
The Effects of Parent and Offspring Gender On Food Allocation in Budgerigars
23
THE EFFECTS OF PARENT AND OFFSPRING GENDER ON FOOD ALLOCATION IN BUDGERIGARS by JUDY STAMPS1), ANNE CLARK, BARBARA KUS and PAT ARROWOOD (Department of Zoology, University of California, Davis, California 95616, U.S.A.) (With 3 Figures) (Acc. 2-VI-1986) Introduction In most diploid organisms, parents and offspring come in two sexes. Hence, whenever both parents provision the young, there are four possi- ble permutations of feeding relationships: father-daughter, father-son, mother-daughter and mother-son. According to current theory, each of these relationships may involve a different amount of resources. On the one hand, selection may favor differential investment in male and female offspring (TRIVERS & HARE, 1976; MAYNARD-SMITH, 1980; CLUTTON-BROCK & ALBON, 1982). For example, in species with pro- nounced sexual size dimorphism, the larger sex may require more food from the parents (e.g. mammals, REITER et al., 1978; CLUTTON-BROCK El al., 1982, or social hymenopterans, TRIVERS & HARE, 1976). Conversely, mothers and fathers may invest different amounts in their offspring (TRIVERS, 1972, 1985; POWER, 1980; WITTENBERGER, 1982; ROSKAFT, 1983; MARTINDALE, 1984). Finally, there might be important interac- tions between parent and offspring gender. For example, if offspring of one sex were more expensive to raise, one parent might be selected to provide the additional care. Studies of interactions between parent and offspring gender in the con- text of food provisioning are still in their infancy. One reason is that mothers provide most or all of the nutrients in many of the taxa with parental care (see TRIVERS, 1985). But even in birds, where biparental provisioning is common, data on gender effects are scarce. They might be expected in species with pronounced sexual size dimorphism, such that one offspring sex presumably costs more to raise than the other (e.g. NEWTON, 1978; HOWE, 1979; FIALA & CONGDON, 1983; BANCROFT, 1) This research has been supported by grants from the Institute of Ecology and the Agricultural Experiment Station, University of California, Davis.
Transcript of The Effects of Parent and Offspring Gender On Food Allocation in Budgerigars

THE EFFECTS OF PARENT AND OFFSPRING GENDER ON FOOD ALLOCATION IN BUDGERIGARS
by
JUDY STAMPS1), ANNE CLARK, BARBARA KUS and PAT ARROWOOD
(Department of Zoology, University of California, Davis, California 95616, U.S.A.)
(With 3 Figures) (Acc. 2-VI-1986)
Introduction
In most diploid organisms, parents and offspring come in two sexes.
Hence, whenever both parents provision the young, there are four possi- ble permutations of feeding relationships: father-daughter, father-son,
mother-daughter and mother-son. According to current theory, each of
these relationships may involve a different amount of resources. On the
one hand, selection may favor differential investment in male and female
offspring (TRIVERS & HARE, 1976; MAYNARD-SMITH, 1980; CLUTTON-BROCK & ALBON, 1982). For example, in species with pro- nounced sexual size dimorphism, the larger sex may require more food
from the parents (e.g. mammals, REITER et al., 1978; CLUTTON-BROCK El
al., 1982, or social hymenopterans, TRIVERS & HARE, 1976). Conversely, mothers and fathers may invest different amounts in their offspring
(TRIVERS, 1972, 1985; POWER, 1980; WITTENBERGER, 1982; ROSKAFT,
1983; MARTINDALE, 1984). Finally, there might be important interac-
tions between parent and offspring gender. For example, if offspring of
one sex were more expensive to raise, one parent might be selected to
provide the additional care.
Studies of interactions between parent and offspring gender in the con-
text of food provisioning are still in their infancy. One reason is that
mothers provide most or all of the nutrients in many of the taxa with
parental care (see TRIVERS, 1985). But even in birds, where biparental
provisioning is common, data on gender effects are scarce. They might be expected in species with pronounced sexual size dimorphism, such
that one offspring sex presumably costs more to raise than the other (e.g. NEWTON, 1978; HOWE, 1979; FIALA & CONGDON, 1983; BANCROFT,
1) This research has been supported by grants from the Institute of Ecology and the Agricultural Experiment Station, University of California, Davis.

178
1984b; ROSKAFT & SLAGSVOLD, 1985). However, out of three studies of
sexually dimorphic species, none found that parents of either sex provi- sioned the larger sex offspring at higher rates (redwinged blackbirds
Agelaius phoeniceus, FIALA 1981a; rooks, Corvus frugilegus, ROSKAFT &
SLAGSVOLD, 1985; sparrowhawks, Accipiter nisus, NEWTON, 1978). How-
ever, HARPER (1985) has reported an intriguing but nonsignificant interaction between parent and offspring gender in European robins, Erithacus rubecula, in which the parents divided the fledglings, and fathers
fed daughters, while mothers fed sons.
Previous work on domesticated budgerigars (Melopsittacus undulatus) in
captive flocks suggested that interesting interactions between parent and
offspring gender might occur in this species. Male budgerigars made
considerable contributions to raising nestlings. Early in the nest cycle
they allofed the females, who then allocated this food to the young; later
some males also fed the nestlings directly (STAMPS et al., 1985). However, there was considerable intra and inter-individual variance in the
tendency of males to directly feed nestlings, and this variance seemed to
be related to the sex ratio of the brood. Hence in this study we have
focused on the following issues: 1 ) Does the feeding rate of either parent
vary according to offspring gender, and if so, why? 2) Is one offspring
gender fed more frequently than the other, and if so, do these extra
feedings have any beneficial effect on the more expensive sex? In order
to answer these questions, we gathered data on feeding rates, fledge
weights and ages, activity and social behavior in the first weeks after
fledging, and indices of reproductive success for males and females in
their first breeding season.
Methods General methods have been described in detail in STAMPS et al., 1985. Domestic budgerigars (Melopsittacus undulatus) were studied in large (2.5 x 3.2 x 6.3 m) outdoor flight cages at Davis, California from 1978 to 1981. Adults and fledglings were marked for easy identification with black Nyanzol-D dye on the head, neck, breast, wings, etc. and were leg banded for permanent identification. Wooden nestboxes equipped with one- way viewers allowed us to observe and record parent and offspring behaviour within the nests. Reproductive data (dates of egg laying, hatch dates, weight and size of young, fledge dates) were determined by checking each nest daily in the early morning; nestlings were marked with gentian violet, weighed and banded during these daily nest inspections.
Male parents rarely fed nestlings younger than 10 days old (STAMPS et al., 1985). Hence, we focused on the last two periods of nestling development: Period 3: day 11-22, and Period 4: day 23 to fledge age (about day 34, see Results, Section II). Day 11 and day 23 were used to demarcate Periods 3 and 4 because preliminary analysis showed that most budgerigar nestlings achieved asymptotic weights at day 22, and day 11 cor- responded to the age at which most nestlings achieved 50% of their asymptotic weight

179
(CLARK, unpubl. data). "Fledge weight" was the last recorded weight, taken within 24 hours of fledging. Fledging was defined as a nestling leaving the nestbox; in the few cases when a nestling subsequently returned to its nestbox, the first date that it left was con- sidered to be its fledge date. Broods which fledged only one offspring were called "singletons", while those fledging two or more offspring were called "families". For analyses involving fledge ages, broods were divided into small broods (one or two fledg- lings) and large broods (three or more fledglings).
Behavioural observations were made in 1978, 1979 and 1981 on 14 nests, of which five were singletons and nine were families (Table 1). These broods were divided into two major categories: female-biased broods (FBB), with 50 to 100% female nestlings, and male-biased broods (MBB) with less than 50% females. Similarly, female-biased families were FBF; while male-biased families were MBF. Sex ratios were specified by Pp, the percent female offspring within the brood.
We determined offspring sex when the birds matured; adult males have blue ceres while those of females are pale to dark brown. Two nestlings from the 14 observed nests were not sexed. One from nest # 501 died accidently when it fell from the nest at age 30, and one nestling from #413 was stolen soon after it fledged (see Table 1). For- tunately, both broods could be classified unambiguously as to sex bias. Four of the five nestlings in # 502 were male, so regardless of the gender of the accident victim, this was a male-biased brood. Similarly, #413 was clearly a female-biased brood (see Table 1). ).
Fledge weight, fledge age and sex ratio data were available for a total of 32 fully-sexed broods. Intervals between the production of one brood and the laying of the subsequent clutch could be calculated for most of these broods. Exceptions were nests which fledged young at the end of the breeding season, when the nestboxes were plugged or removed to prevent breeding into the autumn.
In addition to the 32 fully-sexed broods, six nests with three or more fledglings were partially-sexed (one nestling of unknown sex per nest). Sex ratios for these broods were estimated by averaging two ratios: the sex ratio if the unknown bird were a male, and the ratio if the bird were a female (see Table 1 for nest 413 and 502). These nests were included in Figs 1 and 2 for the sake of completeness, but were not used in any statistical analyses involving sex ratios.
Methods for observation of begging and feeding within the nests have been described elsewhere ((STAMPS et al., 1985). In 1979, juveniles were observed for the first three weeks after fledging. Beginning the morning after fledging (day 1 postfledge) each juvenile was observed daily for 15 minutes. On day 8 postfledge, daily focal samples were reduced to 5 min, and this regime continued through day 21. The order in which juveniles were observed was rotated daily; all observations were made between 0700 and 1130 hr. From these focal samples, we estimated juvenile activity levels by the number of self-initiated flights, which were defined as flights not obviously a result of displacements or attacks by other birds. "Social" self-initiated flights occurred when one bird landed within one body length of another; "nonsocial" flights terminated further than one body length from another bird.
Nonparametric statistical tests were used for analyses involving behavioural variables, since sample sizes were generally too small to determine if assumptions of equal variance and normalcy applied. Parametric tests were used when these assumptions were valid (e.g. for analyses of fledge ages, fledge weights and arcsin transformed sex ratios).

180

181
Results
I. Parental feeding behaviour.
Paternal feeding rates.
There were no significant differences in the rates at which fathers fed
male and female nestlings, when all nestlings were considered together
(see Table 2) or when we calculated the average feed rates for brothers and sisters within the same nests (Period 3: Wilcoxon Test, T = 9, N = 5
with two ties; Period 4: Wilcoxon Test, T = 6, N = 7). However, the sex
ratio within a brood was related to paternal feeding rates. Fathers fed
the offspring in female-biased nests at higher rates than those in male-
biased nests.
TABLE 2. Parental feeding rates (Regurgitations/hr/nestling) for male and female nestlings
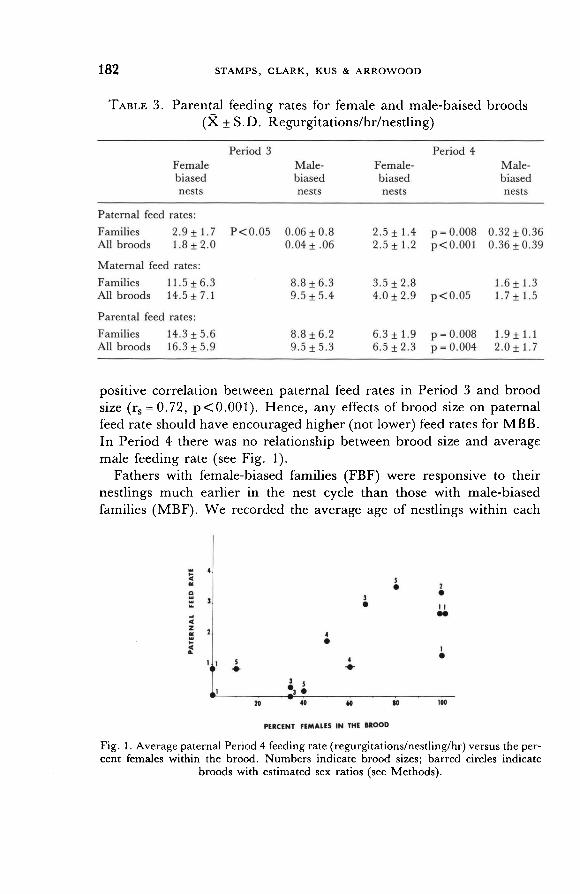
Paternal feeding rates were higher in female-biased families for both Period 3 and 4 nestlings (Table 3). Similarly, there were strong positive correlations between paternal feeding rates and Pq in seven fully-sexed families (Period 3: Spearman rank correlation coefficient rs = 0.87,
p<0.05, Period 4: rs=0.90, p<0.01, Fig. 1).
Adding singletons to the analysis gave a similar picture for Period 4.
There was a highly significant tendency for males to feed FBB more than
MBB (Table 3), and there was a strong positive relationship between P9 and male feeding rate for fully-sexed broods (rs = 0. 73, P < 0.01, Fig.
1). Period 3 singletons were never observed to be fed by their fathers.
Hence, adding singletons to the analysis for Period 3 obscured the rela-
tionship between sex ratio and paternal feeding rate (see Table 3). The
male-biased broods observed in this study were of slightly larger average sizes than the female-biased broods (FBB: X:t S.D. = 2.63 + 1.6 nest-
lings ; MBB: 3.0 + 1.8, Table 1), so we also investigated the relationship between brood size and paternal feeding rates. There was a strong
182
TABLE 3. Parental feeding rates for female and male-baised broods
(R + S.D. Regurgitations/hr/nestling)
positive correlation between paternal feed rates in Period 3 and brood
size (rs=O.72, p < 0.001). Hence, any effects of brood size on paternal feed rate should have encouraged higher (not lower) feed rates for MBB.
In Period 4 there was no relationship between brood size and average male feeding rate (see Fig. 1).
Fathers with female-biased families (FBF) were responsive to their
nestlings much earlier in the nest cycle than those with male-biased
families (MBF). We recorded the average age of nestlings within each
Fig. 1. Average paternal Period 4 feeding rate (regurgitations/nestling/hr) versus the per- cent females within the brood. Numbers indicate brood sizes; barred circles indicate
broods with estimated sex ratios (see Methods).

183
brood on the day the male was first observed to feed the nestlings. Fathers with FBF first fed when their nestlings were on average 7.3 days old (S.D. = 6.2, 6 nests), while those with MBF waited until the nestlings were an average of 25.8 (± 10.4, 4 nests) days old (Mann-Whitney U
Test, p = 0.005). One possible reason for this difference was that all males responded
similarly to offspring begging calls, but that nestlings in FBF began beg-
ging at earlier ages or at higher rates than those in MBF. In order to test
this hypothesis, we determined the highest beg rate ignored by each father
(e.g. the highest beg rate recorded on any day before the male was first
observed to feed, (see also STAMPS et al., 1985). If all males were similarly
responsive to nestling begging, then the highest beg rates on days prior to the first feeding should have been comparable for both types of
families. However, our results suggest that fathers with FBF were far
more responsive to nestling begging than those with MBF. On average, the highest beg rate prior to paternal feeding for FBF was 1.3 begs/hr
(S.D. = 1.4), while the average beg rate prior to first paternal feed for
MBF was 33.5 ± 18.9 begs/hr (Mann-Whitney U Test, p = 0.005). Since
the intensity of begging calls tends to increase as a function of nestling
age (see STAMPS et al., 1985), fathers feeding FBF early in the nest cycle were responding to softer calls than fathers which fed MBF in late Period 3 and 4.
Further analyses focused on Period 4, because all fathers fed younger
offspring much less frequently, and because fathers were responsible for
a large proportion of the food delivered to Period 4 nestlings (see STAMPS
et al., 1985 and Table 2). The first question was whether individual males
varied their feeding rates for broods as a function of sex ratio. Of the 14
broods for which data were available, two males were responsible for three broods each, while the remaining eight males were responsible for
one brood apiece. Male # 16 had three broods with Po = 80 % (Brood size, B = 5), 0 % (B = 1) and 50 % (B = 4). His average Period 4 feeding rates per nestling were 3.5 feeds/hr, 0.9 feeds/hr and 0.9 feeds/hr. Male
# 18's three broods had PQ = 100 % (B = 1), 50 % (B = 4), 0 % (B = 1); his average feeding rates per nestling for those broods were 2.7 feeds/hr, 1.76 feeds/hr and 0 feeds/hr. Though hardly conclusive, these data sug-
gest that an individual male's feeding rate might be related to the sex
ratio of a given brood.
Another question was whether the effect of sex ratio on average pater- nal feeding rate was the same for male and female nestlings. For each
brood we calculated separate average Period 4 feeding rates for male and

184
female nestlings. There were similar correlations between sex ratio and
average paternal feeding rates for each sex (females: rs = 0.77, N = 10,
p<0.01, males: rs = 0. 75, N = 8, p<0.05). Recall that in an earlier
analysis, we found no difference in the rates at which fathers fed sons and
daughters within the same brood. Taken together, these data refute the
hypothesis that males were selectively feeding their daughters.
Maternal feeding behavior.
There was no tendency for mothers to feed daughters more than sons (or to feed sisters more than brothers in the same nests) in either Period 3
or 4 (Table 2). However, there was a slight tendency for females to feed
FBB more frequently in Period 3, a tendency which was weakly signifi- cant in Period 4 (Table 3). Correlations between sex ratio and maternal
feeding rate were far from significant in either period; the highest r, value obtained was 0.27 (N = 12), for broods in Period 4.
The relationship between brood size and maternal feeding rate in
Period 3 was the inverse of the male's: nestlings in small broods were fed more frequently (rs = -0.68, p<0.01). Brood size and sex ratio
apparently had independent effects on maternal feed rate, since com-
parable negative correlations between brood size and maternal feed rate
were obtained for FBB (rs = -0. 74, p < 0.05) and MBB (rs = -0.61, n. s. ). As in the males, there were no significant relationships between brood
size and average maternal feeding rates in Period 4.
Combined parental feeding rates and inter family comparisons.
Even though neither parent fed one sex significantly more often than the
other, similar trends in both parents could yield significant differences in
total food delivery to male and female nestlings. However, there were no
significant differences in total food delivery to nestlings for Period 3 or
4 (Table 2). Within families, brothers and sisters obtained an equivalent number of feedings in both periods (Period 3: Wilcoxon Test, T = 11,
N = 7; Period 4: Wilcoxon Test, T = 6, N = 7). Nor was brood size
related to combined parental feeding rates in either period (Period 3:
rs = -0. 24, N = 14, n.s.; Period 4: rs = -0.26, N = 14). Sex ratios within broods had no effect on combined feeding rates in
Period 3 (Table 3). However, the tendency of both parents to feed Period
4 nestlings more frequently resulted in a significantly higher total feeding rate for FBB versus MBB, and for FBF versus MBF (Table 3). Within
FBF, male nestlings tended to be fed more often than their sisters (male nestlings: X ± S.D. = 7.7 ± 2.8 feeds/hr; female nestlings: 5.2 ± 4.0

185
feeds/hr), again refuting the hypothesis that females were preferentially fed within nests. Elsewhere we have shown that parental feeding rates for Period 4 nestlings were positively related to growth rates, and that fathers and mothers probably deliver comparable amounts of food per regurgita- tion (STAMPS et al., 1985). Hence, the difference in feeding rate for female-biased nests probably reflects a real difference in the amount of
food delivered to nestlings within those nests.
Compensatory feeding within pairs.
Instead of combining maternal and paternal feed rates, we can use the same data to answer a different question. Given a certain family type, do parents compensate for the feeding rate of their mate? If so, we would
expect to see an inverse relationship between maternal and paternal feed rates per nest. Since fathers delivered a low proportion of the total
feedings in Period 3, we concentrated on Period 4 offspring for this
analysis. In female-biased nests, there was a strong reciprocal relationship
between maternal and paternal feeding rates (FBB: rs = -0. 76, p < 0.05 ; FBF: rs = -0.90, p<0.05). Parental feeding rates were also inversely related in MBF (rs = -0.80, n.s.), but this was not true when singletons were added to the analysis (MBB: rs = 0.30, n.s.). The most extreme
example of compensation was observed in the female of nest # 211, who
raised two daughters with no male assistance. Her Period 4 feeding rates
were much higher than those of mothers raising FBB, all of whom had
male assistance (FBB: X:t S. D. = 4.0 :t 2.9 feeds/hr; nest # 211: 16
feeds/hr). These results suggest that parents may have a rough "expectation" of
the required feeding rates for a family, and that either sex can compen- sate for shortfalls by the mate. In addition, the "expectations" for
parents with male and female-biased families apparently differed, since
average combined feeding rates were lower for MBF than for FBF,
(Table 3) and since there was no trace of reciprocity when both types of
families were considered together (rs = 0.02, N = 9). Reciprocity was less
evident when singletons were added to the analysis (see FBB and MBB
above); perhaps subtle adjustments of parental feeding rates per nestling were not as important when only one nestling required provisioning.
II. Fledge weights, fledge ages and nestling gender.
Since the offspring in FBB were fed more frequently than those in MBB, and since parental feed rates were positively related to growth rates in

186
Period 3 and 4 (STAMPS al., 1985), we might expect nestlings in FBB
to fledge earlier or at higher weights than those in MBB.
There were no significant correlations between average weight at
fledging per brood and either sex ratio (arcsin Pq as mean fledge weight, r = 0.13, 30 df) or brood size (r = 0.17, 30 df). Nor were there any signifi- cant correlations when the sexes were considered separately (arcsin Pq and average female fledge weight, r = -0.29, 30 df; arcsin Pq and
average male fledge weight, r = 0.22, 24 df). Looking at it another way, we can ask whether female budgerigars tended to fledge at heavier
weights than males. In fact, an opposite trend was detected
(males = 38.5:t 3.3 g, N = 38, females = 37.5 ± 4.3 g, N = 50, p = 0.33). Since there was considerable inter-familial variation in weight at fledg-
ing, we also compared the fledge weights of brothers and sisters within
the same broods. Again, brothers tended to fledge heavier than their
sisters, but the trend was not significant (t = 1.85, 18 df). In wild
budgerigars there are no significant differences in adult body weight
(WYNDHAM, 1978, 1980), and a similar lack of sexual size dimorphism in adult body weight was found in periodic samples of our captive flocks
(CLARK, unpubl. data).
Age at fledging was related to brood size; small broods fledged an
average of two days older than larger broods (1 or 2 fledglings,
average fledge age: k+ S.D. = 34.9 + 1.6 days; 3 to 7 fledglings: 33.3 ± 1.1 days, t = 3.2, 30 df, p < 0.01 ). Within larger broods there was
no significant correlation between brood size and fledge age (r = 0.34, N = 18). Hence, in analyses of fledge age and sex ratio, we considered
small and large broods separately. In small broods there was no significant correlation between average
age at fledging and arcsin Pq (r = 0.18, N = 14). However, for large
broods, arcsin Pq was strongly correlated with average fledge age (r = -
0.71, N = 18, p <0.001, Fig. 2). All large broods plotted along the same
regression line, and there were strong correlations between arcsin Pq ç and average fledge age for broods with 3 (r = -0. 77), 4 (r = -0.95) and 5
(r = -0.84) fledglings (Fig. 2). In large broods brothers and sisters had
comparable fledge ages (Wilcoxon Test, T = 77, N = 16), and there were
comparable negative correlations between average fledge age and arcsin
P 9 for females (r = -0.44) and for males (r = -0.37). Overall, there was
no difference between the average fledge age of males (33.5 + 1.7 days) and females (33.8 + 1.8 days).
There were no significant correlations between average age at fledging for a large brood and other potentially confounding variables, such as

187
Fig. 2. Average fledge age per brood versus the percent females within the brood. Brood sizes: 3 (dots), 4 (squares), 5 (circles), 6 and 7 (diamonds). Barred symbols indicate
broods with estimated sex ratios (see Methods).
brood size (r = 0.34, 16 df, see also Fig. 2), or average fledge weight
(r = -0.19, 17 df). Nor was there a significant relationship within broods
between hatch order and fledge age. Of 18 large broods, 10 had positive
relationships between hatch order and fledge age, and 6 had negative
relationships.
III. Average age at fledging versus onset of the next clutch.
Fledging a brood early might be advantageous to parents if it allowed
them to begin laying their next clutch at an earlier date. Data on fledge
ages and date of onset of the next clutch were available for 20 large broods. Time to onset of the next clutch was measured several ways: FEGG = days from the first egg of the first clutch to the first egg of the
next clutch, LEGG = days from the last egg of the first clutch to the first
egg of the next clutch, INTFIRST = days from the first fledge of the first
clutch to the first egg of the next clutch and INTLAST = days from the
last fledge of the first clutch to the first egg of the next clutch.
None of these measures of the time to the next clutch were significantly related to the average fledge age of the first brood (FEGG vs X Flyage, r = 0.13; LEGG vs X Flyage, r = 0.06; INTFIRST vs X Flyage, r = 0.10; INTLAST vs X Flyage, r = 0.02). Since previous results had shown a
strong positive linear relationship between brood size and days to the
onset of the next clutch (STAMPS et al., 1985), we re-ran this analysis using
multiple regression to statistically control for the linear effects of brood

188
size on onset of the next clutch. The results of these analyses were also
nonsignificant. Hence, we had no indication that fledging young earlier had any effect on the date that parents initiated their next breeding effort.
IV. Parent and offspring behaviour in the first three weeks after fledging.
Age at fledging and fledgling behaviour.
If budgerigar behavioural development were strictly age-dependent, then
early fledging budgerigars might be behaviourally immature relative to those fledging at later ages. Alternately, a flexible development schedule
might result in fledging whenever juveniles had attained a certain stage of maturation, in which case behaviour would be independent of fledge age. Finally, early fledging might indicate an advanced stage of develop- ment, such that early fledging juveniles were behaviourally precocious.
The frequency of social and nonsocial flights was used as one index of behavioural development in the first three weeks after fledging. While
age at fledging was related to flight activity in both sexes, the signs of these relationships were opposing in males and females.
In females, young fledgers were more active than those fledging at older ages. The frequency of nonsocial flights during the first three weeks
post fledging was negatively related to fledge age (rs = -0.58, N = 17,
p < 0.02, Fig. 3), and the correlation between social flights and fledge age had the same sign (rs = -0.15, N = 17). Combining social and nonsocial
flights (total rate of flights) gave a similar picture: younger fledging females flew more often than older fledging females (r, = 0.52, p < 0.05).
In contrast, younger fledging males tended to fly less often than males
fledging at older ages. This trend was nonsignificant for nonsocial flights (rs = 0.30, N = 13, Fig. 3), significant for social flights (rs = 0.60, N = 13,
p < 0.05) and not significant for total flights (rs = 0.39, N = 13). Another index of social activity was the rate at which juveniles initiated
contacts with various other categories of birds (e.g. siblings, father, adult
females, etc. ). These data will be reported in detail elsewhere (STAMPS et
al., in prep.), but for our purpose, the most important result was that
younger fledging individuals were neither retarded nor precocious with
respect to the rate of social contact with flockmates.
Parental behaviour towards fledglings in their first three weeks.
Since fathers preferentially fed offspring in female-biased broods when the young were still in the nest, we wondered if a similar bias existed after

189
Fig. 3. Rate of nonsocial flights for female (dots) and male (circles) fledglings as a func- tion of fledge age.
fledging. Fathers rarely fed fledglings (X = 0.21 ± 0.38 feeds/hr, N = 36
fledglings), so in addition to actual feedings, we also recorded instances
in which fledglings begged from their fathers, or in which fathers
approached their offspring. If anything, fathers tended to favor male-biased broods during the post
fledging period. For example, there was a negative correlation between
average paternal fledgling feeding rate and Pq for the brood (r = -0.68, N = 7 families). Average fledgling beg rates to their father were unrelated
to sex ratio (rs = 0.04), and there was no tendency for fathers to
approach fledglings more often when they belonged to female-biased
families (rs = 0.05).
Similarly, there was no indication that fathers paid more attention to
female fledglings. Paternal feed rates were comparably low for both sexes
(females: X ± S.D. = 0.35 t 0.67 feeds/hr, males: 0.12 ± 0.90 feeds/hr). Both sexes begged at equivalent rates to their fathers, and fathers
approached male and female offspring at comparable rates.

190
Mothers virtually never fed fledglings. Aside from one male who was fed three times by his mother, the only maternal feedings occurred in a brood of two females raised by their mother with no male assistance.
Fledgling beg rates to their mothers were low (0.25 + 0.48 begs/hr, N = 36), and there was no discernible relationship between Pq or off-
spring gender on rate of begging to the mother or approaches by the mother.
V. Age at fledging and eventual reproductive success.
Information on reproduction was available for nine of the females which
fledged in 1979 and were retained in the flocks to breed in 1980. In
general, younger fledging females were more successful breeders in their first year than were older fledgers. First, young fledging females pro- duced a higher percentage of fertile eggs: egg success (# offspring hatched divided by the total number of eggs laid) was negatively related
to female fledge age (rs = -0.74, N = 9, p < 0.05, Table 4). There were
also negative correlations between female fledge age and the number of
offspring fledged in her first brood (r = -0.76, N = 9, p < 0.05), and the number of offspring she fledged during her first breeding season (r, = -
TABLE 4. Reproduction in the first breeding season
* Mate added after the other birds were paired. * * Mate fledge age unknown.

191
0.75, N = 9, p<0.05, Table 4). None of these three variables was
significantly related to fledge weight (Table 4). Also evident in Table 4 is an interesting correlation between female
fledge age and that of her mate. If we include the 1979 and 1980 pairs for which both fledge ages are known, and in which birds had
unrestricted mate choice (e.g. excluding YF32, see below), there was a
strong correlation between the fledge ages of mated pairs (rs = 0.73,
p<0.05). YF32 was excluded from this analysis because she was
unpaired at the beginning of the 1980 breeding season. A male with an
early fledge age was then added to the flock, and he subsequently paired with YF32. Because of the disparity of fledge ages within this pair, it was
possible to look at the effects of male fledge age on reproductive success.
Fledge age of males was not significantly correlated with either egg suc-
cess (rs = -0.38), number of young fledged from the first brood (rs = -
0.41) or total number of young in the first breeding season (r = 0.17)
(Table 4). Our results also hint that older-fledging females may have difficulty
attracting mates. Despite the presence of unmated males in the flight
cages, three females with known fledge ages failed to obtain mates. All
three of these females fledged at above average ages (Table 4).
Discussion
A complex pattern of budgerigar parental behaviour is suggested by this
and a previous study (STAMPS et al., 1985). Female budgerigars are
primarily responsible for the early care of offspring, during which time
they allocate food provided to them by their mates. When offspring are
small (Period 1 and 2 and 3), mothers allocate food according to offspring size, and generally ignore offspring gender or beg rate. This tactic
ensures that all members of the brood have equivalent growth rates,
despite considerable asynchrony in hatch dates for brood members.
However, by the start of Period 4 most nestlings have achieved asymp- totic mass, so that size is no longer a useful cue for food allocation. At
this point, mothers may attend to the sex ratio of the brood, since
mothers fed offspring in female-biased broods more than those in male-
biased broods.
In contrast, male parents allofeed their mates during incubation,
brooding and the nestling period. They may also feed the young directly, and the male tendency to feed nestlings is positively related to the propor- tion of female offspring in the brood. The immediate cue used for food
allocation after males enter the nest is offspring begging rate, and the

192
most vigorously begging nestlings on any given feeding visit are fed the most by their fathers.
This pattern of parental care raises a host of questions. First, what
behavioural mechanisms could yield the observed strong correlations
between brood sex ratio and paternal care? One possibility is that fathers
might use early sex differences in behavior to determine brood sex ratios.
Young females tend to start begging at earlier ages and beg at higher rates in Period 1 and 2 than do young males (STAMPS et al., in prep.). In
addition, the begging calls of young males and females may differ
acoustically, as seems to occur in zebra finches (BALDA, pers. comm.). Since males tend to enter boxes after they hear begging calls (STAMPS et al., 1985), they may use auditory rather than visual cues to determine
brood sex ratios. Another hypothesis is that pairs are able to adjust the sex ratios of their
broods. For example, mothers might produce female-biased broods when
they had evidence of male condition and interest in allofeeding. They
might even use male allofeeding rates prior to egg laying as an estimate
of male feeding rates for nestlings later in the nest cycle (e.g. NISBET,
1973; NIEBUHR, 1981). Alternately, both sex ratio and male feeding behaviour might covary in response to some external environmental fac-
tor (e.g. date of brood initiation within the breeding season). However, this hypothesis is unlikely for several reasons. First, there is
little evidence for a primary sex ratio bias in other avian species (reviews in NEWTON & MARQUISS, 1979; PATTERSON & EMLEN, 1980; FIALA, 1981b;
RICHTER, 1983; BANCROFT, 1984a). Second, there was little evidence for
either primary or post-hatching sex ratio adjustment in our flocks. Partial
brood mortality was rare (91 % of broods fledged all hatchlings), and
nestling mortality was not skewed toward either sex (CLARK et al.,
unpubl. data). Third, there was no indication of sex ratio bias among
nestlings, in relation to hatch order, identity of one or both parents, prior
breeding experience of parents, date in the breeding season, year, or
when all the data were combined (CLARK, unpubl. data). And finally, two females in our flocks successfully raised broods with no male
allofeeding prior to or after egglaying, and both of these broods were
female-biased. Hence, the correlation between paternal care and brood sex ratio is more likely a result of paternal discrimination than of
adjustments of sex ratios within broods.
A second question is why fathers (and possibly mothers) fed according to brood sex ratio, instead of selectively feeding the female nestlings. We have shown in a previous study that selective feeding slows the rate of

193
food delivery in budgerigars: mothers feed more slowly than fathers, and
fathers choosing between allofeeding their mates and their nestlings feed
more slowly than when they nonselectively feed just the nestlings
(STAMPS al., 1985). Hence, discrimination of each offspring's gender prior to each regurgitation could appreciably reduce the food delivery rate for both mothers and fathers. In addition, budgerigars are hole nesters and it is dark at the bottom of the nest holes. Parents returning from an outside foraging trip would probably have to wait some time
after entering before their eyes dark adapted enough to distinguish individual young. Fathers might always have this problem, since their
visits into the nest hole are brief throughout the nest cycle. In contrast, mothers usually remained in the box for long periods while brooding and
allocating food, so dark adaptation would primarily affect them late in the nest cycle, when they began to make foraging trips to provision older
nestlings. SKUTCH (1976) discusses other evidence that parent birds may have
difficulty discerning offspring under low light conditions. He notes that
nestlings of hole nesting species often have more conspicuous mouthparts
(e.g. larger flanges, brighter gape markings, more colorful mouth linings) than their open nesting counterparts. The pattern of parental care may also affect the conspicuousness of nestling mouthparts. In particular, we
might expect that nestlings in species in which both parents foraged outside of the nest when the nestlings were young might have more con-
spicuous "feeding targets" than related hole nesting species in which one
of the parents remained in the nest for long periods and allocated food
provided by the other parent. Another set of questions relates to the possible ecological or evolu-
tionary significance of the feeding patterns reported here. Clearly, we
can't say anything definitive about the significance of these results for
wild birds under field conditions. However, the domesticated
budgerigars used in these studies were tested in the environment in which
they have lived, and been selected, for the past 50 to 100 generations
(SOSSINKA, 1982; PRICE, 1984). Hence, it is relevant to ask whether the
additional feeding of female-biased broods leads to higher reprodutive success under captive conditions (e.g. food ad lib and close to the
nestboxes, high densities, competition for preferred nestboxes) and leave
aside for the moment the possible significance of such a behaviour pat- tern for wild budgerigars.
Our results suggests that the extra feeding of female-biased broods
was adaptive for all but the smallest broods. Very small broods (one or

194
two nestlings) fledged two days older than larger broods, and in small
broods, sex bias had no discernable effect on fledge age. However, a
strong negative relationship between sex ratio and average fledge age was
obtained for larger brood sizes. Fledge ages in turn were significantly
negatively related to female flight rates within the first three weeks after
fledging, and to female reproductive success (number of young fledged) in the subsequent breeding season. There were few significant relation-
ships between these variables and fledge age for male budgerigars. Indeed, the only significant result was that early fledging males tended
to fly less often in social contexts than did their later fledging counter-
parts. Hence, the parental care pattern may favor offspring of the sex
which can most benefit from extra feedings. At this point it is unclear why higher feeding rates might benefit
female nestlings more than males. In contrast to species with sexual size
dimorphism, there was no significant sex difference in mass for either
fledglings or adults; if anything, males tended to fledge heavier than
females. Another possibility is that females benefit from early maturity because of their competitive role in budgerigar society. Adult females
have primary responsibility for the acquisition and defense of nest sites
(MASURE & ALLEE, 1934; CLARK, unpubl. data). The most severe and
vigorous fights observed in captivity occurred when females contested
preferred nestboxes, and females played the major role in defending nestboxes against conspecifics throughout the breeding cycle (CLARK,
unpubl. data). Perhaps younger fledging females are better competitors
(e.g. because of higher agility or behavioural precocity). Currently we are
investigating the behavioural development of females and males, to see
if fledge age is related to success in dominance interactions, the early
acquisition of a mate, or other behaviours of potential importance for
subsequent reproductive success. The definition for parental investment (TRIVERS, 1972) requires that
parental care increase the reproductive success of the offspring while
decreasing the subsequent reproductive success of the parent. In this
study there was no indication that pairs fledging female-biased broods renested any later than those producing male-biased broods. Of course, other assays of future reproductive success might have revealed an effect.
For example, pairs with female-biased broods might fledge smaller subse-
quent broods than those with male-biased broods. However, it is also
possible that in our field cages, the costs to pairs of providing additional
feedings to FBB were negligible. As RICHTER (1983) has pointed out, if
food is not limited, parental investment in males and females can be
equal even though one sex requires more energy to raise than the other.

195
Assuming that juvenile female budgerigars benefit from extra
feedings, why should the male provide most of this extra cate? One
possibility is that by the time offspring reach Period 4, females are so
stressed by their previous parental efforts that they can not provide the
required extra care to female-biased broods. Unless the male contributed
this extra care, those broods would fledge late, with a consequent reduc- tion in fitness for the female offspring within those broods.
This "female exhaustion" hypothesis seems unlikely to apply to
budgerigars, for two reasons. First, females with FBB were able to com-
pensate for shortfalls in Period 4 food delivery by their mates, and some
females even managed to rear FBB with no male help whatsoever. Sec-
ond, budgerigars share with many other species a parental role division
pattern that SKUTCH (1976) calls the "hornbill" pattern after the group in which it is most highly developed. In species with this pattern, females
specialize in incubation, brooding, guarding the nest, and allocating food
to the young, while males feed their mates during incubation, and later
provision the mate, who transfers food to the young. When the offspring are older, the female may also begin to forage on her own and thus
directly feed the young, but the extent of female provisioning is variable
and often lower than that of the male. The hornbill pattern occurs in
many parrots (references below, see also SMITH, 1972, 1975; SAUNDERS,
1982), birds of prey (hawks, owls, shrikes), and hornbills, is common
among corvids, and sporadic in several other groups, including
passerines (reviews in KENDEIGH, 1952; SKUTCH, 1976; see also NEWTON,
1978; COLLOPY, 1984). An energy budget for two such species suggests that males perform most of the energetically expensive duties, and that
if either parent is stressed midway through the nest cycle, it ought to be
the male (1VIUNGAAS & KING, 1981; ROSKAFT, 1983). Another hypothesis for males taking responsibility for the more expeil-
sive sex is that by doing so, males increase their future success with
subsequent broods. This hypothesis assumes that the success of the current
brood is unaffected by the gender of the parent providing the extra care, but that males which free their mates from this responsibility have
greater success with subsequent broods than do less solicitous males. In
budgerigars, there seems to be an important tradeoff for females between
feeding current young and initiating the next clutch. This was indicated
by a strong positive relationship between brood size and time to the first
egg of the next clutch, and females raising young without male assistance
waited significantly longer before renesting than did females raising com-
parably sized broods with male help (STAMPS et al., 1985). Hence, if

196
females had to provide all of the additional feedings going to the nestlings in FBB in Period 4, they might have to delay the onset of the next clutch.
In consequence, both the female and her mate would experience a
reduced rate of brood production. Note that under the system used by
budgerigars (male provides most of the extra care), there was no signifi- cant delay in renesting for pairs raising FBB, as compared to those rais-
ing MBB.
In view of the strong gender effects on feeding in budgerigars, it is
interesting that no gender effects have been reported in three other avian
species (redwinged blackbirds, FIALA, 1981a; rooks, ROSKAFT &
SLAGSVOLD, 1985; sparrowhawks, NEWTON, 1978, see Introduction). For
good logistical reasons, these and other field studies usually don't follow
parental provisioning patterns during the post fledging period, even
though the period of parental provisioning outside of the nest often
exceeds the period within it. Since in budgerigars the interaction of
parent and offspring gender became most interesting soon before the off-
spring were independent, the same might be true in other birds. It would
be interesting to look at post-fledging provisioning by male and female
parents of male and female young in sexually dimorphic birds such as
rooks, hawks or blackbirds. This approach is especially promising, since
birds often seem to divide the fledglings among the two parents (e.g. NOLAN, 1978; SMITH & MERKT, 1980; MORENO, 1984; review in HARPER,
1985), and in at least one of these cases, the broods seemed to be divided
along sexual lines (HARPER, 1985). Sex-biased brood division might
simplify the delivery of extra investment to one offspring sex, since the
more expensive offspring sex could be "allocated" to the parent sex best
able to deliver the required extra care.
Interactions between parent and offspring gender may occur in many other birds besides budgerigars. One suggestive bit of evidence is that
paternal rates of provisioning are often more variable than maternal rates
(e.g. MAXWELL & PUTMAN, 1972; UTTER & LEFEBVRE, 1973; HOWE, 1979;
SAUNDERS, 1982; BALL, 1983). Some of this variance may occur because
males respond more strongly than females to increases in brood size (e.g. HowE, 1979; FIALA, 1981a). However, it is also possible that males are
more apt than females to respond to the extra needs of a more expensive
offspring sex. Monogamous species with variable paternal provisioning would be opportune subjects for further study of this problem.

197
Summary Parental feeding rates in relation to the sex of parents and offspring were studied in domesticated budgerigars in large flight cages at Davis, California. While neither the male nor the female parent preferentially fed offspring of either sex, paternal feeding rates were strongly related to the sex ratio of the brood. Fathers fed female-biased broods much more frequently than male-biased broods throughout the nest cycle, and male feeding rates were highly correlated with the sex ratios of the broods. Mothers exhibited a similar but much weaker tendency to feed female-biased broods more often, and only at the end of the nest cycle was there a barely significant relationship between sex bias and maternal feeding rate. As a result of the extra parental care, female-biased broods obtained nearly three times more regurgitations in the final pre-fledge period than male- biased broods.
Brood sex ratios were unrelated to fledge weights, and male and female nestlings fledged at comparable weights. However, for all but the smallest broods, there was a strong negative relationship between the percent of females within the brood and the average age of fledging. Fledge age, in turn, was related to post-fledge activity rates. Females fledging at younger ages initiated more flights than those fledging at older ages. Conversely, in males, fledge age was either unrelated, or positively related to the rate of flights. There were also significant negative relationships between female fledge age and reproductive success (e.g. number of young fledged in the first breeding season), but no significant relationships between male fledge age and reproductive success. Hence, male parents may provide extra care to broods biased toward the offspring sex which most benefits from this extra care.
References
BALL, G. F. (1983). Evolutionary and ecological aspects of the sexual division of parental care in barn swallows. � Rutgers Univ.: Ph.D. thesis.
BANCROFT, G. T. (1984a). Patterns of variation in size of boat-tailed grackle Quiscalus major eggs. - Ibis 126, p. 496-509.
�� (1984b). Growth and sexual dimorphism of the boat-tailed grackle. - Condor 86, p. 423-432.
CLUTTON-BROCK, T. H. & ALBON, S. D. (1982). Parental investment in male and female offspring. � In: Current problems in sociobiology (King's college sociobiology group, eds), p. 223-247. Cambridge University Press, Cambridge.
��, GUINESS, F. E. & ALBON, S. D. (1982). Red deer: behaviour and ecology of two sexes. - Chicago University Press, Chicago.
COLLOPY, M. W. (1984). Parental care and feeding ecology of golden eagle nestlings. - Auk 101, p. 753-760.
FIALA, K. L. (1981a). Reproductive cost and the sex ratio in red-winged blackbirds. � In : Natural selection and social behavior (R. D. ALEXANDER & D. W. TINKLE, eds), p. 198-214. Chiron, New York.
�� (1981b). Sex ratio constancy in the red-winged blackbird. - Evolution 35, p. 898-910.
- - & CONGDON, J. D. (1983). Energetic consequences of sexual size dimorphism in nestling red-winged blackbirds. - Ecology 64, p. 642-647.
HARPER, D. G. (1985). Brood division in robins. - Anim. Behav. 33, p. 466-480. HOWE, H. F. (1979). Evolutionary aspects of parental care in the common grackle,
Quiscalus quiscula L. � Evolution 33, p. 41-51. KENDEIGH, S. C. (1952). Parental care and its evolution in birds. - Illinois Biol.
Monogr. 22, p. 1-356. MARTINDALE, S. (1984). Sexual dimorphism and parental role switching in gila
woodpeckers. � Wilson Bull. 96, p. 117-121.

198
MASURE, R. H. & ALLEE, W. C. (1934). Flock organization of the shell parakeet Melopsit- tacus undulatus Shaw. - Ecology 15, p. 388-398.
MAXWELL, G. R. & PUTNAM, L. S. (1972). Incubation, care of young and nest success of the common grackle (Quiscalus quiscula) in northern Ohio. - Auk 89, p. 349-359.
MAYNARD-SMITH, J. (1980). A new theory of sexual investment. - Behav. Ecol. Sociobiol. 7, p. 247-251.
MORENO, J. (1984). Parental care of fledged young, division of labor, and the develop- ment of foraging techniques in the Northern wheatear (Oenanthe oenanthe L.). - Auk 101, p. 741-752.
MUGAAS, J. N. & KING, J. R. (1981). Annual variation of daily energy expenditure by the black-billed magpie: A study of thermal and behavioral energetics. - Studies in Avian Biology 5, p. 1-78.
NEWTON, I. (1978). Feeding and development of sparrowhawk Accipiter nisus nestlings. - J. Zool. Lond. 184, p. 456-487.
- - & MARQUISS, M. (1979). Sex ratio among nestlings of the European sparrowhawk. - Am. Nat. 113, p. 309-315.
NIEBUHR, L. (1981). An investigation of courtship feeding in herring gulls, Larus argen- tatus. - Ibis 123, p. 218-223.
NISBET, I. C. (1973). Courtship-feeding, egg-size and breeding success in common terns. - Nature, Lond. 241, p. 141-142.
NOLAN, V. (1978). Behavior and ecology of the prairie warbler. - Am. Ornithol. Union Monogr. 14, p. 1-595.
PATTERSON, C. B. & EMLEN, J. M. (1980). Variation in nestling sex ratios in the yellow- headed blackbird. � Am. Nat. 115, p. 743-747.
POWER, H. W. (1980). The foraging behavior of mountain bluebirds, with emphasis on sexual foraging differences. - Washington D.C., Amer. Ornithol. Union, Ornithol. Mon. 28, pp. ix + 72.
PRICE, E. O. (1984). Behavioral aspects of animal domestication. - Quart. Rev. Biol. 59, p. 1-32.
REITER, J., STINSON, N. L. & LEBOEUF, B. J. (1978). Northern elephant seal develop- ment : the transition from weaning to nutritional development. � Behav. Ecol. Sociobiol. 3, p. 337-367.
RICHTER, W. (1983). Balanced sex ratios in dimorphic altricial birds: the contribution of sex specific growth dynamics. - Am. Nat. 121, p. 158-171.
RØSKAIFT, E. (1983). Sex role partitioning and parental care by the rook Corvus frugilegus. - Ornis Scand. 14, p. 180-187.
- - & SLAGSVOLD, T. (1985). Differential mortality of male and female offspring in experimentally manipulated broods of the rook. - J. Anim. Ecol. 54, p. 261-266.
SAUNDERS, D. A. (1982). The breeding behaviour and biology of the shortbilled form of the white-tailed black cockatoo, Calyptorhynchus funereus. - Ibis 124, p. 422-456.
SKUTCH, A. F. (1976). Parent birds and their young. � Univ. Texas Press, Austin, Texas. 503 pp.
SMITH, G. A. (1972). Nestling and nestling parrots. - Avicult. Mag. 78, p. 155-164. �� (1975). Systematics of parrots. - Ibis 117, p. 18-68. SMITH, J. N. M. & MERKT, J. R. (1980). Development and stability of single-parent
family units in the song sparrow. � Can. J. Zool. 58, p. 1869-1875. SOSSINKA, R. (1982). Domestication in birds. - In: Avian biology, vol. 6 (D. S. FARNER
& J. R. KING, eds), p. 373-403. Academic Press. STAMPS, J., CLARK, A., Kus, B. & ARROWOOD, P. (1985). Parent-offspring conflict in
budgerigars. - Behaviour 94, p. 1-40. TRIVERS, R. L. (1972). Parental investment and sexual selection. - In: Sexual selection
and the descent of man (B. CAMPBELL, ed.), p. 136-197. Aldine Chicago. �� (1985). Social evolution. - Benjamin Cummings, Menlo Park, Calif.

199
TRIVERS, R. L. & HARE, H. (1976). Haplodiploidy and the evolution of the social insects. - Science 191, p. 249-263.
UTTER, J. M. & LEFEBVRE, E. A. (1973). Daily energy expenditure of purple martins (Progne subis) during the breeding season: estimates using D2O18 and time budget methods. � Ecology 54, p. 597-605.
WITTENBERGER, J. F. (1982). Factors affecting how male and female bobolinks apportion parental investments. � Condor 84, p. 22-39.
WYNDHAM, E. (1978). Ecology of the budgerigar Melopsittacus undulatus (Shaw) (Psit- taciformes : Platycercidae). - Univ. New England Armidale: Ph.D. thesis.
�� (1980). Diurnal cycle, behavior and social organization of the budgerigar Melopsit- tacus undulatus. � Emu 80, p. 25-33.




















