
The feeling of movement: EEG evidence for mirroring activity during the observations of static,...
10
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of The feeling of movement: EEG evidence for mirroring activity during the observations of static,...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Biological Psychology 85 (2010) 233–241
Contents lists available at ScienceDirect
Biological Psychology
journa l homepage: www.e lsev ier .com/ locate /b iopsycho
The feeling of movement: EEG evidence for mirroring activity during theobservations of static, ambiguous stimuli in the Rorschach cards
Luciano Girominia, Piero Porcelli b, Donald J. Viglionec, Laura Parolina, Jaime A. Pinedad,∗
a Department of Psychology, University of Milano-Bicocca, Italyb Psychosomatic Unit, IRCCS De Bellis Hospital, Italyc California School of Professional Psychology, Alliant International University, San Diego, CA 92131, USAd Department of Cognitive Science, University of California, San Diego, La Jolla, CA 92037-0515, USA
a r t i c l e i n f o
Article history:Received 6 March 2010Accepted 11 July 2010Available online 21 July 2010
Keywords:Mirror neuronsAction observationEmpathyRorschach
a b s t r a c t
The mirror neuron system (MNS) is considered the best explanation for the neural basis of embodiedsimulation. To date no study has investigated if it may be activated not only by actual but by the “feelingof movement”. The Rorschach test cards were used to investigate evidence of EEG mu wave suppression atcentral areas, an index of MNS activity, since human movement responses (M) to the Rorschach elicit suchfeelings of movement. Nineteen healthy volunteers observed different sets of Rorschach stimuli duringattribution, identification, and observation of human movements and different scenarios while their EEGwere recorded. Significant mu suppression occurred when subjects perceived movement, regardless ofthe experimental condition. These results show that mirroring can be activated by static, ambiguousstimuli such as Rorschach cards, suggesting that internal representation of the “feeling of movement”may be sufficient to trigger MNS activity even when minimal external cues are present.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Social cognition is a broad concept that refers to the cogni-tive and brain processes that subserve behavior in response toconspecifics (i.e., other individuals of the same species) (Adolphs,1999). There is extensive theoretical discussion concerning the roleof the Mirror Neuron System (MNS) in social cognition (Agnew etal., 2007; Gallese and Goldman, 1998; Iacoboni and Dapretto, 2006;Rizzolatti and Craighero, 2004; Williams et al., 2001), and empiricalevidence to date supports this link. For example, MNS activationis reduced among clinical populations for which social cognitivedeficits represent a core feature (e.g., autism) (Hadijkhani et al.,2006; Oberman et al., 2005). Crucially, a number of authors haverecently begun to investigate in healthy populations the proposedlink between the MNS and social cognitive functions such as empa-thy (Gazzola et al., 2006; Kaplan and Iacoboni, 2006; Pfeifer et al.,2008) and facial emotion processing (Enticott et al., 2008; Sato andYoshikawa, 2007; Wicker et al., 2003). The premotor MNS areasappear to be recruited not only for understanding the purpose orgoal of an action, but also in the interpretation of the intention andthe meaning of an action. It is not surprising, therefore, that the MNShas been implicated in disorders, such as autism spectrum disor-
∗ Corresponding author. Tel.: +1 858 534 9754; fax: +1 858 534 1128.E-mail address: [email protected] (J.A. Pineda).
ders and schizophrenia, in which social cognition deficits are corefeatures (Buccino and Amore, 2008; Dapretto et al., 2005; Obermanet al., 2005).
1.1. The mirror neuron system
Mirror neurons are cortical brain cells that fire during both theexecution and the observation of action and were discovered dur-ing single cell recording in the ventral premotor cortex (F5) ofmacaque monkeys that either performed an action or observed thesame action performed by another monkey or an experimenter (diPellegrino et al., 1992; Gallese et al., 1996; Rizzolatti et al., 1996).Single-unit recording is not typically performed in the humanbrain (although see Mukamel et al., 2010). Nonetheless, indirectpopulation-level measures support the existence of a functionalanalogous system to macaque MNS in the human inferior frontalgyrus (IFG) (Fadiga et al., 1995; Iacoboni et al., 1999) through tran-scranial magnetic stimulation (TMS) (Fadiga et al., 1995; Maeda etal., 2002; Strafella and Paus, 2000), positron emission tomography(PET) (Parsons et al., 1995), functional magnetic resonance imaging(fMRI) (Buccino et al., 2004; Grézes et al., 2003; Iacoboni et al., 1999;Rizzolatti et al., 1996), and electroencephalography (EEG) (Cochinet al., 1998; Muthukumaraswamy and Johnson, 2004; Oberman etal., 2007a,b; Pineda et al., 2000; Pizzamiglio et al., 2005; Rizzolattiet al., 2001). Further, data are consistent with the idea that thefrontal MNS in humans may be part of a broader network of brainregions including the inferior parietal lobule (Buccino et al., 2001;
0301-0511/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.biopsycho.2010.07.008

Author's personal copy
234 L. Giromini et al. / Biological Psychology 85 (2010) 233–241
Parsons et al., 1995), the superior temporal sulcus (Iacoboni et al.,2001), sensorimotor cortex (Pineda, 2008), and regions of the lim-bic system (Morrison et al., 2004; Singer et al., 2004; Wicker etal., 2003). This broader network suggests that the MNS may play amore sophisticated role than pure imitation. It is speculated that thehuman MNS represents not only the physical aspects of an actionbut also the underlying intentions, thoughts, and feelings that moti-vated that action, therefore providing the neural basis for uniquehuman social skills such as empathy, theory of mind, and facialemotion processing (Gallese, 2001, 2006; Gallese and Goldman,1998; Pelphrey and Morris, 2006; Uddin et al., 2007) that alloweffective social cognitive processes and interactions.
Previous studies (Cochin et al., 1998; Oberman et al., 2005,2007a,b; Pineda et al., 2000) have linked activity in the human MNSwith activity in the EEG mu frequency band recorded over senso-rimotor cortex. At rest, these sensorimotor neurons spontaneouslyfire in synchrony (Gastaut, 1952), leading to large amplitude EEGoscillations in the 8–13 Hz (mu) frequency band. When subjectsperform an action, these neurons fire asynchronously, reflectinggreater levels of active processing during motor movement andobservation and thereby decreasing the power of the mu-bandEEG oscillations (Pfurtscheller et al., 1997). Over the past 50 yearsthere have been several theories relevant to the function of themu rhythm (for a review, see Pineda, 2005). Most recently, resultsof several studies have uncovered various properties of mu sup-pression that directly link it to the frontal mirror neuron system.First, mu power recorded from electrodes at scalp locations C3 andC4 is reduced by self-initiated movement and observed movement(Babiloni et al., 1999; Cochin et al., 1998; Gastaut, 1952; Oztop andArbib, 2002). Importantly, similar to mirror neuron activity, the muwave does not respond to nonbiological directional motion such asbouncing balls (Oberman et al., 2005). Furthermore, analogous toprevious fMRI studies of the MNS (Buccino et al., 2001), the pres-ence of a target object increases mu wave suppression as comparedto pantomimed actions (Muthukumaraswamy and Johnson, 2004).
Since the mu rhythm is generated by activity in sensorimotorareas (Gastaut, 1952), and mirror neurons are located in premo-tor cortex, it has been hypothesized that the mu rhythm mayspecifically index downstream modulation of primary sensorimo-tor processing by mirror neuron activity (Muthukumaraswamy andJohnson, 2004; Pineda, 2005).
An intriguing issue related to MNS activity is the role played bythe nature of the perceptual stimuli. The visual feature that acti-vates mirror neurons is the observation of a significant interactionbetween the agent of the action (e.g., seeing a hand grasping) andthe object being the target of it (e.g., a tea cup). Mirror neuronstypically do not respond to the observation either of a hand merelymiming an action or an object alone, even when the object is ofinterest (e.g., food for a monkey). In other words, mirror neurons arepart of a neural matching system that allows the observer, duringaction observation, to place her or himself in the same “internal”situation as when actively executing the same action. An impor-tant experiment showed that the MNS in monkeys responds whenthe final part of an action, most crucial in triggering the responsein full vision, was hidden to the observer, i.e., when the inten-tion of the hand gesture could only be inferred (Umiltà et al.,2001). Similar results were found when the actual stimuli were notmoving but rather static images from which dynamic informationwere extracted (implied motion). For example, the medial tempo-ral/medial superior temporal cortex (MT/MST complex) is thoughtto activate during the visual experience of real (as in movies) or illu-sory motion, namely when the observer can infer the position of anobject in a subsequent moment in time without seeing the actualmovement, such as seeing two photographs of an athlete beforeand after heaving the shot put (Kourtzi and Kanwisher, 2000). Sim-ilarly, a TMS study indicated that the MNS is responsive when
dynamic information about body actions is inferred from static pic-tures of body postures (photographs of pincer grips) (Urgesi et al.,2006).
Accordingly, the MNS would be expected to respond whenactions are mostly generated “internally”, within the individual’sexperience (feeling of motion), and triggered by minimal indirectcues placed “externally”, in the actual static picture itself. To ourknowledge, no empirical study to date has investigated this hypoth-esis.
1.2. Rorschach test stimuli
The inkblot-based water-color stimuli, as developed byRorschach (1921), provide an excellent opportunity to studywhether mirror neurons are activated when subjects “project” thesense of movement during the observation of ambiguous staticimages (Exner, 2003). The 10 Rorschach inkblot cards are composedof ambiguous and contradictory but evocative stimuli and subjectsare instructed to produce suitable representations to interpret thestimuli. The request is: “What might this be?”, and each response,freely given by the respondent, is scored according to the distinctperceptual classes of features that are selected by the subject asrelevant for identifying a specific representation for the stimulus.These features are referred to as “determinants” and include form(contour) of the blot (e.g., “it looks like a person because of theshape”), the color (e.g., “the shape and the color make it look like aperson wearing a red hat”), and shading (e.g., “a person surroundedby smoke because of the shading there”). A fourth characteristicor determinant of the response is the movement, i.e., the subjectidentifies an object not only because of form, color, and/or shadingof the stimulus but also because he or she “experiences” the sen-sation that the object (a human being, an animal, or an inanimateobject) is moving. These movement responses are the type of deter-minant that is not contained in the actual Rorschach stimulus, butadded to the stimulus field as a product of ideational and imaginalactivity of the subject. Responses characterized by the perception ofhuman movement (e.g., “two people dancing together”) are scoredfor human movement, or M, in the Rorschach Comprehensive Sys-tem (RCS) (Exner, 2003).
Based on historical interpretation of human movement andempirical findings, the M responses are considered indices of highercognitive functioning (because M involves the integration of differ-ent perceptual features and individual psychological involvement),ability to imagine (because the actual stimuli do not move), andempathy (because of the implied ability to identify a human being)(Exner, 2003). They are strongly linked to the individual’s abilityto evoke an internal experience of movement from an ambiguousstimulus (Bendick and Klopfer, 1964; Ferracuti et al., 1999; Gallucci,1989; Orlinsky, 1966; Porcelli and Meyer, 2002; Rhue and Lynn,1987; Steele and Kahn, 1969; Wood et al., 2003; Young and Wagner,1993) and to involve this experience of movement in the perceptualprocessing of static pictures. The psychological process eliciting Mresponses resembles theoretical constructs such as Einfühlung (aGerman term meaning literally “feeling-in” which has been used inthe experience of art indicating the immediate physical responsesgenerated in the observer by the exposure to art painting), empathy(the ability to understand others’ actions, sensations, and emo-tions), and also mentalization (the cognitive mechanism that allowsone to ascribe goals and intentions to others). The human capacityto pre-rationally make sense of the actions, emotions and sensa-tions of others depends on “embodied simulation”, a functionalmechanism through which the actions, emotions, or sensations wesee activate our own internal representations of the body statesthat are associated with these social stimuli, as if we were engagedin a similar action or experiencing a similar emotion or sensation(Freeberg and Gallese, 2007). Based on the MNS process, Gallese

Author's personal copy
L. Giromini et al. / Biological Psychology 85 (2010) 233–241 235
speculated that this “shared manifold of intersubjectivity” allowsus to recognize other human beings who are similar to us (Gallese,2003).
If the link between the M responses to the Rorschach stimuli andembodied simulation, as well as the link between embodied simu-lation and the MNS hold true, then Rorschach-based, self-initiatedM responses may be associated with the activation of mirror neu-rons.
1.3. Aim of the study
The purpose of the present study was to characterize specificproperties of the stimuli that produce activation in the human MNSby studying perceptions induced only by internal representations ofhuman movement produced by static, ambiguous stimuli. Althoughto date no study has evaluated the activation of the MNS dur-ing the administration of the Rorschach test, past theoretical andempirical literature suggest that human movement responses tothe Rorschach should be associated with MNS activity. Our exper-iment determined whether the observation of static images fromthese ambiguous stimuli (i.e., static pictures with no actual move-ment) of Rorschach inkblots is sufficient to activate the MNS.
2. Materials and methods
2.1. Participants
Participants consisted of 20 undergraduate students recruited from the Psy-chology Department’s subject pool at the University of California, San Diego. Onesubject’s data were excluded from analysis due to an excess of head movementsand eye blinking (less than 1 s clean data per stimulus was available), resulting in afinal sample of 19 subjects (7 male, 12 female) ranging in age from 18 to 27 years(M = 20.58, SD = 1.98). Participants received class credit for their participation, andall gave written consent. The study was reviewed and approved by the InstitutionalReview Board of the University of California, San Diego Human Research ProtectionsProgram.
2.2. Procedure
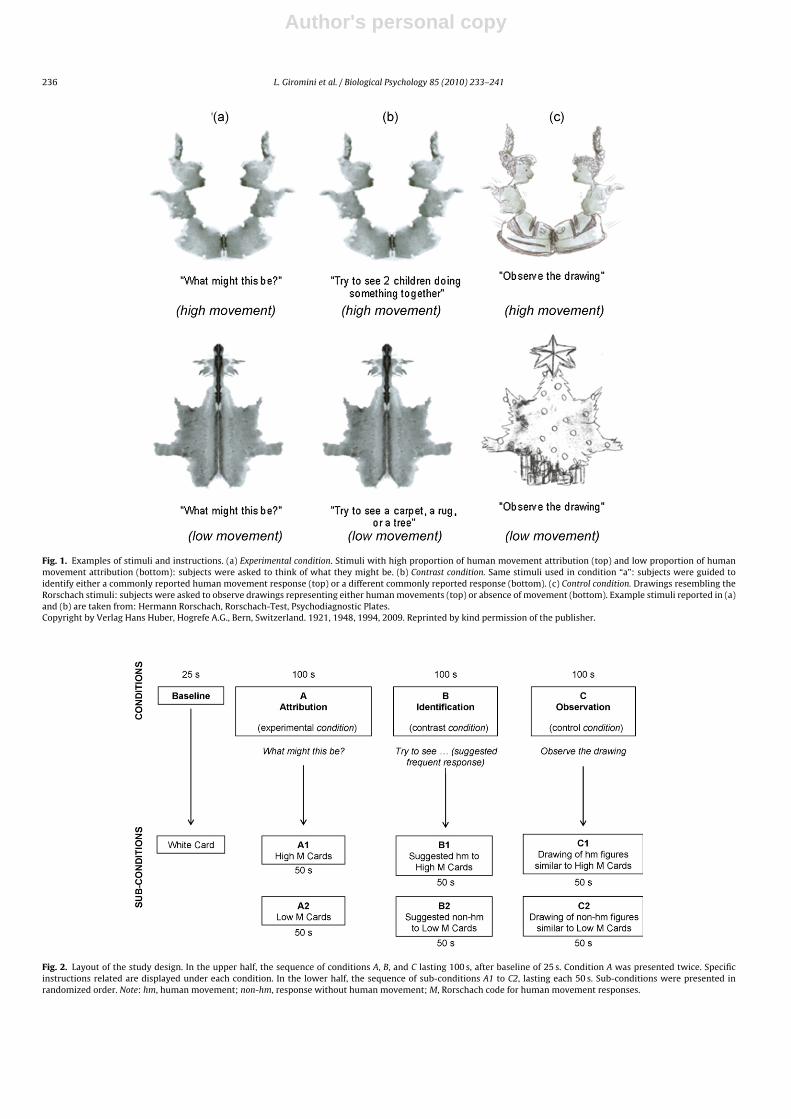
EEG data were collected during baseline and three different experimental, con-trast, and control conditions. (Baseline) This involved the participant looking at themiddle of a white card on a screen. (A) Attribution of human movement (experimen-tal condition) was evaluated by asking the subjects to look at the Rorschach stimuliwith the instruction to think of what they might be (Fig. 1a). This practice is generallyconsistent with the standard instructions of the RCS (Exner, 2003), except for thedemands of the computer and EEG recording. Four Rorschach cards were selectedfor this study, two high M cards (III and VII) with the highest frequency of humanmovement attribution (M responses) and two low M cards (V and VI) with the low-est frequency of human movement in the reference database (Exner and Erdberg,2005). Consistent with standard RCS administration, participants were asked to ver-balize what they saw after first looking at each of the cards and thinking about theirresponse.
(B) Identification of human movement (contrast condition) was evaluated by ask-ing subjects to observe the Rorschach stimuli with suggestions verbally providedby the experimenter during the visual exposure to identify a commonly reportedmovement response (Fig. 1b). The same stimuli as the previous experimental con-dition A were utilized, but in this condition subjects were guided to identify acommonly reported human movement response (e.g., “two children doing some-thing together”, “two persons arguing with each other”, “two women raising up aheavy basket”) on the two cards with the highest proportion of human movementattribution, and a different commonly reported response (e.g., “a tree”, “a totempole”, “X-ray of pelvis”) on the two cards with the lowest proportion of humanmovement attribution. Following the observation period, subjects were asked aboutthe attributions of their responses. We would expect to obtain similar results inboth the experimental and contrast conditions. (C) Observation of human movement(control condition) was evaluated by asking subjects to observe non-ambiguoushand-made drawings specifically created to resemble the Rorschach inkblots shownearlier (Fig. 1c). Two of the drawings represented a commonly reported humanmovement response to the related inkblot (such as “two children doing somethingtogether”) while the other two represented a different commonly reported response(such as “a tree”).
In summary, three main conditions (attribution – A, identification – B, and obser-vation – C), subdivided into six sub-conditions according to the presence or absenceof human movement: attribution (A1), identification (B1), and observation (C1) ofhuman movements, and attribution (A2), identification (B2), and observation (C2)
without human movements. Condition A differed from the other two in that itrequired the respondent to spontaneously attribute a movement in the card forthe corresponding EEG data to be analyzed in the movement condition (A1) andto not attribute a movement for the corresponding EEG data to be analyzed in thenon-movement condition (A2). Therefore, in order to obtain enough clean EEG data,only the experimental A condition was presented twice, instructing the subjects tothink of a different response each time a stimulus was repeated.
All stimuli were shown on a screen situated at a distance of 96 cm, at a size of17 cm × 24 cm, like the original Rorschach cards. For the experimental A and the con-trast B conditions, subjects were instructed to continue focusing on their responsefor the entire period of EGG recording. The baseline condition was presented at thebeginning and among the three other conditions, the experimental A condition waspresented first to insure that the spontaneous responses were not influenced by anyform of priming; contrast B condition was presented second so as not to allow thedrawings to facilitate identification of suggested responses; finally, control C condi-tion was presented last. The presentation order of the stimuli within each condition(i.e., stimuli with and without human movement) was randomized (Fig. 2).
2.3. EEG data acquisition and analysis
Disk electrodes were applied to the face above and below the left eye to moni-tor the electrooculogram (EOG) and behind each ear (mastoids) for use as referenceelectrodes. Data were collected from 13 electrodes embedded in a cap, at the fol-lowing scalp positions: F3, Fz, F4, C3, Cz, C4, P3, Pz, P4, T5, T6, O1, and O2, using theinternational 10–20 method of electrode placement. Following placement of the cap,electrolytic gel was applied at each electrode site and the skin surface was lightlyabraded to reduce the impedance of the electrode–skin contact. The impedanceson all electrodes were measured and confirmed to be less than 5 k� both beforeand after testing. Once the electrodes were in place, subjects were seated inside anacoustically and electromagnetically shielded testing chamber. EEG was recordedand analyzed using a Neuroscan Synamps system (band pass 0.1–30 Hz). Data werecollected for approximately 100 s per condition – around 50 s per each of the 6 sub-conditions A1, B1, C1, A2, B2, and C2 – at a sampling rate of 500 Hz. Given that thebaseline task consisted of looking at the middle of a white card, only 25 s of data wererecorded during this condition in order to avoid distraction or less engagement withthe task because of increasing boredom. Eye blinks as well as eye and head move-ments were manually identified in the EOG recording, and artifacts during theseintervals were removed prior to analysis using traditional methods (Goldensohn etal., 1999). For each cleaned segment the integrated power in the 8–13 Hz range wascomputed using a Fast Fourier Transform. Data were segmented into epochs of 1 sbeginning at the start of the segment. Fast Fourier transforms were performed onthe epoched data (256 points). A cosine window was used to control for artifactsresulting from data splicing. Data were only analyzed if there were sufficient cleandata, i.e., cards with less than 5 valid epochs were removed, and the different con-ditions (composed of the combined valid cards) were only analyzed if a minimumof 20 s were available.
Mu suppression over sensorimotor cortex (scalp locations C3, Cz and C4) wascalculated in all conditions as the ratio of the power during the sub-conditions rel-ative to the power during the baseline condition. As a common procedure for thistype of study (Altschuler et al., 2000; Bernier et al., 2007; Martineau et al., 2008;Oberman et al., 2005, 2007a,b; Pineda and Hecht, 2009; Ulloa and Pineda, 2007), aratio was used to control for variability in absolute mu power as a result of indi-vidual differences such as scalp thickness, electrode placement, and impedance, asopposed to differences in brain activity. Since ratio data are inherently non-normal,as a result of lower bounding, a log transform was used for analysis. A log ratio of lessthan zero indicates suppression, whereas a value of zero indicates no suppressionand a value greater than zero indicates enhancement. Given that one of the subjectshad less than 5 s of clean EEG data at the baseline, his data were excluded from thefollowing analysis.
After combining the different stimuli used for the same sub-conditions (i.e.,card III and VII for the high movement sub-conditions, and card V and VI for the lowmovement sub-conditions), a three-way repeated-measures ANOVA was conductedto compare the mu suppression within-subjects across different tasks (attribution,identification, and observation), scalp locations (C3, Cz, and C4), and movementconditions (high movement and low movement).
3. Results
3.1. Attribution of human movement
Subject statements were transcribed verbatim and codedaccording to standard RCS rules. Three of the authors, who areexperts well trained in the RCS and have been using the RCS in clin-ical and research settings for many years (two of them for decades),independently coded each transcription and their agreement aboutthe presence vs. absence of M was 100%. Of the 19 participants,17 saw at least one M response card from the high M frequency
Author's personal copy
236 L. Giromini et al. / Biological Psychology 85 (2010) 233–241
Fig. 1. Examples of stimuli and instructions. (a) Experimental condition. Stimuli with high proportion of human movement attribution (top) and low proportion of humanmovement attribution (bottom): subjects were asked to think of what they might be. (b) Contrast condition. Same stimuli used in condition “a”: subjects were guided toidentify either a commonly reported human movement response (top) or a different commonly reported response (bottom). (c) Control condition. Drawings resembling theRorschach stimuli: subjects were asked to observe drawings representing either human movements (top) or absence of movement (bottom). Example stimuli reported in (a)and (b) are taken from: Hermann Rorschach, Rorschach-Test, Psychodiagnostic Plates.Copyright by Verlag Hans Huber, Hogrefe A.G., Bern, Switzerland. 1921, 1948, 1994, 2009. Reprinted by kind permission of the publisher.
Fig. 2. Layout of the study design. In the upper half, the sequence of conditions A, B, and C lasting 100 s, after baseline of 25 s. Condition A was presented twice. Specificinstructions related are displayed under each condition. In the lower half, the sequence of sub-conditions A1 to C2, lasting each 50 s. Sub-conditions were presented inrandomized order. Note: hm, human movement; non-hm, response without human movement; M, Rorschach code for human movement responses.
Author's personal copy
L. Giromini et al. / Biological Psychology 85 (2010) 233–241 237
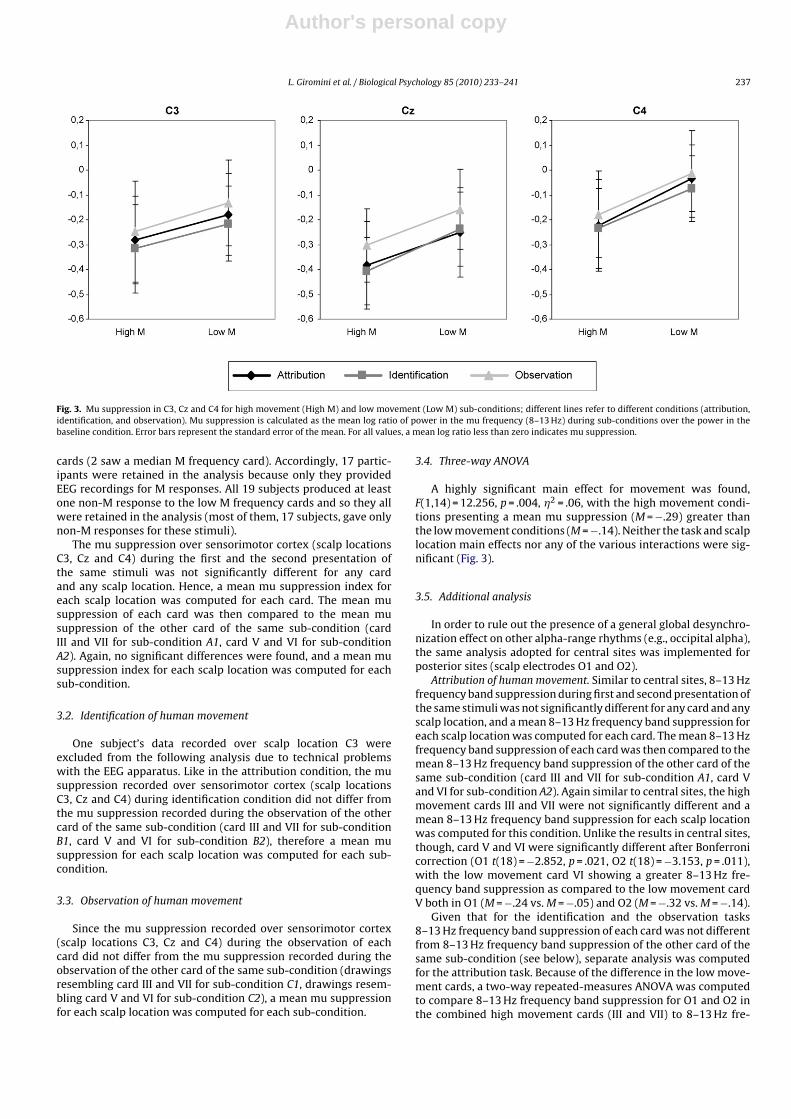
Fig. 3. Mu suppression in C3, Cz and C4 for high movement (High M) and low movement (Low M) sub-conditions; different lines refer to different conditions (attribution,identification, and observation). Mu suppression is calculated as the mean log ratio of power in the mu frequency (8–13 Hz) during sub-conditions over the power in thebaseline condition. Error bars represent the standard error of the mean. For all values, a mean log ratio less than zero indicates mu suppression.
cards (2 saw a median M frequency card). Accordingly, 17 partic-ipants were retained in the analysis because only they providedEEG recordings for M responses. All 19 subjects produced at leastone non-M response to the low M frequency cards and so they allwere retained in the analysis (most of them, 17 subjects, gave onlynon-M responses for these stimuli).
The mu suppression over sensorimotor cortex (scalp locationsC3, Cz and C4) during the first and the second presentation ofthe same stimuli was not significantly different for any cardand any scalp location. Hence, a mean mu suppression index foreach scalp location was computed for each card. The mean musuppression of each card was then compared to the mean musuppression of the other card of the same sub-condition (cardIII and VII for sub-condition A1, card V and VI for sub-conditionA2). Again, no significant differences were found, and a mean musuppression index for each scalp location was computed for eachsub-condition.
3.2. Identification of human movement
One subject’s data recorded over scalp location C3 wereexcluded from the following analysis due to technical problemswith the EEG apparatus. Like in the attribution condition, the musuppression recorded over sensorimotor cortex (scalp locationsC3, Cz and C4) during identification condition did not differ fromthe mu suppression recorded during the observation of the othercard of the same sub-condition (card III and VII for sub-conditionB1, card V and VI for sub-condition B2), therefore a mean musuppression for each scalp location was computed for each sub-condition.
3.3. Observation of human movement
Since the mu suppression recorded over sensorimotor cortex(scalp locations C3, Cz and C4) during the observation of eachcard did not differ from the mu suppression recorded during theobservation of the other card of the same sub-condition (drawingsresembling card III and VII for sub-condition C1, drawings resem-bling card V and VI for sub-condition C2), a mean mu suppressionfor each scalp location was computed for each sub-condition.
3.4. Three-way ANOVA
A highly significant main effect for movement was found,F(1,14) = 12.256, p = .004, �2 = .06, with the high movement condi-tions presenting a mean mu suppression (M = −.29) greater thanthe low movement conditions (M = −.14). Neither the task and scalplocation main effects nor any of the various interactions were sig-nificant (Fig. 3).
3.5. Additional analysis
In order to rule out the presence of a general global desynchro-nization effect on other alpha-range rhythms (e.g., occipital alpha),the same analysis adopted for central sites was implemented forposterior sites (scalp electrodes O1 and O2).
Attribution of human movement. Similar to central sites, 8–13 Hzfrequency band suppression during first and second presentation ofthe same stimuli was not significantly different for any card and anyscalp location, and a mean 8–13 Hz frequency band suppression foreach scalp location was computed for each card. The mean 8–13 Hzfrequency band suppression of each card was then compared to themean 8–13 Hz frequency band suppression of the other card of thesame sub-condition (card III and VII for sub-condition A1, card Vand VI for sub-condition A2). Again similar to central sites, the highmovement cards III and VII were not significantly different and amean 8–13 Hz frequency band suppression for each scalp locationwas computed for this condition. Unlike the results in central sites,though, card V and VI were significantly different after Bonferronicorrection (O1 t(18) = −2.852, p = .021, O2 t(18) = −3.153, p = .011),with the low movement card VI showing a greater 8–13 Hz fre-quency band suppression as compared to the low movement cardV both in O1 (M = −.24 vs. M = −.05) and O2 (M = −.32 vs. M = −.14).
Given that for the identification and the observation tasks8–13 Hz frequency band suppression of each card was not differentfrom 8–13 Hz frequency band suppression of the other card of thesame sub-condition (see below), separate analysis was computedfor the attribution task. Because of the difference in the low move-ment cards, a two-way repeated-measures ANOVA was computedto compare 8–13 Hz frequency band suppression for O1 and O2 inthe combined high movement cards (III and VII) to 8–13 Hz fre-
Author's personal copy
238 L. Giromini et al. / Biological Psychology 85 (2010) 233–241
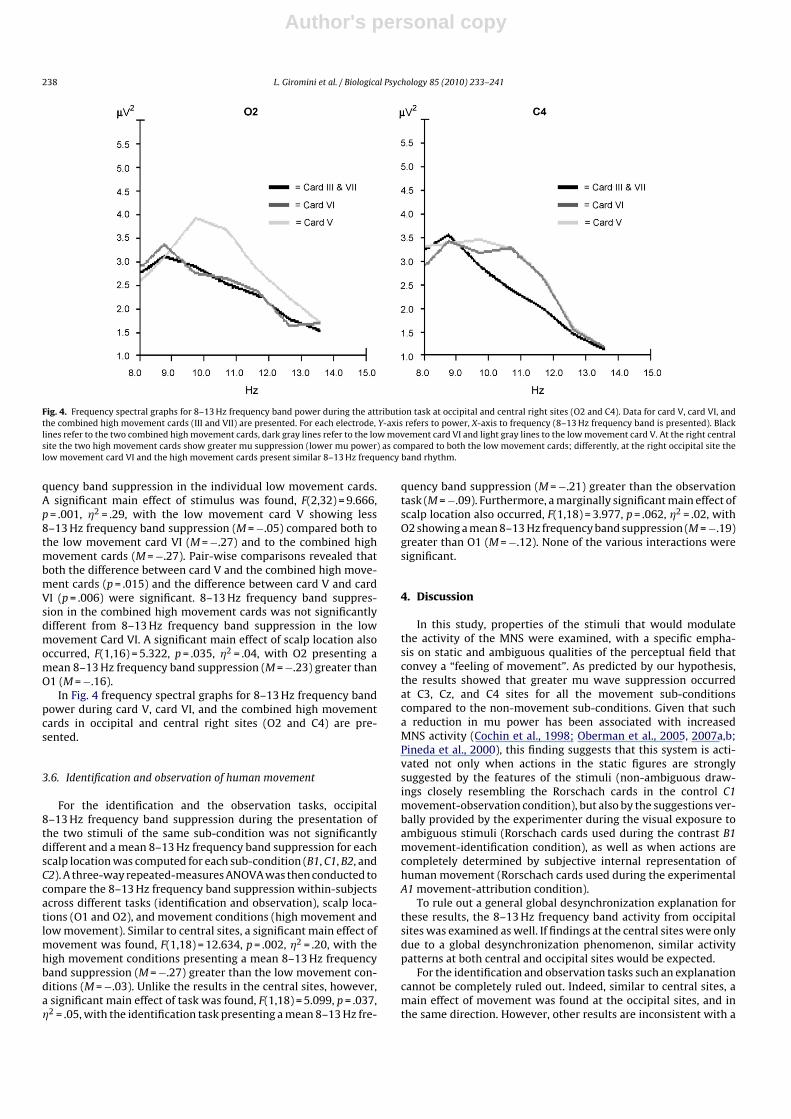
Fig. 4. Frequency spectral graphs for 8–13 Hz frequency band power during the attribution task at occipital and central right sites (O2 and C4). Data for card V, card VI, andthe combined high movement cards (III and VII) are presented. For each electrode, Y-axis refers to power, X-axis to frequency (8–13 Hz frequency band is presented). Blacklines refer to the two combined high movement cards, dark gray lines refer to the low movement card VI and light gray lines to the low movement card V. At the right centralsite the two high movement cards show greater mu suppression (lower mu power) as compared to both the low movement cards; differently, at the right occipital site thelow movement card VI and the high movement cards present similar 8–13 Hz frequency band rhythm.
quency band suppression in the individual low movement cards.A significant main effect of stimulus was found, F(2,32) = 9.666,p = .001, �2 = .29, with the low movement card V showing less8–13 Hz frequency band suppression (M = −.05) compared both tothe low movement card VI (M = −.27) and to the combined highmovement cards (M = −.27). Pair-wise comparisons revealed thatboth the difference between card V and the combined high move-ment cards (p = .015) and the difference between card V and cardVI (p = .006) were significant. 8–13 Hz frequency band suppres-sion in the combined high movement cards was not significantlydifferent from 8–13 Hz frequency band suppression in the lowmovement Card VI. A significant main effect of scalp location alsooccurred, F(1,16) = 5.322, p = .035, �2 = .04, with O2 presenting amean 8–13 Hz frequency band suppression (M = −.23) greater thanO1 (M = −.16).
In Fig. 4 frequency spectral graphs for 8–13 Hz frequency bandpower during card V, card VI, and the combined high movementcards in occipital and central right sites (O2 and C4) are pre-sented.
3.6. Identification and observation of human movement
For the identification and the observation tasks, occipital8–13 Hz frequency band suppression during the presentation ofthe two stimuli of the same sub-condition was not significantlydifferent and a mean 8–13 Hz frequency band suppression for eachscalp location was computed for each sub-condition (B1, C1, B2, andC2). A three-way repeated-measures ANOVA was then conducted tocompare the 8–13 Hz frequency band suppression within-subjectsacross different tasks (identification and observation), scalp loca-tions (O1 and O2), and movement conditions (high movement andlow movement). Similar to central sites, a significant main effect ofmovement was found, F(1,18) = 12.634, p = .002, �2 = .20, with thehigh movement conditions presenting a mean 8–13 Hz frequencyband suppression (M = −.27) greater than the low movement con-ditions (M = −.03). Unlike the results in the central sites, however,a significant main effect of task was found, F(1,18) = 5.099, p = .037,�2 = .05, with the identification task presenting a mean 8–13 Hz fre-
quency band suppression (M = −.21) greater than the observationtask (M = −.09). Furthermore, a marginally significant main effect ofscalp location also occurred, F(1,18) = 3.977, p = .062, �2 = .02, withO2 showing a mean 8–13 Hz frequency band suppression (M = −.19)greater than O1 (M = −.12). None of the various interactions weresignificant.
4. Discussion
In this study, properties of the stimuli that would modulatethe activity of the MNS were examined, with a specific empha-sis on static and ambiguous qualities of the perceptual field thatconvey a “feeling of movement”. As predicted by our hypothesis,the results showed that greater mu wave suppression occurredat C3, Cz, and C4 sites for all the movement sub-conditionscompared to the non-movement sub-conditions. Given that sucha reduction in mu power has been associated with increasedMNS activity (Cochin et al., 1998; Oberman et al., 2005, 2007a,b;Pineda et al., 2000), this finding suggests that this system is acti-vated not only when actions in the static figures are stronglysuggested by the features of the stimuli (non-ambiguous draw-ings closely resembling the Rorschach cards in the control C1movement-observation condition), but also by the suggestions ver-bally provided by the experimenter during the visual exposure toambiguous stimuli (Rorschach cards used during the contrast B1movement-identification condition), as well as when actions arecompletely determined by subjective internal representation ofhuman movement (Rorschach cards used during the experimentalA1 movement-attribution condition).
To rule out a general global desynchronization explanation forthese results, the 8–13 Hz frequency band activity from occipitalsites was examined as well. If findings at the central sites were onlydue to a global desynchronization phenomenon, similar activitypatterns at both central and occipital sites would be expected.
For the identification and observation tasks such an explanationcannot be completely ruled out. Indeed, similar to central sites, amain effect of movement was found at the occipital sites, and inthe same direction. However, other results are inconsistent with a

Author's personal copy
L. Giromini et al. / Biological Psychology 85 (2010) 233–241 239
single global alpha-like explanation. Unlike the findings at centralsites, a significant main effect of task and a marginally significantmain effect of scalp location occurred at the occipital sites.
Stronger evidence against the single global alpha-like expla-nation emerges for the attribution task. Unlike the results at thecentral sites, 8–13 Hz frequency band suppression to the two lowmovement cards at occipital sites differed substantially so thatthey could not be combined. 8–13 Hz frequency band suppres-sion to one of the low movement cards (card VI, sub-condition A2)was identical to the combined high movement cards (card III andVII, sub-condition A1). Accordingly, examination of the frequencyspectral graphs confirmed that the 8–13 Hz frequency band powerduring attributions is consistent with an expected effect (more sup-pression for both high movement cards as compared to both lowmovement cards) on central and not occipital sites, especially atthe right side. A remaining puzzling observation for the attributiontask is the high 8–13 Hz frequency band power found for card V atoccipital sites. A possible explanation may be that card V “pulls”for a specific, simple, popular response (i.e., “a bat” or “a butterfly”)more often than do card III, VI and VII (Exner and Erdberg, 2005),and that for this reason the answer to the question “what might thisbe?” is an easier task to solve than for the other cards. Other expla-nations may be possible as well. Nonetheless, the different patternsobserved at central and occipital sites suggest that, at least for theattribution task, mu suppression at central sites is not just due to aglobal alpha-like effect, but more likely related to MNS activation.Nonetheless, even with some limitations, these results show for thefirst time that the self-initiated “feeling of movement,” internallyperceived and cognitively processed by subjects and identified bythe Rorschach M responses, is a powerful stimulus for modulatingthe MNS activity.
Mirror neurons are premotor neurons that fire when the sub-ject performs an action and also when the subject observes anothersubject performing an action (Buccino et al., 2001; Dapretto et al.,2005; Gallese, 2003; Rizzolatti et al., 2001; Umiltà et al., 2001). Todate, research has shown that the MNS is not a biological mech-anism for action recognition, but that some specific clues in thestimuli are required for the activation of this neural system, suchas a goal, an agent, and a context for the perceived action. The useof the static, ambiguous Rorschach inkblots allows us to clarify thepower of a prerequisite that seems necessary for MNS activation,the identification with the other. The importance of this processis underscored in our experiment by the minimal perceptual cluesprovided by the stimuli that are, nonetheless, presumably sufficientfor the activation of the MNS.
Non-human and static pictures have been used in MNS researchfor ascertaining the role played by the understanding of inten-tions. Oberman et al. (2007a) used videos of robot hands engaged inobject-directed (picking up a ball) and non-object-directed (open-ing and closing hand with no object present) actions as well ashuman hands moving volitionally (opening and closing the righthand) and non-volitionally (hand being moved by a string). Theyreported that both human and robot movement suppressed the muwave to relatively the same extent, suggesting that the “human-ness” and the volitional qualities of the stimulus are not necessaryfor engaging MNS activity. In addition, the observation of staticpictures of dynamic action seems to produce effects on movement-related brain areas (Proverbio et al., 2009) and specifically MNSactivation (Rizzolatti and Craighero, 2004). The medial tempo-ral/medial superior temporal cortex (MT/MST complex) has beenshown to activate while observing implied motion, i.e., dynamicinformation extracted from static images (Kourtzi and Kanwisher,2000). Inferring motion from still images is likely to imply high-level perceptual inference engaging the mirror neuron network,which is highly suitable for matching action observation withaction execution. Enticott et al. (2008) showed that the MNS acti-
vation is associated with static (still facial photographs) but notdynamic (video clips showing the morphing from a neutral to anemotional facial expression) facial emotion processing. Ebisch etal. (2008) found overlapping activation of a mirroring mechanismfor tactile experiences and the sight of touch, regardless of stim-uli being animate and inanimate, independent of the intentionalityof the observed touch. Our results are consistent with this lineof research and confirm that static and non-intentional featuresof even ambiguously shaped pictures such as the Rorschach cardstimuli are sufficient to engage the MNS.
Studies performed so far have used perceptual stimuli in whichthe action is explicitly (moving objects in video clips) or implic-itly (still pictures) placed into the figures. Because of the graphicproperties of the Rorschach cards, we showed that the activationof a purely internal representation of the sense of movement is ableto activate MNS. It is likely that multimodal proprioceptive chan-nels concur to generate the experience of movement. Indeed, it hasbeen shown that the MNS is not merely related to the visual stimu-lation but involve other sensory mechanisms and is active for a richrepertoire of body actions. Therefore, the mirror mechanism seemsto serve several functions that depend on the part of the motor sys-tem that resonates: prefrontal and premotor areas become activein coding actions, subcortical structures as amygdala for emotion-laden stimuli, other circuits that store sensory copies of the actionif action understanding is or is not required in the task (Rizzolattiet al., 2001). For instance, audio-visual mirror neurons in area F5have been found to discharge when an action can only be heard bysound (Kohler et al., 2002) and both animate and inanimate touch(the sight of objects touching each other) activates inner repre-sentations of touch (activation of the MNS responding when thesubject is touched and sees someone else being touched) (Keyserset al., 2004). Furthermore, mirror-touch synesthesia, in which theobservation of touch on other human beings results in tactile sen-sation on the subject’s own body, was significantly associated withempathy scale scores (Banissy and Ward, 2007). Even pain sensa-tion seems to be involved in embodied simulation as activation inthe anterior cingulate cortex and anterior insula has been foundwhen people empathized for another individual in pain (Singer etal., 2004).
A shared process of identification modulated by the network ofmirror neurons may therefore underlay the mechanisms of embod-ied simulation, social cognition, and empathy. In an EEG study,subjects watching videos of interactive social actions showed asignificant suppression of mu wave compared to watching socialnon-interactive and non-interactive, non-social actions (Obermanet al., 2007b) and the magnitude of suppression was similar tothat observed in the present study. Because of the lack of clearactions in the Rorschach stimuli set, our findings seem to con-firm the hypothesis that the internal sense of identification witha living, moving person is among the most important componentsof embodied simulation, even when identification relies more oninner representations than external cues. Support for this hypoth-esis comes from several other data reviewed above that show theMNS activates when watching pain sensations in others (Singeret al., 2004), non-human beings in human-like actions (Obermanet al., 2007a), actions whose crucial part is hidden and can onlybe inferred (Umiltà et al., 2001), implied motion in still images(Urgesi et al., 2006), and even gazing at artwork (Freeberg andGallese, 2007) and alphabet characters passively observed by right-handers (Longcamp et al., 2003) and left-handers (Longcamp etal., 2005). Empathic feelings should not be regarded therefore as amere intuition but as a pre-rational making sense of behavior, emo-tions, and cognition of others. As it occurs when individuals watchothers engaged in a goal-directed action, an ambiguous figure asa Rorschach card can activate embodied simulation enabling “theobserver to use his/her own resources to penetrate the world of the

Author's personal copy
240 L. Giromini et al. / Biological Psychology 85 (2010) 233–241
other without the need of explicitly theorizing about it” (Gallese,2003, p. 174). From a broader viewpoint, this view is consistent withthe “as-if body loop” referred to by Damasio (1999) who specu-lated about the ability of the brain in simulation mode to reproduceactual somatic states when emotions are induced not by observingothers but also by only feeling them in mind, without an externalrelated stimulus. From the point of view of test validity, these find-ings could also suggest that the standard interpretation of humanmovement on the Rorschach may have a neurophysiological foun-dation involving MNS activation. A more rigorous assessment ofthis relationship is currently being investigated.
This study had several limitations. First, as a white card was usedas baseline, the possibility that the subjects’ spontaneous thinkingmay activate the MNS at different levels cannot be ruled out. Thebaseline presentation of the white card for only 25 s was thereforeset for minimizing distraction or spontaneous thinking. Second,this limited duration provided a limited amount of clean EEG dataand consequently high variability or error in the statistical anal-ysis. Third, the baseline condition showed a lot of alpha activityover occipital sites but relatively less activity over central sites,and this may have biased the mu-suppression-like effect at pos-terior sites. Fourth, the sample size was small and the presenceof artifacts in the EEG signal led to an even smaller sample size.Fifth, in order to not influence the participants, condition A (spon-taneous attribution of human movement) was presented first andcondition C (observation of drawings) last. Although sub-conditionsA1 to C2 were presented at random, lack of randomization of theexperimental, contrast, and control conditions may have alteredthe final results. Finally, although mu wave suppression is consid-ered a valid index of mirror neuron activity (Cochin et al., 1998;Muthukumaraswamy and Johnson, 2004; Oberman et al., 2007a,b;Pineda et al., 2000; Pizzamiglio et al., 2005; Rizzolatti et al., 2001),owing to the low spatial resolution of EEG it is difficult to differenti-ate between activity selective to the premotor MNS and activity inother regions that are part of a larger action observation/executionnetwork that may modulate the activity in the premotor MNS(Muthukumaraswamy and Johnson, 2004; Muthukumaraswamyet al., 2004). Further investigations with higher-spatial-resolutiontechniques, such as fMRI and high-resolution EEG, may be able todissociate between these two sources of activation.
Nevertheless, the results were consistent with our hypothesisand support for the first time the idea that MNS activity, as indexedby EEG mu rhythms, can be activated by static, ambiguous stimulisuch as the Rorschach test cards. This supports the hypothesis thatstrong internal representation of the “feeling of movement” maybe sufficient to trigger MNS activity even when minimal externalcues are present.
Conflict of interest
None declared.
Acknowledgments
We thank Laura Giromini for drawing the control conditionstimuli and Jake Feldman for helping with the EEG recording.
References
Adolphs, R., 1999. Social cognition and the human brain. Trends in Cognitive Sciences3, 469–479.
Agnew, Z.K., Bhakoo, K.K., Puri, B.K., 2007. The human mirror system: a motor reso-nance theory of mind-reading. Brain Research Reviews 54, 286–293.
Altschuler, E.L., Vankov, A., Hubbard, E.M., Roberts, E., Ramachandran, V.S., Pineda,J.A., 2000. Mu wave blocking by observer of movement and its possible use asa tool to study theory of other minds. In: Poster session presented at the 30thAnnual Meeting of the Society for Neuroscience, New Orleans, LA, November.
Babiloni, C., Carducci, F., Cincotti, F., Rossini, P.M., Neuper, C., Pfurtscheller, G.,Babiloni, F., 1999. Human movement-related potentials vs. desynchronizationof EEG alpha rhythm: a high-resolution EEG study. NeuroImage 10, 658–665.
Banissy, M.J., Ward, J., 2007. Mirror-touch synesthesia is linked with empathy.Nature Neuroscience 10, 815–816.
Bendick, M.R., Klopfer, W.G., 1964. The effects of sensory deprivation and motorinhibition on Rorschach movement responses. Journal of Projective Techniquesand Personality Assessment 28, 261–264.
Bernier, R., Dawson, G., Webb, S., Murias, M., 2007. EEG mu rhythm and imitationimpairments in individuals with autism spectrum disorder. Brain and Cognition64 (3), 228–237.
Buccino, G., Amore, M., 2008. Mirror neurons and the understanding of behaviouralsymptoms in psychiatric disorders. Current Opinion in Psychiatry 21, 281–285.
Buccino, G., Binkofski, F., Fink, G.R., Fadiga, L., Fogassi, L., Gallese, V., Rizzolatti, G.,Freund, H.J., 2001. Action observation activates premotor and parietal areas ina somatotopic manner: an fMRI study. European Journal of Neuroscience 13,400–404.
Buccino, G., Vogt, S., Ritzl, A., Fink, G.R., Zilles, K., Freund, H.J., Rizzolatti, G., 2004.Neural circuits underlying imitation learning of hand action: an event-relatedfMRI study. Neuron 42, 323–334.
Cochin, S., Barthelemy, B., Lejeune, S., Roux, J., Martineau, J., 1998. Perception ofmotion and qEEG activity in human adults. Electroencephalography and ClinicalNeurophysiology 108, 287–295.
Damasio, A., 1999. The Feeling of What Happens: Body and Emotion in the Makingof Consciousness. Harcourt Brace, New York.
Dapretto, M., Davies, M.S., Pfeifer, J.H., Scott, A.A., Sigman, M., Bookheimer, S.Y.,Iacoboni, M., 2005. Understanding emotions in others: mirror neuron dys-function in children with autism spectrum disorders. Nature Neuroscience 9,28–30.
di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., Rizzolatti, G., 1992. Understand-ing motor events: a neurophysiological study. Experimental Brain Research 91,176–180.
Ebisch, S.J.H., Perrucci, M.G., Ferretti, A., Del Gratta, C., Romani, G.L., Gallese, V., 2008.The sense of touch: embodied simulation in a visuotactile mirroring mechanismfor observed animate or inanimate touch. Journal of Cognitive Neuroscience 20,1611–1623.
Enticott, P.G., Johnston, P.J., Herring, S.E., Hoy, K.E., Fitzgerald, P.B., 2008. Mirror neu-ron activation is associated with facial emotion processing. Neuropsychologia46, 2851–2854.
Exner Jr, J.E., 2003. The Rorschach. A Comprehensive System. Vol. 1. Basic Founda-tions and Principles of Interpretation, 4th edn. John Wiley & Sons, New York.
Exner Jr., J.E., Erdberg, P., 2005. The Rorschach: A Comprehensive System. Vol. 2.Advanced Interpretation, 3rd edn. John Wiley & Sons, New York.
Fadiga, L., Fogassi, L., Pavesi, G., Rizzolatti, G., 1995. Motor facilitation during actionobservation: a magnetic stimulation study. Journal of Neurophysiology 73,2608–2611.
Ferracuti, S., Cannoni, E., Burla, F., Lazzari, R., 1999. Correlations for the Rorschachwith the Torrance tests of creative thinking. Perceptual and Motor Skills 89,863–870.
Freeberg, D., Gallese, V., 2007. Motion, emotion and empathy in esthetic experience.Trends in Cognitive Sciences 11, 197–203.
Gallese, V., 2001. The ‘shared manifold hypothesis’: from mirror neurons to empathy.Journal of Consciousness Studies 8, 33–50.
Gallese, V., 2003. The roots of empathy: the shared manifold hypothesis and theneural basis of intersubjectivity. Psychopathology 36, 171–180.
Gallese, V., 2006. Intentional attunement: a neurophysiological perspective on socialcognition and its disruption in autism. Brain Research 1079, 15–24.
Gallese, V., Goldman, A., 1998. Mirror neurons and the simulation theory of mind-reading. Trends in Cognitive Sciences 2, 493–501.
Gallese, V., Fadiga, L., Fogassi, L., Rizzolatti, G., 1996. Action recognition in the pre-motor cortex. Brain 119, 593–609.
Gallucci, N.T., 1989. Personality assessment with children of superior intelligence:divergence versus psychopathology. Journal of Personality Assessment 53,749–760.
Gastaut, H., 1952. Etude electrogastrographique de la reactivité des rythmesrolandiques (EEG study of rolandic rhythms activity). Revue Neurologique 87,176–182.
Gazzola, V., Aziz-Zadeh, L., Keysers, C., 2006. Empathy and the somatropic auditorymirror system in humans. Current Biology 16, 1824–1829.
Goldensohn, E.S., Legatt, A.D., Koszer, S., Wolf, S.M., 1999. Goldensohn’s EEGInterpretation: Problems of Overreading and Underreading. Futura Publishing,Armonk, NY.
Grézes, J., Armony, J.L., Rowe, J., Passingham, R.E., 2003. Activations related to “mir-ror” and “canonical” neurons in the human brain: an fMRI study. NeuroImage18, 928–937.
Hadijkhani, N., Joseph, R.M., Snyder, J., Tager-Flusberg, H., 2006. Anatomical dif-ferences in the mirror neuron system and social cognitive network in autism.Cerebral Cortex 16, 1276–1282.
Iacoboni, M., Dapretto, M., 2006. The mirror neuron system and the consequencesof its dysfunction. Nature Reviews Neuroscience 7, 942–951.
Iacoboni, M., Koski, L.M., Brass, M., Bekkering, H., Woods, R.P., Dubeau, M.C., Mazz-iotta, J.C., Rizzolatti, G., 2001. Reafferent copies of imitated actions in the rightsuperior temporal cortex. Proceedings of the National Academy of Sciences 98,13995–13999.
Iacoboni, M., Woods, R.P., Brass, M., Bekkering, H., Mazziotta, J.C., Rizzolatti, G., 1999.Cortical mechanisms of human imitation. Science 286, 2526–2528.

Author's personal copy
L. Giromini et al. / Biological Psychology 85 (2010) 233–241 241
Kaplan, J.T., Iacoboni, M., 2006. Getting a grip on other minds: mirror neurons, inten-tion understanding, and cognitive empathy. Social Neuroscience 1, 175–183.
Keysers, C., Wicker, B., Gazzola, V., Anton, J.L., Fogassi, L., Gallese, V., 2004. A touchingsight: SII/PV activation during the observation and experience of touch. Neuron42, 335–346.
Kohler, E., Keysers, C., Umiltà, M.A., Fogassi, L., Gallese, V., Rizzolatti, G., 2002. Hearingsounds, understanding actions: action representation in mirror neurons. Science297, 846–848.
Kourtzi, Z., Kanwisher, N., 2000. Activation in human MT/MST by static images withimplied motion. Journal of Cognitive Neuroscience 12, 48–55.
Longcamp, M., Anton, J.L., Roth, M., Velay, J.L., 2003. Visual presentation of single let-ters activates a premotor area involved in writing. NeuroImage 19, 1492–1500.
Longcamp, M., Anton, J.L., Roth, M., Velay, J.L., 2005. Premotor activations in responseto visually presented single letters depend on the hand used to write: a studyin left-handers. Neuropsychologia 43, 1699–1846.
Maeda, F., Kleiner-Fisman, G., Pascual-Leone, A., 2002. Motor facilitation whileobserving hand actions: specificity of the effect and role of observer’s orien-tation. Journal of Neurophysiology 87, 1329–1335.
Martineau, J., Cochin, S., Magne, R., Barthelemy, C., 2008. Impaired cortical activationin autistic children: is the mirror neuron system involved? International Journalof Psychophysiology 68 (1), 35–40.
Morrison, I., Lloyd, D., di Pellegrino, G., Roberts, N., 2004. Vicarious responses topain in anterior cingulate cortex: is empathy a multisensory issue? CognitiveAffective and Behavioral Neuroscience 4, 270–278.
Mukamel, R., Ekstrom, A.D., Kaplan, J., Iacoboni, M., Fried, I., 2010. Single-neuronresponses in humans during execution and observation of actions. Current Biol-ogy 20, 750–756.
Muthukumaraswamy, S.D., Johnson, B.W., 2004. Primary motor cortex activationduring action observation revealed by wavelet analysis of the EEG. Clinical Neu-rophysiology 115, 1760–1766.
Muthukumaraswamy, S.D., Johnson, B.W., McNair, N.A., 2004. Mu rhythm modula-tion during observation of an object-directed grasp. Cognitive Brain Research19, 195–201.
Oberman, L.M., Hubbard, E.M., McCleery, J.P., Altschuler, E.L., Ramachandran, V.S.,Pineda, J.A., 2005. EEG evidence for mirror neuron dysfunction in autism spec-trum disorders. Cognitive Brain Research 24, 190–198.
Oberman, L.M., McCleery, J.P., Ramachandran, V.S., Pineda, J.A., 2007a. EEG evidencefor mirror neuron activity during the observation of human and robot actions:toward an analysis of the human qualities of interactive robots. Neurocomputing70, 2194–2203.
Oberman, L.M., Pineda, J.A., Ramachandran, V.S., 2007b. The human mirror neuronsystem: a link between action observation and social skills. Social Cognitive andAffective Neuroscience 2, 62–66.
Orlinsky, D.E., 1966. Rorschach test correlates of dreaming and dream recall. Journalof Projective Techniques and Personality Assessment 30, 250–253.
Oztop, E., Arbib, M.A., 2002. Schema design and implementation of the grasp-relatedmirror neuron system. Biological Cybernetics 87, 116–140.
Parsons, L.M., Fox, P.T., Downs, J.H., Glass, T., Hirsch, T.B., Martin, C.C., Jerabek, P.A.,Lancaster, J.L., 1995. Use of implicit motor imagery for visual shape discrimina-tion as revealed by PET. Nature 375, 54–58.
Pelphrey, K.A., Morris, J.P., 2006. Brain mechanisms for interpreting the actions ofothers from biological motion cues. Current Directions in Psychological Science15, 136–140.
Pfeifer, J.H., Iacoboni, M., Mazziotta, J.C., Dapretto, M., 2008. Mirroring others’ emo-tions relates to empathy and interpersonal competence in children. NeuroImage39, 2076–2085.
Pfurtscheller, G., Neuper, C., Andrew, C., Edlinger, G., 1997. Foot and hand area murhythms. International Journal of Psychophysiology 26, 121–135.
Pineda, J.A., 2005. The functional significance of mu rhythms: translating “seeing”and “hearing” into “doing”. Brain Research Reviews 50, 57–68.
Pineda, J.A., 2008. Sensorimotor cortex as a critical component of an ‘extended’ mir-ror neuron system: does it solve the development, correspondence, and controlproblems in mirroring? Behavioral and Brain Functions 4, 47.
Pineda, J.A., Hecht, E., 2009. Mirroring and mu rhythm involvement in social cog-nition: are there dissociable subcomponents of theory of mind? BiologicalPsychology 80, 306–314.
Pineda, J.A., Allison, B.Z., Vankov, A., 2000. The effects of self-movement, observa-tion and imagination on mu rhythms and readiness potentials (RP’s): towarda brain–computer interface (BCI). IEEE Transactions on Neural Systems andRehabilitation Engineering 8, 219–222.
Pizzamiglio, L., Aprile, T., Spitoni, G., Pitzalis, S., Bates, E., D’Amico, S., Di Russo,F., 2005. Separate neural systems for processing action- or non-action-relatedsounds. NeuroImage 24, 852–861.
Porcelli, P., Meyer, G.J., 2002. Construct validity of Rorschach variables for alex-ithymia. Psychosomatics 43, 360–369.
Proverbio, A.M., Riva, F., Zani, A., 2009. Observation of static pictures of dynamicactions enhances the activity of movement-related brain areas. PLoS One 4,e5389.
Rhue, J.W., Lynn, S.J., 1987. Fantasy proneness and psychopathology. Journal of Per-sonality and Social Psychology 53, 327–336.
Rizzolatti, G., Fadiga, L., Gallese, V., Fogassi, L., 1996. Premotor cortex and the recog-nition of motor actions. Cognitive Brain Research 3, 131–141.
Rizzolatti, G., Fogassi, L., Gallese, V., 2001. Neurophysiological mechanisms under-lying the understanding and imitation of action. Nature Review Neuroscience 2,661–670.
Rizzolatti, G., Craighero, L., 2004. The mirror neuron system. Annual Review of Neu-roscience 27, 169–192.
Rorschach, H., 1921. Psychodiagnostik. Bern, Bircher.Sato, W., Yoshikawa, S., 2007. Enhanced experience of emotional arousal in response
to dynamic facial expressions. Journal of Nonverbal Behavior 31, 119–135.Singer, T., Seymour, B., O’Doherty, J., Kaube, H., Dolan, R.J., Frith, C.D., 2004. Empathy
for pain involves the affective but not sensory components of pain. Science 303,1157–1162.
Steele, N.M., Kahn, M.W., 1969. Kinesthesis and the Rorschach M response. Journalof Projective Techniques and Personality Assessment 33, 5–10.
Strafella, A.P., Paus, T., 2000. Modulation of cortical excitability during actionobservation: a transcranial magnetic stimulation study. Neuroreport 11, 2289–2292.
Uddin, L.Q., Iacoboni, M., Lange, C., Keenan, J.P., 2007. The self and social cognition:the role of cortical midline structures and mirror neurons. Trends in CognitiveSciences 11, 153–157.
Ulloa, E.R., Pineda, J.A., 2007. Recognition of point-light biological motion: murhythms and mirror neuron activity. Behavioural Brain Research 183, 188–194.
Umiltà, M.A., Kohler, E., Gallese, V., Fogassi, L., Fadiga, L., Keysers, C., Rizzolatti,G., 2001. I know what you are doing”: a neurophysiological study. Neuron 32,91–101.
Urgesi, C., Moro, V., Candidi, M., Aglioti, S., 2006. Mapping implied body actions inthe human motor system. The Journal of Neuroscience 26, 7942–7949.
Wicker, B., Keysers, C., Plailly, J., Royet, J.P., Gallese, V., Rizzolatti, G., 2003. Both of usdisgusted in my insula: the common neural basis of seeing and feeling disgust.Neuron 40, 655–664.
Williams, J.H.G., Whiten, A., Suddendorf, T., Perrett, D.I., 2001. Imitation, mirrorneurons and autism. Neuroscience and Biobehavioral Reviews 25, 287–295.
Wood, J.M., Krishnamurthy, R., Archer, R.P., 2003. Three factors of the Comprehen-sive System for the Rorschach and their relationship to Wechsler IQ scores in anadolescent sample. Assessment 10, 259–265.
Young, G.R., Wagner, E.E., 1993. Behavioral specificity in the Rorschach humanmovement response: a comparison of strippers and models. Journal of ClinicalPsychology 49, 407–412.




















