
Male workers in the female-dominated long-term care sector: evidence from England
Upload
independentCategory
view
1download
0
J. Insect Physiof. Vol. 37, No. 12, pp. 869-882, 1991 Printed in Great Britain. All rights reserved
0022-1910/91 $3.00 + 0.00 Copyright 0 1991 Pergamon Press plc
THE FEMALE- AND MALE-PRODUCED VITELLOGENINS OF LEUCOPHAEA MADERAE
GRACE DON-WHEELER and FRANZ ENGELMANN
Department of Biology, University of California, Los Angeles, CA 90024, U.S.A.
(Received 21 May 1991; revised 29 July 1991)
Abstract-Vitellogenin synthesis was induced in adult females and males as well as in last-instar nymphs of both sexes of Leucophaea maderae by the juvenile hormone analogue methoprene. Vitellogenic competency in females and males is developed during the last nymphal instar, but maximal vitellogenic competency in females is attained only after emergence. No increase in competency is observed for adult males. Once vitellogenic competency is established, last-instar female nymphs synthesize vitellogenin at a higher rate than either adult or last-instar males. Female- and male-produced vitellogenins are different in their polypeptide compositions. The difference in composition may reflect differences in vitellogenin processing in the female and male fat body. Female- and male-produced vitellogenins are taken up into competent oocytes at different rates in uivo and in uitro, i.e. the female-produced vitellogenin is preferred over the male-produced vitellogenin.
Key Word Index: Vitellogenins; vitellins; methoprene; fat body; vitellogenesis
INTRODUCTION
In many species of insects, juvenile hormones induce the synthesis of vitellogenin in the fat body of adult females (Engelmann, 1979). During the gonotrophic cycle large quantities of vitellogenin are produced and secreted into the haemolymph. It is transported to competent oijcytes where it is taken up via receptor-mediated endocytosis (Riihrkasten and Ferenz, 1985). Once vitellogenin is taken up into the oocyte, it is processed to vitellin, the yolk protein.
The induction of vitellogenin synthesis in insects provides a model system in which to study the regulation of expression of a protein. In insects, juvenile hormone is neither a stage- nor sex-specific hormone yet the induction of vitellogenin synthesis is normally limited to adult females. It is possible, however, to induce vitellogenin synthesis in adult males and nymphs with large doses of juvenile hor- mone or juvenile hormone analogues. This has been demonstrated for Nauphoeta cinerea (Lanzrein, 1974), Diploptera punctata (Mundall et al., 1979), Blattella germanica (Kunkel, 1981) Locusta migra- toria (Dhadialla and Wyatt, 1983), and Leucophaea maderae (della-Cioppa and Engelmann, 1984a; Don- Wheeler and Engelmann, 1989). In these nymphs and males, usually only a fraction of the vitellogenin is made compared to normal vitellogenic adult females. Thus, there are stage- and sex-specific differences in the response of the fat body to juvenile hormone.
Vitellogenesis in females as well as the vitellogenic growth of ovaries implanted in males may be affected by the quantity and the quality of the vitellogenin available for uptake into competent oijcytes (Lamy,
1984). In the cockroach species Diploptera (Mundall et al., 1979) and Leucophnea (Don-Wheeler and Engelmann, 1989), vitellogenesis in males was deter- mined by implanting competent previtellogenic ovaries into adult males. In Leucophaea, implanted oiicytes never grew to full size and eventually were resorbed. This may be due to the low vitellogenin titre of the male haemolymph and to the structure of the vitellogenin produced in males (Don-Wheeler and Engelmann, 1989). It has been shown that implanted oijcytes completed vitellogenesis in Diploptera males despite a low vitellogenin titre of the males (Mundall et al., 1979). However, here too, the number of oocytes that completed vitellogenesis and the vitellin content per oijcyte was less than what is found in vitellogenic females.
Leucophaea is a convenient model in which to examine the competency of the fat body to synthesize vitellogenin as related to sex and developmental age. The development of vitellogenic competency as well as the magnitude of vitellogenin synthesis were deter- mined in adults and nymphs of both sexes. The characterization of the vitellogenin produced in females and males revealed differences in these vitellogenins. The uptake of female- and male- produced vitellogenin into the oijcyte was evaluated in vivo and in vitro.
MATERIALS AND METHODS
Animats
Animals were reared 12 h light-12 h dark cycle, at 26°C 75% r.h. Adults, last- and penultimate-instar

870 GRACE DON-WHEELER and FRANZ ENGELMANN
nymphs were collected shortly after the moult and kept in plastic cages (6-8 animals per cage). Animals were fed rat chow and given water ad libitum.
Allatectomy and ovariectomy
All surgical procedures were performed under carbon dioxide narcosis. Allatectomy was performed on adults on the fourth day after emergence as previously described (della-Cioppa and Engelmann, 1984b). Adult females were ovariectomized 4 days later by removing the ovaries through incisions made in the intersegmental membrane between the fifth and sixth dorsal abdominal segment. In all operations, a few grains of penicillin-streptomycin were placed into the incisions; the wounds were sealed with warm paraffin wax.
Methoprene treatment
The juvenile hormone analogue methoprene (a gift from Zoecon Corp., Palo Alto, Calif.) was used for all hormone treatment. Methoprene was diluted in acetone and applied topically to the dorsal abdominal segments of adults (allatectomized and ovari- ectomized), last- and penultimate-instar nymphs. In all instances, animals received a single 400 pg dose of methoprene while control animals were treated with acetone only. Animals were treated on the twelfth day following either adult emergence, or last- and penultimate-instar nymphal moults.
Vitellogenin uptake into oiicytes in vivo
Previtellogenic competent ovaries were dissected from newly emerged adults and cleaned of the adher- ing fat body. Ovaries were implanted&o the haemo- coel of adults (8 days after emergence) or last-instar nymphs (8 days after the nymphal moult). These animals were treated 4 days later with a 400 ,ug dose of methoprene. On the sixth day following metho- prene treatment, the ovaries were recovered and vitellogenin uptake into oijcytes was evaluated by quantifying the vitellin content of the whole ovary and by measuring the Iength of the terminal oiicytes.
PuriJication of vitellogenin and vitellin
Vitellogenins from adult females and males were purified by QAE ion-exchange column chromatog- raphy. Elution of vitellogenin was effected with 0.2-1.0 M NaCl in phosphate-citrate buffer, pH 6.8 (Engelmann et al., 1976). Vitellogenin was con- centrated by ultrafiltration using a Centriprep Concentrator (Amicon). The protease inhibitor phenylmethylsulphonyl fluoride (PMSF) was added to a final concentration of 0.1 mM.
Vitellin was purified from ovaries implanted into methoprene-treated adults and last-instar nymphs. Yolky oiicytes were homogenized in phosphate- buffered saline (0.01 M NaH,P04 pH 7.0, 0.4 M NaCl, 0.1 mM PMSF) and the cellular debris was pelleted by centrifugation for 1 min in a Beckman
Microfuge B. The supernatant was dialysed exten- sively against distilled water containing 0.1 mM PMSF. Because vitellin precipitates out in low ionic strength solutions, it was possible to separate vitellin from the other egg proteins (Engelmann, 1979). The purities of vitellogenin and vitellin were determined by immunoelectrophoresis (Garvey et al., 1977) or by SDS-PAGE (polyacrylamide gel electrophoresis) according to Laemmli (1970).
Radiolabelled vitellogenin and vitellin
Vitellogenins were metabolically labelled with either 1 ,uCi of [‘4C]leucine (sp. act. 184 mCi/mmol), 50 PCi of Trans3’S-label ([3’S]-methionine and [35S]- cysteine; sp. act. 1082 mCi/mmol), or 0.5 mCi of [32P]orthophosphate (285 Ci/mg, carrier free). All radioisotopes were purchased from ICN Radio- chemicals. The isotope was injected into the haemo- coel of methoprene-treated or control animals with a Hamilton microlitre syringe. After 4 h, the animal was neck-ligated and a puncture wound was made in the dorsal neck membrane. The animal was placed head down in a Kolmer centrifuge tube (Fisher Scientific) and the haemolymph was collected by centrifugation at low speed. Haemolymph samples were routinely stored at -20°C.
Radiolabelled vitellin was obtained by pulse- labelling methoprene-treated adults and last-instar nymphs that had been implanted with ovaries. These hosts were pulsed with 5OpCi of Trans3’S-label for 24 h. The ovaries were recovered and vitellin was isolated as previously described.
Immunological metho&
Antibodies against vitellogenin were prepared by immunizing rabbits with a haemolymph from vitello- genie adult females. Non-sex specific antibodies were eliminated from the antiserum by absorption with haemolymph from adult males. The resulting anti- serum was shown to contain only antibodies against vitellogenin. The specificity of the rabbit anti- vitellogenin was assessed by immunoelectrophoresis (Garvey et al., 1977) and by the Western immuno- blotting method (Burnette, 1981).
Vitellogenin and vitellin were quantified by using rocket immunoelectrophoresis according to Monthony et al. (1978). Samples and known quan- tities of purified vitellogenin were diluted in 0.4 M NaCl and electrophoresed in a 1% (w/v) agarose gel (Bio-Rad Agarose Immunoelectrophoresis Tablet) containing 1.5% (v/v) rabbit anti-vitellogenin. Electrophoresis was performed at 140 V for 18 h at room temperature. The quantitation of vitellogenin and vitellin by this method has been previously described (della-Cioppa and Engelmann, 1984a).
Rates of vitellogenin synthesis
Rates of vitellogenin synthesis in methoprene- treated and control animals were determined by the

Female- and male-produced vitellogenins 871
incorporation of [14C]Ieucine into vitellogenin in a 4 h pulse in uiuo. The immunoprecipitation of radio- labelled vitellogenin from the haemolymph was per- formed as previously reported (della-Cioppa and Engelmann, 1984b). Haemolymph samples were di- luted with phosphate-buffered saline and then pre- cipitated with anti-vitellogenin for 24 h at 4°C. The precipitates were washed four times with immuno- precipitation wash buffer (0.1 M Tris pH 7.5, 0.2 M NaCl, 1% NP-40) and then solubilized in 1 N NaOH. This material was analysed by liquid scintillation spectrometry in a Beckman LS 7000. The amount of radioactivity contained in antibody-precipitated vitellogenin was used as an indicator for the rate of vitellogenin synthesis.
Polyacrylamide gel electrophoresis (PAGE)
A Hoefer SE 600 vertical slab electrophoresis apparatus (Hoefer Scientific Instruments) was used for both non-denaturing PAGE and SDS-PAGE. Acrylamide, N,N’methylene-bisacrylamide, Tris base, sodium dodecyl sulphate were purchased from BRL. TEMED and ammonium persulphate were obtained from Bio-Rad.
The native molecular weights of the adult vitello- genins were determined by non-denaturing PAGE (Davis, 1964). For this, QAE-purified vitellogenins and protein standards of known molecular weights (Sigma) were electrophoresed on 4-8% polyacryl- amide gels. The molecular weights were calculated according to Hedrick and Smith (1968). The proteins were visualized by the use of the ultrasensitive silver staining method for polyacrylamide gels (Van Keuren er al., 1981).
The polypeptide compositions of the vitellogenins were determined by the fractionation of 35S- or 32P-labe11ed vitellogenins by 7.5% SDS-PAGE according to Laemmli (1970). The radiolabelled vitellogenins were immunoprecipitated from the haemolymph as previously described and these pre- cipitates were subsequently solubilized in Laemmli sample buffer (62.5 mM Tris pH 6.8, 2.3% SDS, 5% 2-mercaptoethanol, 10% glycerol). Samples were electrophoresed on 7.5% polyacrylamide gels. Gels containing 32P-1abelled vitellogenins were fixed in 10% methanol/7% acetic acid and dried onto filter paper. Gels containing 35S-labelled vitellogenins were fixed and then fluorographed with 22% 2,5 diphenyl- oxazaole in dimethysulphoxide before drying (Bonner and Laskey, 1974). Kodak X-AR film was used for both 32P and 35S autoradiography.
Protease digestion of vitellogenin polypeptides
The fragment compositions of adult vitellogenin polypeptides were determined by digestion of the polypeptides with StaphyZococcus aureus V-8 protease (Sigma) which cleaves at the C-terminus of aspartic acid and glutamic acid residues (Cleveland et al., 1977). 32 P-labelled vitellogenins were first electro-
phoresed on a 7.5% SDS-polyacrylamide gel and the vitellogenin polypeptides were then visualized by staining with Coomassie blue R-250. The poly- peptides were excised from gels and equilibrated for 1 h in Cleveland sample buffer (0.125 M Tris pH 6.8, 0.1% SDS, 10% glycerol, 0.3% 2-mercaptoethanol, 1 mM EDTA). Individual gel slices were placed into the wells of a 15% polyacrylamide gel and then overlayed with 10 pg of V-8 protease in sample buffer. Proteolysis of the polypeptides was effected in the stacking gel (dye front at the bottom) by turning off the current for 30 min. Low molecular weight protein standards (BRL) were electrophoresed in a separate well. The fragment compositions of the vitellogenin polypeptides were revealed by 32P autoradiography.
Vitellogenin uptake in vitro
32P-1abe11ed vitellogenins were prepared by pulsing methoprene-treated adult females and males with 1.0 mCi [32P]orthophosphate for 2 h. The haemo- lymph was collected and 125 ~1 were diluted (1: 1) with 0.4 M NaCl. The unincorporated 32P-1abel was removed by gel filtration on a Bio-Rad P-10 column (25 cm x 0.6 cm). Under the conditions used for in vivo labelling, 98% of the 32P incorporation is found in vitellogenin (Engelmann and Friedel, 1974). The specific activities of 32P-labelled vitellogenins were ascertained by antibody-precipitation of vitellogenin specific counts and by quantitation of the vitellogenin by rocket immunoelectrophoresis. For the assay, the specific activities of the 32P-labe11ed female and male vitellogenins were adjusted to 1000 cpm/pg vitello- genin by the addition of the respective unlabelled vitellogenin.
Mid-vitellogenic ovaries (terminal oiicyte of 3-4 mm) were collected from vitellogenic adults. Only ovaries with undamaged terminal oiicytes were used to determine the uptake of vitellogenin in vitro. The assay medium was composed of HB 101 (Hana Biologics, Emeryville, Calif.) pH 7.2, supplemented with 10% adult male haemolymph. For each determi- nation, 5 ovarioles were placed in the bottom of a 7 x 1 cm glass tube and then incubated in 250 ~1 of medium containing 100,000 cpm of female or male vitellogenin for 5 h at 30°C. The specificity of 32P- labelled vitellogenin uptake was determined by com- petition with unlabelled vitellogenin or y-globulin. Following the incubation, the vitellin of the oiicytes was collected by first washing the follicles several times with ice-cold cockroach saline (5 mM KCI, 0.4 M NaCl, 1 mM CaCl,) and then homogenizing in distilled water to precipitate the vitellin. Vitellin was pelleted by centrifugation and solubilized in 1 N NaOH. The amount of radioactivity contained in the vitellin pellet was determined by liquid scintil- lation spectrometry and this value was used as index of vitellogenin uptake. Statistical significance for paired differences was ascertained using the Student’s t-test.

872 GRACE DON-WHEELER and FRANZ ENGELMANN
RESULTS
The induction of vitellogenin synthesis in adult females and males
The rates of vitellogenin synthesis and the vitello- genin titres of adult ovariectomized females and males following a single application of methoprene were assessed over a &day period. The incorporation of [‘4C]leucine into vitellogenin during a 4 h period was used as an index for the rate of vitellogenin synthesis in the fat body. Following methoprene exposure, newly synthesized vitellogenin was first detected in females within 24 h, whereas in males, significant vitellogenin synthesis was not detected until after 48 h (Fig. 1). In males, the rate of vitello- genin synthesis increased until day 4 and declined after day 5. Vitellogenin synthesis in females contin- ued to rise during the 6 days. Throughout this period, the rate of vitellogenin synthesis in females was greater than that of males. Differences in the rate of vitellogenin synthesis in adult females and males was also reflected in the vitellogenin titre of the haemo- lymph (Fig. 2). Adult females had a vitellogenin titre of 20 /*g/p1 after 72 h of treatment whereas the titre of males remained low (1 pg/pl) for several days. A marked increase in the vitellogenin titres of males was observed on the fifth and sixth day of treatment (Fig. 2). By the sixth day, the titre of adult ovari-
0 1 2 3 4 5 6
Days after methoprene
Vitellogenin synthesis in adult ovariectomized females and males at days O-6 following the administration of a single 4OOpg dose of methoprene. The animals were injected with 1 nCi of [‘4C]leucine for 4 h at the given days. The vitellogenin in 10~1 of haemolymph was immuno- precipitated with rabbit-anti-vitellogenin. Each value shows the mean + SE for three animals. VgF, female vitellogenin;
VgM, male vitellogenin.
ectomized females was 18 times greater than that of adult males.
Vitellogenin synthesis in nymphs and adults
The inducibility of vitellogenin synthesis during nymphal development was established by treating last- and penultimate-instar nymphs with 400 pg of methoprene. The rate of vitellogenin synthesis and the vitellogenin titre in the haemolymph of these nymphs were assessed on the sixth day following methoprene exposure. Vitellogenin synthesis was not inducible in penultimate-instar nymphs as determined by the negligible incorporation of [‘4C]leucine (rela- tive to controls) and by the absence of vitellogenin in the haemolymph (Table 1). Vitellogenin synthesis was, however, inducible in female and male last-instar nymphs. Last-instar females exhibited a higher degree of vitellogenic competency than did last-instar males (Table 1). The extent of vitellogenin synthesis in last-instar females, however, was far less than that of adult ovariectomized females. A comparison of vitellogenin synthesis in adults and last-instar nymphs revealed that after 6 days of methoprene exposure, the vitellogenin titre of last-instar females was 12 times higher than that of either last-instar or adult males. Adult and last-instar males had similar low rates of vitellogenin synthesis and vitellogenin titres (Table 1).
200
t 180 t
160 t
Days after methoprene
Fig. 2. The time course of vitellogenin accumulation in the haemolymph of adult ovariectomized females and males treated with a single 4OOpg dose of methoprene. The vitellogenin concentration of the haemolymph was deter- mined by rocket immunoelectrophoresis. Each value shows
the mean + SE for three animals.

VaM VoF
540
Fig. 3. Analysis of vitellogenins by native PAGE. Female- and male-produced vitellogenins were isolated from methoprene-treated adults. The vitellogenins were purified by QAE ion-exchange column chroma- tography and then electrophoresed on non-denaturing polyacrylamide gels according to the method of Davis (1964). Molecular weights are indicated on the left. VgF, female vitellogenin; VgM, male
vitellogenin.
Adult Last -instar
‘W V9M VgF WM
x
- 85
Fig. 4. Analysis of vitellogenins by SDS-PAGE. The female- and male-produced vitellogenins of methoprene-treated adults and last-ins& nymphs were labelled with Trans35S-label in a 4 h pulse in viva. The vitellogenins were immunoprecipitated from the haemolymph and then fractionated on a 7.5% SDS-polyacrylamide gel (Laemmh, 1970). The polypeptide patterns were visualized by 35S autoradi- ography. Molecular weight standards are given on the far left. VgF, female vitellogenin; VgM, male
vitellogenin.

Adult
VtF VtM
Last -instar
VtF VW
Fig. 5. Analysis of vitellins from adults and last-instar nymphs by SDS-PAGE. Female- and male- produced vitellins were labelled with Tr~rr.s~~S-label in a 24 h pulse in r&o. The vitellins were isolated by distilled water precipitation. Following electrophoresis on 7.5% SDSpolyacrylamide gels, the polypeptide patterns were visualized by 35S autoradiography. The molecular weights of the vitellin polypeptides are
given in the figure. VtF, female vitellin; VtM, male vitellin.
874

(A)
W=
(B)
112 85 92 54 F M M F M F M
-97
-92
-87/85
-54
43
29
18
6.2
Fig. 6. (A) Autoradiogram of “P-labelled adult vitellogenins fractionated by 7.5% SDSPAGE. Molecular weight standards are shown in the far left lane. The molecular weights of the vitellogenin polypeptides are given on the far right. VgF, female vitellogenin; VgM, male vitehogenin. (F3) The fragment compositions of the respective polypeptides of female- and male-produced vitellogenin. The ‘*P-1abelled polypeptides (shown in A) were digested with S. aureus V-8 protease and the resulting fragments were resolved by 15% SDS-PAGE. The fragment patterns were visualized by 32P autoradiography. The
molecular weight standards are given on the far left. F, female vitellogeniq M, male vitellogenin.

Fig. 7. The uptake of female-produced vitellogenin (A) and male-produced vitellogenin (B) into obcytes in viva. Previtellogenic component ovaries were implanted into the haemocoel of adult ovariectomized females and males. The hosts were treated with a single 4OOpg dose of methoprene and the implanted
ovaries were recovered 6 days later.
876

Female- and male-produced vitellogenins 877
Table 1. The rate of vitellogenin synthesis and the vitellogenin titre of methoprene-treated and control animals
Vitellogenin synthesis (dpm x 10e3 per
5 pl of haemolymph)
Vitellogenin titre of the haemolymph
(pg/kJ) -
Adult female Methoprene Control
Adult male Methoprene Control
Last-instar female Methoprene Control
Last-instar male Methoprene Control
Penultimate-instar female Methoprene Control
Penultimate-instar male Methonrene
4253 f. 374 (6) 70 f 5 (5)
345 f 48 (4) 95 + 23 (4)
2539+ 131 (15) 83+ 11 (6)
328 & 37 (10) 69 + 7 (7)
143 + 15 (4) 153 + 12 (3)
130 * 22 (4)
245 + 29 (6) ND
5.8 + 1.2 (7) ND
62+8 (13) ND
4.lkO.5 (11) ND
ND ND
ND Control 139 + 33 (5j ND
Adults, last-instar and penultimate-instar nymphs were treated with a 400 pg dose of methoprene. Adult females were ovariectomized. Control animals were treated with acetone only. Animals were pulsed with [‘4C]leucine in uivo on the sixth day following treatment. Vitellogenin synthesis was assessed by the incorporation of [“‘Clleucine. Each value represents the mean &SE for the number of samples given in parentheses. ND, not detected.
The native molecular weights of female- and male- produced vitellogenins
Electrophoresis of the vitellogenins by non- denaturing PAGE revealed that female- and male- produced vitellogenins contained two bands with apparent molecular weights of 270 and 540 kDa (Fig. 3). The male vitellogenin contained small quan- tities of two additional bands of molecular weights of 210 and 180 kDa.
The polypeptide compositions of female- and male- produced vitellogenins
Analysis of 35S-labelled vitellogenins by SDS- PAGE revealed that the vitellogenins of adult females and males contained four polypeptides with molecu- lar weights of 112, 97, 92 and 54 kDa (Fig. 4). The vitellogenin of males, however, contained quantitat- ively less of the 112 and 97 kDa polypeptides, and an additional 85-87 kDa polypeptide “doublet”. The female- and male-produced vitellogenins of last- instar nymphs were similar to those synthesized by adults (Fig. 4). The female vitellogenin of last-instar nymphs, however, contained an 85 kDa polypeptide which was not present in the adult.
The polypeptide compositions of the female - and male - produced vitellins
In Leucophaea, the uptake of vitellogenin into the oocyte is followed by processing to vitellin. Because of the apparent structural differences in female- and
male-produced vitellogenins, it was of interest to compare the processing of these two vitellogenins within oijcytes. Oiicytes grown in vitellogenic females and males (adults and last-instar nymphs) contained proteins which were immunologically identical to vitellogenin and were thus identified as female- and male-produced vitellin, respectively. The polypeptide compositions of female- and male-produced vitellins (Fig. 5) were characteristically similar to the respect- ive precursor (Fig. 4). The female vitellin of adults and last-instar nymphs contained four major polypeptides of molecular weights of 112, 92, 90 and 54 kDa. The male vitellin of adults and last-instar nymphs contained the 92 and 54 kDa polypeptides in similar quantities, but less of the 112 and 90 kDa polypeptides. The male vitellin contained an ad- ditional 85 kDa polypeptide as well as several small polypeptides (Fig. 5).
Pro tease digestion of “P-labelled vitellogenin polypeptides
All of the polypeptides of the female- and male-produced vitellogenins were phosphorylated [Fig. 6(A)]. These 32P-labelled vitellogenin poly- peptides were further analysed by protease digestion with S. aureus V-8 protease. The fragments were separated by 15% SDS-PAGE and the pattern visu- alized by autoradiography. The fragment compo- sitions of corresponding polypeptides of the two vitellogenins were similar [Fig. 6(B)]. However, the 92
878 GRACE DON-WHEELER and FRANZ ENGELMANN
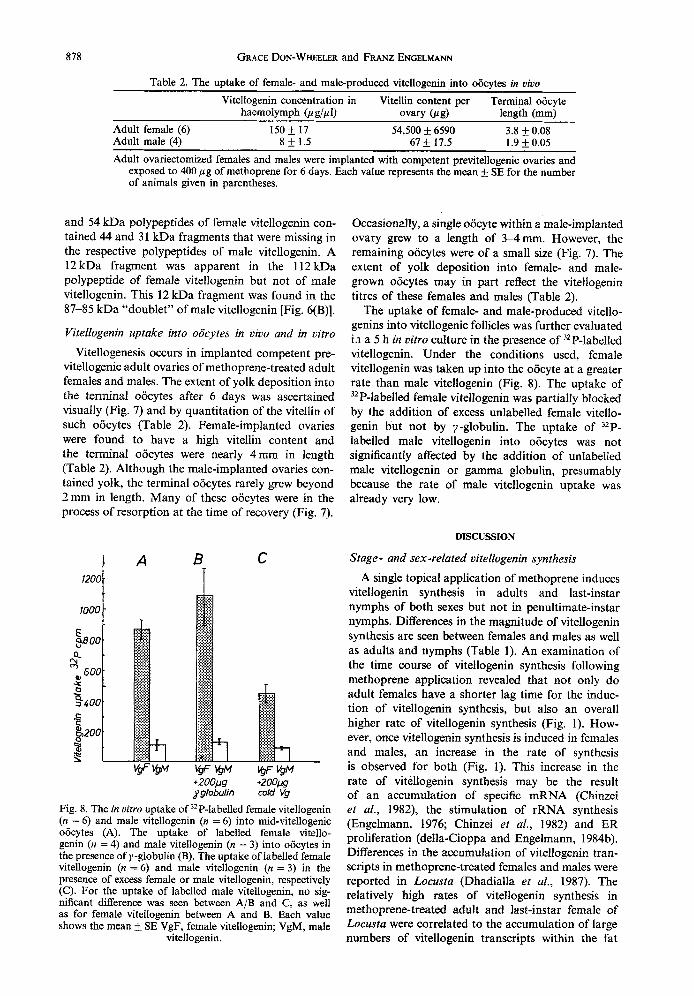
Table 2. The uptake of female- and male-produced vitellogenin into obcytes in vivo
Vitellogenin concentration in Vitellin content per Terminal oocyte haemolymph &g/PI) ovary (rcn) length (mm)
Adult female (6) 150 + 17 54,500 rt 6590 3.8 rf: 0.08 Adult male (4) 8+ 1.5 67 + 17.5 1.9 f 0.05
Adult ovariectomized females and males were implanted with competent previtellogenic ovaries and exposed to 400 pg of methoprene for 6 days. Each value represents the mean rt: SE for the number of animals given in parentheses.
and 54 kDa polypeptides of female vitellogenin con- tained 44 and 31 kDa fragments that were missing in the respective polypeptides of male vitellogenin. A 12 kDa fragment was apparent in the 112 kDa polypeptide of female vitellogenin but not of male vitellogenin. This 12 kDa fragment was found in the 87-85 kDa “doublet” of male vitellogenin [Fig. 6(B)].
Vitellogenin uptake into oBcytes in vivo and in vitro
Vitellogenesis occurs in implanted competent pre- vitellogenic adult ovaries of methoprene-treated adult females and males. The extent of yolk deposition into the terminal oiicytes after 6 days was ascertained visually (Fig. 7) and by quantitation of the vitellin of such oiicytes (Table 2). Female-implanted ovaries were found to have a high vitellin content and the terminal oiicytes were nearIy 4mm in length (Table 2). Although the male-implanted ovaries con- tained yolk, the terminal oiicytes rarely grew beyond 2 mm in length. Many of these oiicytes were in the process of resorption at the time of recovery (Fig. 7).
1200
1000 -
E poo’
4- n, 600. I+ _
5400.
c
+2oopg +zoo/lg ~globulin cold Vg
Fig. 8. The in vitro uptake of 32P-labelled female vitellogenin (n = 6) and male vitellogenin (n = 6) into mid-vitellogenic oiicytes (A). The uptake of labelled female vitello- genin (n = 4) and male vitellogenin (n = 3) into oiicytes in the presence of y-globulin (B). The uptake of labelled female vitellogenin (n = 6) and male vitellogenin (n = 3) in the presence of excess female or male vitellogenin, respectively (C). For the uptake of labelled male vitellogenin, no sig- nificant difference was seen between A/B and C, as well as for female vitellogenin between A and B. Each value shows the mean + SE VgF, female vitellogenin; VgM, male
vitellogenin.
Occasionally, a single oiicyte within a male-implanted ovary grew to a length of 3-4mm. However, the remaining oijcytes were of a small size (Fig. 7). The extent of yolk deposition into female- and male- grown oiicytes may in part reflect the vitellogenin titres of these females and males (Table 2).
The uptake of female- and male-produced vitello- genins into vitellogenic follicles was further evaluated i.1 a 5 h in vitro culture in the presence of “P-labelled vitellogenin. Under the conditions used, female vitellogenin was taken up into the oijcyte at a greater rate than male vitellogenin (Fig. 8). The uptake of “P-1abelled female vitellogenin was partially blocked by the addition of excess unlabelled female vitello- genin but not by y-globulin. The uptake of 32P- labelled male vitellogenin into oijcytes was not significantly affected male vitellogenin or because the rate of already very low.
by the addition _ of unlabelled gamma globulin, presumably male vitellogenin uptake was
DISCUSSION
Stage- and sex-related vitellogenin synthesis
A single topical application of methoprene induces vitellogenin synthesis in adults and last-instar nymphs of both sexes but not in penultimate-instar nymphs. Differences in the magnitude of vitellogenin synthesis are seen between females and males as well as adults and nymphs (Table 1). An examination of the time course of vitellogenin synthesis following methoprene application revealed that not only do adult females have a shorter lag time for the induc- tion of vitellogenin synthesis, but also an overall higher rate of vitellogenin synthesis (Fig. 1). How- ever, once vitellogenin synthesis is induced in females and males, an increase in the rate of synthesis is observed for both (Fig. 1). This increase in the rate of vitellogenin synthesis may be the result of an accumulation of specific mRNA (Chinzei et al., 1982), the stimulation of rRNA synthesis (Engelmann, 1976; Chinzei et al., 1982) and ER proliferation (della-Cioppa and Engelmann, 1984b). Differences in the accumulation of vitellogenin tran- scripts in methoprene-treated females and males were reported in Locusta (Dhadialla er al., 1987). The relatively high rates of vitellogenin synthesis in methoprene-treated adult and last-instar female of Locusta were correlated to the accumulation of large numbers of vitellogenin transcripts within the fat

Female- and male-produced vitellogenins 879
body of these females, whereas, induced last-instar males produced only small amounts of vitellogenin and ‘a correspondingly small number of transcripts was found. The sex-specific differences in the rate of vitellogenin synthesis in Leucophaea may likewise reflect differences in the level of vitellogenin gene transcription in females and males although this still needs to be shown.
We considered the possibility that vitellogenin secretion in males is slower than that in females and consequently larger quantities of vitellogenin would accumulate within the male fat body. However, when fat body from methoprene-treated females and males were pulse-labelled in vitro, a greater quantity of radiolabelled vitellogenin was found in the fat body homogenates of females than of males (data not shown). We conclude that the rate of vitellogenin synthesis of the female fat body is indeed greater than that of males.
Juvenile hormone III-induced vitellogenin syn- thesis in adult females of Leucophaea was found to peak on the third day after treatment and declined afterwards (della-Cioppa and Engelmann, 1984b), whereas a decline in methoprene-induced vitellogenin synthesis was not observed during a 6-day period (Fig. 1). The high dose of methoprene used and its stability may explain why a high rate of vitellogenin synthesis is sustained for several days. The extra- ordinarily high titre of vitellogenin found in the haemolymph of these females (Fig. 2) is due to the removal of the ovaries which normally drain vitello- genin from the haemolymph.
Penultimate-instar nymphs of Leucophaea could not be induced to synthesize vitellogenin, whereas both sexes of last-instar nymphs were inducible (Table 1). The lack of vitellogenic competency in the penultimate-instar nymphs of Leucophaea is in con- trast to reports for Blczttelh (Kunkel, 1981) and Locusta (Dhadialla and Wyatt, 1983). Blattella and Locusta penultimate-instar nymphs produce only low and variable amounts of vitellogenin, and in Bluttellu, the inducibility of vitellogenin synthesis in the penul- timate-instar nymphs develops with age within the instar. In Leucophaea, the competency to produce vitellogenin appears to develop later than in Blattella and Locusta.
Once vitellogenic competency is established in last instar nymphs of Leucophuea, females synthesize more vitellogenin than either last-instar or adult males (Table 1). Like Locusta (Wyatt et al., 1984), the development of maximal vitellogenic competency in females of Leucophuea is attained after the adult emergence. Vitellogenic competency in males was not greatly affected by the adult moult, i.e. an increase in vitellogenic competency was not as evident in males as it was in females (Table 1). In contrast to Locusta, males of Leucophaea do not lose their vitellogenic competency after emergence.
Adult females of Leucophuea have a greater haemolymph volume than either adult males or last-
instar nymphs, and therefore, any injected radiolabel is more diluted in the adult female than in adult males or nymphs. In spite of this dilution, the vitellogenin content of the haemolymph of adult ovariectomized females was greater and more highly labelled than either in adult males or last-instar nymphs (Table 1). This shows that adult females have indeed a higher degree of vitellogenic competency than adult males or last-instar nymphs.
Although the fat body of last-instar females has the capacity to synthesize substantial amounts of vitello- genin, large doses of methoprene are required. The stage-dependent vitellogenic response of the female fat body is similar to the stage-dependent response of the fat body to adipokinetic hormone in Locusta (Van der Horst et al., 1989). The degree of adipokinetic hormone-induced lipid mobilization in the fat body of azadirachtin-produced overage nymphs is less than that of adults. While these data suggest that the adult moult is not required for the development of com- petency in the case of juvenile hormone-induced vitellogenin synthesis or adipokinetic hormone- induced lipid mobilization, maximal competency is achieved only after emergence in either case. Perhaps a period of low juvenile hormone titres coupled with ecdysone exposure is required for the metamorphic change of nymphal female fat body into a state of full vitellogenic competency (Wyatt et al., 1984).
Female- and male -produced vitellogenins
The apparent molecular weights of female vitello- genin (Fig. 3) are in agreement with what has been previously reported for Leucophueu (Engelmann, 1979). The 270 kDa band represents the vitellogenin monomer while the 540 kDa band corresponds to the vitellogenin homodimer. The male-produced vitello- genin is virtually identical to the female. The nature of the 210 and 180 kDa bands found in male vitello- genin is unclear; these may be either degradation products or contaminants.
The polypeptide compositions of the female- and male-produced vitellogenin were similar but with quantitative differences in some of the polypeptides (Fig. 4). The origin of the 87-85 kDa polypeptide “doublet” of male vitellogenin is unclear. It is striking that the male vitellogenin contains quantitatively less of the 112 and 97 kDa polypeptides and it is possible that this 87-85 kDa “doublet” is related to one of these polypeptides.
All of the polypeptides of the two vitellogenins were phosphorylated [Fig. 6(A)]. The 112 and 97 kDa polypeptides of male vitellogenin gave a markedly stronger signal on 32P autoradiograms than on 35S autoradiograms. The difference in signal intensity may be due to quantitative differences in the incor- poration of 35S- and 32P-label into the 112 and 97 kDa polypeptides of male vitellogenin.
The protease digestion of 32P-labelled polypeptides of female and male vitellogenins revealed both similarities and differences in the composition of

880 GRACE DON-WHEELER and FRANZ ENGELMANN
fragments of the respective polypeptides [Fig. 6(B)]. The differences in the compositions suggest differ- ences in the phosphorylation and processing of vitellogenin in the female and male fat body. The 12 kDa fragment which is present in the 112 kDa polypeptide of female vitellogenin but absent from that of male vitellogenin, may in fact be a contami- nant from the 97 kDa polypeptide. The 112 and 97 kDa polypeptides are not completely resolvable by 7.5% SDS-PAGE. In support of this idea, we have found that V-8 protease digestion of 112 kDa polypeptides of 32P-labelled female and male vitellin did not produce a 12 kDa fragment (data not shown). Contamination of the 112 kDa polypeptide of vitellin would not be a factor since these vitellins contain small quantities of the 97 kDa polypeptide (Fig. 5). The presence of the 12 kDa fragment in the 97 kDa polypeptide needs to be confirmed, however. Finally, the 87-85 kDa “doublet” of male vitellogenin has in common some of the fragments found in the 112 and 92 kDa polypeptides. This suggests that the “doublet” is derived from one of the larger vitello- genin polypeptides.
The underlying reasons for the different poly- peptide compositions of female- and male-produced vitellogenins are as yet unclear. Both female and male vitellogenins of Leucophaea appear to be synthesized as 210 kDa phosphorylated precursors (data not shown). In females, the pro-vitellogenin is processed to yield four major vitellogenin polypeptides (Fig. 4) which are assembled into the mature protein (della- Cioppa and Engelmann, 1987). It is possible that differences in the glycosylation of the vitellogenin precursor in the female and male fat body could lead to differences in proteolytic processing and further post-translational modifications of vitellogenin. For example, in Blattella (Wojchowski et al., 1986) and in Aedes aegypti (Dhadialla and Raikhel, 1990), it has been shown that the glycosylation of vitellogenin occurs co-translationally and that the disruption of glycosylation with tunicamycin profoundly affects vitellogenin processing. In Blattella, tunicamycin treatment disrupted the proteolytic processing and post-translational modification of the vitellogenin precursor. In Aedes, tunicamycin-treated animals produced an aglycosylated vitellogenin precursor which was not phosphorylated, but was proteo- lytically processed. However, these aglycosylated vitellogenin subunits did not undergo further modifi- cations and subsequently could not be chased into mature subunits (Dhadialla and Raikhel, 1990). The role of glycosylation in the synthesis and processing of the vitellogenin of Leucophaea will be reported later.
Female - and male -produced vitellins
Female-produced vitellogenin and vitellin of Leucophaea were found to be similar in their poly- peptide compositions (Figs 4 and 5). However, vitellin contained quantitatively less of the 97 kDa
polypeptide than vitellogenin and an additional 90 kDa polypeptide. It is possible that the 97 kDa polypeptide of the vitellogenin is further processed within the oiicyte to yield the 90 kDa polypeptide of vitellin (Engelmann, 1979).
The polypeptide composition of male vitellin was characteristically similar to the corresponding male vitellogenin (Figs 4 and 5). Although the male vitello- genin is different from the female vitellogenin, the male vitellogenin is taken up by the oocyte and processed. The additional low molecular weight polypeptides found in male vitellin may be due to proteolytic degradation which could have occurred within the oiicyte or during the isolation procedure. However, female vitellin isolated in an identical man- ner did not contain these additional polypeptides. The female vitellin may be more stable than the male vitellin.
The uptake of female- and male-produced vitellogenin into the oiicyte
In Leucophaea, vitellogenic oijcytes of methoprene- treated males rarely grew beyond 2 mm whereas oiicytes implanted into methoprene-treated adult ovariectomized females grew to nearly full size within a 6-day period (Fig. 7). The accelerated growth of oiicytes in these females may be due to the high vitellogenin titre of these hosts (Table 2). The rela- tively low vitellogenin titre of males (8 pg/pl) may in part explain why males produced only small yolky oiicytes within the observed period. However, this does not explain why these oijcytes fail to complete vitellogenesis since normal mid-vitellogenic females of Leucophaea have a vitellogenin titre of only 2-3 pg/pl (Engelmann, 1978) and normal vitello- genesis is completed within a 3-week period. Differ- ences in the structure of the female and male vitellogenin, or factors which are either present or missing in the female and male milieu may affect the in vivo uptake of vitellogenin into the compotent oiicyte.
For some insects, it has been shown that vitello- genin is taken up into the oiicyte by receptor- mediated endocytosis (Rdhrkasten and Ferenz, 1985) and that the structure of vitellogenin is important for its recognition and uptake. For example, chemically modified vitellogenin of Locusta is no longer taken up into the oocyte (Rohrkasten and Ferenz, 1986). In Blattella, vitellin which has been deglycosylated is not taken up into oiicytes as well as intact vitellin (Gochoco et al., 1988). For Leucophaea, it is shown that the female vitellogenin is taken up into the oocyte at a greater rate than male vitellogenin (Fig. 8). The uptake of labelled female vitellogenin was significantly lowered in the presence of unlabelled female vitellogenin but not of y-globulin. Under the given assay conditions, the uptake of labelled male vitellogenin into oiicytes was not affected by y-globulin and only slightly inhibited (not sig- nificantly) by excess unlabelled male vitellogenin.

Female- and male-produced vitellogenins 881
In a separate experiment, labelled male vitellogenin of twice the specific activity was used (data not shown). Although the uptake of labelled male vitellogenin into 06cytes was greater, it was still significantly lower than the uptake of labelled female vitellogenin. Since male vitellogenin is taken up at a very low rate, the system was perhaps not sensitive enough to detect significant competition with unlabelled male vitellogenin.
For this in vitro assay, we used haemolymph that contained 32P-labelled vitellogenin. Lipophorin is a major phospholipoprotein of the haemolymph of females and males. However, vitellogenic females and males of Leucophaea synthesize lipophorin at a very low rate relative to vitellogenin (unpublished data). For example, the lipophorin synthesized in vitello- genie females during a 1 h 32P pulse contained only 1% of the total incorporated 32P, whereas 98% of the 32P was associated with vitellogenin (Engelmann and Friedel, 1974). Vitellogenic males have an even lower rate of lipophorin synthesis than females (data not shown). In addition, we know that the lipophorin content of mid-vitellogenic oijcytes of Leucophaea is only 1% of the total protein (unpublished data). These data suggest that the contribution of 32P-labelled lipophorin to the total radioactivity recovered from the cultured oijcyte is negligible.
In summary, our data demonstrate that once vitellogenic competency is established in Leucophaea, sexual differences in vitellogenin synthesis become apparent. Rates of vitellogenin synthesis in adults and nymphs may be influenced by the rate of vitello- genin gene transcription, or at the level of translation and secretion. The rate of transcription could be affected by the structure of the vitellogenin gene, the copy number of vitellogenin genes, or by the presence or absence of factors that normally regulate the stage- and sex-specific expression of the vitellogenin gene. Sex affects not only the quantity of vitellogenin produced, but also the quality of the vitellogenin. Possible differences in the quantity and composition of processing enzymes in the female and male fat body could ultimately affect the structure of the vitellogenin produced in females and males. Further investigations into the proteolytic processing and post-translational modification of vitellogenin in females and males may give us further insight as to the nature of these vitellogenins.
Acknowledgements-The research reported here has been supported in part by a grant from the National Science Foundation (DCB 86-02887), a Biomedical Science Support Grant, a University Research Grant, and a Sigma Xi Grant. We thank Dr E. C. Mundall for reading a draft of this paper and for making helpful suggestions for improvement.
REFERENCES
Bonner W. M. and Laskey R. A. (1974) A film detection method for tritium-labelled proteins and nucleic acids in polyacrylamide gels. Eur. J. Biochem. 46, 83-88.
Burnette W. N. (1981) Western blotting: electrophoretic transfer of protein from sodium dodecyl sulfate- polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Analyt. Biochem. 112, 195-203.
Chinzei Y., White B. N. and Wyatt G. R. (1982) Vitello- genin mRNA in locust fat body: identification, isolation, and quantitative changes induced by juvenile hormone. Can. J. Biochem. 60, 243-251.
Cleveland D. W., Fischer S. G., Kirschner M. W. and Laemmli U. K. (1977) Peptide mapping by limited proteolysis in sodium dodecyl sulfate and analysis by gel electrophoresis. J. biol. Chem. 252, 1102-l 106.
della-Cioppa G. and Engelmann F. (1984a) Phospholipid synthesis in the fat body endoplasmic reticulum during primary and secondary juvenile hormone stimulation of vitellogenesis in Leucophaea maderae. Roux’s Arch. Dev. Biol. 193, 78-85.
della-Cioppa G. and Engelmann F. (1984b) Juvenile hormone regulation of phospholipid synthesis in the endoplasmic reticulum of vitellogenic fat body cells from Leucophaea maderae. Insect Biochem. 14, 27-36.
della-Cioppa G. and Engelmann F. (1987) The vitellogenin of Leucophaea maderae: synthesis as a large phosphory- lated precursor. Insect Biochem. 17, 401-415.
Davis B. J. (1964) Disc electrophoresis-II. Method and application to human serum proteins. Ann. N.Y. Acad. Sci. 121, 404-427.
Dhadialla T. S. and Raikhel A. S. (1990) Biosynthesis of mosquito vitellogenin. J. biol. Chem. 265, 9924-9933.
Dhadialla T. S. and Wyatt G. R. (1983) Juvenile hormone- dependent vitellogenin synthesis in Locusta migratoria fat body: inducibility related to sex and stage. Dev. Biol. 96, 436444.
Dhadialla T. S., Cook K. E. and Wyatt G. R. (1987) Vitellogenin mRNA in locust fat bodv: coordinate induc- tion ofiwo genes by a juvenile hormone analog. Dev. Biol. 123, 108-l 14.
Don-Wheeler G. and Engelmann F. (1989) Vitellogenin synthesis in the female and male cockroach, Leucophaea maderae. Am. Zool. 29, 49A.
Engelmann F. (1976) Induction of the insect vitello- genin in vivo and in vitro. In The Juvenile Hormones (Ed. Gilbert L. I.), pp. 470-485. Plenum Press, New York.
Engelmann F. (1978) Synthesis of vitellogenin after long- term ovariectomy in a cockroach. Insect Biochem. 8, 149-154.
Engelmann F. (1979) Insect vitellogenin: identification, biosynthesis, and the role in vitellogenesis. Adv. Insect Phy&o/. 14, 4%108.
Engelmann F. and Friedel T. (1974) Insect volk urotein precursor, a juvenile hormone induced phoipho&otein. Life Sci. 14, 587-594.
Engelmann F., Friedel T. and Ladduwahetty M. (1976) The native vitellogenin of the cockroach Leucophaea maderae. Insect Biochem. 6, 21 l-220.
Garvey J. S., Cremer N. E. and Sussdorf D. H. (1977) Methods in Immunology, 3rd edn. Benjamin/Cummings, Reading, Mass.
Gochoco C. H., Kunkel J. G. and Nordin J. H. (1988) Experimental modifications of an insect vitellin affect its structure and its uptake by oocytes. Archs Insect Biochem. Physiol. 9, 179-199.
Hedrick J. L. and Smith A. J. (1968) Size and charge isomer separation and estimation of molecular weights of pro- teins by disc gel electrophoresis. Archs Biochem. Biophys. 126, 155-164.
Kunkel J. G. (1981) A minimal model of metamorphosis: fat body competence to respond to juvenile hormone. In Current Topics in Insect Endocrinology and Nutrition (Eds Bhaskaran G., Friedman M. and Rodriquez J.), pp. 107-128. Plenum Press, New York.

882 GRACE DON-WHEELER and FRANZ ENGELMANN
Laemmli U. K. (1970) Cleavage of structural proteins Van der Horst D. J., Van Doom J. M., Pener M. P., Van during the assembly of the head of the bacteriophage T4. den Broek A. Th. M., Van Marrewijk W. J. A. and Nature, Lond. 227, 680-685. Beenakkers A. M. Th. (1989) Development of imaginal
Lamy M. (1984) Vitellogenesis, vitellogenin and vitellin in competence to adipokinetic hormone in Locusta: males of insects: a review. Int. J. Invert. Reprod. Dev. 7, lipophorin conversions in precocene-induced adultiforms 311-321. and in azadirachtin-induced overage nymphs. Comp. Bio -
Lanzrein B. (1974) Influence of a juvenile hormone analogue them. Physiol. 92B, 133-136. on vitellogenin synthesis and oogenesis in larvae of Van Keuren M. L., Goldman D. and Merril C. R. (1981) Nauphoeta cinerea. J. Insect Physioi. 20, 1871-1885. Detection of radioactively labeled proteins is quenched by
Monthony J. F., Wallace E. G. and Allen D. M. (1978) silver staining methods: quenching is minimal for i4C and A non-barbital buffer for the immunoelectrophoresis and partially reversible for 3H with a photochemical stain. zone electrophoresis in agarose gels. Clin. Chem. 24, Analyt. Biochem. 116, 248-255. 1825-1827. Woichowski D. M.. Parsons P.. Nordin J. H. and Kunkel
Mundall E. C., Tobe S. S. and Stay B. (1979) Induction of vitellogenin and growth of implanted oocytes in male cockroaches. Nature, Land. 282, 97-98.
Rijhrkasten A. and Ferenz H.-J. (1985) In vitro study of selective endocytosis of vitellogenin by locust oocytes. Roux’s Arch. Dev. Biol. 194, 411-416.
Riihrkasten A. and Ferenz H.-J. (1986) Solubilization of the vitellogenin receptor. Biochim. Biophys. Acta 860, 577-582.
J: G. (1986) Processing of pro-vitellogenin in insect fat body: a role for high mannose oligosaccharide. Dev. Biol. 116, 422-430.
Wyatt G. R., Dhadialla T. S. and Roberts P. E. (1984) Vitellogenin synthesis in the locust fat body: juvenile hormone-stimulated gene expression. In Biosynthesis, Metabolism and Mode of Action of Invertebrate Hormones (Eds Hoffman J. and Porchet M.), pp. 475484. Springer, Berlin.
Copyright © 2022 FDOKUMEN




















