenulations. The mesial marginal ridge is cut medianly by the vertical groove. The marginal ridge in the posteroinfer- ior corner is cut by a sloping extension of the vertical groove, becoming a small notch.
The lower molars have five primary cusps arranged in a typical dryopithecine or Y-5 pattern. The occlusal surfaces have complex crenulations. M2 is much larger than MI. The endoconid of M1 protrudes a bit lingually. The vertical groove that separates the metaconid and hypoconulid passes through the posterior fovea and then cuts across the distal marginal ridge. This is not the case with Ma. Between the protoconid and hypoconid of M1 there is downwardly inclined buccal cingulum which converges at the groove between the two cusps to create a small pit. M2 also has buccal cingulum but not nearly as evident as in MI. There is a distinct notch on the posterolingual margin of the apex of the molar metaconid. Neither of the distal surfaces of the M2’s have interstitial wear or contact facets, so it can be inferred that the M3’s had not yet erupted.
Comparison and discussion From the morphological description it is evident that both in size and morphology
the PA548 mandible is clearly distinct from R. lufengensis (PA580). The difference is most noticeable in the shape of the dental arcade, the morphology of the anterior

Etler] FOSSIL HOMINOIDS OF LUFENG 21
teeth including P3, the proportional size of M1 and Ma, and the general character of the mandible. These differences in size and morphology are also encountered in sivapithecine and ramapithecine material from Pakistan, India, and Turkey.
The Lufeng PA548 mandible most closely resembles that of S. indicus in size and morphology and somewhat less so S. siualensis. It differs considerably from Dryopi- thecus. This discussion will concentrate, therefore, on comparisons of Sivapithecus materials.
A typical mandible of S. indicus (YPM 13828), in terms of mandibular robustness, and the morphology and size of the cheek teeth, is very similar to PA548. In both cases molars are bunodont, Mz is clearly larger and broader than MI, M2 has a comparatively large posterior fovea that lacks an anteroposterior vertical groove passing through it, and there are thickly enameled dental crowns. The P4 talonid of YPM 13828, however, does not protrude posterointeriorly, the molar buccal cin- gulum is not obvious, the posterior lingual margin of the metaconid lacks traces of a notch, and the distance between the base of the anterior margin of the mandibular ramus and the facing buccal surface of the molars is relatively large. These are all characteristics unlike those of PA548. Even though the cheek teeth of the Indian specimens are not heavily worn, there are no visible crenulations on their occlusal surfaces. Other dental specimens of S. indicus also show crenulations that are slight and undefined. The occlusal surfaces of the cheek teeth of the Lufeng PA548 man- dible, in contrast, have a degree of complexity corresponding to that seen in the South China fossil orangutan of the early and middle Pleistocene. In addition, unlike other Sivapithecus mandibles, PA548 has a wide, flat fossa digastrica.
Isolated P3’s of S. indicus at times resemble those of PA548. A minute swelling or cuspulid (not yet a small cusp) appears in the center of the primary (metaconid) ridge. This character also exists in S. siualensis. Gregory and Hellman (1926) recog- nized this small, cuspulid-like swelling as an incipient metaconid. This characteris- tic is also encountered in present-day large-bodied apes, but is clearly unlike the bicuspid P3 of R. lufengensis.
The anterior mandibular fragment (AMNH19411) and the left mandibular corpus (AMNH19412) discovered in India were at first designated D. pilgrimi and D. cautleui, respectively. They were later ascribed to S. siualensis and served as the basis for reconstructing the mandible of this species. These two fragmentary speci- mens are clearly much smaller than the PA548 mandible. Morphologically, there are also some major differences. For example, according to the description of Gregory and Hellman the digastric fossa of AMNH19411 divides into two elongated, finger- shaped fossae on either side of the spina digastrica and the lower margin of the symphysial region is very narrow and lacks a posteriorly extended simian shelf. These conditions are quite unlike those seen in PA548.
With respect to the morphology of the AMNH19412 mandible, the tooth row from C1 to M3 is straight, C1 is in a slightly buccal position, and the basal cross section of C1 crown is almost circular. P3 lacks anterior buccal cingulum and has a narrow distal lingual surface. P4 crown is rhomboidal, molar buccal cingulum is not marked, and the lingual margin of the metaconid lacks a notch. The occlusal surface of the molars has only slight crenulations. These features all differ from the PA548 speci- men and are noticeably more primitive.
Geochronologically these two mandibular fragments are from late Miocene depos- its. The PA548 and Indian YPM 13828 mandibles are from early Pliocene deposits. The latter are morphologically closer to the Pleistocene fossil orangutan and extant orangutan, and reflect a developmental trend in that direction. Gregory et al. (1938), in summarizing late Miocene to early Pliocene fossil hominids of India, had already stated that “When looking at the whole picture it can be conceived that a graded series exists from S. sivalensis to S. indicus . . . with the modern orangutan possibly being a branch of this lineage.”
In 1957, at the Xiaolongtan colliery, Kaiyuan, Yunnan, China, five lower teeth from the right tooth row (P3-M3) (PA82:1-5) of one individual were discovered. These teeth are relatively close in size and morphology to those of S. siualensis

22 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
(AMNH19412). The teeth of the Kaiyuan specimen are, however, a little bit larger. P3 has strong anterior buccal cingulum, and M3 has a small metaconulid and sixth cusp. The latter two traits are unlike the Indian specimens and closer to the Lufeng specimens. This suggests that the Kaiyuan and Lufeng specimens could have a phylogenetic relationship.
In the last few years, Pilbeam et al. have discovered Sivapithecus fossils in Pakistan which have been assigned to S. sivalensis and S. indicus. In regard to the former species, Pilbeam et al. did not at first make a morphological description of it. For the latter species, they have raised a number of important characteristics, among them: the tooth row is subparallel, canines are projecting, P3’s are closely approximated to the canine, there is thick enamel on the occlusal surface, and there is a long symphysial region with a prominent interior transverse torus (simian shelf). These characteristics resemble those of the Lufeng PA548 mandible. Other characteristics of the Pakistan specimens, i.e., postcanine tooth rows that are mark- edly concave buccally and a superior transverse torus, are different from that in the Lufeng specimen. Pilbeam et al. raise the idea that the Pakistani material can be classified into two subfamilies: Sivapithecinae and Ramapithecinae. Moreover, they recognize that Sivapithecinae have dental traits of A ustralopithecus and early Homo. Because of this, they place it and Ramapithecinae together in the family Ramapithe- cidae. In looking at the dental morphology of PA548, however, it can be seen to fundamentally differ from Australopithecus or early Homo. On the contrary, it appears extremely close to the south China early- to middle-Pleistocene fossil oran- gutan. Based on this assessment, we recognize the situation of Sivapithecus to be exceedingly complex. This genus in the early Pliocene may have already differen- tiated into a number of types with the Lufeng PA548 species perhaps developing toward Pongo.
Sivapithecus darwini fossil teeth from the previously mentioned Pasalar site in Turkey are described by Andrews as having cheek teeth morphologically similar to the Ramapithecus material from the same locality. However, its other characteris- tics, i.e., P4’s compressed trigonid and possession of buccal cingulum, lower molars with noticeable buccal cingulum, and lower molars in general proportionally similar to Proconsul, indicate that the Pasalar specimen is in comparison a bit more primi- tive than the Lufeng material. Similarly, its geologic age is relatively older.
Bodvapithecus altipalatus (Kretzoi, 1975) from early Pliocene deposits of Hungary is now recognized as a type of Siuapithecus. Looking at its dental characteristics, the Rud-14 right mandible has premolars with noticeable posterior buccal cingulum and molars with obvious buccal cingulum and relatively robust crenulations on occlusal surfaces. These traits are clearly unlike those of the Lufeng specimen.
European dryopithecine fossils, e.g., the French D. fontani mandible, have widely spaced lower incisors with low crowns, a relatively large diastema between C1 and P3, and cheek teeth with relatively developed cingulum and laterally flaring molar cusps. These characteristics are absolutely different from PA548, and moreover, obviously differ from other Sivapithecus species. The Lufeng specimen is then com- paratively distant from Dryopithecus.
Finally we should bring up the late-Miocene discovery of Ouranopithecus macedon- iensis (de Bonis and Melents, 1977) fossil jaws from Greece. The type is based on four relatively complete lower jaws and other mandibular fragments. The morphological characteristics of the four mandibles include strong, large jaws; robust dentition surpassing that of other sivapithecine specimens in size; a noticeable simian shelf; and an arcuate dental arcade. The angle of divergence of the dental arcade among males is relatively small and close to “U”-shaped (RPI75). The teeth are widely spaced, there being large diastemata between the canines and its anterior and posterior teeth. [Reviewer’s note: this is due to postmortem distortion of the mandi- bles.] The canines show a large degree of sexual dimorphism. The male canine projects strongly; P3 is unicuspid, having noticeable mesial, lingual cingulum which merges with the posterior margin. The primary ridge of the female specimen P3 (RPI54) is developed and runs transversely, becoming a horizontal broad talonid, but it does not become bicuspid as in Ramapithecus lufengensis.

Etler] FOSSIL HOMINOIDS OF LUFENG 23
Lower molars have obvious buccal cingulum, and M3 has a metaconulid and sixth cusp producing a very complex pattern. M3 is proportionally largest among the molars. These basic morphological features, although comparable in a number of ways to Sivapithecus and Ramapithecus, are in sum actually more similar to Gigan- topithecus. As de Bonis and Melentis (1977) point out: “This is a species close to Sivapithecus, Bodvipithecus and Ramapithecus but also very close to an ancestor of Gigantopithecus.”
Based on the above comparisons, we offer the following conclusions: (1) In size and morphology, the Lufeng PA548 mandible is very close to Asian
Sivapithecus. Its differences with European Dryopithecus are relatively great. Among Sivapithecus species, the Lufeng specimen has “specific characteristics’) which dis- tinguish it from others. We have, therefore, assigned it to S. yunnanensis sp. nov.
(2) In a morphological comparison with already known Neogene fossil pongids, S. yunnanensis most closely approximates the Pleistocene fossil orangutan. It could be, therefore, an ancestor to the widely distributed south China and southeast Asian fossil orangutan and modern Pongo.
(3) Throughout Eurasia, wherever Sivapithecus has been found, it has always been associated with Ramapithecus except in Africa. In its early occurrence, i.e., the early middle Miocene Turkish specimen, it has certain morphological traits of Proconsul and Ramapithecus. This could very possibly indicate that both Sivapithecus and Ramapithecus arose from Proconsul.
In the early and middle Pliocene species, the Pakistani material is recognized to possess certain characteristics of A ustralopithecus. The Lufeng specimen, however, is close to the fossil orangutan, reflecting the tendency toward differentiation of Sivapithecus at this time.
LU QINGWU, XU QINGHUA AND ZHENG LIANG (1981) PRELIMINARY RESEARCH ON THE CRANIUM OF SIVAPITHECUS YUNNANENSIS. VERTEBRATA PALASIATICA. 19(2):101-106.
ABSTRACT This is a descriptive and comparative account of the first fossil hominoid skull assigned to the species Sivapithecus yunnanensis discov- ered at the Shihuiba colliery site, Lufeng county, Yunnan, in December 1978. Morphologically the skull retains certain primitive characteristics of Procon- sul africanus while sharing many derived features with the modern orangu- tan. The authors suggest that this fossil type may be a link between the two species. At the same time the authors recognize that the skull shows some close similarities to the robust australopithecines and they suggest that S. yunnanensis may have an ancestral relationship to A ustralopithecus robustus.
In December 1978, a joint excavation team of the IVPP (Academia Sinica) and the Yunnan Provincial Museum discovered a relatively complete fossil skull of Sivapi- thecus at the Shihuiba colliery site, Lufeng county, Yunnan (WPP # PA644). The fossil (not including mandible) was found lying in a ventral position compressed within a brown lignite stratum. The facial portion of the skull is basically well preserved and intact. The cranium, however, is somewhat damaged. The forehead shows a trigonum frontale; the parietals are lost; all that remains of the occiput is a small area encircling the foramen magnum. The portion above the zygomatic process of the temporal bone is missing. Specimens of the different dental morphs are preserved; the crown of the left canine is lacking. As the fossil was subject to compressive forces within the lignite stratum it has been crushed flat. The facial bones are tightly packed with the cranium and difficult to separate. At the time of writing, the fossil is still contained within the lignite block. The following is a preliminary account and discussion of the skull.
Comparison The teeth of this specimen match well with those of the type specimen of the
species (PA548) and are comparatively different from those of Ramapithecus lufen-

24 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
gensis. As far as its dental characteristics are concerned there is no doubt that the specimen should be referred to S. yunnanensis. According to its dental wear patterns and the morphology of the canines, it is inferred that the specimen was an adult male.
The Lufeng hominoids lived during the early Pliocene extending to the early middle Pliocene. Up to today there have been no finds of fossil hominoid skulls as complete as this one from the end of the Miocene to the early Pliocene.
Comparison with Proconsul africanus First, we will compare the skull with the early Miocene skull of l? africanus from
Kenya, East Africa. The Proconsul skull is round and small with a light construction lacking robust
bony ridges. It has a sharply pointed snout very much like that seen in catarrhine monkeys. The S. yunnanensis skull is, on the contrary, much larger, of robust construction, with developed bony ridges (although its supraorbital ridge is not very evident, retaining a primitive form as in Proconsul), and a broad, foreshortened face.
Upper incisors Proconsul incisors are spatulate and the incisive edge is relatively thin. S. yunna-
nensis has large, broad incisors with a thick incisive edge and relatively developed basal lingual tubercles. The upper lateral incisors are much smaller than the central incisors. The labial surface of the incisors has a number of transverse ridges.
Upper canines The dental crowns of both Proconsul and S. yunnanensis are pyramidal, but those
of Proconsul are smaller and shorter with blunted tips; the mesial edge has a rather deep vertical groove; the superior portion of the groove terminates in a small fovea and there is a pronounced anterior cingulum close to its bottom. The upper canine of S. yunnanensis has a similar construction, but the crown is more robust, larger, sharper, and more projecting.
There is a diastema between the upper incisors and the upper canines in S. yunnanensis; this is not seen in l? africanus. l? africanus is a gracile-bodied early hominoid; other than its teeth, which have a variety of hominoid features, its construction is much like that of a monkey. As its phylogenetic position is relatively primitive, P africanus still does not have the secondarily derived characteristics of later hominoids (i.e., developed supraorbital ridges). S. yunnanensis, however, al- ready has a variety of characteristics seen in later hominoids.
Comparison with A ustralopithecus robustus We will now compare the skull specimen with the later A. boisei (Zinjanthropus). The two specimens have the following clear similarities: the forehead has a
trigonum frontale, there is a broad interorbital septum, and the zygomatic archs and the zygomatic processes of the maxilla flare outward laterally; therefore the temporal fossa is massive, yielding a noticeably broad face (Table 1).
Forehead and supraorbital region The supraorbital ridge of “Zinjanthropus” is more robust than that of S. yunna-
nensis. The glabellar protuberance of “Zinjanthropus” is also rather developed, forming a supraorbital torus at the junction of the lateral supraorbital ridges. The forward projection of the supraorbital torus greatly exceeds that of the lateral
TABLE 1.
Upper face width (Ft-Ft) Face width (Zy-Zy) Interorbital width CLa-La)
“Zinjanthropus” 115.4 mm 160 mm 32.5 mm S. vunnanensis 103 mm 175 mm 34 mm

Etler] FOSSIL HOMINOIDS OF LUFENG 25
supraorbital ridges. The relevant portion of the S. yunnanensis skull was caved in during the depositional process. In the hollowed-out area that resulted, a small ridge joining together the lateral supraorbital ridges can still be made out. It can be inferred that before the glabellar region was deformed, there was a ridgelike protu- berance, but it was probably lower than the two lateral supraorbital ridges.
The temporal ridges of “Zinjanthropus” are curved and approach the midsagittal plane of the skull vault sooner than do those of S. yunnanensis. The temporal ridges of S. yunnanensis are straighter. The apical angle of the trigonum frontale of “Zinjanthropus” (30”) is smaller than that of S. yunnanensis (50”). The point of intersection of the temporal ridges in the midsagittal plane is situated more ante- riorly in “Zinjanthropus” than in s. yunnanensis; therefore, the surface area of the trigonum frontale is larger in the latter than the former. Since the temporal ridges of “Zinjanthropus” are not as robust or projecting as those of S. yunnanensis, the surface of its trigonum frontale is only slightly concave. That of S. yunnanensis has a relatively deeper concavity. The temporal ridges of “Zinjanthropus” run parallel to the orbital margins and extend medially almost as far as the superomedial angle of the orbit before deviating posteriorly, in this manner utilizing the maximum of space anteriorly for the attachment of the temporalis muscle (Tobias, 19671, and expressing the fact that this musculature was very well-developed.
The temporal ridges of S. yunnanensis are also very robust and similar in nature, but posterior to the bend in direction they are relatively straight, and the bend takes place at about one-half of the supraorbital ridges. It can be seen that the placement and degree of development of the masseter muscles in both species is rather similar.
Nasal aperture and subnasal region The apertura piriformis of S. yunnanensis is elongate, while that of “Zinjanthre
pus” is horizontally broad and somewhat spheroid and, moreover, has an anterior nasal spine. This is not evident in S. yunnanensis.
The corpus of the “Zinjanthropus” maxilla is more prominent than the zygoma and constitutes a relatively large proportion of the facial skeleton. This proportion is comparatively less in S. yunnanensis. Therefore, the nasoalveolar clivus of the maxilla of “Zinjanthropus” is longer than that of S. yunnanensis. It is also slightly concave and the area on either side is relatively high. The anterior portion of the maxilla of “Zinjanthropus” is somewhat constricted and steep. This area in S. yunnanensis, on the other hand, is relatively prognathous.
Anterior maxillary teeth. There is a great disparity between the relatively small lateral incisors of S. yunnanensis and its large, broad central incisors. This disparity does not exist in “Zinjanthropus.” The superior portion of the incisors of S. yunna- nensis have obvious ridges on their lateral lingual margins. These are not evident on the inferior portion. The lingual surface has fairly well-developed basal tubercles. The labial surface of the incisors have a number of transverse ridges. The incisors of “Zinjanthropus” are weakly to moderately shovel-toothed and chisellike in form, their labial surfaces are smooth, without transverse ridges.
The canines of “Zinjanthropus” are incisorform and do not project beyond the plane of the other tooth crowns. The canines of S. yunnanensis, however, do project beyond the occlusal surface of the other tooth crowns and, moreover, there are diastemata between them and the lateral incisors. The occlusion of the upper and lower jaws forms a “canine-premolar complex” in S. yunnanensis. “Zinjanthropus” lacks diastemata and the above-mentioned dental complex.
Comparison with the present-day orangutan In our appraisal of the S. yunnanensis mandible (PA,548) we noted that “This
species was perhaps widely distributed over southern China and southeast Asia and ancestral to both the fossil and extant orangutan.” Presently, we will compare the skull specimen with that of an orangutan (Indonesian specimen #925).
The frontal portion and facial skeleton of both specimens are, in general, con- structed along the same lines; i.e., both have a frontal trigone, a relatively broad

26 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
face, a long, narrow piriform aperture, and outwardly flaring canines. Below are more detailed comparisons.
Forehead and orbital region The convergence of the lateral temporal ridges toward the midsagittal plane is
more pronounced in the orangutan than in S. yunnanensis. The apical angle of the orangutan trigonum frontale is 60°, that of S. yunnanensis 50”. The point of inter- section of the temporal ridges is more anteriorly placed in the orangutan than in S. yunnanensis so that the area of the frontal trigone is less in the former than the latter specimen.
The two lateral temporal ridges of the orangutan start out tangent with the left and right outer orbital margins. The temporal ridges and the rugged bony ridges of the supraorbital margins gradually converge toward the frontal-sphenoidal process of the zygomatic; i.e., the two ridges merge as a rugose distal protuberance at the zygomatic process of the frontal bone. Below the zygomatic-frontal suture, the surface of the frontal process of the zygomatic process is very smooth.
The lateral sides of the frontal trigon of S. yunnanensis intersect the midpoint of the lateral supraorbital margins. There is no obvious bony supraorbital ridge of the narrow rugose type seen in the orangutan. The ridge goes along the supraorbital margin laterally to the zygomatic-frontal suture. The bony surface along this route is comparatively flat and smooth.
W.E. Le Gros Clark and L.S.B. Leaky (1951) pointed out, “In the absence of a strong supra-orbital torus the orang clearly preserves a primitive feature in which it resembles Proconsul more closely than do the African apes, though, in fact the morphology of the orbital region in this Asiatic ape is very different in other details from that of P. africanus.”
Neither the orang nor S. yunnanensis have a glabellar prominence which links together their supraorbital margins into a robust supraorbital torus as seen in present-day african pongids. This indicates that the two former species may share a source closer to that of Proconsul.
From Table 2, which compares different elements of facial breadth, it can be seen that the face of S. yunnanensis is broader than the orangutan, especially as regards interorbital distance.
Piriform aperture and anterior maxillary region The piriform aperture of the orang is unlike the broad aperture of the African
apes, being basically long and narrow. Its lower portion is somewhat wider than in S. yunnanensis. The lateral margins of the orang nasal aperture are rather rounded and blunt, especially the lower portion. The upper portion in S. yunnanensis is relatively sharp and the lower portion rounded and blunt.
The subnasal aspect of the anterior maxilla of the orang extends more clearly downward anteriorly than in S. yunnanensis. The axis of the canines slopes down- ward and spreads out into an inverted “V” shape. The angle formed by these axes is larger in the orang. The distance between the alveolar process of the canine region is also larger in the orangutan than in S. yunnanensis (orang 68 mm, S. yunnanensis 53 mm).
Anterior maxillary teeth. The teeth of the orangutan, within all dental morphs, are larger than in S. yunnanensis; however, the disparity in size between the central and lateral incisors is more clearly evident in S. yunnanensis than in the orang. The orangutan canines are longer and more robust, and the diastemata between them
TABLE 2.
Upper face width (Ft-Ft) Face width (Zy-Zy) Interorbital width (La-La)
Orangutan 101 mm 165 mm 18 mm S. yunnanensis 103 mm 175 mm 34 mm

Etler] FOSSIL HOMINOIDS OF LUFENG 27
and the lateral incisors is larger than in the Lufeng sivapithecine (orang left lateral diastema 8 mm, S. yunnanensis right lateral diastema 6-7 mm).
Le Gros Clark and Leaky (19511, in analysing the present-day African apes, noted a strengthening of their bony ridges, tori, and anterior dentition which they attrib- uted to “secondary developments which appeared at a later stage of evolution” (p. 113). In comparing the skulls of the orang and S. yunnanensis a similar phenomena can be discerned, i.e., the supraorbital ridge, incisors, and canines of the former are more robust than those of the latter, as is subnasal prognathism. The modern orangutan’s robust character is, therefore, the result of autonomous secondary evolutionary processes.
Discussion In the 1950s Le Gros Clark and L.S.B. Leaky pointed out that the orang, in lacking
a strong supraorbital projection, had a feature quite analogous to that of Proconsul. S. yunnanensis, likewise, has an underdeveloped supraorbital projection; moreover, its upper canines have a relatively deep mesial vertical groove and anterior cin- gulum. These features are also held in common with Proconsul. Andrews, in his study of maxillary remains from the lower Miocene Songhor Formation in Kenya, noted that the specimen (KNM-SO 700), preserving a complete dentition except for the incisors, had features reminiscent of both Proconsul and Pongo, and placed it as a terminal species of Proconsul, possibly ancestral to the orangutan. This would be the earliest example of a relationship between Proconsul and the orang.
Similarly, the skull of S. yunnanensis clearly maintains a number of features in common with the orangutan; looking at the frontal and facial aspects, these are the lack of a glabellar protuberance linking the supraorbital margins into a rugose supraorbital torus; a relatively broad, expansive face; a long, narrow nasal aperture; and laterally flaring canines. In studying the mandible of S. yunnanensis (PA.5481, we discovered that its dental system was very similar to that of the fossil and extant orang. At that time we observed that it could be ancestral to both forms. Given that S. yunnanensis lived much later than Proconsul, it can be estimated that it repre- sented a fairly late type, relating Proconsul to Pongo, i.e., S. yunnanensis can be seen as a missing link between Proconsul and the fossil (south China Pleistocene) and living orangutan.
In our morphological comparison of the skulls of S. yunnanensis and the robust australopithecines, we also observed certain common characteristics, i.e., a trigonum frontale formed by robust temporal ridges, a relatively wide interorbital septum, a broad face, expansive temporal fossae, and laterally protruding zygomatic arches and zygomatic processes of the maxilla. In addition, in Pilbeam et al.’s (1977) research into the hominoid fossils of Pakistan, it is strongly stated that the hominid- like apes of the Siwaliks all have cheek teeth which are obviously thickly enameled and truly megadont, being very large relative to body size. In these two respects, the hominid-like apes of the Siwaliks resemble the upper Pliocene and lower Pleis- tocene hominids and are unlike the pongids. In describing the hominid-like ape subfamily Sivapithecinae, it was pointed out that they are, on one hand, similar to the australopithecines and early hominids, and on the other hand, they have simi- larities to the pongids and dryopithecine apes. We, therefore, realize that the S. yunnanensis skull clearly shows certain affinities to those of robust australopithe- cines and could possibly have a close relationship.
WU RUKANG, HAN DEFEN, XU QINGHUA, QI GUOQIN, LU QINGWU, PAN YUERONG, CHEN WANYONG, ZHANG XINGYONG, AND XIAO MINGHUA (1982) MORE RAMAPITHECUS SKULLS FOUND FROM LUFENG, YUNNAN. REPORT ON THE EXCAVATION OF THE SITE IN 1981. ACTA
ANTHROPOLOGICA SINICA 1(2):101-108 (IN CHINESE WITH ENGLISH SUMMARY).
ABSTRACT Initial observations on the excavations carried out between October and December 1981 at the Lufeng fossil ape site are reported, includ- ing comments on the stratigraphy and the hominoid and other primate fossils recovered. During this excavation three crania of Ramapithecus were discov-

28 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
ered in the third stratum. Of these, two represent juvenile individuals. In the fifth stratum, a relatively complete mandible of Sivapithecus was uncovered. The discovery of this material will be of assistance in determining the phylo- genetic position of Ramapithecus and in reconstructing the morphology of the Sivapithecus skull.
S urn mary The Joint Excavation Team of the Institute of Vertebrate Paleontology and Pa-
leoanthropology (Academia Sinica) and the Yunnan Provincial Museum excavated at the fossil ape site of Shihuiba, Lufeng County, Yunnan Province (Locality of IVPP 75033) from October to December of 1981.
The excavation area of the 1981 field season is next to that of 1980. The range of the excavation is 20 m in length and 5 m in breadth and in depth. In addition, we also excavated a part of the sixth layer of the excavation area of 1980 (19 m in length, 10 m in breadth, and 0.3-0.5 m in depth). A test pit (2 m in length, 1 m in breadth, and 3 m in depth) was dug at the northern end of the excavation section.
The excavation area is higher than that of 1980 because of topographical differ- ence. It was divided into two areas: northern area (K) and southern area (4. The thickness, composition, and structure of every layer of the new excavation section is somewhat different from that of the excavation section of the last field season.
The excavation section of 1981 is described as follows. Excavation thickness is about 5 m. Trend NE 30", from top downward in turn: (1) Yellow sandy clay with sand and gravel lenses. The clay was contaminated by
ferromagnesium and contains small quartz gravel. These materials came from the slope of a hill nearby. There is a thin sandy layer on the bottom of layer 1. It thickens westward. An erosional surface exists between layer 1 and layer 2. Thickness is 1.3- 0.6 m. Dip 220". Dip angle 5". More than ten kinds of mammalian fossils (Hipparion etc.) were collected.
(2) Blackish-grey carbonaceous clay and greyish white sand interbedded with thin lignite intercalation. The thickness of every small sandy layer is about 10-15 cm. Sandy layers are thin and thin out eastward, but organic materials increase west- ward. Sand and clay form clear, striped layers. Thickness is 2.4-1.5 m. Dip 210". Dip angle 5 "-10". With abundant fossils of primates, especially hylobatids and lorisids, and more than 20 kinds of other mammalian fossils; in addition, with fish, reptile, bird, mollusc, and plant (tree trunk) fossils, etc.
(3) Massive lignite. There are thin carbonaceous clay intercalations in the upper and lower part of the layer. Some burrow existed in the lignite. Thickness is 1.2-0.5 m. Lots of ape fossils and more than 30 kinds of mammalian fossils were found. Fish, reptile, and mollusc fossils were also common. Dip 210"-220". Dip angle 5"-10".
(4) Laminated carbonaceous clay and greyish-white silt interbedded with clear fine stratifications and lignite intercalations. Thickness is 1.0-0.3 m. Dip 210". Dip angle 5"-10". There are fossils of apes and of more than 30 kinds of mammals. A piece of tree leaf and some tree trunk fossils were found.
(5) Greyish-white sand with laminated lignite and carbonaceous clay, without clear fine and cross bedding. Thickness is 2.3-1.3 m. Dip 210". Dip angle 5"-10". With ape and more than 20 kinds of mammalian fossils.
(6) Lignite with quartz gravel, sandy clay, and laminated carbonaceous clay. Thickness is about 0.5 m. With ape and more than ten kinds of mammalian fossils. Fish, reptile, bird, and mollusc fossils were also found.
(7) Greyish-white clay with nodules of pyrite. Quartz gravel exists in the lower part of the layer. Thickness is about 1.6 m.
(8) Yellowish-brown, red clay. The weathering crust of Kunyan Group of Presinian Period. Thickness is about 0.8 m.
The layers mentioned above could be deposited in three different environments. The first layer belongs to shallow lacustrine deposit; the second to sixth layers, to lacustrine-swamp deposit; and the seventh layer, to deep lacustrine deposit. The eighth layer is the red weathering crust of Kunyan Group of Presinian Period.

Etler] FOSSIL HOMINOIDS OF LUFENG 29
We collected fossil specimens strictly according to natural layers and paid atten- tion to observing differences of number, element, and burying of fossils from each natural layer in the excavation of 1981. There were no ape and other primate fossils in the first layer, The second and third layers contained abundant fossil teeth of apes and other primates, but few skulls, maxillae, and mandibles. The fossils of skull and limb bones of large-size mammals were concentrated in the northern area (K) of the lower part of the second layer. A complete skull of hylobatids was collected from the fourth layer. Fossils decreased in number from the fifth layer to the sixth layer. The seventh and eighth layers did not have any animal fossils in the excava- tion of this field season. According to the preliminary observation, we collected in this field season some new kinds of Carnivora and Artiodactyla besides old types reported before.
Hominoid and other primate fossils-initial observations Abundant fossil remains of Ramapithecus and Sivapithecus were recovered during
the 1981 excavations. An initial assessment follows: Ramapithecus: three incomplete crania, one mandible, three mandibular frag-
ments, six upper and lower tooth rows, and 87 isolated teeth. Sivapithecus: two incomplete crania, two mandibles, one mandibular fragment,
one maxillary fragment, two upper and lower tooth rows, and 55 isolated teeth. A number of postcranial remains, possibly associated with the above-mentioned
hominoids, were also recovered. Brief descriptions of the most important specimens follow.
Ramapithecus specimens PA828. A well-preserved, nearly complete, undistorted juvenile frontal bone. Its
inner and outer surfaces are smooth and it is fairly thick walled. The frontal surface bulges. The supraorbital ridges also clearly project. There is a broad, oblique depres- sion between the supraorbital and glabellar regions. The temporal ridges are very weak, arising from the lateral margins of the zygomatic processes. They extend posteriorly in a bow shape. This condition is very unlike that seen in adult specimens and has features in common with the temporal line of modern man. The cerebral surface is concave, and cerebral grooves (impressions) and juga cerebralia are very clearly seen. The frontal crest projects strongly. There are four or five small fovea to either side posteriorly.
PA844. A largely preserved cranium which, unfortunately, is severely crushed and damaged. A very weak temporal ridge can be observed on the frontal surface. There is a very large zygomaticofacial foramen on the zygomatic surface at a point below the inferior angle of the left orbit. The osseous surface features of the basi- cranium are largely damaged, but the foramen magnum, occipital condyles, left mandibular fossa, and external auditory meatus are all very well preserved. The alveolar process of the maxilla preserves left C, P3, dM2, M1, M2; and right P3, M1, and M2. Right M3, and left are not fully erupted. The canine is small. The tooth row on the right is distorted; the left side retains its original appearance. The two sides of the dental arcade diverge posteriorly.
A very incomplete cranium which is severely fractured and fragmented. In norma verticalis, one can see a clear right temporal ridge which comes into close proximity, but does not converge with, the remains of the left temporal ridge medially at the crown of the skull. The maxilla is split and deflected downward to the base of the skull. All the teeth except 1''s and left M3 are preserved. I2 is especially small. The canine is also small.
A mandible in which the mandibular corpus is largely preserved. The mandibular corpus and symphyseal region are both rather shallow, and the basal portion of the symphyseal region has a weak digastric spine. All the teeth are in their original position in their respective alveoli. As the mandibular corpus is twisted, the tooth row is a bit distorted. The lateral sides of the dental arcade diverge posteriorly. This mandible is extremely close in size and morphology to the PA580 mandible discovered in the winter of 1976.
PA846.
PA848.

30 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
Sivapithecus specimens PA845. A severely crushed and damaged cranium which preserves all teeth
except right C. The teeth are extremely large, especially the canine and central incisor. The morphology of the tooth crowns and the complexity of the occlusal crenulations are very close to conditions seen in the Pleistocene fossil orangutan from southern China.
PA814. Cranium-other than a wholly preserved left parietal, the remainder of the skull is fragmentary. The left parietal surface is very smooth and very thinly walled. In superior aspect, one can observe a very weak, bow-shaped temporal ridge. Distinct cerebral impressions, juga cerebralia, and arterial features can be made out on the cerebral surface. A fragmentary piece of the left maxilla preserves in place three deciduous teeth (dc-dM ) and a permanent M1. Five unerupted permanent teeth, left I - p , can be seen in the damaged portion of the maxilla. Other than this, there are an isolated right dM1 and an unerupted right M2.
PA820. Mandible-the right mandibular corpus includes a relatively complete mandibular ramus giving important evidence for determining its height and breadth. The mandibular ramus has been lacking in previous mandibular specimens. The left mandibular corpus is fractured and twisted between P3 and MI, producing severe distortion. All in all, this mandible is particularly tall and robust; the symphyseal region is very deep and is very thick in basal section. Unfortunately, the surface characteristics of the bone have been largely destroyed and morphological details are hard to analyse. All the teeth are in their original alveolar position and their crowns are high and large.
Other primate specimens This excavation has produced a number of other primate fossils in addition to the
two hominoid types already discribed. These include a number of hylobatids and numerous lorisoids. Among the former specimens, there is a relatively complete cranium (not including mandible), five upper and lower jaws, and 21 isolated teeth. Among the latter, there are 23 upper and lower jaws, 31 tooth rows, and 117 isolated teeth.
The gibbon skull (PA860) is the first such discovered. The left orbit is basically preserved; the right side retains only the inferior margin of the orbit. The nasal bone is completely preserved and the left nasal aperture is damaged. The zygomatic process of the left and right maxillary corpus are incomplete. The greater part of the anterior aspect of the right maxillary corpus is preserved. The palate is rather well preserved. All the teeth are intact except that the I2 crown is incomplete.
The lorisoid maxilla (PA861) is the most complete among the lorisoid specimens. The body of the maxilla is almost totally lost and only left C - p and M2 are preserved among the teeth. The canine is shapely cusped, projecting well beyond the surface of the tooth row.
Among this group of lorisoid fossils there are typically three upper and lower premolars. P2 and P3 are both single cusped; P2 is noticeably diminutive while P3 is comparatively robust and short. p is molarized with three cusps, its morphology being very much like that of a molar.
The most complete lorisoid mandible is PA862. It preserves relatively intact the left and right mandibular corpus and on either side a remnant of the posteroinferior portion of the linea obliqua of the ramus. The alveolar margin and the inferior margin of the mandibular corpus are perfectly preserved. The whole mandible is shallow, long, and gracile. The linea obliqua projects into a crest. The mental foramina on either-side of the mandible are perfectly preserved and positioned inferiorly between C and P2. The mandibular dental arcade is of an elongated “V” shape. Only left 12 and e are preserved of the anterior teeth. The cheek teeth preserve left Pz-Ml and right P3, M1-3. P2 and P3 are similar in character to their maxillary analogues in being single cusped. P4 is, likewise, molarized. The occlusal surface of the molars shows definite signs of attritional wear as the dentine is exposed. Enamel crenulations are patterned like butterfly wings.

Etler] FOSSIL HOMINOIDS OF LUFENG 31
Discussion This was a comparatively large-scale excavation in which a great number of new
primate and other mammalian fossils were discovered. Advances have been made in ongoing studies of the area's geomorphology and paleoecology with the recording of new information and the recovery of more detailed samples. The stratigraphy and depositional environment of the fossil ape site have been studied in greater depth and stratigraphic correlations with the surrounding area have advanced.
Among the three newly discovered Ramapithecus skulls, two are of juveniles. These are the first such discoveries for this type and will help in further defining the systematic position of Ramapithecus. Among the Sivapithecus fossils, the man- dibular specimen, which preserves a relatively complete mandibular ramus (hitherto unknown), will help in the reconstruction of Sivapithecus's cranial morphology. The discovery of these additional fossil apes will have important implications for the study of human origins.
Since the 1960s, some anthropologists have recognized Ramapithecus as the earli- est representative of the hominids. Moreover, the earliest occurrence of ramapithe- cine fossils goes back some 14 million years ago. Since the most important discoveries have been in Asia, some have thought that mankind could have an Asian origin prior to 14 million years ago. In the last few years, however, based on molecular anthropology research and the discovery of very early Australopithecus remains in East Africa, there have been increasing doubts raised about the phylogenetic posi- tion of Ramapithecus. The data suggest that mankind could have diverged from its pongid ancestors 5 or 6 million years ago. Ramapithecus may not be, therefore, the earliest hominid representative.
The question of Miocene hominoid ancestors, which is also the question of the phylogeny of Ramapithecus, was addressed at a seminar held in Florence, Italy, before the commencement of the 8th International Primatology Conference in July 1980. Since the fossil material of Ramapithecus and other similar apes is meager, it is still difficult to determine their exact systematic position. The prevailing opinion was, the original idea, that man and the apes had clearly differentiated by Miocene times and that the-then members of the Hominoidea had direct ancestralldescendant relations with extant apes and man, had to be revised. Differences between Miocene hominoids are not so clear-cut and their relationships to modern forms are unclear. To determine answers to these questions we must wait for more ramapithecine fossils to be discovered and for further research.
Since 1975 there have been continuous and abundant discoveries of Ramapithecus, Sivapithecus, and associated animal fossils a t Shihuiba, Lufeng county, Yunnan, from 8 million-year-old late Miocene deposits. The 1980 discovery of a Ramapithecus skull was the first such find in the world. The 1981 excavation also revealed portions of three Ramapithecus skulls, a number of jaws, a great quantity of teeth, and fossils of the intimately associated Sivapithecus. The abundance of material far exceeds that from any other fossil ape site of this kind. Investigative results will certainly add to our knowledge of the phylogenetic position of Ramapithecus and help to resolve questions relating to mankind's time and place of origin, greatly increasing our understanding of human origins.
WU RUKANG (WOO JU-KANG), XU QINGHU AND LU QINGWU (1983) MORPHOLOGICAL FEATURES OF RAMAPITHECUS AND SIVAPITHECUS AND THEIR PHYLOGENETIC
RELATIONSHIP-MORPHOLOGY AND COMPARISON OF THE CRANIA. ACTA ANTHROPOLOGICA SINICA 2U): 1-10,
ABSTRACT According to concrete morphological comparison, Ramapithe cus and Sivapithecus could be considered the female and male of one species. They are more similar to the present-day orangutan than to either the gorilla or chimpanzee. As such, they can be seen as an ancestor of the orangutan. Accordingly, the orangutan split off from the hominoid branch leading to man and the African pongids approximately 12 million years B.P.

[Vol. 27, 1984 32 YEARBOOK OF PHYSICAL ANTHROPOLOGY
Foreword The most controversial question regarding human origins at present is the phylo-
genetic position of Ramapithecus, i.e., is Ramapithecus a hominid or not. Since the 1960s anthropologists have universally recognized Ramapithecus to be the earliest- known representative of the Hominidae. This is still the considered opinion of most anthropologists. In the last few years, however, especially the last 3 or 4 years, there have been increasing doubts raised by a number of workers.
Ramapithecus was first described in 1934 by G.E. Lewis (1934), based on a frag- mentary maxilla discovered in India. Since it had certain characteristics of man, it was suggested that the find could possibly be that of a hominid. This supposition did not, however, generate much scientific interest.
In the early 1960s, E.L. Simons once again raised the question of the hominid affinities of Rarnapithecus, In 1965 he and D.R. Pilbeam did a comprehensive investigation of the then-known Tertiary fossil apes and emphasized that Ramapi- thecus represented the earliest-known member of Hominidae. Moreover, they in- cluded a portion of the fossil ape material from Kenya and Kaiyuan, China, in the genus Ramapithecus. Since then, Ramapithecus has become more and more recog- nized by anthropologists as the earliest hominid. At present, ramapithecine-like fossils have been discovered in India, Pakistan, Turkey, Kenya, Greece, Hungary, and China. Their geochronological age is between 14 million and 8 million years B.P. Accordingly many think that mankind first separated from the evolutionary lineage of the apes before 14 million years B.P.
In the 1970s, however, with advances in molecular biology and the use of the molecular clock to estimate evolutionary divergence times, it became clear that the earliest that man and the apes could have separated was on the order of 5 to 6 million years B.P. This was in great contradiction to the fossil evidence as then interpreted. At the same time, Australopithecus afarensis, with an approximate age of 3.6 million years B.P. and having already evolved to the level of a hominid, was discovered in Africa. It, moreover, had morphological features a t variance with Ramapithecus. This discovery produced doubts among many anthropologists as to the ancestral hominid status of Ramapithecus.
In the past few years a t relevant international meetings, the phylogenetic position of Ramapithecus, vis-a-vis human origins, produced heated debate. As there was, however, insufficient physical fossil evidence to rely on, this question could not be settled.
Other than in China, the fossil material of Ramapithecus as yet discovered throughout the world consists of fragmentary mandibles and maxillas and isolated teeth. On December 1, 1980, a t the Shihuiba colliery site, Lufeng county, Yunnan province, a fossil skull of Ramapithecus was discovered. This raised the interest of anthropologists around the world. From 1975, when this fossil ape site was first discovered, we have conducted eight excavations, yielding a large number of ape fossils consisting of the following:
Ramapithecus: three crania, four mandibles, four cranial fragments, 16 man- dibular fragments, 20 upper and lower tooth rows, and 278 isolated teeth.
Siuapithecus: two crania, five mandibles, two cranial fragments, 20 upper and lower jaw fragments, seven tooth rows, and 290 isolated teeth.
Additionally, a small number of postcranial materials have been recovered. This article is a synthesis of important material evidence of the Ramapithecus and
Sivapithecus crania, mandibles, and teeth, makes an investigative comparison, and arrives at some new conclusions.
Cranial morphology and comparison There are five fossil ape crania discovered at Lufeng that are reasonably complete.
Of these, two represent Sivapithecus and three Ramapithecus. In addition, there are

Etler] FOSSIL HOMINOIDS OF LIJFENG 33
six fragmentary crania of these two types and a dozen or so maxillary fragments. The important craniofacial characteristics are as follows (Figs. 4-7):
Face-general features The cranium of the Lufeng Siuapithecus specimen (PA6441 is relatively well pre-
served. After an initial restoration, we were able to piece together the frontal and facial structure. It can, therefore, be considered the type cranial specimen of the species. It is an adult male. A notable feature in frontal view is its short, broad face with laterally flaring cheek bones. This is morphologically very similar to the condition seen in the male orangutan skull. In contrast, the face of both the chim- panzee and gorilla is relatively long and narrow, and the cheek bones do not flair as dramatically. The skull of 5'. indicus (GSP15000), discovered in Pakistan in 1979, is likewise that of an adult male (Pilbeam and Smith, 1981). Its face is particularly long and narrow and leaves a gracile impression. The PA644 skull, in contrast, is extremely robust.
Viewed laterally, the midfacial contour of PA644 is concave from the supraorbital ridges to the nasoalveolar clivus; the anterior region of the maxilla is, however, markedly raised as a result of strongly procumbent central incisor jugae. The face of the male orangutan is also concave in lateral view with procumbent central incisors, much as in the PA644 cranium. The supraorbital region of both the chim- panzee and gorilla is comparatively steep; the maxillofacial region turns abruptly downward and the face is inclined straight downward anteriorly. The anterior region of the maxilla is not raised prowlike as in the Lufeng specimen. The Pakistan skull in lateral view closely approximates PA644.
Of the three Lufeng Ramapithecus crania, PA677 is the most complete; unfortu- nately, it has been severely crushed and its facial dimensions are hard to determine.
Temporal ridges and sagittal crest The temporal ridges of PA644 are well formed. They arise from the lateral margins
of the supraorbital ridges and move mesially in a parallel fashion to the central orbital region and then turn strongly backward to form a median sagittal crest at
Fig. 4. Cranium of Siuapithecus from Lufeng (PA644). Frontal view.
34 YEARBOOK OF PHYSICAL ANTHROPOLOGY
Fig. 5. Cranium of Rumapithecus from Lufeng (PA677). Top view.
Fig. 6. Cranium of Siuupithecus from Lufeng (PA644). Left latera1 view.
pol. 27, 1984
Etler] FOSSIL HOMINOIDS OF LUFENG 35
Fig. 7. Cranium of Ramupithecus from Lufeng (PA677). Basal view
the vertex of the skull vault. The left and right temporal ridges convergence to form a large, flat trigonum frontale anteriorly.
The temporal ridges of the male orangutan also arise a t the outside corner of the supraorbitals conjoint with the supraorbital ridges. A thinly incised line separates the ridges, which run parallel to each other until near the central portion of the supraorbital margin, when they clearly separate. The temporal ridges bow backward and merge at the midsagittal plane to form an elongated, strongly projecting, yet thin, sagittal crest similar to the PA644 Sivupithecus cranium.
The temporal ridges of the male gorilla start from the robust posterosuperior margin of the zygomatic process of the frontal bone. Its starting point is a bit more medial than in the orangutan. What differs most between it, PA644, and the orangutan is that once they start, the temporal ridges separate from the supraorbi- tals and immediately begin to extend posteromedially to form a very prominent and comparatively thick sagittal crest. The point of convergence of the temporal ridges is noticeably anterior approximately in the lower one-third of the cranium; hence the trigonum frontale is markedly small. As the supraorbital ridges project strongly, the trigonum frontale of the gorilla is deeply depressed creating a supraorbital sulcus.
The temporal ridges of the male chimpanzee are relatively weak and tend toward the midsagittal plane. The sagittal crest, situated on the posterior portion of the skull vault is correspondingly weak. The temporal ridges originate in the same manner as in the gorilla, i.e., they very quickly separate from the supraorbital ridge

36 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
soon after they begin. The surface posterior to the supraorbital ridge is very concave but not to the extent seen in the gorilla.
The temporal ridges of the PA644 Sivapithecus skull are comparable to those seen in the three ape species, but are clearly most similar to the male orangutan.
Only the left side of the Pakistan Siuapithecus skull GSP15000 preserves the portion of the supraorbital margin where the temporal ridge begins. Its morphology largely corresponds with that of the male orangutan and PA644 skull.
The temporal ridges of Ramapithecus can be very clearly seen on the PA677 specimen. They are quite a bit weaker than in Sivapithecus and do not converge into a sagittal crest. They originate in the same manner as in Siuapithecus. They project a little a t first, but after separating from the supraorbital ridge they gradually begin to taper off within 2 or 3 cm. The ridges approach each other medially and slightly posteriorly to within 1.3 cm and then gradually diverge. This condition is also clearly seen in the YV 652 Ramapithecus skull. As the posterior part of the skull vault is well preserved in the latter specimen, the arch of the temporal ridge can be seen to undulate posteriorly and form a very minute sagittal crest.
The temporal ridges of the female orangutan and Ramapithecus skulls are very similar; neither has a sagittal crest.
The temporal ridges of the female gorilla differ from both Ramapithecus and other female apes in that they converge. After the temporal ridges start out from the lateral corners of the supraorbitals, they separate from the supraorbital ridge and quickly weaken and flatten out, converging slightly anteriorly and sagittally, and extending posteriorly as a low-lying, flat, broad sagittal crest straight to the occipital protuberance. The anterior portion of the trigonum frontale is extremely concave.
The temporal ridges and sagittal crest of the male chimpanzee have certain similarities with Ramapithecus (especially YV 652) but the anterofrontal portions of the temporal ridges are more curvilinear in the chimpanzee and the supraorbital sulcus is deeper. The temporal ridges of the female chimpanzee are very weak and widely separated, totally unlike the Lufeng specimen. Supraorbital ridge and glabella
A notable feature of the Lufeng PA644 Sivapithecus skull is that the left and right supraorbital ridges do not form a continuous torus a t glabella. Of particular note is the broadness and deep concavity of the glabellar region. The lateral supraorbital ridges of the male orangutan are also distinctly separate, but the glabellar region is only slightly concave and it is not nearly as broad. The supraorbital ridges of the male gorilla are joined together and form a robust and strongly projecting supraor- bital torus at glabella. Its glabella region is broad and large, and projects noticeably forward. The chimpanzee closely approximates the gorilla in these characteristics, but is not nearly as robust as the latter.
The Pakistan Sivapithecus skull GSP15000, like the PA644 specimen, has discon- tinuous supraorbital ridges, but its glabellar region is flat and narrow, very different than in PA644.
The morphology of the Ramapithecus PA677 skull supraorbital ridge and glabellar region is exactly like that seen in Sivapithecus PA644 except that it is much more gracile. These two characteristics are analogous to the condition in the male and female orangutan.
In both the female gorilla and chimpanzee, the supraorbital ridges join together, but due to the effects of sexual dimorphism they are weaker and more gracile, The glabellar region does, however, project forward in the female gorilla. Orbits and interorbital region
The orbits of the PA644 Sivapithecus specimen are subrounded in contour. The interorbital region is very wide and concave. The orbits of the orangutan are ovoid with the long axis of the orbit vertical (i.e., somewhat taller than wide). Its interor- bital region, however, is relatively narrow and flat, especially in the female. The orbits of the gorilla are rhomboid, broader than tall. Its interorbital region is also very broad but protuberant, especially the nasal bone. The internasal suture forms

Etler] FOSSIL HOMINOIDS OF LUFENG 37
a ridgelike crest which gradually widens inferiorly. The chimpanzee orbits are also rhomboidal, being even larger in breadth. Its interorbital region projects slightly.
The orbits of the Pakistan GSP15000 specimen are ovoid with height greater than breadth. Its interorbital region is very narrow. It is only 10 mm at its narrowest point, less than half the breadth of the PA644 Sivapithecus. In addition, its interor- bital region protrudes in a bow-shape, forming a thin ridge on the superior portion of the nasal bone. The inferior portion of the nasal bone is very flat.
The orbits of Ramapithecus and the condition of its interorbital region are as in Sivapithecus. The orbits of the two Lufeng specimens are closest to the Pakistan GSP15OOO and the orangutan in that they are all basically circular in shape. The orbits of the gorilla and chimpanzee can be classified as rectangular. In the mor- phology of the interorbital region, each has its own particular character and it is difficult to sort them out.
Infraorbital margin and position of the superior margin of the nasal aperture In the Lufeng Sivapithecus skull (PA644), the superior margin of the nasal aper-
ture is slightly higher than the infraorbital margin. This is also the case in the chimpanzee. In the orangutan it is somewhat lower and in the gorilla much lower. It is also particularly lower in the Pakistan GSP15000 specimen, quite a t variance to the Lufeng skulls.
The Lufeng PA677 Ramapithecus skull also has a superior margin of the nasal aperture that is slightly higher than the infraorbital margin. In this regard it approximates the condition seen in the female orangutan and chimpanzee, but differs substantially from what is seen in the GSP15000 specimen and the female gorilla.
Nasal aperture and nasoalveolar clivus The nasal aperture of PA644 is particularly narrow, long, and pear-shaped, being
widest in its lower quarter. The inferior lateral borders of the nasal aperture are bluntly rounded. The nasoalveolar clivus is foreshortened and concave with a slight anterior nasal spine. The shape of the orangutan nasal aperture and that of the Lufeng specimen are nearly identical, differing only in that the former is a bit wider. The nasoalveolar clivus of the orangutan is long and directed outward anteriorly, often with a weak anterior nasal spine. The nasal aperture of the gorilla is very broad and elliptical in shape. The inferior lateral margin is relatively sharp and gradually draws in medially to the distal side of the central incisor root. Its nasoal- veolar clivus is broad and short, and contracts inferiorly, without an anterior nasal spine. The nasal aperture of the chimpanzee is, comparatively speaking, the short- est, forming an abbreviated pear shape. The inferior lateral margin is bluntly rounded. Its nasoalveolar clivus is short and broad, flat and smooth, sloping anteriorly.
The Pakistan GSP15000 nasal aperture is also elongated, but relatively wider, being widest a t its base. Its inferior lateral border is blunt, and its nasoalveolar clivus is long and flat with a fairly obvious anterior nasal spine.
The nasal aperture and nasoalveolar clivus of the Lufeng Ramapithecus PA677 are as in Sivapithecus. In view of the above, the nasal morphology of the Lufeng specimens are most like that of the Pakistan Sivapithecus and the orangutan with clear differences from the gorilla and chimpanzee.
Canine jugae and canine fossa PA644 has noticeable canine jugae and very deep canine fossae. From the frontal
view, the canines diverge laterally downward to form an inverted “V”. These characteristics are strongly reminiscent of the orangutan and GSP15000 skulls and clearly distinct from the gorilla or chimpanzee. The canine jugae of the latter two are only slightly noticeable and the canine fossae are comparatively shallow. The angle of divergence of the canines is not as great, especially in the chimpanzee, where they are close to being parallel.

38 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
The canines of the Lufeng Ramapithecus are much smaller than in Sivapithecus but, as they had been lost before deposition and the canine jugae and canine fossae were crushed and distorted during fossilization, comparison is difficult.
Central incisor jugae and sulcus The PA644 Sivapithecus skull has pronounced central incisor jugae and a corre-
sponding deep central incisor sulcus, very close to the condition seen in the orangu- tan. In contrast, the central incisor jugae of the gorilla are not very prominent, although there is a t times a narrow intervening sulcus. The central incisor jugae and sulcus of the chimpanzee are somewhat more conspicuous.
The central incisor roots of GSP15000 are strongly curved. As a result the central incisor jugae are extremely well developed and the sulcus is very deep. As the lateral incisors are particularly small and bounded by strongly projecting central incisor and canine jugae, a very broad lateral incisor sulcus is created. Its superior portion passes through to the nasoalveolar clivus. This condition can also be seen in the orangutan and Lufeng crania but not to such an extent.
The anterior maxillary region of the Ramapithecus PA677 skull is crushed flat and distorted. This region in other skulls is not well preserved, so that it is difficult to observe and make comparisons.
Maxillary arcade The maxillary dental arcade of the Lufeng Siuapithecus skull is close to being
“U”-shaped. It is widest between the canines and diverges a bit posteriorly. This is seen most clearly in the PA845 skull. The arcade of the male orangutan is also “U”- shaped, with its widest dimension between the canines, but it draws in somewhat posteriorly. The arcades of the gorilla and chimpanzee are typically “U”-shaped. The chimpanzee, at times, is slightly constricted posteriorly. The GSP15000 arcade is likewise “U”-shaped with a very wide expanse between the canines. The extent of breadth between the canines in the arcades of the Lufeng, Pakistan, and orangu- tan specimens is related to the growth of the canines in an inverted “V” fashion.
The palate of Ramapithecus is sub “V” in shape, and its anterior portion is relatively rounded. The two tooth rows gradually diverge posteriorly. The arcades of the female orang, gorilla, and chimpanzee, however, are all “U”-shaped and M2 and M3 are drawn in medially.
Maxillary dentition A prominent feature of the Lufeng Sivapithecus central incisors is their particu-
larly large labiolingual diameter of the crown. The length-breadth indices of some teeth at times exceed 100, i.e., the labiolingual breadth exceeds the mesial-distal length. In present-day apes and the Pakistan specimen the labiolingual breadth of the central incisors is less than the mesiodistal length. The absolute mesial-distal length of the central incisor of the Lufeng Siuapithecus is a great deal smaller than seen in present-day apes or GSP15000. The central incisors of the orangutan, chim- panzee and GSP15000 are moderate in length, and that of the gorilla is the largest.
The root of the PA644 central incisor is strongly curved, giving the central incisor a tendency to be more vertical. This is most similar to the orangutan. The central incisors of the chimpanzee are more strongly projecting, this being even more evident in the gorilla. The central incisor root of GSP15000 is especially curved.
The lateral incisors are relatively small, only about one-half the size of the central incisors. This is most comparable to the orangutan. The lateral incisors of the gorilla and chimpanzee are clearly larger. The disparity in size between incisors is greatest in GSP15000. The Lufeng Ramapithecus incisors are morphologically identical to Sivapithecus, differing only in being smaller in size.
The canines of PA644 are relatively large in relation to the tooth row. The canines of present-day apes are all especially large, particularly in the gorilla. It can be said, in general, that the size and morphology of the Lufeng Sivapithecus canines are comparable to the orangutan and most similar to GSP15000.

Etler] FOSSIL HOMINOIDS OF LUFENG 39
The upper canines of the Lufeng Ramapithecus are very different morphologically from those of Sivapithecus. They are comparatively small and low. Compared to the extant apes, they are closest to the female orangutan, but the latter are compara- tively larger and taller.
As regards the cheek teeth (P's and M's), the occlusal surface enamel crenulations of the Lufeng Sivapithecus are fairly complex, cusps are moderately high, and enamel is comparatively thick. In sum, the dental morphology is most similar to the orangutan. The enamel ridges of the gorilla cheek teeth are most robust, their cusps high and sharp with developed cingulum, which is much different than the Lufeng specimen. In the chimpanzee, crenulations are complex but thin, the cusps are very low, and there is frequent evidence of lingual cingulum. This is quite unlike the Lufeng material. The cheek teeth of the Lufeng Ramapithecus are morphologically basically the same as Siuapithecus, differing only in size.
In summary, the Lufeng Sivapithecus skull in its full facial aspect, i.e., temporal ridges, supraorbital ridges, shape of orbits, nasal aperture, canine fossae, central incisor, and canine orientation and jugae, and in basic dental character is closest to the orangutan and GSP15000. It differs to a greater extent from the gorilla and chimpanzee. Similarly, the Lufeng Ramapithecus crania is relatively similar to the female orangutan in its morphological features and is more distinct from either the gorilla or chimpanzee. These similarities and differences reflect their phylogenetic relationships.
Discussion-Based on the preceding comparisons, three questions are addressed:
The relationship between Ramapithecus and Sivapithecus These two archaic apes were initially named on the basis of a few fossil fragments
discovered in India. Later fossils of the two types gradually accumulated. What is most striking is that these fossils, be they from India, Pakistan, Turkey, Kenya, Greece, Hungary, or China, are always found together a t the same localities and from the same deposits. Because of this, a number of scholars (Pilbeam, Andrews, etc.) have put forward, in the last few years, the position that, based on morphologi- cal similarities of the dentition and differences to be expected relative to individual size, the two could represent different sexes of a sexually dimorphic species. Green- field (1979) has most clearly pointed out that there is a great concordance between Ramapithecus and Sivapithecus, and that the former should be subsumed under the latter. This outlook, however, was not widely accepted until now, given the lack of sufficient evidence, especially cranial material.
In our morphological comparisons we can clearly see that Ramapithecus and Siuapithecus have the same basic facial characteristics. Their morphological differ- ences, be they in size, temporal ridge, sagittal crest, or canines, are like those between the male and female orangutan. This expresses the fact that Ramapithecus and Sivapithecus could be the female and male, respectively, of a single sexually dimorphic species.
It is notable that the size difference between Ramapithecus and Siuapithecus, be they in the skull, mandible, or tooth size, are very great. According to Napier and Napier (1976), however, the extant female orangutan is estimated to be only 47% the size of the male. Therefore, the difference in size between the Lufeng specimens can be understood as a difference in size between male and female.
If the differences between Ramapithecus and Sivapithecus are merely sexual in character, they cannot be differentiated by name. As Sivapithecus has priority over Ramapithecus, according to the rules of international nomenclature, Ramapithecus would be subsumed into the former and be revised as Siuapithecus.
Systematic position of Sivapithecus For a long time Sivapithecus was viewed as a species of dryopithecine ape consid-
ered ancestral to the extant pongids. In the last few years, however, a number of people have recognized Sivapithecus to possess certain traits, such as thick tooth

40 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
enamel and megadont molars, similar to A ustralopithecus. They, therefore, advo- cated that Sivapithecus be seen as a hominid ancestor.
Most recently, Pilbeam (1982), in further investigating the Pakistan S. indicus skull GSP15000, pointed out many similarities between it and the orangutan which possibly originated as shared, derived characters. Although Pilbeam still doubts that S. indicus is itself the ancestor of the orangutan, he considers it very likely that a similar late Miocene Asian species could be. Andrews (1982) forcefully advocates including Rarnapithecus with Siuapithecus. He suggests that Sivapithecus and the orangutan are closely related and have a much more intimate relationship with each other than either has with man, the chimpanzee, or gorilla.
Is Sivapithecus, therefore, really the ancestor of the orangutan? After going over the above systematic comparisons of craniofacial morphology it can be seen that the Lufeng Sivapithecus (including Ramapithecus) crania are comparatively most simi- lar to the extant orangutan and relatively distinct from the extant chimpanzee and gorilla. At the same time, the Lufeng specimens are craniofacially very similar to the GSP15000 Sivapithecus skull from Pakistan. In conclusion, the Lufeng and Pakastani crania and the orangutan have the following important shared characters:
(1) The facial contour is concave and the premaxillary region is upturned. (2) The temporal ridges start out conjoint with and run parallel to the supraorbital
(3) The left and right supraorbital ridges do not meet to form a continuous supraor-
(4) Orbits are circular. (5) The nasal aperture is long and narrow. (6) The canine pattern of orientation is into an inverted “V” shape. (7) The upper central incisor roots are strongly curved, with clear alveolar jugae. (8) Molar cusps are moderately high, the occlusal surface enamel is relatively
These shared characters are clearly distinct from the gorilla and chimpanzee. They strongly suggest that the Lufeng and Pakistan Sivapithecus and the extant orangutan have a close phylogenetic relationship and are more distantly removed from the gorilla and chimpanzee. Therefore, it is apparent that Sivapithecus should be viewed as an ancestor of the present-day orangutan. The time of divergence of Sivapithecus from the hominoid lineage leading to the African apes and man
The deposits from which the Lufeng ape fossils have been discovered are geochron- ologically from the late Miocene; their estimated age is 8 million years B.P. The fossil ape teeth from the Xiaolongan colliery site, Kaiyuan, Yunnan, were deposited a bit earlier. The Pakistan and Indian material comes from deposits that are 12 to 8 million years old. Other regions where fossils of this type have been discovered are all between 14 and 8 million years B.P. At present, only the specimens from China and Pakistan can be thoroughly compared. The material from Africa and Europe is still too limited, and the morphology of the skulls is still not well understood. In order to estimate the time when the ancestor of the orangutan diverged from the hominoid lineage, we must at present rely on the material from China and Pakistan. The branch of the orangutan seems then to be divergent from the main stock of the hominoids a t approximately 12 million years B.P.
ridges.
bital torus.
thick, crenulations are comparatively complex, and buccal cingulum is lacking.
&I GUOQIN (1983) DESCRIPTION OF CARNIVORA FOSSILS FROM LUFENG. ACTA ANTHROPOLOGICA SINICA 2(1): 11-21.
ABSTRACT Since 1975 the Sivapithecus (Rarnapithecus) fossil site at Lufeng has undergone extensive excavations. Besides yielding a great many hominoid and primate fossils these late Miocene lignite beds have also pro- duced abundant genera and species of other mammals. Descriptions of fossil Carnivora, including Proputorius lufengensis and Epirnachairodus fires dis- covered between 1975 and 1976, are given.

Etler] FOSSIL HOMINOIDS OF LUFENG 41
Fossil descriptions
Carnivora Bowdich 1921 Canoidea Simpson 1931 Mustelidae Swainson 1935 Proputorius Filhol 1890 Proputorius lufengensis sp. nov.
Holotype-Left mandibular corpus with P3-M1 and socket for P2 and M2 (V6885.1). Paratype-Fragment of left mandibular corpus with MI and M2 alveolus (V6885.2).
Diagnosis The mandible is lightly constructed, lacks PI, P2 is single rooted, P4 has a small
accessory cusp posteriorly on the outer edge of the talonid. M1 has metaconid, P2-P4 tooth row length greater than M1 length.
Description Gracile mandible; two mental foramina are situated anterior to P3 and P4 on the
horizontal corpus; the symphysis and vertical ramus are broken off and lost. Mandi- bular height at the anterior margin of M1 is 6 mm. I and C missing; lacks PI; P2 is missing but the alveolus shows it to be single rooted; P3 is basically conical with a steep anterior margin, the posterior margin recedes, it is double rooted, and the anterior surface develops clear cingulum. P4 is larger than P3 but basically of the same form; a small but obvious metastylid develops posterior and lateral to the primary cusp, double rooted. MI has a clear metaconid; the talonid is hollowed out but the hypoconid is shearing, the breadth less than that of the trigonid. M2 is missing but the alveolus demonstrates it to be smaller than MI. Moreover, it is single rooted. P2-P4 tooth row length (10.5 mm) is greater than MI length (7.4 mm).
Discussion The two Lufeng mustelid specimens, in terms of size and M1 having a metaconid,
indicate that they undoubtedly belong to the martin subfamily. In China five genera of fossil martins have so far been discovered. Among these only Charronia is restricted to the Pleistocene. The other four genera are spread throughout Miocene and Pliocene deposits. Only one species of Sinictis has been discovered, S. dolichog- nathus. It is a relatively large type and retains PI. Five species of Martes have been recovered from Plio-Pleistocene deposits. These are M. anderssoni, M. palaeosinensis, M. crassa, and M. pachygnatha; all are quite a bit larger than the V6885.1 specimen; and M. zdanskyi, which is relatively smaller but retains PI. Vormela is represented by the small V prisca. This diminutive type lacks a PI; P4 has a metastylid; M1 talonid basin and P2-P4 arcade length are, however, much smaller than in V6885.1 (Table 3). The above analysis indicates that V6885.1 and 2 must be placed outside of Sinictis, Martes, and Vormela. In size and other important characteristics it con- forms to Proputorius and should be so assigned. Two species of this genus have already been discovered in China: I! minimus from Baode, Shanxi, and F! yaogouen- sis from Zhunge’er district, Inner Mongolia. In the former, the lower premolar and molar size approximates V6885.1, but P2-P4 tooth row length is comparatively greater, P2 is double rooted, and P4 lacks a metastylid. In comparison to P yaogouen- sis the V6885.1 mandibular height is less; it retains only two mental foramena, the lower premolars are relatively more developed, tooth row length is relatively greater, and M1 is comparatively smaller. The French species I? sansaniensis is, in compari- son, closer to F! yaogouensis than V6885.1. Considering the above morphological differences and taking into consideration aspects of geographical distribution, it is felt that V6885.1 and 2 should be designated a new species.

42 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
2
Left mandible with P3-Ml of Proputorius Zufengensis sp. nov from Lufeng V6885.1 X 3. 1. labial face; 2. occlusal face; 3. lingual face
TABLE 3.
Tooth dimensions (mm)
I! ninimus P: yaogouensis I! sansaniensis V. prisca Lu feng DeRijin Qiu Zhuding Filhol DeRijin
V6885.1 (1945) (1979) (1891) (1945)
p3 L' 3.7 3.8 W' 2.1 2.2
3.1 1.8
p4 L 4.8 4.7 3.3 W 2.3 2.4 2.6
4.0 2.0
5.0 3.0
MI L 7.4 1.5 8.7 10.0 W 3.4 3.1 3.4 4.0
P,-P,L 10.5 11.8 7.8
4.0
7.0 2.5
M1 6.0 6.0 7.6
'L, length; W, wldth.
....m
Sivaonyx Lydekker 1884 Sivaonyx bathygnathus Lydekker
Material
ment preserving part of (V6886.3); isolated left
Left anterior maxillary fragment with I2 and I3 (V6886.1); right maxillary frag- and P3 (V6886.2); right maxilla preserving P-M1
and M1 (V6886.4,5) and two damaged right M's (V6886.6,7);

Etler] FOSSIL HOMINOIDS OF LUFENG 43
right mandible with C-P4 (V6886.8); left mandible containing P3, P4, and part of MI cV6886.9); one isolated left P4 fv6886.10); one damaged left M1 (V6886.11); and a complete right MI (V6886.11); and a complete right M1 (V6886.12). Description
1' diminutive, I3 is somewhat larger, and both are single rooted. Upper canine is missing but the preserved alveolus indicates it was relatively large. Pz and P3 are, without exception, degenerate, single cusped, and double rooted; P3 does, however, retain a small degree of cingulum on its posterior surface.
is squared with an external shear blade, and the outside cusps are taller than the inside cusps (i.e., paracone taller than metacone and protocone taller than hypocone). There is a developed paraconule and clear internal and external cingulum.
M1 is broad interiorly but narrow exteriorly, and is wider than long; the paracone and metacone are both relatively low and lophodont. M1 has a metastyle and a developed cingulum blade at the location of the hypocone. The whole tooth has pronounced anterior lingual cingulum.
The lower canine is fairly long and sharply cusped P1 is lacking; Pz and P 3 retain only a primary cusp. Both narrow anteriorly and broaden posteriorly, and are double rooted and encircled by cingulum.
P4, likewise, narrows anteriorly and broadens posteriorly. Moreover, the outer posterior surface has a strongly developed metastylid and a relatively broad talonid. The base of the crown is girdled by a pronounced cingulum.
M1 is broad with a paraconid positioned longitudinally 0.67 cm along the central axis. The metaconid and paraconid are moderately tall. The talonid is broader than the trigonid, but of similar length. The talonid is basin-shaped and encircled by small cusps. The hypoconid is fairly tall, a small cuspulid develops posterior to the protoconid, and there is pronounced cingulum anteriorly and on the outside surface (Table 4). Discussion
Sivaonyx was a large-bodied otter. It was named by Lydekker in 1884 based on specimens recovered from Dhok Pathan deposits in the Siwaliks. Later a closely related type, S. hessica, was discovered in Neogene deposits of Europe and Africa
Tooth dimensions (mm) S. bathygnathus
No. 19509 Lufeng Amer. Mus,
V6886.2 V6886.3 V6886.4 V6886.5 V6886.8 V6886.9 V6886.10 V6886.12 Colbert, 1935
P3 L 7.3 W 5.7
P4 L 13.0 12.2 13.7 W 13.8 13.5 13.3
w 14.0 14.6 C L 6.4
5.0 $1 16.7
W 3.6
W 4.0 4.1
W 6.0 6.0 6.5 M' L 15.9 16.5
w 9.0 9.5
M' L 9.3 9.5
P2 L 4.3
P3 L 5.6 5.3
P L 8.8 8.4 8.8
'H, height.

44 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
(Eppelsheim). It differs from the Siwalik specimen in having a comparatively shal- low mandible and narrow M1 and P4 posterior root. S. hessica material is, however, very scarce and lacks reliability by itself in establishin a species. The Lufeng
and P4, greatly expanding the scope of Siuaonyx material. The Lufeng specimens (especially P4 and MI), in both size and morphology, correspond well with S. bath- ygnathus of the Siwaliks. The two are basically the same, differing only in the Lufeng specimen having a somewhat shallower mandible. Mandibular depth at M1, and M1 length, are basically the same.
specimens constitute not only P4 and MI, but also 12, 13, P 5 , p, M1, and C, Pa, P3,
Lutra Erxleben 1777 Lutra sp.
The specimen consists of a left mandible preserving P4-M1 (V6887). The primary cusp of P4 is damaged and all that can be seen is that there was developed posterior cingulum. The paraconid on M1 trigonid is low, comparatively blunt, and centrally positioned. The protoconid is tall, the talonid basin-shaped, and about two-fifths the length of the trigonid. This specimen has certain similarities to the Indian specimen discovered in 1932 by Pilgrim and described as Lutrine genus indet. Furtivus sp. nov. These include: small size, robust mandible which is deeper than MI is long, M1 relatively short with greatest width medially, relatively short talonid about one-half the length of trigonid, and P4 somewhat broad posteriorly with a fairly obvious metastylid and posterior cingulum. The Lufeng specimen has a deeper mandible, the metaconid in the trigonid center is fairly small, and M1 is larger than in the Siwalik specimen (Table 5).
eeeee
Feloidea Simpson 1931 Hyaedae Gray 1869 Ictitherium Wagner 1848 Ictitherium gaudryi Zdansky
Material Four isolated left p s (V6888. 1-41; among these the exteroposterior surfaces of
V6888.1-3 roots are lost; the entire root of V6888.4 is lost. Five isolated right P% (V6888.5-9), exteroposterior surfaces of V6888.6,7 roots are lost; the third blade of V6888.8 is lost. On one isolated right P3 (V6888.101, the hypoconulid and anterior face of the root are lost. The root of one isolated right M1 (V6888.11) is completely lost. Two left mandibles preserving P2-M1 (V6888.12,13), I, C, and Ma, and vertical ramus are lost. One right mandible preserving P2-M1 (V6888.141, I, C, Ma, and vertical ramus is lost.
Description p4 protocone is large and elongated mesiobucally; the second and third blades are
relatively sharp and inclined posteriorly. p4 has pronounced lingual cingulum and is triple rooted. The mandible is relatively deep and thick. Tooth size is moderate and cusps are low and blunt. Anterior cusps of P2 and P3 are very weak or lacking,
TABLE 5.
Tooth dimensions (mm) Lufena Suecimen V6887
L 8.0 12.8 W 4.2 10.5

Etler] FOSSIL HOMINOIDS OF LUFENG 45
but the posterior cusp is noticeable. The inner margin of the posterior cusp has a cingula-derived talonid. M1 trigonid length is approximately two times the talonid length. The M1 paraconid is long and carnassial; the protoconid is high and before abrasion carnassial. The metaconid is small, as high or somewhat lower than the paraconid; the talonid broadens into a basin; the endoconid is taller than the hypoconid. In unabraded specimens a hypoconulid can be seen. The alveolus of P1 and M2 indicate that they were single rooted. Discussion
In size and morphology the Lufeng specimens are clearly distinct from the Siwalik species I. siualensis and I. indicum. In comparison to I. siualensis, the Lufeng mandibles are rather deep (mandibular depth at MI is 25-26 mm) and the teeth rather small (MI length 16-18 mm). Crowns lack robust enamel wrinkles and cingulum is not as developed. The Lufeng P4 length is much less than MI, and P4 is without a noticeable paraconid. M1 total length is relatively small, but the talonid is comparatively enlarged. In comparison to I. indicum the Lufeng specimens are smaller in body size, have mandibles that are not as deep, and lower premolar and molar cusps not as tall. I . indicum P4 parastylid is more pronounced than in the Lufeng material, M1 is longer in length, and the talonid is comparatively short, just one-fourth the total length of M1. In comparison to European species, the Lufeng specimens are typologically closest to I . robustus, but the I. robustus M1 is compara- tively long, its talonid longer, and its metaconid quite a bit lower. Four species of Ictitherium have been described from north China: I. gaudryi, I. sinensis, I. wongi, and I. hyaenoides. Of these I. sinensis is based on a possibly distorted specimen and its species-designation is questionable (Qiu Zhanxiang et al., 1979). I. hyaenoides is obviously much larger than the Lufeng specimens and has been designated a sub- species of I. hipparionum by Qiu. I. wongi, although similar in build to I. hyaenoides and close to the Lufeng material, has a comparatively shallow mandible, and P4 and lower dental cusps that are comparatively much higher and sharper. The total morphological pattern and dimensions of the Lufeng mandibles, @, and lower dentition, however, all demonstrate clear affinities to I. gaudryi (Table 6). .....
Felidae Gray 1821 Epimachairodus Kretzoi 1929 Epimachairodus fires sp. nov.
Holotype A pair of upper canines (V6889.1,2), possibly from the same individual; the greater
part of the left canine crown is lost; left lower canine (V6889.3); left mandibular fragment preserving P3 and P4 and right mandibular fragment preserving masseter fossa, possibly from same individual (V6889.4,5). Diagnosis
The upper canine is especially huge and robust, with denticulations on anterior and posterior borders. The lower canine is small but fairly robust with denticulations on the lingual aspect of the crown. P3 is degenerate with both an anterior and posterior stylid. P4 has two noticeable posterior stylids. Cusps of P3 and P4 are slanted posteriorly with denticulate edges to the blades. Description
The upper canine is extremely long, broad and thick, curved posteriorly, with sharp-edged anterior and posterior margins, and clear denticulations along the entire edge. The lower canine is degenerate, short, and relatively small, but robust and conical. The two sharp edges of the lingual aspect of the crown have obvious denticulations. P3 is degenerate; the primary cusp slants posteriorly. It has a rela- tively small parastylid and metastylid which both slant anteriorly; it is double rooted and has pronounced lingual and buccal cingulum. P4 is bigger than P3 but of
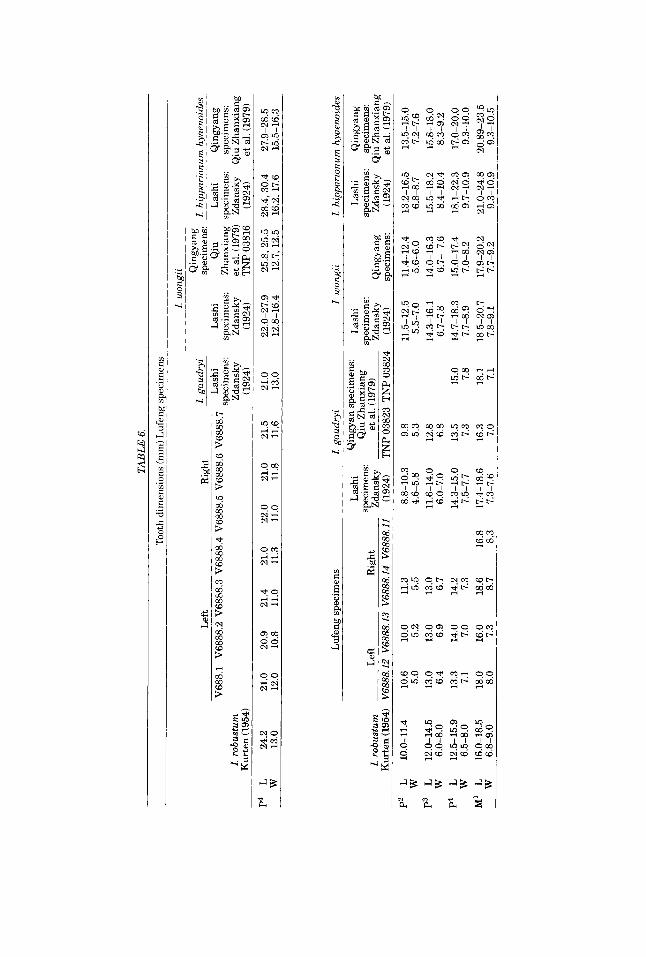
TAB
LE 6
.
Too
th d
imen
sion
s (m
m) L
ufen
g sp
ecim
ens
I. w
onjz
ii Qin
gyan
g sp
ecim
ens:
I.
hipp
ario
num
hya
enoi
des
I. ga
udry
i L
eft
Rig
ht
V68
8.1
V68
88.2
V68
88.3
V68
88.4
V68
88.5
V68
88.6
V68
88.7
sp
zE
ns:
sp
ecim
ens:
L
ashi
Z
hanx
iang
Q
iu
spec
imen
s:
Las
hi
spec
imen
s:
Qin
gyan
g
Kur
ten
(195
4)
(192
4)
(192
4)
TN
P 0
3816
(1
924)
et
al.
(197
9)
I. ro
bust
urn
Zda
nsky
Z
dans
ky
et a
l. (1
979)
Z
dans
ky
Qiu
Zha
nxia
ng
p4
L 24
.2
21.0
20
.9
21.4
21
.0
22.0
21
.0
21.5
21
.0
22.0
-27.
9 25
.8, 2
5.5
28.4
, 30.
4 27
.9-2
8.5
W
13.0
12
.0
10.8
11
.0
11.3
11
.0
11.8
11
.6
13.0
12
.8-1
6.4
12.7
, 12.
5 16
.2, 1
7.6
15.5
-16.
3
I. w
ongi
i L
ufen
g sp
ecim
ens
I. ga
udry
i L
ashi
Q
ingy
an s
peci
men
s:
Las
hi
spec
imen
s:
Qiu
Zha
nxia
ng
spec
imen
s:
Zda
nsky
et
al.
(197
9)
Zda
nsky
Q
ingy
ang
Kur
ten
(195
4)
V68
88.1
2 V
6888
.13
V68
88.1
4 V
6888
.11
(192
4)
TN
P 0
3823
TN
P 0
3824
(1
924)
sp
ecim
ens:
L
eft
Rig
ht
I. ro
bust
urn
P2
L
10.0
-11.
4 10
.6
10.0
11
.3
8.8-
10.3
9.
8 11
.5-1
2.5
11.4
-12.
4 W
5.
0 5.
2 5.
5 4.
6-5.
8 5.
3 5.
5-7.
0 5.
6-6.
0 L
12.0
-14.
5 13
.0
13.0
13
.0
11.6
-14.
0 12
.8
14.3
-16.
1 14
.0-1
6.3
W
6.0-
8.0
6.4
6.9
6.7
6.0-
7.0
6.8
6.7-
7.8
6.7-
7.
6 p4
L
12.5
-15.
9 13
.3
14.0
14
.2
14.3
-15.
0 13
.5
15.0
14
.7-1
8.3
15.0
-17.
4 W
6.
5-8.
0 7.
1 7.
0 7.
3 7.
5-7.
7 7.
3 7.
8 7.
7-8.
9 7.
0-8.
2 M
' L
16.0
-18.
5 18
.0
16.0
18
.6
16.8
17
.4-1
8.6
16.3
18
.1
18.5
-20.
7 17
.9-2
0.2
W
6.8-
9.0
8.0
7.3
8.7
8.3
7.3-
7.6
7.0
7.1
7.8-
9.1
7.7-
9.2
I. hi
ppar
ionu
m h
yaen
oide
s
Las
hi
Qin
gyan
g sp
ecim
ens:
sp
ecim
ens:
Z
dans
ky
Qiu
Zha
nxia
ng
(192
4)
et a
l. (1
979)
13.2
-16.
5 13
.5-1
5.0
6.8-
8.7
7.2-
7.6
15.5
-18.
2 15
.8-1
8.0
8.4-
10.4
8.
3-9.
2 18
.1-2
2.3
17.0
-20.
0 9.
7-10
.9
9.3-
10.0
21
.0-2
4.8
20.8
9-23
.5
9.3-
10.9
9.
3-10
.5

Etler] FOSSIL HOMINOIDS OF LUFENG 47
similar form with an obvious parastylid, two metastylids, and developed internal and external cingulum. Both P3 and P4 have denticulations on the blade edge of the primary cusp (protoconid), parastylid, and metastylid. The crown surfaces have minute enamel crenulations. Discussion
The Lufeng sabre-toothed cat is represented only by upper and lower canines, P3, Pq, and some mandibular fragments. The skull, mandibular symphysis, and mandi- bular joint, all of which are important diagnostically, are not known, so it is difficult to make a full and complete comparison with contemporary Chinese and foreign genera and species. An initial observation of the size and character of the upper canines, however, shows them to belong to an undoubted species of large-bodied sabre-toothed cat with affinities to Machairodus aphanistus from European Eppels- heim Pikermi deposits and Epimachairodus palanderi from northern China. For example, the Lufeng specimen upper canines are very large, robust, and elongated, with sharp edges both anteriorly and posteriorly, the whole edge having clear denticulations; the lower premolar cusps slant posteriorly. These traits are very similar to M. aphanistus, but the upper canines of M. aphanistus are more strongly denticulated both anteriorly and posteriorly; P3 is more degenerate, having only a strong parastylid but lacking a metastylid. The Lufeng specimen P3, while degener- ate, is unlike M. aphanistus in having both a parastylid and metastylid.
The Lufeng specimen has long, broad upper canines, degenerate P3, and P3 and P4 blade margins with noticeable denticulations. These traits are similar to those seen in E. palanderi. The upper canines of E. palanderi, however, while having denticu- lations on the anterior and posterior borders of the tooth crowns have an increas- ingly broad basal portion of the anterior edge of the crown on which denticulations do not appear. The basal portion of the anterior crown of the Lufeng specimen still has denticulations which are, moreover, separated by inverted “Y”s (serrated) yield- ing two rows of comparatively minute denticulations. The lower canine size is not much different in E. palanderi, but the Lufeng upper canines are much larger than
A) Right upper canine of Epimachairodus fires sp.nov. V6889. 1 X %. Left side, lingual face; Right side, labial face. B) Left lower canine of Epimachairodus fires sp. nov. V6889. 3 x 1. Left side, lingual face; Right side, labial face.

48 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
those of E. palanderi. The biggest difference is that the Lufeng specimen’s P 3 have a relatively obvious parastylid and metastylid. Considering the above, we conclude the Lufeng specimen is a new species of Epimachairodus (Table 7).
..... Pseudaeilurus Gervais 1848-1852 Pseudaeilurus sp.
One right P 3 (V6890.1) with primary cusp somewhat inclined posteriorly, no parastylid, notable metastylid, developed internal cingulum, minute enamel crenu- lations on tooth crown surface, crown length 15 mm, breadth 8.8 mm. One left MI (V6890.21, posterior blade lost, preserves mesiodistal blade, without talonid, anterior face of one tooth root preserved, crown length 20 mm, breadth 8.4 mm. These two dental specimens differ from species of the north China Hipparion fauna and Inner Mongolia mainly in size. Three species of Pseudaeilurus have been discovered in Chinese Hipparion fauna. One, I? major, is from Yushi, Shanxi. Another P minor has been found in Shanxi, Gansu, and Henan. The Pgs of both of these species have noticeable metastylid and lingual fovea; M1 at times has metaconid. P major and I? minor differ only in size; the latter has recently been sunk into €? parvulus (Table 8).
One other species is from Tong gu’er, Inner Mongolia, I? mongoliensis. Its lower premolars have parastylid, degenerate MI talonid, and it lacks a metaconid. As the Lufeng specimens can only be evaluated based on the size of P3 and MI, it is closest to I? major. The sparsity of material, however, leaves it best unassigned at the species level.
TABLE 7.
Tooth dimensions (mm) M. aphanistus E. palanderi
Roth & Wagner Wagner Gaudrv Shanxi,
Lufeng (1885) (1860) (1873) Zdansky Baode V6889.1 V6889.2 V6889.3 V6889.4 Pikermi Pikermi Mt. Leberon (1924) (1957)
- C L 42.0 43.0
w 19.0 H 122.0
- C L
W 14.0 11.4
35.0 14.0
14.0 15.0
38.0-45.0 36.0 13.5-16.5 13.0
105.0
15.0 14.3 10.3
H P3 L 15.0 20.0 19.0 15.1
W 8.6 7.4 P L 28.6 27.0 27.0 25.7
W 11.4 10.4
TABLE 8.
Tooth dimensions (mm) Lufeng l? major l? ct: l? paruulus l? mongoliensis
major Zhang Xiti
and Zdansky Teiln Liv Houyi Zdansky Kpokoc Tenius Colbert
V6890.1 V6890.2 (1924) (1945) (1964) (1924) (1939) (1951) (1939)
P3 L 15.0 15.5 14.0 12.5 9.9 11.2 10.3 12.0 w 8 8 8.4 8.0 6.0 5.3 4.5-5.7 5.1 5.7 _ _ ..
M’ L 20.0 23.2 24.0 21.5 18.1 17.8-19.2 17.2-17.8 17.3 W 8.4 10.1 10.0 10.0 7.2 7.0- 7.5 7.0- 7.5 7.2

Etler] FOSSIL HOMINOIDS OF LUFENG
HAN DEFEN (1983) A NEW SPECIES OF LOPHOCHOERUS FROM LUFENG. ACTA ANTHROPOLOGICA SINICA 2(1):22-25.
49
ABSTRACT A description and comparative analysis of Lophochoerus dis- covered at the Lufeng fossil ape site in Yunnan Province during the winter of 1981 is presented. The premolars and molars of the Lufeng Lophochoerus have definite differences in comparison with the several species of the genus already known from the Siwaliks. Because of this a new species, Lophoche erus lufengensis sp. nov., has been established which represents the first discovery of this particular fossil type from late Miocene strata of China.
Specimens of Lophochoerus were first discovered in the Siwaliks of India and Pakistan. Pilgrim’s (1926) analysis included three species: L himalayensis, L exguus, and L. nagrii. The material, however, was limited to only a couple of fragmentary mandibles and a few isolated cheek teeth. Matthew (1928) summarized the charac- teristics of the genus as follows: a diminutive animal with simple cusps, little or no extra cuspules, and a slight tendency to transverse cresting. The Lufeng Lophoche erus fossil was collected in 1981 from the sixth stratum of the excavated section (thin lignite stratum). Previously, this type of fossil had been recorded in China from undetermined provenance. Koenigswald (1963) described two Lophochoerus teeth which had been obtained from Chinese apothecaries in Hong Kong. These included a M3 in which the front row of cusps had already joined together to form a crest. The back row of cusps, however, had not formed a true transverse crest. Koenigswald, accordingly, bestowed a new species on the specimen, L. pilgrim, which he thought to possibly be a member of the Chinese Hipparion fauna. Although the L. lufengensis sp. nov. consists of only a fragmentary mandible, its lower premolars are well preserved and have distinctive features. The transverse crests of the molars are relatively well expressed and the specimen was produced from a definite strati- graphic location. For these reasons, the discovery of this fossil from the Lufeng fossil ape site not only helps to elucidate the geographic distribution of this animal at the time in question, but also has definite implications for comparing the Lufeng fauna with those of other contemporaneous sites. With this in mind, the following descrip- tion and discussion are given.
Description of the specimen
Artiodactyla Owen, 1868 Suina Gray, 1868 Suidae Gray, 1821 Lophochoerus Pilgrim, 1926 Lophochoerus lufengensis sp. nov.
Holotype -Two pieces of a fragmentary mandible; left mandible possesses P1-P4, Mz, and M3; right mandible possesses PI, right lower canine (V6891). Analysis
The posterior crests of P3 and P4 divide into inner and outer branches. The median valley which forms between them is blocked off by the talonid. The inner cusps of M3 are close to being conical. The outer cusps taper posteriorly and are “V”-shaped.
The anterior arm of each of the outer cusps extends forwardly, differentiating into an anterior cingulum and a central part of the forward row of cusps which form a transverse crest. The talonid, likewise, possesses a forwardly directed crest which reaches to the middle part of the transverse crest formed by the posterior row of cusps. The enamel layer is smooth and without wrinkles. Description
The fragmentary mandible (V6891) was discovered in the sixth stratum of the Lufeng fossil ape site lignite beds (see Wu et al., 1981, for cross-sectional figure). The
50 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
Top: Labial and occlusal views of the left mandible of L. lufengensis sp. nov. Bottom: L. lufengensis sp. nov.
Lingual view of
Internal surface and external surface view of the right lower canine of L. lufengensis sp. nov

Etler] FOSSIL HOMINOIDS OF LUFENG 51
left side of the mandible preserves a part of the mandibular corpus and ascending ramus. The mandible had been crushed flat and its thickness cannot be measured. Its height posterior to M3 is 27.3 mm. The teeth are small. Part of the crown surface of the left and right sides of P1 has been damaged. P1 lacks both cingulum and anterior and posterior cuspules. Pz and P3 are unworn. The primary cusp of Pz is quite tall and comparatively sharp, with anterior and posterior ridges that are downwardly inclined and without cuspules. Toward the inside, anteroinferior side, there is an anterior cingulum cusp. In general P3 is morphologically similar to Pa. The lower portion of the posterior crest of the primary cusp is approximately two- thirds the height of the tooth crown. It divides into inner and outer branches. The talonid cusps are small, only about one-third the height of the tooth. They become pointed anteriorly and extend to the depression formed by the two posterior branches of the primary cusp. P4 has been abraded and is relatively wider than Ps. It has well-developed anterior cingulum and its posterior construction is similar to P3. The outer branch posterior to the worn surface links up with talonid, and the inner branch stops at the anteroinferior edge of the talonid. P1-P3 all have two roots; P4 has an undivided root. The anterior and posterior faces of P1-P3 and the anterior face of P4 all lack contact facets. M1 is missing. The two outer cusps of M2 are broken and the tooth has been severely worn. Its anterior cingulum is well expressed and the lingual cusps are conical in shape. M3 is well preserved and only lightly worn. The anterior cingulum is positioned at a little less than one-half the tooth height. The lingual cusps are conical and separated from each other, and their inner walls are steep and straight. There is a wide basin and no cuspules. Buccal cusps are worn posteriorly and taper into a “V” shape. The anterior arm of the protoconid is developed and extends inwardly and forwardly to the middle of the anterior cin- gulum. The posterior arm is weak and links up with the metaconid to form the first transverse crest. The anterior arm of the hypoconid extends to the middle of the first transverse crest. The second transverse crest is analogous to the first, except that it is somewhat narrower in width. As the anterior arm of the hypoconid links with the first transverse crest, the basin between the two buccal cusps remains unopened. The talonid is relatively small and slightly lower than the anterior face of the primary cusp. Its upper portion is very lightly worn and the anterior aspect has developed into a ridge passing through the basin to the middle of the second transverse crest. The anterior aspect of the lingual portion of the talonid has an obvious cuspule, blocking off the basin between the entoconid and the talonid. The upper portion of the four primary cusps of M3 are clearly higher than the crests formed by the cusps.
The right lower canine crown is completely preserved. The upper portion has, however, been worn to a slant. The inner and outer faces are covered with enamel. The outer distal side has a small concave groove. The posterior surface enamel is lost and it is somewhat narrower than the outer surface. The root is fragmentary (Table 9).
Comparison and discussion Pilgrim (1926) created the genus Lophochoerus based on some of the small-sized
suid fossils from the Siwaliks of India and Pakistan. The material of this genus was
TABLE 9.
Tooth dimensions (mm) Measurements PI Pz p3 p4 Mz M3 C1
Length 6.4 10.8 11.8 11.8 13.0 16.4 outer width 6.6 inner width 9.3
Width 3.9 4.6 5.8 6.8 9.2 9.5 posterior width 6.1 crown height 31.4’
‘Measured from crown base. 2Measured from lingual surface

[Vol. 27, 1984 52 YEARBOOK OF PHYSICAL ANTHROPOLOGY
divided into three species: L. himalayensis (a right mandible possessing P4-M2 from Nagri zone), L. exguus (a posterior lower molar, from Chinji zone), and L. nagrii (a left mandible of a juvenile, from Nagri zone). According to the description of the author, the important distinguishing characteristics of the genus are as follows: the cusps of M1 are semicircular in structure, unlike other suid molars from India; the outside of the inner cusps and the inside of the outer cusps form a somewhat flat plane; each blade of the cusps is linked up with its opposite; the two blades are separated by a transverse valley; the cusps are high and rise above the crest which link them, and in this it is similar to Listriodon. A low blunt crest passes through the transverse valley from the posteroexterior cusp to the anterointerior cusp. Tooth structure is very simple. There are no cuspules, tubercles, or enamel crenulations. There is anterior and posterior cingulum with some small tubercles positioned at the middle of the mouth of the outside of the valley. P4 is almost like that of Propotamochoerus.
The L. lufengensis specimen consists of a fragmentary mandible. At the time of discovery, the fragments were collected together. Moreover, the left and right P1 are alike in size and morphology and should be assigned to the same individual. Impor- tant features of the teeth indicate that the Lufeng specimen is without doubt that of Lophochoerus. Nevertheless, the posterior crest of the primary cusp of P3 and Pq, the structure of the talonid, lack of buccal tubercles on M3, and the developed anterior crest of the third blade, etc., are characteristics of the new Lufeng species that differ from the three Lophochoerus species discovered in the lower and middle Siwaliks. The lengths of P4 and M2 of the Lufeng specimen are slightly longer than in L. himalayensis (M2 length 9.5 mm, cf. Koenigswald, 1963, p. 195). The outline of P4 is not conical in shape. The length and width of P4 and M2 in L. himalayensis is about the same (Pilgrim, 1926: plate X, fig. 51, but the length and width of the P4 of the Lufeng specimen is smaller than that of M2. From the above comparison, we conclude that the discovery of the Lufeng material represents a new species, i.e., L. lufengensis sp. nov.
As stated earlier, the Lophochoerus mandibular fossils collected earlier in the Siwaliks were incomplete. Because of this we still do not know the tooth row pattern of this suid’s premolars. Recently, there have been reports of Lophochoerus fossils (Pilbeam, 1979; Badgley and Behrensmeyer, 19801, although we have not seen analytic studies of any specimens.
The mandible of L. lufengensis had been fragmented and the true position of the four premolars in the mandible cannot be made out. The anterior and posterior faces of P1-P3 and the anterior face of P4, however, all lack contact facets, which seems to indicate that there were definite diastemata between the premolars and that they were not in direct contact with each other. It is true that P1-P3 have not been worn, but that does not necessarily prove that the lack of contact facets is due to the fact that the specimen is of a juvenile individual. Although our diagram indicates the presence of diastemata between the premolars, this characteristic must be verified by the discovery of more specimens.
Koenigswald (1963) described two Lophochoerus teeth obtained from Chinese apothecaries in Hong Kong. Among them, one was a very deeply worn tooth that could not be assigned to a species or be subject to comparison. The other, an M2 (L. pilgrimi Koenigswald), is a slightly worn tooth (Koenigswald, 1963: plate 1, Fig. 12). The opposite cusps of the anterior part of this upper molar have joined to form a crest, but the opposite cusps of the posterior part are still separate. As the Koenig- swald specimen is an M2 and our specimen is of the lower dentition, the two cannot be directly compared. The two rows of cusps, both anterior and posterior of the M3 of L. lufengensis, however, have formed clear transverse crests, which differs from L. pilgrim.
In regard to the systematic position of Lophochoerus, Colbert (1935, p. 8) writes as follows: “The pattern of relationship between Lophochoerus and other suids is still a difficult question.” Pilgrim proposes that it split off from the main stem of Propotamochoerus-Potamochoerus. The discovery of L. lufengensis fossils in China

53 Etler] FOSSIL HOMINOIDS OF LUFENG
advances evidence that the associated fauna of the Lufeng hominoids and the Siwalik fauna have a number of features in common. In comparing the P 4 and M 2 of the new Lufeng species, the latter is longer than the former. This differs from L. himalayensis in which P 4 and M 2 are equal in length and indicates a certain progressive quality. The structure of P3 and P4, however, and the outline of P 4 of L. lufengensis are unlike that seen in L. himalayensis and L. nagrii. Whether or not the new Lufeng species and the several Siwalik species can be considered to have a direct ancestralldescendant relationship requires more material and investigation.
LITERATURE CITED
(The literature cited is collated from the total collection of articles which were translated.)
Andrews, P (1970) Two new fossil primates from the lower Miocene of Kenya. Nature 228r537-540. (Lu, 1981)
Andrews, P (1971) Rarnapithecus wickeri mandible from Fort Ternan, Kenya. Nature 231t192-194. (Xu, 1978,1979)
Andrews, P (1982) Hominoid evolution. Nature 295185-186. (Wu, 1983)
Andrews, P, and Cronin, JE (1982) The relation- ships of Siuapithecus and Ramapithecus and the evolution of the orang-utan. Nature 297:541-546. (Wu, 1983)
Andrews, P, and Tekkaya, I (1976) Ramapithecus in Kenya and Turkey. IXe Congr. Union Int. Sc. Prehist. Protohist., Coll. VIt7-25. (Xu, 1979; Wu, 1983)
Andrews, P, and Tobien, H (1977) New Miocene locality in Turkey with evidence on the origin of Ramapithecus and Siuapithecus. Nature 268r699- 701. (Xu, 1978,1979)
Badgley, C, and Behrensmeyer, AK (1980) Paleo- ecology of Middle Siwalik sediments and fauna, Northern Pakistan. Paleogeogr. Paleoclim. Pa- leoecol. 30:133-155. (Han, 1983)
Banetice, D (1968) Siwalik microflora from Punjab, India. Rev. Paleaoboton. Palynol. 6:171-176. (Sun, 1980)
Bi, Z, et al. (1977) Division of the upper Tertiary and mammalian fossils in the vicinitv of Naniina “ I
Vert. PalAsiat. 15(2). (Qi, 1979) Bonis, L de, and Melentis, J (1976) Les Dryopithe- cines de Macedoine (Grece): Leur place dans L’ev- olution des Primates hominoides du Miocene. IX“ Congr. Union Int. Sc. Prehist. Protohist. Coll. VE26-38. (Xu, 1979)
Bonis, L de, and Melentis, J (1977) Les Primates hominoides du Vallesian de Macedoine (Grece). Etude de la Machoire inferieure. Gkobws No. 10. fasc. 65349-885. (Xu, 1979)
Colbert, EH (1935) Distributional and phylogenetic studies of Indian fossil mammals (4): Am. Mus. Novitatus N. 799. Wan, 1983)
Colbert, EH (1935) Siwalik mammals. Trans. Am. Philos. Soc. 26t82-126. (Qi, 1979,1983)
Colbert, EH (1939) Carnivora of the Tung-Gur for- mation of Mongolia. Bull. Am. Mus. Nat. Hist. 76t47-81. (Qi, 1983)
Conroy, GC, and Pilbeam, D (1975) Ramapithecus: A review of its hominid status. In R Tuttle (ed) Paleoanthropology, Morphology, and Paleoecol- ogy. The Hague: Mouton. (Qi, 1979)
Ewer, RF (1955) The fossil carnivores of Transvaal caves: Machairadontinae. Proc. Zool. SOC. Lond. 125t587-615. (Qi, 1983)
Folhol, MN (1891) Etudes sur les mammiferes fos- siles de Sansan. Ann. Soc. Geol. 21:94-117. (Qi, 1983)
Forsten, AM (1968) Revision of the paleartic H i p parion. Acta Zool. Fenn. 119:. (Qi, 1979)
Ghosh, AK (1977) Palynology of Siwaliks. Adv. Pol- len Spore Res. 2:14-19. (Sun, 1980)
Ginsburg, L (1961) La faune des carnivores Mio- cenes de Sansan. Mem. Mus. Nat. Hist. [C], 9. (Qi. 1983)
Greenfield, LO (1979) On the adaptive pattern of “Ramapithecus”. Am. J. Phys. Anthropol. 50t527- 548. (Wu, 1983)
Gregory, WK (1922) The Origin and Evolution of the Human Dentition. Baltimore: Williams and Wilkins. (Xu, 1979)
Gregory, WK, and Hellman, M (1926) The dentition of Dryopithecus and the origin of man. Anthropol. Pap. Am. Mus. Nat. Hist. 28:l-123. (Xu, 1979)
Gregory, WK, Hellman, M, and Lewis, GE (1938) Fossil Anthropoids of the Yale-Cambridge India Expedition of 1935. Carnegie Inst. Washington 1- 27. (Xu, 1979)
Huang, W (1974) The Pliocene of Huoxian, Shanxi. Vert. PalAsiat. 12(1). (in Chinese) (Qi, 1979)
Hussain, ST (1969) Revision of Hipparion from the Siwalik Hills of Pakistan and India. Bayeriache Akad. Wiss. Math. Natur. Klasse Abhandlungen. Neue Folge, Heft 147. (Qi, 1979)
Koenigswald, GHR, von (1963) Fossil pygmy Sui- dae from Java and China. Proc. Koninkl. Nederl. Akad. van Wetenschappen. Amsterdam. Series B66:192-197. (Han, 1983)
Koenigswald, GHR, von (1972) Ein Unterkiefer eines fossilen Hominoiden aus Unterpliozan Griechlands. Proc. Koninkel. Nederl. Akad. van Wetenschappen. Amsterdam. 75385-394. (Xu, 1979)
Kretzoi, M (1975) New ramapithecines and Pliopf thecus from the Lower Pliocene of Rudabanya in north-eastern Hungary. Nature. 257t578-581. (Xu, 1978,1979)
Kurten, B (1952) The Chinese Hipparion fauna. Comments. Biol. Soc. Sci. Fenn. 13(4): (Qi, 1979)
Kurten, B (1954) The type collection of Ictitheriurn robusturn and the radiation of the ictitheres. Acta Zool. Fenn. 86:l-26. (Qi, 1983)
Kurten, B (1971) The Age of Mammals. New York: Columbia University Press. (Qi, 1979)
Le Gros Clark, WE, and Leakey, LSB (1951) The Miocene Hominoidea of East Africa. Fossil Mam- mals Africa. Br. Mus. Nat. Hist. 1:l-117 (Xu, 1979; Lu, 1980; Wu, 1983)
Leakey, LSB (1962) A new Lower Pliocene fossil

54 YEARBOOK OF PHYSICAL ANTHROPOLOGY [Vol. 27, 1984
primate from Kenya. Ann. Mag. Nat. Hist. 4:689- 696 (Xu, 1979)
Leakey, LSB (1967) An Early Miocene member of Hominidae. Nature 213:155-163. (Xu, 1979)
Leakey, LSB (1968) Upper Miocene primates from Kenya. Nature 218:527-528. (Xu, 1979)
Lei, C, and Lin, Y (1982) New species of sabre-tooth cat from Liyang, Xialin, Jiangsu. Vert. PalAsiat. 20:54-57. (in Chinese) (Qi, 1983)
Lewis, GE (1934) Preliminary notes of new man- like apes from India. Am. J. Sci. 27:161-179. (Xu, 1979; Wu, 1983)
Lewis, GE (1937) Taxonomic syllabus of Siwalik fossil anthropoids. Am. J. Sci. 34:139-147. (Xu, 1978,1979)
Licenti, E, and Trassaert, M (1935) The Pliocene lacustrine series in central Shansi. Bull. Geo. Soc. China. 14: (Qi, 1979)
Liu, D, Li, Z, and Zhai, R (n.d.1 Pliocene Vertebrate Fossils from Lantian, Shaanxi. (in Chinese) (Qi, 1979)
Lu, Q, Xu, Q, and Zheng, L (1981) Preliminary research on the cranium of Siuapithecus yunna- nensis. Vert. PalAsiat. 19(2):101-105. (in Chinese) (Wu, 1983)
Lukose, NG (1969) Microfossils from the Middle Siwaliks of Bihar, India. J. Palgnol. 4:107-112 (Sun, 1980)
Lydekker, R (1884) Siwalik and Narbada Carni- vora. Pal. Ind. Ser. X. Vol. 11, Part 6. (Qi, 1979,1983)
Matthew, WD (1928) Critical observations upon Si- walik mammals. Bull. Am. Mus. Nat. Hist. 54:437-560. man, 1983)
Nandi, B and Bandyopadhvay, NN (1970) Prelimi- nary observations on the microfossils and micro- spores of Siwalik lignite from Himachal Pradesh, India. Sci. Cult. 36:200-242. (Sun, 1980)
Nandi, B (1972) Some observations on the micro- flora of Middle Siwalik sediments of Mohand (east) field, Uttar Pradesh. Proc. Semin. Palaeopalynol. India strat. 275-383. (Sun, 19801
Napier, JR, and Napier, PH (1976) A Handbook of Living Primates. London: Academic Press. (Wu, 1983)
Pearson, HS (1928) Chinese fossil Suidae. Pal. Sin. Ser. C., Vol. 5, Fasc. 5. (Qi, 1979)
Petter, G (1976) Etude d’un nouvel ensemble de petits carnivores du Miocene d‘Espagne. Geol. Med. 3:135-154. (Qi, 1983)
Pilbeam, D (1966) Notes on Ramapithecus, the ear- liest known hominid, and Dryopithecus. Am. J. Phys. Anthropol. 25:l-6. (Xu, 1979)
Pilbeam, D (1969) Tertiary Pongidae of East Africa: Evolutionary relationships and taxonomy. Bull. Peabody Mus. Nat. Hist. 31:l-185. (Xu, 1979)
Pilbeam, D (1969) Newly recognized mandible of Ramapithecus. Nature 222:1093-1094. (Xu, 1978,1979)
Pilbeam, D (1976) Neogene hominoids of south Asia and the origins of the Hominidae. IXe Congr. Union. Int. Sc. Prehist. Protohist. Coll. VI:39-59. (Xu, 1979)
Pilbeam, D (1979) Miocene sediments and fauna of Pakistan. Bull. Peabody Mus. Nat. Hist. Yale Univ. 179:l-45. man, 1983)
Pilbeam, D (1980) Major trends in human evolu- tion. In L.K. Konigsson (ed) Current Argument on Early Man. Oxford: Pergamon, pp. 261-285. (Wu, 1983)
Pilbeam. D (1982) New hominoid skull material
Pilbeam, D, and Smith, R (1981) New skull remains of Siuapithecus from Pakistan. Mem. Geol. Survey Pakistan 11:l-13. (Wu, 1983)
Pilbeam, D, et al. (1977a) Geology and paleontology of Neogene strata of Pakistan. Nature 270t684- 689. (Xu, 1979; Qi, 1979; Lu, 1981; Wu, 1983)
Pilbeam, D, et al. (1977b) New hominoid primates from the Siwaliks of Pakistan and their bearing ~~ .~~~ ~~~~ ~ ~
on hominoid evolution. Nature 270:689-695. (Xu, 1979; Wu, 1983)
Pilbeam, D, et al. (1980) Miocene hominoids from Pakistan. Postilla 181:l-94. N u , 1983)
Pilmim, GE (1915) New Siwalik primates and their bearing on the question of the- evolution of man and the Anthropoidea. Rec. Geol. Surv. Ind. 451- 74. (Xu, 1978,1979)
Pilgrim, GE (1926) The fossil Suidae of India. Geol. Surv. India. 8. (4). (Qi, 1979; Han, 1983)
Pilgrim, GE (1932) The fossil Carnivora of India. Paleontol. Ind. N. Ser. No. 18: (Qi, 1979,1983)
Pilgrim, GE (1931) Catalogue of Pontian Carnivora of Europe. Cat. Br. Mus. Nat. Hist. (Qi, 1979,1983)
Prasad, KN (1975) Observation on the paleoecology of south Asian Tertiary primates. In R. Tuttle (ed) Paleoanthropology, Morphology and Paleoecology. The Hague: Mouton. (Qi, 1979)
Preuss, TM (1982) The face of Szuapithecus zndicus: Description of a new, relatively complete speci- men from the Siwaliks of Pakistan. Folia Prima- tol. 38:141-157. (Wu, 1983)
Qiu, ZD (1979) Pliocene mammalian fossils from several localities in North China. Vert. PalAsiat. 17:222-235. (in Chinese) (Qi, 1983)
Qiu, ZX, et al. (1979) Hyaenidae fossils from the Pliocene of Qingyang, Gansu. Vert. PalAsiat. 17:200-221. (in Chinese) (Qi, 1983)
Ringstrem, NF (1924) Nashorner der Hipparion- Fauna Nord-Chinas. Pal. Sin. [C] 1(4). (Qi, 1979)
Sefve, I(1927) Die Hipparion Nord-China. Pal. Sin. Ser. C 1(4): (Qi, 1979)
Simons, EL (1961) The phyletic position of Rama- pithecus. Postilla. 57:l-10. (Wu, 1983)
Simons, EL (1964) On the mandible of Ramapithe cus. Proc. Natl. Acad. Sci. USA 51t528-535. (Xu, 1979)
Simons, EL (1968) A source for dental comparison of Ramapithecus with Australopithecus and Homo. S. Afr. J. Ski. 64:92-112. (Xu, 1978,1979)
Simons, EL (1976) Relationships between Dryopi- thecus, Siuapithecus and Ramapithecus and their bearing on hominid origins. IXe Congr. Union Int. Sc. Prehist. Protohist. Coll. VIr60-67. (Xu, 1978,1979)
Simons, EL (1977) Ramapithecus. Sci. Am. 236(5):28-35. (Xu, 1978,1979)
Simons, EL (1981) Man’s immediate forerunners. Philos. Trans. R. Soc. Lond. [Biol.] 292:21-41. (Wu, 1983)
Simons, EL, and Pilbeam, D (1965) Preliminary revision of the Dryopithecinae. Folia Primatol.
Teilhard de Chardin, P, and Leroy, P (1945) Les Feledes de Chine. Public. Inst. Geobiol. No. 11. (Qi, 1979)
Teilhard de Chardin, P, and Leroy, P (1945) Les Mustelides de Chine. Public. Inst. Geobiol. No. 12. (Qi, 1979)
Teilhard de Chardin, P, and Trassaert, M (1937) Pliocene Camelidae, Giraffdae, and Cervidae of South-Eastern Shansi. Pal. Sin. New Ser. C, 1. (Qi. 1979)
3:81-152. (XU, 1978,1979; WU, 1983)
, y - 1 ~- - - from the Miocene of Pakistan. Nature 295:232- 234. (Wu, 1983)
Teilhard de Chardin, P, and Trassaert, M (1938) Caviconia of South-Eastern Shansi. Pal. Sin. New

Etler] FOSSIL HOMINOIDS OF LUFENG 55
Ser. C, 6. (Qi, 1979) Teilhard de Chardin, P, and Young, CC (1921) Fos. sil mammals from the Late Cenozoic of North China. Pal. Sin. Ser. C, 9(1). (Qi, 1979)
Teilhard de Chardin, P, and Young, CC (1933) The late Cenozoic Formation of S. E. Shansi. Bull. Geo. Soc. China. 12. (Qi, 1979).
Tekkaya, L (1974) A new species of Tortonian an- thropoid (Primates, Mammalia). Bull. Min. Res. Exp. Inst. Turkey. 83:148-165. (Xu, 1979)
Tobias, PV (1967) Olduvai Gorge, Vol. 2. The Cran- ium and Maxillary Dentition of Australopithecus (Zinjanthropus) boisei. Cambridge University Press. (Lu, 1981)
Tong, Y, et al. (1975) Hipparion fauna of Huoxian, Shanxi. Vert. PalAsiat. 13(1). (Qi, 1979)
Walker, A, and Andrews. P (1973) Reconstruction of the dental arcades of Ramupithecus wickeri. Nature 244:313-314. (Xu, 1979)
Wu, R (1957) Dryopithecus teeth from Keiyuan, Yunnan. Vert. PalAsiat. 1(1):25-32. (Xu, 1978,1979; Wu, 1983)
Wu, R (1958) New materials of Dryopithecus from Keiyuan, Yunnan. Vert. PalAsiat. 2(1):38-43. (Xu, 1979; Wu, 1983)
Wu, R, et al. (1978) The History of Human Devel- opment. Beijing: Science Publishing House. (Sun, 1980)
Wu, R, et al. (1981) Ramapithecus skull found first time in the world. Sci. Bull. 26:1125-1127. (Chinese edition) 26(11):1018-1020. (English edition) (Wu, 1983)
Wu, R, et al. (1982) More Rarnapithecus skulls found from Lufeng, Yunnan. Acta Anthropol. Sin. 1(2):101-105. (in Chinese) (Han, 1983; Wu, 1983)
Yan, D (1978) Duodaoshi formation of Jinxiang, Hubei. Vert. PalAsiat. 16(1). (in Chinese) (Qi, 1979)
Xu, Q, et al. (1978) Fossil mandible of the Lufeng Ramapithecus. Science Bull. 9:554-556, 546. (in Chinese) (Xu, 1979; Lu, 1980)
Xu, Q and Lu, Q (1979) The mandibles of Rarnapi- thecus and Siuapithecus from Lufeng, Yunnan. Vert. PalAsiat. 17(1):1-18. (Lu, 1980)
Zdansky, 0 (1924) Jungtertiare Carniviren Chinas. Pal. Sin. Ser. [C1 Vol. 1, Fasc. 1. (Qi, 1983)
Zdansky, 0 (1927) Weitere Bemerkungen uber fos- sile Carnivoren aus Chinas. Pal. Sin. Ser. C, Vol. 4, Fasc. 4. (Qi, 1979)
Zhang, X (1957) Sabre-tooth tigers of Pliocene Pon- tian deposits, Shanxi. Vert. Zool. 1:193-200. (in Chinese) (Qi, 1983)
Zhang, X, and Liu, H (1964) Metuilurus fossil from Xiakou, Yushi, Shanxi. Vert. PalAsiat. 8:182-185. (in Chinese) CQi, 1983)
POSTSCRIPT
Interested readers should refer to a number of other papers detailing the Lufeng hominoids and their physical environment. These include:
Flynn, L and Qi, G (1982) Age of the Lufeng, China hominoid locality. Nature 298:746-748.
Wu, R, et al. (1981) Ramapithecus skull found first time in the world. Science Bulletin 26(11):1018-
Wu, R, et al. (1984) Morphological features of Ra- mapithecus and Siuapithecus and their phyloge- netic relationship- morphology and comparison of the mandibles. Acta Anthropologica Sinica 3(1):1-
1020 (English edition) 10. Wu, R and Oxnard, C (1983) Rarnapithecus and Siuapithecus from China: some implications for higher primate evolution, Am. J. Primatol. 5303- 344.
Zhang, X, et al. (1981) Stratigraphy of Ramapithe- cus bearing Pliocene of Lufeng, Yunnan. Memoirs of Beijing Nat. Hist. Museum 10.
The presence of Miocene hominoids of a primitive aspect in Sihong county, Jiangsu province, along China’s eastern seaboard north of Shanghai has also recently come to light. Excavations at this latest site along with new excavations at Lufeng and Kaiyuan in Yunnan will yield new information on hominoid origins and evolution in East Asia during the Neogene and play an increasingly important role in our understanding of the diversity and dispersal of the higher primates.
I would like to thank the authors for their review of my translations and many helpful suggestions.




















